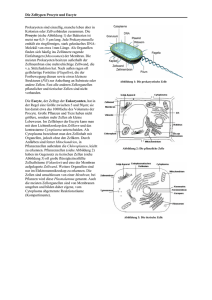
BRT Skript
Werbung

BRT Skript 1.Einleitung Vergleicht man die heutige medizinische Versorgung mit den Zuständen noch vor 100150 Jahren, so ist alleine durch die technische Entwicklung im Laufe dieser Zeit, ein riesiger Fortschritt erreicht worden. Krankheiten, die damals noch in vielen Fällen das Todesurteil für den Patienten bedeuteten, können heute oft komplikationslos behandelt werden. Alleine die Entwicklung von Impfstoffen rettete Tausende von Menschenleben und es war durch flächendeckende Impfmaßnahmen sogar möglich, manche Krankheiten weitgehend auszurotten. Die Mediziner in unserem hochtechnisierten Zeitalter können auf Diagnoseverfahren zurückgreifen, die vor 100 Jahren nicht vorstellbar gewesen wären. Auch die rasante Entwicklung in der Gentechnik öffnet neue Möglichkeiten in der Behandlung von bislang unheilbaren Krankheiten. Den Menschen in der heutigen Zeit kommen diese Fortschritte zugute und es müsste davon ausgegangen werden, dass wir eigentlich im Vergleich zu unseren Ahnen ein recht sorgenfreies Leben führen könnten. Jedoch haben sich auch die Ansprüche an den Menschen von heute, bei im Durchschnitt gesteigerter Lebensqualität, drastisch verändert. Der Mensch in einer hochzivilisierten Gesellschaft ist gezwungen zu funktionieren und verlangt dabei von seinem Körper oft Höchstleistungen. Es ist deshalb nicht verwunderlich, dass in den westlichen Industrieländern etwa ein Drittel der Menschen, die mit gesundheitlichen Problemen einen Arzt aufsuchen, keinerlei Anzeichen einer organischen Erkrankung aufweisen (v. UEXKÜLL und WESIAK 1988). Dies liegt oft darin begründet, das die konventionelle Medizin auch heute noch von einem linearen monokausalem Zusammenhang zwischen den physiologischen Zuständen eines Organismus und einem Krankheitsbild ausgeht. Es ist aber hinlänglich bekannt, dass Organismen, auch auf niedrigerem Organisationsniveau als der Mensch, funktionell nicht in linearen Zusammenhängen zu beschreiben sind. Es ist eine irrige Annahme ein komplexes biologisches Geschehen mit einem entsprechend komplexem multifaktoriellen Krankheitsgeschehen durch z.B. gentechnologische Analysen kausal erklären und behandeln zu wollen. In der konventionellen Medizin wird dem "kranken" Organismus eine passive Rolle zugedacht und dabei versucht, krankhafte Veränderungen oder die ihnen zugeschriebene Ursachen direkt zu beseitigen. Durch eine operative oder chemische Ausschaltung, durch pharmakologische Lenkung und Gegensteuerung zur Wiederherstellung einer kybernetischen definierten Norm oder durch künstlichen Ersatz von fehlenden oder mangelnden Körperwirkstoffen wird versucht, die Krankheitssymptome zu beseitigen. In vielen Fällen sind diese herkömmlich medizinischen Verfahren sinnvoll und notwendig. Kann jedoch die Ursache einer Krankheit nicht definitiv festgestellt werden, können diese Verfahren eine Sackgasse darstellen, aus der Jahrzehnte andauernde Patientenkarrieren resultieren. Im Gegensatz dazu entspricht die biologische Medizin nicht dem kausalanalytischen Denkstil der konventionellen Medizin. In der biologischen Medizin wird von dem Organismus als offenes nicht lineares System ausgegangen, das bestrebt ist, sich und sein funktionelles Gleichgewicht zu erhalten um energetisch am effektivsten zu arbeiten. Therapieweisen der biologischen Medizin wirken 1 dementsprechend indirekt als Reaktion bzw. Regulationsleistung auf eine entsprechende Reizbelastung hin und fördern damit die Anregung endogener Eigenleistungen, die unter den jeweils individuellen Verhältnissen eine Gesundheit des Organismus ermöglichen können. Verfahren die im Rahmen der biologischen Medizin arbeiten, d.h. die die nichtlineare Arbeitsweise eines Organismus berücksichtigen sind z.B. die Homöopathie, sämtliche Akupunkturverfahren und die Bioresonanztherapie. Während es sich bei der Homöopathie um eine klassische Regulationstherapie handelt, zielen Akupunktur und Bioresonanzverfahren in ihrer Wirkungsweise direkt auf die Grundregulation eines Organismus. Um jedoch die Wirkungsweise und die Interpretation der Bioresonanzverfahren verstehen zu können, reicht ein rein schulmedizinisches Verständnis nicht aus. In erster Linie ist es notwendig die funktionellen Zusammenhänge innerhalb der Bestandteile eines Organismus zu verstehen, die für die Regulation und die Aufrechterhaltung der Homöostase notwendig sind. Zusätzlich sind Exkurse in biophysikalische, biokybernetische und physikalische Gesetzmäßigkeiten erforderlich um Bioresonanzverfahren gezielt einsetzen zu können. Die Schulung ist deshalb in zwei Schwerpunkte aufgeteilt. Im ersten Abschnitt werden die biologischen Strukturen eines Organismus behandelt, die in erster Linie an der Aufnahme und der Verarbeitung der Bioresonanzfrequenzen beteiligt sind. Dabei werden alle biokybernetischen Faktoren sowie chronobiologische Aspekte, die im Körper eine entscheidende Rolle spielen behandelt. Im zweiten Abschnitt der Schulung werden die physikalischen Grundlagen von elektromagnetischen Feldern erklärt, sowie an verschiedenen Systemen die Arbeitsweise von Bioresonanzverfahren erläutert. 2 2.Biologische Grundlagen der Bioresonanztherapie 2.1. Zelle und Zellmembran Der Grundbaustein aller lebender Organismen ist die einzelne Zelle. Sie ist die kleinste lebende Einheit und tritt als eigenständige Organisationsform als Einzeller (Protozoen) auf. Bei mehrzelligen Lebewesen bilden die Zellen mehr oder weniger ausgedehnte Verbände, die funktionelle Einheiten einer übergeordneten Struktur darstellen. Alle tierischen Zellen besitzen ein einheitliches Grundmuster, trotz großer Unterschiede in ihrer Aufgabenverteilung. Entsprechend ihrer Funktion sind einzelne Zellen in ihrem Aufbau spezialisiert. Für den Transport von Sauerstoff sind die roten Blutkörperchen zuständig, während über Nervenzellen die schnelle Informationsübertragung (Erregungsleitung) im Organismus gewährleistet werden kann. Der menschliche Körper besteht etwa aus 75.000 Milliarden Zellen. Den größten Anteil bilden mit 25.000 Milliarden die Erythrozyten, gefolgt von den Nervenzellen mit 25.000 Millionen. Die Größe von einzelnen Zellen schwankt zwischen 5 µm (einige Bindegewebszellen) und 150 µm (weibliche Eizelle), manche Nervenzellen erreichen mit ihren Fortsätzen die beträchtliche Länge von 1 m. Die Zellen eines Organismus, egal welcher Organisationsstufe, haben eine Reihe von Grundeigenschaften, auch wenn sie im Hinblick auf die Übernahme bestimmter Aufgaben differenzierter sind. 2.1.1. Zelleigenschaften: Stoffwechsel und Energiegewinnung Jede Zelle ist darauf angewiesen aufgenommene Stoffe zu zelleigenen, dem Aufbau der Zelle dienende Verbindungen umzuwandeln und die anfallenden Endprodukte wieder abzugeben. Zur Aufrechterhaltung ihrer Funktion muss jede Zelle Nährstoffe, in Form von Fette, Eiweiße und Kohlenhydrate aufnehmen und diese zur Energiegewinnung chemisch umwandeln. Die anfallenden Endprodukte werden an die die Zelle umgebende Flüssigkeit abgegeben. Vermehrung und Lebenserwartung Fast alle Zellen vermehren sich lebenslang durch Zellteilung (Mitose). Dadurch werden zugrunde gegangene Zellen ersetzt und die Regeneration von Geweben und Organen gewährleistet. Das menschliche Knochenmark bildet stündlich etwa 160 Millionen rote Blutkörperchen und die männlichen Keimdrüsen (Hoden) etwa 85 Millionen Spermien pro Tag. Im Gegensatz zu manchen Nerven und Muskelzellen, beträgt die durchschnittliche Lebenserwartung der Schleimhautzellen des Dünndarm nur 30 - max. 100 h und weisen dementsprechend eine hohe Zellteilungsrate auf. Reizaufnahme und Reizbeantwortung Die meisten Zellen stehen durch spezifische Zelloberflächenstrukturen mit ihrer Umgebung in Verbindung und sind damit in der Lage, Reize aufzunehmen (über spez. Rezeptoren), auszuwerten und entsprechend zu beantworten oder weiterzuleiten. Zusätzlich zu diesen basalen Eigenschaften besitzen bestimmte Zellen spezifische Eigenschaften, die mit ihrer Funktion zusammenhängen. So haben z.B. Abwehrzellen im Bindegewebe sowie männliche Spermien die Fähigkeit sich zu bewegen. Auch werden durch bestimmte Oberflächendifferenzierungen, wie z.B. Flimmerhaare auf 3 Schleimhautepithelzellen des Atemtrakts oder Bürstensaum auf Schleimhautepithelzellen des Dünndarmes die funktionellen Eigenschaften der Zellen gewährleistet. 2.1.2. Grundbauplan der Zelle Abb.1: Vereinfachte Darstellung des Grundbauplans einer Zelle 2.1.2.1. Zellmembran Zellen sind umgeben von einer Zellmembran, die auch als Plasmamembran (Plasmalemm) bezeichnet wird. Grundsätzlich gilt, dass alle Lebensprozesse zu einem wesentlichen Teil auf den strukturellen und funktionellen Eigenschaften von Membranen beruhen. Alle biologischen Membranen wirken als selektive Barrieren, die einen gerichteten Stoff-, Energie- und Informationsaustausch zwischen Zellen und Umwelt einerseits und zwischen bestimmten Kompartimenten innerhalb der Zelle andererseits ermöglichen. Die Zellmembran besteht aus einer Lipiddoppelschicht, wobei 2 Lagen von Lipidmolekülen (Phospholipide, Glycolipide und Sterine wie Cholesterin) derart angeordnet sind, dass ihre fettlöslichen Anteile (Fettsäuren) einander zugewandt sind, während die wasserlöslichen Anteile an die Innen- bzw. die Außenmembran grenzen. Die Lipiddoppelschicht wird mosaikartig von Eiweißmolekülen (Proteinen) durchsetzt, die als Poren für Wasser und Salze dienen oder als Rezeptorproteine regulatorische Aufgaben übernehmen. Die an die Außenseite der Zelle grenzenden Membranproteine sowie zum Teil auch die wasserlöslichen Anteile der Phospholipide werden von einer dünnen Schicht komplexer Zuckermoleküle (Kohlenhydrate), der Glycokalyx 4 überzogen. Die chemische Zusammensetzung der Glykokalyx ist spezifisch für jede Zelle und genetisch festgelegt. Durchschnittlich besteht die Plasmamemran etwa 40% aus Fetten, 50% aus Proteinen und 8% aus Kohlehydraten. Abb.2a: Phospholipidstruktur (Ch = Cholin, Fe = Fettsäuren, Gl = Glycerol, Ko = hydrophile Kopfgruppe, P = Phosphat, Sch = lipophile Schwänze) b: Membranproteine. Abb.3: Schematischer Aufbau der Zellmembran (1 = Oligosaccharidkette, 2 = Glycoprotein, 3 = Glycolipid, 4 = globuläres Protein, 5 = Kanal, 6 = lineares Protein, 7 = Cholesterin, 8 = Phospholipid) 2.1.2.2. Transportmechanismen an der Zellmembran Die Zellmembran wirkt als selektive Barriere zur Aufrechterhaltung des intrazellulären Milieus und ermöglicht den gerichteten Austausch von Ionen, niedermolekularen Substanzen und Makromolekülen zwischen Zelle und Extrazellularraum. Es bestehen große Unterschiede bezüglich der Ionenkonzentration zwischen dem äußeren und dem inneren Milieu einer Zelle. So haben fast alle tierischen Zellen eine höhere intrazelluläre [K+]- und eine niedrige [Na+]- Konzentration im Vergleich zum Außenmedium. Dadurch besteht zwischen innen und außen ein Ladungsunterschied und ein Konzentrationsgefälle für bestimmte Ionen. Diese Spannungsdifferenz wird als 5 Membranpotential bezeichnet und liegt je nach Zelltyp zwischen - 20 bis - 200 mV. Die Innenseite der Zellmembran ist gegenüber der Außenseite immer negativ geladen. Für lipophile und kleine ungeladene polare Moleküle wie Wasser, CO2 und Harnstoff ist die Lipiddoppelschicht durchlässig (permeabel). Diese Stoffe durchqueren die Membran durch einfache Diffusion in Abhängigkeit vom Konzentrationsgefälle, während sie für große polare Moleküle wie Glucose und für Ionen undurchlässig ist. Ionen können die Zellmembran nur über spezielle Transportmechanismen passieren. Dabei unterscheidet man zwischen passivem, entlang dem Konzentrationsgefälle und aktivem Transport. Aktive Transportmechanismen arbeiten immer entgegen einem Ionengradienten und benötigen deshalb Energie. Der passive Transport erfolgt über Ionenkanäle, die aus integralen Membranproteinen bestehen und verschließbar sind, während aktive Transportmechanismen über Ionenpumpen betrieben werden. Diese Ionenpumpen bestehen ebenfalls aus integralen Membranproteinen, die durch reversible Änderung ihrer dreidimensionalen Struktur (Konformationsänderung) spezifische Ionen auf der einen Seite der Membran binden und auf der anderen Seite wieder entlassen. Die Konfirmationsänderung des Membranproteins erfordert Energie und beruht auf einer reversiblen Phosphorylierung durch Adenosinphosphat. Niedermolare ungeladene Substanzen wie Glucose sind an den Ionentransport gekoppelt, d.h. er verläuft über die gleichen Trägerproteine. Abb.4: Aktiver und passiver Transport durch die Zellmembran Für Makromoleküle die die Zellmembran passieren müssen, gibt es komplizierte Transportmechanismen, bei denen Membranfusionsprozesse die entscheidende Rolle spielen. 6 Bei der Endozytose (Transport ins innere der Zelle) bildet sich an der Außenseite der Membran eine Einbuchtung. Dann binden die äußeren Teile der Lipiddoppelschicht aneinander und schnüren ein von der Membran umgebendes Bläschen (Vesikel) ab, das ins innere der Zelle wandert. Bei diesem Vorgang bleibt die Polarität der Lipiddoppelschicht erhalten: Die gegen das Zellinnere gerichtete Schicht der Membran, umschließt das Vesikel von außen, während die in den Extrazellulärraum gewandte Seite den Vesikel von innen auskleidet. Man bezeichnet die Endocytose von großen Partikeln als Phagozytose und die von kleinen Partikeln als Pinocytose. Bei der rezeptorvermittelten Endocytose werden spezifische Substanzen durch Membranrezeptoren an die Plasmamembran gebunden und ins Zellinnere eingeschleust. Bei der Exozytose (Transport in den Extrazellularraum) läuft der Vorgang in umgekehrter Richtung ab: Ein Vesikel aus der Zelle fusioniert mit der Innenseite der Membran und entlässt seinen Inhalt in den Extrazellulärraum. Abb.5: Membranfluß: 1 = Endozytose und 2 = Exozytose (Ce Centriol, Cs Cytosol, En endocytotisches Vesikel, Es Endosom, Es exocytotisches Vesikel, rER raues endoplasmatisches Reticulum, Go Golgi-Apparat, Ly Lysosom, Mi Mitochondrium, No Nucleolus, Nu Nucleus, Pe Peroxisom, Pm Plasmamembran, Ps Polysomen, Sv Sekretvesikel) Zusammenfassend ist festzustellen, das die unterschiedlichen spezifischen Transportmechanismen bei der Aufrechterhaltung der Homöostase zwischen Zelle und Umgebung die entscheidende Rolle für die Funktion spielen. Passive Transportmechanismen sind die freie Diffusion z.B. für O2, CO2 und H2O, die erleichterte Diffusion z.B. für Glukose und Aminosäuren in den Zellen der Darmschleimhaut und Osmose und Filtration z.B. für Glukose und Aminosäuren in den Gewebekapillaren. Über aktive, d.h. energieabhängige Transportmechanismen sind zum einen die Ionenpumpen sowie die Endo- und Exotzytose für Proteine. Diese unterschiedlichen, teilweise sehr spezifischen Transportmechanismen sind essentiell für die Funktionsweise der Zelle. Durch Verletzungen der Membran können diese Mechanismen nicht mehr ablaufen, d.h. der Stoffwechsel der Zelle ist erheblich gestört und es kann zum Absterben von einzelnen Zellen und im weiteren Verlauf zu erheblichen Funktionsstörungen des jeweiligen Gewebes kommen. 7 2.1.2.3. Zytoplasma und Zytoskelett Tierische Zellen sind durch intrazelluläre Membranen, die die gleiche Struktur wie die Plasmamembran haben, in eine Vielzahl von geschlossenen Kompartimenten unterteilt. Diese Kompartimente, in denen verschiedenen Enzymaktivitäten gleichzeitig ablaufen können, bilden die Zellorganellen. Dieses innere Membransystem führt einerseits zu einer Vergrößerung der funktionellen Membranoberfläche, anderseits aber auch zu einer strukturellen und funktionellen Differenzierung der Membranen. Das gesamte intrazelluläre Membransystem, das die Zellorganellen bildet ist umgeben von der gel-artigen Masse des Zytoplasmas, das etwa 50% des Zellvolumens aus macht. Das Zytoplasma enthält ein vielmaschiges Netz aus Faserproteinen, die das Zytoskelett bilden. Dieses Grundgerüst von Filamenten ist in der Zellmembran verankert und garantiert Stütz- und Bewegungsfunktion. Es können drei verschiedene Filamente unterschieden werden: Die Actinfilamente (Durchmesser 7 -9 nm), die Mikrotubuli (Durchmesser 30 nm), Röhrenelemente die der Beweglichkeit der Zelle dienen und den intermediären, stabileren Filamenten (Durchmesser 8 - 14 nm) die hauptsächlich eine Stützfunktion haben. Die Actinfilamente bilden in Verbindung mit dem Myosin die kontraktilen Elemente der Zelle. Amöbide Bewegungen, Plasmaströmungen im Zellinneren und Muskelkontraktionen sind auf ihr Wirken zurückzuführen. Die Mikrotubuli, die vom Centriol der Zelle strahlenförmig ins Zytoplasma ziehen, gewährleisten den Transport der Bläschen und der zytoplasmischen Organellen. Auch bilden sie die Spindelfasern die die Chromosomen bei Zellteilungsvorgängen (Mitose und Meiose) auftrennen. Sie bewirken ebenfalls zusammen mit dem Verbindungsprotein Dynein die Bewegungen der Wimpern und Geißeln von Zellen. 2.1.2.4. Zellorganellen Mit Ausnahme des Zellkerns oder Nucleus können die zytoplasmatischen Organellen in drei Kategorien eingeteilt werden: 1) Energieorganellen wie die Mitochondrien, die die meisten Moleküle des Adenosintriphosphates (ATP) erzeugen. ATP ist der universelle Energielieferant der Zelle und ist beteiligt an allen biochemischen Reaktionen. Die Herstellung von ATP aus den drei Grundnahrungsstoffen, Proteine, Fette und Kohlenhydrate, findet nahezu ausschließlich in den Mitochondrien statt. Je nach Größe und Energiebedarf der Zelle kann die Zahl der Mitochondrien sehr unterschiedlich sein. Mit Ausnahme der roten Blutkörperchen hat jede Zelle mindestens ein, aber es gibt auch Zellen in denen bis zu 100 000 Mitochondrien vorkommen. Mitochondrien sind längliche Zylinder von 2 - 8 µm Länge und einem Durchmesser von 0,5 - 1 µm. Jedes Mitochondrium ist von einer Doppelmembran umgeben, wobei die innere Membran sich in Form von Röhren, Falten oder Säckchen in den Innenraum des Organells einstülpt. Dadurch kommt es zu einer Unterteilung des Mitochondriums in vier Kompartimente: der äußeren Membran, des Intermembranraumes, der inneren Membran und der Matrix oder auch Stroma. Die äußere Membran besteht aus einer Lipiddoppelschicht, die Enzyme des Lipidstoffwechsels und ein Transportprotein enthält. Dieses Protein bildet hydrophile Kanäle, durch die kleinere Moleküle und Proteine in den Innenmembranraum eindringen können. Die innere Membran ist für die meisten dieser Moleküle undurchlässig. Sie enthält ein spezielles Phospholipid (Cardiolipin) und ist zur Oberflächenvergrößerung zu sogenannten Christae aufgefaltet. In ihr liegen die 8 Enzyme der Atmungskette und der ATP-Synthease-Komplex, der durch den elektrochemischen Gradienten angetrieben wird und in der Matrix bzw. im Stroma ATP produziert. Die Matrix enthält Hunderte von Enzymen, die zur Oxidation von Pyruvat und Fettsäuren für den Zitronensäurezyklus und für die Biosynthese der Aminosäuren benötigt werden. Zudem enthält sie mehrere Kopien der ringförmigen mitochondrialen DNA, mitochondriale Ribosomen, transfer-RNA (t-RNA) sowie alle Enzyme die für die Replikation der mitochondrialen Gene notwendig sind. Abb.6: Dreidimensionale Rekonstruktion eines Mitochondriums (äM äußere Membran, iM innere Membran, Cr Crista, Ma Matrix, Pp Protonenpumpen). 2) Syntheseorganellen, wie das endoplasmatische Reticulum (ER) und der Golgi-Apparat. Das endoplasmatische Reticulum durchzieht das Zytoplasma in Form von röhren - und bläschenförmigen Strukturen, die von Einheitsmembranen umgeben sind und an die äußere Membran des Zellkernes grenzt. Es hat zwei Funktionsbereiche: zum einen das raue ER (rER) oder auch Ergastoplasma, das innerhalb der Zelle ein weitläufiges System bildet und in dem die Proteinsynthese stattfindet. Zum anderen das glatte ER (gER), das vor allem in den auf den Abbau oder die Synthese von Lipiden spezialisierten Zellen vorkommt. Der proteinsynthetisierende Teil des ER oder raue ER ist mit Ribosomen besetzt, die an der Außenseite (zytoplasmatischen Seite) sitzen. Die Ribosomen des rER, die auch frei in der Zelle vorkommen können, stehen im Dienste der Eiweißsynthese. Am rER gebunden sind sie für die Produktion von Exportproteinen (Drüsensekrete), den Proteinen und Glykoproteinen der Membranen des ER selbst und den Proteinen der extrazellulären Matrix verantwortlich, während sie als freie Ribosomen zelleigene Proteine (Enzyme und Strukturproteine) produzieren. Man findet raues ER besonders ausgeprägt in Bauchspeicheldrüsenzellen. Das glatte ER (gER) ist vor allem auf den Fettstoffwechsel spezialisiert und in den Zellen stark entwickelt, die wie Leberzellen große Mengen von Lipoproteine exportieren. Auch hat es Entgiftungsfunktion in Bezug auf Medikamente, Drogen und toxischen Stoffwechselprodukten. Im glatten ER findet auch die Synthese von Steroidhormonen statt und ist deshalb besonders in hormonproduzierenden Zellen anzutreffen. Der Golgi-Apparat ist ein weiteres Hohlraumsystem, das Lipide und Proteine aus dem endoplasmatischen Reticulum enthält. Er setzt sich aus mehreren Golgi-Feldern (Zisternen oder auch Dictyosomen) zusammen, die an der Aufnahme und an der Ausschleusung von Stoffen in Form von membranbegrenzten Sekretvesikeln beteiligt ist. Die Golgi-Zisternen besitzen eine Aufnahme- und eine Abgabeseite. Vorstufen von Eiweissekreten wandern aus dem rauen ER zur Aufnahmeseite der GolgiZisternen, wo sie in Transportvesikel verpackt und über die Ausgabeseite aus der Zelle geschleust werden. Die an das ER angrenzende Seite wird deshalb als Bildungs - oder cis-Kompartiment bezeichnet, die Gegenseite als Sekretions- oder trans9 Kompartiment. Bei der Ausschleusung der im Golgi-Apparat gebildeten Vesikel verschmilzt die Vesikelmembran mit der Zellmembran. Die Erneuerung der Zellmembran ist daher eine wichtige Aufgabe des Golgi-Apparates. 