17.2 Die Signalverarbeitung in der Retina
Werbung

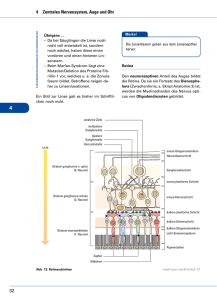
17 Visuelles System Die Signalverarbeitung in der Retina 323 erweiterung mit Mydriatika (Sympathomimetika 17.2.1 Überblick und Funktion oder Parasympatholytika) zur Beurteilung des Augen- Die Retina (Netzhaut) ist entwicklungsgeschichtlich hintergrunds möglichst verzichten. Da eine Pupillenverengung zur Erweiterung des Kammerwinkels und ein Teil des Diencephalons und besteht aus einem lichtempfindlichen Teil (Pars optica) sowie einem so zum besseren Abfluss des Kammerwassers führt, lichtunempfindlichen Teil (Pars caeca mit Pars iridica werden therapeutisch beim Glaukom Miotika verab- und Pars ciliaris) (Abb. 17.4.). Ihr Sinnesepithel bietet reicht (z. B. Parasympathomimetika wie Pilocarpin). die anatomische Voraussetzung für die Sinnesfunksetzung von elektromagnetischen Wellen in Memb- Die Tränenflüssigkeit wird in den Tränendrüsen (Gl. ranpotenziale, als auch die ersten neuronalen Verar- lacrimalis) als Ultrafiltrat des Blutes gebildet und durch den regelmäßigen Lidschlag über die Hornhaut beitungsprozesse der optischen Information. Die optischen Signale werden in den Photosensoren, den Stäbchen und Zapfen, erfasst. (Kornea) verteilt. Sie schützt die Kornea vor Austrocknung und schwemmt Staubteilchen und andere Substanzen weg. Zudem enthält sie sekretorisches IgA zur 17.2.2 Der Aufbau der Netzhaut Erregerabwehr. Die Tränenflüssigkeit ist leicht hyper- Bei Betrachtung der Schichten von außen nach innen ton (salziger Geschmack) mit einem höheren Kalium- ist die Netzhaut folgendermaßen aufgebaut: und niedrigeren Natriumgehalt als das Blutplasma. Pigmentzellschicht Fremdkörper im Auge reizen über mechano- und nozizeptive Endigungen in Hornhaut und Bindehaut Photosensoren (Zapfen und Stäbchen) Horizontalzellen den N. trigeminus (V. Hirnnerv), der so über das Ggl. Bipolarzellen pterygopalatinum und parasympathische Fasern die amakrine Zellen Tränensekretion steigert. Ganglienzellen (die Axone der Ganglienzellen bilden den N. opticus). ✔ ✔ Check-up Die Außenglieder der Photosensoren ragen in das Pig- Wiederholen Sie die Fehlsichtigkeiten, ihre Pathophysiologie und wie man sie korrigieren kann. Machen Sie sich nochmals klar, wie die Pupille auf Lichteinfall reagiert und durch welche Pharmaka eine Pupillenerweiterung erreicht werden kann. mentepithel, das die Innenseite des Bulbus auskleidet, hinein und werden ständig regeneriert (s. u.). Das Pigmentepithel hat u. a. die Aufgabe, die abgestoßenen, alten Membranscheibchen aus den Außengliedern zu phagozytieren. Bevor das Licht die Photosensoren erreicht, muss es zunächst mehrere Zellschichten (Horizontalzellen, Bipolarzellen, amakrine Zellen und Ganglienzellen) durchdringen. 17.2 Die Signalverarbeitung in der Retina Lerncoach Die Umsetzung eines optischen Reizes in ein elektrisches Signal (Aktionspotenzial) ist in der Retina mit einer Reihe komplizierter Sensorprozesse verbunden, deren Ablauf Sie sich Schritt für Schritt klarmachen sollten. Beachten Sie dabei, dass die Photosensoren des Auges anders als die meisten anderen Zellen auf einen adäquaten Reiz mit einer Hyperpolarisation reagieren. Die elektrische Information wird dem Lichteinfall entgegen weitergegeben: die Innenglieder der Photosensoren bilden Synapsen mit den Bipolarzellen, die wiederum mit den Ganglienzellen in Verbindung stehen. Dabei kommt es zu einer Konvergenz, d. h. viele Photosensoren verschalten auf mehrere Bipolarzellen, die ihre Information an noch weniger Ganglienzellen weitergeben. Außerdem bestehen auf horizontaler Ebene Querverknüpfungen über inhibitorische Neurone: die Horizontalzellen verschalten die Photosensoren untereinander, die