Kapitel 35, Struktur und Wachstum der Pflanzen
Werbung

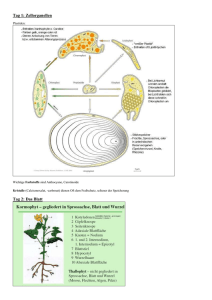
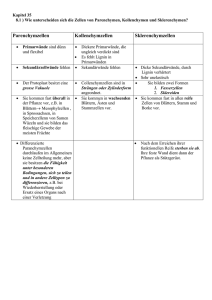
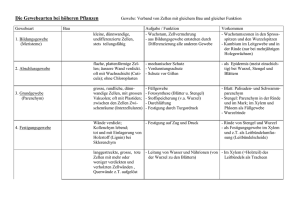
Kapitel 35, Struktur und Wachstum der Pflanzen Die Morphologie und Anatomie der Angiospermen Die Angiospermen werden in zwei Klassen aufgeteilt, in die Monocotyledoneae (Lilien, Orchideen, Gräser, Getreide etc.) und die Dicotyledoneae (Rosen, Sonnenblumen, Erbsen, Ahorn etc.) Sie werden anhand der Samenblätter bei der Embryopflanze unterschieden. Die Basis der Morphologie von Pflanzen reflektiert die Evolutionsgeschichte von Organismen, die auf dem Land leben. Eine Pflanze, die an Land lebt, muss sich gleichzeitig an zwei verschiedene Nährstoffquellen anpassen, an die Erde und and die Luft. Aus der Erde bezieht die Pflanze Wasser und Mineralien, aus der Luft hauptsächlich CO2 und Licht. Deswegen wird die Pflanze in 2 Systeme eingeteilt, unter der Erde Wurzelsystem (Hauptwurzel, Nebenwurzeln) über der Erde Sprosssystem (Stamm, Blätter, Blüte). Beide Systeme können ohne das andere nicht existieren! Fig. 35.4 Die zwei Typen von Gefässgewebe sind das Xylem (transportiert Wasser und Mineralien von den Wurzeln bis in den Trieb) und das Phloem (transportiert Assimilationen) Das Wurzelsystem / root system Die Wurzeln sind verankert in der Erde. Sie absorbieren Mineralien und Wasser, leiten Wasser und mineralische Nahrung und lagern organische Depotstoffe ab. Die Struktur der Wurzeln ist auf diese Funktionen angepasst. Viele Dicotyledoneae haben ein Pfahlwurzelsystem / taproot system, d.h. sie haben eine lange, vertikale Wurzel, welche viele kleinere, seitliche Wurzeln produziert. Die Hautwurzel ist fest und stabil. Ein Beispiel dafür ist die Karotte. Die Wurzel, also die eigentliche Karotte, die wir essen, lagert eine Menge Nahrung für die Pflanze. Die Pflanze braucht diese Reserven dann, wenn sie blüht oder Früchte macht. Deshalb werden solche Gemüse geerntet, bevor sie blühen. Die Monocotyledonen haben ein fasriges Wurzelsystem / fibrous root, d.h. die Wurzeln sind dicht unter der Erdoberfläche und gehen in alle Richtungen, wie Fasern. Ein gut verzweigtes Wurzelsystem verbessert die Erreichbarkeit von Wasser und Mineralien. Diese Wurzeln schützen den Boden sehr gut vor Erosion. Bei beiden Wurzelsystemen bilden an den Spitzen der Wurzeln sog. Wurzelhaare aus. Es sind Ausstülpungen der Rhizodermis. Diese Wurzelhaare wachsen an der Spitze und nicht am Ansatz wie beispielsweise unsere Haare! Es gibt auch Pflanzen, die Wurzeln oberhalb der Erdoberfläche haben. Sie wachsen vom Stamm oder von den Blättern der Pflanze aus. Man sagt, solche Wurzeln seien nicht dazugehörig (adventicus). So werden alle Teile der Pflanze genannt, welche an einer unüblichen Stelle wachsen. Diese nicht dazugehörigen Wurzeln dienen zur Requisite, um hohe Stämme zu unterstützen. Das Sprosssystem / shoot system Die einzelnen Teile des Stamms/Stiels/Stengels, der Blätter und Knospen, was wo lokalisiert ist, bitte Fig. 35.4. (engl. Ausgabe) selber nachschlagen. Die Blätter sind die wichtigsten Organe einer Pflanze um Photosynthese zu betreiben. Blätter sind sehr unterschiedlich in Form, jedoch bestehen alle