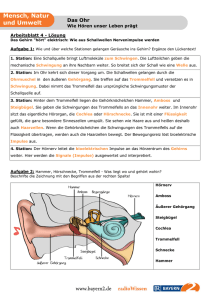
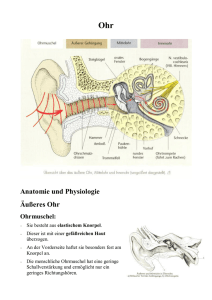
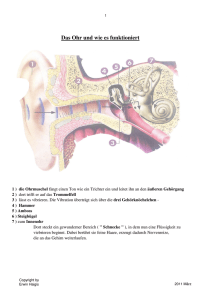
sinne - cipmm
Werbung

SINNE Im Praktikumsteil „SINNE“ lernen Sie die Sinnesorgane AUGE, OHR, VESTIBULARORGAN kennen. Bitte lesen sie noch vor dem Praktikum die entsprechenden Teile der Praktikumsanleitung und machen sie die unten stehenden Hausaufgaben. Wiederholen Sie bitte vor dem Praktikum auch die Kapitel „Wellenlehre“, „Akustik“ und „Optik“ aus der Physikvorlesung sowie die morphologisch-histologischen Kapitel „Auge“, „Ohr“ und „Vestibularorgan“ aus den Anatomie- und Physiologievorlesungen. Zusätzliche vertiefende Informationen finden sie in den Physiologielehrbüchern. Alle dort behandelten Themen sind klausur- und testatrelevant. Zum besseren Verständnis gibt es in den Praktikumsräumen Informationstafeln und Modelle von Auge, Ohr und Vestibularorgan. In der Praktikumsanleitung befinden sich Informationen zur Ophthalmologie und Audiologie, die auf den Praktikumsversuchen basieren und in der klinischen Ausbildung vorausgesetzt werden. Darüber hinaus werden im KlinischPhysiologischen Seminar ausgewählte pathophysiologische Aspekte und Krankheitsbildern behandelt. Die fünf nachstehenden Hausaufgaben sind obligatorisch und werden von den Versuchsleitern vor Praktikumsbeginn kontrolliert Alle zur Lösung der Aufgaben notwendigen Informationen sind in der Praktikumsanleitung enthalten. Hausaufgabe 1: Bitte beschriften Sie diese Zeichnung. Hausaufgabe2: Klassifizieren Sie diese Augen und benennen sie die notwendigen Korrekturlinsen. SINNE Hausaufgabe 3: Beschriften Sie die rot markierten Strukturen des Ohrs- und des Vestibularorgans. Hausaufgabe 4: Beschriften Sie das Hörfeld ( Größenordungen der X + Y Achse, Hauptsprachbereich). Zeichnen Sie zwei Punkte mit gleicher Lautstärke (Kreise) und zwei Punkte mit gleichem Schalldruck (Dreiecke) ein. Hausaufgabe 5: Beschriften Sie die Strukturen des linken Bogengangs und zeichnen Sie den rechten Bogengang daneben. Tragen sie temporal und nasal ein. 2 SINNE 3 SEHEN Autoren: Janine Mohrbach, Prof. Dr. H.-Peter Richter, Dr. Helmut Machulla, Dr. Bernd Bufe Das Auge ist unser wichtigstes Sinnesorgan! Pro Sekunde sendet das Auge etwa 40 Millionen optische Details an das Gehirn. 80 % unseres Wissens haben wir mit den Augen erworben; die restlichen 20 % teilen sich Gehör, Gleichgewichtssinn, Tastsinn, Geruch- u. Geschmacksinn. Unser Sehvermögen wird erst im Laufe von etwa 10 Jahren optimal ausgebildet und ist abhängig von allen strukturellen, opto-physikalischen, sensor-physiologischen, psycho-physikalischen, neuro-anatomischen Parametern der paarigen Sinneskanäle. Inhalts-Übersicht Seite A. Physikalische Grundlagen 4 B. Anatomische Grundlagen 6 1. Aufbau des Bulbus 2. Sehbahn C. Physiologische Grundlagen 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. 7 Brechkraft Transduktion Adaptation Akkommodation OKN, EOG, VEP Primär-Rezeptives Feld Gesichtsfeld Augen-Innendruck Sehschärfe Farbensehen Tiefen-Wahrnehmung D. Anleitungen zu den Aufgaben -- Hausaufgaben A) - D) -- Durchführung der Versuche 1 - 5 -- Klinische Bezüge 17 SINNE 4 A. Physikalische Grundlagen Sichtbares Licht wird als elektromagnetische Strahlung eines Wellenlängenbereiches von 400 nm (Violett-Blau) über Grün bis 750 nm (Rot) wahrgenommen. Ultraviolettes Licht hat Wellenlängen von kleiner als 400 nm; Infrarotlicht größer als 750 nm. Also je kurzwelliger Licht ist, desto energiereicher ist es. Errechnet aus Frequenz f mal Wellenlänge λ beträgt die Lichtgeschwindigkeit c0 im Vakuum 2,99108 [m/s]: c = f λ_ Da die Geschwindigkeit für alle Farben gleich ist, bedeutet das: je größer λ, desto niedriger f, und umgekehrt. Beim Durchtritt durch den optisch dichteren dioptrischen Apparat des Auges wird fokussierend zum Lot hin gebrochen. Refraktion: Je optisch dichter das Material ist, desto größer ist n, desto langsamer breitet sich das Licht in diesem Medium aus, desto stärker wird es abgelenkt = gebrochen. Der Brechungsindex (Brechzahl) n des Mediums ist das Verhältnis der AusbreitungsGeschwindigkeit c im Vakuum zur Geschwindigkeit v im Medium. Sie ist eine Materialkonstante; die von Wellenlänge und Frequenz des einfallenden Lichtes abhängt. n = c/v >1 [ohne Dimension] Es gilt der Satz nach Snellius: „Trifft Licht schräg auf eine Grenzfläche aus einem optisch dünneren Medium mit niedrigerer Brechzahl auf ein optisch dichteres Medium mit höherer Brechzahl, so wird es zum Lot hin gebrochen; tritt es von einem optisch dichteren in ein dünneres Medium, so wird es vom Lot weg gebrochen“. sinα1 n1 = sinα2 n2 (α = Winkel zwischen Strahl u. Lot) Reflexion: An Grenzflächen werden einfallende Strahlen immer nur zum Teil gebrochen, der Rest wird ohne in das Medium einzudringen reflektiert. [Einfallswinkel = Reflexionswinkel ]. Streuung: Ein Teil der eingetretenen Strahlung ändert beim Durchtritt durch Materie aufgrund von Wechselwirkungen seine Richtung (= Tyndall-Effekt). Weichen Wellen von ihrer geradlinigen Ausbreitungsrichtung ab (Fermat-Prinzip), wenn sie auf spaltförmige Öffnungen treffen und sich überlagern (Interferenz), entsteht Beugung (z.B. am Rand der Iris). Abbildungsfehler: Optische Linsen brechen an ihrer Oberfläche mehrfarbiges Licht ungleichmäßig. Diese chromatische Aberration beruht darauf, dass Licht verschiedener Wellenlänge unterschiedlich stark gebrochen wird (Blau wird stärker gebrochen als Rot). Zusätzlich wird Licht am Rand der Linse stärker gebrochen, da sich hier der Krümmungsradius stark ändert sphärische Aberration. Korrigiert wird dieser Augenfehler durch die Blendenwirkung der Iris, die den peripheren Lichteinfall minimiert. Merke: Jedes Auge hat einen durch den Druck der Augenlider bedingten physiologischen Astigmatismus, weil durch diesen die Corneakrümmung in vertikaler Ebene unterschiedlich von der horizontalen Ebene wird. Beim Astigmatismus ist die Abbildung auf der Retina nicht mehr punktförmig, sondern oval (Stabsichtigkeit). SINNE Vereinfacht lässt sich der Strahlengang durch das Auge mit einer Sammellinse darstellen. 5 aus: ÄP Physiologie Der Zentralstrahl einer Linse wird nicht gebrochen. Alle Parallelstrahlen verlaufen zunächst parallel zur optischen Achse und werden nach Auftreffen auf der Linse so gebrochen, dass sie durch den hinteren Brennpunkt (Fokus) verlaufen. Bei den Brennpunktstrahlen verhält es sich genau umgekehrt. Zur Konstruktion eines solchen Strahlenganges reichen in der Regel zwei Strahlen aus, zum Beispiel ein Parallelstrahl und Zentralstrahl. Das entstandene Bild auf der Retina ist verkleinert, umgekehrt, reell. Der Kehrwert der Brennweite f wird als Brechkraft (Brechwert) D einer Linse bezeichnet: D=1/f [1/Entferunng in Meter] in Dioptrie [dpt] Die Gegenstandsweite g (Entfernung des Gegenstandes zur Hauptebene), die Bildweite b (Entfernung b des Bildes von der Hauptebene) und Brennweite f stehen in folgender Beziehung: 1/f = 1/g + 1/b (= Abbildungsgleichung) Grundlegende Beispielrechnungen: 1) Wie stark muss die Linse für ein Objekt in 2 m bzw. 0,2 m Abstand akkomodieren? Es gilt: D=1/f. Für 2 m folgt daraus D=1 / 2 = 0,5 dpt, für 0,2 m folgt daraus 1 / 0,2 = 5 dpt 2) Die Brechkraft der Cornea beträgt 43 dpt. Wo liegt ihr Brennpunkt f? Nach Umformung der Gleichung aus (1) gilt: f = 1/D. Bei 43 dpt folgt daraus 1 / 43 = 0,023 m. 3) Bei einem kurzsichtigen Auge beträgt der Abstand des Fernpunkts zum Auge 2 m. Welchen Brechwert muss eine vorgesetzte Linse haben, um den Fernpunkt auf 20 cm zu verlagern? Sehfehler des myopen Auges: f = 1/D = 1/ 2 m = + 0,5 dpt Erforderliche Brechkraft für 0,2 m: f = 1/D = 1/ 0,2 m = + 5 dpt Differenz: 4,5 dpt (= 5 dpt – 0,5 dpt) SINNE 6 4) Mit einer Linse mit einer Brennweite f von 0,3 m soll ein 2 m entfernter Gegenstand (g = 2 m) scharf abgebildet werden. In welchem Abstand b hinter der Linse muss der Schirm aufgestellt werden? Es gilt: 1/f = 1 / g + 1 /b. Da f und g gegeben sind, wird die Linsengleichung nach b umgestellt (Zwischenschritte: 1/b = 1/f - 1/g, f·1/b= 1-f /g, f·g · 1/b = g-f, 1/b = (g-f / f·g) daraus folgt b = f·g / (g-f). b = 0,3 m · 2 m / (2 m - 0,3m) = 0,35 m. Der Schirm muss also 35 cm hinter der Linse aufgestellt werden. B. Anatomische Grundlagen 1. Aufbau des Augapfels (Bulbus) Der annähernd kugelförmige in der Orbita liegende Augapfel (Ø 24 mm) wird durch die 6 äußeren Augenmuskeln bewegt. Er hat einen dreischichtigen Aufbau: Die äußere Schicht (Tunica fibrosa bulbi) gliedert sich in Lederhaut (14 sklera) als hinteren Teil und in die Hornhaut (2 cornea) als vorderen stärker gekrümmten, lichtdurchlässigen Teil. Bei der Cornea wird die Vorderfläche (5) durch ein mehrschichtiges unverhorntes Nach: R. F.Schmidt, G. Thews, F. Lang Physiologie des Menschen (2000) Plat-tenepithel gebildet, während die Rückfläche (4) durch einschichtiges Plattenepithel gebildet wird. Zwischen den beiden Basallaminae und der Bowman- u. Descemet-Lamina liegen 45-70 Lamellen aus dicht gepackten Kollagen-Fibrillen und vereinzelten Fibrozyten. Die mittlere Schicht des Bulbus ist die Aderhaut (11 chorioidea) als hinterer Teil der aus drei Abschnitten bestehenden Gefäßhaut Uvea Tunica vasculosa bulbi. Die Iris dient der Adaptation: ihre Blendenöffnung (Pupille) wird durch die beiden intraokularen glatten Augenmuskeln verengt. Miosis, durch den ringförmigen, parasympathisch innervierten musculi sphincter pupillae (3) oder erweitert Mydriasis, durch den speichenförmigen, sympathisch innervierten musculus dilatator pupillae (7). Der Ziliarkörper enthält den ringförmigen, parasympathisch innervierten musculus ciliaris (8). Die zugfesten Zonulafasern (9) dienen der Befestigung der Linse (10) am Ziliarkörper. Dessen Epithel filtriert und sezerniert das Kammerwasser als Ultrafiltrat bei einem peripherem Blutdruck von ca. 45 mmHg, wovon 85 % aus der hinteren Augenkammer durch die Pupille in die vordere Augenkammer (6) und über das Trabekelwerk (hier Resorption) in den venösen Schlemmkanal (1) abfließen, der einen Druck von nur ca. 9 mmHg aufweist. Der Rest an Kammerwasser (15 %) wird uveo-skleral resorbiert. Die tunica interna bulbi besteht aus dem Pigmentepithel (13) mit dicker Bruch´scher Lamina und der Retina (12). Hier liegen die Papille (18) als Austrittsort des nervus opticus (19) aus dem Bulbus sowie der gelbe Fleck (16 macula lutea) mit der fovea centralis (17) - seiner zentralen Vertiefung. Diese enthält nur Zapfenzellen, der para-foveale Ring hingegen fast nur Stäbchenzellen. Der gelartige Glaskörper (15) ist im Zentralbulbus. SINNE 7 Die lichtempfindliche Retina besteht aus etwa 9 Struktur-Schichten und ist im Mittel ca. 200 µm dick. Das einschichtige Pigmentepithel versorgt die Photosensorzellen mit Metaboliten und phagozytiert die verbrauchten Segmente der Außenglieder. Zwischen Pigmentepithel und Sensorzellen existieren keinerlei Haftstrukturen! Der Augeninnendruck (OID) verhindert über den Glaskörper die Ablösung der Netzhaut vom Pigmentepithel (Retinitis pigmentosa). Die Sensorzellen stellen anatomisch das 1. aus: U. Schuhmacher e. a. Prometheus Lernatlas der Anatomie (2007) Neuron der Sehbahn dar und sind daher primäre Sinneszellen. Da sie jedoch selbst kein Aktionspotenzial ausbilden, sind sie funktionell sekundäre Sinneszellen. Man unterscheidet zwei Typen: Stäbchen (100-125 Millionen / Auge), für skotopisches = Dämmerungs- u. Nachtsehen und Zapfen (6-7 Mio) für photopisches = Farbensehen. Die Sensoren liegen auf der licht-abgewandten Seite der Retina (inverser Aufbau; Schutz vor zu starker Lichtintensität und für optimale Versorgung mit Nährstoffen). 2. Die Sehbahn Nach der Aufnahme des Lichts durch die Photosensorzellen (1. Neuron) wird die Lichtenergie durch diese in elektrisch-chemische Aktivität transformiert und als via Synapse an die nachgeschalteten Bipolarzellen (2. Neuron) weitergegeben, die wiederum über die nächste Synapse die Ganglienzellen als drittes Neuron (etwa 1,2 Mio) aktiviert. In der Fovea gibt es eine 1:1:1 -Verschaltung für die Zapfen-Photosensorzellen. D.h., dass die Transduktion von nur einer Zapfenzelle via nachgeschalteter Bipolarzelle auf nur eine Ganglienzelle stattfindet. Dies erlaubt eine hohe Auflösung. In der Peripherie werden viele Photosensorzellen über wenige Bipolarzellen verschaltet auf noch weniger Ganglienzellen „gebündelt“ (Prinzip der Konvergenz, geringe Auflösung bei höherer Lichtsensitivität). Ganglienzellen sind die ersten Zellen der Netzhaut, in denen Aktionspotenziale gebildet werden. Die anderen Neuronen der Retina haben hingegen ein graduiertes Rezeptorpotential (elektrotonische Erregungsleitung). Die Axone aller Ganglienzellen verlassen am blinden Fleck den Bulbus und bilden - zum Teil myelinisiert (Oligodendrozyten) - den nervus opticus. Entsprechend ihrer Zielregion, Aufbau und Informationsverarbeitung unterscheidet man drei Haupttypen retinaler Ganglienzellen: Ca. 10% sind achromatische, schnell leitende magnozelluläre Zellen (M-Zellen = α-(Y)Zellen = parasol-Ganglienzellen) “ der Schichten 1 und 2 im corpus geniculatum laterale=CGL) zum Bewegungssehen. Ca. 80% sind kleine parvozelluläre Zellen (P-Zellen = β-(x)- oder midget-Ganglienzellen) der Schichten 3-6 im CGL zur Analyse von Mustern und Farben. Weitere ca. 10% sind koniozellulär Ganglienzellen = K-Zellen (γ-W-Zellen) bistratified (=‚zweifach geschichtet‘).-zur Steuerung der Pupillenweite (Erdinger-Westphal-Kern), der reflektorischen Sakkaden (colliculi superiores) sowie zur Synchronisierung der zirkadianen Tag-Nacht-Rhythmik (nucleus suprachiasmaticus) der Melatoninausschüttung aus der Zirbeldrüse. Sie enthalten Melanopsin, mit einem Absorptionsmaximum im blauen Bereich bei 480 nm. Horizontalzellen und Amakrinzellen bilden zwei lateral wirkende Interneuronsysteme, die hemmende Einflüsse auf die Signalverarbeitung ausüben können. Die Horizontalzellen sind SINNE 8 besonders wichtig für die Adaptation der Lichtintensität. Die Amakrinzellen bewirken u.a. die Umschaltung vom photopischem zum skotopischem Sehen. Die Müllerzellen wirken als Gliazellen der Retina – sie durchziehen alle retinalen Schichten, regulieren den Metabolismus der anderen Retinazellen, sind gleichzeitig deren Stützzellen mit geschmeidiger Stoßdämpferwirkung und transretinale Lichtleiter zu den Außengliedern der Sensorzellen. Dabei wirken sie ähnlich einem Glasfaserkabel, indem sie das Licht von der vorderen Netzhautoberfläche zu den Außengliedern der Sensorzellen lenken. Das ist besonders bei geringer Lichtintensität (Nachtsehen) von Bedeutung. Die Neurone des n. opticus ziehen zum chiasma opticum, wo nur die nasalen Fasern kontralateral kreuzen. Im tractus opticus projizieren ipsi- u. kontra-laterale Fasern zum corpus geniculatum laterale (CGL). Zuvor ziehen einige koniozelluläre Fasern zur reflektorischen Steuerung der Pupillo-Motorik zu den colliculi superiores und der BlickMotorik zur area praetectalis. Im CGL findet die Umschaltung auf das 4. Neuron statt. Dessen Fasern ziehen in der radiatio optica weiter zum primären visuellen Cortex (V1 = Brodman-Areal 17). C. Physiologische Grundlagen 1. Gesamtbrechkraft, Achsenametropie, Refraktionsanomalie Als dioptrischer Apparat ist das Auge ein zusammengesetztes optisches System. Entsprechend ihrer Dichte brechen Cornea, Kammerwasser, Linse, Glaskörper das durch sie durchtretende Licht. Dabei hat das Auge im fern-akkommodierten Zustand eine Gesamtbrechkraft von etwa 59 dpt. Nicht variabel ist die Brechkraft der Cornea von ca. 43 dpt. Die Brechkraft der Corneavorderfläche von Luft auf Stroma beträgt D = +48 dpt sie wrid aber durch die Brechkraft der Corneahinterfläche zum Kammerwasser um –5 dpt reduziert. An der Linse hingegen kann sich die Brechkraft ändern: bei Fern-Akkommodation ist die Brechkraft D = 19 dpt; bei Nah-Akkommodation je nach Linsenkrümmung bis D = 33 dpt). Hinweis: durch diverse Abbildungsfehler reduziert sich die Gesamtbrechung von Coneal+ Linse um -3dpt. Bei Normalsichtigkeit (Emmetropie) liegt der Brennpunkt exakt auf der Retina. Dadurch entsteht in der Fovea centralis ein scharfes Bild. Der Fernpunkt liegt im Unendlichen ( ). Achsenametropie: (= Missverhältnis zwischen Bulbuslänge und Gesamtbrechkraft) 1) Bei Kurzsichtigkeit (Myopie) ist der Bulbus im Verhältnis zur Gesamtbrechkraft zu lang. Bei der Fernakkommodation liegt das scharfe Bild vor der Retina und wird daher unscharf auf der Retina abgebildet. Die Nahakkommodation funktioniert normal. Der Myope muss im Gegensatz zum Emmetropen die Brechkraft seiner Linse nicht so stark erhöhen, um einen gleichnahen Gegenstand scharf zu fokussieren. Der Fernpunkt liegt jedoch nicht im Unendlichen, sondern nimmt einen reellen endlichen Wert an (bei einer Myopie von +2 dpt sind das 0,5 m), wobei der Nahpunkt etwas näher an das Auge heranrückt. Zur Korrektur der für den längeren Bulbus relativ zu hohen Brechkraft werden konkave Zerstreuungslinsen (- dpt) verwendet, welche die Lichtstrahlen entsprechend spreizen und so Nah- u. Fernpunkt an die Norm (Emmetropie) angleichen, indem der Fokus auf die Retina fokussiert wird. 2) Bei Weitsichtigkeit (Hyperopie) ist der Bulbus zu kurz, bei gleicher Gesamtbrechkraft entsteht das scharfe Bild nicht auf, sondern hinter der Retina, d.h., der Fernpunkt ist virtuell. Beim Blick in die Ferne kann durch Nahakkommodation die Bildebene so weit nach vorn verlagert werden, dass sie auf der Retina liegt. Im Nahbereich reicht jedoch die Akkommodationsfähigkeit der Linse nicht mehr aus, um die Gegenstände scharf abzubilden. Hyperope Kinder fallen meist durch Einwärtsschielen auf, was durch Konvergenzbewegung beider Bulbi bei der Nahakkommodation verursacht wird. Der Nahpunkt liegt weiter entfernt vom SINNE 9 Auge als bei Emmetropie mit gleicher Akkommodationsbreite. Zur Korrektur werden konvexe Sammellinsen (+ dpt) eingesetzt, die den Brennpunkt der Lichtstrahlen durch deren „Stauchung“ auf die Retina zurückfokussieren: der Fernpunkt liegt nun wieder im Unendlichen und der Nahpunkt rückt näher ans Auge heran. Die Akkommodationsbreite in Dioptrien ist in beiden Fällen unverändert, nur der Akkommodationsbereich in Metern verschiebt sich in Relation zum Bulbus (siehe Abbildung unten). Die Alters(weit)sichtigkeit (Presbyopie) hingegen ist keine Achsen-Ametropie, sondern beruht auf einem Verlust der Elastizität der Linse, der durch Größenzunahme der Linse infolge von vermehrter Einlagerung proteinärer Kristalline sowie von Wasserverlust verursacht wird. Ist die Akkommodationsbreite von ursprünglich 14 auf 3 - 0 dpt abgesunken, spricht man von Presbyopie. aus: P. Deetjen, E.