amakrinen Zellen die Ganglienzellen. Auf diese Weise kann ein Teil der optischen Information bereits im Auge verarbeitet werden. Erst in den Axonen der Ganglienzellen entstehen Aktionspotenziale, wohingegen die übrigen retinalen 17 Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. tion des Sehens. In der Retina erfolgen sowohl die Um- 17.1.8 Die Tränenflüssigkeit 324 Die Signalverarbeitung in der Retina 17 Visuelles System ein Zilium miteinander verbunden. In den Stäbchenaußengliedern sind etwa 1000 geldrollenförmig angeordnete Membranscheibchen enthalten, bei den Zapfen findet man funktionell gleichwertige Einfaltungen der Zellmembran. In den Membranscheibchen befindet sich der Sehfarbstoff. Das Verhältnis von Zapfen : Stäbchen beträgt etwa 1 : 20, allerdings ist die Verteilung sehr unterschiedlich: in der Fovea centralis findet man ausschließlich Zapfen, im Randbereich der Netzhaut nur Stäbchen. Die etwa 6 Millionen Zapfen der Retina eines Auges befinden sich im Bereich der Fovea centralis und angrenzender zentraler Retinabezirke. Durch ihre enge Verschaltung mit den Ganglien-Zellen (in der Fovea centralis bis zu 1:1-Verschaltung) ermöglichen sie eine besonders gute Auflösung. Die Zapfen enthalten jeweils einen von drei möglichen Sehfarbstoffen (Zapfen-Opsine), deren Absorptionsmaximum für Licht in unterschiedlichen Wellenlängenbereichen liegt. Der eigentliche Farbeindruck ergibt sich aus der Summe der Erregung der drei Zapfensorten. Abb. 17.4 Aufbau der Retina (blau = Ganglienzelle mit ihrem rezeptiven Feld) (nach Lang) Die Stäbchen Die etwa 120 Millionen Stäbchen sind in der Netzhautperipherie lokalisiert (am dichtesten 30° rings 17 Neurone ihre Information elektrotonisch und über um die Fovea centralis). In der Fovea centralis selbst chemische Synapsen weitergeben (s. S. 233). Die gibt es keine Stäbchen. Stäbchen sind lichtempfindli- Axone der Ganglienzellen bilden die innerste Schicht cher als Zapfen und ermöglichen daher auch bei der Retina und verlassen den Bulbus gemeinsam an schwachem der Papilla n. optici, im Gesichtsfeld resultiert so der Sehen“), können aber keine Farben unterscheiden. Licht einen Seh-Eindruck („Nacht- „blinde Fleck“ (s. S. 333). Der Sehfarbstoff der Stäbchen ist das Rhodopsin, sein 17.2.2.1 Die Photosensoren Absorptionsmaximum liegt bei ca. 500 nm und damit zwischen dem der Blau- und Grün-Zapfen. Licht aus Die Retina besitzt zwei unterschiedliche Arten von diesem Wellenlängenbereich (Blau-Töne) wird daher Photosensoren: die Zapfen für das photopische beim skotopischen Sehen heller wahrgenommen als Sehen (Sehen bei Tageslicht, Farbensehen) und die Stäbchen für das skotopische Sehen (Nacht-Sehen, Schwarz-Weiß-Sehen). Ihnen gemeinsam ist das Vorhandensein von Sehpigmenten, deren chemische Konfiguration sich bei Lichteinfall in einer photochemischen Reaktion verändert und so den Signalprozess in Gang setzt (s. u.). Stäbchen und Zapfen bestehen aus den erneuerungsfähigen Außensegmenten und den permanenten Innensegmenten. Außen- und Innensegment sind über Licht anderer Wellenlängen (z. B. Rot-Töne) (Purkinje-Erscheinung). 17.2.3 Die Signaltransduktion in den Photosensoren Anders als die meisten anderen Körperzellen sind die Photosensoren unter Ruhebedingungen relativ stark depolarisiert (Ruhemembranpotenzial ca. –30 mV). Grund dafür ist die im Dunkeln verhältnismäßig hohe Na+- und Ca2+-Leitfähigkeit, die durch geöffnete, Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. Die Zapfen cGMP-abhängige Na+-Ca2+-Kanäle bedingt ist und das Meta-Rhodopsin II stimuliert seinerseits das G-Pro- Membranpotenzial in Richtung Na+- und Ca2+-Gleich- tein Transducin. Transducin aktiviert eine Phospho- gewichtspotenzial verschiebt (vgl. S. 10). Ein Lichtreiz führt über das Schließen dieser Kanäle diesterase, die cGMP zu GMP hydrolysiert. Die Abnahme der cGMP-Konzentration bewirkt eine Schlie- zu einer Hyperpolarisation, dadurch verringert sich ßung der cGMP-abhängigen Kationenkanäle. Die da- die Freisetzung von Transmitter (Glutamat) an den Sy- raus resultierende Abnahme der Na+- und Ca2+-Leit- napsen und in den nachgeschalteten Bipolar- und Ho- fähigkeit führt zu einer Hyperpolarisation der Stäb- rizontalzellen kommt es zu einer Potenzialänderung. chenzelle, weil nun, wie in anderen Körperzellen, Die Ganglienzellen setzen schließlich das Sensorpo- der Einfluss der K+-Leitfähigkeit auf das Membranpo- tenzial in eine entsprechende AP-Frequenz um, tenzial überwiegt. Die Hyperpolarisation der Zelle deren Höhe mit dem Anstieg der Hyperpolarisation korreliert. hemmt die Ausschüttung des Transmitters Glutamat an die Bipolarzellen und beeinflusst so die Potenziale der nachgeschalteten Neurone. MERKE Beendet wird diese Lichtreaktion durch die Neusyn- Anders als die meisten anderen erregbaren Zellen reagieren Photosensoren auf einen adäquaten Reiz mit einer Hyperpolarisation (nicht mit einer Depolarisation). these von cGMP: durch die Schließung der Na+-Ca2 -Kanäle sinkt der intrazelluläre Ca2+-Gehalt. Dadurch + wird eine Ca2+-empfindliche Guanylatzyklase enthemmt, die nun verstärkt cGMP produziert. Durch Die Außenglieder der Stäbchen enthalten Scheibchen das cGMP können die Na+-Ca2+-Kanäle wieder geöffnet werden, der Na+-Ca2+-Dunkelstrom setzt wieder mit dem Sehfarbstoff Rhodopsin, der aus Opsin (Pro- ein und die Zelle depolarisiert, bis sie ihr normales teinkomponente) Dunkelpotenzial wieder erreicht hat. und 11-cis-Retinal (Vit.-A-Ab- kömmling) besteht. Die durch einen Lichtreiz ausge- Das All-trans-Retinal wandert in das Pigmentepithel löste photochemische Reaktion mit Isomerisierung und wird dort enzymatisch in mehreren Schritten des 11-cis-Retinals setzt die Signalkaskade in Gang: wieder zu 11-cis-Retinal umgewandelt. Anschließend Die Belichtung des Sehfarbstoffs führt zu einer Kon- gelangt es zurück in die Sensorzelle und verbindet formationsänderung des 11-cis-Retinals. Durch Umlagerung einer Molekülbindung entsteht das gestreckte sich dort mit Opsin erneut zu Rhodopsin. Die Signaltransduktion in den Zapfen verläuft ähnlich Molekül All-trans-Retinal und das Rhodopsin wandelt wie in den Stäbchen, statt Rhodopsin kommen jedoch sich über mehrere schnelle Zwischenschritte in Meta- die Zapfen-Opsine mit ihrer unterschiedlichen spekt- Rhodopsin II um (Abb. 17.5). ralen Empfindlichkeit zum Einsatz (s. o.). Abb. 17.5 Signaltransduktion in den Photorezeptoren (nach Silbernagl/Despopoulos) 17 Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. 17 Visuelles System Die Signalverarbeitung in der Retina 325 17.2.4 Die neuronalen Verarbeitungsprozesse in der Retina 17.2.4.1 Die Ganglienzellen Die Retina eines Auges besitzt etwa 130 Millionen Photosensor-Zellen, aber nur ca. 1 Million Ganglienzellen. Die Information muss also schon auf retinaler Ebene gebündelt werden. Dies geschieht, indem viele Sensorzellen auf mehrere Bipolarzellen verschalten, die ihre Signale wiederum an eine Ganglienzelle weitergeben. Die Information aus einem Netzhautareal mit mehreren Sensoren läuft also in einer Ganglienzelle zusammen (Signalkonvergenz). Auch eine Signaldivergenz findet in der Netzhaut statt, wenn eine Bipolarzelle nicht nur mit einer, sondern mit mehreren Ganglienzellen Synapsen bildet. Insgesamt überwiegt jedoch bei weitem die Konvergenz. Die Ganglienzelltypen empfindlichkeit im entsprechenden Bereich. Ein kleines rezeptives Feld hat den Vorteil, dass der Lichteinfall genau lokalisiert werden kann: wäre das Licht nur ein klein wenig mehr nach rechts oder links gefallen, hätte es bereits ein anderes rezeptives Feld getroffen und eine andere Ganglienzelle erregt oder gehemmt. Dagegen verschalten bei einem großen rezeptiven Feld, das viele Sensoren umfasst, auch die Sensoren rechts und links noch auf dieselbe Ganglienzelle, so dass man anhand der Potenzialänderung dieser Ganglienzelle das beleuchtete Gebiet nicht so genau bestimmen kann. Bei der Lichtempfindlichkeit verhält es sich umgekehrt: Während bei schwachem Licht die Erregung einiger weniger Sensorzellen nicht ausreicht, um eine Potenzialänderung in der Ganglienzelle zu erreichen, ist durch die Summation vieler, auch schwacher Reize, doch noch eine Potenzialänderung der Ganglienzelle möglich. Die Retina enthält drei Ganglienzelltypen zur Erfassung des visuellen Gesamteindrucks. Die großen α- MERKE (Y-)Zellen (magnozelluläres System) machen etwa net sind. Bei den zugehörigen Photosensoren handelt Kleine rezeptive Felder haben ein gutes Auflösungsvermögen auf Kosten der Lichtempfindlichkeit, große rezeptive Felder dagegen eine gute Lichtempfindlichkeit, dafür aber ein schlechtes Auflösungsvermögen. es sich hauptsächlich um Stäbchen, sie können daher keinen Farbeindruck vermitteln. Aus diesem Grund findet man auch dort, wo eine gute Ca. 80 % der retinalen Ganglienzellen sind kleine Auflösung besonders wichtig ist (Fovea centralis), β-(X-)Zellen (parvozelluläres System), die die Auf- sehr kleine rezeptive Felder (bis zu 1:1-Verschaltung gabe haben, Farbe und Gestalt wahrzunehmen. Sie zwischen Sensor- und Ganglienzelle!) und dort, wo besitzen kleine rezeptive Felder und dünnere Axone, insbesondere eine hohe Lichtempfindlichkeit erreicht die Leitungsgeschwindigkeit ist entsprechend etwas werden soll (Peripherie), große rezeptive Felder. 10 % der retinalen Ganglienzellen aus. Sie besitzen schnell leitende Axone, die besonders zur Erfassung von Bewegung und Entfernung von Objekten geeig- 17 geringer. Die γ-(W-)Zellen (koniozelluläres System) dienen der Steuerung der Pupillomotorik und reflektorischer On-/Off-Zentrums-Neurone Augenbewegungen. Während die α- und β-Zellen in larisation oder eine Hyperpolarisation der zugehöri- die primäre Sehrinde projizieren, ziehen die dünnen, gen Ganglienzelle zur Folge haben. Rezeptive Felder markarmen Axone der γ-Zellen ins Mittelhirn. Die Belichtung eines Netzhautareals kann eine Depo- haben eine kreisförmige Gestalt, dabei liegen einige Sensoren im Zentrum, andere am Rand dieser Fläche. 17.2.4.2 Die rezeptiven Felder V. a. durch die laterale Verschaltung über die Horizon- Als rezeptives Feld eines visuellen Neurons bezeich- talzellen kommt es zu einer gegensätzlichen Reaktion net man den Netzhautbereich, dessen Reizung zu einer Aktivitätsänderung dieses Neurons führt. Für der Ganglienzelle, je nachdem, ob nur das Zentrum oder die Peripherie beleuchtet wird: eine Ganglienzelle z. B. umfasst dieser Bereich die Ge- On-Zentrums-Neurone: Werden die Photorezeptoren im Zentrum der On-Zentrums-Ganglienzelle beleuchtet, geht sie in den „On-Zustand“ über, d. h. die Ganglienzelle depolarisiert und er- samtheit aller Photosensoren, die auf eine Ganglienzelle verschalten. Die Größe der rezeptiven Felder hat Einfluss auf das Auflösungsvermögen und die Licht- Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. 