Fortpflanzungsorgane, weiblich
Werbung

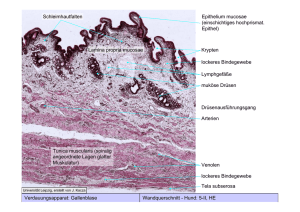
Fortpflanzungsorgane, weiblich 119) Ovar mit Corpus luteum, Katze, Zenker, HE, 8 μm -Oberfläche wird bedeckt von einem einschichtigen platten bis kubischen Epithel mit Mikrovilli -> Peritonealepithel. Tunica albuginea: straffes, dichtes Bindegewebe unter dem Oberflächenepithel, das kontinuierlich in das Stroma des Ovars übergeht. Es gliedert sich in einen Cortex ovarii (Rinde) und eine Medulla ovarii (Mark), die beide aus zellreichem Bindegewebe (mit interstitiellen Zellen) bestehen. Stroma ovarii, dichtes spinozelluläres Bindegewebe und in der Rinde außerdem Folliculi ovarii (Ovarialfollikel), die die heranreifenden Eizellen, Oozyten enthalten. Das Bindegewebe im Mark ist locker und enthält zahlreiche Gefäße und Nerven, die durch das Hilum ovarii ins Ovar eintreten, dort, wo ein kurzes Mesovar am Eierstock ansetzt. In der Rinde befinden sich die Follikel in ihren verschiedenen Stadien. 1.Primordialfollikel: -Dicht unter der Kapsel gelegen und besteht aus einem Oozyt, der von einer Schicht flacher Follikelepithelzellen umgeben ist. Es hat einen großen, exzentrisch gelegenen Kern mit großem Nukleolus 2.Primärfollikel: -der Oozyt hat sich vergrößert, das Follikelepithel ist einschichtig kubisch bis hochprismatisch geworden und setzt sich etwas vom Oozyt ab -> Es bildet sich ein Spaltraum, in den sich amorphe Grundsubstanz einzulagern beginnt 3.Sekundärfollikel: -deutlich größer als Primärfollikel, liegen in verschiedenen Entwicklungsstadien vor. Es kommt zur: Vergrößerung des Oozyt, Ausbildung einer Zona pellucida, Vermehrung des Follikelepithels (Granulosazellen, die an der Synthese von Steroiden (Östrogene, Progesteron) beteiligt sind) und Entstehung einer Theca folliculi externa und interna vom Follikelepithel durch Basallamina (Glashaut) getrennt 4.Tertiärfollikel, Bläschenfollikel: -die Schichtenzahl der Follikelepithelzellen hat weiter zugenommen, flüssigkeitsgefüllte Räume zwischen den Follikelepithelzellen sind im LM sichtbar. Die Theca folliculi gliedert sich in Theca folliculi interna und Theca folliculi externa. Der Oozyt wächst weiter, verharrt jedoch unverändert im Diktyotänstadium. Antrum folliculi: immer mehr und weitere vergrößerte Spalträume, die mit fortschreitendem Follikelwachstum entstehen, zusammenfließen und schließlich zu einer gemeinsamen großen Höhle werden. Liquor folliculi: hyluronsäurereiche Flüssigkeit, die Spalträume und Antrum folliculi füllt. Cumulus oophorus: entsteht dadurch, daß durch weitere Flüssigkeitszunahme das Follikelepithel zu einer mehrschichtigen randständigen Zellage wird, die an einer Stelle einen exzentrisch gelegenen, in die Follikelhöhle vorgewölbten, kleinen Zellhügel, Cumulus oophorus, bildet, der den Oozyt enthält. Die Follikelepithelzellen in unmittelbarer Nähe des Oozyt sind hochprismatisch, im übrigen aber verzweigt und flach 4.1.präovulatorische Follikel, reife Follikel, Graaf Follikel -dieses Endstadium wird nur von wenigen Primordialfollikeln erreicht. Wölben die Oberfläche des Ovars vor und erscheinen als durchsichtiges Bläschen. Die Follikelepithelschicht wird niedriger, erhalten bleibt jedoch ein dichter mehrschichtiger Kranz von hochprismatischen Follikelzellen um den Oozyt (Corona radiata). Zur Zeit der Ovulation befindet sich der Oozyt mit der Corona radiata frei in der Follikelhöhle. Bei der Ovulation kommt es zur Ruptur des präovulatorischen Follikels, die der das Ovum freigesetzt und von der Tuba uterina aufgefangen wir Follikelatresie: ihr unterliegen die meisten Primordial- und Primärfollikel, kann aber jeden Follikeltyp ergreifen Corpus luteum: -Hervorgegangen aus