Mustertexte 273 - Beck-Shop
Werbung

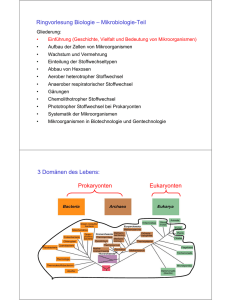
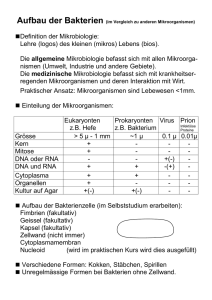
Biologie und molekulare Medizin für Mediziner und Naturwissenschaftler Bearbeitet von Monica Hirsch-Kauffmann, Manfred Schweiger, Michal-Ruth Schweiger 7. Auflage 2009. Buch. XIII, 416 S. Kartoniert ISBN 978 3 13 706507 4 Format (B x L): 19 x 27 cm Gewicht: 1350 g Weitere Fachgebiete > Medizin > Vorklinische Medizin: Grundlagenfächer > Physik, Chemie, Biologie für Mediziner Zu Inhaltsverzeichnis schnell und portofrei erhältlich bei Die Online-Fachbuchhandlung beck-shop.de ist spezialisiert auf Fachbücher, insbesondere Recht, Steuern und Wirtschaft. Im Sortiment finden Sie alle Medien (Bücher, Zeitschriften, CDs, eBooks, etc.) aller Verlage. Ergänzt wird das Programm durch Services wie Neuerscheinungsdienst oder Zusammenstellungen von Büchern zu Sonderpreisen. Der Shop führt mehr als 8 Millionen Produkte. 273 Mikrobiologie Zu den Mikroorganismen werden in der Mikrobiologie neben den Bakterien auch Viren, Pilze und unter Umständen auch größere Parasiten gerechnet. Mikroorganismen erlangen immer größere Bedeutung als Produzenten von wichtigen Wirkstoffen, als Komponenten der ökologischen Gleichgewichte, bei der Entwicklung der modernen Biotechnologie, aber auch für die Pathologie (Tab. 10.1). Wie aus dem Namen ersichtlich, beschäftigt sich die Mikrobiologie mit den Mikroorganismen (Abb. 10.1). Ganz sicher gehören dazu die Bakterien. Schon bei den Viren ist die Definition fraglich, da Viren keine autonomen Organismen sind. Trotzdem wird die Virologie zur Mikrobiologie gerechnet. Auch Pilze gehören hierher und Algen. Bei den Pilzen gibt es einige, die eine beträchtliche Größe erreichen können. Einzellige Algen können bis 20 cm groß sein (Acetabularia major). Auch Mehrzeller können zur Mikrobiologie gehören. Da viele der bisher erwähnten Organismen Parasiten des Menschen sind, die Größe eines Organismus aber kein gutes systematisches Kriterium ist, werden häufig auch die großen Parasiten der Mikrobiologie zugeschlagen (z. B. der Rinderbandwurm, der mehr als 10 Meter lang werden kann). Diese Tab. 10.1 Gruppen von Mikroorganismen mit medizinischer Bedeutung Einzeller Mehrzeller ▬ Bakterien ▬ Pilze ▬ DNAViren ▬ intrazelluläre Parasiten: Rickettsien Chlamydien Mycoplasmen ▬ mehrzellige Parasiten Würmer Insekten ▬ RNAViren Tumorviren ▬ Protozoen Protisten Thermo- Methanogene Halophile und anaerob abhängig von Acidophile hohem Salz Archaebakterien ▬ Protozoen: Amöben Flagellaten Ciliaten Sporozoen Zugehörigkeit wird auch dadurch dokumentiert, dass in fast allen Instituten für Mikrobiologie Abteilungen für Parasitologie existieren. Wegen der wachsenden Bedeutung der Parasitologie, auch im Zusammenhang mit Infektiologie und Tropenmedizin, wurde diesem Gebiet im vorliegenden Buch ein eigenes Kapitel gewidmet. Organismen der Mikrobiologie Animalia Protophyten pflanzenähnlich Viren Plantae Algen, Moose, Farne, blühende Pflanzen Abb. 10.1 Stellung der Mikroorganismen im Stammbaum der Organismen. Nur höhere Pflanzen und Tiere mit Ausnahme der Parasiten gehören nicht zur Mikrobiologie. Fungi Schleimpilze, Hefe Entwicklung der Chloroplasten