grundsätzlich aus einem Blattstiel, dem Petiolus und einer Blattspreite. Bei Gräser und anderen Monocotyledoneae fehlt der Stengel, stattdessen formt das Blatt eine Umhüllung um den Stengel. Ein einfaches Blatt besteht aus einem ganzen Blatt, einem Stengel und einer zusätzlichen Knospe. Es gibt auch Blätter, die aus mehreren Blättchen bestehen und keinen „richtigen“ Stengel haben. Die Blätter der Monocotyledoneae (Monocots) und der Dicotyledoneae (Dicots) unterscheiden sich in der Anordnung der Adern Blattnervatur. Bei den Monocots verlaufen die Hauptadern parallel, der Länge vom Blatt entlang. Im Gegensatz zu den Monocots verlaufen die Adern bei den Dicots netzwerkartig von der Hauptader weg. Strukturelle Anpassungen von pflanzlichen Zellen an ihre Funktion Durch solche Anpassungen an ihre Funktion gibt es heutzutage einige verschiedene Zelltypen. Beispielsweise gibt es Unterschiede mit dem Protoplast. (Der Protoplast umfasst den Inhalt einer Zelle ausgenommen der Zellwand) Nur bei solchen Zellen, die mit der Photosynthese zu tun haben, finden man im Protoplast Chloroplast. Es gibt aber auch wichtige Modifikationen bei den Zellwänden. 1. Parenchymzellen/ Parenchyma Cells Die ganze Zelle ist wenig spezialisiert, wodurch sie als „typische“ pflanzliche Zelle gilt. Ihre Wände sind dünn und flexibel, meist fehlt die sekundäre Zellwand. Der Protoplast hat meist eine grosse zentrale Vakuole. Diese Zellen verrichtet die meisten metabolischen Funktionen, synthetisiert und lagert verschiedene organische Stoffe. Die Photosynthese läuft in den Chloroplasten der Mesophyllzellen (das sind die Parenchymzellen der Blätter) ab. 2. Collenchymzellen/ Collenchyma Cells Diese Zellen haben eine dickere Zellwand als die Parenchymzellen. Sie sind vor allem bei jungen Pflanzen oder bei den jungen Teilen einer Pflanze zu finden. Sie verlängern sich mit dem Stiel oder den Blättern, die sie unterstützen. 3. Sclerenchymzellen/ Sclerenchyma Cells Auch hier ist die Funktion der Zellen Unterstützung. Mit ihrer dicken und gestärkten (mit Lignin: druckfestes Material, welches Festigung ergibt) zweiten Zellwand jedoch sind sie einiges starrer als die Collenchyma Cells. Je nachdem sind die Zellen bereits schon tot bevor sie funktionell reif wären, da der Protoplast fehlt. Deswegen sind sie auch bei Teilen der Pflanze zu finden, die nicht mehr wachsen. Es gibt zwei Sorten von Sclerenchyma Cells Faser Zellen Sind langgezogene Sclerenchyma Cells, gleichmässig angeordnet Sclereiden Haben sehr dicke, lignifizierte zweite Zellwände, ungleichmässig geformt 4. Tracheiden und Gefässe/ Tracheids, Vessel Elements Das Xylem besteht aus Tracheiden und Gefässe. Beide Zelltypen sind tot bei ihrer funktionalen Reife, sie produzieren jedoch sekundäre Zellwände, bevor der Protoplast abstirbt. Bei den Gymnospermen haben die Tracheiden eine Doppelfunktion, Wassertransport und strukturelle Unterstützung. Bei den Angiospermen haben sie nur die Funktion, Wasser zu transportieren. Für den strukturellen Support sind die Fasern zuständig. Die sekundären Zellwände sind unterbrochen durch Tüpfeln/Pits, das ist eine dünnere Region, wo nur die primäre Zellwand vorhanden ist. Tracheiden sind lange dünne Zellen mit sich zuspitzenden Enden. Das Wasser fliesst durch die Pits von Zelle zu Zelle. Die Gefässe sind kürzer, haben einen grösseren Durchmesser und sind dünnerwandig als die Tracheiden. Sie sind so ausgerichtet, dass sie sich mit den Enden berühren und so lange Mikroröhren bilden, sog. Xylemgefässe. 