Musterseiten 075-077
Werbung

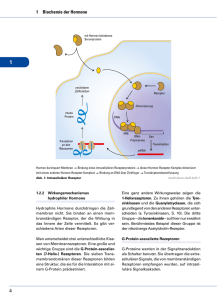
3.2 Synapsen – Rezeptoren – Wirkungen von Rezeptorbesetzung 3.2 Synapsen – Rezeptoren – Wirkungen von Rezeptorbesetzung ■■ Struktur von Synapsen. Als Synapse bezeichnet man die Verbindung einer Nervenzelle mit einer anderen Zelle, z. B. einem weiteren Neuron. Die Endknöpfchen des ersten legen sich an der Membran eines zweiten (postsynaptischen) Neurons an; dies geschieht zumeist an dessen Dendriten (axodendritische Synapsen), seltener am Körper des Neurons (axosomatische Synapsen) oder an seinem Axon nahe der Endknöpfchen (axoaxonale Synapsen); letztere bilden die anatomische Grundlage der präsynaptischen Hemmung. ■■ Allgemeines zur synaptischen Übertragung, zu Rezeptoren und Ionenkanälen. Bei den hier allein betrachteten chemischen Synapsen ist der Spalt zwischen der Membran des zuerst erregten (präsynaptischen) Neurons und der postsynaptischen Nervenzelle so breit, dass die elektrische Erregung nicht unmittelbar überspringen kann. Die Übertragung geschieht chemisch mithilfe von Neurotransmittern (kürzer: Transmitter; zu Deutsch Botenstoffe). Diese werden im präsynaptischen Neuron aufge­baut und in den Vesikeln der Endknöpfchen gespeichert. Kommt eine Erregung (das Ak­tions­potenzial) dort an, so werden die Ve­ sikel nach vorne gestoßen und entleeren die Moleküle in den synaptischen Spalt. Die frei­ gesetzten Botenstoffe diffundieren zu einem gewissen Prozentsatz zur Membran des postsynaptischen Neurons, wo sie sich an Rezeptoren (genau auf die Art des Transmitters zugeschnittene Bindungsstellen) anlagern können. Üblicherweise schüttet eine Nervenzelle nur einen Typ von (niedrigmolekularem) Transmitter aus, und nur für diesen liegen an der postsynaptischen Zellmembran passende Bindungsstellen bereit. Diese Anlagerung ist ein sehr kurzfristiger Vorgang; ausgesprochen irreführend ist die 75 Vorstellung, dass Transmitter und Rezeptor nun eine unlösliche Verbindung eingehen würden, dass der Botenstoff vom Rezeptor regelrecht »verschluckt« würde. Der kurze Kontakt reicht jedoch aus für Veränderungen der direkt benachbarten Zellmembran. Diese Rezeptoren kontrollieren nämlich Ionenkanäle, Öffnungen in der Membran, durch die elek­ trisch geladene Teilchen nach innen oder außen strömen können; die wichtigsten beweglichen Ionen sind Na+, K+, Ca2+ und Cl–. Die dafür geeigneten Kanäle heißen entsprechend Natrium-, Kalium-, Calcium- oder Chloridkanäle, wobei es unter einer Synapse immer nur einen Typ gibt, der von diesem einen dort vorhandenen Rezeptortyp kontrolliert wird. Rezeptoren, durch deren Besetzung sich Na+Kanäle und Ca 2+-Kanäle öffnen, sind erregend: Die vornehmlich extrazellulär lokalisierten, positiv geladenen Natrium- und Calciumionen strömen dann ein und machen das negative Zellinnere geringfügig weniger negativ (Depolarisation oder exzitatorisches postsynaptisches Potenzial = EPSP); die Erregbarkeit der Zelle nimmt damit zu (daher die Bezeichnung »erregend« für den zugehörigen Rezeptor) Werden hingegen K+-Kanäle geöffnet, so können die vermehrt intrazellulär zu findenden Kaliumionen ausströmen und das Zellinnere wird noch stärker negativ (Hyperpolarisation, inhibitorisches postsynaptisches Potenzial = IPSP). Chloridionen befinden sich großteils extra­zellulär; die Öffnung von Chloridkanälen mit folgendem Einstrom führt daher ebenfalls zur Hyperpolarisation. K+- und Cl–-Kanäle kontrollierende Rezeptoren sind also hemmend. Da an