3) Verdauungsorganellen: Die kugelförmigen Lysosomen sind die Verdauungsorgane der Zelle, die im GolgiApparat gebildet werden. Sie enthalten Verdauungsenzyme besonders saure Hydrolasen und Phosphatasen, mit deren Hilfe sie aufgenommene Fremdkörper und zelleigene Abfallprodukte abbauen und dem zellulären Stoffwechsel in Form von Ausgangsstoffen wieder zur Verfügung stellen können. Die Lysosomenmembran schützt die intakte Zelle von einer unkontrollierten Wirkung der lysosomalen Enzyme. In geschädigten Zellen können die freigesetzten Enzyme zur Autolyse (Selbstauflösung) der Zelle beitragen. Peroxisomen sind lysosomähnliche Vesikel, die jedoch nicht im Golgi-Apparat gebildet werden. Sie enthalten Oxidasen, die verschiedene Substrate wie Aminosäuren, Purine, Milchsäure, Fettsäuren usw. mit molekularem Sauerstoff oxidieren. Bei diesem Vorgang entsteht Wasserstoffperoxid, das vom Enzym Katalase zu Wasser reduziert wird. 2.1.2.5. Zellkern Jede Säugetierzelle mit Ausnahme der roten Blutkörperchen besitzt einen Zellkern oder Nucleus, der von einer doppelten Membran, der Kernhülle umgeben ist. Die äußere Membran grenzt an das endoplasmatische Reticulum (ER), während die innere Membran zum Kern hin von der fibrillären Lamina bedeckt ist. Innerhalb des Zellkerns liegt das Kernkörperchen oder der Nucleolus, dessen Aufgabe die Produktion ribosomaler RNA ist. Bei inaktiven Zellen ist der Nucleolus unauffällig, während er bei stoffwechselaktiven Zellen mit einer erhöhten Proteinsynthese deutlich hervortritt. Die fibrilläre Lamina stabilisiert die Kernhülle und verbindet den Nucleus mit dem Chromatin, dem inneren Kernbestandteil. Die beiden Membranen der Kernhülle bilden zusammen Kernporen mit einem Durchmesser von 30 - 100 nm, über die ein intensiver Stoffaustausch zwischen Kern und Zytoplasma erfolgen kann. Das Chromatin besteht aus DNA und den Histonen (basische Chromosomenproteine). Die DNA ist auf die Chromosomen verteilt, fadenförmige Strukturen, die nur während der Mitose (Kernteilung) sichtbar sind. In der Interphase (zwischen den Teilungen) ist das Chromatin diffus im Kern verteilt. Da die Chromosomenzahl beim Menschen 46 beträgt und die Chromosomen aus einer kontinuierlichen DNA-Kette bestehen, sind die DNA-Moleküle in den Chromosomen mehrere Zentimeter lang. Die Verpackung dieser Riesenmolelküle in den Zellkern, der nur einen Durchmesser von etwa 10 µm besitzt, und ihre gleichmäßige Verteilung in der Mitose wird durch die dynamische Struktur des Chromatins ermöglicht, die sowohl eine dichte Verpackung (in der Mitose) als auch eine aufgelockerte Anordnung (in der Interphase) - und damit sowohl Replikation als auch Transkription ermöglicht. Kern- und Zellzyklus Zellen entstehen in einem zyklischen Prozeß von Wachstum und Teilung. Der Zellzyklus gliedert sich in zwei Hauptphasen, die Interphase und die Teilungsphase. In der Interphase nimmt die Zelle an Masse zu und repliziert ihre DNA , während die Teilungsphase aus der Kernteilung, der Mitose, und der Teilung des Zytoplasmas besteht. 10 Die Struktur der DNS-Einzelstränge ermöglicht bei der Zellteilung eine identische Verdopplung. Hierbei trennen sich die Basenpaare der Doppelhelix wie ein Reißverschluss in der Mitte und an jedem Einzelstrang wird ein exakt komplementärer Strang synthetisiert. Mitotische Zellteilungen sind die Vorraussetzung für die physiologische Zellerneuerung und führen zur Regeneration von Gewebe nach Verletzungen. Mit Ausnahme weniger Zellen geht die Teilungsfähigkeit während des gesamten Lebenszyklus nicht verloren. Allerdings ist die Teilungsfähigkeit von Zellen unterschiedlich ausgeprägt, d.h. je hochdifferenzierter das Gewebe desto seltener ist die Mitoserate. Die Mitose lässt sich in vier verschiedene Phasen unterteilen: Prophase (pro = vor) In der Prophase rundet sich die Zelle ab und im Kern erscheinen die Chromosomen als fädige Strukturen. Gleichzeitig verschwindet die Kernmembran und die beiden Zentralkörperchen (Zentriolen) rücken auseinander. In dem sie zu den Zellpolen wandern bilden sie die Zentralspindeln aus. Metaphase (meta = mitte) In der Metaphase verkürzen und verdicken sich die Chromosomen, die beiden chomatiden werden sichtbar und sind in Größe und Form deutlich zu unterscheiden. Dann ordnen sich die Chromosomen zwischen den beiden Polen in der sogenannten Äquatorialebene an und zwar so, dass ihre jeweiligen Einschnürungen (Zentromere) zur Mittelachse gerichtet sind. Anaphase (ana = aufwärts) Zu Beginn der Anaphase wandern die beiden Chromosomenhälften (Chromatide) auseinander und es entstehen zwei sternförmige Figuren (Diaster). Durch die Wanderung der Tochterchromatide zu jeweils einem der beiden Pole wird das gesamte genetische Material identisch auf beide Tochterzellen verteilt. Telophase (telos = Ende, Ziel) In der Telophase versammeln sich die Chromatiden, die jetzt die Chromosomen der Tochterzellen bilden in der Nähe des Zentriols und entspiralisieren sich. Mit der Bildung zwei neuer Kernmembranen sind zwei neue Inerphasenkerne entstanden. Mit der Durchschnürung des Zellleibes entstehen zwei gleichgroße Tochterzellen. Die Mitose dauert durchschnittlich 60 Minuten, wobei die Anaphase mit etwa 3 Minuten die kürzeste Phase darstellt. 11 Abb.7: Schema der Mitose (a Prophase, b frühe Metaphase, c späte Metaphase, d und e Anaphase, f Telophase) Das Wachstum und die Differenzierung von Geweben hängt in erster Linie von der Regulation der Zellteilung ab. Die Länge der einzelnen Phasen eines Zellzyklus variiert bei den verschiedenen Zelltypen eines Organismus. Viele Nervenzellen, Muskelzellen und Erythrocyten (rote Blutzellen) teilen sich nach der Reifung in der Regel nicht mehr. Die Epithelzellen des Darmes, der Lunge, der Haut und die blutbildenden Zellen des Knochenmarks teilen sich dagegen ständig und werden laufend ersetzt. Viele Zelltypen teilen sich kaum, behalten aber ihre Teilungsfähigkeit. So können Leberzellen nach operativer Entfernung von einem Teil des Organs, diesen Ausfall vollständig regenerieren. Nach erfolgreicher Regeneration stellen sie dann ihre Teilungsaktivität wieder ein. Auch Untersuchungen mit Zellkulturen belegen, dass bei gesunden Zellen Rückkopplungsmechanismen bestehen, die das Zellwachstum beschränken. Es kommt zu einer Kontaktinhibition der Zellteilung, d.h. die Position der Zelle in der Kolonie bzw. im Gewebe bestimmt ihren Zellzyklus. Diese Rückkopplungsmechanismen die das Wachstum gesunder Zellen beschränken, sind bei Krebszellen unwirksam. Krebszellen wachsen unbegrenzt und können daher fremde Gewebe infiltrieren und Metastasen bilden. Die Zellvermehrung (Zellpoliferation) wird durch verschiedene Wachstumsfaktoren gefördert. Dazu gehören einerseits lokal wirkende chemische Mediatoren wie Prostaglandine und Histamin, anderseits Hormone die im Blut zirkulieren wie z.B. der epidermale Wachstumsfaktor (EGF) und der Nervenwachstumsfaktor (NGF) die ein beschleunigtes Wachstum von Epidermiszellen bzw. Nervenfasern bewirken. Polypeptid- und Steroidhormone kontrollieren das Wachstum derjenigen Zellen, die dafür entsprechende Rezeptoren dafür aufweisen. 12 Abb.8: Zellzyklus (Interphasen: G1, S und G2, M Mitose) Ein Zellzyklus umfasst eine Wachstums- und eine Teilungsphase. Allgemein ist er in vier Phasen (G1, S, G2, M) unterteilt. Nach der Mitose (M) treten die Tochterzellen in die Interphase (G1, S, G2) eines neuen Zellzyklus ein. In der G1-Phase spricht die Zelle besonders auf die verschiedenen Wachstumsfaktoren (EGF, IGF1, IGF2 und Insulin) an. 2.2. Gewebe als Verbände gleichartig differenzierter Zellen Geweben sind Verbände gleichartig differenzierter Zellen und ihrer Abkömmlinge, den Interzellularsubstanzen, die funktionell eine Einheit bilden. Grob kann man vier verschiedene Gewebetypen unterscheiden: Epithelgewebe, Binde- und Stützgewebe, Muskelgewebe und Nervengewebe. 2.2.1. Epithelgewebe oder Grenzflächengewebe Morphologisch sitzen alle Epithelgewebe einer Balslmembran oder Glashaut auf, die dem Epithel mechanischen Halt gibt. Funktionell lassen sich Epithelgewebe in die oberflächenbildende Epithelien, Drüsenepithelien und Sinnesepithelien aufteilen. Alle oberflächenbildende Epithelien umschließen die äußere und innere Oberfläche des Körpers und haben in erster Linie Schutzfunktion. Auch steht der Körper über die oberflächenbildende Epithelien mit der Umwelt in Verbindung und regelt darüber die Stoffausscheideung (Sekretion) und Stoffaufnahme (Resorption). Nach ihrer Gestalt unterscheidet man das Plattenepithel, das kubische- oder isoprismatische Epithel und das Zylinder- oder hochprismatische Epithel. Einschichtiges Plattenepithel wird von abgeplatteten Zellen gebildet, bei denen der kernhaltige Abschnitt etwas verdickt ist. Dieses Gewebe findet man in erster Linie dort, wo eine besondere Durchlässigkeit erforderlich ist. Es bildet z.B. die oberflächliche Schicht (Mesothel) von Bauch- und Brustfell, die innerste Schicht (Endothel) des gesamten Gefäßsystems sowie die Auskleidung der Lungenaveolen. Mehrschichtige Plattenepithelien kommen an den Grenzflächen vor, die einer mechanischen Beanspruchung und Abnutzung ausgesetzt sind: der Haut (Epidermis), Teilen des Verdauungsrohres die noch einer mechanischen Beanspruchung ausgesetzt sind wie der Mundhöhle und der Speiseröhre, der Hornhaut des Auges (Cornea) und der Vagina. Das mehrschichtige Plattenepithel besteht nur in der obersten Schicht aus abgeflachten Zellen. Die basalen, an das Bindegewebe grenzenden Zellen sind länglich und senkrecht zur Oberfläche orientiert. Von diesen basalen Zellen geht die 13 Regeneration des Gewebes aus. Es schieben sich ständig Zellen zur Oberfläche des Epithels. Während dieses Vorganges verändert sich die Zellform: die prismatischen Zellen des Stratum basale werden in den mittleren Schichten polygonal und platten sich nahe der Oberfläche mehr und mehr ab. Die Grenze zwischen Plattenpithel und Bindegewebe, die eine geringe mechanische Belastung ausgesetzt sind, hat das Plattenepithel eine glatte ebene Form (Cornea). Bei stärkerer mechanischen Belastung sind Plattenepithel und Bindegewebe miteinander verzahnt um eine größere Kontaktfläche zu erreichen. Außerdem sind sie über zahlreiche Haftzonen (Desmosomen) miteinander verbunden. Bei kubischen oder isoprismatischen Epithelien sind die Zellen senkrecht zur Oberfläche etwa ebenso hoch wie breit. Die Schilddrüsenfollikel und das Epithel der Nierenkanälchen werden von kubischen Epithelien gebildet. Die Zellen des Zylinderepithels oder hochprismatischen Epithels sind deutlich höher als breit. Das Zylinderepithel kann ein- oder mehrschichtig sein. Das Schleimhautepithel des Dünndarms wird von einem einschichtigen Zylinderepithel mit einem Bürstensaum als Oberflächendifferenzierung gebildet. Das Übergangsepithel besteht aus Basal- , Intermediär- und Deckzellen. Die Deckzellen sind große, teilweise mehrkernige Zellen, die an der Oberfläche des Epithels eine gut abdichtende Schicht bilden. Die Basal- und Intermediärzellen sind für die starke Dehnbarkeit des Epithels verantwortlich. Gewebe die starken Volumenschwankungen ausgesetzt sind, wie die Harnblase, der Harnleiter, die Nierenkelche und das Nierenbecken sind mit Übergangangsepithel ausgekleidet. Das Flimmerepithel hat seinen Namen von Zellen, deren freie Oberfläche mit feinen 5 10 µm langen, 0,2 µm dicken sich bewegenden Härchen (Kinocilien) gleichmäßig besetzt ist (z.B. Bronchialepithel 109 Cilien je 1 cm²). Nach Anordnung der Zellen unterscheidet man ein- und mehrschichtiges Flimmerepithel. Bei mehrschichtigen Flimmerepithelien bilden die Flimmerepithelzellen zusammen mit Becherzellen (schleimbildende Zellen) die Oberfläche des Epithels. Zwischen den Flimmerepithelzellen sind ein oder mehrere Reihen kegelförmiger Basal- und Intermediärzellen eingeschoben, die nicht bis zur Epitheloberfläche reichen und keine Cilien tragen. Die teilungsfähigen Basalzellen liefern Intermediärzellen nach, die sich dann zu Flimmerepithel- oder Becherzellen ausdifferenzieren können und so für den Ersatz abgestoßener Epithelzellen sorgen. Drüsengewebe stellen innerhalb des Epithelgewebes besonders spezialisierte Vertreter dar. Exokrine Drüsenzellen geben ihre Sekrete an der inneren und äußeren Körperoberfläche ab, während endokrine Drüsenzellen ihre Sekrete über die Gewebsflüssigkeit in die Blut- und Lymphbahnen abgeben. Drüsenzellen kommen als Einzelzellen innerhalb von Epithelgewebe ( z.B. als Becherzellen im Dickdarm) oder in Form von epithelialer Organe (Schweißdrüsen, Speicheldrüsen, Tränendrüsen, Bauchspeicheldrüse) vor. Die von den Drüsen gebildete Sekrete werden im Falle der exokrinen Drüsen über Ausführgänge an den Bestimmungsort transportiert, während endokrine Drüsen (alle hormonproduzierende Drüsen wie die Schilddrüse und die Hirnanhangdrüse) ihre Sekrete direkt an das Blut abgeben. Nach Art der Sekretabgabe unterscheidet man zusätzlich merokrine, apokrine und holokrine Drüsenzellen. Bei merokrinen Drüsenzellen erfolgt die Sekretabgabe als Exozytose. Die meisten endokrinen Zellen sind merokrin. In apokrinen Drüsenzellen sammelt sich das Sekret am apikalen Zellpol und wird dann zusammen mit etwas Zytoplasma und einem Teil der Plasmamembran abgeschnürt. Apokrin wird das Milchfett von den Brustdrüsenzellen abgegeben. Talgdrüsen geben ihre Sekrete holokrin ab. Wenn sich die Drüsenzellen mit Sekret gefüllt haben, stellen sie die Regeneration des Zytoplasmas ein. Die Zellen sterben ab und ihre Reste gehen in das Sekret mit ein, d.h. die ganze Zelle wird abgegeben. 14 Sinneszellen innerhalb von Epithelzellverbänden dienen der Reizaufnahme, wobei die auftreffenden Reize (Licht, chemische Stoffe, mechanische Reize) in elektrische Signale umgewandelt werden (Transduktion) und als Erregung an die angeschlossenen Nervenfasern weitergeleitet werden. Nur sekundäre Sinneszellen sind Epithelzellen, die mit Rezeptoren versehen sind (Geschmacksknospen, Rezeptorzellen des Innenohres, Netzhaut) und ihre Erregung über Synapsen auf Nervenzellen übertragen. Abb.9: Verschiedenen Funktionen von Epithelzellen (a Epidermis der Haut, b Dünndarmzotten, c exokrine Drüse, d Netzhaut im Auge) 2.2.2. Binde- und Stützgewebe Alle Formen von Binde- und Stützgewebe sind Abkömmlinge des embryonalen Bindegewebes (Mesenchym). Man unterscheidet zwischen zellreichem Bindegewebe zu dem das retikuläre Bindegewebe und das Fettgewebe gehört. Dem faserreichen 15 Bindegewebe zu dem das lockere und das straffe Bindegewebe gehört. Und dem Knochen- und Knorpelgewebe, dass das Stützgewebe bildet. Binde- und Stützgewebe besteht aus Zellen und zwischenzellige Substanzen (Extrazellularmatrix), die flüssig, halbflüssig oder fest sein können. Diese extrazelluläre Matrix sind am Aufbau von Binde- und Stützgewebsstrukturen in qualitativer und quantitativer unterschiedlicher Weise beteiligt. Je weniger die Stützfunktion ausgeprägt ist desto mehr tritt die Stoffwechselfunktion in den Vordergrund. Bindegewebe ist gut durchblutet, es verbindet die Organe mit den Blutgefäßen und ist dementsprechend an der Versorgung der Organe beteiligt. 2.2.2.1. Bindegewebe Die sehr unterschiedlichen Funktionen des Bindegewebes machen es zu einem der wesentlichen Geweben in Bezug auf die Homöostase innerhalb eines Organismus und ist deshalb auch für die Bedeutung der Bioresonanztherapie von größter Relevanz. Innerhalb seiner Bindefunktion umhüllt es die Organe, Gefäße und Nerven und verbindet alle Komponenten untereinander. In Form von Bändern stabilisiert es Gelenke und in Form von Sehnen dient es der Kraftübertragung von Muskeln auf die Knochen. Die Stoffwechselfunktion werden in erster Linie von fixierten Bindegewebszellen übernommen, während der Stoffaustausch innerhalb der Interzellularsubstanz erfolgt. Die aus dem Blut austretenden Nährstoffe diffundieren über die Interzellularsubstanz zu den Zellen, wobei das Bindegewebe der Nährstoffverteilung dient. Umgekehr gelangen die von Zellen ausgeschiedenen Stoffe über das Bindegewebe zu den abführenden Blut- und Lymphgefäßen. Auch für den Wasserhaushalt des Körpers spielt das Bindegewebe eine wichtige Rolle. Ein Großteil der Extrazellulären Flüssigkeit befindet sich in den Zwischenräumen des lockeren Bindegewebes, wo es auch gespeichert werden kann. Dies wird deutlich bei Herz- oder Nierenerkrankungen wo Aufgrund von Fehlfunktionen Ödeme im Gewebe auftreten können. Bei der Wundheilung bilden sich spezielle Bindegewebszellen sogenanntes Granulationsgewebe, das später in Narbengewebe übergeht. Freie Bindegewebszellen mit einer amöboiden Eigenbewegung, wie die weißen Blutkörperchen (Leukozyten), sind für das Abwehrsystem des Organismus verantwortlich. Durch die Aufnahme von geformten Bestandteilen wie Krankheitskeime und Fremdkörper mittels Phagozytose unterstützen sie durch Bildung von Antikörpern die körpereigene Abwehr. Fettgewebe, das auch zu den Bindegeweben gehört dient dem Organismus als Kalorienspeicher. Zellreiches Bindegewebe Retikuläres Bindegewebe Das retikuläre Bindegewebe steht hinsichtlich seiner Zellanordnung und dem Zellaussehen dem embyonalen Bindegewebe nahe. Einem Zellgitter lagern sich retikuläre Fasern an, die selbst wieder ein räumliches Netzt aufbauen. Dabei legen sich die langen Fortsätze der Reticulumzellen den Gitterfasern so an, dass letztere fast vollständig eingeschlossen werden. Retikuläres Bindegewebe tritt als Grundgerüst im Knochenmark, in der Milz, in den Lymphknoten und den Lymphfollikeln sowie als Begleitgewebe von Nerven und Gefäßen auf. Die freien Bindegewebszellen der lymphatischen Organe liegen in den Maschenrämen des retikulären Bindegewebes und im Knochenmark befinden sich zwischen dem räumlich angeordneten Verband von Retikulumfasern die 16 Blutbildungszellen. Auf diese Weise bildet das retikuläre Bindegewebe mit den freien Zellen eine Funktionsgemeinschaft. Fettgewebe Fettgewebe kann als eine spezielle Form des retikulären Bindegewebes angesehen werden. Das Fettgewebe dient der mechanischen und thermischen Isolierung, es hat Speicherfunktion und wird als Füllgewebe eingesetzt. Mit seiner Interzellularsubstanz und seines großen Volumens spielt es eine wichtige Rolle bei der Wasserregulation im Körper. Als Baufett sind die Fettkörper mit bindegewebiger Umhüllung umgeben und es liefert das Polstermaterial für die Fußsohle, den Handteller, die Wangen, den Fettkörper der Augen, die Capsula adiposa der Niere und für das Gesäß. Es wird erst im fortgeschrittenen Hungerzustand eingeschmolzen und der Kalorienbildung zugeführt. Demgegenüber ist das Speicherfett eine Stoffwechselreserve, das einen doppelt so hohen Brennwert wie Kohlenhydrate oder Eiweiß besitzt. Die speichernde Fettzelle nimmt Fettsäuren über pinozytoseartige Einstülpungen der Plasmamembran auf. Die Entspeicherung beginnt mit der Esterspaltung an der Oberfläche des Fetttropfens. Freie Fettsäuren werden in Vesikeln zur Plasmamembran transportiert und ausgeschleust. Auch bei ausgewogener Kalorienbilanz findet in der Fettzelle ein ständiger Umbau des gespeicherten Fetts statt. Die Zellen des braunen Fettgewebes, deren Durchmesser nur etwa 30 µm beträgt, werden ebenfalls von einer Basalmembran umschlossen. In ihrem Zellleib gruppieren sich zahlreiche kleine Fetttropfen um den zentralliegenden Kern. Das Zytoplasma enthält zahlreiche Mitochondrien mit hohem Cytochromgehalt (dadurch braune Farbe) und reichlich Glycogengranula. Braunes Fettgewebe ist stoffwechselintensiver als normales Speicherfett. Es wird adrenerg innerviert und spielt eine Rolle bei der Wärmeregulation des Körpers. Es kommt bei Neugeborenen vor allem zwischen den Schulterblättern vor und ist in den ersten Lebensmonaten wichtig für die Wärmeregulation. Bei Erwachsenen kann es kaum noch festgestellt werden. Faserreiches Bindegewebe Lockeres Bindegewebe Das lockere Bindegewebe ist im Körper weit verbreitet. Es füllt Lücken zwischen Organen und verbindet sie verschieblich miteinander. Im Inneren der Organe kommt es als intersitielles Bindegewebe oder Stroma vor und lagert spezifisches Organgewebe oder Parenchym ein. Durch das lockere Bindegewebe ziehen Nerven und Blutgefäße und die wässrige Phase dient als Transportmedium zwischen den Kapillaren und den Parenchymzellen. Darüber hinaus dient es als Wasserspeicher und Verschiebeschicht. Straffes Bindegewebe Das straffe Bindegewebe tritt an Orten stärkerer mechanischer Beanspruchung auf. Es besteht aus Bündeln kollagener Fasern, denen mehr oder weniger elastische Fasern beigemischt sind. Im Vergleich zum Faseranteil bleibt der Gehalt an Zellen und Grundsubstanz im straffen Bindegewebe gering. Entsprechend der Zugbelastung in verschiedenen Richtungen ist straffes Bindegewebe oft geflechtartig aufgebaut. Innerhalb des geflechtartigen Bindegewebes verlaufen die Kollagenfasern in Bündeln, die filzartige Geflechte bilden. In dieser Ausbildung findet man es in Fascien, im Tarsus, im Pericard, in Organkapseln, im Stratum reticulare der Lederhaut und in der 17 harten Hirnhaut (Dura mater). Parallelorientierte kollagene Fasern sind als Ausdruck einseitiger Zugbelastung in den Sehnen und Bändern zu finden. Sie regulieren die Kraftübertragung von Muskeln auf Knochen. Durch Muskelzug werden die Fasern zunächst gestreckt, um so eine ruckartige Übertragung der Zugwirkung auf den Knochen zu verhindern. 2.2.2.2. Stützgewebe Typische Stützgewebe sind das Knorpel- und das Knochengewebe. Sie enthalten vorwiegend kollagene Fasern und besitzen damit die Zugfestigkeit von Bindegewebsstrukturen. Durch eine besondere Ausbildung der extrazellulären Matrix beim Knorpel und durch die Einlagerung von Kalksalzen beim Knochen wird zusätzlich die Druckfestigkeit erhöht. Knorpelgewebe Nach der Ausbildung und den Eigenschaften der Interzellularsubstanz unterscheidet man hyalinen, elastischen Knorpel und Faserknorpel. Alle drei Knorpelarten sind gefäßfrei, d.h. die Ernährung des Knorpels erfolgt durch Gefäße, die in einer bindegewebigen Hülle (Perichondrium) verlaufen. Der hyaline Knorpel überzieht als Gelenkknorpel die Gelenkflächen, bildet den Rippenknorpel, einen Teil der Nasenscheidewand, das Kehlkopfskelett und die Spangen der Luftröhre sowie die großen Bronchien. Der hyaline Gelenkknorpel besitzt als einziger keine Knorpelhaut (Perichondrium), daher kann nach seiner Zerstörung durch z.B. entzündliche oder degenerative Gelenkerkrankungen, kein funktionsfähiger Knorpel mehr aufgebaut werden. Die Interzelluklarsubstanz des hyalinen Knorpels besteht etwa aus gleichen Teilen aus kollagenen Fibrillen und Grundsubstanz, dem Chondromucoid. Die Grundsubstanz oder Chondromucoid ist ein hydratisiertes Proteoglykangemisch. Es enthält PAS-positive Glycoproteine und verschiedene Glycoaminoglykane. Mit dem Alter nimmt der Chondromucoidgehalt des Knorpels ab. Infolgedessen sinkt sein Wassergehalt (normal etwa 70%) und die Permeabilität. Der elastische Knorpel ist aufgebaut wie der hyaline Knorpel enthält aber zusätzlich ein Netzwerk aus feinen grob elastischen Fasern, die um die Chondrozyten ziehen und in das angrenzende Perichondrium einstrahlen. Elastischer Knorpel kommt in der Ohrmuschel , in Teilen des Kehlkopfskeletts, im Kehldeckel sowie in der Ohrtrompete vor. Der Faser- oder Bindegewebsknorpel besteht überwiegend aus kollagenen Fasern mit deutlicher Querstreifung und wenig Chondromucoid. Faserknorpel findet sich überall dort, wo Sehnen oder Bänder durch Druck beansprucht werden, so z.B. in den Zwischenwirbelscheiben sowie in den Gelenkscheiben (Discus und Meniscus). 