Page as PDF - Max-Planck
Werbung

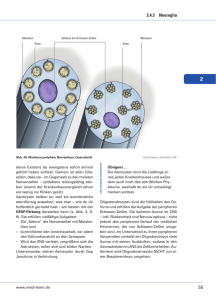
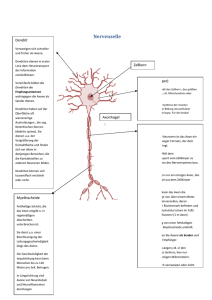
Jahrbuch 2013/2014 | Nave, Klaus-Armin | Neu entdeckte Funktion von Oligodendrozyten im zentralen Nervensystem Neu entdeckte Funktion von Oligodendrozyten im zentralen Nervensystem Novel function of oligodendrocytes in the central nervous system Nave, Klaus-Armin Max-Planck-Institut für experimentelle Medizin, Göttingen Korrespondierender Autor E-Mail: [email protected] Zusammenfassung Die Myelinisierung neuronaler Axone durch Oligodendrozyten dient der Erhöhung der Leitgeschw indigkeit und gehört zu den komplexesten Interaktionen von Zellen im zentralen Nervensystem. W issenschaftler des MPI entdeckten eine neue Funktion der Oligodendrozyten, die Unterstützung des Stoffw echsels von Axonen. Genetische Experimente in der Maus zeigen, dass es sich dabei um den Transfer von energiereichen Metaboliten handelt, die von glykolytischen Oligodendrozyten am Axon freigesetzt w erden. Bei erblichen und erw orbenen Myelinerkrankungen spielt dieser Transfer eine w ichtige Rolle. Summary One of the most spectacular interactions of cells in the central nervous system is the myelination of axons by oligodendrocytes, w hich serves the rapid impulse propagation. Max Planck scientists have discovered a novel function of oligodendrocytes in the metabolic support of myelinated axons. Genetic evidence suggests that oligodendrocytes are glycolytic cells that release lactate as an energy-rich metabolite. Loss of this trophic function of oligodendrocytes may play a critical role in inherited and acquired myelin diseases. Einleitung Das Nervensystem ist ein Netzw erk aus Milliarden von Nervenzellen (Neuronen). Diese kommunizieren über lange Fortsätze (Axone) und die Information w ird in Form von elektrischen Impulsen übertragen, die entlang der Axone zu w eit entfernten Synapsen auf Zielneuronen gelangen. Myelin, eine mehrlagige Membran, isoliert das Axon elektrisch und erhöht damit seine Leitgeschw indigkeit auf bis zu 100 Meter pro Sekunde [1]. Myelin w ird von hochspezialisierten Gliazellen, den Oligodendrozyten, gebildet, die ihre eigene Zellmembran spiralig um jew eils kurze Abschnitte des Axons w ickeln – diese Umw icklung bildet dann die kompakte Struktur der Myelinscheide [2]. Die Myelinscheiden können bis zu einem halben Millimeter lang w erden und noch deutlich länger im peripheren Nervensystem, w o eine vergleichbare Aufgabe von den sog. Schw annzellen übernommen w ird. Zw ischen den einzelnen myelinisierten Segmenten © 2014 Max-Planck-Gesellschaft bleiben "Ranvier'sche w w w .mpg.de Schnürringe" offen, an denen elektrische 1/5 Jahrbuch 2013/2014 | Nave, Klaus-Armin | Neu entdeckte Funktion von Oligodendrozyten im zentralen Nervensystem Aktionspotenziale entstehen. Die Myelinisierung langer Axone durch Oligodendrozyten erlaubt es dem Gehirn, sehr viel schneller zu arbeiten und gleichzeitig eine sehr kompakte Bauw eise zu behalten. Ohne Myelinisierung w äre die Leistung des menschlichen Gehirns nicht denkbar. Ein Großteil der myelinisierten Axone erkennt man als "w eiße Substanz" unterhalb der Großhirnrinde (Cortex), die etw a 50 Prozent des menschlichen Gehirns ausmacht [1]. Störungen der Myelinisierung führen zu schw ersten Entw icklungsstörungen (Leukodystrophien) und zur Beeinträchtigung w esentlicher motorischer, sensorischer und kognitiver Fähigkeiten. Krankheiten, die das Myelin in der w eißen Substanz angreifen, können auch später erw orben w erden, w ie zum Beispiel die Multiple Sklerose. Die Erforschung der