-J. Speckmann, J. Hescheler Physiologie (2005) Beim Astigmatismus weisen zwei Ebenen der Cornea oder der Linse unterschiedliche Krümmungsradien und damit unterschiedliche Brechkräfte auf (Refraktionsanomalie). Die Abbildung eines Punktes ist auf der Retina dann nicht mehr punkt- sondern strichförmig (Stabsichtigkeit). Stehen die Ebenen senkrecht zueinander, spricht man von regulärem Astigmatismus, Abweichungen zwischen horizontaler und vertikaler Ebene von bis zu 0,5 dpt sind noch physiologisch und werden durch den mechanischen Druck des Lides auf die Cornea verursacht. Dem irregulären Astigmatismus liegen komplexere Abweichungen zugrunde, wie z.B. Verletzungen oder angeborene Asymmetrien der Cornea. Astigmatische Augen werden mit zylindrisch eingeschliffenen Plus- oder Minus-Brillengläsern oder Kontaktlinsen korrigiert. Beim Katarakt (= Grauer Star = Linsentrübung) kann das Licht aufgrund von Ausfällungen im kristallinen Gefüge der Linse nicht mehr ordnungsgemäß gebrochen werden. Objekte werden dadurch zunehmend grau und verschwommen wahrgenommen, Farben wirken blass bis anders-farbig, oft sind die Patienten geblendet. Auch scheinen Enzymdefekte und UVStrahlung bei der Entstehung des Altersstars Cataracta senilis eine Rolle zu spielen. Angeborene Formen (Cataracta totalis u.a. nach intra-uteriner Rötel-Infektion) führen unbehandelt zur Erblindung. Therapeutisch wird die trübe Linse entfernt (= aphages Auge) und durch eine Kunststofflinse ersetzt oder durch eine „Star“-Brille nah-korrigiert. 2. Die Akkommodation (= Anpassung an die veränderte Entfernung eines Gegenstandes) Abhängig vom Krümmungsradius der Linse können nahe oder ferne Objekte in der Fovea scharf abgebildet werden: beim Blick in die Ferne (= Desakkommodation) ist der zirkuläre m. ciliaris entspannt und die Sklera zieht ihn nach außen. Die Zonulafasern sind gespannt. SINNE 10 Der Zug an der passiven Linse bewirkt deren Abflachung; wodurch sich ihre Krümmungsradien vergrößern und ihre Brechkraft abnimmt (33 dpt 19 dpt). Bei Kontraktion der m. recti laterales bewegen sich beide Augen auseinander (Divergenz). Die Nah-Akkommodation dagegen ist ein aktiver Vorgang: durch Kontraktion des parasympathisch innervierten m. ciliaris werden die Zonula-Fasern entspannt und die Linse kann sich aufgrund ihrer Eigenelastizität „abkugeln“, wodurch sich ihre Krümmungsradien verkleinern und die Brechkraft ansteigt (19 dpt 33 dpt) und der Brechungsindex sich von von 1,414 auf 1,424 (also um 0,01) erhöht. Zusätzlich wird die Pupille zu Erhöhung der Tiefenschärfe reflektorisch verengt und beide Augen führen Konvergenz-Bewegungen (Kontraktion der Mm. recti mediales) aus, um in der fovea centralis die gleichen Bilder zu fixieren. Der Nahpunkt ist die kürzeste Entfernung (individuell zwischen 0,07-0,3 m), bei der ein Gegenstand gerade noch scharf abgebildet werden kann. Er entfernt sich mit zunehmendem Alter vom Auge in Richtung Fernpunkt (Presbyopie). Der Fernpunkt ist die Entfernung, bei der bei Fern-Akkommodation gerade noch scharf gesehen wird. Beim emmetropen Normalsichtigen liegt dieser Punkt im Unendlichen. Nah- u. Fernpunkt stellen die Grenzen des Akkommodationsbereichs in Metern dar. Die Akkommodations-Breite (AB) ist die Differenz der Brechkraft bei Einstellung von Nahu. Fernpunkt (AB = 1/NP-1/FP) und wird in Dioptrie angegeben. Sie beträgt im Kindesalter etwa 14 dpt und kann im Alter auf Null abnehmen (Presbyopie = durch Verlust der Elastizität der Linse). Siehe Praktikum-Versuch Nr. 1 Als Pseudo-Akkommodation wird die Fähigkeit bezeichnet, sowohl Gegenstände in der Ferne als auch in der Nähe ohne aktive Brechkraftänderung des Auges hinreichend scharf erkennen zu können. 3. Der Transduktionsprozess (chemische und elektrische Veränderungen in den Photosensoren bei Belichtungsänderung) Die Photosensoren haben ein Außen- und ein Innensegment. Im Außensegment befinden sich bei Stäbchen etwa 1.000 Membran-Scheibchen (Disks) und bei Zapfen MembranEinfaltungen, die die photonen-perzipierende Membranoberfläche vergrößern. In beiden sind die Sehfarbstoffe enthalten: in den Stäbchen das Rhodopsin - bestehend aus dem transmembranären Glycoprotein Opsin und dem lichtempfindlichen Chromophor 11-cisRetinal. Die drei verschiedenen Zapfentypen enthalten das „Zapf- oder Iod-Opsin“, die sich in der Aminosäuresequenz ihres Opsins unterscheiden und dadurch verschiedene spezifische Absorptionsmaxima haben: 420 nm für Blau, 535 nm für Grün und 565 nm für Rot. Alle Zapfopsine enthalten das gleiche 11-cis-Retinal wie Stäbchenrodopsin. Pigment-Epithelzellen regenerieren das anfallende all-trans-Retinal über Retinol zur aktiven isomeren 11-cis-Form. Bei Dunkelheit (siehe Abbildung) liegt das Ruhemembranpotenzial der Photosensoren bei ca. –30 mV. Im Wesentlichen wird diese Dauerdepolarisation durch erhöhte Na+-Leitfähigkeit von cGMP-abhängigen Na+Kanälen verursacht. Dabei wirkt das durch konstitutiv aktiv wirkende Guanylyl(at)zyklase produzierte cGMP als second messenger, wodurch sich in der Zellmembran unspezifische, cGMP-abhängige Kationenkanäle öffnen, aus: R. Klinke, H.-C. Pape, S. Silbernagl Physiologie (2005) also viel Na+ und etwas Ca2+ SINNE 11 einströmen = Dunkelstrom. Das in die Zelle eindringende Ca unterbricht über ein Modulatorprotein die Arbeit der Guanylatzyklase, hemmt somit die Bildung von cGMP. „Überschüssiges“ Ca2+ wird dann wieder über 1Na+/3Ca2+-Antiporter nach extra-zellulär transportiert. Durch den Einstrom von Kationen (= Anstieg positiver Ladungen) wird in der negativ geladenen Zelle eine Membrandepolarisation erzeugt, die sich als graduiertes Rezeptorpotential vom Außen- zum Innenglied der Photosensorzelle fortpflanzt. In der Nähe der Präsynapse werden dadurch mehr spannungsgesteuerte Calciumkanäle aktiviert, was einen Anstieg der intrazellulären Ca2+-Konzentration auslöst. Infolgedessen erhöht sich die Transmitterfreisetzung von Glutamat. 2+ Bei Belichtung kommt es zur Stereo-Isomerisierung des 11-cis-Retinals in die all-trans-Form. Das Rhodopsin ändert dabei seine Konformation über mehrere Zwischenstufen in das aktivierte Metarhodopsin II. Dieses setzt eine Signalkaskade in Gang, die zu einer SignalVerstärkung um den Faktor 106 führt. Zusammen mit GTP aktiviert es das heterotrimere GProtein Transducin. Dessen α-Untereinheit aktiviert Phosphodiesterase-5, die cGMP zu 5´GMP hydrolysiert. Die cGMP-abhängigen Kationenkanäle schließen sich infolgedessen vermehrt und weniger Natriumionen (weniger positive Ladungen) können einströmen. Dadurch kommt es zur Hyperpolarisation bis auf etwa –70 mV (bei max. Lichteinfall); wobei die Ca2+ -Konzentration in der Zelle infolge der Arbeit der Na+-Ca2+-Antiporter absinkt und die Transmitterfreisetzung (Exozytose von Glutamat) abnimmt. Der Effekt des von der Photorezeptorzelle abgegebenen Transmitters Glutamat auf die nachgeschalteten Bipolarzellen hängt von deren Typ ab. Es gibt ON- und OFF-Bipolarzellen: OFF-Bipolarzellen haben ionotrope Glutamatrezeptoren (= Kanäle) und werden daher durch viel Glutamat aktiviert. Bei Belichtung scheidet die Photorezeptorzellen aber nur wenig Glutamat aus und es folgt eine Hyperpolarisation der nachgeschalteten OFF-Bipolarzelle. An den Ganglienzellen bilden sich ein geringes EPSP, wodurch sich die AP-Frequenz verringert. ON–Bipolarzellen werden dagegen bei Dunkelheit durch viel Glutamat von der Photorezeptorzelle gehemmt und durch wenig Glutamat aktiviert. Unter dem Einfluss von viel Glutamat wird über ihre Gi-gekoppelten metabotropen Glutamat-Rezeptoren eine Phosphodiesterase aktiviert. Diese hydrolysiert das von einer konstitutiv aktiven Guanylatzyklase produzierte zyklische Nukleotid cGMP, wodurch sich infolge dieses weniger zur Verfügung stehenden cGMPs mehr cGMP-abhängige Na+-Kanäle der ON-Bipolarzelle verschließen, d. h. weniger Kationen (Na+) in ihr Zellinneres gelangen. Die Bipolarzelle ändert deshalb ihr Membranpotenzial - sie hyperpolarisert auf etwa –70 mV. Dies bewirkt eine geringere Freisetzung von Glutamat an der Präsynapse zu der ihrer Nachgeschalteten Ganglienzelle. Die Frequenz der APs wird geringer. Bei Belichtung der Photorezeptorzelle wird weniger Glutamat freigesetzt. Daher entfällt der hemmende Effekt der Gi-gekoppelten metabotropen Rezeptoren. Der cGMP-Spiegel steigt und mehr cGMP-abhängigen Na+-Kanäle öffnen sich. Sukzessive wird die Bipolarzelle depolarisiert, mehr spannungsabhängige Calciumkanäle öffnen und lassen Ca2+ in die Präsynapse einströmen, was zur vermehrten Glutamat-Freisetzung in den synaptischen Spalt zur ON-Ganglienzelle führt. In den Nachgeschalteten Ganglienzellen entstehen mehr EPSPs und schließlich am Axonhügel eine erhöhte AP-Frequenz. 4. Die Adaptation (= Anpassung an veränderte Licht-Reiz-Amplituden) Die Hell-Adaptation dauert nur wenige Sekunden; wobei über den musculus sphincter pupillae die Pupille reflektorisch verengt wird (Miosis). Bei > 107 Lux ist der Unterschied in der Leuchtdichte zu groß – Folge ist eine vorübergehende Blendung (evt. Schmerz). SINNE 12 Zur Dunkel-Adaptation werden hingegen längere Zeiträume benötigt. Dabei zunächst die Pupille vom musculus dilatator pupillae erweitert (Mydriasis). Nach 8-10 min. wird im Kohlrausch-Knick retinal vom photopischen auf das skotopische Sehen umgeschaltet. Die wesentlich lichtempfindlicheren Stäbchen erreichen ihre maximale Lichtem-pfindlichkeit erst nach 180 min. Bei unveränderter Gesamtgröße der primärphotopisch---- ------------------------------------- skotopisch rezeptiven Felder vergrößern sich dabei aus: R. Klinke, H.-C. Pape, S. Silbernagl Physiologie (2005 deren Zentren, wodurch sich die räumliche Summation erhöht, während sich Auflösungsvermögen und Kontrast-Wahrnehmung verschlechtern. Gemeinsam haben alle drei Zapfentypen ein mittleres Absorptionsmaximum bei 555 nm, die Stäbchen jedoch bei 500 nm (Purkinje-Verschiebung: Blaues Licht reflektierende Flächen werden dadurch als heller und Rot reflektierende Flächen wahrgenommen). Bei Dunkeladaptation, d. h. nach Umschaltung auf Stäbchensehen, wird die nur Zapfen besitzende Foveola zum blinden Fleck (Skotom). Die Sehschärfe ist dann im parafovealen Bereich mit der höchsten Stäbchendichte am größten. Während der Dunkelphase werden die Photosensorzellen über verschiedene biochemische Regenerationsprozesse auf die erneute Verarbeitung von Photonen vorbereitet. Zudem hyperpolarisieren Stäbchen langsamer als Zapfen. Siehe auch „primär-Rezeptives Feld“. Folge: unvollständige, unscharfe, langsamere Wahrnehmung („Nachts sind alle Katzen grau!“) Nachtblinden Menschen (Nyktalopie = Nachtsichtigkeit; med. Hemeralopie = Tagessichtigkeit) steht für die Dunkeladaptation nur ihr Zapfensystem zur Verfügung. Die ersten 8-10 Min unterscheiden sich nicht von normaler Adaptation. Eine weitere Adaptation ist dann aber aufgrund der insuffizienten Stäbchen nicht mehr möglich, z.B. durch Gendefekte oder Vitamin A-Mangel. Messung mittels Adaptometer. Stäbchen-monochromate Menschen besitzen keine funktionstüchtigen Zapfen. Bei Tageslicht werden sie geblendet. Die Dunkeladaptation läuft aber schneller ab, da der hemmende Einfluss des Zapfensystems auf das ausschließlich verwendete Stäbchensystem fehlt. 5. Nystagmus, Okulogramm, Evoziertes Potenzial Der optokinetische Nystagmus (OKN) ist eine reflektorische Augenbewegung und wird z.B. durch Blickfixierung aus einem fahrenden Zug oder auf eine sich drehende Streifentrommel ausgelöst. Die großflächige Bewegung des Bildes auf der Retina heißt optischer Fluss. Dieser löst entgegen der eigenen Bewegungsrichtung langsam folgende Kompensationsbewegungen (Deviation) der Bulbi aus. Schnelle Rückstellbewegungen (Sakkaden) in Bewegungsrichtung unterbrechen diese Deviation. Die Sakkade gibt die Richtung des Nystagmus an und wird durch eine erhöhte AP-Frequenz bestimmt. Augenbewegungen sind mittels Elektro-Okulo-Gramm (EOG) objektiv darstellbar. Zwischen Pigmentepithel (positiv geladen) und Photosensoren (negativ) besteht bei Belichtung eine Potenzialdifferenz = belichtungs-abhängiges corneo-retinales Bestandspotenzial. Elektroden an Schläfe und Nase registrieren Bewegungen dieses okularen Dipols. Visuell Evozierte Potenziale (VEP) helfen bei Beurteilung der Sehbahnfunktion: dazu wird über den visuellen Kortex okzipital ein spezielles EEG abgeleitet und gemittelt, währenddessen dem Patienten intermittierende Lichtreize präsentiert werden. Der typische Verlauf zeigt ein mehrgipfliges Potenzial mit einer ausgeprägten positiven P100-Welle (Latenz 90-120 ms). Pathologisch sind erhöhte Latenzzeiten, erniedrigte Amplituden oder signifikante Latenzdifferenzen zwischen beiden Augen. Eine Latenzverlängerung der P100-Welle auf >120 SINNE 13 ms bei gleichzeitiger Abnahme der Amplitude ist beispielsweise bei Entzündungen des nervus opticus oder Multipler Sklerose zu verzeichnen. 6. Das primär-Rezeptive Feld (pRF) Durch Verschaltung von retinalen Zellen entsteht ein primäres Rezeptives Feld: dieses wird von Gruppen solcher Sensorzellen gebildet, die die Aktivität eines gemeinsamen afferenten Neurons beeinflussen. Die pRFs sind in der Fovea centralis sehr klein und werden zur Peripherie (= Ora serrata) hin zunehmend größer. Sie sind rund, überlagern sich nicht und untergliedern sich in ein dominantes Zentrum und eine antagonistische Peripherie. Bei ON-Zentrum-Bipolaren wirkt eine Belichtung des Zentrums erregend, die Belichtung der Peripherie hingegen hemmend. Umgekehrt hemmt Licht das Zentrum von OFF-ZentrumBipolaren und erregt die Peripherie. Beachte Bei Adaptationsvorgängen ändert sich lediglich die Größe des Zentrums, nicht aber die Größe des gesamten Feldes! Bei maximaler Dunkeladaptation können die Zentren die Größe eines gesamten Feldes erreichen; während sie sich bei Helladaptation verkleinern. Horizontalzellen vermitteln eine Rückkopplung zwischen Photorezeptorzellen. Sie dienen der Einstellung des Helligkeitsbereichs. Amakrine Zellen sind sehr funktionsvariabel. Sie haben verschiedene Transmitter und wirken auf verschiedene post-synaptische Rezeptoren der Ganglienzellen. Sie sind wichtig für Bewegungssehen, für intermittierende Reize, für Verschaltung der pRFs mit den ON/OFFGanglienzellen. Funktion: Die primäres Rezeptives Feld bewirken mittels lateraler Hemmung der Interneurone eine Kontrastverstärkung zwischen den hellen und dunklen Strukturen, so z. B. der Schwarz-Weiß-Übergang eines Buchstaben oder am Rand einer Struktur) sowie zwischen verschiedenfarbigen Strukturen (= ZweipunktDiskriminierung). Im Dunkeln ist der Antagonismus aufgehoben folglich sinkt der Visus (Unschärfe durch räumaus: R. F. Schmidt , G. Thews, F. Lang Physiologie d. Menschen (2000) liche Summation). Und durch verlängerte zeitliche Summation der Photonen und der Aktivierung des rezeptiven Feldes nehmen wir Objekte um bis zu 30 % langsamer bzw. verzögert wahr. Hinzu kommt, dass Stäbchen langsamer hyperpolarisieren als Zapfen. Simultankontrast oder Sukzessivkontrast (letzterer auch als Nachbilder bekannt) beruhen auf einer komplexen lokalen (in der Fovea), sensorisch-photochemischen (Isomerisierung des Retinals = zeitliche Sättigung der Empfindlichkeit der Zelle) sowie neuronalen (Singleopponent Cells = Gegenfarben-Neurone) Adaptation. SINNE 14 7. Das Gesichtsfeld Das Gesichtsfeld ist der Bereich der Umwelt, der mit dem unbewegten Auge wahrgenommen wird. Mit einem Perimeter kann es für beide Augen getrennt bestimmt werden. Bei der Untersuchung wird das nicht untersuchte Auge abgedeckt und mit dem zu untersuchenden Auge ein Punkt in der Mitte des Perimeters fixiert = Mittelpunkt des Gesichtsfeldes (0°). Zur Bestimmung der Gesichtsfeldgrenzen werden von der Peripherie her Lichtpunkte ins Gesichtsfeld hineinbewegt (kinetische Prüfung). Sobald der Lichtreiz wahrgenommen wird, ist die Grenze erreicht. Bei einer statischen Perimetrie leuchten Lichtpunkte an verschiedenen Stellen nacheinander auf. Die Ausdehnung des Gesichtsfeldes reicht ca. 90° nach temporal, 60° nach nasal, 60° nach oben und 70° nach unten. Für weißes Licht ist es am größten und entsprechend der Verteilung der für die 3 Grundfarbenkodierenden Zapfentypen für Blau größer als fürRot sowie am kleinsten für aus: R. Klinke, H.