18 Abb.10: Verschiedene Knorpelgewebe (links: hyaliner Knorpel, mitte: elastischer Knorpel, rechts: Faserknorpel). Knochengewebe Die Härte und Festigkeit von Knochengewebe kommt durch den hohen Gehalt an Kalziumsalzen zustande, die in der Interzellularsubstanz eingelagert sind. In den Knochen befinden sich 99% des Kalziums und 80% des Phosphats des menschlichen Körpers, woraus die Bedeutung des Stoffumsatzes im Knochen für den Kalzium - und Phosphatstoffwechsel ersichtlich wird. Die einzelnen Knochenzellen (Osteozyten) haben lange Ausläufer und sind über diese netzartig untereinander verbunden. Sie werden allseitig von Knochengrundsubstanz (Extrazellularmatrix) eingeschlossen. Die Extrazellularsubstanz ist reich an Kollagenfasern, die in einer mit den anorganischen Salzen angereicherten Grundsubstanz verlaufen. In den fertig ausgebildeten Knochen liegen die Osteozyten in kleinen Aussparungen der Interzellularsubstanz, den Knochenhöhlen. Die Osteozyten entsenden zahlreich, z.T. verzweigte Fortsätze, die in feinen, radiär orientierten Knochenkanälchen verlaufen und mit den Fortsätzen benachbarter Zellen in Kontakt stehen. Als Knochenkapsel kleidet eine 0,1 - 2 µm dicke, nicht mineralisierte Grundsubstanzschicht mit eingelagerten Fibrillen die Knochenhöhle und -kanälchen aus. Auf diese Weise wird die Knochensubstanz, dessen mineralisierte Interzellularsubstanz die Diffusion hemmt, von einem feinen Kanalsystem (VolkmannKanal) durchzogen und so den Knochen mit Nährstoffen versorgt. Nach der Anordnung von Osteozyten und Fibrillen unterscheidet man Geflechtknochen und Lamellenknochen. Der Geflechtknochen ist die phylo- und ontogenetisch ältere Form des Knochengewebes, bei dem die Fibrillen geflechtartig angeordnet sind. Das Skelett niederer Wirbeltiere (wie z.B. Amphibien) besteht vollständig aus Geflechtknochen. Er wird bei Menschen in der Embryonalperiode und auch nach einem Knochenbruch gebildet und dann in Lamellenknochen umgewandelt. Nur an wenigen Stellen z.B. an den Schädelnähten und der knöchernen Labyrintkapsel, bleibt Geflechtknochen zeitlebens erhalten. Der Lamellenknochen ist jene Art des Knochengewebes, die beim Erwachsenen den meisten Teil des Skelettes bildet. Im Aufbau eines Lamellenknochen unterscheidet 19 man einen äußeren dichten Knochen (Substantia compacta oder Compacta) von einem inneren Bälkchenknochen (Substantia spongiosa oder Spongiosa). Während die Compacta im gesamten äußeren Bereich eines Röhrenknochens vorhanden und im Schaft (Diaphyse) besonders ausgeprägt ist, ist die Spongiosa vor allem an den Enden (Epiphysen) des Knochens ausgebildet. Funktionell ist dies von Vorteil, da so mit einem Minimum an Material ein Maximum an Festigkeit erreicht wird. Durch die Ausrichtung der Spongiosabälkchen (Tabekel) kann der Knochen sich funktionell anpassen. Die Tabekel bilden sich unter dem Einfluss von Biegebeanspruchung, und zwar in Form von Druck- und Zugtabekel. Zwischen den Spongiosatabekel befindet sich das rote, blutbildende Knochenmark. Das gelbe Knochenmark (Fettmark) kommt hingegen im Bereich der Markhöhle der Diaphyse vor. Die meisten Röhrenknochen entstehen indirekt über eine knorpelige Vorstufe (chondrale oder indirekte Ossifikation), nur wenige Knochen (einige Schädelknochen sowie das Schlüsselbein) entstehen desmal d.h. durch direkte Ossifikation. Als Apophysen bezeichnet man die aus besonderen Knochenkernen hervorgehenden Anbauten von Knochen, an denen sich die Sehnen von Muskeln anheften. Abb.11: Vereinfachte Darstellung der Entwicklung eines Röhrenknochens (Pfeil nach links: Dickenwachstum, Pfeil nach oben und unten: Längenwachstum). Im Vergleich zwischen Knochen- und Knorpelgewebe, zeigt es sich, dass der gefäßreiche Knochen gut regeneriert und sich durch stetigen Umbau veränderten statischen Bedingungen anpassen kann. Demgegenüber kann der gefäßfreie Knorpel kaum regenerieren, da er nicht mit Nährstoffen versorgt werden kann. 2.2.3. Muskelgewebe Muskelgewebe ist dadurch gekennzeichnet, dass in den länglichen Zellen Myofibrillen verlaufen, die die Kontraktionsfähigkeit der Muskelzellen bewirken. Aufgrund der funktionellen Morphologie lassen sich drei Arten von Muskelgewebe unterscheiden: Quergestreiftes- oder Skelettmuskelgewebe, Herzmuskelgewebe und die glatte Muskulatur. 20 2.2.3.1. Skelettmuskulatur Aufbau Skelettmuskulatur Die Skelettmuskulatur ist mit einem Anteil am Gesamtkörpergewicht von 40 - 50% das weitaus am stärksten ausgebildete Organ des Menschen. Die skelett- oder quergestreifte Muskulatur besteht aus Muskelfasern, die 10 - 100 µm dick und bis zu 15 cm lang werden können. Muskelfasern sind vielkernige (bis zu 7000 Kerne) zylindrische Plasmastränge mit einer Vielzahl von Myofibrillen. Die Kerne liegen mit ihrer Längsachse in Richtung der Muskelfaser, und zwar unmittelbar unter der Oberfläche. Jede einzelne Muskelzelle ist vom einer Zellmembran (Sarkolemm) umgebener Zytoplasmaschlauch. Die einzelnen Muskelfasern sind zu Faserbündel zusammengeschlossen, die von Bindegewebe (Endomysium) umgeben sind und untereinander über lockeres Bindegewebe (Perimysium internum) in Verbindung stehen. In regelmäßigen Abständen ziehen tubuläre Einstülpungen vom Sarkolemm transversal in das Innere der Muskelfaser, sog. t-Tubuli (transversal Tubuli). Eine weitere Besonderheit von Muskelzellen sind die zwischen den transversalen Tubuli in Längsrichtung zu den Myofribrillen angeordnete longitudinalen Tubuli, sog. l-Tubuli (longitudinales System). Die l-Tubuli sind eine Röhrensystem des endoplasmatischen Retikulums (in Muskelzellen = sarkoplasmatisches Retikulum), das ein Reservoir für Kalziumionen darstellt, die bei einer Kontraktion schnell freigesetzt werden können. Innerhalb einer Muskelfaser liegen zwischen 40 und 500 Rezeptororgane, so genannte Muskelspindeln, die spezifische Dehnungsrezeptoren enthalten. Über diese Rezeptoren werden Längenänderungen der Faser registriert und über spezifische Nervenfasern ans Rückenmark weitergeleitet. Zusätzlich zu den Muskelspindeln enthalten Muskeln am Muskel-Sehnen-Übergang sogenannte Sehnenrezeptoren (Golgi-Sehnenorgane), die den Muskel vor Überspannung schützt. Molekulare Mechanismen der Kontraktion Skelettmuskeln enthalten pro Gramm Gewicht etwa 100 mg kontraktile Moleküle, das Actin und das Myosin. Die kontraktilen Proteine Actin und Myosin bilden die dicken und dünnen Myofilamente der Myofibrillen. Zwischen den schlauchförmigen Myofibrillen sind Mitochondrien oder Sarkosomen eingelagert. Die Myofibrillen sind durch Trennwände in zahlreiche etwa 2,5 µm lange Fächer, den Z-Scheiben oder Sarkomere aufgeteilt. Die Querstreifung der Myofibrillen entseht aufgrund der regelmäßigen Anordnung der Actin- und Myosinfilamente. In der Mitte jedes Sarkomers liegen dicke Filamente aus Myosin mit einem Durchmesser von etwa 10 nm. Dem Bündel stehen zu beiden Seiten des Sarkomers die Actinfilamente gegenüber die nur einen Durchmesser von 5 nm haben und wie die Borsten einer Bürste an den Z-Scheiben befestigt sind. Die Bündel der 1,6 µm langen Myosinfilamente in der Mitte des Sarkomers erscheinen im Lichtmikroskop als dunkle, im polarisierten Licht doppelbrechende, d.h. anisotrope Bande (A-Banden). Diese grenzen zu beiden Seiten an die dünneren, und deshalb im Lichtmikroskop hell erscheinenden, d.h. isotrope Bande (I-Banden) der Actinfilamente. 21 Abb.12: A Bandstruktur der Myofibrillen. Links im erschlafften, rechts im kontrahierten Zustand. B Anordnung der Myosin- und Actinfilamente im erschlafften und kontrahierten Zustand. Im ruhenden Muskel überlappen sich die Enden der Actin- und Myosinfilamente an der Grenze zwischen A- und I- Band nur wenig. Die Muskelverkürzung resultiert aus der Verkürzung unzähliger Sarkomere. Dabei gleiten die dünnen Actinfilamente über die Myosinfilamente, zwischen denen sie sich hindurchschieben, und so tief in das Bündel Myosinfilamente hinein, bis sie die Sarkomermitte erreichen. Bei der Verkürzung eines Sarkomers gleiten die Myosin- und Actinfilamente so übereinander ohne sich selbst zu verkürzen. Auch bei der Dehnung des Muskels ändert sich die Filamentlänge nicht. Vielmehr wird das Bündel der Actinfilamente aus der Anordnung der Myosinfilamente herausgezogen, wobei der Grad der Filamentüberlappung abnimmt. Dieser Gleitmechanismus, bei der bei einer Muskelkontraktion bzw. Muskeldehnung, nicht die beteiligten Filamente verkürzt oder dehnt, kommt durch die Querbrücken zwischen den Actin- und Myosinfilamenten zustande. Die Querfortsätze eines Myosinfilaments werden aus den etwa 20 nm langen Köpfen von etwa 150 Myosilmolekülen gebildet, die in einer bipolaren Anordnung zum Filament gelagert sind. Ein jeder Myosinkopf kann als Querbrücke im Kontraktionsprozess ein Myosinfilament mit einem benachbarten Actinfilament verbinden. Durch die "Kippbewegung" der Köpfe ziehen diese die Actinfilamente in Richtung Sarkomermitte. Abb. 13: Verschiedene Muskelgewebe im Längsschnitt (links: glattes Muskelgewebe, mitte: quergestreifte Skelettmuskulatur, rechts: Herzmuskulatur) 22 Regulation der Muskelkontraktion Muskel werden durch Aktionspotentiale (siehe Kapitel Nervengewebe) der innervierenden Motorneurone erregt, die - via neuromuskulärer Übertragung an den motorischen Endplatten - Muskel-Aktionspotentiale auslösen. Das Auslösen einer Muskelzuckung kann in folgenden Schritten zusammengefasst werden: Reizung der Muskelfaser Aktionspotential bzw. Membranerregung des Muskels Elektromechanische Kopplung Erregungsleitung im T-System Kalziumfreisetzung aus dem L-System Kalziumeinwirkung auf die Myofibrillen Kontraktion der Myofibrillen Bei der elektromechanischen Kopplung des Signals von der erregten Zellmembran zu den in der Tiefe der Zellen liegenden Myofibrillen spielt Kalzium die entscheidende Rolle. Das Muskel-Aktionspotential breitet sich entlang der Membranen des T-Systems in das Innere der Zelle aus. Von dort springt sie auf das L-System über und bewirkt die Freisetzung der in den Terminalzisternen gespeicherten Kalziomionen in die Zellflüssigkeit um die Myofibrillen und löst dadurch eine Kontraktion aus. Die Kontraktion hört auf, wenn die aktivierenden Kalziumionen durch einen aktiven Transportmechanismus, der Kalziumpumpe, wieder in das sarkoplasmatische Retikulum zurücktransportiert werden, was eine Hemmung der Actomyosin-ATP-ase und damit eine Muskelerschlaffung bewirkt. Abb.14: Wirkung der Kalziumionen a. und Funktionsweise Querbrücken b. 23 Der Verlauf und die Dauer von Muskelzuckungen variiert bei der quergestreiften Skelettmuskulatur beträchtlich. Schnelle phasische Muskelfasern arbeiten mit einer Zuckungsdauer von 30 - 40 ms, langsame tonische Fasern mit 100 ms. Phasische Muskeln werden aufgrund ihres geringen Myoglobingehaltes (ähnlich Hämogobin, sauerstoffspeicherndes Protein) als "weiße Muskeln", während tonische Muskeln mit hohem Myoglobingehalt als "rote Muskeln" bezeichnet werden. Tonische oder "rote" Muskulatur ist zu Dauerleistung wie z.B. die Haltearbeit beim Stehen fähig, während die phasische oder "weiße" Muskulatur kurzfristig Höchstleistungen erbringen kann, aber schnell ermüdet. Das Phänomen des "Muskelkaters" ist offenbar nicht, wie man lange vermutet hat, auf lokale Anhäufungen von Milchsäure oder andere Stoffwechselprodukte im Muskel zurückzuführen. Vielmehr scheint es bei der Überbeanspruchung von Muskel zu kleinen Verletzungen (Mikroläsionen) von Muskelfasern zu kommen, die dann die Schmerzen verursachen. Bei der Leichenstarre, die meist 4 - 10 Stunden nach Eintritt des Todes auftritt, fehlt aufgrund des nicht mehr vorhandenen Stoffwechsels das ATP, das im Anschluss an eine Muskelkontraktion, die Verbindung der Actin- und Myosinfilamente wieder löst. 2.2.3.2. Herzmuskulatur Eine besondere Form der quergestreiften Muskulatur ist das Herzmuskelgewebe. Im Unterschied zu den randständigen Kernen des Skelettmuskels liegen die einzelnen voluminöseren Kerne der Herzmuskelzelle meist zentral. Außerdem sind die sich verzweigende Herzmuskelzellen im Querschnitt kleiner als Skelettmuskelzellen. Über die Verzweigungen treten die Herzmuskelzellen miteinander in Kontakt und bilden so eine Art Netzwerk. Die Kontaktstellen zwischen den Zellen verlaufen senkrecht zur Längsachse der Zelle und werden als "Glanzstreifen" bezeichnet. Der Zusammenhalt der Kontaktstellen wird durch eine Kittsubstanz gewährleistet, was dem Glanzstreifen funktionell Ähnlichkeit mit Desmosomen gibt. An der Innenfläche der verhefteten Membranareale sind die Actinfilamente der Myofibrillen befestigt. Über zytoplasmatische Verbindungen zwischen benachbarten Herzmuskelzellen, die Nexus, wird die Erregungsausbreitung im Herzmuskelgewebe garantiert. Sonst entspricht der Aufbau der Herzmuskelzelle dem des Skelettmuskels. Actin- und Myosinfilamente sind ebenso angeordnet, Myofibrillen gruppieren sind und Mitochondrien, sarkoplasmatisches Retikulum sowie t-Tubuli sind ihnen zugeordnet. Die Besonderheit des Herzmuskels liegt in der Fähigkeit eines Teils seiner Zellen, Erregung nicht nur als Antwort auf einen von außen kommenden Reiz, sondern spontan auszubilden. Morphologisch und funktionell sind zwei Typen von Herzmuskelzellen zu unterscheiden: Die Fasern der Arbeitsmuskulatur (Arbeitsmyocard) der Vorhöfe und der Ventrikel, die die Hauptmasse des Herzens ausmachen und die die mechanische Pumparbeit leisten. Die Fasern des spezifischen Erregungsbildungs- und -leitungs-Systems, die im Dienste der Autorhythmie des Herzens stehen (Sinuskoten, AV-Knoten, His-Bündel und Purkinje-Fasern). Die rhythmischen Pulsationen des Herzens werden durch Erregungen ausgelöst, die im Herzen selbst entstehen. So geht der Anstoß zu einem Herzschlag vom Sinusknoten aus, der im rechten Vorhof liegt. Er treibt das Herz bei Körperruhe mit einer Frequenz von ca. 70 Impulse/min an (ungefähr 1 Hz = 1 Impuls pro sec). Vom Sinusknoten breitet sich die Erregung über die Arbeitsmuskulatur beider Vorhöfe aus. Bei der Passage des 24 Erregungsleitungssystems erfolgt im Atrioventrikularknoten (AV-Knoten) zunächst eine Verzögerung. Das anschließende His-Bündel, die Schenkel und ihre Endaufzweigungen, die Purkinje-Fäden leiten die Erregung mit einer Leitungsgeschwindigkeit von ca. 2 m/s weiter, so dass die verschiedenen Regionen der Herzkammer rasch nacheinander erregt werden. Von den Purkinje-Fasern breitet sich die Erregung dann mit einer Geschwindigkeit von 1 m/s über die Kammermuskulatur aus. Abb.15: Schema der Anordnung der Erregungsbildungs- und -leitungsfasern des Herzens (Frontalansicht). Die Autorhythmie des Herzens steht und fällt nicht mit der Funktionsfähigkeit des Sinusknotens, da auch die übrigen Teile des Erregungsleitungs-Systems die Fähigkeit zur automatischen Erregungsbildung besitzen. Diese nimmt jedoch mit der Entfernung zum primären Schrittmacher, dem Sinusknoten beträchtlich ab. Fällt die Erregungsbildung im Sinusknoten aus, so kann ersatzweise der AV-Knoten als sekundäres Erregungsbildungs-Zentrum Schrittmacher-Funktion übernehmen (AVKnoten Rhythmus = 40 - 60 Impulse/min) . Im Falle einer kompletten Unterbrechung der Erregungsleitung von den Vorhöfen auf die Herzkammern (totaler Herzblock) auch ein tertiäres Zentrum im ventriculären Erregungsleitungs-System als Schrittmacher der Kammerautonomie einspringen. Im Gegensatz zur Skelettmuskelfaser hat die Herzmuskulatur eine relativ Lange Refraktärperiode. Die Refraktärperiode bezeichnet den Zeitraum indem die Ansprechbarkeit auf Reize während bestimmter Phasen des Erregungsablaufs aufgehoben oder vermindert ist. Durch ihre langdauernde Refraktärzeit wird die Muskulatur des Herzens vor einer zu schnellen Wiedererregung geschützt, die ihre Pumpfunktion beeinträchtigen würde. 2.2.3.3. Glatte Muskulatur Glatte Muskelzellen kommen hauptsächlich in der Wand von Eingeweiden MagenDarm-Trakt, Gallenblase, harnableitende Wege, Geschlechtsorgane und Gefäßen vor. Außerdem kommt es in den tiefen Atemwegen, am Auge und im Drüsengewebe vor. Die glatte Muskulatur steht unter dem Einfluss des autonomen (vegetativen) Nervensystems, d.h. es kann im Gegensatz zur Skelettmuskulatur nicht willentlich gesteuert werden. Außerdem wird die glatte Muskulatur in vielen Organen durch passive Dehnung (myogene Erregung) erregt. 25 Glatte Muskelzellen sind spindelförmig, etwa 50 - 400 µm lang und 2 - 10 µm dick. Verknüpft durch Desmosomen bilden sie ein mit Kollagenfasern durchsetztes Netzwerk. Da die Myosin- und Actinfilamente nicht regelmäßig angeordnet sind, fehlt den glatten Muskelzellen die für Skelettmuskeln typische Querstreifung. Die Kontraktion von glatten Muskelzellen erfolgt ebenfalls durch ein Übereinandergleiten der Myofilamente; die Filamentverschiebung und die damit einhergehende Spaltung des ATP erfolgt jedoch 100 bis 1000mal langsamer als bei quergestreiften Muskeln. Dadurch sind glatte Muskeln besonders geeignet für energiesparende Halteleistungen. Bezogen auf den Muskelquerschnitt unterscheidet sich die Kontraktionsspannung und die Halteleistung der glatten Muskulatur nicht von der Skelettmuskulatur. Im Darm kann durch gleichmäßige Kontraktionswellen (Peristaltik) der glatten Muskulatur der Darminhalt transportiert werden, aber auch verharren in einem bestimmten Kontraktionszustand (Tonus) z.B. am Schließmuskel des Übergangs MagenZwölffingerdarm. Die myogene Erregung der glatten Muskulatur entseht in Schrittmacherzellen, die sich von anderen Muskelzellen nicht morphologisch, aber aufgrund ihrer elektrophysiologischen Eigenschaften unterscheiden. Die Ausbreitung der Erregung erfolgt über besondere Nahtstellen (Nexus) zwischen den Zellmembranen aneinandergrenzender Muskelzellen. Die Erregung breitet sich mit einer Geschwindigkeit von 5 - 10 cm/s im glatten Muskel aus. Myogene Rhythmen sind periodische Schwankungen im Sekunden- bzw. Minutenbereich des myogenen Tonus, die durch spontane Aktivitätsänderungen der Schrittmacherzellen ausgelöst wird. 2.2.4. Nervengewebe Das Nervensystem, bei dem man zentrale und periphere Abschnitte unterscheidet, dient der Aufnahme und Verarbeitung von Reizen, die mit der Ausbildung und Weiterleitung von Erregung beantwortet werden können. Seine materielle Grundlage ist die Nervenzelle bzw. das Neuron. Das menschliche Gehirn oder Zentralnervensystem (ZNS) besitzt etwa 25 Milliarden solcher Nervenzellen. Nur etwa 25 Millionen Nervenzellen befinden sich in der Peripherie oder verbinden die Peripherie mit dem Zentralnervensystem. Die Nervenzellen sind über Synapsen vielfältig miteinander verknüpft, und deren Zahl ist 1000fach größer als die der Nervenzellen. Über synaptische Kontakte sind Nervenzellen auch mit anderen Zelltypen verknüpft, hauptsächlich mit Rezeptoren für die Aufnahme von Information (in den Sinnesorganen), oder mit Effektoren z.B. den Muskelzellen. Die Nervenzelle besteht aus einem Zellkörper, dem Soma (Perikaryon) mit dem Zellkern, sowie verschiedenen Fortsätzen, den Dendriten und dem Axon oder Neurit. Abb.16: Vereinfachte Darstellung eines motorischen Neurons 26 Die Dendriten (bis zu 1000 pro Zelle) empfangen Reize aus vorgeschalteten Nervenzellen oder rezeptiven Strukturen über Synapsen und leiten sie zum Soma oder Perikaryon weiter. Vom Soma aus wird der Reiz über das Axon zu dem entsprechenden Erfolgsorgan (z.B. Skelettmuskel) oder zu einer anderen Nervenzelle weitergeleitet. Die Richtung oder Polarität der Erregung einer Nervenzelle geht also immer vom Dendriten (Aufnahme = rezeptive Struktur) über das Soma (Zellleib und Stoffwechselzentrum) zum Axon der effektorischen Struktur. Aufgrund ihres Aufbaus kann man verschiedene Nervenzellen unterscheiden. Apolare Nervenzellen sind modifizierte Neurone die weder einen Dendrit noch ein Axon ausbilden. Sie bestehen nur aus einem Soma das am Rezeptorpol der Nervenzelle Härchen ausgebildet hat, worüber die Information aufgenommen wird. Apolare Nervenzellen kommen als Sinneszellen im Innenohr und in den Geschmacksknospen vor. Unipolare Nervenzellen haben Fortsätze entwickelt, die als Axon ausgebildet werden. Auch hier wird über den Rezeptorpol des Soma der Reiz aufgenommen und über das Axon zum Effektororgan weitergeleitet. Man findet sie in den Stäbchen und Zapfen der Netzhaut und als Riechzellen. Bipolare Nervenzellen haben jeweils einen Dendriten und das Axon ausgebildet, die sich an den gegenüberliegenden Seiten des Soma befinden. Bipolare Nervenzellen kommen besonders im Ganglion retinae (1. Verschaltung der Netzhaut) sowie in den beiden Ganglien des Nervus vestibulo-cochlearis vor. Pseudounipolare Nervenzellen entstehen aus bipolaren Zellen, indem der Dendrit und das Axon unmittelbar am Soma miteinander verschmelzen. Bei multipolaren Nervenzellen führen mehrere, weit verzweigte Dendriten zum Soma während am gegenüberliegenden Teil des Somas das Axon ansetzt. Im Gehirn und Rückenmark sind alle Nervenzellen multipolar. Ist das Axon lang und zweigt sich erst entfernt vom Soma auf, spricht man von Neuronen des Deiters-Typs. Bei Neuronen des Golgi-Typs verzweigt sich das kurze Axon nahe am Soma. Die Purkinje-Zellen der Kleinhirnrind und die Pyramidenzellen der Großhirnrinde sind besondere Formen multipolarer Zellen. 