dem Corpus rubrum, einer leeren Follikelhöhle, die mit Blut gefüllt war. Gelbkörper, eine temporäre endokrine Drüse, die nach der Ovulation von den Follikelepithelzellen, die im Ovar verbleiben und den Zellen der Theca interna gebildet wird. Liegt in der Rinde des Ovars und sezerniert von den im Ovar synthetisierten Steroidhormonen hauptsächlich Progesteron, aber auch Östrogene. Die Follikelepithelzellen und die Zellen der Theca interna wandeln sich postovulatorisch um in: 1.Granulosaluteinzellen: leiten sich vom Follikelepithel ab, ihr Volumen nimmt erheblich zu und sie bekommen den Feinbau von Steroidhormon bildenden Zellen. In Ihren Zytoplasma treten außerdem zahlreich Fetttropfen auf -> vakuoliges (wabiges) Aussehen. Das Zytoplasma enthält Lipochrom, ein Pigment, das in Fett löslich ist und für die gelbe Farbe des Corpus luteum eines frisch gewonnenen Ovars hervorruft 2.Thekaluteinzellen: hervorgegangen v.a. aus den Zellen der Theca interna und zu einem sehr geringen Teil aus denen der Zona externa sowie des umgebenden Stromas. Größer als ihre Herkunftszellen, jedoch kleiner und dunkler gefärbt als die Granulosaluteinzellen. Zeigen in ihrem Feinbau ebenfalls die für steroidbildende Zellen typische Ultrasruktur, nämlich tubuläre Mitochondrien und viel sER Corpus albicans: Hat keine Befruchtung der Eizelle stattgefunden, bildet sich das Corpus luteum zurück. Es verbleibt eine „Narbe“ 129) Tuba uterina quer, M, F, HE, 8 μm -Die Wand der Tuba uterina besteht aus: 1.Tunica mucosa: -Die der Ampulle besitzt zahlreiche verzweigte longitudinale Falten, die das Lumen einengen -> im QS Aussehen eines Labyrinths 1.1.Epithel: - einschichtig iso- bis hochprismatisch - 3 Zelltypen: 1.1.1.Flimmerzellen: - tragen an ihrer Oberfläche Kinozilien 1.1.2.sezernierende Zellen: - liegen einzeln oder in Gruppen zwischen den Flimmerzellen. Schlanker basaler aber apikal erweiterter Zellteil. Zeigen alle morphologischen Kennzeichen gesteigerter Proteinsynthese. Sezernieren merokrin und produzieren einen neutralen bis schwach sauren Schleim, der Substanzen, die für die Eizell- und Spermienreifung sowie –ernährung Bedeutung haben enthält 1.1.3.Stiftchenzellen: - vermutlich erschöpfte sezernierende Zellen. Schmale dunkle Zellen, die über die Epitheloberfläche hinausragen 1.2.Lamina propria mucosae: - lockeres Bindegewebe mit zahlreichen freien und mobilen Bindegewebezellen, die sich zu Deziduazellen umbilden 2.Tunica muscularis: 2.1.innere tubeneigene (autochtone) Muskulatur: - liegt unter der Tunica mucosa und ist v.a. im Isthmus deutlich gegliedert, wo sich mehr oder weniger deutlich inne und außen jeweils eine longitudinale und dazwischen eine ringförmige Schicht unterscheiden lassen 2.2.subperitoneale sowie peri- bzw. intravaskuläre Muskulatur: - im gesamten Eileiter vorhanden und hat verschiedene Verlaufsrichtungen -immer glatte Muskulatur, die vom Infundibulum, wo autochtone Muskulatur fehlt, bis zum Isthmus hin zunimmt. Alle Muskelsysteme des Eileiters sind durch Faseraustausch miteinander verbunden 3.Tela subserosa, Tunica serosa: -An ihrer freien Seite wird die Oberfläche des Eileiters vom Peritonealepithel (Mesothel) bedeckt, dem subseröses Bindegewebe unterlagert ist. Auf der Seite der peritonealen Befestigung (Mesosalpinx) verlaufen die ernährenden Gefäße, die Arkaden bilden und zahlreiche vegetative Nerven 121) Uterus (Beginn der Sekretionsphase), F, HE, 10 μm -von außen nach innen sind folgende Schichten zu unterscheiden: 1.Tunica serosa, Perimetrium: -besteht aus Peritonealepithel (Mesothel) 2.Bindegewebeschichten: Tela subserosa, Parametrium, Tunica adventitia: -Tela subserosa ist eine dünne Bindegewebeschicht unter der Tunica