-C. Pape, S. Silbernagl Physiologie (2005) Grün.Auch innerhalb des Gesichtsfeldes können Bereiche auftreten, in denen Lichtreize nicht wahrgenommen werden. Einen solchen Gesichtsfeldausfall bezeichnet man als Skotom. Ein physiologisches Skotom ist z.B. der blinde Fleck (Papilla nervi optici) ca. 15° temporal des Fixationspunktes. Er repräsentiert den sensorfreien Bereich über dem Abgang des n. opticus Bei Dunkelheit wird die fovea centralis zum physiologischen Skotom. Durch Kenntnis des. Sehbahnverlaufs kann bei auftretenden Skotomen auf die Lokalisation der Schädigung geschlossen werden. Besteht ein monokularer Gesichtsfeldausfall, so liegt dieser vor der Sehnervenkreuzung und betrifft die Retina oder den Sehnerven selbst (z.B. vollständige Erblindung eines Auges = Amaurose = Schwarzer Star = Pupillenstarre nach einseitigem Ausfall des n. opticus). Werden die nasalen Fasern im Bereich des Chiasmas geschädigt, tritt eine bitemporale Hemianopsie (ScheuklappenBlindheit) auf, da die Fasern nur die temporalen Gesichtsfeldhälften repräsentieren (z.B. bei Hypophysen-Tumoren). Seltener werden die temporalen Fasern geschädigt, was zur binasalen Hemianopsie führt. Bei Schädigungen hinter dem Chiasma opticum treten homonyme Gesichtsfeldausfälle auf. Diese betreffen dann jeweils nur die rechten oder linken Teile der Gesichtsfelder. Bei Schädigung in der linken Hemisphäre ist die rechte Gesichtsfeldhälfte betroffen. Skotome, die im Rahmen einer Schädigung der Sehnerven durch erhöhten Augeninnendruck (Glaukom) entstehen, liegen anfänglich in der mittleren Peripherie und werden deshalb oft nicht wahrgenommen. 8. Der Augeninnendruck (Intra-Okular-Druck) Der normale Augeninnendruck beträgt ungefähr 10-20 mmHg. Abhängig von der Tageszeit schwankt dieser Wert um etwa 5 mmHg, wobei er morgens niedriger als abends ist. Die Produktion und der Abfluss des Kammerwassers gleichen sich unter normalen Umständen aus (2 mm3/min). Das Kammerwasser wird aus dem uveal-versorgten (peripherer Blutdruck ca. 40 mmHg) Epithel des Ziliarkörpers gebildet und stellt ein Ultrafiltrat und Sekretionsprodukt dar (getrieben durch die Carboanhydrase). Von der hinteren Augenkammer fließt es durch die Pupille in die vordere Kammer, um im Kammerwinkel über das Trabekelwerk in den Schlemm-Kanal resorbiert zu werden und schließlich in den venösen Abfluss (8-9 mmHg) zu gelangen. Durch den intraokularen Druck -- der sich durch den gelartigen Glaskörper homogen verteilt - - wird die Form des Bulbus konstant gehalten. Zur Messung des Drucks stehen verschiedene Tonometrie-Verfahren zur Verfügung, die alle die Verformbarkeit der Cornea in Abhängigkeit vom herrschenden Augeninnendruck und der SINNE 15 Dicke der Cornea nutzen. Bei der Applanations-Tonometrie wird die Kraft gemessen, die zum Abflachen einer definierten Corneafläche benötigt wird, während bei der ImpressionsTonometrie deren Eindellung infolge der Einwirkung eines definierten Druckes gemessen wird. Für beide Verfahren ist Anästhesie und direkter Kontakt zur Cornea erforderlich. Umgangen wird dieses Problem durch die Non-contact = Pneumo-Tonometrie, wozu ein Luftstoß verwendet wird. Zum Erhalt verlässlicher Ergebnisse (Lidschlussreflex, Schreckreaktionen) sollte diese Messung des Drucks jedoch mehrmals wiederholt werden. Bei pathologisch erhöhtem Augeninnendruck liegt ein Glaukom (Grüner Star) vor. Ursache ist entweder eine Behinderung der Resorption des Kammerwassers oder dessen gesteigerte Produktion. Der erhöhte Druck bewirkt eine Schädigung von Sehnervenfasern und Ischämie, was zu Gesichtsfeldausfällen und unbehandelt später zur Erblindung führt. Wenn der Kammerwinkel durch die Iris verlegt wird (akutes Winkelblock-Glaukom), ist der Kammerwasserabfluss behindert und es kommt zum plötzlichen Druckanstieg mit schmerzhafter Asthenopie. Zur Therapie werden Miotika getropft, um den Abfluss zu verbessern, sowie Medikamente, die die Produktion des Kammerwassers hemmen (β-Blocker, Carboanhydratase-Blocker wie Acetazolamid). Zusätzlich kommt eine operative Entlastung in Betracht. Das chronische Offenwinkelglaukom (Glaucoma simplex) verläuft oft schmerzlos und schleichend. Trotz offenem Kammerwinkel ist der Abfluss stetig behindert und erzeugt so einen chronisch erhöhten Augeninnendruck. Therapiert wird mit drucksenkenden Medikamenten und gegebenenfalls wird der Abfluss des Kammerwassers operativ verbessert. 9. Die Sehschärfe (Visus) Der Visus V ist als der Kehrwert des in Winkelminuten gemessenen räumlichen Auflösungsvermögens α (Visus = 1/α [1/Winkelminute]) definiert, also die Fähigkeit, noch zwei Punkte als getrennt erkennen zu.können. Ein Auflösungsvermögen von 1 Winkelminute bedeutet, dass eine Lücke mit einer Größe von 1 Winkelminute (Sehwinkel) in einer Optotype (Landoldt-Ring, Buchstabe) noch erkannt wird. In diesem Fall beträgt der Visus 1 (= 100 %). Werden auch Abstände unter einer Winkelminute erkannt, ist V >1 (= Übervisus). Sind die Abstände größer als eine Winkelminute, ist V <1. Anstelle des Sehwinkels kann auch die Entfernung bestimmt werden, in der eine Lücke bestimmter Größe noch erkannt wird (V = Messentfernung/ Sollentfernung bei V = 1 oder 100 %). Der Visus von Jugendlichen beträgt 0,8-1,5. Beim photopischen Sehen ist die Sehschärfe in der Fovea centralis am größten und nimmt mit der Zapfendichte zur Peripherie hin ab. Mit abnehmender Helligkeit verschlechtert sich der Visus. Bei skotopischen Bedingungen erreicht er para-foveal die höchsten Werte. Da sich in der Fovea centralis keine Stäbchensensorzellen befinden; ist dort der Visus Null. Zur Bestimmung werden bei standardisierter Beleuchtungs-Intensität sogenannte Sehprobentafeln für die Nähe mit Optotypen (Buchstaben, Landoldtringe, Bilder bekannter Gegenstände) verschiedener Größen in z.B. 5 m Entfernung projiziert. Unser Sehvermögen ist von allen strukturellen, opto-physikalischen, sensor-physiologischen, psycho-physikalischen, neuro-anatomischen Parametern der beiden Sinnesorgane und der Sehbahn abhängig! Siehe Praktikum-Versuche Nr. 2 + 5. Weitere klinische Aspekte werden im Klinisch-Physiologischen Seminar behandelt. 10. Das Farbensehen Für die Farbwahrnehmung spielen Farbton u.-Sättigung (= Farbart) und Helligkeit eine Rolle. Durch Kombination dieser Komponenten sind wir in der Lage, über 2 Millionen Farbvariationen wahrzunehmen. Die 3 Zapfentypen unterscheiden sich im Absorptionsmaximum ihrer Sehfarbstoffe in Zapfen für Blau (420 nm), Grün (535 nm) und Rot (565 nm). Nach der trichromatischen Farbtheorie kann durch additive Farbmischung dreier monochromatischer Farben jede beliebige Farbe erzeugt oder die Farbe aufgehoben werden (Weiß). Diese Mischung entsteht durch neuronale Verarbeitung der Antworten der drei Zapfentypen SINNE 16 auf einen Lichtreiz. Entlang der Sehbahn werden die Zapfensignale so verschaltet, dass Gegenfarben-Neurone (Ganglienzellen Neurone im CGL) entstehen. Komplementärpaare sind Rot + Grün, Blau + Gelb (Gelb wird aus Grün und Rot gemischt). Solche Neurone werden im Zentrum des pRFs durch einen Zapfentyp angeregt und aus der Peripherie durch Zapfen der Gegenfarbe antagonistisch gehemmt (ON-Zentrum für Rot, Grün oder Blau). Bei OFF-Zentrum-Ganglienzellen ist die Farbe im Zentrum hemmend, die Gegenfarbe in der Peripherie hingegen erregend. Diese Neurone sind farbempfindlich, aber noch nicht farbspezifisch, denn sie reagieren auf Farb- und Helligkeitsunterschiede ähnlich. DoppelGegenfarben-Neurone befinden sich im visuellen Kortex und sind farbspezifisch. Hier sind nicht nur die Gegenfarben, sondern auch Zentrum und Peripherie antagonistisch verschaltet. Sie entstehen durch Zusammenschalten von ON- und OFF-Gegenfarbenneuronen eines Gegenfarbenpaares. Dadurch wirkt eine Farbe im Zentrum erregend und die zugehörige Gegenfarbe hemmend. In der Peripherie ist es umgekehrt: die Komplementärfarbe wirkt erregend und die andere Farbe hemmend. So werden Farbkontraste zwischen den betreffenden Farben registriert. Farbsinnesstörungen beeinträchtigen die Farbwahrnehmung. Sie werden durch meist X-chromosomal vererbbare Mutationen im Sehpigment verursacht. Wenn ein einzelner Zapfentyp in seiner Funktion reduziert ist, dann liegt eine Farbanomalie (= Farbschwäche) vor. So kann der rot-schwache Mensch (mit Protanomalie) nicht so viele Rottöne wie ein Gesunder unterscheiden und benötigt zum Mischen eines Gelbtons aus Grün und Rot im Anomaloskop einen höheren Rotanteil. Weiterhin existieren Grünschwäche (Deuteranomalie) und Blau-Gelb-Schwäche (Tritanomalie). Fehlt ein Pigment völlig, kann eine Farbe gar nicht erkannt werden (= Farbblindheit). Protanope sind rot-blind, Deuteranope grün-blind und Tritanope blauviolett-blind. Beim Fehlen aller Zapfenpigmente liegt eine totale Farbenblindheit (Achromatopsie) vor. Nur noch anhand der Helligkeit können etwa 500 Graustufen unterschieden werden. Zur Testung auf Farbsinnesstörungen werden neben dem Anomaloskop auch pseudo-isochromatische Tafeln eingesetzt. Sie bestehen aus Punkten unterschiedlicher Farbart, aber gleicher Helligkeit. Dabei sind versteckte Zahlen oder Muster nur für Menschen ohne Farbsinnesstörungen vollständig erkennbar. 11. Die Tiefen-Wahrnehmung (= mono- und bin-okulares räumliches Sehen) 1) Die monokulare Tiefenwahrnehmung erfolgt bei Objekten, die weit entfernt sind, oder wenn nur ein Auge zur Verfügung steht. Bei Verdeckung eines Objekts durch ein anderes befindet sich logischerweise das verdeckte hinter dem anderen. Bewegt man sich, so scheinen sich nahe Gegenstände schneller als weiter entfernt liegende zu bewegen (Bewegungs-Parallaxe). Durch Vergleich von Objekten bekannter Größe kann die Entfernung abgeschätzt werden. Parallele Linien scheinen sich in der Ferne zu schneiden. Gleichgroße Objekte erscheinen je nach Entfernung verschieden groß (Größen-perspektive). Verteilung von Licht und Schatten sowie Farbsättigung sind auch hilfreich. Nahe binokulare Tiefenwahrnehmung Objekte haben gesättigte Farben, ferne aus: R. Klinke, H.-C. Pape, S. Silbernagl Physiologie 2004 erscheinen bläulich. 2) Die binokulare Tiefenwahrnehmung funktioniert bis zu einer Entfernung von 100 m (optimal jedoch zwischen Greifraum und dem 6 m Panum-Raum). Im Nahbereich stimmen SINNE 17 die Abbildungen in beiden Augen nicht völlig überein. Wird ein Punkt F (siehe obige Abbildung) fixiert, wird er in jedem Auge in der Fovea abgebildet. Netzhaut-Areale, auf denen jeweils gleiche Bildpunkte abgebildet werden, heißen korrespondierende Netzhautareale. Außer dem Fixationspunkt F werden also noch andere Punkte auf korrespondierenden Stellen abgebildet. Sie liegen alle auf einer geometrischen Figur, dem Horopter. Liegt ein Punkt nicht auf dem Horopter, verschiebt sich die Abbildung auf der Retina in der Horizontalen (Querdisparation). Liegt ein Gegenstand weiter entfernt als F, so weicht seine Abbildung nach nasal ab. Weicht die Abbildung nach temporal ab, dann liegt das entsprechende Objekt näher als der Fixationspunkt. Weichen die fovealen Abbildungen beider Augen zu stark voneinander ab (Querdisparation mehr als 16 Winkelminuten), können die beiden Bilder nicht mehr zu einem Bild verschmolzen werden (Fusion): man sieht folglich Doppelbilder. Beim Schielen (Strabismus) wird der Gegenstand nur von einem Auge fixiert, während das andere abweicht. Wenn dies bei Kindern nicht rechtzeitig durch Behebung der Ursache (Hyperopie, Fehlstellung) und Fixierungsübungen mit dem abweichenden Auge korrigiert wird, kann es zur Ausbildung eines dominanten Auges kommen. Die Sinneseindrücke des anderen Auges werden unterdrückt und es entwickelt sich eine irreversible Sehschwäche (Amblyopie). PRAKTIKUMVersuche AUG Versuch 1 E + SEHEN Bestimmung von NAH- u. FERN-PUNKT Mit dem Optometer nach DONDERS werden Nahpunkt (NP) und Fernpunkt (FP) des Auges subjektiv ermittelt. Auf einem 0,5-1 m langen Messlineal befindet sich eine verschiebbare, von seitlich oder oben beleuchtete Nadel, welche mit dem Auge der Versuchperson durch die zwei kleinen Löchern der Lochblende zu fixieren ist. Messung des Nahpunktes NP: Die Nadel wird durch Verschieben dem Auge genau so weit angenähert, bis sie nicht mehr einfach, sondern doppelt und unscharf erscheint. Die kürzeste Entfernung für noch einfaches Sehen der Nadel ergibt den Nahpunkt, der am Lineal direkt abzulesen ist. Berechnung des Fernpunktes FP: Der Fernpunkt liegt beim normalsichtigen (= emmetropen) Auge in unendlicher Entfernung, beim kurzsichtigen (= myopen) Auge in endlicher Entfernung vor dem Auge, beim weitsichtigen (= hyperopen ) Auge ist er virtuell (F liegt hinter dem Auge). Der Fernpunkt ist demnach mit obiger Apparatur allein, nicht zu messen. Den Fernpunkt FP ermittelt man, indem hinter die Blende zusätzlich eine Sammellinse z.B. einer Brechkraft D = +3 dpt gesteckt wird. Dadurch wird das Auge künstlich myop gemacht und der Fernpunkt in endliche Entfernung gerückt. Nun wird die größte Entfernung bestimmt,, in der die Nadel noch einfach und scharf erscheint, indem man die Nadel vom Auge weg bewegt. Dies müsste beim emmetropen Auge der Fall sein, wenn sich die Nadel im Abstand der Brennweite der Zusatzlinse befindet, also in 20 cm Abstand. Dann sind die Strahlen nach der Brechung durch die Zusatzlinse parallel, als ob sie aus unendlicher Entfernung kämen. Ein hyperopes Auge wird die Nadel erst in mehr als 20 cm Entfernung einfach sehen. Aus diesen abgelesenen Werten wird schließlich die Lage des tatsächlichen Fernpunktes mittels folgender Formel errechnet: 1 1 ---- = ---- + D FP` FP oder für den Nahpunkt 1 1 ---- = ---NP` NP + D wobei FP`= der mit der vorgesetzten Linse gemessene Fernpunkt, FP = der gesuchte Fernpunkt Ihres Auges, D = die Brechkraft der vorgesetzten Linse ist. Aus den gemessenen SINNE 18 Entfernungen ergibt sich die Änderung der Brechkraft, welche die Akkommodation herbeiführen kann (= die Akkommodationsbreite in Dioptrien). Tragen Sie die VersuchsErgebnisse des getesteten Auges in die Tabelle ein: NP = Lage des Nahpunktes; Fp' = Lage des Fernpunktes nach Vorschalten von +3 dpt. Fp = Lage des Fernpunktes ( nach Abzug der +3 dpt). *NP’ NP 1/NP FP’ 1/FP' 1/FP Akkommodations- (m) (m) (dpt) (m) (dpt) (dpt) Breite (dpt) rechts links * bei tragen von Kontaktlinsen. Beim nicht korrigierten Auge ist Np’ = Np Berechnen Sie die Akkommodationsfähigkeit Ihres getesteten Auges mittels der Formel: Akkommodationsbereich (in m) (oder –breite in dpt) Versuch 2 AB = 1 1 ---- – ---NP FP Bestimmung der SEHSCHÄRFE a) Der Patient wird der genormt gut beleuchteten Tafel in genau 5 m Entfernung gegenüber gestellt. Ein Auge wird durch eine undurchsichtige Scheibe im Testbrillengestell verdeckt (wenn das Auge mit der Hand verdeckt wird, darf kein Druck auf den Bulbus ausgeübt werden, warum?). Der Patient liest nun die Optotypenreihe, die er noch gut erkennen kann. Das Ergebnis wird in Form eines Bruches protokolliert: Entfernung, in der die Prüfung vorgenommen wird im Zähler = IST, die auf der Tafel rechts angegebene Zahl der noch korrekt gelesenen Reihe im Nenner = SOLL. Also: wenn aus 5 m Entfernung die Reihe gut scharf gesehen wird, die der Normalsehende noch aus 8 m Entfernung lesen kann, beträgt die Sehschärfe = 5/8. Der Normalsehende hat meist die Sehschärfe > 5/5 oder > 100 % , wobei maximale Werte bis zu 200 % erreicht werden können. Der Wert 5/8 würde eine unternormale, der Wert 5/4 eine übernormale Sehschärfe bedeuten. Der Bruch wird nicht gekürzt, um aus ihm stets die Untersuchungsentfernung entnehmen zu können. Fehlsichtige Studierende überprüfen mit dieser Methode ihre vom Augenarzt verordnete korrigierende Brillenglasstärke (bitte Brillenpass mitbringen) durch ausprobierendes Vorsetzen von: sphärischer Linsen (minus = konkav = weiß) oder (plus = konvex = schwarz) b) Wiederholung der Prüfung bei einem Weit- oder Normalsichtigen nach Vorsetzen einer schwarz-gefassten Linse von +4 dpt: das Auge ist dadurch künstlich extrem „kurzsichtig“. Nun korrigiert man durch Addition von Zerstreuungslinsen solange, bis