2.2.4.1. Grundlagen der Informationsübertragung von Nervenzellen Die Nervenzellen sind im Organismus auf die Verarbeitung von Information spezialisiert. Diese Informationsverarbeitung als im Organismus universelles Kommunikationsmittel, lässt sich aufteilen in die Aufnahme der Information, ihre Fortleitung über größere Distanz, sowie deren Weitergabe an eine oder mehrere andere Zellen. Die Informationsübertragung erfolgt über elektrische Potentialdifferenzen. Dabei stellt die Ladungsverteilung an der Membran die Grundlage der Funktion von Nerven- und Muskelzellen dar. 2.2.4.2. Das Ruhepotential Nahezu alle Zellen weisen ein durch elektrische Spannungsdifferenzen zwischen der Zelloberfläche und dem Zellinneren erzeugtes negatives Ruhepotential auf. D.h. das Zellinnere ist durch einen Überschuss an negativen Ionen (Anionen = A-) im Gegensatz zum Extrazellulärraum, mit einem Überschuss an Kationen (positiv geladene Ionen), negativ geladen. Das Ruhepotential zwischen Zellinnerem und -äußerem stellt sich ein, weil die Zellmembran kein vollkommener Isolator ist, und für gewisse Ionenarten beschränkt durchlässig (permeabel) ist. Dies gilt hauptsächlich für die relativ kleinen K+- Ionen, die 27 durch Poren die Membran passieren können. Aufgrund der höheren Konzentration der K+-Ionen im Zellinneren wird aufgrund der osmotischen Druckverhältnisse der NettoAusstrom von K+-Ionen angetrieben. Dieser Ausstrom würde schnell zu einem Ausgleich des osmotischen Drucks für K+ bzw. der K+-Konzentration führen, wenn dies nicht durch eine entgegengerichtete gleich große Kraft verhindert würde. Diese Gegenkraft entsteht aufgrund der elektrischen Ladung der K+-Ionen. Strömt, getrieben durch die osmotische Druckdifferenz, ein K+ aus der Zell, so nimmt es eine positive Ladung mit und erzeugt am Membrankondensator eine positive Aufladung der Außenseite, der eine gleich große negative Aufladung der Innenseite entspricht. Dieses so entstandene Membranpotential ist so gerichtet, dass es dem weiteren Ausstrom weiterer Kationen entgegenwirkt: Positives Potential stößt positive Ionen ab. Das Membranpotential wächst so lange, bis die dem K+ Ausstrom entgegenwirkende Kraft gleich groß wird wie der osmotische Druck der K+-Ionen. Bei diesem Potential sind der Ein- und Ausstrom der K+ im Gleichgewicht, man nennt es deshalb das K+Gleichgewichtspotential (EK). Das K+-Gleichgewichtspotential wird also bestimmt durch das Konzentrationsverhältnis der Kaliumionen innerhalb und außerhalb der Zelle. Das Ruhepotential (etwa -55 bis -100 mV) einer Nervenzelle entspricht weitgehend dem K+Gleichgewichtspotential (-97 mV), das durch die Nernst-Gleichung für Diffusionspotentiale beschrieben wird: R x T Eion = z x F x In R T z F extrazell. Konz. des Ions intrazell. Konz. des Ions = Gaskonstante = absolute Temperatur = die Wertigkeit des Ions = Faraday-Konstante Daraus wird ersichtlich das relativ kleine Änderungen der extrazellulären Kaliumionen Konzentration zu drastischen Veränderungen des Ruhepotential führen können, was die Funktion der Zelle beträchtlich beeinflussen kann. Das Ruhepotential ist bei normalem niedrigeren extrazellulären K+-Konzentrationen bis zu 30 mV weniger negativ als das K+-Gleichgewichtspotential. Der Grund dafür ist, dass die Membran nicht vollständig impermeabel für Natriumionen (Na+) ist. Für Na+ besteht ein starkes Konzentrationsgefälle von außen nach innen (etwa 10 : 1) und ein Einstrom von Na+ in die Zelle wird zusätzlich durch das negative Membranpotential im Zellinneren gefördert. Selbst bei einer geringen Permeabilität für Na+ der Membran werden diese in die Zelle strömen und die Amplitude des negativen Ruhepotentials verringern. Abb.17: Membranladung beim Ruhepotential 28 Um wiederum die Funktion der Zelle zu erhalten muss dieser, wenn auch geringe, Na+ Einstrom kompensiert werden. Dies geschieht über einen aktiven Transportmechanismus der Natrium-Kalium-Pumpe, der Na+ aktiv mit Hilfe von Stoffwechselenergie, gegen das Konzentrationsgefälle aus der Zelle pumpt. Der aktive Transport von Na+ aus der Zelle hat eine Komponente die an den Einstrom von K+ in die Zelle gekoppelt ist und damit energiesparend arbeitet. Dies ist für den Energiehaushalt und damit für die Funktion der Zellen immens wichtig: Für Muskelzellen wird geschätzt, dass 10 - 20% des Ruhestoffwechsels für die Aufrechterhaltung des Ruhepotentials, d.h. für den aktiven Ionentransport verbraucht wird. Die Natrium-Kalium-Pumpe funktioniert nach folgendem Schema: An der Innenseite der Membran verbinden sich Na+ mit einem Trägermolekül Y zu einem Molekül NaY. NaY diffundiert durch die Membran und zerfällt an der Membranaußenseite zu Na+ und Y . Das Trägermolekül Y wird an der Außenseite in das Trägermolekül X umgewandelt, und verbindet sich mit K+ zu KX. KX diffundiert dann an die Innenseite der Membran, wo wiederum K+ entlassen wird. Das Trägermolekül X wird über die Spaltung von ATP zu ADP wieder in das Trägermolekül Y verwandelt und nimmt Na+ auf. 2.2.4.3. Das Aktionspotential Folgender Mechanismus liegt einem Aktionspotential zugrunde: Durch eine Erregung die auf die Nervenzelle trifft, werden kurzfristig Membrankanäle (spannungsabhängige Kanäle) in der Zellmembran geöffnet, die für Na+-Ionen durchlässig sind. Dadurch diffundieren Natriumionen entlang ihres Konzentrationsgefälles in das Zellinnere, was dort zu einem Überschuss an positiv geladenen Ionen führt. Das Membranpotential verändert sich von z.B. -90 mV auf +20 mV, d.h. die Polarisation der Membran verändert sich und man spricht von der Depolarisationsphase. Diese schnelle Potentialänderung wird als Aufstrich oder "overshoot" des Aktionspotentials bezeichnet. Der Aufstrich oder "overshoot" dauert nur 0,2 -0,5 ms. Danach schließen sich die Natriumkanäle wieder und für kurze Zeit öffnen sich jetzt mehr Kaliumkanäle. Dadurch strömen mehr Kaliumionen aus der Zelle und repolarisieren (Repolarisationsphase) die Zellmembran wieder hin zum Ruhepotential. Nachdem sich die Kaliumkanäle ebenfalls wieder geschlossen haben, werden unter ATP-Verbrauch die sich im zellinneren befindenden Natriumionen mittels der Ionenpumpen nach außen transportiert. Erst wenn das Ruhepotential der Membran wieder erreicht ist, kann die Zelle erneut gereizt werden. Während der Zeit zwischen Aufstrich und Repolarisation, kann die Zelle nicht erregt werden. Dieser Zeitraum wird als Refraktärzeit bezeichnet. Er begrenzt die Erregbarkeit einer Zelle und beeinflusst damit auch die Leitungsgeschwindigkeit. Die Refraktärszeit einer Nervenzelle beträgt im Durchschnitt 1 ms, die einer Skelettmuskelzelle 2 - 4 ms und die einer Herzmuskelzelle 200 ms. Die Weiterleitung des Aktionspotentiales auf der Oberfläche der Nervenzelle bzw. entlang des Axons beruht darauf, dass das lokal angestiegene Membranpotential benachbarte Ionenkanäle öffnet, wodurch sich die Membranerregung als Depolarisationswelle über die ganze Zelle ausbreitet. Die Geschwindigkeit, mit der sich solch eine Erregung ausbreitet, liegt zwischen wenigen Metern (vegetative Nervenzellen) bis zu 120 m/s (motorische Nerven der willkürlich innervierten Muskeln). 29 Abb.18: Zeitverlauf eines Aktionspotentials Um diese Leitungsgeschwindigkeit zu garantieren sind die Axone von höheren Organismen mit einer unterschiedlich dicken Isolierschicht, der sogenannten Myelinoder Markscheide, auch Schwann-Scheide umgeben. Diese phospholipidhaltigen Membranen sind in bestimmten Abständen durch Einschnürungen, den RanvierSchnürringen durchbrochen. Die Bedeutung der Markscheide (neben der Eigenschaft als Isolationsschicht) für die Erregungsleitung liegt darin, dass bei markhaltigen Nervenfasern eine sprunghaft von Schnürring zu Schnürring ablaufende saltatorische Erregungsleitung möglich ist. 2.2.4.4. Synapsen Die Übertragung der Erregung von einer Nervenzelle zu einer anderen Nervenzelle oder zu einer Muskel oder Drüsenzelle erfolgt über Synapsen. Hierbei erfolgt die Übertragung in den meisten Fällen mit Hilfe von chemischen Überträgerstoffen, den so genannten Neurotransmittern. Im Bereich von Synapsen liegen die Zellen meistens eng beieinander. Man unterscheidet drei Komponenten: Die präsynaptische Membran, den synaptischen Spalt und die postsynaptische Membran. Im Bereich der präsynaptischen Membran ist am Axon meist eine kolbenförmige Verdickung, der synaptische Endkopf (von Nervenzelle zur Skelettmuskelzelle = motorische Endplatte) zu erkennen, indem zahlreiche Vesikel, den so genannten synaptischen Bläschen enthalten sind. Trifft ein Aktionspotential am synaptischen Endkopf ein, werden die in den Vesikeln gespeicherten Neurotransmitter durch Exozytose in den synaptischen Spalt ausgeschüttet. Die Transmitter diffundieren durch den 10 - 50 nm breiten synaptischen Spalt zur postsynaptischen Membran und werden dort an entsprechende Rezeptoren gebunden. Diese Bindung führt entweder zu einer Depolarisation der postsynaptischen Membran und damit zu einer Weiterleitung der Erregung (exzitatorische Synapse) oder zu einer Hyperpolarisation und damit zu einer Hemmung der Weiterleitung (inhibitorische Synapse). 2.2.4.5. Neurotransmitter Als Derivat der Essigsäure ist das Acetylcholin der Neurotransmitter von neuromuskulären Synapsen des vegetativen Nervensystems und des ZNS von allen Wirbeltieren. Bei Dopamin, Noradrenalin und Adrenalin (Derivate der Aminosäuren) handelt es sich um Catecholamine, die ebenfalls sowohl im vegetativen Nervensystem als auch im ZNS vorkommen. Ebenfalls ein Derivat der Aminosäuren, das Monoamin Serotonin, kommt nur im ZNS vor. Der Neurotransmitter Glutaminsäure kommt in erregenden Synapsen, GABA (Gamma-Amino-Buttersäure) und Glycin in hemmenden Synapsen des ZNS vor. 30 An einer Nervenzelle können viele Dutzend bis einige tausend Synapsen enden. Je nachdem ob das Axon an einem Dendriten, einem Soma oder am Axon einer anderen Nervenzelle endet, spricht man von axodentritischen, axosomatischen oder axoaxonischen Synapsen. Elektrische Synapsen so genannte Gap junctions kommen vereinzelt nur bei niederen Wirbeltieren vor. Über einen direkten zytoplasmatischen Kontakt zwischen zwei Nervenzellen der nur 2 -4 nm breit ist, können Ladungsträger in Form von anorganische Ionen passieren und den Nervenimpuls ohne nennenswerte Verzögerung (< 0,1 ms) weiterleiten. 2.2.4.6. Gliazellen Ursprünglich sind Gliazellen lediglich als sogenannte Ernährungs- und Stützzellen der Neurone angesehen worden. Die häufigste Form von Gliazellen im Gehirn sind die Astrozyten. Sie versorgen die Nervenzellen, stellen Verbindungen zwischen ihnen her und umkleiden die Synapsen. Oligodendrozyten umhüllen die Axone von Hirnneuronen während im peripheren Nervensystem die Schwannzellen diese Aufgabe übernehmen. Glia- und Nervenzellen arbeiten im Gehirn und Rückenmark eng zusammen. Glia umkleidet Synapsen und umhüllt und verbindet die langen Axone miteinander, über die die Nervenimpulse gesendet werden. An all diesen Kontaktpunkten "belauschen" die Gliazellen die Signale der Neuronen und obwohl sie selbst nicht depolarisierbar sind, können sie eigene Signale versenden und darüber z.B. die Signalstärke an Synapsen beeinflussen. Mittlerweile wird sogar die Hypothese aufgestellt, dass Gliazellen besonders die Astrozyten, eine entscheidende Rolle bei Lern- und Gedächnissvorgängen einnehmen. Abb.19: Neurone und Gliazellen 31 2.3. Biologische Medizin als Grundlage für Bioresonanzverfahren Die biologische Medizin befasst sich mit den biochemischen und biophysikalischen Grundlagen von Regulationsvorgängen im Körper. Dabei werden diese Regulationsvorgänge bis zu den Mechanismen zwischen einzelnen Gewebszellen und innerhalb der extrazellulären Matrix berücksichtigt. Mit dem Methodenspektrum dieser Fachrichtung ist es möglich, die Ursachen von Störungen der regulatorischen Vorgänge innerhalb eines Organismus zu finden und diese gezielt zu behandeln. Es werden also nicht nur Krankheitssymptome behandelt, sondern Krankheitsursachen. In der biologischen Medizin wird von der Grundlage ausgegangen, das Körpervorgänge durch Regelkreise bestimmt werden, deren funktionieren die Voraussetzung dafür bildet, dass ein Organismus gesund ist. Die Funktionselemente eines kybernetischen Regelkreises sind: 1. Das Messwerk (der Sensor, z.B. eine Sinneszelle in der Haut), mit dessen Hilfe der Istzustand in der Messstrecke erfasst und in Signale transformiert wird (elektrische Erregbarkeit von Axonen der Nervenzellen), die an den Regler (Somata der Nervenzellen im Ganglion oder im Gehirn) weitergegeben werden. 2. Im Regler wird der Istzustand mit einer meist extern vorgegebenen Führungsgröße (von einer Drüse oder einem Organ) verglichen und Abweichungen zwischen Sollwert und Istwert in Signale umgewandelt, die als Stellgröße an das 3. Stellglied (andere Drüse oder Organ) weitergegeben werden. Die Aktion des Stellgliedes korrigiert nun das vom Sensor gemessene Signal, bis Ist- und Sollwert übereinstimmen. In der klinischen Forschung ist es nicht möglich die Funktionsweise eines solchen Regelkreises zu beobachten, wohl aber Regelsysteme, die nach dem gleichen System arbeiten. Ein Regelsystem setzt einen Mechanismus aufeinander bezogener Sollwerte voraus. Aufgabe eines derartigen Wirkungsgefüges ist die Aufrechterhaltung der Homöostase. Diese stellt einen vieldimensionalen Sollwert dar, um den ein Istwert als zulässige Differenz (Toleranz) schwingt. Dadurch ist dieses System in der Lage, eine durch Störgrößen (von außen oder innen) ausgelöste Abweichung mit geringem Energieverlust zu korrigieren. Gesundheit entspricht daher aus kybernetischer Sicht einer Norm, die ein Organismus bestrebt ist, aufrecht zu erhalten. Die Funktion eines Regelkreises - oder Systems ist durch die Regelgüte charakterisiert, die durch die Eigenschwingung einer oder mehrerer Parameter bestimmt wird. Einer optimalen Regelgüte entspricht ein gedämpftes Eigenschwingverhalten oder eine asymptotische Rückkehr zum Sollwert. Dadurch wird das Regelziel schnell und mit geringem Energieverlust erreicht. Dies entspricht dem Toleranzbereich von Gesundheit. 32 12 Stellglied Sollwert 10 Toleranzbereich 8 6 4 Istwert 2 Zeit 0 0 2 4 6 8 10 12 14 Abb.20:Beispiel für ein "gesundes Regelsystem". Der vom Messwerk (Sensor) gemessene Istwert wird vom Stellglied auf den Sollwert hoch reguliert. 12 Stellglied Istwert 10 8 6 Toleranzbereich 4 Sollwert 2 Zeit 0 0 2 4 6 8 10 12 14 Abb. 21: Beispiel für ein "gesundes Regelsystem". Der vom Messwerk (Sensor) gemessene Istwert wird vom Stellglied auf den Sollwert herunter reguliert. Bei krankhaften Veränderungen in einem Organismus kommt es zu Regelformen mit einem periodisch entarteten, labilen Eigenschwingverhalten. Durch schnelles und überschießendes Auslenkung von Parametern innerhalb eines Regelsystems wird das Regelziel überschritten und kann erst in mehreren Nachschwankungen wieder korrigiert werden (z.B. akute Entzündung). 33 Eine Sonderform der periodisch entarteten Reaktion ist die Aufklingreaktion, bei der die Nachschwankungen um den Sollwert zu erreichen immer höher werden, bis keine Kompensation mehr möglich ist und das System in einer Kippreaktion zusammenbricht (z.B. alle Formen der Schockreaktion). Bei einer aperiodisch-trägen entarteten Reaktion wird das Regelziel verspätet oder gar nicht erreicht (z.B. bei chronischen Entzündungen und Malignome). Kann sich wiederum außerhalb des Toleranzbereich des Sollwertes eine Gleichgewicht etablieren, so wird das Beibehalten des falschen Sollwertes auf Dauer den Organismus schädigen und es kommt zu chronischen Erkrankungen. Viele Regelsystemstörungen können vom Organismus über eine kurzzeitige Sollwertverschiebung kompensiert werden. Treten verschiedene Störungen auf so kommt es zu Summationseffekten, die eine bestimmte Symptomatik auslösen. Die Behandlung von lediglich einzelnen Störungen verlagert nur die aufgetretene Symptomatik und es kann keine endgültige Heilung erreichen werden. 11 Stellglied Sollwert 9 Toleranzbereich 7 5 Istwert 3 Zeit 1 0 2 4 6 8 10 12 14 Abb. 22: Beispiel für ein "krankes Regelsystem". Der vom Messwerk (Sensor) gemessene Istwert kann vom Stellglied nicht auf den Sollwert hoch reguliert werden. 34 12 Stellglied Istwert 10 8 6 Toleranzbereich 4 Sollwert 2 Zeit 0 0 2 4 6 8 10 12 14 Abb. 23: Beispiel für ein "krankes Regelsystem". Der vom Messwerk (Sensor) gemessene Istwert kann vom Stellglied nicht auf den Sollwert herunter reguliert werden. Die Symptomatik einer akuten Erkrankung mit der prinzipiellen Rückkehr zur Norm und die verstellte Norm einer chronischen Krankheit, ohne die Möglichkeit zur völligen Genesung, dürfen nicht miteinander verwechselt werden. Leider wird in der Schulmedizin der diffuse, unklare Zustand einer "Befindungsstörung" dem am nächst liegenden klinischen Syndrom zugeordnet und entsprechend behandelt. Es wird dabei nicht berücksichtigt, dass dabei ein chronisches Leiden, das durch die "Verstellung" eines Regelsystems zustande gekommen ist, behandelt wird wie eine akute Krankheit. Es gibt mehrere Hauptursachen für eine regulatorische Entartungen der Regelsysteme. Zu einem Defekt in einem Regelsystem, zählt nicht nur die Erkrankungen der Regelstrecke, sondern auch Defizite der Mediatoren (oder Stellglieder), wie Hormone, Enzyme usw. sowie von Substanzen, die an deren Aufbau beteiligt sind. Auch die Blockierung von Elektrolyten durch pathogene Schwermetallbelastung gehören dazu. Die Folge dieser Defizite ist meist eine aperiodische Entartung des Systems (siehe Abb. 24). Anderseits führt die Vorbelastung der Regelsysteme durch Fremdenergie zur einer Übersteuerung, die zur Labilität der Eigenschwingvorgänge führt. Dabei kommt es kommt es nicht so sehr auf die Stärke der einwirkenden Fremdenergie an, sondern auf die Dauer der Wirkung. Das typische Beispiel für solch eine Störung ist das Adaptationssyndrom, bei der durch das Fehlen von Erholungsphasen ein Erschöpfungszustand erreicht wird. In diesem Zusammenhang muss auf die Rolle der Vernetzung der Regelsysteme bei der Reizbeantwortung hingewiesen werden, die vor allem bei der Beurteilung von Regulationstests zum Tragen kommt: Eine in das System eingegebene Störgröße pathogener Funktionen kann an verschiedenen Ausgängen völlig verschiedene Eigenschwingvorgänge auslösen. So können bei einer Dysregulation die Elektrolyte aperiodische Entartungen zeigen, während die Gefäßwände oder die Leukozytenreaktion zwar ein entartetes, aber doch periodisch labiles Verhalten zeigen. In jedem Fall stört eine Störung von einem oder mehreren Regelsystemen die Homöostase und die Ökonomie eines Organismus. Arbeitet ein Organismus 35 teilweise unökonomisch, d.h. verbraucht er zu viel Energie, erreicht er sein Leistungslimit zu früh. Die dadurch hervorgerufenen metabolischen Störungen begünstigen dabei degenerative Veränderungen in jenen Organen, von denen erhöhte Arbeitsleistung gefordert wird. Dank der Erkenntnisse über die grundlegenden Wirkungsmechanismen von Regelsystemen eines Organismus und den Fortschritten in der Messtechnik, konnten im Laufe der Zeit Diagnose und Therapieverfahren entwickelt werden, mit deren Hilfe die Funktion entarteter Regelsysteme innerhalb eines Organismus erkannt und behoben werden können. Wie jedes andere energetisch offene System ist auch der menschliche Organismus in allen seinen Substanzsystemen schwingungsfähig. Es gibt daher keine starr eingestellten Sollwerte, sondern diese schwingen in einer bestimmten Bandbreite. Aufgrund der komplexen Vernetzung der Subsysteme können durch ein und denselben Reiz an verschiedenen Ausgängen Signale mit verschiedenen Amplituden und Frequenzen generiert oder auch gefiltert werden. Die Ausgangsgröße eines Subsystems kann andere Systeme beeinflussen und ihnen als Führungsgröße oder auch als Störgröße dienen. Die Möglichkeit der Messung der energetischen Vorgänge innerhalb eines Organismus bekommt eine immense Bedeutung, weil dadurch Reaktionsabläufe als Funktionsdiagnose untersucht werden können. Fehlerregulationen können so schon im Beginn ihres Entstehens erkannt werden. Entsprechend können dann noch vor dem Auftreten von z.B. Gewebsveränderungen therapeutische Maßnahmen unternommen werden. 