25.2 Organisation des Cortex cerebri
Werbung

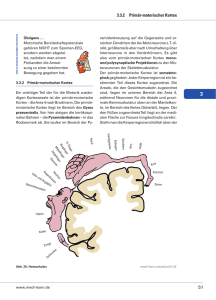
25 Integrative Funktionen des Gehirns 25.2 Organisation des Cortex cerebri 25.2.1 Gliederung des Cortex cerebri in Areale, Schichten (Laminae) und Säulen (Kolumnen) Der Cortex cerebri (zerebraler Kortex, auch als „Hirnrinde“ bezeichnet) nimmt im Menschen mit ca. 500 cm3 nahezu die Hälfte des gesamten Hirnvolumens ein. Regional spezifische histologische Merkmale ermöglichten bereits zu Beginn des 20. Jahrhunderts eine Untergliederung in 52 zytoarchitektonische Areale, die nach dem Erstbeschreiber auch als „Brodmann“-Areale bezeichnet werden (Abb. 25.2). Viele dieser histologisch definierten Areale haben sich auch als funktionell distinkt herausgestellt. Trotz aller Unterschiede besitzen die kortikalen Areale eine Reihe von Gemeinsamkeiten: ▬ einen Aufbau aus Schichten (Laminae) etwa parallel zur Oberfläche der Hirnrinde, ▬ neuronale Verbindungen in vertikaler Achse, innerhalb funktioneller Säulen (Kolumnen), ▬ neuronale Verbindungen in horizontaler Achse, die zum Beispiel die funktionellen Säulen untereinander verbinden, sowie ▬ definierte Eingangs- und Ausgangsstationen, die die primären Quellen und Ziele der afferenten und efferenten Verbindungen des jeweiligen kortikalen Areals bilden. Stammesgeschichtlich werden der Allokortex (mit Archikortex und Paläokortex), der Mesokortex und der Neokortex unterschieden. Der Neokortex macht als phylogenetisch jüngster Teil den größten Bereich des zerebralen Kortex im Menschen aus. Ein auffälliges histologisches Merkmal des Neokortex ist die Strukturierung in 6 Schichten (Laminae), die von der Oberfläche der Hirnrinde ausgehend mit fortlaufenden römischen Zahlen benannt werden (Abb. 25.3). Diese sechsschichtige Struktur ist in allen neo- 3 Lateralansicht 4 6 8 9 10 Gut untersucht sind die Säulen des primären visuellen Kortex (s. Kap. 21.5.3, S. 730 f.), in dem zum Beispiel in den so genannten Orientierungssäulen die Orientierung von Lichtreizen repräsentiert ist. Innerhalb einer Säule erfolgt die Verarbeitung der thalamokortikalen Signale durch ein dichtes synaptisches Netzwerk sowohl in serieller als auch in paralleler Weise von der Eingangschicht IV über die Schichten II/III in die Schichten V und VI, und von dort zurück in die Schicht IV (Abb. 25.3). Pyramidenzellen vor allem aus den Schichten II/ III vermitteln über lateral verzweigte, zum Teil weit reichende Axone kortiko-kortikale Verbindungen. Die Folge dieser Signalverarbeitung ist eine zunehmende Extraktion definierter Merkmale der Sinnesreize. Pyramidenzellen der Schicht VI projizieren in den Thalamus zurück, während die der Schicht V mit anderen subkortikalen Regionen, z. B. Striatum und Colliculi superiores, sowie dem Hirnstamm verbunden sind (Abb. 25.3). Im Gegensatz zum spezifischen thalamokortikalen System innervieren die Axone des unspezifischen thalamokortikalen Systems Kerngebiete diffus verzweigt bevorzugt die Schicht I. Im Neuropil der relativ zellarmen Schicht I sind darüber hinaus synaptische Kontake mit den cholinergen und monoaminergen Axonterminalen des aufsteigenden Hirnstammsystems ausgebildet, die eine Regulation der neuronalen Aktivität in primäre sensorische und motorische Areale 6 46 42 22 38 21 20 19 32 19 37 23 10 18 47 Medianansicht 7 31 24 39 41 3 1 5 9 40 44 4 8 7 2 45 11 1 5 kortikalen Regionen vorhanden, mit Ausnahme der Areale 4, 6 und 8 des motorischen Kortex (der als „agranulärer“ Kortex dem „granulären“ Kortex gegenübergestellt wird; s. Kap. 23.5.1, S. 779 ff.). Jede Schicht besitzt charakteristische funktionelle und anatomische Eigenschaften. In den primären sensorischen Kortizes ist die Schicht IV reich an Sternzellen. Hier enden die Afferenzen aus den spezifischen thalamischen Kerngebieten, den Umschaltstationen der Sinnessignale von den peripheren Sinnesorganen. Die Innervation ist topographisch organisiert, das heißt die Afferenzen benachbarter Orte der Sinnesorgane enden in benachbarten kortikalen Regionen. Dieses Innervationsmuster bildet eine wichtige Grundlage für den Aufbau vertikal orientierter funktioneller Säulen (Kolumnen), in denen definierte Aspekte der Sinnesinformation vermittelt werden. 