einzellig mehrzellig Eukaryonten Monera (Prokaryonten) Cyanobakterien (Blau-Grün-Algen) Bakterien Eubakterien einzellig Entwicklung der Mitochondrien einzellig Ursprung aus: Hirsch-Kauffmann u.a., Biologie und molekulare Medizin (ISBN 9783137065074) 2009 Georg Thieme Verlag KG Mikrobiologie 10 10 274 10 Mikrobiologie 10.1 Prokaryonten sind kernlose Zellen Bakterien und Cyanobakterien (Blaugrünalgen) haben keinen Zellkern und werden Prokaryonten genannt. Organismen, deren Zellen Kerne besitzen, heißen Eukaryonten. Diese beiden Gruppen unterscheiden sich noch in anderen zellulären Strukturen (Rep. 10.1) und biochemisch z. B. in ihrer Empfindlichkeit gegenüber einigen Antibiotika. Repetitorium 10.1 Unterschiede zwischen Prokaryonten und Eukaryonten Prokaryonten Eukaryonten DNA im Zellkern nein ja Membran-begrenzte Organellen (Mitochondrien, Chloroplasten) nein ja Nucleosomen nein ja Zellwand (Murein-haltig) ja nein Flagellen Pili DNA Kapsel 70S Ribosomen Mesosom Plasmamembran Zellwand Reservestoffe (Glycogen, Lipide) Abb. 10.2 Aufbau einer Bakterienzelle. Aufgaben einiger Bestandteile: Flagellum: Fortbewegung Pili: Anheftung an Oberflächen Sex-Pili: Parasexualität Mesosom: Anheftungsstelle der DNA bei der Replikation, Konzentration von Nährstoffen Plasmamembran-Innenseite: Atmungsenzyme, DNA-Polymerase 10.1.1 Die Bakterienzellen haben Murein-haltige Zellwände Charakteristisch für die Bakterienzelle (Abb. 10.2, Abb. 10.3) ist die Zellwand (Tab. 10.2, Abb. 10.4) und in ihr ein zweidimensionales, sackartig die Zelle umgebendes Makromolekül, das Murein (Abb. 10.5). Beim Murein sind Kohlenhydratketten über Peptide miteinander verbunden. Die Kohlenhydratketten bestehen alternierend aus N-Acetylglucosamin und N-Acetylmuraminsäure. Die Peptidketten enthalten, neben den üblichen Aminosäuren L-Lysin und Glycin, auch D-Isoglutamin und D-Alanin, wobei die biologisch unüblichen D-Aminosäuren das Murein wohl vor dem Zugriff von Peptidasen schützen sollen. Diese komplexe Struktur wurde von dem deutschen Biochemiker Weidel aufgeklärt, der dem Sacculus den Namen Murein gab. Grampositive Bakterien besitzen neben einer besonders dicken, mehrschichtigen Mureinschicht Teichonsäure, ein ebenfalls komplexes Makromolekül (Abb. 10.4). Sie trägt die antigenen Eigenschaften der Bakterien. Die Aufgaben der Zellwand (Tab. 10.3) sind es, die Zelle zu schützen, die äußere Struktur der Zelle zu fixieren und sehr differenziell bestimmte Stoffe hineinzulassen und andere auszuschließen. Auch der osmotische Druck und die Ionenkonzentrationen im Inneren der Zelle werden durch die Zellwand aufrechterhalten. Während die mechanischen Schutzfunktionen von Murein ausgeübt werden, sind für die differenzielle Aufnahme bzw. Abgabe von Stoffen die Zellmembranen verantwortlich. Besonders deutlich werden die Aufgaben der Membranschichten in gramnegativen Zellen (Abb. 10.6). Die Phospholipid-Doppelschicht der Membranen ist praktisch undurchgängig für alle polaren Verbindungen wie z. B. für Ionen. Für deren Durchtritt gibt es in der äußeren Membran Poren und in der inneren Membran spezifische Transportproteine, die unter Energieverbrauch die notwendigen Stoffe aufnehmen