serosa mit etwas glatter Muskulatur (Stratum subserosum) -Tunica adventitia besteht aus Bindegewebe, das dem Myometrium unmittelbar aufliegt und mit dem myometralen Bindegewebe zusammenhängt -Parametrium ist der Teil des Bindegewebe, der die Verbindung zur seitlichen Beckenwand herstellt. Es besteht teilweise aus Faserbündeln mit gerichtetem Verlauf (Ligamenta), enthält glatte Muskelzellen und viele elast. Fasern. Außerdem leitet es dem Uterus Nerven und Gefäße zu 3.Tunica muscularis, Myometrium: -dickste Schicht der Uteruswand. Ein Gefüge aus glatten Muskelzellen, Bindegewebe und Gefäßen. Auch das Bindegewebe besteht teilweise aus kontraktilen Zellen (Myofibroblasten), die bald faserbildend-sekretorisch, bald als Muskelzellen tätig sind. Es lassen sich 3 durch Bindegewebe voneinander getrennte Schichten unterscheiden: Im Bereich des Corpus uteri verlaufen die Muskelbündel der inneren und äußeren Schicht hauptsächlich longitudinal. Die stärkste mittlere Schicht ist das Stratum vaskulosum, das bes. gefäßreich ist und dessen Muskelbündel bevorzugt zirkulär orientiert sind. Das Bindegewebe besteht im wesentlichen aus kollagenen und argyrophilen Fasern, die teilweise die glatten Muskelfasern umgeben 4.Tunica mucosa, Endometrium: 4.1.Epithel: -einschichtig hochprismatisches Oberflächenepithel, das aus Zellen mit Kinozilien (Flimmerzellen) und sezernierenden Zellen mit Mikrovilli besteht 4.2.Stratum functionale endometrii: -die Schicht des Endometriums, die zyklischen Veränderungen unterliegt und während der Menstruation abgestoßen wird. Sie besteht aus zellreichem faserarmen Bindegewebe (Stroma endometrii) und enthält Drüsen (Glandulae uterinae) sowie Gefäße und vereinzelte Nervenfasern. Das Bindegewebe ist mesenchymartig und bildet die Lamina propria mucosae. Interzellulär kommen ein Netzwerk aus kollagenen und ret. Fasern (Typ-I- bzw. –III-Kollagen), metachromatische Grundsubstanz aus Glykoproteinen sowie freie Nervenzellen vor. Die Glandulae uterinae sind tubulär und erreichen das Stratum basale, wo sie verzweigen können. Ihr Epithel besteht im wesentlichen aus sezernierenden Zellen, besitzt aber auch Flimmerzellen -Die Gefäße verlaufen stark gewunden, weshalb sie auch als Spiralarterien bezeichnet werde, und setzen sich über Arteriolen in ein dichtes Kapillarnetzwerk fort 4.3.Stratum basale endometrii: -wird während der Menstruation nicht abgestoßen. Von hier aus wird in der Proliferationsphase das Stratum functionale wieder aufgebaut. Es ist bindegewebereich und enthält verzweigte Endabschnitte der Glandulae uterinae. Es kommen versorgende Basalarterien vor. Die Glandulae uterinae weisen eine für die Sekretionsphase typische Sägeblattzahnung auf. 121a) Uterus-Vagina, M, F, HE, 10 μm -Portio vaginalis: bedeckt von mehrschichtigem unverhornten Plattenepithel der Vagina. Die Grenze zum einschichtigen hochprismatischen Epithel des Uterus ist deutlich. Ebenfalls im Grenzbereich liegen unter dem Epithel im Myometrium des Uterus homogen erscheinende Gebilde, Retentionszysten (Ovula nabothi). Die Tunica mucosa der Vagina ist völlig drüsenfrei. 122) Placenta reif, M, Müller-F, HE, 10 μm -Sie gliedert sich in: 1.Chorionplatte: -wird an ihrer Oberfläche von einem einschichtigem hochprismatischen bis kubischen Amnionepithel bedeckt. Subamnial folgen mehrere nicht deutlich voneinander abgrenzbare Bindegewebeschichten, die sich während der Frühentwicklung aus extraembryonalem Mesenchym des Amnions und des Chorions entwickelt haben. Dicht unter dem Amnionepithel verlaufen im Bindegewebe der Chorionplatte zahlreiche Äste der Nabelschnurgefäße, die der Blutversorgung des Zottenbaums dienen. Es folgt eine aufgelockerte Schicht aus