5. Siebröhrenglieder/ Sieve-Tube Member Saccharose und andere organische Komponenten werden mit dem Phloem durch sog. Siebröhren transportiert. Die Siebröhren sind lebend bei ihrer funktionalen Reife. Bei den Angiospermen heissen die Enden der Siebröhrenelemente Siebplatten. Zwischen zwei Siebplatten liegt eine Zelle, ohne Zellkern und ohne Vakuolen, (trotzdem ist sie lebend!) jedoch mit einer funktionstüchtigen (selektiv permeabel) Plasmamembran. An der Längsseite von jedem Siebröhrenelement liegt eine Geleitzelle mit Kern. Die drei Gewebesysteme einer Pflanze Jedes Blatt, jede Wurzel oder jeder Stiel ist unterteilt in die folgenden drei Systeme: Abschlussgewebe, Leitgewebe, Grundgewebe Das äussere Abschlussgewebe, oder auch Epidermis genannt, schützt die junge Pflanze wie eine Haut. Neben dieser Hauptfunktion (Schutz) weist sie auch verschiedene charakteristische Funktionen aus, je nachdem, wo die Zelle lokalisiert ist. Die Epidermis von Blättern und den meisten Sprossachsen sondert einen wachsigen Überzug ab, der Cuticula. Das Leitgewebe bilden das Xylem und Phloem, welche immer Transport- und Stützfunktionen erfüllen. Das Grundgewebe macht die Grösse einer Pflanze aus. Es füllt die Lücke zwischen der Epidermis und dem Leitgewebe aus. Das Basisgewebe besteht hauptsächlich aus Parenchyma Cells, wobei auch die Collenchyma - und die Sclerenchyma Cells vorhanden sind. Die Funktionen sind unteranderem Photosynthese, Festigung und Speicherung. Das Basisgewebe ist zuständig für die meisten metabolischen Prozesse einer Pflanze. Meristeme erzeugen während der gesamten Lebensspanne einer Pflanze Zellen für neue Organe: eine Übersicht Die meisten Pflanzen wachsen ihr Leben ( Lebensspanne bestimmt durch Umwelteinflüsse) lang unbeschränktes Wachstum, hingegen wir Menschen und Tiere nicht. Hingegen weisen bestimmte Pflanzenorgane wie Blätter und Blüten doch beschränktes Wachstum auf. Einjährige vollenden ihren Lebenszyklus (von der Keimung über Blüte und Samenproduktion zum Tod) innerhalb eines Jahres. Die Pflanze wird zweijährig genannt, wenn sie zwei Jahr lebt. Im ersten Jahr wächst die Pflanze vegetativ und bildet die Blüte erst im zweiten Jahr. Lebt eine Pflanze mehrere Jahre wird sie mehrjährig oder ausdauernd genannt. Pflanzen sind zu unbeschränktem Wachstum fähig, weil sie fortdauernd embryonale Gewebe besitzen, die man als Meristeme bezeichnet. Sie sind nicht spezialisiert und teilen sich um weitere Zellen zu bilden. Die Quelle neuer Zellen im Meristem sind die Initialzellen. Jene, die aus dem Meristem ausscheren und sich spezialisieren, heissen Dauerzellen. Das Muster des Wachstums hängt von der Lokalisation der Meristeme ab. Apikalmeristeme befinden sich an den Wurzelspitzen und in den Sprossvegetationspunkten. Sie liefern die Zellen für das Längenwachstum. Diese Streckung wird auch Primärwachstum (nur krautige Pflanzen) genannt. Sekundäres Dickenwachstum