2.3.1. Der Organismus als energetisch offenes System mit endogener Rhythmik Alle biologischen Systeme, vom Einzeller bis zum hochorganisierten Körper des Menschen, sind energetisch offene Systeme und daher darauf angewiesen, mit ihrer Umgebung Materie und Energie auszutauschen. Solche offenen Systeme zeigen keinerlei Linearität und die einzelnen wirksamen Regelkreise sind hochvernetzt und unterliegen einem biologischen Fließgleichgewicht. Eine wichtige Voraussetzung für das Funktionieren solcher Systeme sind Rückkopplungsprozesse die bedingen, dass Wechselwirkungen mit der Umgebung nicht einfach addiert werden können, sondern dass das Ganze stets mehr als seine Einzelteile ist (ein funktionierender Organismus ist mehr als die Summe seine Zellen). Rückkopplungen treten in Organismen vielfältig auf (z.B. laterale Hemmung beim Sehvorgang) und haben den Vorteil, dass die Funktionszustände eines Systems nicht fest fixiert sind, sondern als Ausgangspunkt immer neuer Entwicklungen werden können. Dabei verändern sich die Anfangs- und Randbedingungen ständig, wobei Auslese- und Optimierungsprozesse eine Selbstorganisation ermöglichen, die das energetisch effizienteste funktionieren des Organismus garantiert. Die dabei kurzzeitig auftretenden Ordnungszustände sind jedoch nicht stabil, sondern schwingen fernab von einem thermischen Gleichgewicht, d.h. die Zufuhr von geeigneter Energie (z.B. Nahrungsstoffe) bereitet sich im Organismus schneller aus, als es durch enzymatisch gesteuerte Reaktionen zu erwarten wäre. Trotzdem ist die Regelung der Homöostase eines Organismus unter physiologischen Bedingungen so stabil, dass trotz verschiedenster äußerer und innerer Einwirkungen der Normalzustand meist sehr schnell wieder hergestellt werden kann. 2.3.1.1. Chronobiologische Rhythmen Die regulatorische Selbstorganisation von Organismen liegt in erster Linie an periodisch auftretenden Prozessen in biologischen Systemen, die vor allem der 36 zeitlichen und räumlichen Organisation bestimmter Lebensvorgängen dienen. Diese endogenen Oszillatoren ermöglichen eine zeitlich und räumlich getrennte Organisation von biologischen Abläufen, wodurch besonders die zeitliche Koordination verschiedener inkompatibler biologischer Prozesse gewährleistet wird. Taktgeber sind intrazelluläre biochemische Prozesse wie die rhythmische Synthese von ATP durch die Mitochondrien einer Zelle. Extrazellulär sind es die thermischen Schwingungen von Molekülen der Grundsubstanz oder Extrazellulären Matrix. Ein zellmembrangebundener Taktgeber ist das Sympathikus-assoziierte zyklische Adenosinmonophosphat (cAMP), das eine Zirkardianrhythmik (24 Stunden) aufweist. Die Rhythmen auf molekularer Ebene gehen schließlich in große übergeordnete Rhythmen mit Sekunden- und Minutendauer, tages- und jahreszeitlicher Dauer über. Nahezu alle menschlichen Körperfunktionen unterliegen dem Einfluss der zirkadianen Uhr. Der tageszeitlich abhängige Wechsel von Schlafen und Wachen, Körpertemperatur sowie geistiger und körperlicher Leistungsfähigkeit sind jedem bekannt. Der Taktgeber oder Schrittmacher der menschlichen zirkadianen Uhr sitzt im Zentralnervensystem (ZNS) und zwar an der Hirnbasis direkt über der Sehnervkreuzung im SCN (= Suprachiasmatischer Nucleus). Die innere Uhr des Menschen tickt angeborenermaßen mit einer Periodenlänge von durchschnittlich 25 Stunden und wird durch äußere Signale, sogenannte Zeitgeber (Hell- Dunkel- Wechsel), jeden Tag synchronisiert. Alle Lebewesen der unterschiedlichsten Organisationsstufen besitzen viele Oszillatoren, die erst gemeinsam eine innere Referenzzeit bestimmen. Nicht nur die bilateralsymetrisch angeordneten "Hauptuhren" des SCN, sondern diesen untergeordnete "Nebenoszillatoren" bestimmen das endogene Rhythmusgefüge eines Organismus. Die Rolle des äußeren Zeitgebers ist dabei, diese verschiedenen Oszillatoren in einen gemeinsamen Takt zu zwingen. Fehlen diese externen Signale oder können sie nicht wahrgenommen werden, kann dieses Oszillatorengefüge intern desynchronisiert werden, d.h. in viele einzelne Komponenten mit eigenen Rhythmen zerfallen. Eines der Beispiele, welche Auswirkung das Fehlen bzw. das zu schwache Signal eines äußeren Zeitgebers auf das Funktionieren eines Organismus haben kann ist die Winterdepression oder die Seasonal Affective Disorder (SAD) des Menschen. Auch der sogenannte Jet-lag, der bei Reisen über Zeitzonen auftritt, hat seine Ursache in einer desynchronisation des inneren Zeitgefüges. Bei allen Wirbeltieren ist der Botenstoff Melatonin für die innere synchronisation der Zeit verantwortlich. Melatonin ist normalerweise in der Nacht hoch konzentriert und sinkt unter Lichteinwirkung am Tag ab. Es wird hauptsächlich von der Epiphyse (Pinealorgan) synthetisiert. Das Pinealorgan liegt auf dem Dach des Zwischenhirns unter dem Großhirn und ist z.B. bei Vögeln, Fischen und Amphibien wie ein drittes Auge organisiert, d.h. es kann noch direkt Lichtreize von außen wahrnehmen und damit als Eingang für Zeitgeberreize der inneren Uhr verstanden werden. Beim Menschen ist es nur noch Umschaltstation für über das Auge eintreffende Lichtreize, die als Zeitgeber dienen und Produktionsstätte des Melatonins. Durch Krankheit kann dieses innere rhythmische Ordnungsgefüge gestört werden, anderseits kann ein Störung dieses inneren Ordnungsgefüges durch äußere Einflüsse zur Krankheit führen. Im Falle einer überwundenen Krankheit bedeutet dies jedoch nicht, dass das System zum Ausgangspunkt zurückkehrt. Vielmehr bleibt eine Art "Erinnerung" zurück (z.B. zelluläre immunologisch oder veränderte Grundsubstanz wie Narben), die das weitere rhythmische Verhalten der Grundregulation beeinflusst. Zusammenfassend kann festgestellt werden, dass die zeitliche und räumliche Organisation eines Organismus sowie dessen Leistungsfähigkeit von seinem endogenen Biorhythmus bestimmt wird. Über diese endogenen Oszillator steht der 37 Organismus mit seiner Umwelt in Resonanz. Daher sind auch Resonanzen mit technischen Frequenzen möglich. Zum Beispiel sind die Schumannwellen und das Erdmagnetfeld bioenergetische Ordnungsparameter mit Einfluss auf das psychische Verhalten sowie der raum-zeitlichen Ordnung eines Organismus. Die Selbstheilungskräfte eines Organismus sind von der rhythmischen Ordnung vegetativer Funktionen abhängig. 2.3.2. Rolle der Extrazellularmatrix oder der Grundsubstanz (nach HEINE und PISCHINGER) Um die Funktionsweise des Organismus zu verstehen der nach den Prinzipien eines kybernetisch offenen Systems arbeitet ist es notwendig, neben den einzelnen Zellen und Organen, die Arbeitsweise der Grundsubstanz oder der Extrazellulären Matrix zu erläutern. Das Leben eines höheren mehrzelligen Organismus ist an die Grundsubstanz oder Extrazellularmatrix gekoppelt. Die der Zelle oder des Organs als "Molekularsieb" vorgeschaltete Grundsubstanz ist als integraler Teil des energetisch offenen Systems Organismus von übergeordneter Bedeutung. Die normale als auch die pathologische Reaktionsfähigkeit des genetischen Materials in einem Zellkern ist von der Funktionsfähigkeit des Grundsubstanz abhängig. Entsprechend durchzieht die Grundsubstanz die Extrazellulärräume des gesamten Organismus, erreicht jede Zelle und registriert stets die Kommunikation zwischen Zellen bzw. Geweben. 2.3.2.1. Morphologie der Grundsubstanz Die unterschiedliche Struktur der Zwischenzellsubstanz oder Extrazellularmatrix (Grundsubstanz) macht den Bindegewebsraum einerseits zur Transitstrecke zwischen Blutgefäßen und den Organen, anderseits zum eigentlichen Bindegewebe. Die Extrazellularmatrix besteht im wesentlichen aus intersitieller Flüssigkeit, Proteoglykanen und Glykoproteinen. Die Proteoglykane verleihen der intersistellen Flüssigkeit je nach Aufkommenvisköse bis feste Eigenschaften. Aufgrund ihrer Wasserbindungseigenschaften sind sie z.B. für die Formbeständigkeit des Gelenkknorpels und die Durchsichtigkeit der Cornea verantwortlich. Die Glykoproteine bilden an der Zelloberfläche die Glykocalix und sind Bestandteile der Basalmembran. Sie haben teilweise mechanische Aufgaben innerhalb der Zell-ExtrazellularmatrixHaftung und bilden regulierende Stofftransportschranken zwischen interstitiellen Räumen und den anliegenden Zellen. Die eigentlichen Bindegewebsfasern unterteilt man im Wesentlichen in drei unterschiedliche Typen: die Kollagenfasern, die elastischen Fasern und die Retikulinoder Retikulumfasern. Kollagenfasern sind zugfest und entstehen unter Zugbeanspruchung (Sehnen und Bänder), Retikulinfasern sind biegungselastisch und bilden in Lymphknoten und in der Milz das Grundgerüst als räumliche Netze aus. Die elastischen Fasern (Blutgefäße) sind zugelastisch und können auf das 1,5fache reversibel gedehnt werden. 38 Abb.24 :Schema der Regulation innerhalb der Grundsubstanz (Pfeile). Die Funktion und der Differenzierungsgrad der Bindegewebszellen wird wesentlich durch die jeweils aktuelle Zusammensetzung der Grundsubstanz gesteuert. Dabei spielt das retikuläre Bindegewebe eine wichtige Rolle, da dort die meisten Aufbau- und Umbauprozesse ablaufen. Die jeweils spezifischen rückkoppelnden Informationen zwischen den einzelnen Komponenten der Grundsubstanz wie den Makrophagen / Monozyten, den Fibroblasten, Mastzellen und den Immunozyten werden über das Zytokinnetz weitergeleitet. Gleichzeitig treten enge Beziehungen zu vegetativen Nervenfasern auf. Aufgrund ihrer dynamischen Eigenschaften, was Aufnahme und Weiterleitung innerhalb der Gewebe eines Organismus angeht, stellt die Grundsubstanz den Spiegel des zeitlichen und räumlichen Verhaltens der Zelle (bzw. der Gewebe), insbesondere ihrer Genaktivierung dar. Sie ist sozusagen das metabolische Gedächtnis der Zelle. Dadurch, dass sie verschiedene Signalsubstanzen in Form komplexer räumlicher Muster speichern kann und diese an Zellen freigibt, ist die Grundsubstanz in der Lage Informationen wahrscheinlich als zukünftige Handlungsanweisungen weiterzugeben. 39 2.3.2.2 Energiefluss innerhalb der Grundsubstanz In einem Organismus dienen elektrische Impulse geringer Intensität der schnellen Informationsübertragung (Nerven, Muskeln). Diese Eigenschaft der Gewebe werden als bioelektrische Untersuchungsverfahren, wie EKG, EEG und EMG, in der klinischen und praktischen Medizin genutzt. Auf der anderen Seite wird aber bei der Anwendung von Therapieverfahren häufig nur der biochemischen Reaktion eines Organismus Rechnung getragen und zur Behandlung von Krankheiten die entsprechenden pharmakologischen Präparate eingesetzt. Die Tatsache, dass jede biochemische Reaktion in einem Organismus von einer biophysikalischen, in Form von elektromagnetischen Feldwechselwirkungen begleitet wird und umgekehrt, wird dabei außer acht gelassen. So wird z.B. die Synthesefähigkeit von Zellen (z.B. Fibroblasten), in vivo wie in vitro durch elektromagnetische Felder bzw. elektrische Reize angeregt. Menschliche Fibroblasten reagieren in vitro auf biphasische, sinusförmige elektromagnetische Felder (20 Hz bei 6 mT) innerhalb von 3 Wochen mit einer Ausdifferenzierung und einer Erhöhung ihrer Grundsubstanzsynthese. Der Organismus stellt also ein energetisch offenes hochvernetztes System dar, in dem die Informationsübertragung über biophysikalische und biochemische Prozesse gesteuert werden. Die Grundsubstanz stellt innerhalb des Organismus eine Art "Biosensornetzt" dar, das sich anhand seiner Eigenschaften am besten mit dem physikalischen Begriff der "Unschärfelogik" (fuzzy-logic) beschreiben lässt. Dieses arbeitet gegenüber der klassischen JA-NEIN-Logik mit unscharfen (fuzzy) WENNDANN-Regeln. Gegenüber "neuronalen Netzen", die schon beim Ausfall eines Elementes unbrauchbar werden, ist ein sogenanntes "fuzzy-neuronales Netz", wie die Grundsubstanz, völlig robust. Eine Beschädigung bewirkt hier keinen plötzlichen , sondern stufenweisen Ausfall von Komponenten, so dass durch Selbstorganisation oder therapeutischen Einfluss das System prinzipiell regeneriert werden kann. Komponenten der Grundsubstanz arbeiten nicht nach dem digitalen JA-NEIN-Prinzip vielmehr wie ein "Sieb", das verschiedene, wenig voneinander abweichende Stromstärken nacheinander durchlassen kann. Dabei kommt es nicht auf eine absolute Genauigkeit an, denn es geht in der Grundregulation nicht um die Einhaltung exakter Regeln, sondern um Mustererkennung. In einem "fuzzy-neuronalen Netz" führen Informationen (wie z. B. nur geringe Veränderungen des pH-Wertes oder der Temperatur) zu einer schnelleren Verarbeitung, als digitale JA-NEIN- Entscheidungen. Dadurch können alle Veränderungen in einem fuzzy-neuronalen Netz sofort reguliert werden. Störungen der Grundsubstanz und damit der Grundregulation führen zu einer schnellen elektromagnetischen Informationsverarbeitung über das fuzzy-neuronale Netz und es kommt zu einer entsprechenden Zellreaktion. Aus diesen Ansätzen heraus können neue Untersuchungs- und Therapieverfahren entwickelt werden, die funktionelle Störungen eines Organismus aufdecken und Regulationsvorgänge einleiten. Dermatome und Head-Zonen Hautareale (sowie die dazugehörigen Abschnitte der Grundsubstanz), die von dem gleichen Spinalnerven versorgt werden, werden als Dermatome bezeichnet. 40 Dabei wird jedes Hautareal einem Rückenmarkssegment mit den dazugehörigen Spinalnerven zugeordnet. Man unterscheidet nach den einzelnen zervikalen (Halswirbelsäule, C1-C7), den thorakalen (Brustwirbelsäule T1 - T12)), den lumbalen (Lendenwirbelsäule, L1 - L5), den sakralen (Kreuzbein) Spinalnervenwurzeln. Headschen Zonen beschreiben wiederum die Hautareale, in denen bei einer Erkrankung der inneren Organe eine Überempfindlichkeit für Schmerz- , Temperaturund Berührungsreize, oder eine gesteigerte Schmerzempfindlichkeit auftreten und die in ihrer Ausdehnung dem jeweiligen Dermatom entspricht. Abb.25: Headsche Zonen 41 So kann es zu einer gesteigerten Schmerzempfindlichkeit des 8ten Brustwirbeldermatoms bei Magenstörungen und des 11ten Brust- bis 1sten Lendenwirbeldermatoms bei Dickdarmproblemen kommen. Das Verständnis für den Zusammenhang von Head-Zonen und Funktionsstörungen der Organe kann auf rein biochemischen Weg nicht erklärt werden. Dazu ist es notwendig des elektrische Verhalten der beteiligten Abschnitte der Grundsubstanz zu verstehen: Während Arterien und Venen als Leiterbahnen den Blut-Zulauf und den Blut-Ablauf von Geweben bzw. Organen regulieren, kommt es erst im Bereich der Kapillaren zum Stoffaustausch. Dabei sind die treibenden Kräfte je nach Gewebe der aktive Transport, die Filtration, die Diffusion oder Differenzen im osmotischen Druck. Daran ist das Vascular-Interstitial Closed Electric Circuit System (VICC - System) mit selektivem, elektrisch gesteuertem Transport zwischen Blut und Grundsubstanz maßgeblich beteiligt (NORDENSTRÖM, 1985). Es stellt neben dem Blut- und Lymphkreislaufsystem ein zusätzliches Zirkulationssystem dar. Das VICC-System wird dann aktiviert, wenn lokale anabole (stoffwechselbedingter Aufbau) oder katabole (stoffwechselbedingter Abbau) Prozesse eine elektrochemische Polarisation des Gewebes verursachen. Durch Depolymerisation der Grundsubstanz in einem verletzten Gewebebezirk, z.B. einem Entzündungsherd, wird katabole Energie freigesetzt, die als elektrisches Potential im Verhältnis zum umgebenden Gewebe gemessen werden kann. Diese messbaren Potentialdifferenzen schwanken von positiven über Null zu negativen Werten gegenüber dem gesunden Gewebe. Zusätzlich treten unregelmäßige Oszillationen auf, die bei der Interpretation der Prozesse mitberücksichtigt werden müssen. So zeigen nekrotische Gewebsveränderungen zeitabhängige Phasen in Größe und Polarität der Potentialveränderungen und sichern dadurch verletzten Gewebe durch Zu- und Abstrom die notwendige Stoffwechselversorgung. Durch den Zusammenhang zwischen den involvierten Strukturen innerhalb der Grundsubstanz könne diese, z.B. durch Entzündungen hervorgerufene elektrische Potentialdifferenzen von Geweben über die jeweiligen Head-Zonen festgestellt und mit entsprechenden elektromagnetischen Frequenzen ausgeglichen werden. Akupunkturpunkte und Meridiane Morphologisch lässt sich der Akupunkturpunkt, wobei der Begriff: "Punkt" eigentlich nicht korrekt ist, immer als ein perforierendes Gefäß-Nervenbündel darstellen, das in eine Scheide aus lockerem Bindegewebe eingehüllt ist. Bei rund 82% der klassischen 361 Akupunkturpunkte kann die Perforation der oberflächlichen Körperfaszie festgestellt werden. Dementsprechend müsste man korrekterweise nicht von einem "Punkt", sondern von einem "Loch" sprechen. Der Durchmesser der Akupunkturpunkte variiert von 2 mm bis 8 mm. 42 Abb.26.: Schema eines Akupunkturpunktes (1 Körperfazie, 2 große Vene, 3 kleine Vene, 4 kleine Aterie, 5 Lymphgefäß, 6 Nerv, 7 Vater-Pacinische lamellenkörperchen, 8 Meissnersche Tastkörperchen, 9 Epidermis, 10 subepitheliales Bindegewebe) Zwischen einem Akupunkturpunkt und seiner Umgebung lassen sich mit unterschiedlichen elektrischen Messverfahren Potentialdifferenzen sowie eine erhöhte Leitfähigkeit am Akupunkturpunkt selbst nachweisen. Im Akupunkturpunkt werden Zug-, Druck- und Perfusionsänderungen lokaler und systemischer Art registriert und autonom über Axonreflexe, bei stärkeren Reizen über Zuschaltung übergeordneter Zentren reguliert. Die klassischen Akupunkturpunkte sind entlang einer kinetischen Muskelfaszie (fasziomyo-tendinöser Kette) angeordnet, die mit dem Verlauf der Meridianen identisch sind. 43 Abb.27.: Akupunkturpunkte des Lungenmeridians (Lu 1 - 11). Die Akupunkturpunkte liegen entlang einer faszio-myo-tendinösen Kette. Aufgrund ihrer morphologischen Besonderheiten stellen die Akupunkturpunkte eine Art "Fenster" zur Grundsubstanz dar, worüber regulatorische Vorgänge über Reizung (Nadeln = Schmerzreiz) der Punkte angeregt werden können. 44 3. Physikalische Grundlagen der Bioresonanztherapie Innerhalb des großen Frequenzbereiches der elektromagnetischen Strahlung bestehen enorme Unterschiede hinsichtlich des physikalischen Verhaltens der Strahlung als auch auf ihre mögliche Wirkung auf Lebewesen. Die pauschalisierte Aussage: "Alles was natürlich ist, ist dem Menschen zuträglich, alles was künstlich erzeugt wird, schadet ihm", ist so nicht zutreffend und kann in dieser vereinfachten Form nicht getroffen werden. So kann zum Beispiel die natürliche Sonnenstrahlung, ohne die das Leben auf der Erde nicht möglich wäre, bei längerem ungeschütztem Aufenthalt, für den Menschen zu akuten (Sonnenbrand) als auch zu später auftretenden gesundheitlichen Schäden (Hautkrebs) führen. Auf der anderen Seite werden seit Jahren die Folgen des sogenannten "Elektrosmogs" diskutiert, dem der Mensch in unserer hochtechnisierten Gesellschaft ausgesetzt ist. Mit dem Begriff "Elektrosmog" werden alle künstlich hergestellten elektromagnetischen Felder zusammengefasst und als Gesundheitsschädlich eingestuft ohne zu berücksichtigen, dass es immense Unterschiede in ihrer Wirkungsweise auf Organismen in Abhängigkeit von der Frequenz und der Intensität des Feldes gibt. Es ist einleuchtend, dass elektromagnetische Felder einen Organismus beeinflussen können, gerade im Hinblick darauf, dass viele funktionelle Vorgänge in einem Körper über Ionenverschiebungen an Membranen (Nervenimpulse, Muskelkontraktion, Herzschlag) gesteuert werden. Betrachtet man einen Organismus als offenes System, bei dem die Aufrechterhaltung der Homöostase ein essentieller Bestandteil ist, so muss die Wirkung von elektromagnetischer Strahlung auf die Gesundheit berücksichtigt werden. Gleichwohl kann der gezielte Einsatz von dem Körper "dienlichen" elektromagnetischen Felder helfen, die Kommunikation der Gewebe innerhalb des Organismus zu verbessern und damit die notwendigen funktionellen Einheiten zu regulieren. Aufgrund dieser Vielzahl von Wechselwirkungen zwischen elektromagnetischer Strahlung und Organismen und der Frequenzabhängigkeit dieser Wechselwirkungen können keine allgemeingültige Schlussfolgerungen in Bezug auf Schädigung oder Nutzen gezogen werden. Betrachtet man die Wirkungen elektromagnetischer Strahlung auf den Körper, ergeben sich drei wichtige Fragen: - Gibt es Schwellenwerte für die Expositionsbedingungen, ab denen biologische Wirkungen einsetzten? - Können irreversible Veränderungen auftreten, die sich im Laufe der Zeit zu schwerwiegenden Wirkungen aufsummieren können oder tritt eine Wirkung nur auf solange man der Strahlung ausgesetzt ist? - Ist die Strahleneinwirkung von der Dosis, d.h. vom Produkt aus Strahlungsstärke und Einwirkungsdauer abhängig? Die Antworten auf diese Fragen fällt für die verschiedenen Frequenzbereiche unterschiedlich aus. Teilt man den Frequenzbereich hinsichtlich der biologischen Wirkung auf, so ergeben sich zwei Hauptbereiche: die ionisierende und die nichtionisierende Strahlung. Bei der energiereichen ionisierten Strahlung besitzt bereits die kleinstmögliche Strahlungsmenge, ein Strahlungsquant, genug Energie, um den chemischen Zustand eines Moleküls zu verändern. Die ionisierte Strahlung schließt die UV-Strahlung, die weiche und harte Röntgenstrahlung, die radioaktive Gamma-Strahlung sowie die radioaktive Höhenstrahlung mit ein. Bei der nichtionisierten Strahlung besitzen die Strahlungsquanten weniger Energie, so dass ihre biologische Wirkungen davon abhängen, wie viel Strahlungsquanten pro Fläche gleichzeitig auftreten. Je nach Strahlungsintensität sind stark schädigende Wirkungen, wie z.B. im Nahbereich einer Radaranlage möglich. 45 Um eine elektromagnetische Strahlung positive oder negative Auswirkungen hat, hängt also von ihrem Feldverhalten ab. Die Möglichkeiten elektromagnetische Felder im Bereich der Bioresonanztherapieverfahren liegen eher in ihrer Eigenschaft als Informationsübermittler. Um jedoch die verschiedenen Frequenzspektren, die bei der Bioresonanztherapie zum Tragen kommen zu erläutern ist es zunächst notwendig, die physikalischen Grundlagen von elektromagnetischen Feldern zu verstehen. 3.1. Elektromagnetische Felder 3.1.1. Allgemeine physikalische Grundlagen der Wellenausbreitung Bei der Ausbreitung einer Welle in einem materiellen Medium schwingen die Teilchen des Mediums um ihre Gleichgewichtslagen. Die Energie der Welle wird weitertransportiert, nicht jedoch das Medium selbst. Der Wellenausbreitung können zwei Typen zugeordnet werden: transversale und longitudinale Ausbreitung. Bei einer transversalen Welle schwingen die Teilchen des Mediums senkrecht zur Ausbreitungsrichtung der Wellenenergie. Bei einer longitudinalen Welle schwingen die Teilchen des Mediums parallel oder antiparallel zur Ausbreitungsrichtung der Welle. Die Periode eines schwingenden Teilchens ist die Zeit, die für eine Schwingung benötigt wird. Die Frequenz (gemessen in Hz = Hertz) ist die Zahl der Schwingungen pro Sekunde (Frequenz = 1/Periode). Die Wellenlänge (gemessen in m) ist der kürzeste Abstand zwischen zwei schwingenden Teilchen, die die gleiche Phase besitzen. Die Amplitude ist die maximale Auslenkung des schwingenden Teilchens aus seiner Ruhelage. Frequenzen: 1 Hz = eine Schwingung pro Sekunde 1 kHz = 10³ oder 1000 Schwingungen pro Sekunde 1 MHz = 106 oder 1 Million Schwingungen pro Sekunde 1 GHz = 109 Schwingungen pro Sekunde Geschwindigkeit einer Welle = Frequenz x Wellenlänge c Q = x O Die Ausbreitungsgeschwindigkeit c einer elektromagnetischen Welle beträgt (nach der Maxwellschen Theorie) im Vakuum ~ 300 000 km/sec (= Lichtgeschwindigkeit). Dabei ist das Verhältnis zwischen Wellenlänge O und Frequenz Q einer Strahlung die sich im Vakuum ausbreitet O = c/Q wobei c die Lichtgeschwindigkeit ist. 46 Beispiel: Ein Mittelwellensender mit einer Frequenz von 800 kHz sendet Wellen mit einer Länge von O 3.1.1.1. = 300 000 km : 800 000 = 0,375 km = 375 m aus. Einheiten Aufgrund ihrer physikalischen Eigenschaften unterscheidet man elektromagnetische Felder in niederfrequente elektromagnetische Wechselfelder und elektromagnetische Hochfrequenzstrahlung. Elektrische Felder bewegen elektrische Ladung, bewegte elektrische Ladung wiederum verursachen Magnetfelder, Änderungen magnetischer Felder induzieren ihrerseits elektrische (Wirbel-)Felder. Die Frequenzbereiche von niederfrequenten Feldern liegen zwischen 3 und 30 000 Hz. Diese Felder haben einen elektrischen und einen magnetischen Anteil, die getrennt voneinander betrachtet werden können. Diese Felder sind die Quelle ihrer Entstehung gebunden und ihre Feldstärke nimmt mit zunehmender Entfernung von der Quelle ab. Elektrische Wechselfelder : Stromdichte in A/m² Feldstärke in V/m Magnetische Wechselfelder: Feldstärke in T (Tesla) magnetische Induktion B Die Frequenzbereiche elektromagnetischer Hochfrequenzstrahlung liegen zwischen 30 kHz bis 300 GHz. Aufgrund der höheren Frequenz, d.h. der schnelleren Feldänderung, sind die magnetische und die elektrische Komponente nicht mehr zu trennen, da sie sich gegenseitig bedingen. Außerdem löst sich elektromagnetische Strahlung von einer Antenne ab und breitet sich in Abhängigkeit von der Frequenz im Raum aus. Die bei der elektromagnetischen Strahlung pro Flächeneinheit wirkende Strahlungsleistung ist die Strahlungsintensität. Elektromagnetische HF-Strahlung: Strahlungsintensität in W/m² (Leistungsflussdichte) Die spezifische Absorptionsrate SAR beschreibt die in Wärme umgewandelte Strahlungsleistung bezogen auf die Körpermasse: SAR : spezifische Absorptionsrate in W/kg 3.1.1.1 Resonanz Jedes schwingungsfähige Gebilde ob Molekül, Organ, Gebäude usw. besitzt mindestens eine Eigenfrequenz. D.h. erhält ein schwingungsfähiger Körper einen Anstoß, so kommt stets eine freie Schwingung der Eigenfrequenz f0 zustande. Regt man ein System mit der Eigenfrequenz f0 mit einer Zwangsfrequenz f1 an, bei der die Zwangsfrequenz f1 mit dem Wert der Eigenfrequenz f0 übereinstimmt, so wächst die Amplitude der erzwungenen Schwingung an. Die erzwungene Schwingung f1 tritt in Resonanz mit der Eigenschwingung f0. 