verschieden strukturierten Zytotrophoblastzellen, zwischen denen viel Fibrinoid und Bindegewebe liegen. Zum intervillösen Raum hin wird sie von einem lückenhaften, von Fibrinoid unterbrochenen Synzytiotrophoblast bedeckt. Seitlich, am Plazentarand, gehen die Trophoblastschichten von Chorionplatte und Basalplatte ineinander über. Der Plazentarand wird von Dezidua umfaßt. Die Chorionplatte bildet die apikale Begrenzung des intervillösen Raums, dient der Verankerung der Nabelschnur und ist Leitstruktur für Gefäße 2.Zottenbäume (fetale Kotyledonen): -die reife Plazenta besitzt ca. 200, von denen 50 – 60 voll entfaltet sind. Sie sind mit der Chorionplatte verbunden und hängen in den plazentaren Blutraum. Jeder Baum besteht aus: 2.1.Stammzotten: 2.2.Endverzweigungen: 2.2.1.Intermediärzotten: 2.2.1.1.unreife Intermediärzotten: - kommen überwiegend in der unreifen Plazenta vor - typisches ret. Stroma mit zahlreichen Hofbauer-Zellen - besitzen nur ganz vereinzelt Endzotten 2.2.1.2.reife Intermediärzotten: - Stroma ähnlich locker und faserarm - Stromakanäle sowie Hofbauer-Zellen kommen kaum vor - gehen aus unreifen Intermediärzotten hervor - an ihrer Oberfläche entsteht die Hauptmasse der Endzotten 2.2.2.Endzotten: - die Endverzweigungen des Zottenbaumes - beherrschen das histologische Schnittbild durch die Plazenta 3.Basalplatte: -bildet den Boden der Plazenta und besteht aus fetalem und maternem Bindegewebe . Wird zum intervillösen Raum hin von Synzytiotrophoblast bedeckt, worunter sich eine schmale Bindegewebeschicht befindet, der ein unterschiedlich dickes Fibrinoid folgt. Unter dem Fibrinoid liegt eine Schicht mit wenigen hellen und vielen sehr großen basophilen Zytotrophoblastzellen (X-Zellen). An der Grenze zum maternen Teil der Basalplatte und darunter kommen vielkernige Riesenzellen und Deziduazellen vor. Eine Trennung zwischen maternem und fetalem Teil der Basalplatte ist nicht möglich –> Durchdringungszone. Es kommen zahlreiche mütterliche Gefäße vor, die Blut zum intervillösen Raum hin- bzw. von dort ableiten. Die Arterien haben hier einen gewundenen Verlauf (Spiralgefäße) und verlaufen gemeinsam mit den Venen, weshalb ebenfalls ihre Öffnungen an der Oberfläche der Basalplatte dicht benachbart sind. In der Randzone der Plazenta fallen die Venen durch sinusartige Erweiterungen ihrer Öffnungen auf (Randsinus). 4.Sonderstrukturen: 4.1.Inseln: - kugelig bis bizarr geformte Trophoblastzellkomplexe, die mit dem Zottenbaum verbunden sind. Sie bestehen aus modifizierten Zytotrophoblastnestern, die im inneren und an der Oberfläche Fibrinoidmassen aufweisen. Bedeckt von diskontinuierlichem Synzytiotrophoblasten und gefäßfrei 4.2.Zellsäulen: - verbinden und verankern die Haftzotten mit der Basalplatte. Sie bestehen aus Zytotrophoblastzellen 4.3.Septa placentae: - Verwerfungen der Basalplatte, die von ihr aus in den intervillösen Raum ragen. Ihr Bau entspricht dem der Basalplatte. Hier befinden sich häufig Spalten und Pseudozysten mit kolloidartigem Inhalt -zwischen den Zotten befindet sich der von mütterlichem Blut durchströmte intervillöse Raum -> die menschliche Plazenta ist hämochorial. Zotten sind für den Gasaustausch zwischen Mutter und Kind die wichtigsten Strukturen. Zotten bestehen aus: Synzytiotrophoblast: -bildet einen zusammenhängenden, stellenweise kernreichen Zytoplasmaschlauch (ohne Zellgrenzen) an der Oberfläche aller Zotten. Eine Regeneration ist nicht möglich, hierzu bedarf es des Zytotrophoblasten Zytotrophoblast: -ohne Trennung durch eine Basalmembran unter dem Synzytiotrophoblasten gelegen. Besteht aus einzeln oder in kleinen Gruppen gelegenen Zellen, die auch als Langhans-Zellen bezeichnet werden. Kommt