wird bei Holzpflanzen beobachtet. D.h., die Wurzeln und Triebe, welche vom Primärwachstum stammen, werden verdickt. Das sekundäre Dickenwachstum ist das Produkt lateraler Meristeme. Sie bilden einen Cambium genannten Zylinder teilungsaktiver Zellen, der sich der Länge nach durch Wurzeln und Triebe zieht. Die Epidermis wird ersetzt durch sekundäres Abschlussgewebe, das dicker und widerstandsfähiger ist. Holz ist sekundäres Xylem, das sich über die Jahre angesammelt hat. In holzigen Pflanzen finden das Primärwachstum und das sekundäre Wachstum zur gleichen Zeit, aber an verschiedenen Orten statt. Apikalmeristeme verlängern Wurzeln und Sprosse (Primärwachstum): Das Primärwachstum erzeugt einen primären Pflanzenkörper, der aus drei Gewebesystemen besteht, den Abschlussgeweben, Leitbündeln und Grundgeweben. Krautige Pflanzen und jüngsten Holzpflanzen repräsentieren einen primären Pflanzenkörper. Primärwachstum der Wurzeln An der Wurzelspitze befindet sich die Wurzelhaube. Sie schützt das zarte Meristem, während sie sich in den Boden hinein verlängert. Diese Haube sondert auch ein Schleim ab, der den Boden im Bereich der wachsenden Wurzelspitze schmiert. Das Längenwachstum konzentriert sich auf die Spitze. Man unterscheidet drei Stadien des Primärwachstums. Von der Wurzelspitze aus gesehen findet man zuerst die Teilungszone, Streckungszone und Differenzierungszone. Die Zone der Zellteilung schliesst das Apikalmeristem ein. Das Apikalmeristem im Innern dieser Zone bildet die Zellen der primären Meristeme und ersetzt auch Zellen der Wurzelspitze, die „abgeschliffen“ sind. In der Mitte liegt das ruhende Zentrum, eine Zellpopulation, die sich langsamer als die anderen Zellen teilt. Sie dienen wahrscheinlich als Reserven, falls die anderen Zellen der Meristems in irgendeiner Weise geschädigt werden. Genau über dem Apikalmeristem bilden sich drei primäre Meristeme: Protoderm, Procambium und Grundmeristem Aus denen gehen die drei primären Gewebesysteme der Wurzel hervor (Leitgewebe, Abschlussgewebe und Grundgewebe) Die Teilungszone geht in die Streckungszone über. Hier verlängern sich die Zellen um mehr als das Zehnfache. Dort wo die Streckungszone in die Differenzierungszone übergeht, beginnen die Zellen mit ihrer Spezialisierung in Struktur und Funktion. Primärgewebe der Wurzel Von den drei primären Meristemen geht die Bildung der drei Primärgewebe der Wurzel aus. Das Protoderm ist verantwortlich für die Bildung der Wurzelepidermis, einer einfachen Zellschicht, welche die Wurzel umkleidet. Aus dem Procambium geht ein zentraler Gefässzylinder hervor, der sog. Stele oder auch Zentralzylinder. Darin entwickeln sich Xylem und Phloem. Zwischen Protoderm und Procambium liegt das Grundgewebemeristem, aus dem das Grundgewebesystem hervorgeht. Das Grundgewebe bildet das Cortex, oder die Wurzelrinde (liegt zwischen Stele und Epidermis) Die Grundgewebezellen speichern Nährstoffe. Die Endodermis ist die innerste Schicht des Cortex. Sie stellt eine selektive Barriere dar, die den Durchgang von Substanzen aus der Bodenlösung in das Leitgewebe der Stele kontrolliert. Eine etablierte Wurzel kann Seitenwurzeln bilden, die aus der äussersten Schicht der Stele, dem Perzykel