47 Ein Beispiel für die Auswirkung bei der Entstehung von Resonanzfrequenzen ist, dass beim Überqueren einer Brücke nicht im Gleichschritt marschiert werden darf, da die Anregung der Eigenfrequenz zu einer Resonanz führen kann und die Brücke dabei einstürzen könnte. Eine bestimmte Tonlage kann ein Glas zum Platzen bringen, wenn die Tonlage mit der Eigenfrequenz des Glases zusammentrifft. 3.1.1.2 Schwingungs- und Modulationsformen Elektromagnetische Felder können in verschiedenen Formen appliziert werden: Sinusförmig, rechteckförmig und als Sägezahn. Die meisten technisch erzeugten Felder, wie die Frequenzen im Rundfunkbereich und die Wechselspannung der Stromversorgung sind sinusförmige Welle. Abb.28: Spektren verschiedener Schwingungsformen. Sinusform mit unterschiedlicher Amplitude (links) Rechteckform (a) und Sägezahnform (b) (beide rechts) Elektromagnetische Wellen können Informationen über weite Strecken transportieren. Bei Radio - und Fernsehsendern wird die akustische Frequenz (von 16 Hz bis 15 kHz) nicht direkt übertragen, da dabei der erreichbare Wirkungsgrad zu gering wäre. Das Audio- bzw. Videosignal wird einer Trägerwelle aufmoduliert. Die Frequenzen der Trägerwelle liegen im Rundfunkbereich zwischen 30 kHz und 300 MHz und beim Fernsehen zwischen 50 MHz und 1 GHz. Bei der Amplitudenmodulation (AM) schwankt die Trägerwelle im Takt der niederfrequenten Schwingung, wohingegen bei der Frequenzmodulation (FM) die Frequenz der Schwingung im niederfrequenten Takt mitschwingt. 48 Abb.29.: Amplituden(AM) - und Frequenzmodulation (FM) Radiosendungen im Mittel- und Langwellenbereich sind amplitudenmoduliert, während Sendungen im Kurz- und Ultrakurzwellenbereich sowie Übertragungen im Fernsehbereich frequenzmoduliert ausgestrahlt werden. Bei der Übertragung von Information im Mobilfunkbereich wird eine Pulsmodulation angewendet. So arbeitet z.B. das D-Netz der Telekom im 900 MHz Frequenzbereich bei einer Pulsmodulation von 214 Hz. 3.1.2 Niederfrequente elektrische Wechselfelder Als "niederfrequent" werden elektrische Wechselfelder mit einer Frequenz bis zu 30 kHz bezeichnet. Die Stromversorgung liefert die elektrische Energie mit Wechselspannung, d.h. mit elektrischem Strom, der seine Stärke und seine Richtung zeitlich verändert. Dabei beträgt die sinusförmige Frequenz des Versorgungsnetzes in Deutschland 50 Hz (in der USA z.B. 60 Hz). Der Niederfrequenzbereich wird in die folgenden Teilbereiche untergliedert: ULF - Bereich (ultra low frequency) : bis zu 3 Hz (Wellenlänge bis 100 000 km) ELF - Bereich (extremly low frequency) : 3 Hz bis 3000 Hz (Wellenlänge von 100 000 km bis 100 km) VLF - Bereich (very low frequency) : 3000 Hz bis 30 000 Hz (30 kHz) (Wellenlänge von 100 km bis 10 km) Es besteht zwischen zwei Punkten ein elektrisches Feld wenn diese gegeneinander eine Spannung aufweisen. Dabei ergibt sich die elektrische Feldstärke eines niederfrequenten Wechselfeldes aus der Spannung pro Abstand und hat die Einheit Volt per Meter. Im Gegensatz zu den magnetischen Feldern sind elektrische Wechselfelder nicht an den Verbrauch elektrischer Energie gebunden, sie treten bereits auf, wenn diese lediglich bereitgestellt wird (z.B. Steckdose). Im Gegensatz zum hochfrequenten Bereich, wo sich elektromagnetische Wellen von Antennen ablösen und sich in den Raum ausbreiten, tritt dies bei niederfrequenten Feldern nicht auf: Die Felder sind an ihre Quelle gebunden, die Feldstärke nimmt mit 49 zunehmender Entfernung ab. Man spricht daher in diesem Frequenzbereich nicht von Strahlung, sondern von Feldern. Die biologischen Wirkungen von niederfrequenten elektromagnetischen Feldern kommen in erster Linie durch die auftretenden magnetischen Wechselfelder zustande. Diese werden durch bewegte elektrische Ladung (elektrische Ströme) verursacht. Dabei ist die magnetische Feldstärke H um so stärker, je stärker der Strom und je geringer die Entfernung zum fließenden Strom ist. Die magnetische Flussdichte oder magnetische Induktion B ist proportional der magnetischen Feldstärke. Das Produkt aus magnetischer Flussdichte und der, durch eine Fläche hindurchtretenden magnetischen Feldlinien, wird als magnetischer Fluss bezeichnet. Durch die zeitliche Änderung des magnetischen Flusses bei niederfrequenten magnetischen Wechselfeldern können im Inneren eines Organismus elektrische Ströme (Wirbelströme) induziert werden. Die Beträge von technisch erzeugten magnetischen Wechselfeldern überschreitet vielfach die Werte der durch natürliche elektrische Entladungserscheinungen (Ionosphärenströme oder Sferics) induzierten Felder. Die biologische Relevanz zur Kontrolle der Stärke von niederfrequenten magnetischen Wechselfeldern besteht darin, dass diese Felder im Körperinneren nahezu gleich stark sind wie außerhalb des Organismus. Deutliche biologische Effekte der durch magnetische Wechselfelder induzierte Wirbelströme sind auftretende Seheindrücke (z.B. Flimmern oder Leuchterscheinungen), sogenannte Magnetophosphene. 3.1.2.1. Sferics - natürliche elektrische Wechselfelder Die wichtigsten felderzeugenden Vorgänge in der Natur sind Blitzentladungen. Jeder auf der Welt erzeugte Blitz verursacht nicht nur in seiner unmittelbaren Umgebung kurzzeitig hohe Feldstärken, (z.B. noch in 300 m Entfernung über 100 kV/m) sondern trägt auch weltweit zum Auftreten der natürlichen elektrischen Wechselfelder, der sogenannten Sferics, bei. Sferics besitzen einen charakteristischen zeitlichen Verlauf, der durch ein höherfrequentes Anschwingen mit einem nachfolgendem langwelligen Ausklingen gekennzeichnet ist. Die durch Blitzentladungen entstandenen elektromagnetischen Schwingungen müssen sich in den Raum ausbreiten, der durch die leitende Erdoberfläche und die leitfähige Atmosphärenschicht, die Ionosphäre, begrenzt wird. Bei der Ausbreitung werden jene Wellen bevorzugt, die nach einem Erdumlauf wieder so an ihrem Entstehungsort ankommen, dass sie sich verstärken. Man kann daher den Zwischenraum Erdoberfläche - Ionosphäre als Resonanzraum ansehen, der durch den Blitz zur Schwingung wird und worin sich Resonanzschwingungen ausbilden. Entsprechend dieser Überlegung entspricht die Grundschwingung jener Wellenlänge, die mit der Länge des Resonanzraumes, also etwa dem Erdumfang übereinstimmt. Setzt man in die Gleichung c : O = Q c = 300 000 km/s (Lichtgeschwindigkeit) O = 40 075 km (Erdumfang) so erhält man tatsächlich aus Ausbreitungsgeschwindigkeit und Erdumfang die festgestellte Grundschwingung von etwa 7,5 Hz. Nach ihrem Entdecker W.O. Schumann, werden diese Resonanzschwingungen auch Schumann-Resonanzen genannt. 50 Die Feldstärken der Schumann-Resonanzschwingungen sind niedrig und liegen z.B. bei 10 Hz bei ca. 3 mV/m. Abb.30.: Frequenzverteilung der Schumann-Resonanzschwingungen Die Grundfrequenz der Schumann-Resonanzschwingung liegt im Frequenzbereich der menschlichen Hirnpotentiale und ihr wird innerhalb der Bioresonanztherapieverfahren eine regulierende Wirkung zugeschrieben. 3.1.2.2. Bedeutung von niederfrequenten elektrischen Wechselfeldern in der Bioresonanztherapie Wie in Kapitel 2.2.4. beschrieben ist die Reaktivität biologischer Membranen von dem Aufrechterhalten des Ruhepotentials abhängig. Gerade bei Geweben, deren Funktion von ihrer Erregbarkeit abhängen, wie Nerven- und Muskelgewebe, kann eine Verschiebung des Ruhepotentials in Richtung Hyperpolarisation oder Depolarisation funktionelle Auswirkungen haben. Die Stromdichten, die bei der Erregung einer Zelle auftreten (z.B. Aktionspotential der Nervenzelle) betragen etwa 100 µA/cm², bei einer Frequenz von ~ 10 Hz. Mit einer Stromdichte von nur etwa 0,75 µA/cm² können "lokale Antworten" der Membran ausgelöst werden. Bei diesen schwachen Stromdichten wird die Erregungsschwelle einer Membran zwar nicht überschritten, jedoch kann die Auslenkung des Membranpotentials hin zu positiveren Werten verstärkt werden, was wiederum die Erregbarkeit der Membran erhöhen kann. Die Steigerung der Erregbarkeit ist der einwirkenden Stromdichte direkt proportional. Dabei können selbst kleinste Stromdichten noch zu Verschiebungen des Membranruhepotentials führen, deren Ausmaß von der Richtung und der Frequenz des applizierten Feldes abhängt. Ihre biologische Bedeutung hängt davon ab, wie groß diese Veränderungen im Vergleich zu den körpereigenen physiologischen Schwankungen ist. Sind jedoch die physiologischen Zustände aufgrund von funktionellen Störungen instabil, ist z.B. die Verfügbarkeit von Kalzium das für die Erregung von Muskelzellen notwenig ist, herabgesetzt, so kann schon ein sehr schwaches niederfrequentes elektrisches Wechselfeld regulierend eingreifen. Biologische Feldwirkungen im niederfrequenten Bereich sind stark Frequenzabhängig. Es kann zu einer "Informationsübertragung" auf grund einer spezifischen 51 Feldeinwirkung kommen. Allerdings ist der Informationsgehalt von Signalen nur dann groß, wenn sich ihr Wert ständig auf unvorhersagbare, also eher zufällige Weise verändert. So ist der Informationsgehalt einer harmonischen Schwingung, bei der ja bereits nach einer Periodendauer nichts Neues mehr geschieht, daher klein. Auch wenn der Informationsgehalt eines niederfrequenten Wechselfeldes, das in einer harmonischen Schwingung appliziert wird relativ gering ist, so können aber Resonanzeffekte von Bedeutung sein. Ähnlich wie beim Anstoßen einer Schaukel durch die zeitgerechte Wirkung kleiner Energie, kann es durch Aufsummieren im Organismus zu einem Effekt kommen. Gerade die sogenannten Schrittmacherzellen in einem Organismus, die in der Lage sind, sich periodisch immer wieder selbst zu erregen. Derartige Zellen wirken z.B. als Taktgeber für den Herzschlag (siehe Kapitel 2.2.3.2.). Die Reaktion dieser Zellen auf eine Stimulationsfrequenz ist besonders stark, je ähnlicher diese Frequenz der Eigenerregungsfrequenz ist. Dabei liegen die dazu erforderlichen Stromdichten bei nur 1 µA/cm². Je weiter jedoch die Stimulationsfrequenz von der Eigenerregungsfrequenz entfernt ist, um so höhere Stromdichten sind erforderlich. 3.1.3. Niederfrequente magnetische Wechselfelder Magnetische Wechselfelder werden durch bewegte elektrische Ladungen, also elektrische Ströme verursacht. Dabei ist die magnetische Feldstärke um so stärker, je stärker der Strom und je geringer die Entfernung zum fließenden Strom ist. Die magnetische Induktion (magnetischer Flussdichte) B ist proportional der magnetischen Feldstärke. Unter magnetischem Fluss wird die Anzahl der durch eine Fläche hindurchtretenden magnetischen Feldlinien verstanden. Er kann im einfachsten Fall als Produkt aus magnetischer Flussdichte (Induktion) und der Fläche berechnet werden. Er kann sich daher nicht nur mit dem Magnetfeld ändern, sondern auch, wenn sich die Größe der Fläche oder deren Lage relativ zum Feld ändern. Durch die zeitliche Änderung des magnetischen Flusses werden in einem Organismus elektrische Ströme induziert, die in geschlossenen Strombahnen verlaufen und als Wirbelströme bezeichnet werden. Die Wirbelströme sind umso stärker, je rascher die zeitliche Flussänderung erfolgt, d.h. sie nehmen mit steigender Frequenz zu. Darüber hinaus sind sie auch vom Verlauf der zeitlichen Änderung abhängig: Rechteckförmige Schwingungen induzieren auf grund der steileren Flanken höhere Stromspitzen als sinusförmige. Alle ermittelten Effekte von niederfrequenten magnetischen Wechselfeldern weisen darauf hin, dass ein Organismus mit einer Induktion von über 5 mT beeinflusst werden kann. Im Vergleich dazu: Die Feldstärke des Erdmagnetfeldes beträgt nur 0,05 mT. Magnetische Wechselfelder unterscheiden sich hinsichtlich ihrer biologischen Wirkung in folgenden Punkten wesentlich: - Der menschliche Körper ist gegen Magnetfelder nicht geschützt: Die Felder sind im Körperinneren nahezu gleichstark wie außerhalb. Auch kommt es nicht zu Feldverzerrungen und die damit einhergehenden Feldstärkenerhöhungen an der Körperoberfläche. - Der ungünstigste Fall ist nicht die Orientierung des Feldes parallel zur Körperlängsachse, wie bei elektrischen Feldern, sondern quer dazu, da hier die Querschnittsfläche und damit die induzierte Stromdichte maximal werden. 52 - Während elektrische Felder immer auftreten wenn Strom bereitgestellt wird, treten die dabei induzierten Magnetfelder nur auf wenn tatsächlich Strom fließt. 3.1.3.1 Bedeutung von niederfrequenten magnetischen Wechselfeldern in der Bioresonanztherapie Seit langem wird in der Schulmedizin die Heilung von Knochenbrüchen durch das Einwirken von Magnetfeldern unterstützt. Durch den Einsatz magnetischer Wechselfelder werden gezielt elektrische Ströme im Körper erzeugt, die an der Frakturstelle das Knochenwachstum stimulieren. Ähnlich wie bei elektrischen Wechselfeldern können die durch ein magnetisches Wechselfeld induzierten Ströme innerhalb des Körpers regulierend wirken. Dabei ist jedoch darauf zu achten wie die Wirbelströme innerhalb des Körpers induziert werden. Dabei hängt die Stärke der in einem Körper induzierten Stromdichten wesentlich von der Form und der Größe der Querschnittsfläche und damit auch mit der Orientierung des Körpers zum Magnetfeld ab. Die maximalen Stromdichten treten an den Einschnürungen der Querschnittsflächen im Bereich des Halses, der Hüften und der Genitalien auf, wenn ein Mensch im horizontalen Magnetfeld aufrecht steht. Abb.31.: Strombahnen induzierter Wirbelströme bei längsorientiertem (links) und querorientiertem (rechts) Magnetfeld. Magnetmatten, die innerhalb der Bioresonanzverfahren eingesetzt werden arbeiten mit gepulst applizierten magnetischen Wechselfeldern, bei denen die im Körper induzierten Ströme zur Regulierung der Körperfunktionen eingesetzt werden. Auf grund der unterschiedlichen Verteilung der Wirbelströme im längsorientiertem bzw. querorientiertem Feld, ist auf die Spulenanordnung innerhalb der Matten zu achten. 53 3.1.4. Hochfrequente elektromagnetische Strahlung Im Gegensatz zu niederfrequenten Feldern, bei der die biologische Wirkung hauptsächlich durch frequenzabhängige magnetische Induktionsprozesse auftritt, ist bei hochfrequenter Strahlung keine physikalische Unterscheidung zwischen der elektrischen und der magnetischen Komponente des Feldes möglich. Aus diesem Grunde verhält sich hochfrequente elektromagnetische Strahlung, besonders was deren Wechselwirkung mit Organismen angeht, grundsätzlich anders als niederfrequente magnetische oder elektrische Wechselfelder. Die biologische Wirkung von HF-Strahlung auf den Organismus beruht im wesentlichen auf der Reaktion des Körpers durch die Absorption von Strahlungsenergie. Die Höhe der Aufnahme von Strahlungsenergie ist frequenzabhängig. Die Energieabsorption erreicht ihr Maximum, d.h. ihren Resonanzbereich, wenn die Wellenlänge der Strahlung in gleicher Größenordnung liegt wie die Körperabmessung. Oberhalb dieses Resonanzbereichs konzentriert sich die absorbierte Energie auf grund der geringeren Eindringtiefe auf immer kleinere Bereiche, so dass nur kleinere Teile des Organismus betroffen sind. Unter dem Begriff hochfrequente elektromagnetische Strahlung wird der sich über 7 Größenordnungen erstreckende Frequenzbereich zusammengefasst, dessen untere Grenze etwa mit der Erregbarkeit von Körperzellen zusammenfällt und dessen obere Grenze den Übergang zur optischen Strahlung bei Wellenlängen unter 1 mm bestimmt. Der Frequenzbereich reicht somit von den Radiowellen im Langwellenbereich (30 kHz) bis zu den Mikrowellen im Millimeterbereich (300 GHz). Langwelle (LW) Mittelwelle (MW) Kurzwelle (KW) 30 kHz (O = 10 km) - 300 kHz (O = 1 km) 3 MHz (O = 100 m) - Ultrakurzwelle (UKW) 30 MHz (O = 10 m) Dezimeterwellen Zentimeterwellen Millimeterwellen 300 MHz (O = 1 m) 3 GHz (O = 10 cm) - 30 GHz (O = 1 cm) - 300 kHz (O = 1 km) 3 MHz (O = 100 m) 30 MHz (O = 10 m) 300 MHz (O = 1 m) 3 GHz (O = 10 cm) 30 GHz (O = 1 cm) 300 GHz (O = 10 mm) Beim Durchgang durch Materie wird die Strahlungsenergie der elektromagnetischen Welle absorbiert, d. h. in Wärme umgewandelt. Durch die Eindringtiefe wird jene Wärmeleistung angegeben, nach der die Strahlungsenergie auf ca. 37% ihres Ausgangswertes abgenommen hat. Je größer die Eindringtiefe, desto gleichmäßiger ist die Wärmebelastung des Körpers. Das Abstrahlverhalten einer Strahlungsquelle wird wesentlich vom Verhältnis der Antennenabmessung zur Wellenlänge (O) bestimmt. Ist die Antennenabmessung wesentlich kleiner als die Wellenlänge, wird die Strahlungsenergie nach allen Seiten gleichmäßig als Kugelwelle abgestrahlt (Dipolantenne). Entspricht die Antennenabmessung näherungsweise der Wellenlänge, so findet nur eine geringe Bündelung der abgestrahlten Welle statt. Wenn die Antennenabmessung wesentlich größer ist als die Wellenlänge, so erfolgt eine starke Bündelung in eine Richtung (Richtantenne). 54 Als Fernfeld bezeichnet man den Bereich, in dem der Abstand zur Strahlungsquelle gleichgroß oder größer ist als die Wellenlänge der Strahlung. Im Fernfeld verhält sich die elektromagnetische Strahlung wie eine ebene Welle. Dabei stehen elektrische und magnetische Feldanteile senkrecht aufeinander und verändern sich gleichzeitig und in gleicher Weise. Die elektrische Feldstärke einer ruhenden Ladung nimmt mit dem Quadrat der Entfernung ab. Im Fernfeld einer Sendeantenne ist jedoch die Abnahme der elektrischen und dadurch bedingt auch der magnetischen Feldstärke nicht mehr so stark und erfolgt linear mit der Entfernung. Im Nahfeld einer Strahlungsquelle ist das elektromagnetische Strahlungsfeld sehr inhomogen, d.h. elektrische und magnetische Feldkomponente sind nicht "in Phase". Im Fernfeld einer Antenne ist die Leistungsflussdichte in W pro m² dem Quadrat der elektrischen und auch der magnetischen Feldstärke proportional und nimmt daher mit dem Quadrat der Entfernung ab. Die Wechselwirkungen elektromagnetischer Strahlung mit einem Organismus ist von der Schwingungsform der Welle und ihrer Orientierung relativ zum Organismus abhängig. Eine linear polarisierte Welle liegt vor, wenn sich die elektrischen und magnetischen Feldstärkevektoren immer in einer Ebene bewegen. D.h. in Ausbreitungsrichtung der Welle gesehen, bewegen sich die Spitzen der Feldvektoren entlang einer Geraden auf und ab. Abb.32: Schwingungsform des elektrischen (E) und magnetischen (H) Feldanteils einer ebenen Welle (K = Ausbreitungsrichtung) Zirkular polarisiert ist eine Welle, deren Feldvektorspitzen in Ausbreitungsrichtung einen Kreis durchlaufen. Bei der elliptischen Polarisation durchlaufen die Feldvektorenspitzen in Ausbreitungsrichtung eine Ellipse. 3.1.4.1 Bedeutung von elektromagnetischer HF-Strahlung in der Bioresonanztherapie Im Gegensatz zu niederfrequenten magnetischen oder elektrischen Wechselfeldern, wo eine biologische Wirkung in erster Linie davon abhängig ist mit welcher Stärke das Feld appliziert wird, sind die HF-Effekte hauptsächlich Frequenzabhängig. 55 Abb.33: Spektrum der elektromagnetischen Strahlung Die dominierende Wirkung elektromagnetischer HF-Strahlung beruht auf der Umwandlung von Strählungsenergie in Wärmeenergie (Absorption). Das Ausmaß der durch Strahlung aufgenommene Wärmemenge erlaubt jedoch keine Rückschlüsse auf deren biologische Wirksamkeit. Die aufgenommene Strahlungsenergie hängt besonders stark von der Frequenz ab. Dies ist dadurch zu erklären, dass sich ein Organismus wie eine Antenne verhält und dann am meisten Energie aufnimmt, wenn die Frequenz auf ihn "abgestimmt" ist. Grundsätzlich können drei Frequenzbereiche unterschieden werden: Der untere Hochfrequenzbereich (Lang-, Mittel- und Kurzwellenbereich) mit Wellenlängen von 10 km bis 10 m. Hier ist die Wellenlänge wesentlich größer als die Körperabmessung. Das Absorptionsvermögen, d.h. die Wärmeentwicklung im Körper ist gering, da die Strahlung den Körper passieren kann. Dabei ist die Verteilung der Wärmeenergie im inneren des Organismus ungleichmäßig, da die durch die Magnetkomponente verursachten Wirbelströme und die durch die elektrische Feldkomponente auftretenden Ströme teilweise entgegengesetzt gerichtet sein können. Im Resonanzbereich (Ultrakurzwelle und Dezimeterwellen), wo die Wellenlänge (10 m - 10 cm) in der gleichen Größenordnung liegt wie die Körperabmessungen erreicht die Energieabsorption ihr Maximum. Die Resonanzfrequenz hängt daher von der Körperabmessung und von der Orientierung der einfallenden Strahlung zum Körper ab. Ist der Organismus parallel zur elektrischen Feldkomponente der Strahlung orientiert (E - Polarisation), liegt die Resonanzfrequenz zwischen 60 und 120 MHz. Oberhalb des Resonanzbereiches, nimmt das Absorptionsvermögen des Körpergewebes mit zunehmender Frequenz zu. Dabei konzentriert sich die Energie jedoch immer mehr auf kleinere Bereiche, so dass die Teilkörper- gegenüber der Ganzkörperbestrahlung immer mehr zunimmt. Als Maß für die Tiefenwirkung für die Strahlungsenergie wird die Eindringtiefe angegeben. Sie ist um so geringer, je höher der Wassergehalt des Gewebes ist. Aus 56 diesem Grund ist die Eindringtiefe für Knochen und Fett etwa 3 - 5fach höher als für Muskelgewebe. Dezimeterwellen im Frequenzbereich von 300 MHz bis 3 GHz, haben eine Eindringtiefe in Gewebe mit hohem Wasseranteil von 4 cm, dringen aber bis zu 26 cm tief in Gewebe mit geringem Wassergehalt ein. Bei Zentimeterwellen zwischen 3 - 30 GHz liegt die Eindringtiefe bei 2 cm (hoher Wassergehalt) und bis zu 11 cm (geringer Wassergehalt). Im Millimeterwellenbereich zwischen 30 - 300 GHz liegt die Eindringtiefe nur noch zwischen 0,2 - 0,6 mm, in Richtung Infrarotbereich bei 5 cm aufgrund der höheren Strahlungsenergie. 