11 17 30 Assoziationscortices Abb. 25.2 Strukturelle Organisation des Neokortex. Zytoarchitektonische Areale von Brodmann, in Lateral- (links) und Medianansicht (rechts) des menschlichen Gehirns. Blaue Farbtöne kennzeichnen As- 18 25 12 34 38 20 17 28 37 19 18 soziationskortizes, rote Farbtöne markieren primär sensorische und motorische Areale. Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. 818 25.2 Organisation des Cortex cerebri 819 Neokortex Afferenzen unspezifischer Thalamus Efferenzen I II andere kortikale Areale 2 kontralaterale kortikale Areale III spezifischer Thalamus IV kortikale Areale VI bis 4 4 Verarbeitungssequenz Thalamus weiße Substanz Abb. 25.3 Strukturelle Organisation des Neokortex. Zelluläre Organisation der 6 Schichten (I–VI) des Neokortex, mit Hauptafferenzen (links) und -efferenzen (rechts). Die Zahlen (1 – 4) zeigen die sequen- zielle Informationsverarbeitung thalamokortikaler Signale innerhalb einer vertikalen Säule (Kolumne). Abhängigkeit von Verhaltenszuständen wie zum Beispiel Aufmerksamkeit, Wachheit und Schlaf (s. Kap. 26.3, S. 860 ff.) ermöglichen. Haupteingangs- und Ausgangssystemen. Zum einen erhalten die Assoziationskortizes subkortikalen Zustrom nicht aus thalamischen Kerngebieten, die an der Weiterverschaltung primärer sensorischer und motorischer Information beteiligt sind, sondern aus solchen Kernen des Thalamus, die bereits verarbeitete Signale aus dem Kortex erhalten und diese an den Assoziationskortex zurückschalten. Die phylogenetisch alten Regionen des Archikortex, zu dem der Hippokampus gehört, und des Palaeokortex, zu dem die kortikalen Regionen des Riechhirns zählen, sind in 3 – 4 bzw. 3 Schichten gegliedert. Die funktionelle Bedeutung der unterschiedlichen Gliederung von Neo-, Archi- und Palaeokortex ist nicht genau bekannt, steht vermutlich aber in Beziehung zur Komplexität der Informationsverarbeitung in den jeweiligen Regionen. 25.2.2 Organisation und Funktion der assoziativen Areale des Kortex Die Areale mit primären sensorischen und motorischen Aufgaben belegen nur einen kleinen Teil der Hirnrinde im Vergleich mit denjenigen Arealen, die integrative Wechselwirkungen zwischen den verschiedenen Sinnessystemen und den motorischen Arealen vermitteln, und die demzufolge als Assoziationsareale (Assoziationskortizes) bezeichnet werden (Abb. 25.2). Entsprechend der anatomischen Lage werden frontaler, parietaler, temporaler und okzipitaler Assoziationskortex unterschieden. Der okzipitale Assoziationskortex nimmt insofern eine Sonderposition ein, als seine Funktionen zwar für kognitive Prozesse von Bedeutung sind, allerdings nahezu ausschließlich für solche des visuellen Systems (s. Kap. 21.5.3, S. 730 ff.), die hier nicht weiter behandelt werden. Die Organisation der Assoziationskortizes zeigt bei allen aufgeführten neokortikalen Gemeinsamkeiten wesentliche Unterschiede zu den primären sensorischen und motorischen Kortizes. Auffällig sind die Unterschiede in den Relevante Kerngebiete des Thalamus sind das Pulvinar mit Hauptprojektion in den parietalen Assoziationskortex, der Nucleus lateralis posterior mit Projektion in den temporalen Assoziationskortex und die Nuclei mediales thalami mit Projektion in den frontalen Assoziationskortex. Zum anderen sind die assoziativen Areale durch umfangreiche direkte Projektionen aus anderen kortikalen Regionen, insbesondere auch interhemisphärische kortiko-kortikale Verbindungen, gekennzeichnet. Eine dichte Innervation der Assoziationskortizes durch das aufsteigende Aktivierungssystem aus Hirnstamm (s. Kap. 26.2.3 und 26.2.4, S. 857 ff.) und basalem Vorderhirn bildet die Grundlage für die Regulation der Signalverarbeitung