in reifen Zotten fast ausschließlich in Nachbarschaft von Kapillaren oder Sinusoiden vor. Wird als Proliferationszone angesehen Bindegewebe: -besteht aus einem Netz ortsständiger Bindegewebezellen (Mesenchymzellen, Fibroblasten) sowie aus ungeformter und geformter Interzellularsubstanz, ret. Fasern und Kollagenfasern. Wird vom Trophoblasten durch eine Basalmembran getrennt. Seine Maschen enthalten fetale Gefäße und weinige freie Bindegewebezellen (Plasmazellen, Mastzellen, Makrophagen (HofbauerZellen)) Gefäße: -größere treten nur in den Stammzotten auf, pro Stammzotte je eine Arterie und eine Vene, denen das Mikrozirkulationssystem, das aus Arteriolen bzw. Venolen, Prä- und Postkapillaren sowie Kapillaren und venösen Kapillarabschnitten besteht. Die Gefäßstrecken des Mikrozirkulationssystems kommen v.a. in den Endverzweigungen des Zottenbaumes vor, für die sinusartig erweiterte Kapillaren charakteristisch sind. Dort, wo sie sich dem Trophoblast unmittelbar anlegen, verschmelzen die Basalmembranen von Trophoblast und Kapillare miteinander, wodurch kurze Diffusionsstrecken zwischen mütterlichem und kindlichem Blut entstehen. An der Oberfläche der Basalplatte verläßt mütterliches Blut die Spiralarterien und tritt in den intervillösen Raum ein. wo es sich zwischen Basal- und Chorionplatte verteilt. Das Blut gelangt zur Basalplatte zurück und wird von Venen abgeleitet Plazentaschranke: -wird gebildet von den Grenzschichten zwischen mütterlichem und kindlichem Kreislauf. Sie besteht aus: -Synzytiotrophoblast mit Basalmembran -dem Zottenbindegewebe -der Basalmembran und dem Endothel der Kapillaren -Grundsätzlich erfolgt der Transport in beide Richtungen. Transportiert werden alle Stoffe, die zur Ernährung des Kindes erforderlich sind, und Stoffwechselendprodukte (Atemgase durch Diffusion). Ebenfalls Hormone werden ausgetauscht. Für Blutzellen ist sie im Prinzip undurchlässig 124) Mamma, Macaca mulatta, Bouin, HE, 8 μm -Die verzweigten tubuloalveolären Drüsen der Glandulae mammaria haben eine typische Gliederung in Alveolen, Ductus lactiferi, Ductus lactifer colligens und Sinus lactifer, aus denen jede Einzeldrüse besteht. In den Ductus lactifer colligens münden viele. unterschiedlich gestaltete, verzweigte Ductus lactiferi ein. Kurz vor der Mündung erweitert sich der Ausführungsgang zum Milchsäckchen (Sinus lactifer, Durchmesser bis 8 mm) und öffnet sich schließlich an der Spitze der Papilla mammae (Brustwarze). Die Ausführungsgänge besitzen nahe an ihren Öffnungen mehrreihiges Plattenepithel. Zur Drüsentiefe hin nehmen die Zelllagen jedoch fortschreitend ab, bis schließlich nur noch zwei Schichten kubischen oder hochprismatischen Epithels vorliegen. In den sekretorischen Anteilen der Drüse, den Alveolen und Anfangsteilen der Ductus lactiferi, ist das Epithel einfach kubisch und steht mit einer Basalmembran bzw. einer nicht zusammenhängenden Schicht kräftig entwickelter Myoepithelzellen in Verbindung. Sowohl das Bindegewebe, das die Drüsen bzw. die Drüsenendstücke umgibt, als auch das eingelagerte Fettgewebe sind kräftig entwickelt. Im Bereich der Brustwarze kommen im Bindegewebe zahlreiche glatte Muskelzellbündel vor, die zirkulär angeordnet sind. Außerdem verlaufen glatte Muskelfasern parallel zu den Ausführungsgängen und setzen am Epithel der Papillae mammae an. Das Binde- und Fettgewebe zwischen den Drüsen, teilt die Brustdrüse in Lappen, Lobi, auf. Eine weitere Unterteilung in Drüsenläppchen, Lobuli, kommt durch bindegewebige Septa interlobularia zustande, die von den gröberen Bindegewebezügen ausgehen. Brustdrüse und Bindegewebeapparat zusammen bilden die Mamma. 125) Mamma lactans, M, KFE, Masson, 10 μm -siehe 124