der Proband eine Zeile scharf sieht. Eine Differenz in Dioptrien zeigt den Grad der Weitsichtigkeit an. c) Betrachten Sie die Sehprobentafel nun auch durch die mittels eines Striches orientierte zylindrische konkave (bei Myopie) oder konvexe (bei Hyperopie) Linsen und versuchen Sie, die Sehschärfe durch Kompensation eines möglichen Astigmatismus durch langsames Drehen dieser Linse zu optimieren. Tragen Sie die Versuchsergebnisse Ihres getesteten Auges in die Tabelle ein (Grad° = Winkel der Orientierung = Strich auf der Linse des Zylinders im Auge, % = Sehschärfe in Prozent). Bei normalen Augen ist häufig, bedingt durch den Liddruck, der senkrechte Meridian der Cornea etwas stärker gekrümmt als der waagerechte (physiologischer Astigmatismus: bis 0,5 dpt). sphärisch zylindr. SINNE 19 % sphärisch zylindr. % rechts links Grad° Grad° Das Sehen ohne Linsenkorrektur (Visus sine correctione) bezeichnet man als "Sehleistung" oder “Rohvisus“; die Gesamtleistung des optischen, neuronalen und kortikalen Sinnessystems bezeichnet man als “Sehvermögen“. Die Sehschärfe ist abhängig von vielen Faktoren !! Dioptrischer Apparat - Transparenz der brechenden Medien (Xerophthalmien, Cataracta) - Abbildungsfehler (Refraktions-Anomalien) - Lage des Fernpunktes (Achsen-Ametropien) - Pupillen-Durchmesser (Schärfentiefe) Netzhaut - Orte wie Foveola, Fovea, para-, perifoveal - Status der Retina, Sensoren-Dichte u. -Verteilung, Größe primär-rezeptiver Felder - Gesichtsfeld und Sehbahn Messmethode - Untersuchungs-/ Untersucherstrategie, Patientenalter u. -ansprechbarkeit - Optotypen (Art, Ausrichtung, Abstand, Größen, Kontrast, Leuchtdichte) - Raumbeleuchtung, Prüfentfernung, Darbietungszeit Parameter wie: - Binokulares Sehen (Schielen) - Augeninnendruck (Glaukom) - Augendurchblutung, Pigmentierung (Retinopathien) - Metabolische Störungen (Diabetes mellitus) Versuch 3 Messung des GESICHTSFELDES Das Gesichtsfeld ist der Bereich der Umwelt, der mit dem unbewegten Auge wahrgenommen wird. Mit dem Perimeter kann es für beide Augen getrennt bestimmt werden. Zur Untersuchung wird der Kopf an Kinn und Stirn abgestützt, das nicht untersuchte Auge wird abgedeckt und mit dem zu untersuchenden Auge der Punkt in der Mitte des Perimeters fixiert. Dieser stellt den Mittelpunkt des Gesichtsfeldes dar (0°). Zur Bestimmung der Gesichtsfeldgrenzen werden von der Peripherie her kommend Lichtpunkte ins Gesichtsfeld hineinbewegt (= kinetische Prüfung). Dies geschieht unter verschiedenen Winkeln. Mit Wahrnehmung des Lichtreizes ist die Grenze erreicht. Sie werden von der Assistenz in den Gebrauch des Gerätes eingewiesen. Bestimmen Sie die Grenzen des Gesichtsfeldes des rechten Auges Ihres Praktikumpartners für einen permanenten weißen Lichtpunkt (kinetische Perimetrie). Verbinden Sie die 8 Stichmarken und kleben Sie das Diagramm in Ihre Praktikumanleitung ein. Wie ist ein Perimeter aufgebaut? Warum ist das Gesichtsfeld nicht kreisrund? Ändert sich das Gesichtsfeld nach Vorschalten von Farbfiltern für Blau, Rot, Grün? Was versteht man unter „Skotom“; welche verschiedenen Formen von Skotomen gibt es? Versuch 4 Prüfung des FARBEN-SEHENS SINNE 20 Überprüfen Sie zusätzlich Ihre Farbsehtüchtigkeit mit den iso-chromatischen Farbtafeln nach Ishihara (Führerschein-Test) zur Aufdeckung von extremen Farbseh-Störungen. Farben-Fehlsichtigkeiten: Trichromasie: = ein Zapfentyp ist partiell defekt Protanomalie (rot-schwach) Deuteranomalie (grün-schwach) Tritanopie (blau-gelb-schwach) Dichromasie: = es fehlt ein Zapfentyp total Protanopie (rot-blind) Deuteranopie (grün-blind) Tritanopie (blau-gelb-blind) Wie häufig sind Farben-Fehlsichtigkeiten bei Frauen und Männern? Versuch 5 Prüfung des SEHVERMÖGENS a) Das Gerät „Rodatest“ wird u.a. generell in der betriebsärztlichen Diagnostik eingesetzt und ermöglicht die subjektive Messung: 1.) des Visus bei Hyperopie mit Landoldt-Ringen (im Vergleich zur Visus-Bestimmung mittels Sehprobentafel), 2.) des Gesichtsfeldes (im Vergleich zur kinetischen Perimetrie), 3.) des stereo-skopischen Sehvermögens, 4.) der Farbsehtüchtigkeit und 5.) des Kontrast-Sehens mit und ohne Blendung eines Probanden. SINNE 21 HÖREN Autoren: Zsoka Schwab, Prof. Dr. Hans-Peter Richter Dr. Helmut Machulla, Dr. Bernd Bufe das Ohr ist unser zweit-wichtigstes Fern-Sinnesorgan!! Das Hören ist bereits im Kleinkind optimal funktionsfähig. Es dient u.a. zur frühen Sprachbildung, zum Orientieren im Raum, zum Schutz vor Gefahren Wichtig -- zum besseren Verständnis siehe Vorlesung im C.I.P.-Pool !! unser Hörvermögen ist abhängig von allen strukturellen, akusto-physikalischen, sensor-physiologischen, psycho-physikalischen, neuro-anatomischen Parametern der beiden Sinneskanäle !! Inhalts-Übersicht A. Physikalische Grundlagen Seite 24 1. Schall 2. Werte, Definitionen, Begriffe B. Psychophysik des Hörens 21 1. Hörfeld / Hörfläche C. Funktionen des Hörorgans 23 1. Anatomischer Überblick 2. Schall-Weg über Außen-, Mittel-, Innen-Ohr Reiz-Leitung u. -Verstärkung, Cochlea, Corti-Organ, Frequenz-Dispersion 3. Signaltransduktion 4. Erregungs-Weiterleitung über die Hörbahn D. Pathophysiologie des Hörens 28 1. Hörschäden 2. Subjektive Hörtests 3. Objektive Hörtests E. Anleitungen zu den Aufgaben -- Hausaufgaben A) - F) -- Durchführung der Versuche 1. - 5. -- Klinische Bezüge 31 SINNE 22 A. Physikalische Grundlagen 1.) Was ist Schall? Als Schall bezeichnet man Druckwellen, die sich in einem elastischen Medium ausbreiten. Die Moleküle schwingen dabei hin und her und stoßen sich gegenseitig an; so dass es zu einer abwechselnden Komprimierung und Verdünnung des Mediums in Ausbreitungsrichtung (= Longitudinalwelle) kommt. Die zwei Variablen einer Schallwelle sind die Frequenz (Tonhöhe in Hz) und die Amplitude (Schalldruck in Pa, mit dem subjektiven Korrelat Lautstärke in Phon). Die Leistung des Ohrs besteht darin, mechanische Energie der Druckwellen aufzunehmen, weiterzuleiten, in elektrische Impulse (Änderung vom Membranpotenzial = Sensorpotenzial) umzuwandeln, Transmitter auszuschütten und schließlich in Form neuronaler Aktionspotenziale als Erregung zum auditorischen Kortex des ZNSs zu leiten. Dabei kann es komplexe Klänge und Geräusche in ihre Tonbestandteile auftrennen (= Dispersion) und so einer weiteren Analyse entlang der Hörbahn zugänglich machen. Töne und Klänge haben eine regelmäßige Periode, Geräusche nicht. 2.) Werte, Definitionen, Begriffe des Schalls Schall g e sch w i n d i g k e i t e n: in Luft 333; in Wasser 1.428; in Knochen 5.000 m/s. Schall d r u c k: wird durch die Amplitude der Schallwelle bestimmt [Pa] Schall d r u ck p e g e l: logarithmisches Maß für den Schalldruck in Dezibel [dB] SPL (Sound Pressure Level) = 20 log Schalldruck/Referenzschalldruck (mit Filter A) Referenzschalldruck = 1,5 10-5 Pa (= Hörschwellendruck des Ohres bei 1.000 Hz) Verzehnfacht sich der Schalldruck, nimmt der SPL um 20 dB zu Schall i n t e n s i t ä t: Schalldruck (P) Schallgeschwindigkeit (V) = N/m2 m/s = W/m2 = Schallleistung, die bei senkrechtem Einfall der Welle durch eine Fläche von 1 m2 dringt Charakteristika des hörbaren Schalls: Frequenzen: 16 – 20.000 [Hz] (S/s) (unterhalb = Infraschall, oberhalb = Ultraschall) Wellenlängen: 21 – 0,17 [m] Amplituden: 1,5 10-5 Pa (= absolute Hörschwelle) bis ca. 28 Pa (= Schmerzschwelle) Energie: 10-19 [J] (an der absoluten < Hörschwelle) Formen von Schall-Ereignissen: Ton: einfache, reine Sinusschwingung, erzeugbar nur mit Tongenerator, (Stimmgabel ist idealisiert, normalerweise geringe Obertöne) hat Klang: ein Grundton plus Obertöne, deren Frequenzen ein ganzzahlig Vielfaches vom Grundton sind. Grundton = Klanghöhe Obertöne = Klangfarbe Geräusch: wenn verschiedene hörbaren FreR. Schmidt, G. Thews, F. Lang quenzen gleichzeitig schwingen. Weißes Rauschen = alle hörbaren Frequenzen schwingen gleichzeitig. . „Physiologie d. Menschen“ 2000 Interferenz: Interferenz ist die Überlagerung von Wellen durch Superposition, d.h. durch die Addition ihrer Amplituden. Bei einem geringen Frequenzunterschied zweier Schwingungen kommt es zur Schwebung: zu hören ist dabei ein Ton mit einer Frequenz, die dem arithmetischen Mittel der beiden überlagernden Schwingungen entspricht. Die Lautstärke schwankt periodisch mit der Schwebungsfrequenz (Differenz der Frequenzen der beiden Schwingungen). Das nutzt SINNE 23 man beim Stimmen von mehreren Musikinstrumenten (die Lautstärkeempfindung schwankt nicht mehr, wenn alle Instrumente identisch gestimmt sind und klingen). Reflexion: Treffen Schallwellen auf ein Hindernis, so werden sie teilweise reflektiert. Je „schallhärter“ das Medium, desto größer wird der reflektierte Anteil (= hohe Impedanz /Wellenwiderstand/; z.B. in leeren Räumen „hallt“ es). „Schallweich“ wird es dann, wenn der absorbierte Schallanteil dementsprechend größer ist und die dämpfende Eigenschaft überwiegt (= niedrige Impedanz; z.B. Teppichboden). Resonanz: Ein schwingungsfähiges System kann durch einen Erreger zum Schwingen angeregt werden. Die Schwingungsfrequenz des Erregers zwingt dem Resonator ihre Frequenz auf. Wenn die Erregerfrequenz mit der Eigenfrequenz des Resonators übereinstimmt, kommt es zur Resonanz. Letztere dient der Verstärkung beim Schalltransport über Außen- u. Mittelohr. Schallausbreitung: Je größer die Dichte des Mediums ist, umso höher ist sein Schallwellenwiderstand, d.h. desto mehr Schallenergie ist zum Durchdringen des Mediums nötig (= Impedanzanpassung). Der Schall wird an der Oberfläche reflektiert, wenn die Schallenergie nicht zum Durchdringen des Mediums ausreicht. Andererseits erfolgt die Schallleitung umso schneller, je höher die Dichte des Mediums ist, da die gegeneinander schwingenden Teilchen näher beieinander liegen. B. Psychophysik des Hörens 1.) unser Hörfeld / Hörfläche aus: R. Klinke, H.-C. Pape, S. Silbernagl Physiologie (2005) Die Empfindung, wie laut wir einen Ton hören, hängt nicht nur vom objektiven messbaren Schalldruckpegel ab, sondern auch von der Frequenz des Tons. Es gibt Frequenzbereiche, in denen das Ohr empfindlicher auf leise Geräusche reagiert als bei anderen. Um dieser subjektiven Komponente gerecht zu werden, führte man die nicht messbaren subjektiven physiologischen Größen Lautstärke (Einheit: Phon) und Lautheit (Einheit: Sone) ein. Der Lautstärkepegel entspricht definitionsgemäß bei einer Frequenz von 1.000 Hz dem Schalldruckpegel. Isophone (siehe Abbildung Hörfeld) sind Kurven jeweils gleicher Lautstärkepegel, also gleicher Lautstärkeempfindungen. Die subjektive Hörschwelle liegt bei ca. 4 Phon. Zum Überschreiten dieser Hörschwelle sind für jede Tonfrequenz verschieden starke Schalldruckpegel nötig: Ganz tiefe Töne benötigen vergleichsweise starke Schalldruckpegel im Vergleich zu mittleren oder höheren Tönen, um noch gehört zu werden. Da die Null-Linie SINNE 24 des Schalldruckes willkürlich auf 2x10-5 Pa festgelegt ist, wird der Schalldruckpegel von 0 dB bei etwa 4.000 zum Erzeugen einer gleichen Lautstärke sogar unterschritten ( in diesem Frequenzbereich rufen hungrige Babys!). Maskierung: Durch gleichzeitig auftretende andere Töne steigt die Hörschwelle eines bestimmten Tons. Die Schmerzschwelle liegt bei ca. 130 Phon (abhängig vom Grad der Adaptation - Disco !). Die Lautheit ist eine noch subjektivere Größe als die Lautstärke. Sie beruht auf der Definition des Lautstärkepegels: 1 Sone = 40 Phon bedeutet, dass ein doppelt so laut empfundener Klang eine Lautheit von 2 Sone (4-mal lauter = 4 Sone; halb-mal so laut = 0,5 Sone) hat. Bei Lärm handelt es sich um Schallereignisse, die subjektiv von Menschen als individuell belästigend empfunden werden oder gesundheitsschädlich sind. Dabei spielt nicht nur die Lautstärke, sondern auch die Art der Geräusche eine Rolle (z. B. Kratzen über die Tafel.). Es gibt Unterschiedsschwellen für 1.) die Intensität (Amplitude): an der Hörschwelle bei 0 dB können 3-5 dB; oberhalb 40 dB sogar nur 1 dB voneinander unterschieden werden und für 2.) die Tonhöhe (Frequenz) a) simultan = zwei Töne gleichzeitig dargeboten: 18 Hz (= Schwebung); b) sukzessiv = 3 Hz bei zwei aufeinander folgend bei 1.000 Hz-Tönen; im generellen Frequenzbereich etwa 0,2-0,3 %. Weber-Fechner-Beziehung: Da sich in der Sinnesphysiologie die meisten Reizintensitäten über mehrere Zehnerpotenzen erstrecken und sich die resultierenden individuellen Empfindungen nicht objektiv messen lassen, sind von Rushton (1969) das Weber- und das Fechner-Gesetz kombiniert worden. Empfindung E als Beziehung wirkender Reizstärke I zur Reizstärke an der Absolutschwelle Io I = Weber-Fechner - Beziehung E = K log Io Die Stevens´sche Potenzfunktion beschreibt dazu noch eine lineare Beziehung von applizierter Reizintensität zur Empfindungs-Intensität für Sinnes-Modalitäten (intermodaler Intensitätsvergleich) wie Druck, Geruchs- u. Geschmacks-Qualitäten, Helligkeit, Lautstärke, Temperatur, Schmerzen. C. Funktionen des Hörorgans 1. Anatomischer Überblick aus: R. F. Schmidt, K..Unsicker www..lehrbuchvorklinik.de. Die Bild-CD-ROM .(2003) SINNE 25 2. Der Weg des Schalls Station: Außenohr (Reiz-Leitung) Die trichterförmige Ohrmuschel (Auricula) bündelt die Schalldruckwelle und leitet sie in den äußeren Gehörgang bis zum Trommelfell; außerdem ist sie wichtig für das Richtungshören. Das Trommelfell wird durch die periodischen Luftdruckschwankungen zum Schwingen gebracht. Im äußeren Gehörgang (Meatus acusticus externus) wird das Cerumen (volkstümlich: Ohrenschmalz) als notwendiger physiologischer Bestandteil der Selbstreinigung sezerniert, der aus dem Sekret von Talgdrüsen vermischt mit abgestoßenen Epithelzellen, Staub, Härchen und Pigmenten besteht, bitter schmeckt und ranzig riecht (vertreibt Insekten), aber auch bakterizid und fungizid ist. Station: Mittelohr (Reizleitung u. -verstärkung) Direkt am Trommelfell befestigt ist der Hammer (malleus). Er überträgt die Schwingungen des Trommelfells auf den Amboss (incus) und dieser auf den Steigbügel (stapes), der über das ovale Fenster mit dem Perilymphraum des Innenohres kommuniziert. Der Schall im Mittelohr wird nicht nur weitergeleitet, sondern auch noch um etwa den Faktor 30 verstärkt. Dies ist deshalb nötig, weil das Innenohr im Gegensatz zum Mittelohr mit Flüssigkeit gefüllt ist, die eine viel niedrigere Kompressibilität und einen viel höheren Schallwellenwiderstand als Luft hat. Folglich würde nur relativ wenig Schallenergie in das Innenohr gelangen können, der größte Teil würde aufgrund der erhöhten Impedanz an der Membran des ovalen Fensters reflektiert (s. Vorlesung). Der Schalldruck wird durch folgende Mechanismen verstärkt (= Impedanz-Anpassung) 1) Die Gehörknöchelchen-Kette hat eine Hebelwirkung einer Verstärkung von 1,3-fach. 2) Der Druck wird von einer relativ großen Fläche (Trommelfell ca. 55 mm2) auf eine relativ kleine Fläche (Membran des ovalen Fensters 3.2 mm2) übertragen (Flächendifferenz), was eine 17-fache Verstärkung bewirkt. 