6. Hemmung der Gluconeogenese
Werbung

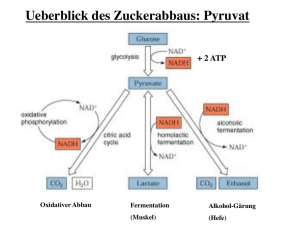
14.3 Stoffwechselwirkungen des Alkohols 6. Hemmung der Gluconeogenese Vorsicht bei Alkoholgenuss frühmorgens auf nüchternem Magen Spätestens vier Stunden nach der letzten Nahrungsaufnahme wird die Blutzucker-Konzentration durch Glykogenolyse der Leber aufrecht gehalten, um alle Zellen, die nicht auf FettsäureVerbrennung umschalten können, vor allem Nervenzellen und Erythrozyten, mit Glucose zu versorgen. Nach weiteren zwei Stunden beginnt allmählich die hepatische Gluconeogenese, diese Aufgabe zu übernehmen. Ohne sie könnten wir nicht das 12-stündige Intervall zwischen Abendessen und Frühstück überstehen. Sie ist somit in der Phase des Hungerns ein lebenswichtiger Prozess. Ist sie gestört, so kommt es zum Versagen von Hirnfunktionen bis hin zum hypoglykämischen Schock und Tod. Gluconeogenese ist eine partielle Umkehrung der Glykolyse. Sie findet statt in der Leber und (nach längerem Hungern) auch in der Niere. Die wichtigsten Substrate sind Lactat (aus Erythrozyten und Muskel), Alanin (aus Muskel) und Glycerin (aus Fettgewebe). Bei ihrer Umwandlung zu Glucose sind mehrere + NAD -abhängige Dehydrogenase-Reaktionen direkt beteiligt: GDH bei Glycerin, GAPDH bei Alanin, LDH und GAPDH bei Lactat; indirekt auch MDH bei Alanin und Lactat. An diesen Reaktionen kann die Gluconeogenese durch das Redox-Chaos, das die Ethanol-Oxidation auslöst, beeinflusst werden. Sir Hans Krebs (siehe Citrat-Zyklus, genannt nach seinem Entdecker Krebs-Zyklus), der die Erforschung der Gluconeogenese anstieß, sah darin einen reduktiven Prozess, der sich – wie er anfangs vermutete – mit Ethanol beschleunigen lässt, weil NADH auf der Stufe der GAPDH verbraucht wird. Seine Experimente ergaben aber genau das Gegenteil. Das nebenstehende Experiment zeigt dies mit der isolierten perfundierten Leber. Nach längerem Nahrungsentzug befand sich die Ratte in Gluconeogenese-Bereitschaft (das Schrittmacher-Enzym PEP-CK wurde durch Glucagon induziert); der vom Gesamtorganismus isolierten Leber aber fehlte das Substrat. Unmittelbar nach Start der Lactat-Infusion wird Glucose ins Perfusat abgegeben. Ethanol-Infusion aber stoppt sofort die Pro duktion. Bis hier simuliert das Experiment eine Situation in vivo (Alkohol, frühmorgens auf nüchternem Magen genossen). Die Hemmung ist reversibel. (Die rasche Reversibilität lässt sich jedoch nur mit nicht-zirkulierendem Perfusat zeigen; denn in vivo verlässt Alkohol nicht schlagartig das Blut.) Was ist die Ursache für die Hemmung der Gluconeogenese? NADH Ethanol NADH Acet– aldehyd Essigsäure Hemmung der Gluconeogenese aus Lactat: Substrat-Limitierung von PC und PEP-CK als Folge einer Verschiebung der LDH- und MDH-Gleichgewichte in Richtung Lactat und Malat PC = Pyruvat-Carboxylase PEPCK = Phosphoenolpyruvat-Carboxykinase Die Negativierung des Redoxpotentials der cytosolischen und mitochondrialen NADH/NAD+-Systeme wirkt sich empfindlich auf die LDH- und MDH-Gleichgewichte aus. Die oxidierten Partner, Pyruvat und Oxalacetat, sinken auf so niedrige Konzentrationen ab, dass den folgenden Reaktionen, katalysiert durch Pyruvat-Carboxylase (PC) und PhosphoenolpyruvatCarboxykinase (PEP-CK), das Substrat fehlt. Die Folge ist, dass der Gluconeogenese-Fluss bereits bei der PC-Reaktion verlangsamt ist. Das wenige Oxalacetat, das doch noch entsteht, reicht nicht aus, um die PEP-CK zu füttern. Zuerst das Brot, dann der Wein! Alkohol nie ohne Kohlenhydrate! „... auf dass der Wein erfreue des Menschen Herz.... und das Brot des Menschen Herz stärke, ... Psalm 104, Vers15 14 – 31