einer Synapse normalerweise nur ein Typ von Rezeptor sitzt und dieser dann entweder erregend oder hemmend wirkt, gibt es erregende und hemmende Synapsen. Hingegen gibt es typischerweise keine erregenden und hemmenden Transmitter; viele sind an einer Synapse erregend, an einer anderen hemmend. Allerdings sind sämtliche bekannten Rezeptoren für Glutamat erregend (sie kontrollieren in 76 3 Erregungsbild und -übertragung in Neuronen – ­Psychopharmakotherapie aller Regel Calciumkanäle), sodass man Glu­ tamat als generell erregenden Transmitter bezeich­ nen kann; Rezeptoren für GammaAmino­buttersäure (GABA) kontrollieren ausschließlich Chloridkanäle; GABA ist also ein hemmender Transmitter. ■■Ionenkanal- und G-Protein-gekoppelte Re- zeptoren. Rezeptoren können den Öffnungsgrad von Ionenkanälen auf zwei verschiedene Arten kontrollieren. Seltener sind die Ionen­ kanal-gekoppelten oder ionotropen Rezeptoren, bei denen eine Andockung des Transmitters unmittelbar zur Öffnung des Kanals führt. Bei den häufigeren G-Protein-gebundenen (Second-Messenger-gekoppelten, metabotropen) Rezeptoren setzt nach Anlagerung des Transmitters ein komplizierter chemischer Prozess ein (auch als nachgeschaltete Signal­ transduktion bezeichnet). Es handelt sich um eine Kettenreaktion (sogenannte Second-Messenger-Kaskade), deren letztes Produkt, der Second Messenger, den Kanal öffnet; erster Schritt ist immer die Abspaltung eines G-Proteins vom Rezeptor (daher der Name G-Pro­ tein-gebundener Rezeptor). Verkürzt ist der Sachverhalt etwa so zu beschreiben: Das abgespaltene G-Protein aktiviert das Enzym Adenylatcyclase (Adenylylcyclase), welches die Bildung von cAMP (zyklisches Adenosinmonophosphat) fördert. Dieses setzt den weiteren Prozess der nachgeschalteten Signal­trans­ ! Nervenzellen sind – als Folge der unterschiedlichen Durchlässigkeit ihrer Mem­ bran für einzelne Ionenarten – gegenüber dem Extrazellulärraum im Ruhezustand negativ geladen (Ruhepotenzial von etwa –70 mV). Dieser Zustand wird jedoch laufend verändert, indem an den Synapsen als Resultat der dort eintreffenden elektrischen Erregungen aus den Endknöpfchen der präsynaptischen Zellen Transmitter freigesetzt werden. Diese gehen kurzfristi- duktion in Gang. cAMP wird wiederum durch das Enzym Phosphodiesterase abgebaut; Phosphodiesterasehemmer (z. B. Sildenafil [Viagra ]) erhöhen die Wirkung der Rezep­tor­be­set­zung, weil cAMP schlechter inaktiviert wird. Die zur Phasenprophylaxe affektiver Störungen eingesetzten Lithiumsalze schwächen umgekehrt durch Eingriff in die nachgeschaltete Signal­ trans­duk­tion die Wirkung von Rezeptorbesetzungen. ® ■■ Präsynaptische Autorezeptoren. Neben den Rezeptoren der postsynaptischen Membran gibt es präsynaptische Autorezeptoren, die an der Membran des präsynaptischen Neurons sitzen und auf den selbst ausgeschütteten Transmitter ansprechen. Die Anzahl der besetzten Autorezeptoren gibt Rückmeldung über die im synaptischen Spalt befindliche Transmittermenge, sodass im Sinne einer Rückkopplung die Produktion oder Ausschüttung von Transmittermolekülen verstärkt oder vermindert werden kann. Werden präsynaptische Autorezeptoren mit Stoffen besetzt, die dort keine Wirkung ausüben, können sich weniger Moleküle anlagern und das präsynaptische Neuron geht fälschlicherweise von einem Transmittermangel im Spalt aus. Auf einer Blockade präsynaptischer Autorezeptoren basiert teils die Wirkung von Antidepressiva, die so indirekt die Konzentrationen von