3) Die Resonanzeigenschaft des mit Luft gefüllten Außen- u. Mittelohrs trägt ebenfalls zur Schallverstärkung bei. 4) Das Mittelohr kann Schallintensitäten aktiv durch unsere zwei kleinsten Skelettmuskeln musculus tensor tympani und musculus. stapedius modulieren (abschwächen): a) bei zu hohem Schalldruckpegel kontrahieren sie reflektorisch und „versteifen“ dadurch Trommelfell und Fußplatte des Stapes, so dass diese nicht mehr so stark schwingen können. b) bei sehr niedrigem Schalldruckpegel dilatieren sie, um die Schallübertragung zu verbessern. Die beiden Reflexbögen laufen über die colliculi inferiores der Hörbahn). Klinik: überlaute Hörempfindungen (Hyperakusis) tritt bei Läsion des n. facialis auf, wenn die Läsion oberhalb der Abzweigung des n. stapedius liegt. Station: Innenohr (Reizleitung, Reizverstärkung, Signaltransduktion) Die Schwingungen der Fußplatte des Stapes werden über die Membran des ovalen Fensters auf die flüssigkeitsgefüllten Räume des Innenohres übertragen. In der Cochlea wird das Schallereignis auf Frequenzen und Intensitäten analysiert. a.) Die Cochlea = Schnecke Die Cochlea ist schneckenförmig mit 2,5 Windungen aufgebaut. Über einen knöchernen Kanal in ihrer Windungsachse (modiolus) wird sie mit Nerven und Blutgefäßen versorgt. Sie enthält drei flüssigkeitsgefüllte, etwa 3,5 cm lange, parallel laufende Schläuche, die sich einer Wendeltreppe (Treppe vom lat. scala) gleich winden und voneinander durch eine „Membran“ getrennt sind. E.P. van het Schip Bildatlas Das Innenohr (1983) SINNE 26 Auf histologischen Querschnitten sind oben ab dem ovalen Fenster die scala vestibuli und unten zum runden Fenster die scala tympani dargestellt, dazwischen liegt keilförmig der Querschnitt der scala media. Die „Reissner-Membran“, eine doppelte Epithelschicht, trennt Scala vestibuli und Scala media voneinander. Eine weitere doppelte Epithelschicht mit der eingelagerten „Basilarmembran“ (Basallamina mit verschieden langen Kollagenfibrillen) trennt die scala media von der scala tympani. Sie entspringt einem knöchernen Vorsprung des Modiolus und verbreitert sich zur Schneckenspitze hin, was sie immer schwingungsfähiger werden lässt. Auf der Basilarmembran sitzt das Sensorepithel des Corti-Organs. Scala vestibuli und scala tympani gehen am Apex der Cochlea ineinander über (diese lochförmige Verbindung heißt Helicotrema); während die scala media blind endet. In den scalae vestibuli und tympani befindet sich Perilymphe, die in ihrer Ionenzusammensetzung etwa dem Ultrafiltrat des Blutplasmas gleicht (ca. 145 mmol/l Na+Cl-). Die in der scala media befindliche Endolymphe hat dagegen etwa die ionale Zusammensetzung einer intrazellulären Flüssigkeit (ca. 145 mmol/l K+, wenig Cl- und Na+), was für die Funktion der Haarzellen von großer Bedeutung ist. Die Endolymphe wird von der stria vascularis, einem zweischichtigen, durchbluteten Epithel an der äußeren Wand der scala media produziert. Durch Na+/K+-ATPasen, Kanäle für K+ (KCNQ4 u. Kir4.1) und für Cl- (ClC-K) sowie den 1Na+/2Cl-/1K+-Kotransporter wird die hohe K+-Konzentration aufrechterhalten. b.) Das Corti-Organ Das Corti-Organ sitzt auf der Basilarmembran. Es besteht aus zwei Typen Scala von sekundären Sinneszellen - den ±vestibuli 0 mV Haarzellen: Die inneren Haarzellen (iHz) sind in einer Reihe auf dem Corti-Organ angeordnet, die äußeren (äHz) in drei Reihen weiter außen. Die Haarzellen besitzen etwa 80 40 mV apikale Stereozilien = besser Stereovilli (mit Aktinfilamenten versteift), die orgel-pfeifenartig von lang nach kurz angeordnet sind. Nur die längsten Stereovilli der äHz (!) Abb. kombiniert nach: F. Lang, P.Lang kontaktieren die „Tektorial-Membran“ + 80 mV 60 mV ± 0 mV Basiswissen Physiologie, 2007 (eine gelartige Matrix!), die vom Modiolus ausgehend das Corti-Organ überdacht. Ihr Rand berührt lymphdicht das Corti-Organ (Achtung: in Abbildungen oft unkorrekt dargestellt; = Fixierungsartefakt). Schwingt nun die Basilarmembran aufgrund des Amplitudenmaximums an der charakteristischen Stelle auf und ab, so verschieben sich dort die auf der Basilarmembran sitzenden äHz und die Tektorial-Gallerte relativ zueinander. Dies führt zu einer Ablenkung (Deflektion) der Stereovilli an ihrem Ursprung. Jeder Stereovillus ist mit dem daneben liegenden kürzeren durch einen nicht dehnbaren Proteinfaden (tip link) so verbunden, dass bei Ablenkung der längsten Stereovilli die kürzeren auch mitbewegt werden. c.) Die Frequenzdispersion Wird durch den Stapes eine Schwingung auf das ovale Fenster übertragen, trifft sie als erstes auf die Perilymphe der scala vestibuli. Dort entsteht durch „Eindellung“ des ovalen Fensters nach innen eine Druckänderung, die die Reissner- und dann die Basilar-Membran frequenzabhängig nach unten bzw. oben schwingen lässt. Die Membran des runden Fensters wölbt sich zum Druckausgleich in Richtung Paukenhöhle aus. Schwingt der Stapes mit gegebener Frequenz zurück, entsteht ein Unterdruck in der scala vestibuli, der die Basilarmembran wieder nach oben zieht, so dass sich runde Fenster in Richtung Innenohr wölbt. Der so ausgelöste frequente Wechseldruck bringt auch die Basilarmembran zum Schwingen. SINNE 27 Da sie von der Cochleabasis zu Cochleaspitze hin immer breiter, dünner und elastischer wird, schwingt sie nicht symmetrisch auf und ab, sondern es bildet sich eine sogenannte Wanderwelle, die sich von der Basis zur Spitze über die Basilarmembran ausbreitet. Dabei erreicht jede Welle zwischen 16 Hz und 20.000 Hz an einer ganz bestimmten, frequenzabhängigen Stelle ihr Amplitudenmaximum und flacht danach sofort wieder ab (rote und grüne Flächen der nachstehenden Abbildung). nach M. Euler Biol. Unsere Zeit 4 (2004) Die Amplitudenmaxima der höheren, energiereicheren Frequenzen sind in Nähe der Cochleabasis, wo die Basilarmembran noch relativ steif ist; die Maxima der niedrigen, energieärmeren Frequenzen treten dagegen näher zur Cochleaspitze auf, wo die Basilarmembran immer elastischer wird (siehe Vorlesung). Nur durch dieses Amplitudenmaximum kann einzig und allein eine optimale Erregung der Haarzellen hervorgerufen werden. Man könnte quasi das gesamte Corti-Organ mit er Harfe vergleichen: die Wanderwellen-Maxima wären die Anschläge, die Saiten wären die unterschiedlich langen Kollagenfibrillen der Basilarmembran. 3. Die Signaltransduktion (elektrische Veränderungen in den Haarzellen bei Änderung der Deflektion) Bei gegebener Schalladaptation, führt eine Deflektion in Richtung der größten Stereovilli durch Anspannen der tip links zu einer erhöhten mechanischen Öffnung von K+-Kanälen in der Membran des Stereovillus (= Transduktionskanäle), durch die Kalium-Ionen aus der Endolymphe in die Zelle einströmen und eine Depolarisation der Haarzellmembran bewirken. Deflektion in Gegenrichtung bewirkt eine Erschlaffung der tip links und führt zu vermehrtem Schließen von K+-Kanälen und dadurch zu einer Hyperpolarisation. Die treibende Kraft für den K+-Einstrom ist hier lediglich die Potenzialdifferenz zwischen den äußere Haarzellen (– 70 mV) bzw. der inneren Haarzellen (– 40 mV) und der etwa +80 mV geladenen Endolymphe (= endo-cochleäres Potenzial). Das Membranpotenzial und das endo-cochleäre Potenzial ergeben eine Potenzialdifferenz von etwa 150 bzw. 120 mV, die die K+ aus der Endolymphe in die Haarzelle treibt (siehe auch Abbildung unter Abschnitt 2b und Vorlesung). Die Änderung des Membranpotenzials führt bei den äußeren Haarzellen in erster Linie zu schwingungsverstärkenden Bewegungen (cochleärer Verstärker u. Modulator), vermittelt durch das spannungs-aktivierbare membranständige Motorprotein Prestin, das hochkonzentriert und massenhaft in der lateralen Zellmembran vorkommt und sich bei Depolarisation kontrahiert bzw. bei Hyperpolarisation entspannt. Bei Depolarisierung strömen Chlorionen aus einer Tasche in den Prestinmolekülen aus, wodurch diese kleiner werden und sich somit die ganze Zelle verkürzt. Bei anschließender Hyperpolarisierung ist es dann umgekehrt. Infolge dieses Prozesses werden auch die Stereovilli der eigentlich „hörenden“ inneren Haarzellen abgelenkt, was auch hier zu einer Depolarisation oder Hyperpolarisation (= Sensor-Potenzial) führt. Auf der basolateralen Seite der Zellen befinden sich K+-Kanäle, durch die das Kalium wieder extrazellulär gelangt und über die Stützzellen und deren Gap junctions in die Marginalzellen der Stria vascularis recycelt werden kann. Nach erhöhter SINNE 28 Beschallung bewirkt die Depolarisation nachfolgend eine Öffnung von spannungsgesteuerten Ca2+-Kanälen, woraufhin es durch die erhöhte Ca2+-Konzentration intrazellulär zur Exozytose Glutamat-haltiger Vesikel mit der basolateralen Membran kommt. Glutamat löst an den Postsynapsen der afferenten Neurone (deren Zellkörper sitzen als Ganglionspirale im Modiolus) ein exzitatorisch-postsynaptisches Potential aus. Ein EPSP reicht aus, um am Axonhügel der Zelle ein Aktionspotenzial (AP) zu generieren. Diese APFrequenzen werden über die Axone des n. cochlearis (n. vestibulo-cochlearis) zu den Ncll. cochleares im Hirnstamm geleitet (siehe Vorlesung). Die verschiedenen Frequenzkomponenten von Klängen haben ihre Amplitudenmaxima an verschiedenen spezifischen Stellen auf der Basilarmembran, so dass sie nur durch verschiedene spezifische Haarzellen registriert werden. Dadurch kommt es zu einer Auftrennung (Dispersion) des Klanges/Geräusches in seine einzelnen Frequenzen (= Fourier-Analyse). Die Amplitude hingegen wird über die AP-Frequenz kodiert. Bei einer gegebenen Adaptation besteht eine Grundentladungsfrequenz von etwa 20-40 AP/s im Neuron, obgleich die Haarzelle nicht direkt gereizt wird: 1) da die tip links immer ein wenig vorgespannt sind, so dass einige K+-Kanäle immer offen sind, 2) da bereits das Rauschen des Blutes der carotis interna stets Schall auf die Lymphräume appliziert (Wir haben noch nie nichts gehört !!). Je größer die Schallintensität, desto mehr Deflektion entsteht. Bei erhöhter Reizung der Zelle öffnen, sich mit dem Schalldruckpegel korrelierend, weitere K+-Kanäle. Entsprechend stärker sind die Depolarisation und die Ausschüttung des Transmitters sowie die damit im Neuron generierte AP-Frequenz. Bei einer Ablenkung in Gegenrichtung erschlaffen mehr tip links, d.h. die meisten K+-Kanäle schließen, und es kommt zur Abnahme der AP-Frequenz. Ist die Frequenz der Deflektion so hoch, dass die Entladungsrate eines einzelnen Neurons gesättigt ist, werden immer mehr benachbarte Neurone rekrutiert. Dies ermöglicht eine Erweiterung des Dynamikbereichs. Worin unterscheiden sich innere und äußere Haarzellen? In der Cochlea gibt es viel weniger innere (iHz) als äußere (äHz) Haarzellen; das Verhältnis beträgt etwa 3.500 : 12.500. Die iHz werden von fast 90 % der Afferenzen des Nervs innerviert (die äHz nur zu wenigen %) und sind so fast allein für die Vermittlung der Erregung zum auditorischen Kortex zuständig. Im Gegensatz zu den Stereovilli der äHz haben die Stereovilli der iHz keinen Kontakt zur Tektorial-Gallerte und sind somit auf die Verstärkung der Bewegung der Endolymphe durch die äHz (durch das membranassoziierte Motorprotein Prestin) angewiesen. Diese Verstärkung geschieht beim Schwingen der Basilarmembran periodisch: Die äHz führen Längenänderungen durch, die die Endolymphbewegung soweit verstärken, dass auch die Stereovilli der iHz deflektiert werden. Dieser cochleäre Verstärker zeigt sich umso stärker, je niedriger der Schalldruckpegel ist. Er ermöglicht eine noch feinere Frequenzanalyse. Die äHz werden hauptsächlich von cholinergen efferenten Neuronen innerviert, die in Richtung colliculi inferiores ziehen und Teil der Reflexbahnen zu den zwei quergestreiften Mittelohrmuskeln sind. 4. Die Hörbahn (Erregungsfortleitung und Kodierung) Die Hörbahn ist mit ihren vielen verschiedenen Faserverläufe extrem komplex (siehe Anatomie). In Kürze: eine gereizte Haarzelle erregt synaptisch etwa 10-20 nachgeschaltete, afferente Neurone; jedes dieser Neurone innerviert also nur diese eine Haarzelle! Jedes Neuron kodiert für ein Frequenzoptimum, bei welchem es schon auf sehr niedrige Schallintensitäten reagiert. Dieses Phänomen lässt sich durch Tuning-Kurven (Empfindlichkeitskurven) verdeutlichen. Ab dem ganglion spirale gelangt über myelinisierte Axone die APFrequenz zum ncl. cochlearis im Hirnstamm. Hier wird auf das zweite Neuron verschaltet. Vom ncl. cochlearis laufen Fasern zum ncl. olivaris superior der ipsi- u. kontralateralen Seite (corpus trapezoideum), die für das Richtungshören von entscheidender Bedeutung sind SINNE 29 (Verschaltung auf das 3. Neuron). Auf dieser Ebene wird auch der Stapedius-Reflex verschaltet. Von hier laufen Fasern zum colliculus inferior der Vierhügelplatte (lemniscus lateralis), wo auf das 4. Neuron verschaltet wird. Vom colliculus inferior weiter zum corpus geniculatum mediale des Thalamus (5. Verschaltung). Hier entspringt die radiatio auditiva, die die Erregung bis zur primären Hörrinde, den Heschl-Querwindungen im dorsalen Teil des Temporallappens leitet. Im Verlauf der Hörbahn bleibt die Abbildung von Teilfrequenzen eines Schallereignisses an bestimmten Stellen des auditorischen Kortex (cochleäre Tonotopie) erhalten. Wichtig ist auch, dass von den ncll. cochleares sowohl kontralateral als auch ipsilateral Fasern aufsteigend laufen. Jede Hemisphäre erhält also Fasern von beiden Nuclei, was sich bei einseitigem Hörschaden positiv auswirkt (s. akustisch-evoziertes Potenzial AEP). D. Pathophysiologie des Hörens 1. Hörschäden Hörschäden induzieren Hörverlust. Grundsätzlich unterscheidet man zwischen Schallleitungs- und Schallempfindungsstörungen. Schallleitungsstörungen betreffen das Außenoder Mittelohr. Sie können z.B. durch einen Cerumenpropfen im Außenohr, eine starke Mittelohrentzündung oder eine otosklerotische Versteifung der Gehörknöchelchenkette verursacht werden. Schallempfindungsstörungen hingegen betreffen das Innenohr (cochleär) oder die nachgeschalteten neuronalen Strukturen (retro-cochleär). Ursache für letztere kann z.B. ein sog. Akustikusneurinom (Tumor) sein. Bei Verlust der Haarzellen oder Läsion der Hörnervenfaser sind die Schäden irreversibel. Diese Schäden können durch langzeitige Lärmexposition oder mechanisch durch kurzzeitige sehr hohe Schalldruckpegel (Knalltrauma) oder durch oto-toxische Substanzen (wie Schleifendiuretika, Aspirin, Streptomycinsulfat) entstehen. Besonders alte Menschen sind von Hörschäden betroffen (Presbyakusis). Durch Sinnesüberflutung, z.B. durch Discos (über 130 dB), Handys, portable music player gibt es bereits unter Jugendlichen bis zu 30 % Hörgeschädigte. Daher sollen hier die wichtigsten Formen der Hörschäden sowie die entsprechenden Testverfahren erklärt werden. Bei den Hörtests unterscheidet man zwischen objektiv / subjektiv, hörschwellig / über-hörschwellig sowie cochleär / retro-cochleär. Die subjektiven Tests haben den Vorteil, dass sie einfach durchzuführen und preiswert sind. Sie setzen jedoch immer die Kooperation des Probanden voraus. Diese ist z.B. bei Säuglingen, Kleinkindern, Debilen und Bewusstseinsgestörten nicht gewährleistet. Hier helfen objektive Tests weiter. 2. Subjektive Hörtests a.) RINNE- Test siehe Praktikumsversuch Nr. 2. Er dient der Diagnose einer möglichen Schallleitungsstörung. Durchführung: eine Stimmgabel wird angeschlagen und auf den processus mastoideus der Versuchperson gepresst. Dadurch werden die Schallwellen direkt vom Knochen auf die Cochlea übertragen (Knochenleitung). Sobald der Proband nichts mehr hört, wird die Stimmgabel vor sein Ohr gehalten (Luftleitung). Bei intakter Schallleitung kann er jetzt dank der Schallverstärkung durch das Mittelohr den Klang wieder hören. Ist dies der Fall, so ist er Rinne-positiv. Er hat also ein normales Gehör. Hört er die Stimmgabel über die Luftleitung gar nicht, ist die Schallleitung gestört. Er ist Rinne-negativ. b.) WEBER-Test siehe Praktikumsversuch 3. Er dient der Differenzierung zwischen einer möglicher Schallempfindungsstörung und Schallleitungsstörung des jeweiligen Rinne-getesteten Ohres durch Prüfung auf eine Lateralisierung der Hörempfindung. SINNE 30 Durchführung: eine schwingende Stimmgabel wird dem Probanden auf die Schädelsagittale gepresst, wobei der Schall über Knochenleitung in beide Innenohren gelangt. Hörgesunde hören den Klang in der Kopfmitte. Empfindet ein Patient den Klang auf der einen Seite lauter (er lateralisiert), hat er eine einseitige Hörstörung. Bei einer Schallempfindungsstörung wird der Klang logischerweise im gesunderen Ohr lauter wahrgenommen. Bei einer Schallleitungsstörung wird der Ton im kranken Ohr lauter empfunden. Dafür gibt es zwei mögliche Ursachen: 1) Bei einer Schalleitungsstörung wird weniger Schall auf die Cochlea des kranken Ohrs übertragen. Die äußeren Haarzellen der Cochlea im kranken Ohr sind daher an niedrige Amplituden adaptiert. Da die Schallleitung beim Weber-Test über den Knochen erfolgt, wird er gleichgut auf beide Ohren übertragen. Da die Cochlea des geschädigten Ohrs aber efferent mehr sensitiviert wird, werden die Schwingungen dort lauter wahrgenommen. 2) Der Schallabfluss kann gestört sein. So führt z.B. ein Cerumenpfropfen oder eine durch Entzündungsherde beschwerte Gehörknöchelchen-Kette zu einer erhöhten Reflektion des aus dem Ohr austretenden Schalls. c.) Hörschwellen-Audiometrie (dritter hörschwelliger Test) Praktikumsversuch Es wird ein Tonaudiogramm erstellt, bei dem die Hörschwellen-Isophone (siehe Hörfeld oben) linearisiert ist, und eine Hörstörung als Hörverlust ermittelt wird. Durchführung: dem einzelnen Ohr werden über einen Kopfhörer acht reine Sinustöne zwischen 60 Hz und 12 kHz sukzessive angeboten. Der Schalldruckpegel wird von unter Null langsam erhöht. Der Proband signalisiert dann, sobald er den Ton hört (was seiner individuellen Hörschwelle für diesen Ton entspricht). Abweichungen von der Normkurve werden nach unten abgetragen (Hörverlust). Das Schwellenaudiogramm liegt beim Hörgesunden bei ± 0 dB Hörverlust über den gesamten getesteten Frequenzbereich. d.) Short Increment Sensitivity Index (standardisierter, über-hörschwelliger S.I.S.I.-Test) Untersucht wird, ob der Proband kleine Lautstärkeänderungen erkennt. Bei Vorliegen einer cochleären Schallempfindungsschwerhörigkeit erkennt der Proband solche Änderungen besser, weil die äußeren Haarzellen (die beim Hörgesunden leisen Schall verstärken und lauten dämpfen) bei ihm insuffizient sind. Durchführung: dem Probanden wird für jedes Ohr separat in 20 kurzen 1-s-Intervallen ein 1.000 Hz-Ton angeboten, der sukzessive um 1-5 dB lauter wird. Er signalisiert, wenn er eine Lautstärkenänderung erkannt hat. Bei 60 % erkannter Intervalle ist er „S.I.S.I.