in Abhängigkeit eines breiten Kontinuums mentaler Zustände. Trotz des hohen Grades an wechselseitigen und zum Teil überlappenden Verbindungen sind die verschiedenen Areale der Assoziationskortizes durch distinkte Eingangs- und Ausgangssysteme charakterisiert, die im Sinne der oben genannten Module spezifische Funktionen begründen. Schlussfolgerungen bezüglich der funktionellen Bedeutung der kortikalen Assoziationsareale im Menschen sind demzufolge durch klinische Beobachtungen von Patienten mit Funktionsstörungen dieser Areale in besonderer Weise möglich. Integrative Funktionen des Gehirns 1 subkortikale Strukturen 3 V Sternzelle Pyramidenzelle 1 25 Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. kortikale Areale 25 Integrative Funktionen des Gehirns 25.2.3 Klinische Konsequenzen lokaler Funktionsstörungen des Assoziationskortex Schädigung des parietalen Assoziationskortex – Aufmerksamkeitsdefizite. Patienten mit unilateralen Läsionen in Bereichen des parietalen Kortex fallen häufig durch eine Verminderung ihrer Fähigkeiten zur bewussten Wahrnehmung auf. Die Patienten reagieren nicht oder nur reduziert auf sensorische Reize, die in Bereichen des Körpers oder des Gesichtsfeldes kontralateral der parietalen Läsion dargeboten werden, und sie haben Schwierigkeiten bei der Ausführung motorischer Aufgaben in diesen Bereichen (Abb. 25.4 A). Ihre generellen sensorischen und motorischen Leistungen sind nicht messbar verändert. Die Symptome reichen von kurzzeitigen Phasen der verminderten kontralateralen Aufmerksamkeit bis zu einer relativen Vernachlässigung der kontralateralen Körperhälfte oder Umgebung, die in individuellen Fällen so ausgeprägt sein kann, dass die Existenz der Körperhälfte geleugnet wird und sich der Patient zum Beispiel weigert, diese zu waschen oder zu bekleiden. Die oft halbseitige Vernachlässigung des eigenen Körpers oder der Umgebung wird als Neglekt bezeichnet, und die Symptome kollektiv als kontralaterales Neglektsyndrom. Die Symptome spiegeln eine Reduktion der selektiven Aufmerksamkeit wider, derjenigen Fähigkeit, unter mehreren Ereignissen nach bestimmten Kriterien auszuwählen und damit eine wichtige Voraussetzung für die bewusste Wahrnehmung zu schaffen. Die Interpretation dieser klinischen Befunde, die dem parietalen Assoziationskortex eine entscheidende (allerdings nicht exklusive) Funktion für die selektive Aufmerksamkeit zuerkennt, ist durch vielfältige Studien mit Hilfe der Elektrophysiologie und der nicht invasiven Bildgebung bestätigt worden (Abb. 25.4 B). Die kontralaterale Ausprägung ist Ausdruck der kontralateralen Grundorganisation der sensorischen (s. Kap. 18.8, S. 664 ff. und 21.5.1, S. 727 f.) und motorischen Systeme (s. Kap. 23.5.4, S. 784). Auffällig allerdings ist die Ausprägung des kontralateralen Neglektsyndroms vorwiegend bei Schädigung des rechtsseitigen Parietalkortex. Sie wird durch eine Lateralisation der Aufmerksamkeitssysteme erklärt (s. Kap. 25.8): Bei vorwiegend kontralateraler Anlage der linksseitigen parietal-kortikalen Aufmerksamkeitssysteme kann die nicht betroffene rechte Hemisphäre funktionell-kompensatorisch wirken, während bei bilateraler Anlage der rechtsseitigen Systeme diese Kompensation nicht greifen kann. Zum Verständnis des Begriffes „Aufmerksamkeit“ ist es sinnvoll, einen selektiven und einen globalen Aspekt zu unterscheiden. Die Grundlage für die Selektion von Information (selektive Aufmerksamkeit) ist ein ausreichender Grad an globaler Wachheit und Aufmerksamkeit, der über das System Hirnstamm-Thalamus-Kortex reguliert wird (vgl. Kap. 26.2.3, S. 857 ff.). Funktionsstörung des temporalen Assoziationskortex – Beeinträchtigung im Wiedererkennen. Als Agnosie wird eine Störung des (Wieder-)Erkennens bezeichnet, die nicht durch eine Störung der elementaren Sinnesleistungen verursacht ist, und bei der – im Unterschied zum Neglektsyndrom – der Patient die Existenz von sensorischen Ereignissen