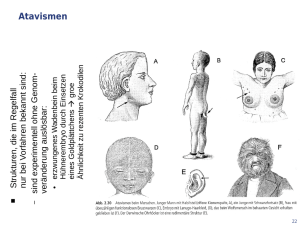
Document
Werbung

2.2 Ursachen von Veränderungen des Genpools: Evolutionsfaktoren Prinzipiell gilt: Jede Veränderung des Genpools bedeutet Evolution. Was kann nun Veränderungen des Genpools, d. h. der Allelhäufigkeiten bewirken? 2.2.1 Mutationen: Sie führen zum Eintrag von neuen Allelen. Wenn mehr als zwei Allele pro Genort in einer Population existieren, spricht man von multipler Allelie. (Wiederholung: Welche Arten von Mutationen gibt es und wie werden sich die verschiedenen Mutationsarten auf die Evolution bzw. auf den Genpool auswirken?) Die Mutationsrate pro Einzelgen ist gering (10-5 bis 10 -6) auf den Gesamtorganismus bezogen allerdings recht hoch: ca.10 % der menschl. Keimzellen tragen Neumutationen) Fazit: Mutationen liefern das Rohmaterial für die Evolution 2.2.2 Rekombination Sexuelle Fortpflanzung sorgt auf drei Ebenen für eine Neukombination der Allele: a) durch zufällige Verteilung der Chromosomen in der Meiose I bei der Keimzellenbildung (=interchromosomale Rekombination) b) durch Stückaustausch beim Crossing-Over (=intrachromosomale Rekombination) c) durch zufälliges Zusammentreffen von Gameten Prinzipiell verändert die Rekombination nicht den Genpool einer Population, sie sorgt nur für eine optimale Durchmischung. Rekombination über Populations- und Rassengrenzen hinweg wird als Hybridisierung bezeichnet und gilt auch als Evolutionsfaktor. 2.2.3 Selektion: 2.2.3.2 Grundsätzliches Selektion greift am durch Mutation und Rekombination gebildeten Phänotyp an. Bestimmte Phänotypen (d. h. Allelkonstellationen) haben in einer bestimmten Umwelt Überlebensvor- oder Überlebensnachteile, bzw. haben größere oder kleinere Chancen sich fortzupflanzen. Die Selektion gibt der Evolution eine Richtung. Beispiel: Oft kommen homozygot rezessive Träger einer Erbkrankheit (z. B. Mucoviszidose) gar nicht oder mit stark verringerter Wahrscheinlichkeit zur Fortpflanzung, da ihre Fitness gering ist. Die Fitness (Kriterium für den Evolutionserfolg) hängt ab von: 1. Lebensdauer des Individuums (hängt oft ab vom Gesundheitszustand) 2. Chance, einen Geschlechtspartner zu finden (geschlechtliche Zuchtwahl) 3. Fertilität, d. h. Zahl der Nachkommen (hoch bei R-Strategen, vgl. Cornelsen S. 337)) 4. Überlebensrate der Nachkommen (hoch bei K-Strategen, vgl. Cornelsen S. 337)) Das klassische Beispiel der Selektion beim Birkenspanner (Cornelsen S. 244) zeigt, dass veränderte Umweltbedingungen (mehr oder weniger Ruß auf Birken) erheblichen Einfluss auf die Richtung der Selektion haben können. 2.2.3.2 Selektionsarten: (Cornelsen S. 245) - stabilisierende Selektion - gerichtete = transformierende = verschiebende Selektion - disruptive = aufspaltende Selektion Merke! Selektion bewirkt keine Variation, sondern sorgt nur für die Bevorzugung schon vorher bestehender Varianten (Präadaptation oder Prädisposition). 2.2.3.3 Selektionsfaktoren: (Cornelsen S. 246 f. und S. 299-340 = Einschub Ökologie als Expertenrunde) abiotische: - Temperatur (Klimaregeln; Wüstenlebewesen, C4 und CAM-Pflanzen) - Luftfeuchtigkeit - Lichtmenge und Lichtqualität - Salzkonzentration (im Wasser) - Sauerstoffgehalt - CO2-Gehalt,pH-Wert - Wind (Bsp. Kerguelen-Fliege, Cornelsen S. 246), usw…. biotische: - intra- und interspezifische Konkurrenten, z. B. Nahrungskonkurrenten - Beutetiere oder Räuber (→ Tarnung, Flucht, Warnung, Mimikry etc., z. B. Abendpfauenauge erschreckt mit Augenpaar, Hornissenschwärmer ahmt Hornisse nach = Mimikry) - Futterpflanzen - „Bestäuber“ bei Pflanzen (vgl. Koevolution, Die Evolution von Blüten und Insekten ist gekoppelt und geschieht durch wechselseitige Selektion, z. B. Fingerhut bietet Landeplatz für Hummeln, Cornelsen S. 246) - Parasiten (Koevolution) - Geschlechtspartner (oft Sexualdimorphismus, Weibchen und Männchen einer Art sehen unterschiedlich