-positiv“. Ein Hörgesunder benötigt einen Unterschied von bis zu 5 dB, um überhaupt eine Veränderung zu registrieren; einem Innenohrgeschädigten reicht 1 dB (dieser klinisch wichtige Test kann im Praktikum leider nicht durchgeführt werden). e.) Recruitment-Test (über-hörschwelliger Test nach J. FOWLER) Eine Methode zum Vergleich der Lautstärke-Empfindlichkeit beider Ohren bei einer einseitigen oder erheblich seiten-unterschiedlichen Schwerhörigkeit (nach dem S.I.S.I-Test durchzuführen). Durchführung: beiden Ohren wird simultan ein Ton angeboten. Die Schalldruckpegel werden auf jeder Seite so eingestellt, dass der Proband den Ton auf beiden Ohren als gleich laut empfindet (real ist der Pegel am kranken Ohr höher). Dies wiederholt man mehrfach mit immer höheren Schalldruckpegeln. Bei einer Schallleitungsschwerhörigkeit bleibt das Verhältnis zwischen den beiden eingestellten Schalldruckpegeln immer gleich -- das Mittelohr leitet dann einfach auf jedem Schalldruckniveau gleich „schlecht“. Bei einer Schallempfindungsstörung mit sog. „Positivem Recruitment“ hingegen, gleicht sich der Hörverlust mit zunehmender Lautstärke aus. Im gesunden Ohr beginnen die äußeren Haarzellen mit der Dämpfung bzw. Verstärkung der angebotenen Intensität. Benachbarte innere Haarzellen SINNE 31 werden dann in den Transduktionsprozess einbezogen (die Assistenz skizziert eine TuningKurve). Bei einem bestimmten Pegel empfindet der Proband auf beiden Ohren den Ton als gleich laut = Lautheits-Ausgleich (dieser klinisch wichtige Test wird im Praktikum erklärt). 3. Objektive Hörtests a.) Oto-Akustische Emissionen (OAE) Oto-akustische Emissionen sind Schallereignisse, die durch die oszillatorischen Bewegungen der äußeren Haarzellen entstehen und retrograd (also über ovales Fenster, Gehörknöchelchen und Trommelfell) aus dem Ohr hinausgeleitet (emittiert) werden. Sie können spontan entstehen oder durch Beschallung mit „Klick“-Geräuschen provoziert werden. Sie werden über Mikrophone registriert und lassen Rückschlüsse speziell auf die Funktion des Corti-Organs [ aus: schwerhoerigenforum.de ] zu (= wichtigster Hörtest beim postnatalen Hör-Screening. Dieser Test ist klinischer Standard beim Hörscreening bereits von Säuglingen, um frühzeitig Hörstörungen zu diagnostizieren und therapeutisch gezielt die Sprachentwicklung zu fördern. b.) Tympanometrie Dieser Test misst die Compliance (Schwingungsfähigkeit) des gesamten Mittelohrapparates mit einem in den Gehörgang eingeführten Lautsprecher und Mikrophon (= Tympanometer). Durchführung: ein Testton wird auf das Trommelfell gerichtet und dort reflektiert. Diese Reflexion ist abhängig vom akustischen Schallwellen-Widerstand (Impedanz) des Systems Trommelfell-musculus tensor tympani bzw. von der Flexibilität des Trommelfell-Gehörknöchelchen-Apparates.Der reflektierte Anteil der Schallwellen wird umso größer, je höher die Impedanz (Schallwellenwiderstend) des Trommelfells ist. c.) Stapedius-Reflex-Prüfung Beim Stapedius-Reflex handelt es sich einen Schutzmechanismus zur Schalldämpfung gegen kurze dem Innenohr gefährlich werdende Töne. Bei zu hohen Schalldruckpegeln wird über den Stapediusreflexbogen (N. cochlearis – Colliculi inferiores – N. facialis) der musculus stapedius kontrahiert und die Membran der Stapesplatte aus dem ovalen Fenster gezogen und damit versteift. Dadurch wird die Impedanz der Trommelfell-Gehörknöchelchen-Kette erhöht. Bei der Messung kommt das Tympanometer zum Einsatz. Beim normalen Stapediusreflex eines Gesunden wird der reflektierte Anteil des Schalls bei kontinuierlicher Erhöhung des Schalldrucks immer größer. Im Falle einer Erkrankung bleibt der Reflex aus. d.) Akustisch Evoziertes Potenzial (AEP) Dieser vierte, für das postnatale Hörscreening wichtige objektive Test basiert auf der Messung eines speziellen Elektro-Enzephalogramms (EEG). Durch ca. 2.000 „Klicks“ hervorgerufene Veränderungen im EEG-Muster werden von einem Mittelwertrechner herausgehoben. Die zufällig entstandenen Potenzialschwankungen der Ruheaktivität gleichen sich dadurch gegenseitig zur Nulllinie aus und die wesentlich geringeren, durch den Schallreiz hervorgerufenen Hörnerven- u. Hirnstammpotenziale (Feldpotenzialschwankungen) werden nach Fourier-Transformation als 5-7 charakteristische positive und negative Wellen mit Latenz u. Amplitude sichtbar. Auf diese Weise kann man neben Erkrankungen wie Multiple Sklerose auch Reifegrad und Läsionen der Hörbahn untersuchen. SINNE 32 PRAKTIKUM Ohr + Hören Versuche Versuch 1 einfache physikalische und physiologische Tests 1.) Stellen Sie mit Hilfe von Mikrophon u. Oszilloskop den Zeitverlauf von Schallwellen dar: Durch welche Schwingungsmuster werden Töne, Klänge, Geräusche charakterisiert? Wie haben Sie im Physikpraktikum die Frequenz eines Stimmgabelklangs bestimmt? 2.) Demonstrieren Sie mit zwei Stimmgabeln auf Klangkörpern das Phänomen der Resonanz. 3.) Erzeugen Sie mit den Stimmgabeln f1 (440 Hz) und f2 (430 Hz) eine Schwebung. Welche Frequenzen sind nach den Formeln: FS = f1+f2 / 2 und FS = f2–f1 / 2 beteiligt? Versuch 2 RINNE-Versuch Durchführung: eine a1-Stimmgabel (440 Hz) wird mit einem Klöppel angeschlagen und mit ihrem Fuß auf den processus mastoideus des Probanden gepresst (Knochenleitung!, d. h. die Schallwellen werden vom Knochen direkt auf die Cochlea übertragen). Sobald der Proband nichts mehr hört, wird die Stimmgabel vor sein Ohr gehalten (Luftleitung). Bei intakter Schallleitung hört er jetzt dank der Schallverstärkung durch das Mittelohr den Klang. Er ist „Rinne-positiv“. Wenn er die Stimmgabel über die Luftleitung gar nicht hört, ist die Schallleitung stark gestört (>20dB). Er ist „Rinne-negativ“. Anmerkungen: 1) Bei einer völlig intakten Schallleitung sollte der Testton noch 30 Sekunden zu hören sein. Ist dies nicht der Fall, so kann man von „Rinne-eingeschränkt“ sprechen. 2) Der Rinne-Test ist ein subjektiver Test der Schalleitung. Fällt der Rinne-Test positiv aus, liegt keine Schallleitungsstörung vor, eine Schallempfindungsstörung ist damit jedoch nicht ausgeschlossen. Probanden mit einem mäßigen Mittelohrschaden sind normalerweise in diesem Test unauffällig und werden als Rinne positiv diagnostiziert. aus H.-G. Boenninghaus, T. Lenarz: H-N-O-Heilkunde (2001) SINNE 33 Versuch 3 WEBER-Versuch Er dient der Differenzierung zwischen einer möglicher Schallempfindungsstörung und Schallleitungsstörung des jeweiligen Rinne-getesteten Ohres durch Prüfung auf eine Lateralisierung der Hörempfindung. Durchführung: eine schwingende Stimmgabel wird dem Probanden auf die Schädelsagittale gepresst, wobei der Schall über Knochenleitung in beide Innenohren gelangt. Hörgesunde hören den Klang in der Kopfmitte. Empfindet ein Patient den Klang auf der einen Seite lauter (er lateralisiert), hat er eine einseitige Hörstörung. Bei einer Schallempfindungsstörung wird der Klang im gesundem Ohr lauter wahrgenommen. Bei einer Schallleitungsstörung wird der Klang jedoch im kranken Ohr lauter wahrgenommen dafür gibt es zwei Gründe: 1) gestörter Schallabfluss: der Schall wird infolge der durch Entzündungsherde beschwerten Gehörknöchelchenkette vermindert nach außen abtransportiert, 2) efferent gesteuerte Adaptation: die Haarzellen sind an niedrige Amplituden adaptiert, so dass die Schwingungen über die Knochenleitung lauter wahrgenommen werden. Dieses Phänomen können Sie an sich selbst durch Hören der eigenen Sprache simulieren: laut zählen „ein-zwei-drei“ und dabei ein Außenohr verschließen. Führen Sie gegenseitig in Ihrer Praktikumszweiergruppe den RINNE-Weber-Versuch durch: -- Simulieren sie beim Weber-Test eine Schalleitungsstörung, indem sie mit dem Finger ein Ohr verschließen -- Sie werden von der Assistenz detailliert eingewiesen. -- Sorgen Sie für optimale Ruhe im Test-Studio !! aus H.-G. Boenninghaus, T. Lenarz: H-N-O-Heilkunde (2001) Aufgabe, ohne die obigen Erklärungen noch einmal zu lesen: Rinne-Test links positiv = normal und rechts negativ. Der Patient lateralisiert nach rechts. Welcher Hörschaden liegt in welchem Ohr vor ? SINNE 34 Versuch 4 Tonschwellen-AUDIOMETRIE: Die Hör- bzw. Tonschwellen-Audiometrie: - ist ein subjektiver Test der frequenzabhängigen Hörempfindlichkeit. - beschreibt das subjektive Hörvermögen für Töne, also die individuelle frequenzabhängige Hörempfindlichkeit eines Menschen unter Verwendung mehrerer definierter Tonfrequenzen bei unterschiedlichen Lautstärken (Schalldruckpegeln). - wird als Hörverlust einer jeweiligen Person dargestellt. - erlaubt die Unterscheidung zwischen Funktionsverlusten in der SchallÜbertragung über Luftleitung (den verstärkenden Mittelohrapparat) und in der Schallempfindung über Knochenleitung (die direkte Schallleitung via Knochen unter Umgehung des Mittelohrapparates). - identifiziert nicht nur den Gesamtzustand des Innenohrs, sondern auch vorhandene partielle Schäden an der Gehörschnecke (Frequenzdispersion), da die Frequenzskala des Tonschwellen-Audiogrammes (x-Achse) die Anordnung der Frequenzempfindlichkeit auf der Cochlea widerspiegelt. - Gleichzeitig zeigt sich anhand der benötigten Tonstärke der Grad der Zerstörung der Hörzellen innerhalb einer definierten Tonfrequenz. Zur Vereinfachung werden die Schalldruckpegel der Isophone des Hörfeldes im Tonschwellen-Audiogramm linearisiert, d. h. die Hörschwelle des Hörfeldes wird auf eine gerade Grundlinie gebracht (- quasi entzerrt). Durchführung: Die für die Tonschwellen-Audiometrie benutzten Testtöne sind genau definierbar und haben - 12 festgelegte Frequenzen (= Tonhöhen), für die je ein Hebel am Gerät vorhanden ist - Lautstärken von leise (-20 bis 0 dB) bis laut (130 dB), die durch Verschieben des Hebel kontinuierlich ineinander übergehen. Die Untersuchung erfolgt sowohl über Prüfung der Luftleitung als auch der Knochenleitung bei definierten Tonfrequenzen. Die Luftleitung prüft die Schallleitungsverstärkung durch die Gehörknöchelchen-Trommelfell-Kette im Mittelohr. Die Knochenleitung prüft die Schallempfindlichkeit des Innenohrapparates. Für die Tonschwellen-Audiometrie muss ein möglichst schalldichter Raum vorhanden sein. Für den Test selbst sollte eine entspannte Atmosphäre vorherrschen. Zur Erstellung eines Tonschwellen-Audiogrammes spielt ein Proband die Testperson und einer den Arzt, danach werden die Rollen gewechselt. Zu testende Gehörseite einstellen (rechts, links) und den bereit liegenden Audiogrammbogen eingelegt. Dann wird jede Person praktisch zweimal getestet: zuerst wird bei allen Frequenzen die Luftleitung (LL) und dann erst die Knochenleitung (KL) gemessen. Für die LL muss die Testperson einen über Kopfhörer eingespielten Ton hören. Der Untersucher spielt zuerst jedes Mal, nachdem er der Testperson der Messbeginn mitteilt, der Reihe nach bestimmte Töne in steigender Frequenz über den LL-Kopfhörer ab, indem er kontinuierlich und sehr langsam die Lautstärke des jeweiligen Tones gleicher Frequenz durch Bewegen des entsprechenden Hebels am Gerät solange erhöht, bis der Ton von der Testperson gehört wird. Die Testperson zeigt durch Drücken eines Signalknopfes sofort an, dass sie den Ton zum ersten Mal hört, während der Untersucher dies auf dem Audiogrammbogen durch Markierung der Hebelstellung ( = Lautstärke) vermerkt. Da der Test die Mitarbeit der Testperson benötigt, also subjektiv ist, sollten zur Kontrolle alle oder bestimmte Töne ein weiteres Mal überprüft SINNE 35 werden bis die Hörschwelle genau festgestellt ist. Zur Untersuchung des Innenohrs (Knochenleitung KL) wird der Ton dann über einen Spezialkopfhörer abgegeben, der auf den Processus masteideus (Mastoid) hinter der Ohrmuschel des geprüften Ohrs aufgesetzt wird. Dieser Spezial-Kopfhörer besteht aus einem Metallvibrator, von welchem ein definierter Ton an die Schädelknochen zur Weiterleitung an die Hörzellen abgegeben wird. Der festgestellte Dezibel-Wert ist bei Gesunden für die LL und KL bei jeder Frequenz immer äquivalent zur normalen Hörschwelle. Er liegt also bei ca. 0-10 dB (siehe Abb. A). Bei verschlossenem Gehörgang ist die Schallleitungsverstärkung im Mittelohr verringert, was sich im Absinken der LL-Hörschwelle zeigt (Abb. B). Das komplette Fehlen von Trommelfell-Gehörknöchelchen wäre der Maximalfall, da damit die Schallleitungsverstärkung im Mittelohr nicht nur geschwächt ist, sondern entfällt, d.h. ca. 30-40 dB Hörverlust (Abb. C). In beiden Fällen bleibt die Schallempfindlichkeit als Innenohrfunktion unverändert, weshalb die Knochenleitungshörschwelle an der Nulllinie verbleibt. Im Falle eines Innenohrschadens (Hörzellenverlust) senkt sich die KL-Hörschwelle (Abb. D). Da die LL-Hörschwelle nicht über der KL-Schwelle befinden kann, wandert sie dabei mit. Hinweis: Im Falle einer hier nicht gezeigten zusätzlich gestörten Schallleitung würde die LLHörschwelle sich aufsummieren und entsprechend tiefer unter der KL-Hörschwelle liegen (ähnlich Abb. B). Die geprüfte Frequenz als Hörschwelle ist in ein genormtes Formular des TonschwellenAudiogramms einzutragen. Auf der horizontalen (x-) Achse des Audiogramms ist die Tonhöhe (Frequenz in Hertz) angegeben, während die senkrechte Achse die Lautstärke in Dezibel angibt, bei der die Hörschwelle liegt. Auf den Formularen wird eingetragen, welches Ohr gemessen wurde (rechts bzw. links) und die durch Verbindung der Messpunkte erhaltenen Kurven mit LL (Luftleitung) und KL (Knochenleitung) beschriftet. Ein Tonschwellen-Audiogramm besteht somit aus Luftleitungshörschwelle (Leitung der Schallsignale über das Außenohr) und Knochenleitungshörschwelle (Leitung der Schallsignale über die Schädelknochen). Merke: Da die LL von der Schallempfindlichkeit des Innenohres abhängt, muss der Hörverlust bei LL immer unterhalb der KL liegen. Ist dies nicht so, dann liegen technische Fehler vor. Ein Ton kann selbst bei intakter Schallleitung nicht wahrgenommen werden, wenn die Schallempfindung defekt ist. Die LL-Hörschwellenkurve liegt also in keinem Fall über der KL-Hörschwellenkurve, sie wandert bei Erkrankung entweder mit dieser mit (Schallempfindungsstörung) oder sie trennt sich von ihr (Schallleitungsstörung). Achtung – beides kann auch kombiniert auftreten. nach: R. Schmidt, F. Lang Physiologie des Menschen (2007) SINNE 36 Ermitteln Sie Ihre Hörschwelle: -- Testen Sie sowohl die Luftleitung als auch die Knochenleitung eines Ohres. -- Welche diagnostischen Parameter liefert ein Hörschwellen-Audiogramm ? -- Warum liegen beim Gesunden Knochen- und Luftleitung auf der gleichen Grundlinie im Audiogramm? Versuch 5 Impedanz-Audiometrie: Tympanometrie und Stapedius-Reflex-Prüfung dienen: - der objektiven Untersuchung der Funktion des Mittelohrs: - dem indirekter Nachweis pathologischer Flüssigkeiten, - der Beurteilung der Druckverhältnisse und der Gehörknöchelchenkette. Zur Durchführung des Versuches dient das Tympanometer, Dessen Messsonde (siehe Abbildung) besteht aus: einem Tongenerator gibt Sondentöne mit definierter Frequenz und Schalldruckpegel in Richtung Trommelfell ab, einem Mikrofon, das an ein Messinstrument angeschlossen ist, zur Messung des vom Trommelfell reflektierten Schalls. einer Druckpumpe baut im Gehörgang definierte Drücke auf, über einen Schlauch werden Drücke aufgenommen bzw. abgeführt. aus: Welleschik: http://commons.wikimedia.org/wiki/File:Tympanometrie_Schema1.svg?uselang=de 5a Tympanometrie (= Trommelfellbeweglichkeitsmessung): Unter normalen Verhältnissen entsprechen sich die Druckverhältnisse zwischen Gehörgang und Mittelohr, wodurch sich am Trommelfell ein normaler akustischen Widerstand (akustische Impedanz) ergibt (Nullpunkt), d. h. in dieser Konstellation ist die Schallreflektion am Trommelfell am geringsten, die Schallweiterleitung also optimal. Mittels Druckpumpe kann der Druck im Gehörgang geändert werden (Über- oder Unterdruck), wodurch sich das Trommelfell mehr spannt und sich demzufolge auch dessen akustische Widerstand ändert. Infolgedessen erhöht sich auch die Schallreflexion am Trommelfell, die mittels Messmikrofon ähnlich einem Echo aufgenommen wird, da sich der Schalldruckpegel des reflektierten Tons im Gehörgang erhöht. Durchführung: Die Verarbeitung der Messwerte zum Tympanogramm erfolgt im Gerät automatisch und kann über den Interndrucker ausgedruckt werden. Der Test funktioniert nur, wenn ein auf die Messsonde aufzusetzende Stöpsel den Gehörgang dicht abschließt, da Undichtigkeiten keinen Druckauf- oder -abbau zulassen. Dafür kann entsprechend der Größe und Form des Gehörgangs der Untersuchungsperson zwischen 3 Stöpselgrößen gewählt werden. Wenn man nach Begutachtung des Gehörgangs die Sonde leicht an den Gehörgang andrückt, zeigt eine grüne Signalleuchte die Dichtigkeit an. Bei schlechtem Sitz und Undichtigkeit verändert sich die Farbe der Sonde in gelb. Undichtigkeiten werden auch am Gerätemonitor auch angezeigt. SINNE 37 Die am Trommelfell durch kontinuierlich erzeugte Änderungen des Gehörgangdruckes induzierten Schallreflexionen definierter Töne werden als Compliance in einem für Gesunde typischen Kurvenverlauf (Tympanogramm) dargestellt. Das Tympanogramm beschreibt also die Beziehung zwischen dem Gehörgangsdruck und der Compliance (= Elastizität, Kehrwert des akustischen Widerstandes [Volumendehnbarkeit in ml]) des Trommelfells. Veränderungen des Tympanogramms (Verschiebung oder Abflachung der Normalkurve) werden zur Erkennung von Krankheitsbildern benutzt: Tympanogramm eines Gesunden (Abb. a) Tubenkatarrh (Abb. b): infektbedingter Verschluss der, der Eustachischen Röhre durch dünnflüssiges Sekret verhindert Druckausgleich. Resorption des Sekrets im Mittelohr bewirkt Unterdruck, Trommelfell wird nach innen gezogen. Mittelohrvereiterung (Abb. c) führt zu Paukenhöhlenerguss. Da die Flüssigkeit die Schwingfähigkeit des Trommelfells stark dämpft, kann sich die Kurve im y-Achsenbereich nicht aufbauen, flacht also ab. fehlende(s) Gehörknöchelchen (Abb. d) = Trommelfell nicht gespannt. Compliance größer als der Messbereich. Trommelfell perforiert (keine Abb.) = Methode nicht anwendbar, da kein Druckaufbau möglich. 