326 Die Signalverarbeitung in der Retina 17 Visuelles System 17 Visuelles System Die Signalverarbeitung in der Retina 327 quenz). Die Belichtung der Photorezeptoren der 17.2.4.3 Die Anpassung des Auges an unterschiedliche Lichtverhältnisse Peripherie hat den gegenteiligen Effekt, es kommt zu einer Hyperpolarisation mit Abnahme Das Auge ist in der Lage, sich an Lichtreize von ganz unterschiedlicher Leuchtdichte anzupassen und sie der AP-Frequenz. Wird das gesamte rezeptive wahrzunehmen. Die Anpassung des Auges an die je- ihre Aktionspotenzialfrequenz (AP-Fre- Feld belichtet, so kommt es ebenfalls zu einer Erhö- weilige Leuchtdichte nennt man Adaptation. Dieser hung der AP-Frequenz, allerdings fällt diese gerin- Vorgang nimmt etwas Zeit in Anspruch: kommt ger aus als bei alleiniger Belichtung des Zentrums. man beispielsweise aus dem Sonnenlicht in ein abge- Off-Zentrums-Neurone: Die Off-Zentrums-Neurone reagieren genau umgekehrt. Die AP-Frequenz der Ganglienzelle nimmt bei Belichtung des Zentrums ab („Off-Zustand“), bei Belichtung der Peripherie zu. Die antagonistische Reaktion von Zentrum und Peripherie hat eine Verstärkung der Kontrastwahrnehmung zur Folge, weil von den rezeptiven Feldern, die an der Grenzfläche zwischen hell und dunkel liegen, nur Teile belichtet werden. Möglich ist beispielsweise, dass die Peripherie noch belichtet wird, während das Zentrum bereits im Dunklen liegt. Die Ganglienzelle reagiert aber gerade auf eine unterschiedliche Belichtung von Zentrum und Peripherie besonders stark. Durch die laterale Hemmung (Hemmung benachbarter Elemente) erscheint so die dunkle Seite noch dunkler, die helle noch heller. Aus diesem Grund wirkt beispielsweise eine graue Fläche vor schwarzem Hintergrund heller als vor weißem, dieses Phänomen nennt man Simultankontrast (Abb. 17.6). Bei Dunkeladaptation vergrößert sich das Zentrum des rezeptiven Feldes auf Kosten der Peripherie, dadurch nimmt die Kontrastverstärkung deutlich ab, bis sie schließlich ganz aufgehoben wird. On-Off-Ganglienzellen reagieren sowohl auf Lichtreize als auch auf Verdunklung mit kurzen AP-Frequenzsteigerungen. Aus diesem Grund eignen sie sich besonders gut zur Wahrnehmung von Bewegungen, bei denen sich Hell-Dunkel-Reize schnell über die rezeptiven Felder bewegen. dunkeltes Zimmer, erscheint zunächst alles schwarz. Abb. 17.6 Simultankontrast Nach einiger Zeit gewöhnt sich das Auge jedoch an die schwächere Lichtintensität und Einzelheiten sind erkennbar, die Reizschwelle hat sich gesenkt. Auch die Flimmer-Fusionsfrequenz (zeitliches Auflösungsvermögen), d. h. die Anzahl von Bildern pro Sekunde, ab der sie nicht mehr als getrennte Bilder, sondern als Bewegungsablauf wahrgenommen werden, sinkt dabei von 65–90/sec beim photopischen Sehen auf 20–25/sec beim skotopischen Sehen ab. Den zeitlichen Verlauf der Dunkel-Adaptation zeigt Abb. 17.7. Die Kurve gibt für jeden Zeitpunkt der Adaptation an, welche Lichtintensität das Auge gerade noch wahrnehmen kann. Zu Beginn benötigt das helladaptierte Auge hohe Lichtintensitäten, um erregt zu werden. Je länger man ihm jedoch Zeit gibt, sich an die Dunkelheit zu adaptieren, desto niedriger muss die Leuchtdichte sein, um noch eine Empfindung auszulösen. Die Adaptationskurve weist in ihrem Verlauf einen Knick auf, den sog. Kohlrausch-Knick. Er kommt da- Abb. 17.7 Adaptation von Stäbchen und Zapfen bei unterschiedlichen Lichtverhältnissen (nach Silbernagl/Despopoulos) 17 Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. höht durch zustande, dass das Sehen ab einer gewissen sinkonzentration und damit die Lichtempfindlich- Lichtintensität von Zapfen- auf Stäbchen-Sehen übergeht. Auf Höhe des Kohlrausch-Knicks ist die minimale Schwelle der Zapfen erreicht, d. h. noch schwä- keit ansteigt. Die Organisation der rezeptiven Felder kann in gewissem Umfang an die Lichtverhältnisse angepasst chere Lichtintensitäten führen zu keiner Erregung werden. Bei abnehmender Beleuchtungsstärke der Zapfen mehr, sondern können nur noch von den nimmt auch die laterale Hemmung ab, was zu Stäbchen registriert werden. Der