nicht leugnet. Er erkennt deren Zusammenhang, Bedeutung oder Bekanntheit allerdings nicht. Die Störung betrifft häufig komplexe Reizsituationen und beinhaltet lexikalische und ge- dächtnisbezogene Komponenten (Schwierigkeiten bei der Zuordnung von Sinnesreizen zu verbalen oder kognitiven Symbolen und beim Abruf von Informationen bei Präsentation bekannter Reize). Klinische Untersuchungen zeigen Läsionen oder Funktionsstörungen in Agnosie-Patienten in inferioren Regionen der temporalen Assoziationskortizes (Abb. 25.4 B). Linksseitige Störungen sind vorwiegend mit Schwierigkeiten der Wiedererkennung von Sprache assoziiert, und rechtsseitige Störungen haben visuelle Agnosien zur Folge (s. a. Kap. 25.8). In Abhängigkeit von Ort und Größe der Schädigung des inferioren temporalen Assoziationskortex können verschiedene, zum Teil objektspezifische Agnosien auftreten. Prosopagnosie (s. Kap. 21.5.4, S. 732) ist eine Form der visu- A posttraumatische Läsion, links-parietal leichtes Neglektsyndrom mit Extinktionsphänomen B lokale Schädigungen parietale Läsion Aufmerksamkeitsdefizite, Neglektsyndrom temporale Läsion Wiedererkennungsdefizite, Agnosie frontale Läsion Defizite in Handlungsplanung, Persönlichkeitsänderungen Abb. 25.4 Schädigungen des Assoziationskortex und resultierende kognitive Defizite. A Magnetresonanztomographische Aufnahmen (3 verschiedene Horizontalschnitte) des Gehirns eines männlichen, 34-jährigen Patienten mit posttraumatischer Läsion links-parietal (markiert durch roten Kreis). Der Patient zeigt ein leichtes Neglektsyndrom rechts mit Extinktionsphänomen. Zum Beispiel werden visuelle Reize in der rechten Gesichtsfeldhälfte nur vermindert wahrgenommen, wenn gleichzeitig links ähnliche Reize dargeboten werden. (Aufnahmen Prof. Dr. E. Düzel, Otto-von-Guericke-Universität Magdeburg.) B Regionen des (rechten) Assoziationskortex, die bei bestimmten kognitiven Defiziten am häufigsten betroffen sind. Markiert sind die Regionen, in denen klinische Studien eine lokale Schädigung (z. B. eine Läsion) bei entsprechendem kognitiven Defizit zeigten. Zu beachten ist, dass bei Funktionsstörungen des frontalen Assoziationskortex vielfältige Beeinträchtigungen kognitiver Leistungen bekannt sind. Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. 820 25.3 Kognition versus Emotion – Das limbische System Schädigung des frontalen Assoziationskortex – vielfältige Veränderungen der Persönlichkeit. Bei Funktionsstörungen des frontalen Assoziationskortex (Abb. 25.4 B) und der präfrontalen kortikalen Areale sind vielfältige Beeinträchtigungen kognitiver Leistungen bekannt. Sie sind Ausdruck der re- lativen Ausdehnung des frontalen Kortex im menschlichen Gehirn und dessen Bedeutung für höhere integrative Funktionen, wie zum Beispiel Handlungsplanung, Antizipation, Problemlösung, Informationsselektion und Steuerung sozialer Interaktionen (der oben geschilderte Fall Phineas Gage veranschaulicht die komplizierte Symptomatik bei Schädigung des frontalen Kortex). Eine hirnbiologisch spezifische Struktur/Funktionskorrelation ist demzufolge in den wenigsten Fällen möglich. Ein unrühmliches Beispiel für fehlgeleitete Korrelationen und resultierende medizinische Praxis ist die in den 30er- und 40er-Jahren vor allem in den Vereinigten Staaten von Amerika in Tausenden von Patienten zur Behandlung von Symptomen „geistiger Verwirrtheit“ durchgeführte neurochirurgische Resektion des Frontallappens bzw. Durchtrennung fronto-thalamischer Bahnen („Lobotomie“, „Leukotomie“). Die Eingriffe hatten vielfältige hirnorganische Psychosyndrome und Persönlichkeitsveränderungen zur Folge. Zusammenfassung Kap. 25.2 Der Cortex cerebri (zerebraler Kortex, „Hirnrinde“) wird nach Brodmann in 52 zytoarchitektonische Areale untergliedert. Die kortikalen Areale sind aus horizontalen Schichten (Laminae) und vertikal zur Oberfläche orientierten funktionellen Säulen (Kolumnen) aufgebaut. Der Neokortex enthält 6 