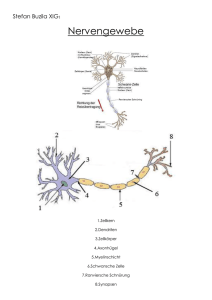
Plastizität des auditorischen Mittelhirns
Werbung

7. DGA Jahrestagung 2004 Die Plastizität des auditorischen Hirnstamms nach einseitiger Schwerhörigkeit oder Ertaubung Illing R.-B., Meidinger M.A., Kraus K.S. Neurobiologisches Forschungslabor, Hals-Nasen-Ohrenklinik, Universität Freiburg, Killianstr. 5, 79106 Freiburg, E-mail: [email protected] Hintergrund und Fragestellung Das auditorische System des Menschen ist plastisch. Für die Identifikation der Richtung, aus der ein Geräusch kommt, müssen wir die Signale, die uns von beiden Ohren vermittelt werden, vergleichen und verrechnen. Eine wichtige physikalische Größe für diese Verrechnung ist der binaurale Lautstärkeunterschied. Bei Versuchspersonen kann man diese Größe dadurch stören, dass man den Außenohrkanal einer Seite mit einem schalldämmenden Ohrenstöpsel ausfüllt. Dies hat zur Folge, dass die Versuchsperson eine unsichtbare Schallquelle zunächst falsch lokalisiert. Erst im Laufe von etwa einer Woche hat sich das zentrale auditorische System den veränderten Bedingungen angepasst und die Versuchsperson lokalisiert Schallquellen wieder richtig (Florentine, 1976). Welche Vorgänge dabei im Gehirn ablaufen wissen wir jedoch nicht. Auch an Patienten, die postlingual ertaubt sind und ein Cochlear Implantat erhalten haben, kann man binaurale Plastizitätsphänomene beobachten. Haben diese Patienten ein Resthören auf dem anderen Ohr, das sie z.B. vermittels eines Hörgerätes nutzen, entscheidet sich meist binnen einer Woche, ob die Signale beider Ohren miteinander integriert werden können oder ob der betreffende Patient bevorzugt, nur eines der Geräte anzuschalten. Auch die Einbeziehung der Elektroden im Hochtonbereich ist für die Patienten unmittelbar nach der Implantation oft schwierig; ob sie genutzt werden können erweist sich zumeist innerhalb von zwei Wochen. Beide Plastizitätsphänomene betreffen die binaurale Integration. Die erste Station entlang der aufsteigenden Hörbahn, in der binaurale Integration stattfinden kann, ist der Obere Olivenkomplex. Die Olive empfängt nicht nur Signale vom Ohr über den Cochleariskern, sie projiziert auch zurück ins Ohr, und zwar entlang zweier Hauptsysteme: dem lateralen olivocochleären oder LOC System und dem medialen olivocochleären oder MOC System. Material und Methoden Wir haben unsere Untersuchungen an der adulten Ratte durchgeführt und zwei Ertaubungsmodelle eingesetzt: das einseitige Schalltrauma (Michler und Illing, 2002) und die Cochleotomie (Illing et al., 1997). Immunhistochemie wurde unter Einsatz von Antikörpern gegen Neurofilamente, Calretinin, Synaptophysin und das wachstumsassoziierte Protein GAP43 ausgeführt (Illing et al., 1997). Der Nachweis von GAP-43 mRNA erfolgte durch in situ-Hybridisierung mit einer etwa 1 kb langen Sonde (Illing et al., 1999). Als axonale Tracer wurden Meerrettich-Peroxidase (Illing et al., 1997), Fast Blue und Diamidino Yellow (Horváth et al., 2000) verwendet. Läsionen der Oberen Olive wurden durch stereotaktische Injektionen von Kainat erzielt (Kraus und Illing, 2004). Ergebnisse Eine partielle Schädigung der Cochlea, wie sie durch ein Schalltrauma entstehen kann, führt zu einem Verlust eines Teils ihrer zentralen Projektion (Michler und Illing, 2003). Eine Cochleotomie führt dagegen zu einem vollständigen Verlust des Spiralganglions und seiner Axone im Cochleariskern, also zu einer totalen Taubheit. Im Cochleariskern erkennt man dies an der