Serotonin oder Noradrenalin im synaptischen Spalt erhöhen. ge Verbindungen mit bestimmten, auf die Transmitterart zugeschnittenen Eiweißkomplexen (den Rezeptoren) ein, worauf sich (postsynaptisch) ein für den Rezep­ tor­typ charakteristischer (von ihm kon­ trol­lierter) Ionenkanal öffnet. In der Umgebung der Rezeptoren verändern sich durch Ein- oder Ausströmen von Ionen die Ladungsverhältnisse. Wird beispielsweise ein Calciumkanal geöffnet (wie an bestimmten Glutamatrezeptoren), wan- 3.3 Transmitter ! dern positiv geladene Calciumionen ins Zellinnere und vermindern dort die Ne­ gativität (Depolarisation bzw. EPSP = ex­ zitato­risches postsynaptisches Potenzial). An anderen Rezeptoren kann als Folge der Ionenbewegungen eine Verstärkung der Negativität (Hyperpolarisation) resultieren (IPSP = inhibitorisches postsynaptisches Potenzial); die von den zahllosen Rezeptoren stammenden EPSPs und ­IPSPs wandern längs der Membran zum Axonhügel, wo sie miteinander verrechnet werden. Ist die Depolarisation dort genügend groß, kehren sich durch Einströmen von positiven Natriumionen kurzzeitig die Ladungs­ verhältnisse um (Aktionspotenzial = AP); dieses Aktionspotenzial wandert längs des Axons und seiner Kollateralen zu den Endknöpfchen (schnell und sprunghaft in myelinisierten Axonen, langsam und kontinuierlich in unmyelinisierten). 3.3Transmitter 3.3.1Allgemeines ■■ Überblick. Augenblicklich sind etwa 100 verschiedene Botenstoffe zwischen Nervenzellen bekannt; man kann sie grob in zwei Klassen einteilen: hochmolekulare Neurotransmitter niedrigmolekulare Neurotransmitter • • Die der ersten Klasse sind aus mehreren Molekülen zusammengesetzt; ihre wichtigste Gruppe bilden die Peptidtransmitter, welche aus aneinander geketteten Aminosäuren bestehen. Dazu gehören u. a. die endogenen Opioide – die bei der Schmerzkontrolle eine wichtige Rolle spielen – sowie die ebenfalls im Schmerzsystem als Transmitter fungierende Substanz P 77 Typischerweise setzen die Endknöpfchen der präsynaptischen Neurone an den Den­ driten oder Perikaryen der postsynaptischen Zelle an (axodendritische und axosomatische Synapsen), teils auch am Ende der Axone (axoaxonale Synapse; Grundlage der präsynaptischen Hemmung). Neben den postsynaptischen Rezeptoren sind Bindungsstellen am ausschüttenden Neuron zu finden; diese Autorezeptoren registrieren die Transmitterkonzentrationen im synaptischen Spalt und sind damit bei der Regelung der Ausschüttung von Bedeutung. Bei den Rezeptoren unterscheidet man ionotrope, bei denen eine Besetzung unmittelbar zur Öffnung des Ionenkanals führt, von den häufigeren metabotropen oder G-Protein-gekoppelten. Bei letzteren setzt die Andockung der Transmitter einen komplizierten (pharmakologisch gut beeinflussbaren) chemischen Prozess in Gang, der schließlich auf die Weitstellung des Kanals wirkt. und das auch als zentralnervöser Transmitter wirksame Cholecystokinin (welches zudem ein Hormon im Verdauungstrakt ist). Die niedrigmolekularen Transmitter bestehen aus einem einzigen Molekül bzw. sind, wie Acetylcholin, durch Verbindung zweier kleiner Moleküle hervorgegangen. Sie werden folgendermaßen eingeteilt: Aminosäuretransmitter (Glutamat, GABA, Glycin) Monoamintransmitter bzw. biogene Amine (Serotonin, Dopamin, Noradrenalin und das diesbezüglich weniger bedeutsame Adrenalin) Acetylcholin (das eine eigene Kategorie bildet) lösliche Gase • • • • Die Transmitter innerhalb der einzelnen Gruppen sind strukturell verwandt und wer-