bzw. abgeben. Bakterielle Proteine, die durch die innere Membran in den periplasmatischen Raum transportiert werden sollen, folgen ähnlichen Prinzipien wie die der Eukaryonten: die Peptide sind durch eine Signalsequenz markiert und werden im ungefalteten Zustand unter ATP-Verbrauch mittels eines Translocons transloziert. Einen speziellen Mechanismus haben pathogene Bakterien entwickelt: sie sind in der Lage, Toxine durch die innere und die äußere Membran durch eine „Röhre“, die u. a. von Pili gebildet wird, hinauszutransportieren. Proteine am äußersten Pilusende ermöglichen dann, z. B. beim Pestbakterium, den Kontakt zur Membran von Makrophagen, die durch das Toxin inaktiviert werden. Zwischen beiden Membranen liegt der periplasmatische Raum, in dem sich das Murein befindet. Ein Lipoprotein verankert die äußere Membran an dem Murein. Der Lipidanteil ist in die lipophile Schicht der Phospholipid-Doppelschicht eingelagert und der Proteinteil ist mit dem Murein verbunden. Auf der äußeren Seite der äußeren Membran sind Lipopolysaccharide (LPS) fixiert. Ein solches Lipopolysaccharid ist z. B. das Endotoxin, das die Bildung von Pyrogen induziert und dadurch Fieber hervorruft. aus: Hirsch-Kauffmann u.a., Biologie und molekulare Medizin (ISBN 9783137065074) 2009 Georg Thieme Verlag KG 275 10.1 Prokaryonten sind kernlose Zellen a Abb. 10.3 Bakterien im Elektronenmikroskop. a Gramnegatives Bakterium (Escherichia coli) mit Flagellen und Pili; dazu LambdaViren (Aufnahme: B. Menge, K. G. Lickfeld, Basel; M: Balken = 0,5 m). b Grampositive Bakterien (Bacillus subtilis) (Aufnahme: J. C. Benichon, Basel; M: Balken = 0,5 m). Kapsel oder Schleimschicht Zellwand (grampos., gramneg.) Cytoplasma Zellmembran Zellwandaufbau grampositiv gramnegativ Teichonsäure Lipopolysaccharid äußere Membran Murein Lipoprotein Abb. 10.4 Aufbau der Zellwand. Der Aufbau der Zellwand unterscheidet sich bei verschiedenen Bakterien. Grampositive: Eine Schicht aus einem Polysaccharid, der Teichonsäure, ist für Ionentransport und Antigenität verantwortlich. Eine mehrlagige Mureinschicht bietet mechanischen Schutz. Die Zellmembran, eine Phospholipid-Doppelschicht, bildet den Anschluss zum Cytoplasma hin. Bei den gramnegativen Bakterien übernimmt eine Schicht aus Lipopolysacchariden die Schutzfunktion, vermittelt Antigenität und enthält Toxine. Eine äußere Phospholipid-Doppelschicht wird über Lipoproteine mit der einlagigen Mureinschicht verankert. Alle drei äußeren Schichten behindern das Eindringen von Substanzen wie z. B. Penicillin oder Farbstoffe. Der Zellwand kann (nicht obligat) durch Sekretion eine Kapsel oder Schleimschicht aufgelagert werden, die Schutz vor Phagocytose bietet, wodurch das Bakterium seine Virulenz erhöht (s. z. B. Pneumokokken). Murein Zellmembran Zellmembran Das beim Zerfall der Bakterien frei werdende Endotoxin reagiert als exogenes Pyrogen mit Rezeptoren der Makrophagen, die auf diesen Reiz hin endogenes Pyrogen, das Interleukin I, ausschütten. Interleukin I setzt die Empfindlichkeit des Temperatur-Regulationszentrums im Hypothalamus herab, was zur Fieberreaktion führt. Noch weiter außen kann sich zum weiteren Schutz der Zelle eine Schleimschicht aus Polysacchariden