weitere Kurvenver- einer Ausdehnung der Zentren der rezeptiven lauf wird daher von den lichtempfindlicheren Stäbchen bestimmt. Felder auf Kosten der Peripherie führt (räumliche Summation) : je größer das Netzhautareal ist, Die unterschiedliche Anordnung von Zapfen und durch das eine Ganglienzelle erregt wird, desto Stäbchen hat zur Folge, dass bei Dunkelheit nicht mehr richtig fixiert werden kann: der Punkt schärfs- höher ist auch die Lichtempfindlichkeit. Unterschwellige Reize können, wenn sie länger an- ten Sehens liegt in der ausschließlich Zapfen enthal- dauern, doch noch überschwellig werden und ein tenden Fovea centralis. Zapfen können aber durch Aktionspotenzial in der Ganglienzelle auslösen nur schwach leuchtende Objekte (z. B. einen kleinen (zeitliche Summation). Diese Reizverlängerung Stern am Nachthimmel) nicht erregt werden. Ver- kann durch „längeres Hinschauen“ erreicht wer- sucht man also dieses Objekt zu fixieren, ist es nicht 17 den. zu sehen. Blickt man dagegen an dem Objekt Eine lokale Adaptation der Netzhaut lässt sich beim vorbei, fällt sein Bild auf den Randbereich der Retina, wo es durch die Stäbchen wahrgenommen werden sog. Sukzessivkontrast beobachten. Blickt man einige Zeit auf ein schwarz-weißes Muster und danach auf kann. eine weiße Fläche, sieht man ein umgekehrtes Nach- Bei der Nachtblindheit (Nyktalopie) z. B. infolge eines Vitamin A-Mangels sind die Stäbchen in bild: die vormals dunklen Anteile erscheinen heller, ihrer Funktion gestört, die Dunkeladaptationskurve tritt dieses Phänomen auf, die Nachbilder entstehen verläuft also als reine Zapfen-Kurve. Sinkt die Licht- jeweils in der Komplementärfarbe. Ursache für die intensität unter die Schwellenintensität der Zapfen, Nachbilder ist eine kurzfristige Anpassung der Emp- kann dann kein visueller Eindruck mehr entstehen. Folgende Mechanismen sind neben dem Wechsel findlichkeit der jeweiligen Netzhautareale: primär hell belichtete Netzhautareale werden dadurch un- zwischen Zapfen- und Stäbchensehen für die Hell- empfindlicher, nicht belichtete dagegen empfind- Dunkel-Adaptation verantwortlich: Die Pupille kann die Menge des einfallenden Lichts licher. die vormals hellen dunkler. Auch beim Farbensehen durch ihre Weite kontrollieren. Bei Weitstellung 17.2.4.4 Die Visusbestimmung gelangt mehr, bei Engstellung weniger Licht ins Der Visus (Sehschärfe) ist ein Maß für das räumliche Auge, daher ist die Pupille bei Dunkelheit weiter Auflösungsvermögen des Auges, also die Fähigkeit, zwei Punkte gerade noch getrennt wahrnehmen zu können. Der Visus wird in 1/α (α in Winkelminuten [= 1/60°]) angegeben und mit Hilfe von Sehprobetafeln bestimmt. Als Testobjekt werden neben Bildoder Buchstabentafeln häufig sog. Landolt-Ringe verwendet: es handelt sich dabei um unterschiedlich große Ringe mit einer Lücke, deren Lage erkannt werden muss. Je nach Größe und Entfernung entspricht die Breite der Aussparung einer oder mehreren Winkelminuten. Ein normalsichtiger Proband kann bei guten Lichtverhältnissen noch eine Lücke der Breite 1 Winkelminute wahrnehmen, sein Visus ist damit V = 1. Muss die Lücke doppelt so breit sein, damit er sie noch wahrnehmen kann, beträgt der Visus V = 0,5. als bei Helligkeit. Die Pupillenreaktion ermöglicht so eine schnelle Anpassung an einen plötzlichen Helligkeitswechsel. Durch die Änderung der Rhodopsinkonzentra- tion ist die chemische Empfindlichkeitsanpassung der Sensoren an die Lichtverhältnisse möglich, denn die Lichtempfindlichkeit der Stäbchen hängt von der Verfügbarkeit des Sehfarbstoffs ab. Fällt viel Licht ins Auge, zerfällt ein großer Teil des Rhodopsins und steht für den Transduktionsprozess nicht mehr zu Verfügung, die Lichtempfindlichkeit nimmt ab. Bei schwachen Lichtverhältnissen wird Rhodopsin regeneriert ohne gleich wieder verbraucht zu werden, so dass die Rhodop- Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. 