hervorgehen. Seitenwurzeln werden im Perzykel angelegt, strecken sich und durchstossen das Cortex bis sie als Primärwurzeln hervortreten. Das Leitgewebe ist in der Wurzel ein durchgehendes System. Primärwachstum des Sprosses Das Apikelmeristem eines Sprosses ist eine kuppelartig geformte Masse sich teilender Zellen. Es befindet sich an der Spitze einen Endknospe, auch Terminalknospe genannt. Wie in der Wurzel bildet das Apikalmeristem der Spross-Spitze, die Primärmeristeme (auch Protoderm, Procambium und Grundmeristem) die sich weiter in die drei Gewebesysteme differenzieren. Blätter entstehen aus Blattprimordien an den Seiten des Apikalmeristems. Achselknospen entwickeln sich aus Inseln meristematischer Zellen, die vom Apikalmeristem an der Basis der Blattprimordien zurückgelassen worden sind. Der Hauptanteil der eigentlichen Spross-Streckung erfolgt beim Wachstum der etwas älteren Internodien unterhalb des Spross-Scheitels. Dieses Wachstum erfolgt durch Zellteilung / Zellstreckung innerhalb des Internodiums. Bei Gräsern setzt sich die Streckung über die Gesamtlänge des Sprosses über längere Zeit fort. Das ist möglich, da an der Basis von jedem Internodiums bleibende meristematische Regionen sitzen, sog. Intercalarmeristeme. Die Zweige des Sprosses stammen aus den Achselknospen, die auf der Oberfläche des Hauptsprosses angesiedelt sind. Die Leitbündel einer Sprossachse befinden sich nahe der Oberfläche. Daher können sich Seitenzweige mit einer Verbindung zum Leitgewebe entwickeln, ohne aus der Tiefe des Hauptsprosses hervorgehen zu müssen. Primärgewebe der Sprossachse In der Sprossachse verläuft das Leitgewebe in mehreren Strängen, die Leitbündel genannt werden. Dies unterscheidet sich von der Wurzel, wo das Leitgewebe eine einzige Stele bildet, die aus dem vereinigten Satz der Leitbündel besteht. In der Übergangszone, dem Wurzelhals, wo Spross und Wurzel ineinander übergehen, laufen die Sprossachsenleitbündel zusammen und bilden die Wurzelstele. In den meisten Dicots sind die Leitbündel ringförmig angeordnet, mit Markparechym innerhalb des Rings und Rindenparenchym (Cortex) ausserhalb. Beide sind Teile des Grundgewebesystems. Das Xylem der Leitbündel liegt jeweils innen zum Mark, ihr Phloem verläuft entlang der Rinde. In den Monocots liegen die Leitbündel im Grundgewebe über den Querschnitt verstreut. Aus dem Protoderm der Endknospe geht die Epidermis hervor, die Sprossachse und Blätter als Teil des durchgehenden Abschlussgewebes überkleidet. Die Gewebeorganisation der Blätter Das Blatt wird von einer Epidermis bedeckt, deren Zellen wie Puzzleteile eng miteinander verzahnt sind. erste Verteidungslinie gegen Schäden Die wachsige Cuticula stellt ausserdem eine Barriere gegen Wasserverlust dar. Diese epidermale Barriere wird nur durch die Stomata (Spaltöffnungen) unterbrochen, winzigen Poren, die von den Schliesszellen gebildet werden. Die Stomata erlauben den Gasaustausch zwischen Aussenluft und den photosynthetisch aktiven Zellen im Blatt. Sie sind auch die Hauptroute für den Wasserverlust der Pflanze durch Verdunstung. Transpiration Das Grundgewebe des Blattes, das zwischen oberer und unterer Epidermis liegt, ist das Mesophyll. Es besteht