5b Stapediusreflex: Diese Messung erfolgt unmittelbar nach der Tympanometrie automatisch, weshalb man die Sonde weiterhin am Gehörgang angedrückt lassen muss. Bei normalem Druck im Gehörgang wird zuerst der normale und relativ leise Ton zur Impedanzmessung zugeführt. Daraufhin wird für einen kurzen Augenblick ein lauterer Ton bei einer Frequenz von 500 Hz relativ nahe unterhalb der Unbehaglichkeitsgrenze (95 Phon, siehe Hörfeld) zugeschaltet, was die Versuchsperson auch wahrnimmt, also Versuchsperson nicht erschrecken), woraufhin der Stapes und damit das Trommelfell kurz angespannt werden, das Mikrofon in der Sonde misst die sich daraus ergebende kurze Impedanzänderung (= Schallwellenwiderstandsänderung, Compliance nimmt ab). Dieser Versuch wird automatisch bei einer zweiten höheren Frequenz (4000 Hz) wiederholt. Die Versuche 5a und 5b sind an beiden Ohren der Versuchperson zu messen. Für die Messungen am linken bzw. rechten Ohr leuchtet zur besseren Erinnerung an SINNE 38 der Messsonde eine entsprechende LED in blau bzw. rot, wobei man die eingestellte Seite auch auf dem Gerätedisplay ablesen kann. Die Verschaltung des Stapediusreflexes ist nachfolgend dargestellt: Das für 5a und 5b erstellte Protokoll ist auszudrucken und auf Abweichung zu beurteilen. Letztere kann auf Fehler in der Versuchdurchführung fußen oder auf eine Erkrankung hinweisen. Ein ausbleibender Stapediusreflex deutet auf Erkrankungen hin, wie behinderte bzw. unbewegliche Gehörknöchelchen (z. B. Otosklerose), Erguss in Paukenhöhle (Eiter), Tumorbildung der Hörnerven (Akustikusneurinom) oder im Stammhirn. Der Reflex ist nicht nur in dem Ohr messbar, in dem er ausgelöst wurde (ipsilateral), sondern ebenfalls im nicht stimulierten Ohr (kontralateral). Ergebnisse hier einkleben SINNE 39 Versuch 5 Otoskopie (Ohrenspiegelung) Untersuchung des äußeren Gehörgangs und des Trommelfells Das Video-Otoskop (hier: MS101 von APPLE Bio Medical Inc.) ist eine Weiterentwicklung des veralterten Ohrtrichter-Stirnreflektor/-spiegel-Systems (arbeitet noch immer nach dem ab 1860 eingeführten Anton von Tröltsch-Prinzip). Das Otoskop setzt sich aus einem Korpus (bestehend aus Gerätedisplay, Ohrtrichter, integriertem Kamerateil sowie Kaltlichtquelle) und einem Griff mit Trockenzellbatterie (reicht für 4 Stunden Betrieb) zusammen. Es verfügt außerdem über einen Videoanschluss und zeigt den Batterieverbrauch an (grünes Licht = 0 geladen, rot = verbraucht). Es dient Ohrenärzten und Hörgeräteakustikern, da es relativ unkompliziert eine gute Einsicht in den äußeren Gehörgang bis direkt auf das Trommelfell gewährt, wobei das Bild direkt auf dem Display wiedergegeben wird. Das Bild könnte im Bedarfsfall auch auf einen größeren Bildschirm übertragen werden. Da das Trommelfell durchscheinend ist, lassen sich zu einem gewissen Grad Aussagen zur Paukenhöhle machen. von: D. Descouens http://commons.wikipedia.org/wiki/File:Conduit-auditif-externe.jpg?uselang=de 2007 Durchführung: Zu Beginn eines jeden Einsatzes des Otoskops wird über den Ohrentrichter eine Einweghülle aufgezogen bis diese mit einem Klick einrastet. Hinweis: Ist die Einweghülle nicht richtig eingerastet, bekommt man auf dem Gerätedisplay unscharfe Bilder. Dann schaltet man das Otoskop nur durch ca. 2 sec Drücken der AN-AUS-Taste ein. Wenn das Display aufleuchtet, ist das Gerät einsatzbereit. Dazu führt man den Ohrtrichter mit aller gebotenen Vorsicht ins Ohr der sitzenden Untersuchungsperson ein bis man auf dem Display das Trommelfell scharf erkennen kann. Diese Einsicht ins Außenohr dient einer ersten Begutachtung bei der Untersuchung des Ohres. Bei einem gesunden Probanden sind Gehörgang und Trommelfell weder gerötet, noch geschwollen und, abgesehen vom Cerumen (Ohrenschmalz), sind keine flüssigen oder eitrigen Ansammlungen vorhanden – das Trommelfell zeigt sich glatt, glänzend und weiß. Außer Paukenhöhlenerguss, Trommelfellschädigungen (Perforation, Ruptur), Wucherungen oder Entzündungen, lassen sich so auch ein Ohrschmalzpfopf, Fremdkörper oder Parasiten feststellen. Nicht nur ein Loch im Trommelfell, sondern auch ein eingezogenes oder vorgewölbtes Trommelfell kann ein Hinweis auf krankhafte Vorgänge sein. Hinweis: Da das Gerät batteriebetrieben läuft, ist es durch Druck der AN-AUS-Taste unbedingt abzuschalten, wenn es nicht in Benutzung ist. ACHTEN SIE AUF SICHERHEIT IM UMGANG MIT DEM INSTRUMENT!! An diesem vom übrigen Praktikumsraum abgetrennten Teil dürfen sich nur die beiden am Versuch beteiligten Personen aufhalten, um Verletzungen durch Stoßen oder Schubsen zu vermeiden. SINNE 40 GLEICHGEWICHT Autoren: Zsoka Schwab, Prof. Dr. H.-Peter Richter Dr. Bernd Bufe Inhalts-Übersicht Seite A. Allgemeines 38 B. Anatomische Grundlagen 39 C. Physiologie der Macula-Organe 40 D. Physiologie der Bogengang-Organe 41 E. Regulation des Gleichgewichts 42 1. Vestibularis-Neurone 2. Vestibularis-Bahnen 3. Vestibularis-Reflexe F. weitere Anleitungen zu den Aufgaben 46 -- Hausaufgaben A) –F) -- Durchführung der Versuche 1 - 4 -- Klinische Bezüge A. Allgemeines Unser Gleichgewichtssinn ist ein im Alltag oft vergessener Sinn, dessen Wichtigkeit uns aber spätestens dann extrem bewusst wird, wenn er plötzlich nicht mehr richtig funktioniert. Solange er es jedoch tut, gewährleistet er uns einen sicheren Gang und Stand. Darüber hinaus versorgt er uns mit wichtigen Informationen über Bewegungen, denen wir ausgesetzt sind oder die wir selbst verursachen, sowie über die Stellung unseres Körpers im Raum. Um all dies einwandfrei bewerkstelligen zu können, ist die fein abgestimmte Zusammenarbeit von Gleichgewichtsorganen, Kleinhirn, visuellen und senso-motorischen Systemen erforderlich. SINNE 41 B. Anatomische Grundlagen Das Gleichgewichtsorgan (Vestibularorgan, Vestibularapparat, Labyrinth) gehört anatomisch zum Innenohr, hat jedoch völlig andere Funktionen. Es besitzt einen mit Endolymphe gefüllten Bereich, das häutige Labyrinth. Dieses Labyrinth wiederum sitzt im knöchernen Labyrinth, einer gleichgeformten Aushöhlung im Felsenbein pars petrosa (es ist der härtester Knochen im Organismus: warum und wozu ?). Zwischen dem häutigem und dem knöchernem Labyrinth befindet sich die Perilymphe, getrennt durch ein Doppelepithel. Der Endolymphraum des Gleichgewichtsorgans ist über den ductus reuniens mit der scala media der Cochlea verbunden. Die Endolymphe wird in der Stria vascularis gebildet und fließt über den ductus endolymphaticus in den sog. saccus endolymphaticus ab, wo sie resorbiert wird. Die Endolymphe hat auf jeder Seite ein relativ konstantes Volumen von ca. 0,07 ml. Der Vestibularapparat besteht auf jeder Seite aus je fünf zusammenhängenden EndolymphRäumen mit kleinen Sinnesorganen: 1) die Statolithenorgane im Utriculus horizontal zur Körperachse mit der macula utriculi und im Sacculus vertikal mit der macula sacculi sowie 2) die drei Bogengangsorgane mit jeweils einer Ampulle, die das Sensorepithel enthält. Die Sensorzellen sind ebenfalls Haarzellen, besitzen jedoch im Unterschied zu denen der Cochlea außer den 60-80 Stereovilli noch jeweils ein großes Kinozilium. Jedes Macula-Organ besteht aus einem flächigen Sinnesepithel mit Haarzellen, deren Stereovilli und das Kinozilium vollständig von einer endolymph-mucopolysaccharid-haltigen und dadurch gelartigen Masse (entsprechend der Tektorial-Gallerte des Corti-Organs) umgeben sind. Auf dieser Gallerte eingebettet liegen kleine Calcit-Kristalle (Otolithen oder Statokonien), denen die gesamte Masse den Namen „Otolithen-Membran“ gibt, und damit eine höhere Dichte als die Endolymphe besitzt. (Klinik: sie können sich lösen und einen benignen paroxysmalen Lage-Nystagmus-Schwindel hervorrufen !!). Jedes Bogengangsorgan besteht aus drei Bogengängen (ducti semicircularis) = dünne Röhrchen eines Durchmessers von 0,8 mm: der Canalis ist knöchern, der Ductus häutig. Je ein horizontaler (lateralis), vorderer vertikaler (anterior), hinterer vertikaler (posterior) Bogengang steht mit dem großen Volumen des Utriculus in Verbindung. Der Endolymphschlauch ist jedoch in der Ampulle funktionell durch die Cupula getrennt. Die Cupula ist (wie Tektorial- u. Otholithen„Membran“) eine gallertige Masse (jedoch ohne Kristalleinlagerungen), die am Epithel der Ampullenwand endolymph-dicht veranskert ist und vollständig dem Sinnesepithel, der sogenannten crista ampullaris aufliegt. SINNE 42 C. Physiologie der Macula-Organe Für die Reizweiterleitung in den Maculae sowie den Bogengangsorganen sind auch hier die Stereovilli zuständig. Die Signaltransduktion erfolgt entsprechend der der inneren Haarzellen der Cochlea: aus einer adaptiven Ruhephase heraus kommt es zu einer relativen Verschiebung der gallertigen „Membranen“ und Deflektion der längsten Stereovilli zum Kinozilium hin / vom weg mit vermehrtem / verringertem Kalium-Einstrom aus der endolymphhaltigen Gallerte und somit zur Depolarisation / Hyperpolarisation sowie vermehrtem / verringertem Calcium-Einstrom vermehrter / verringerter Ausschüttung von Transmitter erhöhter / erniedrigter AP-Frequenz im ersten n. vestibularis-Neuron. Die beiden Maculae stehen im Raum senkrecht aufeinander und reagieren auf Translations= Linear-Beschleunigungen (= linear gerichtete Beschleunigung: = Änderung der Geschwindigkeit pro Zeit). Durch die Perzeption der permanent wirkenden Erd- = Gravitations-Beschleunigung (1 g = 9,18 m/s2) geben uns die Macula-Organe Aufschluss darüber, wo „unten u. oben“ ist (statische Komponente). Die Erdbeschleunigung wird unbewusst im Stehen über die macula sacculi, im Liegen über die macula utriculi registriert. Wird das Sinnesepithel der macula utriculi aus der Waagerechten durch Neigen des Kopfes etwas gekippt, dann folgt die darüber befindliche dichte Otolithen-Gallerte der Schwerkraft, so dass die in sie hineinragenden Stereozilien des apikal befindlichen Sensorepithels etwas nach schräg unten abgeschert (Deflektion) werden. Deflektion ist der adäquate Reiz auch für diese Haarzellen mit folgender Transduktion: a) wenn die Deflektion in Richtung zum Kinozilium hin erfolgt, dann werden die tip links gespannt und öffnen so vermehrt mechanisch-gesteuerten K+-Kanäle; wodurch ein K+Einstrom aus der Endolymphe erfolgt, die hier nur ein positives Bestandspotenzial von ca. 510 mV hat; die Membran der Haarzelle wird aus dem adaptiven Ruhepotenzial depolarisiert und die Glutamat-Ausschüttung wird erhöht. b) erfolgt umgekehrt die Deflektion vom Kinozilium weg, werden die Haarzellen hyperpolarisiert und somit die Erregungsweiterleitung gehemmt. Zu beachten ist, dass im Gegensatz zu den cochleären Haarzellen die vestibulären Haarzellen sogenannte „Ruhesensoren“ sind, d.h., dass sie durch die permanente Eigenbewegung des Kinozilium stets Änderungen der Ausschüttung von Transmitter bewirken und damit die hohe Ruhe-APFrequenz der Neurone auf ca. 50-90 AP/s induzieren. Auch Änderung der Linear-Beschleunigungen anderer Ursache werden hier registriert (dynamische Komponente), sowohl in der Senkrechten (z.B. Aufzug-Fahren) als auch in der Horizontalen (z.B. Bahn-Fahren). Die Otolithen auf der Gallerte haben aufgrund ihrer Trägheit das Bestreben, bei jedem Anfahren (positive Beschleunigung) etwas zurück zubleiben bzw. sich bei jedem Anhalten (negative Beschleunigung) geradlinig weiter zu bewegen. Dabei werden die verschiedenen Haarzellpopulationen erregt oder gehemmt. In den Maculae sind die Haarzellen entsprechend der Position ihres Kinoziliums nicht alle gleich, sondern in verschiedenen Richtungen ausgerichtet -- hierdurch und durch das Vorhandensein zweier senkrecht zueinander stehenden Macula-Organe, werden alle Ebenen des Raumes erfasst. Aus dem Muster, das die Erregungszustände der verschiedenen Gruppen je gleich ausgerichteter Haarzellen bilden und die daraus resultierenden AP-Frequenzen, kann das Gehirn genau errechnen, in welche Richtung der Kopf (Körper) gerade beschleunigt wird und wie stark diese Beschleunigung ist. SINNE 43 D. Physiologie der Bogengang-Organe Die drei Bogengänge pro Kopfseite sind so ausgerichtet, dass sie etwa senkrecht aufeinander stehen. In jeweils einer geometrischen Ebene liegen 1.) die beiden horizontalen Bogengänge, 2.) der rechte vordere und linke hintere vertikale Bogengang und 3.) der linke vordere und rechte hintere vertikale Bogengang. Die Kinozilien der Haarzellen sind hier im Gegensatz zu den Macula-Organen alle gleichförmig ausgerichtet: entweder alle zum Utriculus hin im horizontalen Bogengang oder alle vom Utriculus weg in den beiden vertikalen Bogengängen. Da die Cupula genau so dicht ist wie die Endolymphe, können lineare Beschleunigungen hier nicht wirken. Die Funktion wird dagegen durch Eindellen oder Ausbeulen der Cupula-Gallerte innerhalb der Ampulle ermöglicht. Da die endolymph-haltigen Röhren kreisförmig gebogen sind, kann die Endolymphe nur durch Rotations- = Winkelbeschleunigungen ( α = rad / s2 ) bewegt werden. Durch ihre Trägheit neigt Endolymphe bei Beschleunigungen dazu, ihren momentanen stationären Zustand beizubehalten - d.h., zu Beginn einer Bewegung zurück zubleiben und sich beim Anhalten weiter zu bewegen. Die Cupula würde das auch tun, doch sie ist mit der Bogengangswand verbunden und muss sich zusammen mit ihr bewegen. Dadurch stößt bei einer Rotationsbeschleunigung die Endolymphe auf der einen Seite gegen die Cupula, während sie sich auf der anderen Seite zurückzieht: die Cupula wird dabei wie ein elastisches Segel entgegen die Rotationsrichtung gedrückt, wobei auch die in sie hineinragenden Stereovilli und Kinozilien der Haarzellen abgelenkt werden. Für die horizontalen Bogengängen gilt: die Haarzellen sind auf jeder Seite so ausgerichtet, dass das Kinozilium zum Utriculus weist. Eine Ablenkung der Cupula in Richtung Utriculus (= utriculo-petal, Abb. links) führt demnach zu einer Deflektion der Stereovilli in Richtung Kinozilium und somit zur erregenden Depolarisation der Haarzelle; wohingegen eine utriculo-fugale Ablenkung (Abb. rechts) eine hemmende Hyperpolarisation hervorruft. In den vertikalen Bogengängen sind die Kinozilien der Haarzellen genau umgekehrt angeordnet: eine utrikulo-fugale Deflektion der Cupula führt hier zur Erregung der Haarzellen; eine utriculo-petale Deflektion zur Hemmung. Zu Nase beachten ist, dass die den Reiz perzipierenden Haarzellen stets ein adaptives depolarisiertes Ruhepotenzial von ca. –50 bis -60 mV haben (siehe die nur ca. –40 mV Ruhepotenzial der cochleären Haarzellen) und ebenfalls de- oder hyperpolarisiert werden können !! Erfolgt keine Beschleunigung mehr, wird durch Reibung der Endolymphe am Bogengangsepithel und durch die elastische Rückstellkraft der Cupula die Endolymphe wieder in ihre ursprüngliche Ruhelage zurück bewegt. Aus diesen zunächst etwas verwirrend anmutenden Konstellationen ergibt sich folgendes: bei einer Rotationsbeschleunigung in Richtung A werden die Haarzellen des entsprechenden Bogenganges auf der Seite A erregt, die seines Gegenspielers auf der anderen Seite, welcher in der selben Ebene liegt, hingegen gehemmt. Was am Ende zählt, ist die Differenz der APFrequenzen beider Seiten ! Wird die eine Seite erregt, während die andere auf denselben Reiz hin gleichzeitig gehemmt wird, verdoppelt das die Differenz, wodurch die Empfindlichkeit des ganzen Apparates enorm gesteigert wird (dabei sind auch Kommissurenfasern beteiligt ). Hinweis: die Bogengangsorgane verstoßen etwas in der Form gegen die Regel, dass die Vestibularorgane nur auf Beschleunigungen reagieren können: sie können auch auf Drehgeschwindigkeiten reagieren - allerdings nur bei sehr kurzzeitigen Drehbewegungen. Bei länger andauernder Rotation reagieren auch sie nur auf Beschleunigung. SINNE 44 E. Regulation des Gleichgewichts 1.) Vestibularis-Neurone Ausgeschüttetes Glutamat öffnet in den Postsynapsen der nachgeschalteten Neurone des n. vestibulare liganden-gesteuerte Ionenkanäle (Na+-Kanäle), und es entsteht ein EPSP. Dieses wird elektrotonisch zum Soma im ganglion vestibulare weitergeleitet und generiert am Axonhügel ein Aktionspotenzial. Wie die Neurone des Ganglion spirale zeigen auch die des Ganglion vestibulare eine Ruheaktivität (50-90 AP/s). Wird die Haarzelle depolarisiert, steigt die AP-Frequenz über diese Ruhefrequenz an, wird sie hyperpolarisiert, sinkt sie dementsprechend darunter. Die Stärke der Beschleunigung wird (wie bei der Cochlea der Schalldruckpegel) über die AP-Frequenz codiert. Die AP werden über den vestibulären Teil des n. vestibulo-cochlearis zu den Vestibularis-Kernen in der Medulla oblongata geleitet. Wie oben bereits angedeutet, ist die Differenz zwischen den AP-Frequenzen beider Seiten entscheidend. 