Schichten (I – VI), die afferenten Hauptzustrom aus dem spezifischen (IV) und unspezifischen Thalamus (I) sowie aus kortikalen Arealen (II–VI) erhalten. Sie sind efferent mit anderen kortikalen Arealen (II, III), subkortikalen Strukturen (V) und dem Thalamus (VI) verbunden. Vertikale und weitreichende horizontale kortikale Verbindungen ermöglichen eine zunehmende Extraktion definierter Merkmale von Sinnesreizen. Den Hauptteil der Hirnrinde belegen Assoziationsareale (Assoziationskortizes), die Wechselwirkungen zwischen den Arealen mit primär sensori- 25.3 Kognition versus Emotion – Das limbische System 25.3.1 Lobus limbicus und Papez-Kreis – Grundlagen des limbischen Systemkonzepts In einer bereits 1878 veröffentlichten Arbeit bemerkte der französische Neurologe Paul Broca, dass nahezu alle Säugetiere an der zur Mittellinie orientierten Oberfläche des Gehirns eine Gruppe von Arealen besitzen, die in einer ringförmigen Anordnung untereinander verbunden zu sein scheinen. An den lateinischen Ausdruck „Limbus“ (Rand, Ring) angelehnt nannte er diese Anordnung Lobus limbicus (Abb. 25.5 A). Wichtige Areale des Lobus limbicus sind der Gyrus cinguli und der Gyrus parahippocampalis mit dem Hippokampus. Broca entwickelte allerdings keinerlei Vorstellungen über die mögliche funktionelle Bedeutung dieser schen und motorischen Aufgaben vermitteln. Die Assoziationskortizes stellen demzufolge wichtige Module für integrative, so genannte höhere Hirnfunktionen dar. Die integrative Funktion der Assoziationskortizes zeigt sich an ihrem afferenten Hauptzustrom aus thalamischen Gebieten, die bereits verarbeitete Signale aus dem Kortex erhalten, sowie einem hohen Grad an kortiko-kortikalen Verbindungen. Eine lokale Funktionsstörung des Assoziationskortex hat bestimmte kognitive Defizite zur Folge. Parietale Schädigung führt häufig zu Aufmerksamkeitsdefiziten (z. B. Neglektsyndrom), temporale Schädigung hat Defizite im (Wieder-)erkennen (z. B. Agnosie) zur Folge, und frontale Schädigung resultiert in vielfältigen Defiziten der planend-vorausschauenden Handlung sowie Änderungen der Persönlichkeit Strukturen, und in der nachfolgenden Zeit wurde deren Funktion primär in Verbindung mit dem Geruchssinn vermutet. Ab etwa 1930 vermehrten sich jedoch Hinweise auf einen Zusammenhang zwischen den Strukturen des Lobus limbicus und emotionalem Verhalten. Zu dieser Zeit stellte der amerikanische Neurologe James Papez die Hypothese eines „Emotionssystems“ in Form eines Rückkopplungskreises, des so genannten „Papez-Kreises“, auf (Abb. 25.5 B). In diesem Modell fungiert ein definiertes Areal der Hirnrinde, der Gyrus cinguli, als das Areal für die Empfindung von Emotionen, und Rückkopplungen mit dem Hypothalamus vermitteln die funktionelle Verbindung der emotionalen Empfindung mit der körperlich-vegetativen und hormonell vermittelten emotionalen Reaktion, wie zum Beispiel der Änderung von Blutdruck und Herzfrequenz bei Schreckreaktionen. Die Rückkopplungsschleife im Papez-Modell beinhaltet efferente Fasern des Hippokampus, die über den Fornix das Corpus mamillare (eine Struktur des poste- Integrative Funktionen des Gehirns Organisation des Cortex cerebri 25 Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. ellen Agnosie, bei der ein Gesicht zwar als solches, jedoch nicht als das einer bekannten Person erkannt wird. Diese Patienten können zum Beispiel ihnen seit Jahren vertraute Personen, wie den Ehepartner, nicht anhand des Gesichts identifizieren. Bei Befragung geben sie an, das entsprechende Gesicht nicht zu kennen, und sie erinnern sich nicht an Eigenschaften der zugehörigen Person. Andererseits erkennen sie andere, ihnen bekannte Objekte wieder und können selbst subtile Unterschiede solcher Objekte differenzieren. Sie sind darüber hinaus in der Lage, bestimmte Personen mit Hilfe anderer Hinweise, wie zum Beispiel des Klangs der Stimme, der Körpergestalt oder des Geburtsdatums zu identifizieren. 821