aus hauptsächlich aus Parenchymzellen, die mit Chloroplasten ausgerüstet und spezialisiert sind für die Photosynthese. In den meisten Dicots befinden sich unterschiedliche Mesophylle. In der oberen Hälfte des Blatts findet man ein oder zwei schichtige Palisadenparenchym. In der unteren Hälfte findet man das Schwammparenchym mit einem Labyrinth von Lufträumen. In den Lufträumen zirkuliert Kohlendioxid und Sauerstoff. Sie steigen zu den Palisadenparchym auf. Bei den meisten Pflanzen ist die Zahl der Stomata auf der Blattunterseite deutlich grösser als auf der Blattoberseite. Dadurch wird der Wasserverlust auf ein Minimum herabgesetzt, der auf der Blattoberseite viel schneller erfolgt. Xylem und Phloem der Sprossachse setzen sich ununterbrochen in die Leitbündel des Blattes fort. Die Abzweigungen der Leitbündel sind die Nervatur des Blattes. Sie durchziehen das gesamte Mesophyll und sind so im engen Kontakt mit dem photosynthetisch aktiven Gewebe, das aus dem Xylem Wasser und Mineralien erhält und das Phloem mit Zuckern und anderen organischen Produkten zum Transport in andere Teile der Pflanze schickt. Modularer Sprossaufbau und Phasenveränderungen während der Entwicklung Die fortlaufende Entwicklung von Sprossknoten und Internodien innerhalb einer Triebspitze mit anschliessender Internodienstreckung bringt einen Trieb mit modularem Aufbau hervor. Das Alter eines Pflanzenmodules wächst direkt proportional mit seiner Entfernung zum Apikalmeristem. Das Meristem kann von einer Entwicklungsphase in eine andere wechseln. Einer dieser Phasenwechsler besteht in einem schrittweisen Übergang vom juvenilen vegetativen (blattproduzierenden) zum reifen (adulten) vegetativen Status. Auffäligstes Zeichen dafür ist die Modifikation der Blattmorphologie. Die Blätter eines juvenilen unterscheiden sich von dem eines adulten in Form und anderen Merkmalen. Hat das Meristem einmal juvenile Nodi und Internodien angelegt, behalten diese ihren Status bei, auch wenn die Sprossachsenstreckung einsetzt und das Meristem in die Reifephase übergeht. In einigen Fällen vollzieht die Spross-Spitze einen zweiten Phasenübergang vom reifen vegetativen zum reproduzierenden (blütenbildenden) Status. Im Gegensatz zum vegetativen selbsterneuernden Wachstum beendet die Blütenbildung das Primärwachstum der betreffenden Spross-Spitze. Das Apikalmeristem wird verbraucht bei der Blütenbildung. Laterale Meristeme erweitern den Umfang von Sprossachsen und Wurzel (sekundäres Dickenwachstum): Zusätzlich zum Streckenwachstum führen die meisten Gefässpflanzen auch ein sekundäres Dickenwachstum durch. Der sekundäre Pflanzenkörper besteht aus den Geweben, die während der Durchmesserzunahme entstanden sind. Zwei laterale Meristem spielen im sekundären Dickenwachstum eine Rolle: das Leitgewebe-Cambium (bildet sekundäres Xylem und sekundäres Phloem) und das Korkcambium (bildet widerstandsfähige, dicke Schutzschicht für Spross und Achse). Sekundäres Dickenwachstum zeigen alle Gymnospermen. Bei den Angiospermen findet es bei den meisten Dicots statt, bei Monocots jedoch nur selten. Sekundäres Dickenwachstum der Sprossachse Das Leitgewebe-Cambium geht aus Parenchymzellen hervor, die ihre Teilungsfähigkeit