328 Die Signalverarbeitung in der Retina 17 Visuelles System 17 Visuelles System Die Signalverarbeitung in der Retina 329 Das optimale Auflösungsvermögen wird bei guten Farbmischung : die rote Farbe erscheint rot, weil Lichtverhältnissen in der Fovea centralis, erreicht. sie alle Wellenlängen außer den roten absorbiert, Der Visus bezieht sich auf diesen „Ort des schärfsten Sehens“. In der Netzhautperipherie oder bei Dunkel- die grüne grün, weil sie alle Wellenlängen außer den grünen absorbiert. Mischt man nun beide Far- adaptation nimmt der Visus ab. ben, bleiben nur die Wellenlängen übrig, die weder von der grünen noch von der roten Farbe absorbiert werden, das Spektrum ist also nach dem Mischen kleiner als vorher. Die Wellenlänge des sichtbaren Lichts liegt etwa zwi- Die Fähigkeit, Farben unterscheiden zu können, ist auf schen 400 (blauviolett) und 750 nm (rot). Werden Wellenlängen des gesamten Spektrums gemischt, entsteht der Farbeindruck „weiß“. Ein blauer Gegen- Retina-Ebene an die unterschiedliche spektrale Empfindlichkeit der Zapfen-Opsine gebunden, d. h. in Abhängigkeit von seiner Wellenlänge wird Licht von den stand erscheint uns dagegen blau, weil er alle Wellen- verschiedenen Zapfentypen unterschiedlich stark ab- längen außer den blauen absorbiert. Die „übrig ge- sorbiert und führt dementsprechend auch zu unter- bliebenen“ blauen Wellenlängen werden reflektiert schiedlich starken Sensorpotenzialen (trichromati- und können nun im Auge den Farbeindruck „blau“ sche Therorie des Farbensehens). Das Pigment der vermitteln. Blau-(K-)Zapfen absorbiert kurzwelliges, blauviolet- Folgende zwei Formen von Farbmischungen werden tes Licht (Maximum bei ca. 420 nm), Grün-(M-)Zap- unterschieden: Mischt man Licht unterschiedlicher Wellenlänge fen absorbieren mittelwelliges, blaugrünes bis gelbes Licht (Maximum bei ca. 535 nm) und Rot-(L-)Zapfen (z. B. Farbscheinwerfer im Theater), so entsteht langwelliges, gelbes bis rotes Licht (Maximum bei ca. ein neuer Farbeindruck (Bsp.: rotes Licht + grünes 565 nm). Licht => gelbes Licht). Man spricht hier von additi- Der eigentliche Farbeindruck ist das Ergebnis der Ana- ver Farbmischung, weil zu den roten Wellenlängen grüne Wellenlängen hinzukommen, das Spektrum also nach der Mischung größer ist als vorher. Wenn man dagegen Farben mischt (z. B. in der Malerei), entsteht ebenfalls ein neuer, aber anderer Farbton (Bsp.: rote Farbe + grüne Farbe => braune Farbe). Hierbei handelt es sich um eine subtraktive lyse, in welchem Verhältnis zueinander die verschiedenen Zapfen erregt worden sind. Licht der Wellenlänge 450 nm führt beispielsweise zu einer starken Erregung der Blau-Zapfen, einer geringeren Erregung der Grün-Zapfen und nur noch zu einer sehr schwachen Erregung der Rot-Zapfen (Abb. 17.8). Eine Verbesserung des Farbunterscheidungsvermögens (Farbkontrast) wird auf der Stufe der neuronalen Abb. 17.8 Trichromatisches Farbensehen (Bsp.: 450 nm) (nach Klinke/Silbernagl) 17 Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. 