Abhängigkeiten der Fotosynthese
Werbung

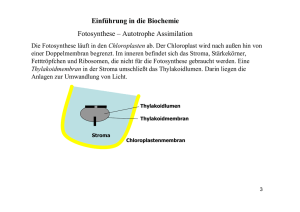
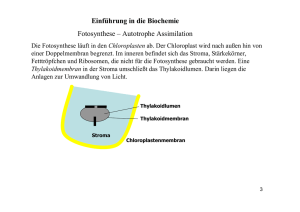
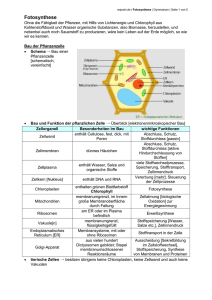
Biologie-Skript 11/12, Herr Blaurock 2. Energiebindung und Stoffaufbau durch Fotosynthese 2.1 Fotosynthese und Kohlenstoffkreislauf [Anm: In der Abbildung fehlen die sog. chemoautotrophen (gr.: chemisch selbsternährend) Organismen, Bakterien die energiereiche organische Substanzen mithilfe von chemischer Energie aus energiereichen anorganischen Substanzen (z.B. H2S) erzeugen. In Abgrenzung davon werden Lebewesen (z.B. Pflanzen) die energiereiche organische Substanzen mithilfe der Lichtenergie erzeugen, fotoautotroph (gr.: mit licht selbsternährend) genannt. Organismen (z.B.: Tiere oder Pilze), die energiereiche organische Substanzen nur abbauen können werden heterotroph (gr.: von anderen ernährt) genannt.] 2.2 Abhängigkeiten der Fotosynthese Experimentell lässt sich feststellen, dass die Fotosyntheseleistung von zahlreichen Faktoren abhängig ist. CO2-Konzentration Lichtintensität Temperatur pH-Wert Die Abhängigkeit von den Faktoren CO2-Konzentration (als Substrat) sowie Temperatur bzw. pHWert lässt sich durch enzymkatalysierte Schritte in der Fotosynthese erklären und die zugehörigen Schaubilder entsprechen den Erwartungen an eine Enzymreaktion. Schaubilder hierzu siehe Buch S. 34 Biologie-Skript 11/12, Herr Blaurock Als neuer Faktor kommt die Lichtintensität hinzu. Mit steigender Lichtintensität steigt auch die theoretische Fotosyntheseleistung, auch reelle Fotosynthese genannt (messbar an der O2-Abgabe bzw. CO2-Aufnahme), bis sie bei Lichtsättigung der Pflanze ein Maximum erreicht. Allerdings die experimentell bestimmte Fotosyntheseleistung, auch apparente Fotosynthese genannt, stets etwas niedriger als der erwartete Wert, bei sehr niedrigen Lichtintensitäten beobachtet man sogar eine "negative" Fotosyntheseleistung (insgesmt eine CO2-Abgabe anstelle einer –Aufnahme)! Dieser Effekt resultiert aus der Zellatmung, der Umkehrreaktion der Fotosynthese, welche die Pflanze ständig betreibt um ihre Zellen (auch bei Dunkelheit bzw. wenn diese gar keine Chloroplasten besitzen, z.B. Wurzelzellen) mit Energie aus den selbst hergestellten Kohlenhydraten zu versorgen. Die Zellatmungsrate ist bei gleichbleibender Temperatur konstant. Insgesamt ist die apparente Fotosynthesekurve daher gegenüber der reellen Fotosynthese um die Zellatmungsrate nach unten verschoben. Die Lichtintensität, bei der sich Zellatmung und Fotosyntheseleistung die Waage halten (also CO2 insgesamt weder aufgenommen noch abgegeben wird), wird Lichtkompensationspunkt genannt. 