wiedererlangt haben und wieder meristematisch werden. Der Übergang zur meristematischen Aktivität erfolgt in einer Zone, die zwischen primären Xylem und Phloem eines jeden Leitbündels liegt. Das Cambium das innerhalb der Leitbündel liegt, heisst Fascikularcambium, der andere Teil, der sich in den Markstrahlen zwischen den Leitbündeln befindet, bildet das Interfascikularcambium. Zusammen bilden sie einen geschlossenen Cambiumzylinder. Die meristematischen Zellen des interfascikulären Cambiums werden als Markstrahlinitialen bezeichnet. Sie bilden radiale Reihen parenchymatischer Zellen, die auch als Xylemstrahlen und Phloemstrahlen unterschieden werden. Die Cambiumzellen innerhalb der Leitbündel heissen fusiforme Initialen. (fusiform = zugespitze Enden) Entlang der Sprossachse sind sie gestreckt. Sie bilden neues Leitgewebe und erzeugen so sekundäres Xylem (Holz) und sekundäres Phloem (Bast). Durch dauerndes Dickenwachstum entsteht Holz. Es besteht hauptsächlich aus Tracheiden, Gefässgliedern und Holzfasern. Diese Zellen, die bei der Erreichung der funktionalen Reife tot sind, geben dem Holz Stärke und Härte. (durch die lignifizierten Wände der Zellen) Jedes Jahr wird in den gemässigten Klimazonen der Welt das sekundäre Dickenwachstum unterbrochen, wenn das Cambium Winterpause macht. Im Frühling wird das sekundäre Dickenwachstum wieder aufgenommen, so entstehen die Jahrringe. Das sekundäre Phloem häuft sich im Laufe der Jahre nicht so beträchtlich wie das Xylem. Während der Baum an Umfang zunimmt, entwickelt sich das ältere sekundäre Phloem zur Rinde, die schliesslich aufplatzt und abblättert. Die Epidermis, die während dem Primärwachstum gebildet wurde, reisst und trocknet aus und fällt vom Stamm ab. Ersetzt wird sie von neuem Gewebe, das vom Korkcambium gebildet wird. Das Korkcambium ist ebenfalls ein Zylinder aus meristematischen Zellen. Aus der Teilung der Initialen im Korkcambium gehen Parenchymzellen hervor, die Phelloderm heissen, falls sie auf der Innenseite liegen, aussenliegende heissen Korkzellen. Die Korkzellen lagern in ihre Zellwände ein wachsähnliches Material, Suberin genannt ein und sterben ab. So ist auch hier eine Barriere da, die den Spross vor physischem Schaden und Pathogenen schützt. Phelloderm, Korkcambium und Korkschichten bilden zusammen das Periderm. Das ist ein Schutzmantel des sekundären Pflanzenkörpers, der die Epidermis des primären Sprosses ersetzt. Die Grösse des ursprünglichen Zylinders des Korkcambiums ist festgelegt. Nach kurzer Zeit verliert das Korkcambium seine meristematische Aktivität und die verbleibenden Zellen werden zu Kork. Die zunehmende Ausdehnung der Sprossachse lässt das ursprüngliche Periderm reissen. Neuer Korkcambium bildet sich tiefer in der Rinde. Schliesslich bleibt keine Rinde übrig, und das benötigte Korkcambium entsteht dann im sekundären Phloem aus Parenchymzellen. Falls weitere Korkcambien den Schutzmantel der Pflanze aufbauen, heisst es Borke. Die Borke bezeichnet alle Gewebe ausserhalb des innersten, aktiven Korkcambiums. Nur das jüngste Phloem ist im Zuckertransport tätig. Man kann das Ergebnis jahrelangen sekundären Dickenwachstums erkennen, wenn man einen alten Baustamm betrachtet.