17.2.5 Die retinalen Mechanismen des Farbensehens 330 Die Informationsverarbeitung in der Sehbahn 17 Visuelles System Check-up Signalverarbeitung durch die Verschaltung farbantagonistisch organisierter rezeptiver Felder (Rot-Grün, ✔ Blau-Gelb, Schwarz-Weiß) erreicht (Gegenfarbentheorie). Dabei verarbeiten die sog. Gegenfarbenneurone Rezeptorsignale im Zentrum und in der Periphe- ✔ rie ihres jeweiligen rezeptiven Feldes antagonistisch. Diese Gegenfarbenneurone findet man auch im CGL. ✔ Wiederholen Sie den mikroskopischen Aufbau der Retina und die Funktion der einzelnen Sensortypen. Machen Sie sich klar, warum man einen grünen Ball grün sieht. Verdeutlichen Sie sich nochmals die Signaltransduktion in den Photosensoren. Ist eine Zapfensorte in ihrer Funktion gestört oder 17.3 Die Informationsverarbeitung in der Sehbahn Tritanomalie). Ein kompletter Ausfall einer Zapfen- Lerncoach sorte führt zur Protanopie (Rotblindheit), Deuterano- Um sich den Verlauf der Sehbahn klarzumachen, sollten Sie sie auch einmal selbst aufzeichnen. So lassen sich auch die charakteristischen Gesichtsfeldausfälle bei Läsionen der Sehbahn leicht ableiten. Denken Sie dabei daran, dass durch die optische Brechung das Gesichtsfeld umgekehrt auf der Retina abgebildet wird: das temporale Gesichtsfeld wird also auf der nasalen Retinahälfte abgebildet und umgekehrt. pie (Grünblindheit) oder Tritanopie (Blauviolettblindheit). Da die Farbwahrnehmung dann nur noch über zwei Sehpigmente stattfindet, spricht man auch von einer dichromaten Störung. Störungen des Farbensinnes sind häufig (ca. 9 % der Männer und 0,5 % der Frauen). Die Gene für das Zapfenpigment der mittelund langwelligen Zapfen befinden sich auf dem XChromosom, deshalb sind Männer wesentlich häufiger von Störungen des Rot-Grün-Sehens betroffen als Frauen. 17 Ein kompletter Ausfall aller Zapfen führt dagegen 17.3.1 Überblick und Funktion nicht nur zu einem fehlenden Farbempfinden, son- Über die Sehbahn erreicht die in der Retina erfasste dern auch zu einer deutlich herabgesetzten Sehschärfe und erhöhten Blendungsempfindlichkeit. und verarbeitete Information das ZNS, wobei die zentrale Signalverarbeitung der optischen Information Zur Diagnostik eines gestörten Farbensehens verwen- sehr komplex ist. Die verschiedenen Teilinformatio- det man ein sog. Anomaloskop oder Farbtafeln (z. B. nen des optischen Eindrucks (Form, Farbe, Bewegung) Ishihara-Tafeln). werden getrennt durch spezialisierte Neurone verarbeitet. Klinischer Bezug Retinopathia (Retinitis) pigmentosa: Eine Funktionsstörung des Pigmentepithels beider Augen mit Verlust der Photosensoren wird als Retinitis pigmentosa bezeichnet. Die Netzhautdegeneration schreitet von der Peripherie zum Zentrum fort. Symptome sind ein progredienter (sich verschlechternder) Visusverlust, progrediente Gesichtsfelddefekte und Nachtblindheit. Bei der Beurteilung der Netzhaut (Ophthalmoskopie) zeigt sich eine knochenbälkchenartige Hyperpigmentierung der mittleren und äußeren Netzhautperipherie. Die Erkrankung kann vererbt werden oder im Rahmen einer generalisierten Stoffwechselerkrankung auftreten. Die zusätzlich frühzeitig auftretende Katarakt (Linsentrübung) kann man zur Verbesserung des Visus operativ behandeln. Andere Therapiemöglichkeiten gibt es derzeit nicht. 17.3.2 Der Verlauf der Sehbahn Die Information der Photosensoren (1. Neuron) wird über die Bipolarzellen (2. Neuron) an die Ganglienzellen (3. Neuron) vermittelt, deren Axone gemeinsam als N. opticus das Auge verlassen. Im Chiasma opti- cum kreuzen die aus der nasalen Retinahälfte stammenden Nervenfasern auf die Gegenseite, während die Fasern der temporalen Retinahälfte weiterhin ungekreuzt verlaufen. Zusammen mit den gekreuzten Fasern aus der kontralateralen, medialen Retinahälfte bilden sie den Tractus opticus und ziehen zum Corpus geniculatum laterale (CGL), einem Teil des Thalamus. Der Tractus opticus enthält also jeweils die Informationen aus dem kontralateralen Gesichtsfeld. Im CGL (4. Neuron) erfolgt eine erneute Umschaltung. Heruntergeladen von: Thieme E-Books & E-Journals. Urheberrechtlich geschützt. ganz ausgefallen, kommt es zu Störungen der Farbwahrnehmung. Eine Farbschwäche bezeichnet man als Farbanomalie (Protanomalie, Deuteranomalie,