aus und sind unterschiedlich groß, z. B. Hirschkäfer); Sexuelle Selektion, d.h. Weibchen wählt meistens unter verschiedenen Männchen aus, wobei oft die äußerlich prachtvollsten Männchen das beste Immunsystem haben und sich z. B. den Luxus eines parasitenfreien Pfauenschwanzes leisten können. (auch Bsp. Birkhahn). Außerdem kann es zu Einschüchterung und Kampf unter Konkurrenten um Weibchen kommen (z. B. Hirsche). - Künstliche Zuchtwahl durch den Menschen (Bsp. Taubenrassen) 2.2.4 Isolation: Unterbindung der Paarung und somit des Genaustausches zwischen Teilpopulationen 1. Geografische Isolation = Separation: Trennung von Teilpopulationen durch Meer, Gebirge, Kontinentaldrift… Beispiele: Erdhörnchen am Grand Canyon, Grünspecht und Grauspecht Galapagosfinken (Cornelsen S. 248) 2. Ökologische Isolation = Einnischung: Anpassung an verschiedene ökologische Nischen. Falls viele Nischen zu Beginn frei sind, kann dies zur adaptiven Radiation (vgl. S. 253), d. h. zur breit gefächerten Aufspaltung einer Ursprungspopulation führen. Allerdings sind hierbei vermutlich noch andere Mechanismen wie ethologische Isolation und Hybridisierung der verschiedenen Unterpopulationen von Bedeutung. 3. Zeitliche Isolation: Balz oder Blüte zu verschiedenen Zeiten, z. B. blüht der rote Holunder früher als der schwarze Holunder , Gras- und Wasserfrosch laichen zu verschiedenen Zeiten ab, abhängig von der Wassertemperatur (S. 249). 4. Ethologische Isolation: Teilpopulationen „verstehen“ sich nicht mehr, z. B. unterschiedlicher Gesang von Grundfink und Baumfink auf den Galapagosinseln. 5. Mechanische Isolation: Blüten oder Begattungsorgane passen nicht mehr nach dem Schlüssel-Schloss-Prinzip zusammen. 6. Genetische Isolation: Genetische Veränderungen, z. B. Polyploidisierung oder Hinzukommen oder Wegfallen eines Chromosoms durch Teilen oder Verschmelzen führen dazu, dass es keine fertilen Nachkommen mehr gibt. Hierzu zählt auch die Isolation durch Sterilität, wie sie z. B. bei der Kreuzung von Pferd und Esel zu Muli oder Maultier vorliegt. (Grund: Pferd hat 64, Esel 62 Chromosomen, was beim Mischling die Bildung befruchtungsfähiger Keimzellen verhindert.) 2.2.5 Gendrift (Cornelsen S. 250) Unter Gendrift versteht man das zufällige Verschwinden (oder Hinzukommen) von Allelen bei kleinen Populationen vor allem in Folge von Katastrophen, wenn z. B. zufällig alle Träger eines bestimmten Allels (z. B. alle gelockten Meerschweinchen) sterben. Die drastische Dezimierung von Populationen aufgrund von Katastrophen oder Klimaänderungen bezeichnet man als Flaschenhalseffekt. Die hierdurch erfolgte genetische Verarmung kann auch zum Aussterben der Restpopulation führen. Die Neubesiedlung eines Gebietes durch wenige Individuen führt zum so genannten Gründereffekt, der ebenfalls auf Zufall beruht. Erklärung: Eine Population, die sich aus wenigen Individuen , z. B. einem Kaninchenpaar entwickelt, besitzt in ihrem Genpool nur die Allele dieses Paares, d. h. nur einen Bruchteil der Allele der Ursprungspopulation. (Bsp.: Ein Pärchen glatter Meerschweinchen wird wieder nur glatte Nachkommen bekommen!) Eine geringe Populationsgröße führt auch zu vermehrter Inzucht (=Störung der optimalen Rekombination, keine Panmixie). In kleinen isolierten Gebieten kommt es durch gehäufte Inzucht zum vermehrten Auftreten von Homozygoten, d. h. auch zur gehäuften Ausprägung homozygot rezessiver Erbkrankheiten. Dies gilt auch dort, wo viele Verwandtenehen vorkommen (Bsp. Bluterkrankheit im Adel). Anmerkung: Wenn man allerdings zwei Inzuchtlinien mit unterschiedlichen homozygoten Allelen kreuzt, kommt es zum verstärkten Auftreten von Heterozygoten, die sich durch besondere Fitness auszeichnen. Diesen Effekt bezeichnet man als Heterosiseffekt. Der Hauptgrund ist vermutlich das Fehlen der Ausprägung von homozygot rezessiven Defekten.