2.3 Blattstruktur und Fotosynthese Arbeitsblätter [Lösung auf die Frage nach der Funktion der Kohlenwasserstoffkette des Chlorophyllmoleküls: Der hydrophobe Kohlenwasserstoffanhang des Chlorophyllmoleküls verankert es im hydrophoben Inneren der Thylakoidmembran] 2.4 Fotosynthesepigmente und Lichtabsorption Praktikumsanweisung und Arbeitsblatt (ausgefüllter Lückentext siehe unten) Bei der Beobachtung von Bäumen und anderen Pflanzen fällt die Einheitliche Farbe aller Fotosynthese-betreibenden Pflanzenbestandteile auf: Grün. Biologie-Skript 11/12, Herr Blaurock Bei der Chromatographie eines Blattextraktes wird aber deutlich, dass mehrere Farbstoffe in den Blättern einer Spinatpflanze vorliegen. Zwei grüne Farbstoffe liegen vor, nämlich Chlorophyll a und Chlorophyll b dazu mehrere gelbe Farbstoffe, die sogenannten Carotinoide, darunter β-Carotin und Lutein. Um zu verstehen, warum die Pflanzenfarbstoffe uns ausgerechnet in diesen Farben erscheinen ist ein wenig Physik notwendig und ein berühmtes Experiment des Biologen Theodor Engelmann aus 1882. Das für uns sichtbare Licht kann auf zweierlei Arten aufgeteilt werden: Nach den verschiedenen Farben oder aber nach den dazugehörigen Wellenlängen Die Zuordnung ist auf der Skala dargestellt. Dabei ist es wichtig zu verstehen, dass Licht kleinerer Wellenlängen (im blauen Bereich) deutlich Energiereicher ist als Licht größerer Wellenlängen. Bei der Betrachtung eines farbigen Gegenstands sehen wir übrigens immer nur diejenige Farbe, die vom Gegenstand reflektiert, also nicht absorbiert wird. Ein schwarzer Gegenstand absorbiert also alle Lichtfarben, ein weißer Gegenstand absorbiert keine Lichtfarbe und reflektiert alles Licht. Dabei ist gar nicht klar, ob Pflanzen eigentlich alle Lichtfarben (und –Energien) für die Fotosynthese ausnutzen oder nur bestimmte Lichtteile. Theodor Engelmann hat einen raffiniert einfachen Versuchsaufbau verwendet um genau dieses herauszufinden. Er hat mit einem Prisma das sichtbare Licht in seine Farben aufgetrennt und damit eine lange Fadenalge beschienen. Auf dieser Alge war eine Bakterienkolonie angesiedelt, die zum Überleben Sauerstoff benötigt. Das Ergebnis des Experiments ist rechts dargestellt. Zum Thema NAD+/NADP+ und deren Funktion bzw. Vergleich mit ATP/ADP siehe Buch S. 33 2.5 Reaktionsschritte der Fotosynthese Allgemeine Fotosynthesegleichung bisher 6 CO2 + 6 H2O ------> C6H12O6 + 6 O2 Arbeitsblatt: Der Hill-Versuch ACHTUNG: Bei der folgenden Blackbox-Darstellung hat sich ein Fehler in der Stunde eingeschlichen. Wasser ist kein Ausgangsstoff der Lichtunabhängigen Reaktion!!! Warum nicht, erklärt der folgende Abschnitt, der die Ergebnisse des Arbeitsblatts zur Hill-Reaktion zusammenfasst. Aus Hills Experiment schließen wir, dass die Fotosynthese in eine Lichtreaktion und eine Dunkelreaktion bzw. lichtunabhängige Reaktion aufgeteilt werden kann. Bei der Lichtreaktion, die Hill untersucht hat, wird Wasser in Sauerstoff und H+-Ionen aufgespalten. Dabei werden Elektronen frei, die auf NADP+ übertragen werden. Die entstandenen H+-Ionen werden bei der Elektronenaufnahme auf NADP+ übertragen, die Sauerstoffatome gehen als elementarer Sauerstoff aus der Lichtreaktion hervor. Die Lichtreaktion verbraucht dabei jedoch weder CO2 noch wird Glucose erzeugt. Die Reaktionsgleichung sieht unseren Ergebnissen nach wie folgt aus: Biologie-Skript 