Gluconeogenese
Werbung

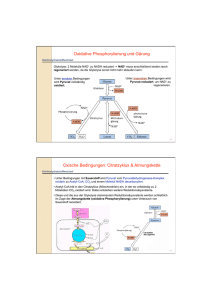
Gluconeogenese Asmaa Mebrad Caroline Mühlmann 06.12.2004 Definition: wichtiger Stoffwechselweg, bei dem Glucose aus Nicht-Kohlenhydrat-Vorstufen synthetisiert wird Ablauf bei längeren Hungerperioden dient der Hochhaltung des Blutglucosespiegels zur Versorgung des Gehirns, der Erythrozyten und der Muskulatur Tagesgesamtbedarf an Glucose beträgt 160g, wovon allein 120g für die Vesorgung des Gehirns benötigt werden Orte der Gluconeogenese 90 % in der Leber 10 % in der Nierenrinde ganz wenig in Gehirn, Skelett- und Herzmuskel Synthesevorstufen Lacat, welches v.a.in den Erythrozyten und bei Sauerstoffmangel in den Muskeln gebildet wird Aminosäuren, die aus Nahrungsproteinen stammen und bei Hunger durch Abbau von Skelettmuskelproteinen entstehen Glycerin, welches beim Fettabbau durch Hydrolyse von Triacylglycerinen in Fettzellen entsteht Vergleich Glykolyse und Gluconeogenese bei der Glykolyse wird Glucose zu Pyruvat abgebaut bei der Gluconeogenese wird Pyruvat in Glucose umgewandelt die Gluconeogenese ist aber keine Umkehr der Glykolyse! Unterschiede zwischen Glykolyse und Gluconeogenese Glucose + ATP Hexokinase Glucose-6-phosphat + ADP Fructose-6-phosphat Phosphofructokinase Fructose-1,6-biphosphat + ADP Phosphoenolpyruvat + ADP Pyruvat-Kinase Pyruvat + ATP die größte Abnahme der freien Enthalpie findet in der Glykolyse in diesen 3 irreversiblen Reaktionen statt Umgehung dieser 3 Schritte bei der Gluconeogenese durch folgende 3 Reaktionen: 1.) Carboxylierung des Pyruvats zu Oxalacetat und anschließender Decarboxylierung und Phosphorylierung zu Phosphoenolpyruvat Enzyme: Pyruvat-Carboxylase PhosphoenolpyruvatCarboxykinase Ablauf in den Mitochondrien 2.) Hydrolyse von Fructose-1,6-BP Zu Fructose-6-phosphat Enzym: Fructose-1,6biphosphatase 3.) Hydrolyse von Glucose-6-Phosphat zu Glucose Enzym: Glucose-6-phosphatase Ablauf im endoplasmatischen Retikumlum Besondere strukturelle, katalytische und allosterische Eigenschaften der PyruvatCarboxylase Bindung des Biotins an die PyruvatCarboxylase über lange, flexible Kette Biotin ist eine prosthetische Gruppe dient als Carrier von aktiviertem CO2 Carboxylierung des Pyruvats in 2 Schritten 1.)Aktivierung der Carboxylgruppe durch Bindung dieser an das N-1-Atom des Biotin-Ringes im CarboxybiotinEnzym-Zwischenprodukt 2.) Übertragung der aktivierten Carboxylgruppe vom Carboxybiotin auf das Pyruvat, wobei Oxalacetat entsteht Aktivität der PyruvatCarboxylase Abhängigkeit der Aktivität von der Gegenwart des Acetyl-CoA ein hoher Acetyl-CoA-Spiegel signalisiert einen höheren Bedarf an Oxalacetat bei ATP-Überschuss wird Oxalacetat in der Gluconeogenese verbraucht bei ATP-Mangel tritt das Oxalacetat durch Kondensation mit Acetyl-CoA in den Citratzyklus ein Oxalacetat Transport von Oxalacetat durch Reduktion zu Malat durch die Mitochondrienmembran Oxidation des Malats zu Oxalacetat im Cytosol Decarboxylierung und Phosphorylierung von Oxalacetat in Phosphoenolpyru -vat Bildung freier Glucose als wichtiger Kontrollpunkt in den meisten Geweben endet die Gluconeogenese beim Glucose-6phosphat (wird v.a.für die Glykogensynthese verwendet), da es nicht wie freie Glucose aus der Zelle diffundieren kann Glucose wird auf 2 Wegen in der Zelle festgehalten, was auf 2 Wegen