Biologie des Myelins ist auch vom W unsch geprägt, entsprechende neurologische und psychiatrische Krankheiten besser zu verstehen. Lange Axone und Myelin entstehen durch extreme zelluläre Spezialisierung Allein aufgrund der Länge ihrer Axone sind die Neurone extrem spezialisierte Zellen. Ein typisches Neuron misst nur 15 Mikrometer im Durchmesser, aber sein nur ein Mikrometer starkes Axon kann einen Meter lang sein, d. h. w eit mehr als 99 Prozent des Zellvolumens sind im Axon lokalisiert – und damit in w eiter Entfernung vom Zellkörper, in dem fortw ährend die genetische Information ausgelesen w ird. Da aber neuronale Informationsübertragungen am Ende des Axons geschehen, muss das Neuron über diese Entfernung Synapsen mit Proteinen beliefern. Die elektrischen Signale des Axons sow ie die Transportvorgänge innerhalb der Axone kosten Energie. Diese muss aus der Verbrennung energiereicher Substanzen (z. B. Glukose) gew onnen w erden, die dem Gehirn über den Kreislauf zufließen. Schon kurzzeitige Energiedefizite können zum Funktionsverlust eines Axons führen. Lange Axone sind deshalb eine Art "Flaschenhals" für die Funktion des Nervensystems. In allen Entw icklungsstufen des Gehirns sind Axone mit Gliazellen assoziiert. Zw ar gab es die Vermutung, dass Gliazellen die Neurone irgendw ie "unterstützen", aber die zugrunde liegenden Mechanismen w aren kaum bekannt [3]. Die Myelinisierung Oligodendrozyten und Erregungsleitung myelinisierter von Schw annzellen Axone Axonen (in w ar praktisch peripheren gehört die Nerven). Die sogar zu den einzige gesicherte "saltatorische", Funktion also bestverstandenen von sprunghafte Konzepten der Neurophysiologie. Aber der genaue Ablauf der Myelinisierung – die vielleicht spektakulärste Interaktion von zw ei Zellen im Nervensystem überhaupt – w ar bis vor kurzem unverstanden. Arbeiten in der Gruppe von Mikael Simons am MPI für experimentelle Medizin haben jetzt durch die Kombination verschiedener licht- und elektronenmikroskopischer Techniken in unterschiedlichen Systemen (Fisch, Maus und Zellkultur) diesen spiraligen Wachstumsvorgang der Oligodendrozytenfortsätze bis ins Detail aufklären können [2]. Dabei ist die Myelinbildung essenziell an eine hohe Konzentration von zellulärem Cholesterin geknüpft [4]. Die Myelinscheide des Axons stellt aber nicht nur eine elektrische, sondern auch eine effiziente physische Isolierung dar. Die axonale Oberfläche ist über 99 Prozent von Myelin abgedeckt. Deshalb ist es nicht leicht verständlich, w ie etw a die Energiegew innung eines myelinisierten Axons aufrechterhalten w ird, w enn der Zustrom von Metaboliten durch das Myelin selbst ausgebremst w ird [3]. Oligodendrozyten produzieren mehr als Myelin Myelin ist eine mehrlagige kompakte Struktur, die aus der oligodendroglialen Zellmembran hervorgeht, aber daneben gibt es ein Netzw erk von Kanälen aus nicht kompaktem Myelin, das die Myelinscheide begleitet und eine Fortsetzung des Zytosols der Oligodendrozyten darstellt [3]. Es handelt sich dabei um die gleichen © 2014 Max-Planck-Gesellschaft w w w .mpg.de 2/5 Jahrbuch 2013/2014 | Nave, Klaus-Armin | Neu entdeckte Funktion von Oligodendrozyten im zentralen Nervensystem Kanäle, die in der Entw icklung den erforderlichen Transport von Membranvesikeln zur Wachstumszone des Myelins an der Spitze des Gliazellfortsatzes ermöglichen, der sich spiralig um das Axon verlängert [2]. Diese Beobachtungen w urden erst durch Methoden der Hochdruck-Kryopräparation von myelinisierten Nerven möglich. Mit dieser Methode lassen sich die meisten Artefakte einer chemischen Fixierung