Degeneration der Nervenfasern (Illing et al., 1997). In beiden Fällen, sowohl nach Schalltrauma als auch nach Cochelotomie, kommt es im Cochleariskern aber nicht nur zu Phänomenen der Degeneration, sondern auch zu Rekonstruktionsprozessen. Um Rekonstruktionsprozesse zu identifizieren bedienen wir uns molekularer Marker. Das wachstumsund plastizitätsassoziierte Protein GAP-43 ist ein solcher Marker, der von Neuronen exprimiert wird, die sich im Zustand der Axogenese und Synaptogenese befinden (Benowitz und Routenberg, 1997). Im Cochleariskern der Ratte auf der Seite einer Cochleotomie ist nach 7 Tagen Überlebenszeit ein deutlicher Anstieg der GAP-43 Immunoreaktivität eingetreten. Diese Zunahme erwies sich nach einer Quantifizierung durch densitometrische Messungen als statistisch signifikant. Natürlich wissen wir damit noch nichts darüber, in welchen zellulären Elementen GAP-43 lokalisiert ist. Bei hoher lichtmikroskopischer Vergrößerung finden wir GAP-43 nicht in Zellkörpern, nicht in Dendriten, auch kaum in Axonen, sondern vor allem in Boutons. Mit dem Lichtmikroskop können wir jedoch nicht nachweisen, ob diese Boutons tatsächlich präsynaptische Endigungen sind, deshalb haben wir das Elektronenmikroskop eingesetzt. Dabei hat sich der Verdacht bestätigt, dass GAP-43 vor allem in präsynaptischen Endigungen lokalisiert ist. Vergleicht man die Dichte der GAP-43 positiven Boutons im Cochleariskern mit der Gesamtpopulation präsynaptischer Endigungen, dargestellt durch das synaptische Protein Synaptohysin, erkennt man, dass der prozentuale Anteil der Synapsen, die sich nach einer Cochleotomie durch GAP-43 anfärben lassen, erheblich ist. Nun war natürlich von Interesse herauszufinden, woher das GAP-43 der präsynaptischen Endigungen im Cochleariskern stammt, d.h. in welchen Zellkör- Die Plastizität des auditorischen Hirnstamms nach einseitiger Schwerhörigkeit oder Ertaubung 1 7. DGA Jahrestagung 2004 pern es produziert wird. Aus dem Cochleariskern selbst stammt es nicht: es gibt dort keine GAP-43 positiven Zellkörper. Es gibt dort auch keine Zellkörper, die nach einer Cochleotomie GAP-43 mRNA produzieren (Illing et al., 1999). Im Gegensatz zum Cochleariskern gibt es aber GAP-43 Produzenten in der lateralen Oberen Olive. Sieben Tage nach einer Cochleotomie waren dort GAP-43 positive Nervenzellkörper erkennbar. Nach einer Cochleotomie war auch die GAP-43 mRNA in einigen Neuronen besonders stark exprimiert. Diese Neurone erwiesen sich in Doppelfärbungsexperimenten als identisch mit den immunreaktiven Neuronen (Illing et al., 1999). Wir konnten auch zeigen, dass es sich bei diesen Neuronen um LOC-Neurone handelt, denn sie konnten gleichzeitig mit dem axonalen Tracer MeerettichPeroxidase aus der Cochlea markiert werden. Als Ursprung für die im Cochleariskern beobachtete ertaubungsabhängige GAP-43 Expression kämen die LOC-Neurone jedoch nur in Frage, wenn sie Kollaterale in dieses Kerngebiet sendeten. Das tun sie im normalen erwachsenen Tier jedoch nicht (Horváth et al., 2000). Wir haben vor einer Cochleotomie einen gelbfluoreszierenden Tracer (Diamidino Yellow) in die Cochlea und einen blaufluoreszierenden axonalen Tracer (Fast Blue) in den Cochleariskern injiziert. Olivocochleäre Zellen mit Axonkollateralen in den Cochleariskern müssten jetzt doppelt gefärbt werden. LOC-Neurone waren aber nur in extrem wenigen Fällen (durchschnittlich nur ein Neuron pro Experiment) doppelt markiert. Nun wäre es denkbar, dass sie solche Axonkollaterale