kontrolliert wird 1.) durch Regulation des für die Umwandlung von Glucose-6-phosphat in Glucose verantwortliche Enzym, die Glucose-6-phosphatase 2.) ist dieses Enzym nur in Geweben zu finden, die für die Homöostase des Blutgluscosespiegels verantwortlich sind, u.z. Leber und in geringem Ausmaß die Niere Erzeugung von Glucose aus Glucose-6phosphat findet im Lumen des endoplasmatischen Reticulums statt T1 transportiert Glucose6-phosphat ins Lumen des ER Hydrolyse des Glucose-6phosphats durch die Glucose-6-phosphatase, für deren Aktivität ein calciumbindendes Protein (SP) unerlässlich ist T2 befördert Pi zurück ins Cytosol T3 befördert Glucose zurück ins Cytosol Nettogleichungen der Gluconeogenese und der Umkehr der Glykolyse Gluconeogenese 2 Pyruvat + 4 ATP + 2 GTP + 2 NADH + 6 H2O zu Glucose + 4 ADP + 2 GDP + 6 Pi + 2 NAD+ + 2 H+ Gleichung für Umkehr der Glykolyse 2 Pyruvat + 2 ATP + 2 NADH + H2O zu Glucose + 2 ADP + 2 Pi + 2 NAD+ • es werden bei der Gluconeogenese 6 Nucleosidtriphosphatmoleküle hydrolysiert, um Glucose aus Pyruvat zu synthetisieren (4 ATP und 2 GTP) • bei der Umwandlung von Glucose in Pyruvat bei der Glykolyse entstehen jedoch nur 2 Moleküle ATP • somit ,,kostet“ die Gluconeogenese für jedes aus Pyruvat gebildetet Molekül Glucose 4 Moleküle mit hohem Phosphorylgruppenübertragungspozenzial extra • diese werden gebraucht, um die energetisch ungünstige Umkehr der Glykolyse in einen energetisch günstigen Prozess, die Gluconeogenese zu verwandeln Gluconeogenese und Glykolyse werden reziprok reguliert innerhalb der Zelle sind beide Prozesse so koordiniert, dass der eine relativ inaktiv ist, während der andere hohe Aktivität zeigt Geschwindigkeit der Gluconeogenese ist abhängig von der Lactatkonzentration und der Konzentration anderer Glucosevorstufen Geschwindigkeit der Glykolyse ist abhängig von der Glucosekonzentration Insulin steigender Spiegel nach Nahrungsaufnahme Stimulation der Expression der Glykolyseenzyme Phosphofructokinase, Pyruvatkinase und des für die Fructose2,6-biphosphat-Herstellung verantwortliche Enzyms Glucagon steigender Spiegel im Hungerzustand hemmt die Expression der Glykolyseenzyme fördert die Bildung der Gluconeogeneseenzyme Phosphoenolpyruvat-Carboxykinase und Fructose-1,6-biphosphatase Substratzyklen Reaktionspaar wie z.B.die Phosphorylierun g von Fructose6-phosphat zu Fructose-1,6biphosphat und dessen Hydrolyse zurück zum Fructose-6phosphat Substratzyklen in den meisten Zellen können nicht beide Reaktionen gleichzeitig aktiv sein in geringem Ausmaß findet jedoch auch während der Gluconeogenese die Phosphorylierung von Fructose-6-phosphat zu Fructose-1,6-biphosphat statt mögliche biologische Bedeutung: Verstärkung der Stoffwechselsignale Wärmeerzeugung durch Hydrolyse von ATP Lactat und Alanin wichtige Synthesevorstufen der Gluconeogenese Erzeugung in aktiver Skelettmuskulatur und in Erythrozyten Lacatbildung verlagert einen Teil der Stoffwechsellast von der Muskulatur auf die Leber • durch die Lactat-Dehydrogenase wird NADH zu NAD+ oxidiert und Pyruvat zu Lactat reduziert • Lacat muss in Pyruvat zurückverwandelt werden, bevor es metabolisiert werden kann • das Pyruvat wird dann durch die Gluconeogenese in der Leber zu Glucose umgewandelt •Damit versorgt die Leber den kontrahierenden Skelettmuskel mit Glucose Cori-Zyklus