und konventionellen Probeneinbettung vermeiden, unter anderem der Kollaps dieses ausgedehnten Systems zytosolischer Kanäle im Myelin [2; 3]. A bb. 1: O ligode ndrozyte n m ye linisie re n Ax one – hie r im Q ue rschnitt ge ze ichne t – und ve rfüge n übe r Monoca rbox yla tTra nsporte r (MC T1) für die Ve rsorgung von Ax one n m it Glyk olyse produk te n. © Ma x -P la nck -Institut für e x pe rim e nte lle Me dizin / Na ve ; a us [9] Durch die zytosolischen Kanäle des Myelins, die auch im adulten Nervensystem zum Teil w eiter bestehen, bleibt die Oligodendrozyte metabolisch mit dem Innenraum der Myelinscheide, der inneren Lippe verbunden, der dem Axon eng anliegt. Dort kommt es zum Übertritt von Metaboliten aus dem Zytosol der Oligodendrozyten in das Lumen des Axons. An dieser Stelle w urden mit Hilfe immun-elektronenmikroskopischer Techniken zum Beispiel Monocarboxylat-Transporter (MCT) lokalisiert, die dem Austausch von Glykolyseprodukten, etw a der Milchsäure (Laktat), dienen. Tatsächlich haben die W issenschaftler um Klaus-Armin Nave in den letzten Jahren eine Modellvorstellung entw ickelt, nach der Oligodendrozyten über zytosolische Kanäle myelinisierte Axone mit energiereichen Metaboliten versorgen können [3]. Sie gehen davon aus, dass die myelinisierenden Gliazellen aus dem Extrazellularraum Glukose aufnehmen, die z. B. über die Bluthirnschranke und Astrozyten dorthin gelangt ist. Glukose kann allerdings nicht direkt an das Axon w eitergegeben w erden, sondern w ird zunächst durch oligodendrogliale Enzyme zerlegt (Glykolyse). Die dabei entstehende Milchsäure diffundiert durch die zytosolischen Kanäle bis an das Axon heran und gelangt durch die Monocarboxylat-Transporter in das Axon hinein, in dem die eigentliche Energiegew innung (ATP) durch neuronale Mitochondrien erfolgt. Der Übertritt von Milchsäure aus der Oligodendrozyte in das myelinisierte Axon ist technisch nur schw er nachzuw eisen, da es sich um Distanzen im Nanometerbereich handelt und die Metaboliten nicht markiert gemacht w erden können. Die Göttinger W issenschaftler konnten aber diesen Mechanismus mit Hilfe eines genetischen Tricks indirekt darstellen. Dazu w urden Mausmutanten erzeugt, in denen Oligodendrozyten selektiv die Fähigkeit verloren haben, die bei der Glykolyse entstehende Milchsäure in eigenen Mitochondrien zur Energiegew innung zu nutzen (Inaktivierung des COX10-Gens). Überraschenderw eise sind diese Mäuse mit © 2014 Max-Planck-Gesellschaft w w w .mpg.de 3/5 Jahrbuch 2013/2014 | Nave, Klaus-Armin | Neu entdeckte Funktion von Oligodendrozyten im zentralen Nervensystem ihren mutanten Oligodendrozyten völlig unauffällig und langlebig, w as den Schluss nahelegt, dass die reifen Oligodendrozyten ohnehin keinen großen Bedarf an mitochondrialer Energiegew innung haben, sondern ihren Energiebedarf in Form von ATP aus der Glykolyse selbst beziehen können. Dafür müssen sie aber in der Lage sein, die entstandene Milchsäure abzugeben, w as durch Diffusion in die myelinisierten Axone geschieht. Wenn man experimentell die w eitere Verarbeitung dieser Milchsäure im Gehirn blockiert, w as z. B. durch die Gabe von bestimmten Anästhetika (Isofluran) der Fall ist, dann erhöht sich der Milchsäurespiegel im Gehirn. Dieser kann durch Methoden der Protonen-Magnetresonanzspektroskopie (3 H-MRS) relativ genau bestimmt w erden und liegt unter den Bedingungen einer solchen Anästhesie bei den Mausmutanten des COX10-Gens fast doppelt so hoch w ie in Tieren ohne Mutation [5]. W ird das Anästhetikum daraufhin gew echselt und die unspezifische Hemmung der mitochondrialen