2.) Vestibularis-Bahnen Auf jeder Seite befinden sich vier verschiedene Vestibulariskerne (nucleus superior, medialis, lateralis, inferior), die jeweils Eingänge für Informationen aus vielen verschiedenen Systemen sind wie z.B. Vestibularorgane, somatosensorische Systeme, visuelles System, Kleinhirn. Darüber hinaus gibt es auch Ausgänge zu Kleinhirn, Augenmuskelkernen, Thalamus, Hypothalamus, Rückenmark. Die Vestibulariskerne beider Seiten stehen über Kommissuren-Fasern miteinander in Verbindung, um Kompensation bei einseitigem Labyrinthausfall zu ermöglichen. Im Folgenden sind die wichtigsten Verschaltungen aufgeführt: 1) zum Somatosensorischen System Unser Gleichgewichtssinn soll sicheren Gang und Stand gewährleisten. Dazu müssen wir ständig ermitteln, wo gerade unten ist, damit wir unsere Gliedmaßen dementsprechend neu positionieren können, da wir ansonsten umfallen würden. Zwischen unserem Kopf und der Fläche, auf der wir gerade stehen oder liegen, befinden sich Gelenke, die über Propriozeptoren die Winkelverhältnisse zwischen Kopf, Hals und Rumpf messen und diese Informationen an die Vestibulariskerne (m. oblongata) weitergeben. Die Vestibulariskerne senden entweder direkt oder über die formatio reticularis Fasern zum Rückenmark. Die so gebildeten Bahnen des tractus vestibulospinalis medialis u. lateralis sowie des tractus reticulospinalis medialis u. lateralis gehören zum efferenten Schenkel der für das Gleichgewicht wichtigen Halte- u. Stellreflexe für die Stützmotorik (Abbildung). aus :J. Sobotta, H. Becher Atlas d. Anatomie d. Menschen III (1973)] SINNE 45 2) zum Visuellen System Eingang: Unsere aktiven und passiven Lageverhältnisse im Raum spüren wir nicht nur, wir sehen sie natürlich auch. Visuelle Informationen sind extrem dominant gegenüber allen anderen Sinnen, werden vom Gleichgewichtssinn genutzt und wirken bei Labyrinthausfällen primär kompensatorisch. Ausgang: Über den fasciculus longitudinalis medialis erreichen Fasern der Vestibulariskerne die Motoneurone des Nucleus abducens und des nuc. oculomotorius und nehmen so Einfluss auf die Okulomotorik. Dies ist nötig, um aus dem visuellen System überhaupt nutzbare Informationen gewinnen zu können. Da wir unseren Kopf stets bewegen, und wenn unsere Augen immer folgen würden, wären verwackelte Bilder das Ergebnis. Um das Problem zu minimieren, sind einige Reflexe etabliert. Unter 3.) sind die wichtigsten Reflex-Aktivitäten aufgeführt: 3) zum Kleinhirn etc. Das Kleinhirn erhält aus allen Vestibulariskernen Eingänge über Moosfasern, die in die Rinde des Vestibulo-Cerebellums (Flocculus, Nodulus, Uvula) ziehen. Die dortigen Purkinjefasern projizieren über die ncll. fastigii wieder zurück auf die Vestibulariskerne sowie auf die formatio reticularis. Das Kleinhirn beeinflusst die Reflexe von sowohl der Blickmotorik als auch der Halte- u. Stellmotorik wie bei der Willkürmotorik als Feinregulator. Natürlich können wir auch bewusst Beschleunigungen wahrnehmen: dazu ist eine Projektion der Vestibulariskerne auf die Großhirnrinde nötig (genauer: auf den parieto-insulären Kortex, Brodman-Area 2 und 7). Wie bei fast allen Sinneseindrücken geht das auch hier nur über das „Tor des Bewusstseins“, den Thalamus. Auch zum Hypothalamus führen Fasern aus den Vestibulariskernen, was den Einfluss des Gleichgewichtssinns auf unsere vegetativen Funktionen erklärt. (siehe unten: Kinetosen !) Die AP-Frequenzen der Vestibularis-Neurone beider Seiten werden vom ZNS ständig miteinander verglichen. In Ruhe sind diese Frequenzen auf beiden Seiten identisch und entsprechen der Ruheaktivität. Verändert sich die AP-Frequenz einseitig, wird dies als Drehschwindel oder Fallneigung empfunden: es erfolgen ein reflektorischer Nystagmus sowie Tonusänderung der Extensoren. Fällt das Labyrinth einer Seite aus, erniedrigt sich auf der kranken Seite die AP-Frequenz der Neurone gegenüber der gesunden Gegenseite (wo relativ hohe AP-Ruhefrequenz vorliegt: es kommt zu Drehschwindel zur gesunden Seite, TonusErniedrigung und damit Fallneigung zur kranken Seite, sowie Nystagmus zur gesunden Seite. 3.) Vestibularis-Reflexe Vestibulo-okulärer Reflex (VOR) Dieser Reflex wird durch die Bogengangsorgane vermittelt: wenn wir unseren Kopf in eine Richtung drehen, bewirken AP-Frequenzen der Vestibulariskerne an die entsprechenden Augen-Muskelkerne, dass sich beide Bulbi automatisch in die entgegengesetzte Richtung drehen. Dadurch wird das ursprüngliche Blickfeld beibehalten und das fixierte Objekt weiterhin auf der Fovea abgebildet: a) bei horizontaler Kopfdrehung werden die kompensatorischen Augenbewegungen vom horizontalen Bogengang gesteuert, b) bei vertikaler Kopfdrehung vom vorderen vertikalen Bogengang gesteuert und c) bei TorsionsBewegungen der Augen vom hinteren vertikalen Bogengang gesteuert. Klinik: dieser prägnante Reflex ist auch bei Komapatienten auslösbar -- ansonsten Zeichen für den Hirntod. SINNE 46 Nystagmus Als Nystagmus bezeichnet man rhythmische Augenbewegungen (Augenzittern), die im Prinzip dazu dienen, das Blickfeld zu stabilisieren (Verschaltung siehe Abbildung), wenn sich das Umfeld relativ zu uns bewegt, wie z. B.: a) im fahrenden Zug sitzend die Landschaft betrachten, b) einen direkt vor uns vorbeifahrenden Zug beobachten (a+b = optisch ausgelöst), c) beim Blicken, wenn man sich um die eigene Körperachse dreht wie beim Tanzen oder d) beim Verfolgen einer sich um uns drehenden Umgebung (c+d = optisch und vestibulär ausgelöst). Das Auge sucht sich dabei immer einen Fixpunkt, den es solang wie möglich verfolgt (langsame kompensatorische Bewegung = Deviation). Noch vor dem Anschlag der Bulbi innerhalb der Orbitae schnellt der Blick dann auf einen neuen Fixpunkt zurück (schnelle Rückstellbewegung = Sakkade), dem er dann wieder folgt usw. Die Richtung des Nystagmus entspricht definitionsgemäß der Richtung der schnellen Sakkade. [siehe Augenbewegungen mit Nystagmogramm in: www.nidcd.nih.gov] aus :J. Sobotta, H. Becher Atlas d. Anatomie d. Menschen III (1973) Physiologische Nystagmen: mit offenen Augen = optokinetisch (wenn sich die Umwelt dreht), und mit geschlossenen Augen = perrotatorisch (wenn wir nur kurz den Kopf drehen), post-rotatorisch (nach Stoppen aus einer längeren Drehung, klinisch), kalorisch (durch Temperatur-Änderung im äußeren Gehörgang, kann bei Tauchern natürliche Desorientierung hervorrufen, wird klinisch eingesetzt). Pathologisch ist der durch Labyrinthitis oder im Kleinhirn neuronal ausgelöste SpontanNystagmus. Spontan-Nystagmen sind oft nicht lange zu beobachten, da sich das ZNS des Patienten kompensatorisch an die Seitendifferenz habituiert und verstärkt das visuelle System zum Einstellen des Gleichgewichts nutzt, was bedeutet, dass betroffene Patienten im Dunkeln größere Gleichgewichtsstörungen haben als im Hellen. Hier ein vereinfachtes Beispiel: zu Beginn einer Drehung nach rechts registrieren die horizontalen Bogengänge eine Beschleunigung nach rechts (AP-Frequenz rechts relativ hoch, links relativ niedrig), die Information „Drehung nach rechts“ wird von den Vestibulariskernen an die Augenmuskelkerne weitergegeben, das Ergebnis ist ein Nystagmus nach rechts. Merke: der per-rotatorische Nystagmus schlägt immer in Drehrichtung. Dreht man sich weiter, jedoch ohne zu beschleunigen, bewegen sich Endolymphe und Cupula bald mit gleicher Geschwindigkeit. Dadurch stellt sich auf beiden Seiten die Ruheaktivität wieder ein, die Augenmuskelkerne erhalten die Information „Keine Drehung“ und der Nystagmus verschwindet. Stoppt man nun die Drehung, dann entspricht das einer negativen Beschleunigung. Die träge Endolymphe, die sich eigentlich weiterbewegen würde, verschiebt jetzt von der anderen Seite die Cupula. Dadurch kommt es auf der linken Seite zu einer utriculo-petalen und auf der rechten Seite zu einer utriculo-fugalen Auslenkung, was der Information „Drehung nach links“ entspricht. Das wird auch so den Augenmuskelkernen signalisiert - die Folge ist ein Nystagmus nach links. Merke: der post-rotatorische Nystagmus schlägt immer entgegen der vorherigen Drehrichtung. SINNE 47 Drehschwindel Im Zusammenhang mit dem Gleichgewichtssinn entsteht Schwindel (Vertigo) immer dann, wenn die Informationen, die aus den verschiedenen Sinneskanälen auf die Vestibulariskerne konvergieren, nicht in Einklang zu bringen sind. Wenn wir uns z.B. mit konstanter Geschwindigkeit um die eigene Achse drehen, melden die Gleichgewichtsorgane: „Körper in Ruhe“; während aber das visuelle System mitteilt: „Körper in Bewegung“, was ebenso von den Propriozeptoren registriert wird, zumindest in dem Fall, dass sich der Körper aktiv dreht. Wenn die Drehung gestoppt wird, registrieren die Gleichgewichtsorgane wieder Bewegung (negative Winkelbeschleunigung), während nunmehr die Propriozeptoren Ruhe signalisieren. Dies bewirkt im ZNS diverse Fehlinformationen, was sich als subjektives Gefühl eines Drehschwindels äußert, der zumeist in Richtung des Nystagmus geht. Kinetosen Bewegungskrankheiten (Kinetosen) = Übelkeit mit Erbrechen auf der Achterbahn, auf See oder beim Rückwärtsfahren im Zug werden unter Umständen bei einer starken Inkongruenz der Informationen erzeugt, die aus den verschiedenen Sinnessystemen oder auch den verschiedenen Vestibular-Organen in den Vestibulariskernen eintreffen. Das Erbrechen bewirkt, dass ein leerer Magen weniger Blutzufuhr benötigt, um im Rahmen einer Stressreaktion mehr Blut für Gehirn und Muskulatur zur Verfügung zu stellen. Morbus Menière 1) Meist durch gestörten Abfluss der Endolymphe bedingte Erkrankung mit spontan auftretenden, starken Schwindelattacken, Übelkeit, Gehörverlust (Tieftonbereich), Tinnitus. Theorie: Resorptionsstörung von Kalium im saccus endolyphaticus Kaliumüberschuss in Endolymphe Wasserzustrom Überdruck Endolymphatischer Hydrops (Leitsyndrom: Tieffrequenzhörsturz) 2) Die epithelialen Membranen zwischen Endolymphraum und Perilymphraum können aufgrund eines Überdrucks (Endolymphatischer Hydrops) reißen, so dass sich die kaliumreiche Endolymphe mit der Perilymphe vermischt. Folge dieser „Kaliumvergiftung“ der Perilymphe ist eine Dauerdepolarisation der Haarzellen und Drehschwindel. Die Membran verschließt sich dann wieder. Wichtig sind folgende Aspekte Charakteristik der adäquaten Reize (Deflektion wird durch welche physikalische Größe erzeugt ??); mögliche Wege der „Reiz“leitung über die einzelnen Strukturen des Ohres, Funktionsprinzip der verschiedenen Sensorflächen (Maculae staticae, Cristae ampullares), die Signaltransduktion in den Haarzellen, Erregungsleitung von den Sensoren zum propriozeptiven Kortex, neuronale Verbindungen zu Augen; Ohren und der Haltemuskulatur, physiologische Testung vom Gleichgewicht. SINNE 48 Übersicht: Auswirkung verschiedener Erregungsformen der Gleichgewichtsorgane (GGO) Durchführen von Rechtsdrehung Durchführen von Linksdrehung Abstoppen einer Rechtsdrehung Abstoppen einer Linksdrehung GGO-Ausfall rechts GGO-Ausfall links Warmspülung*: rechter äußerer Gehörgang Warmspülung*: linker äußerer Gehörgang Kaltspülung*: rechter Äußerer Gehörgang Kaltspülung* : linker Äußerer Gehörgang Bogengang APDrehschwindel, Fallneigung Frequenz Nystagmus, Muskeltonus erregt steigend nach nach Rechts Rechts Rechts Links Links Links Links Rechts Links Links Links Rechts Rechts Rechts Rechts Links Links Links Links Rechts Rechts Rechts Rechts Links Rechts (Rechts) Rechts Links Links (Links) Links Rechts Links (Links) Links Rechts Rechts (Rechts) Rechts Links * Bitte beachten: Eine einseitige Stimulation/ Inhibition durch thermische Spühlung bewirkt einen umgekehrten Effekt auf der anderen Seite (wegen kontralateraler Interneurone). SINNE 49 PRAKTIKUM Versuch GLEICHGEWICHT 1 Untersuchung der vestibulo-spinalen Reflexe Vorgehensweise bei der Prüfung der Funktion der Vestibularorgane a) Anamnese b) Welche Symptome treten bei pathologischen Veränderungen der Labyrinthe auf? c) Diagnostik Folgende einfache Untersuchungen, die den Patienten wenig belasten und keinen apparativen Aufwand erfordern, sollten zuerst durchgeführt werden! Um folgende Versuche (siehe nächste Seite) effizient durchzuführen, müssen die Augen des Patienten geschlossen sein. Warum ? a) b) c) d) Fallversuch nach Romberg Blindgang Tretversuch nach Unterberger (das Knie deutlich anheben ! Warum ?) Zeigeversuch nach Bárány Vorsicht: Welche Hilfestellung muss unbedingt erfolgen? aus H.-G. Boenninghaus, T. Lenarz: H-N-O-Heilkunde (2001) Welche Abweichungen sind bei einem akuten einseitigen Labyrinth-Ausfall zu erwarten ? Welche Vestibularis-Bahnen sind beteiligt ? Wie lassen sich diese 4 Abweich-Reaktionen (= Ausgleichsbewegungen) erklären ? Versuch 2 Einführung in das Labyrinth 1.) Modell eines Bogengang-Organs a) Besprechung des Modells eines horizontalen Bogengangs b) Modellversuch über die Bewegung der Endolymphe bei Drehbewegungen SINNE 50 c) physikalische Vorgänge zu Beginn, während, beim Abstoppen aus der Drehung d) Unterschiede zwischen kurzen und lang-anhaltenden Drehbeschleunigungen 3.) Abbildung linker horizontaler Bogengang von oben gesehen, mit Utriculus, Ampulle, Cupula, 1 Haarzelle, 1 Afferenz Skizzieren Sie Aktivierung oder Inhibierung der Haarzellen beidseits bei Kopfdrehung nach rechts !! Zeichnen Sie die jeweiligen Änderungen farbig ein. Versuch 3 Untersuchung von vestibulo-okularen Reflexen 1.) Okulomotorik: definieren Sie „Nystagmus“ a) Zeichnen Sie hier ein Nystagmogramm (3 Schläge eines horizontalen Rechts-Nystagmus): (rechts) (links) (ms) Nach welcher Komponente wird der Nystagmus benannt ? Wodurch wird die Richtung der Sakkade bestimmt ? Welche Ursachen kann ein Ausfall der vestibulären Systeme haben ? 2.) Simulieren Sie den akuten Ausfall eines horizontalen Bogengangs nach Drehung: a) Induktion des Provokationsnystagmus nach Bárány nach dem Drehstuhlversuch b) Prüfung der Reaktionen eines Probanden nach Drehung (Art, Richtung, Dauer, Palpation) SINNE 51 Vorgehen: Hilfsmittel (beleuchtete Frenzel-Brille mit +16 dpt; warum ?) Hilfsperson !! Ausführung: Wie wird gedreht und warum ? In welcher Kopflage wird der horizontale Bogengang optimal gereizt ? 3) Folgen der Reizung beider Bogengangapparate sind: a) Augen-Bewegungen (= Nystagmus als vestibulo-okularer Reflex) b) Subjektive Empfindungen: Teilen Sie Ihre individuellen Empfindungen nach einem Drehstuhl-Experiment mit. In welche Richtung empfinden Sie ? Wie erklären Sie sich diese Empfindungen ? c) Abweich-Reaktionen (Stellreflexe bzw. vestibulo-spinale Reflexe) Nach Anhalten aus der Drehung erheben Sie die gestreckten Arme (Bárány): Nach welcher Seite erfolgt die Abweichreaktion der Arme und warum ? Zeichnen Sie zum besseren Verständnis die Schemata der beiden horizontalen Bogengänge (siehe Schema oben: Stellung des Kinoziliums ?; erklären Sie "utriculo-fugal" und "utriculo-petal"). Überlegen Sie sich die Knochen-, Endolymph- und CupulaBewegungen, die beim Andrehen (per-rotatorisch), während (nach mindestens 30 sec), und nach Stoppen (post-rotatorisch) eines Drehstuhlexperiments zu beobachten sind. Markieren Sie eine Erhöhung bzw. Verringerung der Entladungsfrequenz der Afferenzen mit den Vorzeichen ‚+’ und ‚–‚. In welcher Richtung erfolgt jeweils der Nystagmus ? Welche neuronalen Verschaltungen spielen bei diesen Abweichreaktionen eine Rolle? Versuch 4 Besprechung der kalorischen Prüfung Durchführung: Proband liegt auf dem Rücken, den Kopf um etwa 30° angehoben, so dass die horizontalen Bogengänge senkrecht stehen. Man spült den äußeren Gehörgang der zu prüfenden Seite für ca. 20 Minuten abwechselnd mit 41°C warmem und 31°C kaltem Wasser. Durch das warme Wasser wird die Endolymphe im Bogengang. erwärmt und steigt nach oben, wodurch sie auf die Cupula in Richtung Utriculus drückt (utriculopetal). Dadurch (und wahrscheinlich auch direkt durch die Wärmewirkung) werden die Haarzellen des betroffenen Bogengangs utriculo-petal deflektiert und es kommt zu einem Nystagmus in Richtung geprüfter Seite. Bei Kaltspülung erfolgt das Gegenteil: die Endolymphe sinkt ab, und die Cupula wird durch den Unterdruck utriculo-fugal wegbewegt ( wodurch quasi ein Ausfall des Bogengangs simuliert wird ); der Nystagmus zeigt von der geprüften Seite weg zur Gegenseite. Worauf muss man achten, bevor der Test durchgeführt werden kann ? (Kunstfehler !) Welches Hilfsmittel wird benötigt ? Warum funktioniert diese Prüfung (anatomische Verhältnisse, physikalisches Prinzip) ?