47 2.8.3 Stoffwechsel der Lipoproteine - Medi
Werbung

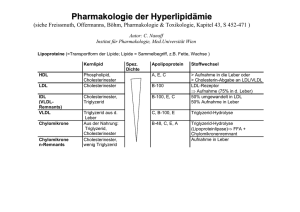
2.8.3 Stoffwechsel der Lipoproteine Kommen wir jetzt zu den VLDL, die in der Leber synthetisiert werden: VLDL bestehen sowohl aus endogen synthetisierten als auch aus − mit der Nahrung aufgenommenen − exogenen Lipiden. Das Verhalten der VLDL im Blut ähnelt dem der Chylomikronen: VLDL verfügen ebenfalls über das Apo CII, durch das die Lipoproteinlipase der Zellen aktiviert wird. Durch die Abgabe ihrer Triacylglycerine schrumpfen auch die VLDL, nur bildet sich dadurch IDL. Dieses Lipoprotein schlägt zwei verschiedene Wege ein. –– EinTeil des IDL gelangt zurück zur Leber und wird durch sie aufgenommen. –– Die restlichen IDL werden im peripheren Blut zu LDL umgewandelt. Die Umformung der IDL zu LDL erfolgt durch die Abspaltung aller Apolipoproteine des IDL bis auf das Apo B100. Und schon sind wir beim nächsten Kandidaten, dem „bösen“ LDL: Das aus IDL gebildete LDL ist sehr cholesterinreich und dient daher der Versorgung peripherer Gewebe mit Cholesterin. Hierbei wird LDL durch rezeptorvermittelte Endozytose in die Zielzellen aufgenommen. Als Ligand für den membrangebundenen LDLRezeptor dient sein Apo B100. Nach Bindung von Apolipoprotein B100 an den Rezeptor kommt es zur Einstülpung des der Zellmembran. Die gebundenen Rezeptoren befinden sich vor allem in besonderen Stellen der Zellmembran (sog. „pits“ = Gruben). Diese Gruben sind mit dem Protein Clathrin ummantelt (sog. „clathrin-coated pits“), das eine korbartige Struktur um die sich einstülpende Zellmembran bildet. Das Happy End bildet das „gute“ HDL: Die HDL werden in Leber und Darm synthetisiert und enthalten als Lipide v. a. Phosphatidylcholin. Von allen Lipoproteinen des Blutplasmas haben sie den höchsten Gewichtsanteil an Apolipoproteinen. HDL dienen dem Transport von Cholesterin aus den extrahepatischen Geweben zur Leber, da nur hier die Cholesterinausscheidung bzw. Cholesterinmetabolisierung zu Gallensäuren möglich ist. Im Plasma binden die HDL das in der Leber synthetisierte und ans Blut abgegebene Enzym Lecithin-Cholesterin-Acyltrans- www.medi-learn.de ferase (LCAT). Die LCAT wird durch das Apolipoprotein A1 der HDL aktiviert und katalysiert die Acylierung von Cholesterin: Cholesterin + Phosphatidylcholin g Cholesterinester + Lysophosphatidylcholin Bei dieser Reaktion wird eine Fettsäure des Phosphatidylcholins genutzt, um mit dem aufgenommenen Cholesterin einen Cholesterinester zu bilden. Das nun um eine Fettsäure ärmere Phosphatidylcholin wird als Lysophosphatidylcholin bezeichnet. Durch diese Reaktion nimmt der Gehalt der HDL an Cholesterin zu, während ihr Gehalt an Phosphoglycerinen abnimmt, da das gebildete Lysophosphatidylcholin abdiffundiert. Das Cholesterin nimmt somit den Platz der Phosphoglycerine ein. Cholesterinreiches HDL gelangt zurück zur Leber, wo das Cholesterin endgültig ausgeschieden bzw. metabolisiert wird. Lipoproteine spielen eine wichtige Rolle bei der Pathogenese der Arteriosklerose. Dabei gilt ein hoher LDL-Spiegel als Risikofaktor für die Entstehung und das Fortschreiten dieser Erkrankung (Abgabe von Cholesterin an die Blutgefäße), während ein hoher HDL-Spiegel diesbezüglich protektiv wirken soll (Einsammeln von Cholesterin und Transport zur Leber). Aus diesem Grund wird LDL auch als „böses“ und HDL als „gutes“ Cholesterin bezeichnet. Zur Abschätzung des kardiovaskulären Risikoprofils wird neben der Bestimmung der Einzelwerte auch der LDL/HDL-Quotient berechnet: Ein LDL/HDL-Quotient unter drei gilt als günstig, während ein LDL/HDL-Quotient über vier einen Risikofaktor darstellt. 2 Übrigens … Der LDL-Rezeptor dient der Aufnahme cholesterinreicher LDL-Partikel und spielt somit eine wichtige Rolle für den Cholesterinspiegel im Blut. Der LDLRezeptor wird im endoplasmatischen Retikulum und Golgi-Apparat synthetisiert. Sinken die Cholesterinanteile in der Membran des ER, so führt dies zu einer verstärkten Transkription des LDL-Gens mit dem Ziel, vermehrt Cholesterin in die Zelle aufzunehmen. 47