3.2 Optische Strahlung Unter optischer Strahlung versteht man den Frequenzbereich elektromagnetischer Strahlung, der sich an den Mikrowellenbereich anschließt und bis an die Grenze der ionisierenden Röntgenstrahlung reicht. Die optische Strahlung beginnt mit dem großen Bereich der Infrarotstrahlung, deren Frequenzbereich sich über drei Zehnerpotenzen erstreckt, gefolgt vom schmalen Bereich des elektromagnetischen Spektrums, den man als sichtbares Licht wahrnehmen kann. Den Abschluss bildet die energiereiche Ultraviolettstrahlung, die sich ebenfalls nur über einen schmalen Bereich erstreckt. Der Frequenzbereich der optischen Strahlung reicht somit über vier Zehnerpotenzen von 300 GHz bis 3000 THz (Tera Hertz = 1012 Hz). Üblicherweise werden diese elektromagnetischen Frequenzbereiche nicht durch ihre Frequenz, sondern durch ihre Wellenlänge beschrieben. Das sichtbare Licht umfasst die Wellenlängen von 780 nm (rot = 3,75 x 1014 Hz) bis zu 380 nm (blau = 7,5 x 1014 Hz), d.h. blaues Licht ist kurzwelliger als rotes. 3.2.1 Infrarotstrahlung Infrarot-Strahlungsquellen sind um so stärker, je höher ihre Temperatur und je größer ihre Ausdehnung ist. Die dominierende Strahlungsquelle ist die Sonne. Ihre Einstrahlung und die wärmeerzeugenden Vorgänge innerhalb der Erde führen dazu, dass auch die Erdoberflächeerwärmt ist und ihrerseits Wärmestrahlung aussendet. Wärmestrahlung wird von allen Materien ausgesendet, dessen Temperatur über dem absoluten Nullpunkt (-273°C) liegt. Das Maximum der Wärmestrahlung liegt bei ca. 3690°C im Infrarotbereich. Obwohl diese Strahlung für uns Menschen nicht sichtbar ist, können wir sie über die Wärmefühler in der Haut wahrnehmen. Die Erwärmung des menschlichen Körpers durch Infrarotstrahlung konzentriert sich auf die Oberfläche und gelangt erst von dort durch die Wärmeleitung ins Körperinnere. Auch wenn die Wärmefühler in der Haut vor einer unbewussten Strahleneinwirkung schützen, kann es doch zu Verbrennungen bei intensiver Sonneneinstrahlung kommen, da die Sonne auch im Infrarotbereich Strahlungsanteile besitzt. Die Eindringtiefe der Infrarotstrahlung nimmt zwar in Abhängigkeit von der Wellenlänge vom fernen zum nahen Infrarotbereich ab, im nahen Infrarot kann die Durchlässigkeit der Haut jedoch selektiv wieder zunehmen. So konzentriert sich die einfallende Infrarotstrahlung einer rotglühenden Fläche, mit einem Strahlungsmaximum bei ca. 4µm auf einen schmäleren Bereich und führt daher zu lokal höheren Erwärmungen und damit früher zu Schmerzempfindungen als die Sonneneinstrahlung gleicher Intensität. Die Schmerzgrenze nimmt bei zunehmender Einwirkungsdauer stark ab. Summiert über die Strahlungsanteile im gesamten Infrarotbereich führen bereits Intensitäten von ca. 150 mW/cm² bei Bestrahlungen über 10 min zu Schmerzempfindungen. Besonders das menschliche Auge ist durch Infrarotstrahlung gefährdet, da die Hornhaut des Auges keine spezifischen Wärmerezeptoren hat, die eine Vermeidungsreaktion wie Lidschluss oder Abwendreaktion auslösen würden. 57 Therapeutisch wird die Infrarotstrahlung aufgrund ihrer Wärmeentwicklung bei Muskelverspannungen und entzündlichen Prozessen eingesetzt. 3.2.2 Sichtbares Licht Für elektromagnetische Strahlung mit Wellenlängen zwischen 780 und 380 nm besitzen wir spezielle Rezeptoren, - die Zapfen und Stäbchen der Netzhaut des Auges. Ein Gegenstand der eine elektromagnetische Strahlung mit einer Wellenlänge von 500 nm reflektiert erkennen wir als "grün". Das zum Farbensehen befähigte Rezeptorsystem in der Netzhaut (photooptische System) besteht beim Menschen aus Blau-, Grün- und Rotzapfen, d.h. aus Rezeptoren, die bei 440 nm (blau), 535 nm (grün) und bei 570 nm (rot) maximal absorbieren. Licht ist ein wesentlicher Bestandteil de Lebens: Es vermittelt nicht nur Information, sondern stellt auch ein wesentliches Stimmungselement dar, das uns sowohl positiv wie negativ beeinflussen kann. Abb.34.: Spektrale Empfindlichkeitskurven der 3 Zapfentypen des Menschen: Blau- (Za1), Grün- (Za2) und Rotrezeptoren (Za3). Jede Wellenlänge ist durch ein bestimmtes Erregungsverhältnis der 3 Zapfentypen charakterisiert. Viele körperinternen Mechanismen, besonders der Wach- Schlaf-Rhythmus werden vom Hell- Dunkel-Wechsel der Sonne synchronisiert. Ungenügende natürliche Lichtzufuhr, wie z.B. in den Wintermonaten, löst bei manchen Menschen eine sogenannte Winterdepression oder die sogenannte "seasonal affective disorder" (SAD) aus. Betroffene können nachts nicht richtig schlafen und sind tagsüber müde. Dies wird oft ausgelöst durch eine nicht mehr dem äußeren Hell- Dunkel- Rhythmus synchronisierte Melatoninsynthese. Melatonin ist ein neurosekretorisches Hormon, das in den Pinealozyten der Epiphyse während der Dunkelheit sezerniert wird. Der Melatoninlevel folgt also einem circardianen Rhythmus, tagsüber sind die Werte niedrig und in den Nachtstunden steigen sie an. Aufgrund einer zu geringen Sonneneinstrahlung, gerade in den Wintermonaten kann dieser Melatoninrhythmus gestört werden und es kommt zu den beschriebenen Störungen. Unter Berücksichtigung dieser chronobiologischen Grundlagen wurden Behandlungskonzepte entwickelt, die ihre Ursache in der Störung des Gefüges der inneren Uhr eines Organismus haben, dem der äußere Zeitgeber zur Synchronisation fehlt: Die Lichttherapie. Seit 1984 wird die Lichttherapie erfolgreich gerade bei Patienten mit Winterdepression eingesetzt. Dabei werden spezielle Lampen verwendet, die in ihrem Strahlungsverhalten annäherungsweise der Intensität und dem Spektrum des Sonnenlichtes entsprechen. Lichttherapiegeräte sollten Beleuchtungsstärken von mindestens 2500 Lux aufweisen, zum Vergleich: herkömmliche Neonlampen arbeiten mit Beleuchtungsstärken von lediglich 500 Lux. Der UV-Anteil der Strahlung muss herausgefiltert sein, um eine 58 Schädigung der Augen zu vermeiden. Die Behandlungsdauer wird vom behandelten Arzt festgesetzt und sollte aber keineswegs in den Abendstunden erfolgen. Abb.35.: Auswirkungen der Lichttherapie bei Depression Das Auge als unser Lichtsinnesrezeptor stellt ein extrem empfindliches Sinnesorgan dar, bei dem schon ein einziges Strahlenquant ausreicht, um eine Sehzelle zu erregen. Darüber hinaus besitzt das Auge einen enormen Dynamikbereich, der von der Wahrnehmbarkeitsschwelle bei 10-18 W/cm² bis zur Blendung bei über 10-6 W/cm² reicht. Erst bei höheren Werten kann eine längere Einwirkdauer zu bleibenden thermischen Schäden der Netzhaut führen. Eine besondere Gefährdung für das Auge stellt Laserlicht dar, da das parallele Laserstrahlbündel auf einen besonders kleinen Brennpunkt (10 bis 20 µm Durchmesser) fokussiert wird, der nur durch die Beugungserscheinungen an der Pupillenöffnung bestimmt wird. Durch zu hohe Lichtintensitäten können thermische, photochemische und mechanische Wirkungen an der Netzhaut auftreten, die auch zu einer Erblindung führen können. Vor einer schädigenden Wirkung von elektromagnetischer Strahlung im Bereich des sichtbaren Lichtes bietet die Funktion der Haut einen gewissen Schutz, der von der Pigmentierung der Haut, d.h. von der Hautfarbe abhängt. Im Gegensatz zu anderen Frequenzbereichen wird das sichtbare Licht zu 45% bis 65% reflektiert. 3.2.3 Ultraviolette Strahlung Bei jedem Material, das über 2500°C erhitzt wird, reicht die ausgesandte Temperaturstrahlung bis in den Bereich des ultravioletten Lichtes. Der kurzwellige Anteil der von der Sonne abgestrahlten UV-Strahlung wird bereits in der Ozonschicht vollständig absorbiert, so dass die auf die Erdoberfläche treffende 59 Strahlung nahezu keine Anteile unter ca. 288 nm mehr besitzt. Die UVStrahlungsintensität unterliegt starken jahreszeitlichen Schwankungen und ist im Herbst bei geringer Luftdichte höher als im Frühjahr. Der UV-Anteil der Globalstrahlung liegt bei etwa 5 mW/cm², wobei die Strahlenbelastung mit der Meereshöhe und dem Reflexionsvermögen der Erdoberfläche ansteigt (hohe Werte deshalb auf Gletscher). UV-Bestrahlungslampen werde aufgrund ihrer photochemischen und photobiologischen Wirkung für kosmetische (Bräunung), hygienische (Desinfektion), medizinische und technische Zwecke verwendet. In der Medizin wird die UV-Strahlung bei der Behandlung von Schuppenflechte, Akne, allergische Hautreaktionen, aber auch bei rheumatischen Erkrankungen und chronischen Lungenentzündungen eingesetzt. Darüber hinaus wird UV-Strahlung zur Vorbeugung gegen Vitamin D-Mangel eingesetzt. Im Hinblick auf ihre biologische Wirkung wird die UV-Strahlung unterteilt in die Bereiche UV-A (380 bis 315 nm), in dem vorwiegend Bräunungswirkungen vorherrschen, UV-B (315 bis 280 nm), in dem die Haut geschädigt wird (Sonnenbrand) und der UV-C Strahlung (unter 280 nm) in dem Keime und Bakterien abgetötet werden können. Im UV- Frequenzbereich besitzen die Strahlungsquanten bereits so viel Energie, dass Quanteneffekte, also photobiologische Wirkungen auftreten können. Dabei können durch die Absorption von Strahlungsquanten Moleküle angeregt und vorübergehend in einen reaktiveren Zustand versetzt, chemische Reaktionen auslösen oder deren Verlauf derart beeinflussen, dass chemische Verbindungen aufgebrochen werden können. Absorbiert ein Molekül ein Strahlungsquant, kann es seine Energie aufnehmen in Form von: - Bewegungsenergie, d.h. Rotations- oder Translationsbewegung (Erwärmung) - Anregung, indem ein Elektron des Moleküls in einen höheren energetischen Zustand versetzt wird (höhere Elektrodenbahn) und damit das Molekül reaktiver wird und neue chemische Verbindungen eingehen kann (photochemische Reaktion) - Änderung des Ablaufs einer chemischen Reaktion - Änderung von Molekülen durch das Aufbrechen chemischer Verbindungen - Befreiung eines Elektrons aus seiner Atomhülle (Ionisation) Die Grenze zwischen der UV-Strahlung kurzer Wellenlänge und der ionisierenden Strahlungen ist unscharf und liegt (je nach Atom) zwischen 319 und 50 nm. Definitionsgemäß wird die UV-Strahlung bis zu einer Wellenlänge von 100 nm angenommen, bis zu der die biologische relevanten Atome C (Kohlenstoff), O (Sauerstoff) und N (Stickstoff) nicht ionisiert werden können. Im UV-Bereich ist vor allem die Absorption durch ungesättigte organische Verbindungen, wie die Desoxiribonukleinsäure (DNA), dem Trägermolekül des Erbgutes, zu beachten. Die destruktive Wirkung der Strahlenquanten auf die DNA kann zu schwerwiegenden Zellveränderungen wie z.B. Krebs führen. Auf der anderen Seite löst die UV-Strahlung in der in der Haut die Umwandlung von 7Dehydrocholesterol in Vitamin D aus. 3.3 Ionisierende Strahlung Obwohl die ionisierende Strahlung als extrem hochfrequente elektromagnetische Strahlung innerhalb der Bioresonanzverfahren keine Rolle spielen, werden sie aus Gründen der Vollständigkeit kurz erläutert. 60 Unter der Bezeichnung ionisierende Strahlung fasst man alle Strahlungsarten zusammen, die eine Ionisation bewirken können, also deren Quantenenergie gleich oder größer der Bindungsenergie eines Elektrons ist. Aus diesem Grund wird diese Form der Strahlung nicht mehr durch Frequenz oder Wellenlänge, sondern durch ihre Quantenenergie charakterisiert. Das Maß für ionisierende Strahlung ist ein Elektronenvolt (eV). Dies beschreibt jene Energie, die ein Elektron in Form von Bewegungsenergie aufnimmt, wenn es mit einer Spannung von 1 Volt beschleunigt wird. Die Bindungsenergie der Elektronen liegt zwischen 2 eV für eine Elektron in der äußeren Schale, z.B. des Kupferatoms, und 88005 eV für ein Elektron in der inneren Schale des Bleiatoms. Unter dem Begriff der ionisierenden Strahlung fallen die elektromagnetische Strahlung und die Teilchenstrahlung. Elektromagnetische ionisierende Strahlung: Obwohl sie aus physikalischer Sicht identisch sind unterscheidet man bei ionisierender elektromagnetischer Strahlung nach der Art ihrer Entstehung zwischen Röntgen- und Gammastrahlung. Die Röntgenstrahlung entsteht außerhalb des Atomkerns durch Vorgänge in der Atomhülle (Eigenstrahlung) oder durch Abbremsvorgänge sich schnell bewegter Elektronen bei Annäherung an den Atomkern (Bremsstrahlung). Demgegenüber entsteht Gammastrahlung durch Vorgänge innerhalb des Atomkerns z.B. beim Zerfall radioaktiver Atomkerne (Radionuklide). Teilchenstrahlung: Alpha-Strahlung: Bestehend aus Atomkernen des Elementes Helium, die sich aus zwei Protonen und zwei Neutronen zusammensetzten. Bei der Aussendung von AlphaStrahlung wandelt sich das Radionuklid um, und es entsteht ein neues Element. Beta-Strahlung, bestehend aus einem Elektron, das ausgesendet wird, wenn ein Neutron des Kerns seine negative elektrische Ladung abgibt und sich in ein positiv geladenes Proton umwandelt. Dadurch entsteht ebenfalls ein neues Element. Neutronenstrahlung entsteht, wenn sich ein instabiles Radioisotop in ein stabiles Isotop umwandelt. Da sich dabei die Kernladung nicht ändert entsteht kein neues Element, jedoch verändert sich die Massenzahl um 1. Bei Spaltprozessen hingegen, wenn ein instabiler Atomkern zerfällt, werden ebenfalls Neutronen frei. Auf der bei diesen Spaltprozessen frei werdenden Energien beruht die zivile Nutzung der Kernenergie. Im medizinischen Bereich wird Röntgenstrahlung zur Diagnose und zur Therapie (Strahlentherapie) eingesetzt. In der Strahlentherapie wird durch eine sorgfältiges Bestrahlungsregime erreicht, dass das den Tumor umgebende gesunde Gewebe weitgehend geschont und hohe Strahlendosen nur auf den Tumorbereich beschränkt bleiben. 61 3.4 Magnetostatische Felder Magnetische Felder treten immer dann auf wenn elektrische Ladungen bewegt werden, d.h. ein magnetostatisches Feld entsteht wenn Gleichstrom fließt. Im Gegensatz zum elektrischen Feld, wo die Feldlinien an positiven Ladungen beginnen und an negativen enden, sind magnetische Feldlinien in sich geschlossen und umschließen den sie erzeugenden Strom. Man spricht daher von Wirbelströmen. Die magnetische Feldstärke H ist um so größer, je größer die Summe der elektrischen Ströme ist. Die magnetische Feldstärke beschreibt die Eigenschaften eines Magnetfeldes nur ungenau, da bei gleicher Feldstärke die Feldeinwirkung durch die magnetischen Materialeigenschaften, die Permeabilität, eines Stoffes bestimmt wird. Die magnetische Induktion oder magnetische Flussdichte B beschreibt die Feldstärke in Bezug auf die Permeabilität des Feldes. Die Einheit der magnetischen Induktion bzw. der magnetischen Flussdichte ist 1 Tesla (1 T = 1 Vs/m²). Je nach dem Verhalten von Materialien im Magnetfeld unterscheidet man: - diamagnetische Stoffe, deren Moleküle sich im Magnetfeld so ausrichten, dass es insgesamt zu einer Schwächung des Magnetfeldes kommt. Paramagnetische Stoffe, deren Moleküle sich im Magnetfeld so ausrichten, dass es zu einer Verstärkung des Magnetfeldes kommt. ferromagnetische Stoffe wie z.B. Eisen, Kobalt und Nickel, die sich qualitativ wie paramagnetische Stoffe verhalten, jedoch eine starke Feldverstärkung hervorrufen. Ähnlich wie Gegenstände hoher elektrischer Leitfähigkeit auf ein Magnetfeld einwirken und dieses verzerren, verursachen auch Gegenstände mit einer hohen Permeabilität (wie z.B. ferromagnetische Stoffe) Verzerrungen des Magnetfeldes. Der menschliche Körper unterscheidet sich in seiner magnetischen Permeabilität nicht im wesentlichen von der, der Luft. Aus diesem Grund kommt es an der Körperoberfläche nicht zu einer Erhöhung der magnetischen Feldstärke, d.h. ein einwirkendes Magnetfeld durchdringt den menschlichen Körper und die magnetische Induktion tritt innerhalb Körpers ungeschwächt auf. 3.4.1.Das Erdmagnetfeld Nach der Dynamo-Theorie erzeugen Konvektionsströme des flüssigen Erdkerns das primäre Magnetfeld, das durch die Erdrotation (Coriolis-Kraft) in der Rotationsachse ausgerichtet wird. Dadurch entsteht ein weitgehend statisches Dipolfeld, dessen Pole sich in der Nähe der beiden geographischen Pole befinden. Als geographischer Nordpol wird allgemein der in der Arktis gelegene Pol bezeichnet, da die nordwärts gerichtete Spitze einer Kompassnadel in dessen Richtung zeigt. Aus physikalischer Sicht ist jedoch der am geographischen Nordpol gelegene Magnetpol der magnetische Südpol. Umgekehrt befindet sich der magnetische Nordpol in der Antarktis, nahe dem geographischen Südpol. Durch die Neigung des Dipols gegenüber der Erdachse ergibt sich eine Abweichung zwischen magnetischem und geographischem Pol, die als Deklination bezeichnet wird. Sie beträgt in den meisten Gebieten der Erde weniger als 20 ° Grad; lediglich in der Nähe der magnetischen Pole kommt es zu größeren Abweichungen. Die magnetischen Feldlinien treten am magnetischen Nordpol aufwärts gerichtet aus der Erdoberfläche und treten am magnetischen Südpol, in der Nähe des geographischen Nordpols wieder ein. Auf der südlichen Hemisphäre verlaufen die magnetischen Feldlinien aufwärts, auf der Nordhalbkugel abwärts gerichtet. Am magnetischen Äquator verlaufen sie parallel zur Erdoberfläche. 62 Die Stärke des magnetischen Feldes (H) wird allgemein in Tesla (nT) angegeben, obwohl dies eigentlich der Einheit der magnetischen Kraftflußdichte bzw. der magnetischen Induktion (B) entspricht. Da B = P x H gilt und die Permeabilität P in Luft und unter Atmosphärendruck 1,000024 beträgt, entspricht diese eigentlich falsche Angabe der Feldstärke in Tesla dem numerisch richtigen Wert. Die Angaben der Totalintensität des Erdmagnetfeldes werden in der Einheit Tesla = T (nano Tesla = nT) angegeben. Am Pol erreicht die Totalintensität ihren Maximalwert von 70 000 nT und wird nur von der Vertikalintensität bestimmt (Horizontalintensität = 0 nT), da die Feldlinien in einem 90° Winkel in die Erdoberfläche eintreten. Im Gegensatz dazu erreicht am Äquator die Horizontalintensität ihren höchsten Wert von ca. 26 000 nT, da die Feldlinien parallel zur Erdoberfläche verlaufen und keinerlei Vertikalkomponente besitzen. Zusätzlich werden durch teilweise in die Erdkruste eingelagerte magnetische Materialien die ursprüngliche Gleichmäßigkeit des Dipolfeldes mehr oder weniger stark durch magnetische Anomalien moduliert. Obwohl das Erdmagnetfeld ein statisches Dipolfeld ist, unterliegt es immerhin so großen Schwankungen, dass es in Abständen von mehreren 105 bis 106 Jahren sogar zu Polaritätsumpolungen kommt. Diese Änderungen erfolgen wahrscheinlich so langsam, dass sie in der Lebensspanne von einzelnen Organismen keine Rolle spielen. Im Gegensatz zu diesen, über lange Zeiträume ablaufenden Prozessen, treten periodische tages- und jahreszeitliche Magnetfeldschwankungen auf, die hauptsächlich durch die Sonne verursacht werden (CAMPBELL 1967). Durch die von der Sonne ausgesandte elektromagnetische Wellen- und Korpuskularstrahlung wird die Ionosphäre der Erde regelmäßig beeinflusst. Die elektrisch geladenen Teilchen der Ionosphäre werden durch die Sonnenwinde bewegt und induzieren ein Magnetfeld. Auf der Nachtseite der Erde wird unter dem Einfluss des Sonnenwindes die Ionosphäre stark auseinandergezogen, während sie auf der Tagseite stark komprimiert wird. Daraus resultiert ein regelmäßiger Tagesgang des Erdmagnetfeldes, bei dem in unseren Breiten die Totalintensität während des höchsten Sonnenstandes am Mittag ein Minimum erreicht. Da der Tagesgang von der Sonne beeinflusst wird, besitzt er je nach geographischer Breite und dem relativen Stand der Erde zur Sonne einen unterschiedlichen Verlauf in Abhängigkeit von den Jahreszeiten. In den Sommermonaten, wenn die Sonneneinstrahlung am stärksten ist, sind die tageszeitlichen Schwankungen bis zu viermal höher als in den Wintermonaten. Zusätzlich verändert sich in einem 11jährigen Zyklus die Sonnenaktivität. Die äußerlich sichtbaren Erscheinungen dieser Aktivität sind die Sonnenflecken. Durch die Oszillation des solaren Magnetfeldes kommt es zu der periodischen Variation der Sonnenfleckenzahlen. Während der maximalen Sonnenaktivität mit einer hohen Anzahl an Sonnenflecken kommt es zu einem vermehrten Ausstoß an Wellen- und Korpuskularstrahlung, der die Ionosphäre der Erde und damit das Erdmagnetfeld beeinflusst. Magnetische Stürme in Zeiten erhöhter Sonnenaktivität beeinflussen die regelmäßigen tages- und jahreszeitlichen Magnetfeldschwankungen. 3.4.2 Biologische Relevanz von magnetostatischen Feldern Das magnetische Feld der Erde spielt für das Leben auf ihr eine entscheidende Rolle: Es schützt weite Bereiche vor der gefährlichen Korpuskularstrahlung (von der Sonne und aus dem Weltraum). Auch sind alle Lebewesen diesem mehr oder weniger statischen Feld ausgesetzt und aufgrund dessen benötigen Organismen das 63 Erdmagnetfeld, ähnlich der Gravitation, zur Entwicklung. Trotz der, gegenüber technisch erzeugten Feldern, geringen Intensität von nur 50 000 nT (in unseren Breiten), sind sogar manche Organismen, wie Vögel und Bienen in der Lage das Erdmagnetfeld zur Orientierung zu nutzen. Sie besitzen dafür spezielle Sinnessysteme, die es ihnen ermöglichen Richtungsinformationen aus den Erdfeldparametern zu nutzen. Auch wenn für den Menschen keine speziellen Magnetrezeptoren nachgewiesen werden konnten, so benötigt unser Organismus doch das geomagnetische Feld zur Aufrechterhaltung seiner Funktion. In einem magnetisch abgeschirmten Raum, in dem die Intensität des Erdmagnetfeldes von 50 000 nT nur noch 300 nT beträgt, können wir uns nicht lange aufhalten. Ähnlich wie in der Schwerelosigkeit, d.h. ohne Gravitation, können essentielle Stoffwechselvorgänge ohne das Vorhandensein von Magnetfeldern in Erdfeldstärke gestört werden. Unter dem Einfluss eines Magnetfeldes werden im Körperinneren elektrische Spannungen erzeugt, da dort elektrische Ladungen bewegt werden. Bewegen sich positiv und negativ geladene Teilchen in gleicher Richtung, wie dies z.B. in den Blutgefäßen der Fall ist, führen einwirkende Magnetfeldkräfte zu einer Ladungstrennung und damit zum Auftreten einer elektrischen Spannung quer zur Bewegungsrichtung. Diese ist um so größer, je höher die Bewegungsgeschwindigkeit ist (Induktion). Biologische Auswirkungen sind also überall dort denkbar, wo in einem Körper schnelle Bewegungsabläufe auftreten oder wenn sich ein Körper schnell in einem Magnetfeld bewegt. Diese Induktionsvorgänge sind jedoch nur unter dem Einfluss technisch erzeugter Felder möglich, die weit über der Intensität des Erdmagnetfeldes liegen. Die größten Bewegungsgeschwindigkeiten im menschlichen Organismus treten am Herzen auf. Durch dessen Kontraktion wird das Blut mit Spitzengeschwindigkeiten von weit mehr als 1 m/s in die Aorta gepumpt. Die Herztätigkeit steigt bei körperlicher Arbeit oder bei sportlicher Betätigung an und die Blutgeschwindigkeit kann dabei bis auf das 5fache ansteigen. Dies bedeutet, dass quer zur ca. 2,5 cm messenden Aorta im ungünstigsten Fall eine Spannung von 125 mV pro Tesla auftreten kann. Dadurch können außerhalb der Aorta im Takt der Herzfrequenz Stromdichten-Spitzenwerte in der Größenordnung von 10 µA/cm² pro Tesla verursacht werden. Die durch ein Magnetfeld verursachten elektrischen Vorgänge können anhand des EKGs durch eine Veränderung der Kurvenform während der Auswurfphase des Ventrikels, bei Induktionen über 300 mT beobachtet werden. 