oder Polypeptiden auflagern. Kapseln aus Polysaccharid-haltigen Schleimen oder manchmal aus Polypeptiden können Bakterien dem Zugriff des Wirtes entziehen. Sie können die Phagocytose verhindern (Abb. 10.4). aus: Hirsch-Kauffmann u.a., Biologie und molekulare Medizin (ISBN 9783137065074) 2009 Georg Thieme Verlag KG Mikrobiologie b 10 276 10 Mikrobiologie Tab. 10.2 Strukturen der Zellwand Tab. 10.3 Funktionen der Zellwand – Antigene – Schutz – Rezeptoren – Gestaltgebung – Sex-Pili – Osmoregulation – Bewegungsorganellen – selektive Stoffaufnahme und -abgabe – Proteine für Oberflächenadhärens a N-Acetylmuraminsäure b N-Acetylglucosamin Lysozym CH2OH O O CH2OH O O O O C C O OH NH H3C CH Kohlenhydratrückgrat NH O C O AS Gly AS Peptid-Brücke Gly Gly Gly Gly Kohlenhydratrückgrat AS AS Gly Glycin-Brücke c AS AS AS Peptid-Brücke CH3 CH3 Gly AS Glycin-Brücke Abb. 10.5 Murein. a Aufbau des Mureins: Kohlenhydratketten, die aus N-Acetylglucosamin und N-Acetylmuraminsäure bestehen, sind über Peptidbrücken zu einem zweidimensionalen Gerüst verbunden. b Chemische Strukturen im Murein. c Isolierte Mureinsacculi von Escherichia coli. Sie spiegeln die Gestalt der Zelle wider, aus der Beim Wachsen der Bakterien müssen natürlich auch die Schichten der Wand mitwachsen. Bei den Membranen werden neue Phospholipide eingelagert. Für die Vergrößerung des Murein-Sacculus müssen das zweidimensionale Netz geöffnet und neue Elementarstrukturen eingesetzt werden. Penicillin und ähnliche Antibiotika dieser Gruppe verhindern das Einsetzen der Elementarstrukturen, indem sie die Bildung der verknüpfenden Peptidbindungen unmöglich machen (Hemmung der Transpeptidase). Berücksichtigt man diese Tatsache, dann werden einige Besonderheiten der Wirkung von Penicillin klar: Nur Prokaryonten werden gehemmt, denn nur diese besitzen Murein. Nur wachsende Zellen sind Penicillin-empfindlich, denn nur diese öffnen das Murein. Sporen und metabolische Ruheformen sind unempfindlich, ebenso sogenannte zellwandlose L-Formen und Mycoplasmen (Rep. 10.2). sie isoliert wurden. Die Sacculi zeigen in der Zellmitte einen scharfen Einschnitt – Resultat des lokalen Abbaus der Zellwand durch zelleigene Enzyme unter Einwirkung von Penicillin G (Aufnahme: H. Frank und U. Schwarz, Tübingen; M: Balken = 1 m). Repetitorium 10.2 Die Zellwand als Angriffspunkt beim Kampf gegen Bakterien Lysozym wirkt besser auf grampositive als auf gramnegative Bakterien sowohl in der Wachstums- als auch in der stationären Phase ▬ es zerstört glycosidische Bindungen des Mureins ▬ die Zellwand löst sich auf ▬ die Zellmembran bleibt zunächst intakt (Protoplast bzw. Sphäroplast) ▬ Zellmembran platzt infolge Osmose (Lyse) Penicilline töten bevorzugt grampositive Bakterien in der Wachstumsphase ▬ Penicilline verhindern das Vernetzen der Peptidbrücken des Mureins (Hemmung der Transpeptidase) ▬ sie zerstören dadurch die Zellwand gramnegative Bakterien: Die äußeren Zellwandschichten behindern den Zutritt des Penicillins! Zellwandlose Bakterien, wie z. B. Mycoplasmen oder L-Formen, werden von Penicillin nicht angegriffen aus: Hirsch-Kauffmann u.a., Biologie und molekulare Medizin (ISBN 9783137065074) 2009 Georg Thieme Verlag KG