11/12, Herr Blaurock Dies hat Folgen für die bisher angenommene Fotosynthesegleichung, denn wenn der komplette elementare Sauerstoff bei der Lichtreaktion entsteht, dann benötigen wir wesentlich mehr Wasser um diesen zu erzeugen. Blackbox-Darstellung der Fotosynthese Wie in der revidierten Fotosynthesegleichung farbig verdeutlicht, stammen die Sauerstoffatome des entstehenden Elementaren Sauerstoffs aus dem Wasser, das als Ausgangsstoff Teil der Lichtreaktion ist. Um die überschüssigen Sauerstoffatome des Kohlenstoffdioxids sowie überschüssige Wasserstoffatome des bei der Lichtreaktion verbrauchten Wassers auszugleichen, entstehen bei der lichtunabhängigen Reaktion zwangsläufig Wassermoleküle. Arbeitsblatt: Elektronentransportkette der Lichtreaktion Die chemiosmotische ATP-Bildung der Lichtreaktion Arbeitsblatt: Chemiosmotische ATP-Bildung… Alle Abläufe der Lichtreaktion erzeugen entweder H+ –Ionen im Thylakoidinnenraum des Chloroplasten (Wasserspaltung, H+-Transport der Elektronentransportkette) oder verbrauchen H+Ionen im Stroma („Cytoplasma“) des Chloroplasten. Hierdurch entsteht ein Ungleichgewicht an H+Ionen entlang der Thylakoidmembran: Der Innenraum ist H+-reich und dementsprechend sauer, der Außenbereit ist H+-arm und dementsprechend basisch. Dieses Ionen-Ungleichgewicht kann vom Chloroplasten ausgenutzt werden: Zur ATP-Synthese. Der Begriff Chemiosmose bezieht sich darauf, dass die Energie mit der die H+-Ionen ausströmen („Osmose“) um die H+-Konzentration auszugleichen ausgenutzt wird um eine chemische Reaktion anzutreiben. Dies wird durch einen Zusammenschluss zweier Membranproteine bewerkstelligt: Ein H+-Ionenkanal gekoppelt an eine ATP-Synthase. Letztere wird beim Ausstrom von H+-Ionen gedreht (man kann sich das vorstellen wie eine Turbine) und erzeugt in der Drehung aus ADP + P ATP. Biologie-Skript 11/12, Herr Blaurock Überblick Lichtreaktion Da bei der Erzeugung eines Sauerstoffmoleküls ein Unterschied von insgesamt 12 H+-Ionen zwischen Thylakoidinnenraum und Stroma entsteht beträgt und mithilfe 4 transportierter H+-Ionen ein ATP-Molekül erzeugt werden kann, beträgt die Gesamtgleichung der Lichtreaktion: 2 H2O + 2 NADP+ + 3 ADP + 3 P ----> O2 + 2 NADPH + H+ + 3 ATP Für einen Fotosyntheseschritt muss diese Gleichung jedoch nochmals versechsfacht werden, da pro erzeugtem Glucosemolekül sechs Sauerstoffmoleküle entstehen: 12 H2O + 12 NADP+ + 18 ADP + 18 P ----> 6 O2 + 12 NADPH + H+ + 18 ATP Biologie-Skript 11/12, Herr Blaurock Zyklischer Elektronentransport Kann ein Chloroplast die folgenden Schritte der Dunkelreaktion nicht durchführen, so würde sich NADPH+H+ im Chloroplasten ansammeln, bis schließlich NADP+ als Elektronenakzeptor aufgebraucht ist. Um dennoch die ATP-Synthese antreiben zu können, verfügen einige Pflanzen über einen alternativen Stoffwechselweg: Den zyklischen Elektronentransport. Biologie-Skript 11/12, Herr Blaurock Die lichtunabhängigen Reaktionen (Calvin-Zyklus) [ergänzt aus dem ausgeteilten Skript] Ausgangspunkt für