infolge einer Ertaubung auswachsen lassen. Wir haben aber zeigen können, dass dies nicht der Fall ist. Ihre Anzahl hat auch nach einer Cochleotomie nicht zugenommen (Kraus und Illing, 2004), also können LOC Neurone nicht für die Zunahme von GAP-43 im Cochleariskern nach einer Ertaubung verantwortlich sein. Anstelle doppelt markierter LOC-Neurone haben wir in diesen Experimenten zahlreiche doppeltmarkierte MOC-Neurone gefunden (Horváth et al., 2000). Diese Zellen erwiesen sich nach einer Cochleotomie auch als positiv für GAP-43 mRNA. Wenn nun MOCNeurone für die im Cochleariskern beobachtete ertaubungsabhängige GAP-43 Zunahme verantwortlich wären, müsste diese ausbleiben, wenn zuvor die MOC-Neurone entfernt würden. Diesen Zusammenhang haben wir durch lokale Injektion des Nervengiftes Kainat in den Oberen Olivenkomplex versucht zu beantworten. Die unmittelbare Wirkung solcher Injektionen lässt sich am Verlust neuronaler Zellkörper überprüfen. In einer Serie solcher Experimente haben wir verschiedene Teile des oberen Olivenkomplexes lädiert, und zwar jeweils vor einer gleichseitigen oder gegenüberliegenden Cochleotomie, oder die Läsion der Olive erfolgte bilateral (Kraus und Illing 2004). Die Wirkung des Verlustes von MOC-Neuronen auf die GAP-43 Expression im VCN war sofort erkennbar. Mit densitometrischen Messungen haben wir zeigen können, dass es vor allem die Projektion von den contralateralen MOC-Neuronen ist, über die GAP-43 in den Cochleariskern gelangt, aber erst im Falle einer bilateralen Läsion der Olive bleibt die GAP-43 Expression infolge einer Cochleotomie völlig aus. Für die Zunahme von GAP-43 im Cochleariskern nach Ertaubung sind also MOC-Neurone verantwortlich. Wir wissen nun, welcher Umstand die ertaubungsabhängige GAP-43 Expression im Cochleariskern induziert und von welchen Neuronen es dorthin gebracht wird. Wir wissen aber noch nicht, ob die GAP43 positiven präsynaptischen Endigungen schon vor der Cochleotomie vorhanden waren und jetzt modifiziert werden, oder ob es Neubildungen synaptischer Kontrakte sind. Wir glauben zeigen zu können, dass das Letztere der Fall ist. Nach einer Cochleotomie erscheinen im gleichseitigen Cochleariskern GAP-43 positive unreife Nervenfasern, die mit Vesikeln gefüllt sind. In günstigen Fällen erkennt man sogar voll ausgebildete Wachstumskegel, deutlich zu erkennen an ihren Filopodien. Zahlreiche dieser Filopodien enthalten GAP-43. Mit diesen Filopodien suchen Wachstumskegel nach geeigneten postsynaptischen Profilen für die Ausbildung synaptischer Kontakte. Können wir schließlich etwas über die postsynaptischen Zellen aussagen, an denen diese neuen präsynaptischen Endigungen sitzen? Wir haben gefunden, dass GAP-43 positive Boutons nur an Zellkörpern sitzen, die den Neurotransmitter Glutamat oder das Kalzium-bindende Protein Calretinin enthalten, aber nicht an GABAergen Neuronen (Illing et al., 1997). Der Verlust sensorischer Erregung infolge einer Ertaubung führt im Cochleariskern also offenkundig zu einer Neubildung von Synapsen vorwiegend an erregenden Neuronen. Diskussion Auf der Grundlage dieser Befunde können wir eine hypothetische Erklärung für das eingangs erwähnte Phänomen der Korrektur des Richtungshörens geben. Richtungshören basiert u.a. auf dem Lautstärkevergleich beider Ohren. MOC-Neurone erhalten Signale von den Cochleariskernen beider Seiten. Wenn die sensorische Aktivierung im Cochleariskern auf der einen Seite nachlässt, bilden die cholinergen MOCNeurone