Atmung fällt w eg, sinkt der messbare Milchsäurespiegel innerhalb w eniger Minuten in allen Gehirnen auf nicht nachw eisbare Konzentrationen herab, und zw ar unabhängig vom Genotyp der Tiere (Abb. 1). Das bedeutet, dass die fortw ährend in Oligodendrozyten entstehende Milchsäure sehr schnell in den Mitochondrien derjenigen Zellen, die mit Oligodendrozyten metabolisch gekoppelt sind, abgebaut w ird. Dies sind zumindest teilw eise die myelinisierten Axone, w ie experimentell an isolierten Präparaten des optischen Nervs gezeigt w urde [5]. Damit w urde eine völlig neuartige Funktion von Oligodendrozyten im zentralen Nervensystem sehr w ahrscheinlich, nämlich die der direkten metabolischen Unterstützung von elektrisch aktiven Axonen. Die Bedeutung dieses Systems für die Funktion und das physische Überleben der myelinisierten Axone im Gehirn w urde in der Arbeitsgruppe von Jeff Rothstein (Baltimore) auch genetisch bestätigt. Heterozygote Mausmutanten, denen eine Kopie des Gens für den Laktat-Transporter MCT1 fehlt und die deshalb 50 Prozent w eniger Laktat an Axone abgeben können, entw ickeln zunehmend axonale Schäden und Degeneration [6]. Diese Neuropathologie erinnert dabei stark an die früheren Befunde im Mausmodell einer Leukodystrophie des Menschen (Pelizaeus-Merzbacher Krankheit), bei der sekundär axonale Schw ellungen auftraten und Degenerationen von Axonen zu einem unheilbar progressiven Krankheitsverlauf führten [7; 8]. Literaturhinweise [1] Nave, K.-A. Myelination and support of axonal integrity by glia Nature 468, 244-252 (2010) [2] Snaidero, N.; Möbius, W.; Czopka, T.; Hekking, L.H.P.; Mathisen, C.; Verkleij, D.; Goebbels, S.; Edgar, J.; Merkler, D.; Lyons, D. A.; Nave, K.-A.; Simons, M. Myelin membrane wrapping of CNS axons by PI(3,4,5)P3-dependent polarized growth at the inner tongue Cell 156, 277–290 (2014) [3] Nave, K.-A. Myelination and the trophic support of long axons Nature Review s Neuroscience 11, 275-283 (2010) © 2014 Max-Planck-Gesellschaft w w w .mpg.de 4/5 Jahrbuch 2013/2014 | Nave, Klaus-Armin | Neu entdeckte Funktion von Oligodendrozyten im zentralen Nervensystem [4] Saher,G.; Brugger, B.; Lappe-Siefke, C.; Möbius, W.; Tozawa ,R.; Wehr, M. C.; Wieland, F.; Ishibashi, S.; Nave, K.-A. High cholesterol level is essential for myelin membrane growth Nature Neuroscience 8, 468-475 (2005) [5] Fünfschilling, U.; Supplie, L. M.; Mahad, D.; Boretius, S.; Saab, A. S.; Edgar, J.; Brinkmann, B. G.; Kassmann, C. M.; Tzvetanova, I. D.; Möbius, W.; Diaz, F.; Meijer, D.; Suter, U.; Hamprecht, B.; Sereda, M. W.; Moraes, C. T.; Frahm, J.; Goebbels, S., Nave, K.-A. Glycolytic oligodendrocytes maintain myelin and long-term axonal integrity Nature 485, 517-521 (2012) [6] Lee, Y . et al. Oligodendroglia metabolically support axons and contribute to neurodegeneration Nature 487, 443-448 (2012) [7] Griffiths, I.; Klugmann, M.; Anderson, T.; Y ool, D.; Thomson, C.; Schwab, M. H.; Schneider, A.; Zimmermann, F.; McCulloch, M.; Nadon, N.; Nave, K.-A. Axonal swellings and degeneration in mice lacking the major proteolipid of myelin Science 280, 1610-1613 (1998) [8] Saher, G.; Rudolphi, F.; Corthals, K.; Ruhwedel, T.; Schmidt, K. F.; Löwel, S.; Dibaj, P.; Barrette, B.; Möbius, W.; Nave, K.-A. Therapy of Pelizaeus-Merzbacher disease in mice by feeding a cholesterol-enriched diet Nature Medicine 18, 1130-1135 (2012) [9] Saab, A.; Tzvetanova, I.; Nave, K.-A. The role of myelin and oligodendrocytes in axonal energy metabolism Current Opinion in Neurobiology 23, 1065-1072 (2013) © 2014 Max-Planck-Gesellschaft w w w .mpg.de 5/5