die Untersuchung der lichtunabhängigen Reaktion der Photosynthese sind die Ausgangsstoffe: Auf der einen Seite stehen die die Produkte der lichtabhängigen Reaktion Bruttogleichung: 12 H2O + 12 NADP+ + 18 ADP + 18 P ----> 6 O2 + 12 NADPH + H+ + 18 ATP Auf der anderen Seite steht CO2 als Kohlenstofflieferant für die Synthese der Glucose Der amerikanische Biochemiker Melvin Calvin untersuchte den genauen Ablauf der Fixierung des gasförmigen Kohlenstoffdioxids in einen Feststoff, der zu Glucose weiterverarbeitet werden kann. Diese Vorgänge finden allesamt im Stroma des Chloroplasten statt. Dabei entdeckte er folgendes (Experimenteller Hintergrund [->TOK] siehe Natura 11 S. 42): CO2 wird auf ein Molekül mit fünf C-Atomen (kurz C5) übertragen, Ribulosebisphosphat (RubP) Nach der Übertragung von CO2 entstehen aus dem entstandenen C6-Zwischenprodukt zwei gleiche Moleküle mit drei Kohlenstoffatomen (C3), Phosphyglycerinsäure (PGS) Kurz darauf verschwindet PGS und es tritt ein neues C3-Molekül auf, das elektronen- und energiereiche Phosphoglycerinaldehyd (PGA) Zwei PGA-Moleküle bilden schließlich ein Glucosemolekül (C6) Aus unserem Vorwissen zu ATP und NADPH + H+ wissen wird, dass diese Stoffe vermutlich für die Umwandlung von PGS zu PGA verantwortlich sind (Abgabe von Energie und Elektronen zur Bildung des Reaktionsprodukts PGA), und so ist es tatsächlich auch. Somit schließt sich auch der Kreis zwischen der Licht- und Dunkelreaktion, die dabei entstandene Energie sowie die Elektronen werden bei der Erzeugung von PGA verbraucht. Bei der Reaktion entsteht als Nebenprodukt Wasser. Die anfängliche Übertragung von CO2 auf RubP wird von einem Enzym übernommen, dass wie üblich nach den Ausgangsstoffen der katalysierten Reaktion benannt ist: Ribulosebisphosphatcarboxylase (Rubisco) In der Übersicht sehen die Prozesse wie folgt aus: Biologie-Skript 11/12, Herr Blaurock An der Stelle verbleibt aber ein Problem: Während der C1-Baustein Kohlenstoffdioxid bei der Fotosynthese kontinuierlich verbraucht wird, besitzt der Chloroplast nur einen begrenzten Vorrat des C5-Bausteins Ribulosebisphosphat. Glücklicherweise kann Ribulosebisphosphat allerdings unter ATPVerbrauch aus PGA gebildet werden. Um die Reaktionen am Leben zu erhalten, muss also ein Überschuss an PGA erzeugt werden, um diesen anschließend in Ribulosebisphosphat umzuwandeln. Die Umwandlung von PGA in Ribulosebisphosphat wird als Akzeptor-Regeneration bezeichnet. Der Gesamtprozess der lichtunabhängigen Reaktionen ist also ein Kreislauf aus CO2-Übertragung auf Ribulosebisphosphat, PGA-Synthese, Umwandlung in Glucose und Regeneration von Ribulosebisphosphat: Der Calvin-Zyklus. Die Gesamtgleichung der lichtunabhängigen Reaktionen beträgt 6 CO2 + 18 ATP + 12 NADPH+H+ ----> C6H12O6 + 18 (ADP + P) + 12 NADP+ + 6 H2O Kombiniert man diese Gleichung mit der Gesamtgleichung der lichtabhängigen Reaktion erhält man wie erwartet die Gesamtgleichung der Photosynthese als Ergebnis. 12 H2O + 6 CO2 ----> C6H12O6 + 6 H2O + 6 O2