dort neue synaptische Kontakte an exzitatorischen Neuronen. Dies könnte dazu dienen, die binaurale Integration wieder auszubalancieren und ein korrektes Muster neuronaler Aktivität trotz asymmetrischen Hörens in der aufsteigenden Hörbahn wieder herzustellen. Dass dieser Vorgang nur dann zum Erfolg führen kann, wenn auf dem beeinträchtigten Ohr ein Resthören besteht, ist offenkundig. Zusammenfassung Nach Schalltrauma oder Cochleotomie kommt es zu einer massiven Expression von GAP-43 im ventralen Cochleariskern. GAP-43 ist in zahlreichen präsynaptischen Endigungen lokalisiert, deren Ursprungszellen nicht im Cochleariskern liegen. Nach Cochleotomie kommt es zur Hochregulation von GAP-43 mRNA nur in den Zellkörpern olivocochleärer Neuro- Die Plastizität des auditorischen Hirnstamms nach einseitiger Schwerhörigkeit oder Ertaubung 2 7. DGA Jahrestagung 2004 nen des Oberen Olivenkomplexes; diese Zellen müssen deshalb Ursprung des GAP-43 Proteins im VCN sein. LOC-Neurone entsenden keine Axonkollaterale in den VCN; sie entwickeln auch keine nach einer Cochleotomie, fallen also als Ursprung für die GAP43 Expression im VCN aus. MOC-Neurone entsenden Axonkollaterale in den Cochleariskern. Werden die MOC-Neurone vor der Cochleotomie entfernt, bleibt die GAP-43 Expression im VCN aus. Dieser Befund veranlasst uns zu der Schlussfolgerung, dass MOCNeurone Ursprung der ertaubungsabhängigen GAP-43 Expression im VCN sind. Infolge einer Ertaubung kommt es zur GAP-43 Expression nicht nur in bestehenden, sondern auch in neu gebildeten präsynaptischen Endigungen, wie das Erscheinen unreifer Neuriten und Wachstumskegeln anzeigt. GAP-43 positive Synapsen finden sich an glutamatergen, aber nicht an GABAergen postsynaptischen Profilen, d.h. die Reinnervation ist zielgerichtet. Die ertaubungsbedingte Reorganisation des Cochleariskerns führt offenbar zu einer selektiven Innervation exzitatorischer Neurone, möglicherweise als Kompensation für die ausbleibende sensorische Stimulation. Literatur Benowitz LI, Routtenberg A. 1997. GAP-43: an intrinsic determinant of neuronal development and plasticity. Trends Neurosci. 20: 84-91. Florentine M. 1976. Relationship between lateralization and loudness in asymmetrical hearing losses. J. Amer. Audiol. Soc. 1: 243-251. Horváth M, Kraus KS, Illing RB. 2000. Olivocochlear neurons sending axon collaterals into the ventral cochlear nucleus of the rat. J. Comp. Neurol. 422: 95-105. Illing RB, Cao QL, Förster CR, Laszig R. 1999. Auditory brainstem: development and plasticity of GAP-43 mRNA expression in the rat. J. Comp. Neurol. 412: 353-372. Illing RB, Horváth M, Laszig R. 1997. Plasticity of the auditory brainstem: effects of cochlear lesions on GAP-43 immunoreactivity in the rat. J. Comp. Neurol. 382: 116-138. Kraus KS, Illing RB. 2004. Superior olivary contributions to auditory system plasticity: medial but not lateral olivocochlear neurons are the source of cochleotomy-induced GAP-43 expression in the ventral cochlear nucleus. J. Comp. Neurol. 480, im Druck. Michler SA, Illing RB. 2002. Acoustic trauma induces re-emergence of the growth and plasticity associated protein GAP-43 in the rat auditory brainstem. J. Comp. Neurol., 451: 250-266. Michler SA, Illing RB. 2003. Modulation of plasticity related molecules in the auditory brainstem of the rat following acoustic overstimulation. Audiol. Neurootol. 8: 190-206. Die Plastizität des auditorischen Hirnstamms nach einseitiger Schwerhörigkeit oder Ertaubung 3