3.5 Erdstrahlung Über die Natur der sogenannten Erdstrahlung existieren unterschiedliche Vorstellungen. Teilweise wird von einer noch unbekannten Form der Energie ausgegangen, teils wird abgenommen, es handelt sich um elektromagnetische Strahlung oder Teilchenstrahlung. Erdstrahlung ist dabei der Sammelbegriff, der alle physikalischen Kräfte die auf die Erde wirken zusammenfasst. Unterschieden wird nach der Ausrichtung der Strahlung, den Gitter- bzw. KubenSystemen, den lageabhängigen Erdstrahlen (z.B. hervorgerufen durch Wasseradern) und den beweglichen Gittersystemen. Dabei haben alle Gittersysteme folgende Eigenschaften: An den Randzonen der bevorzugten Strahlungsrichtung tritt eine verstärkte gerichtete Strahlung auf. Dadurch entstehen sogenannte Reizzonen oder, räumlich betrachtet, Wände von gerichteter Strahlungsdichte, die senkrecht zur Erdoberfläche stehen. Neben dieser senkrechten Strahlung weisen diese Gittersysteme zusätzlich eine Horizontal- sowie eine Querstrahlung auf. 64 3.5.1 Kubensystem nach BENKER Das atomare Kubensystem nach BENKER ist als übergeordnetes System zu betrachten. In ihm können sämtliche anderen Gittersysteme und Reizzonen vorkommen. Das System stellt man sich als aneinandergereihte, gestapelte Quader von 10 m Seitenlänge und einer Wandstärke bis zu 1 m vor. Die Ausrichtung ist magnetisch nach N -S und O - W ausgerichtet. Die einzelnen Kuben sind abwechselnd mit plus und minus polarisiert und weisen in Richtung Zenit. An den Seiten des atomaren Gitters ist die gerichtete Strahlung besonders hoch und ist in der Lage Moleküle zu schädigen. Dabei soll besonders Strahlung der Reizzonen der N -S- Strahlung und die Kreuzpunkte besondere pathologische Wirkung haben. 3.5.2 Hartmann-Netz Das Hartmann-Netz (nach Dr. E. Hartmann) ist ebenfalls nach N - S und O - W ausgerichtet und ist im Benker-Kubensystem enthalten. Die Reizstreifen sind ca. 21 cm breit und haben in N - S Richtung 2 m- und in O - W Richtung 2,50 m Abstand, wobei die Abstände je nach geographischer Lage variieren. Ähnlich des Kubensystems wirken besonders die Kreuzpunkte des Hartmann-Netzes pathologisch, während die Reizzonen das Wohlbefinden negativ beeinflussen. 3.5.3 Curry-Netz Dieses Gitternetz, das ebenfalls in Quader mit entgegengesetzter Polarität aufgeteilt ist, hat Reizstreifen von ca. 60 cm Breite, verläuft diagonal zum Kubensystem und Hartmann-Netz mit einem Abstand von ca. 3,5 m. Hier wirken die Kreuzungspunkte mit dem Kubensystem und dem Hartmann-Netz negativ auf das Wohlbefinden aus. 3.5.4 Ley-Linien Die Ley-Linien hingegen stellen energetisch "Linien der Kraft" dar und haben eine überwiegend positive Wirkung auf Organismen. Viele sakrale Bauten und Kultstätten sind auf Ley-Linien gebaut. 3.5.5 Geostationäre Strahlung und Wasseradern Geostationäre Strahlung sind Reizzonen deren Ursache geologisch bedingt ist. Diese Störzonen, wie z.B. Wasseradern, geologische Verwerfungen und Erdspalten arbeiten als sogenannte Strahlungssammler und haben eine pathologische Wirkung auf Organismen. 65 Abb. 36.:Verschiedene Störzonen mit möglicherweise pathogener Wirkung 66 4. Arbeitsweise der Bioresonanzverfahren Mit Bioresonanzverfahren wird versucht, direkt in die Regulationsvorgänge eines Organismus einzugreifen. Um die Arbeitsweise dieser Verfahren zu verstehen, muss man sich zunächst klarmachen, dass ein Organismus ein energetisch offenes, schwingungsfähiges System darstellt und das alle Subsysteme (Organe usw.) innerhalb eines Organismus miteinander vernetzt sind. Ein auf den Organismus eintreffender Reiz kann auf alle Subsysteme rückkoppelnd aufgeschaltet werden und damit an verschiedenen Subsystemen unterschiedliche Antworten auslösen. Filtermechanismen (die Stellglieder in einem Regelkreis, siehe Kapitel 2.3) können amplitudenabhängig die Reizwirkung über negative Rückkopplung begrenzen. Auf der anderen Seite können geringste Differenzen im Ausgangswert, sich bei fehlender Korrektur im weiteren Verlauf über positive Rückkoppelung zu dramatischen Veränderungen aufschaukeln. Ein Organismus funktioniert im wesentlichen über mehr oder weniger zusammenhängende bzw. kohärente elektromagnetische und mechanische Eigenschwingungen seiner Bausteine und Regelkreise, die fluktuierend ineinander greifen. Gesundheit ist daher aus biophysikalischer Sicht ein "kohärenter Zustand", während Krankheit ein Verlust an Kohärenz darstellt. Anders ausgedrückt ist Krankheit im technischen Sinne ein "Nichtgleichgewichtsphasenübergang". Die Grundbausteine eines Organismus, die elektrische Ladungen tragenden Atome und Moleküle, senden bei Beschleunigung laufend unterschiedliche Energiemengen in Form von Quanten bzw. elektromagnetischer Strahlung verschiedenster Frequenzen aus. Trifft diese Strahlung auf andere Atome, Moleküle, Ionen, Polymere usw. wird diese Energie in Eigenbewegung umgesetzt. Daher findet in einem Organismus ein ständiger Wechsel von Energiequanten in Form von Absorption und Emission statt. Atome schwingen bei menschlicher Körpertemperatur mit mehr als 1015 Hz. Moleküle als Atomverbund haben ihre Hauptfrequenz bei ungefähr 109 Hz. Zellen als Molekülverband reagieren auf 103 Hz. Der Gesamtorganismus Mensch als Organverband schwingt insgesamt mit einer Frequenz von 7 bis 10 Hz (Schumann-Wellen, siehe Kapitel 3.1.2.1.). daraus kann gefolgert werden, dass der Verbund Grundsubstanz - Zellen ebenfalls mit 7 bis 10 Hz schwingt, wie dies z.B. vom Gewebe wichtiger Gehirnzentren abgeleitet wurde. Alle Schwingungen innerhalb eines Organismus sind sensibel für kohärente Schwingungsmomente und die daraus resultierenden Überlagerungen mit der aus der In- und Umwelt erzeugten Oszillationen ist unüberschaubar groß. Jede chemische Reaktion innerhalb des Organismus kann als physikalische Feldeinwirkung interpretiert werden. Zusammenfassend kann daraus geschlossen werden, dass Ordnung oder Homöostase in einem Organismus durch Kohärenz bis in den subatomaren Bereich entsteht, wobei die elektromagnetische Steuerung, z.B. über Ionenverschiebungen an Membranen, den gleichen Stellenwert hat wie biochemische Veränderungen. Biochemische Veränderungen eines Organismus, z.B. durch die Verabreichung von Medikamenten, können zwar die Symptome (z.B. Schmerzen) eines entstandenen Ungleichgewichts (Krankheit) behandeln, in den wenigsten Fällen kann aber die Ursache der Funktionsstörung behoben werden und damit die selbstregulatorischen Prozesse innerhalb des Organismus in Gang setzen. Die "physikalische" Behandlung von Funktionsstörungen (bzw. Krankheiten) mit Hilfe von Bioresonanzverfahren hat den Vorteil, dass wie in der Homöophatie oder bei der Akupunktur, die Nebenwirkungen relativ gering sind. Der Körper empfängt eine 67 elektromagnetisch codierte Information darüber, wie die regulatorische Störung behoben werden kann. Dabei kann es zwar auch teilweise zu einer Erstverschlimmerung der Symptomatik kommen, aber sonstige Nebenwirkungen sind bei der Anwendung von Bioresonanzverfahren nicht festzustellen. Zwar bedeutet der Einsatz von Bioresonanzverfahren nicht, das damit alle "Krankheiten" bzw. Funktionsstörungen eines Organismus behoben werden können, jedoch bietet es Möglichkeiten auf "sanftem" Wege, Symptome zu lindern und dem Körper die Information zur Selbstheilung zu liefern. 4.1. Elektromagnetische Bioresonanz Aufgrund der physikalischen regulatorischen Vorgänge innerhalb eines Organismus ist es naheliegend elektromagnetische Schwingungen wie sie an der Körperoberfläche z.B. in Form des EKG, EEG oder an Akupunkturpunkten gemessen werden können, bei Gesundheitsstörungen gegenläufig wieder einzuspeisen, um damit pathologische Frequenzen auszugleichen. Vorraussetzung dafür ist, dass zum einen pathologische Schwingungsmuster (z.B. Viren, Bakterien, Pilze usw.) bekannt sein müssen und zum anderen, dass über geeignete Elektroden ausgleichende Frequenzen appliziert werden können. Das Problem dabei ist, dass mit Hilfe gängiger Elektroden es nicht möglich ist körpereigene Störschwingungen, die teilweise im Mirkowellenbereich (zwischen 1 und 10 000 GHz) liegen abzugreifen. Deshalb arbeiten die hier vorgestellten Bioresonanzgeräte (QXCI bzw. SCIO oder Life System) mit evozierten Potentialen. 4.1.1. Systemdiagnostik mit Bioresonanzverfahren Beide Systemen arbeiten mit einem Interface in Verbindung mit einem entsprechenden Computerprogramm. Im jeweiligen Computerprogramm sind die unterschiedlichsten pathogenen Schwingungsmuster gespeichert und werden während des Testlaufes über die angeschlossenen Elektroden in den Organismus des zu testenden Körpers gebracht. Die applizierten Schwingungsmuster treten in Resonanz mit den im Körper vorhandenen Frequenzen. Im Rechner wiederum werden die Reaktionen des Körpers auf die applizierten Frequenzmuster analysiert und im Anschluss an den Test in einer Art Funktionsdiagnostik dargestellt. Dabei werden die vom System applizierten Frequenzen, die die größte Übereinstimmung mit den im Körper gefundenen Schwingungen aufzeigen, als jeweilige Funktionsstörung dargestellt. In einer "Ranking"-Darstellung werden die momentan akutesten Störungen in absteigender Reihenfolge aufgezeigt. Die Ergebnisse der Bioresonanzverfahren darf nicht mit der üblichen Zustandsdiagnostik in der Schulmedizin gleichgesetzt werden. Es handelt sich weitgehend um eine Systemdiagnostik die lediglich akute regulatorische Störungen aufzeigen kann. Schwierigkeiten in der Interpretation der Testergebnisse können auftreten, da die gemessenen Frequenzmuster den im Computer gespeicherten Daten zugeordnet werden. So muss ein ermitteltes Testergebnis immer unter der Prämisse der Funktionsstörung betrachtet werden. Die Meldung "Cancer" oder "Tumor" heißt nicht zwingend, dass der getestete Organismus "Krebs" hat. In einem Organismus sind aus regulatorischer Sicht betrachtet immer mehr oder weniger degenerative Prozesse zu beobachten, die aber in einem funktionierendem System ausgeglichen werden können. Kommt bei einem Test mit einem Bioresonanzverfahren also die Meldung z.B. "Lung-Cancer" so bedeutet dies, das die Lunge des betroffenen Organismus Unterstützung im Hinblick auf ihre Selbstregulation benötigt. Auf der anderen Seite wird in den wenigsten Fällen, wo in einem Körper Neoplasien (Krebszellen) festgestellt wurden, dies beim Bioresonanztest auch festgestellt. In 68 diesen Fällen hat der Adaptationsprozess im Körper schon begonnen und der "Krebs" wird nicht als akute Funktionsstörung klassifiziert. Trotzdem kann mit Hilfe der Bioresonanzverfahren der betroffene Organismus unterstützt werden indem vor allem das Immunsystem angeregt wird, um die Selbstheilungskräfte zu verstärken. 4.1.2. Applikation von regulierenden Frequenzmustern In den hier vorgestellten Bioresonanzsystemen ist nicht nur möglich durch das Testverfahren mittels evozierten Potentialen eine Funktionsdiagnostik zu erstellen, sondern auch verschiedenen Frequenzmuster zu applizieren, die bei bestimmten Funktionsstörungen regulierend auf den Körper einwirken können. Diese regulatorischen Frequenzen liegen in der Regel im Bereich von ELF (extrem low frequency) und im VLF-Bereich (very low frequency) der elektromagnetischen Feldern (siehe Kapitel 3.1.2.). Können aber auch in den Bereich der hochfrequenten Strahlung bis 3 GHz reichen. Dabei induzieren die im Niederfrequenzbereich applizierten Felder Wirbelströme im Körper, die regulierend auf den Organismus einwirken können. Allgemein betrachtet werden bei der Anwendung von Bioresonanzverfahren dem Körper Informationen in Form von elektromagnetischen Frequenzmuster geliefert, die seine Selbstheilungskräfte unterstützen. Je chronischer eine Krankheit ist, d.h. je länger eine regulatorische Funktionsstörung vorliegt, um so schwieriger die Beeinflussung durch ein Bioresonanzverfahren. Betrachtet man, wie in der biologischen Medizin, den Körper als offenes System mit untereinander vernetzten Regelkreisen (siehe Kapitel 2.3.), so kann eine chronische Störung zu einer Adaptation innerhalb des Systems führen, die auch mit einem "richtigen" Informationsinput nur schwer aufzubrechen ist. Die Information, die über das Bioresonanzverfahren in den Organismus vermittelt wird, wirkt über den "Regler" des Regelkreises auf das dementsprechende "Stellglied" und es kann - muss aber nicht zu einer Verbesserung des Allgemeinbefindens kommen. Vor allen Dingen die Gewebe, wie Nervensystem und Muskeln, deren Kommunikation innerhalb des Körpers über elektromagnetische Signale abläuft, sind mit Hilfe von Bioresonanzverfahren positiv zu beeinflussen. Gerade innerhalb der Neurologie werden mittlerweile Therapien mit niederfrequenten elektromagnetischen Feldern in klinischen Studien erprobt. Mit der Transkraniellen MagnetStimulation (TMS) werden erfolgreich depressive Patienten behandelt. Das Grundprinzip des Verfahrens ist einfach. Eine Magnetspule wird mit geringem Abstand zum Kopf über das zu stimulierende Hirnareal gehalten - bei depressiven Patienten beispielsweise über die linke Schläfe. Wenn Strom durch den Spulendraht fließt, baut sich ein starkes Magnetfeld mit einer Feldstärke von bis zu 2,5 T (dies entspricht ungefähr der 40 000fachen Stärke des Erdmagnetfelds) auf. Das künstlich erzeugte Feld induziert im Gehirn wiederum einen Stromfluss (Wirbelströme, siehe Kapitel 3.1.2). Auf diese Weise lässt sich die auf elektrischen Impulsen basierende Kommunikation zwischen den Nervenzellen beeinflussen. Der Trick bei der Behandlung depressiver Patienten besteht darin, dass der Strom mit einer Frequenz zwischen 5 und 100 Hz durch die Spule fließt - repititive Magnetstimulation (rTMS) wird diese Variante der Therapie bezeichnet. Dabei ist festzustellen, dass niedrige Frequenzen die neuronale Aktivität hemmen, während eine höhere Impulsrate eine Erhöhung der Nerventätigkeit auslöst. Letztere Applikation kommt insbesondere den depressiven Patienten zugute, deren linkes Vorderhirn chronisch unteraktiv ist. Offenbar fördert die Magnetstimulation in diesem Gehirnareal die Ausschüttung von essentiellen Botenstoffen wie Serotonin und bringt so die aus 69 dem Tritt geratenen Neurone auf ein normales Aktivitätsniveau zurück. Dabei bleibt die Wirkung des Verfahrens über mehrere Tage erhalten, verschwindet also nicht direkt nach dem Entfernen der Spulen. Daraus kann man schließen, dass das Verfahren nicht nur kurzfristig in den Neurotransmitterhaushalt eingreift, sondern auch das Netzwerk der Neurone verändert. Molekularbiologische Studien konnten belegen, dass das Verfahren in den Neuronen zusätzlich einen Transkriptionsfaktor aktiviert, der das Wachstum der Zellen anregt. D.h. die niederfrequente Magnetstimulation fördert nicht nur eine verstärkte Synapsenbildung zwischen bereits vorhandenen Neuronen, sondern lässt auch neue Nervenzellen auswachsen. Ein vergleichbares Verfahren wird bei Knochenbrüchen und Gelenkproblemen angewandt. Auch hier werden über niederfrequent applizierte Magnetfelder Wirbelströme an den Bruchstellen oder am Gelenk induziert, die die Neubildung von Osteozyten - und damit das Knochenwachstum fördern. Aus biophysikalischer Sicht schwingen die Systeme eines Organismen bei Körpertemperatur mit einem bestimmten Muster. Die Frequenz von Zellen als Molekülverband liegt bei ungefähr 1000 Hz und der gesamte menschliche Organismus als Organverbund mit Frequenzen zwischen 7 und 10 Hz. Störungen dieser Frequenzmuster können zu Regulationsstörungen führen, die mithilfe von Bioresonanzverfahren ausgeglichen werden können. 4.1.2.1. Rife- Frequenzen Spezielle Frequenzmuster die besonders pathogene Keime, wie Viren, Bakterien und Pilze inaktivieren, wurden von Royal Raymond Rife (1888 - 1971) erforscht. Bei seinen Untersuchungen im Bereich der Entwicklung neuer mikroskopischer Verfahren erkannt er, dass sich durch die Bestrahlung mit bestimmten Frequenzen Viren und andere pathogene Keime mikroskopisch erkannt werden können. Daraus schloss er, dass es sich dabei um Resonanzphänomene handelt, bei denen spezifische elektromagnetische Strahlung die Membranen der Keime zum Schwingen bringen. Durch diese spezifischen Resonanzschwingung können die Membranen aufgrund der dadurch ausgelösten Verstärkung ihrer Eigenschwingung zerstört werden und damit den jeweiligen Keim inaktivieren (ähnlich die durch hohe Töne erzeugte Resonanzschwingung von Glas, die das Glas zum platzen bringen kann). Dieses Verfahren von Rife ist in den hier vorgestellten Bioresonanzsystemen integriert. Inwieweit tatsächlich durch dieses Verfahren z.B. Viren inaktiviert werden können oder ob bei der Anwendung der Bioresonanz lediglich das Immunsystem stabilisiert werden kann und es dadurch zu einer Verbesserung des Allgemeinbefindens kommt, ist bislang nicht geklärt. Es existieren in Bezug auf den Einsatz der Rife-Frequenzen keine abgesicherten Untersuchungen die ihre Wirkungsweise eindeutig belegen. 4.1.3. Einsatz und Wirkungsweise von Bioresonanzverfahren Die Informationsübertragung innerhalb eines Organismus funktioniert auf biophysikalischem und biochemischem Wege. Dabei ist der schnellere Reaktion auf biophysikalischem Weg zu erreichen, da über elektrische Potentialveränderungen an Membranen sehr viel schneller Information ausgetauscht werden kann. Alle biophysikalischen Reaktionen lösen wiederum biochemische Signale aus, wenn z.B. bestimmte Botenstoffe ins Blut abgegeben werden. Beide Systeme tragen zum Funktionieren eines Organismus bei und sind in ihren Gewichtung gleichberechtigt. 70 Deshalb ist es um so erstaunlicher, dass unserer Gesellschaft in erster Linie auf biochemischem Wege versucht wird, Systemstörungen wie Krankheiten, mit Hilfe von Medikamenten zu beheben. Es ist außer Frage, dass viele Krankheiten nur mit pharmakologischen Mitteln zu behandeln sind. Z.B. sind Diabetiker von der Einnahme von Insulin abhängig und viele akute Entzündungen können nur durch die Einnahme von entsprechenden Medikamenten therapiert werden. Trotzdem sind wir eine Gesellschaft mit einem hohen Medikamentenkonsum. Auch wenn fast alle Medikamente mehr oder weniger unangenehme Nebenwirkungen haben, wird bedenkenlos, selbst bei geringen Befindlichkeitsstörungen, darauf zurückgegriffen. Bioresonanzverfahren, die auf an der biophysikalischen Systemregulierung angreifen sind in unserer Gesellschaft bislang noch unterrepräsentiert. Dies liegt sicherlich darin begründet, dass bei uns die Pharmaindustrie einen großen Einfluss auf das Gesundheitswesen hat. In Russland und den ehemaligen GUS Staaten sind Therapien mit Hilfe von elektromagnetischen Feldern unterschiedlicher Frequenzen auch im klinischen Alltag weit verbreitet. Diese Verfahren werden erst langsam in den westlichen Ländern akzeptiert. Dabei sind bei der Anwendung von biophysikalischen Systemregulierungsverfahren, wie Bioresonanz, bislang von noch keinen gravierenden Nebenwirkungen (bis auf eine leichte Erstverschlimmerungen nach der ersten Therapie) festgestellt worden. Auf der anderen Seite wäre es natürlich fatal, biophysikalische Regulationsverfahren als Allheilmittel darzustellen. Ähnlich wie bei den unterschiedlichen pharmakologischen Therapie dürfen keine "Heilversprechungen" gemacht werden. Jeder Organismus reagiert auf die verschiedenen Verfahren unterschiedlich und es ist grob fahrlässig hier übersteigerte Hoffnungen zu schüren. Biophysikalische Verfahren wie die Bioresonanz können allerdings auf Dauer eine sinnvolle Ergänzung zu etablierten Verfahren, ähnlich der Akupunktur und der Homöopathie darstellen, da auch hier die Nebenwirkungen bei sachgemäßer Anwendung der Systeme, vergleichbar gering sind. Um dies zu verdeutlichen nachfolgend eine kleine Studie, die mit den hier vorgestellten Bioresonanzsystemen (QXCI / SCIO und Life System) mit insgesamt 34 Probanden durchgeführt wurde, die mehr als zweimal mit einem der Bioresonanzsystemen behandelt wurden. Aufgrund der geringen Anzahl der Probanden besteht natürlich kein Anspruch auf eine repräsentative Normalverteilung durch die Bevölkerung, aber es sind doch Tendenzen zu erkennen, das Bioresonanzverfahren regulierend auf Funktionsstörungen einwirken können. Die 22 weiblichen und 12 männlichen Probanden konnten in folgende Altersklassen unterteilt werden: männlich weiblich 0-5 Jahre 3 0 10-15 Jahre 1 1 15-20 Jahre 1 2 20-30 Jahre 0 1 bis 50 Jahre 3 9 bis 70 Jahre 3 7 über 70 Jahre 1 2 insgesamt 12 22 71 n = 34 Es wurden in den verschiedenen Altersklassen unterschiedliche Funktionsstörungen behandelt, die dem jeweiligen Alter der Probanden entsprach. Auswertungskriterium war: - für eine Verschlechterung des Befindens, 0 für kein Unterschied zu vorher und + für eine Verbesserung des Befindens. Bei keinem der Probanden trat eine Verschlechterung ein ( - = 0). Bei 3 (entspricht 25%) der männlichen Probanden konnte keine (0 = 3), und bei 9 (entspricht 75%) eine Verbesserung (+ = 9) der Symptome festgestellt werden. Bei 7 (entspricht 32%) der weiblichen Probanden konnte keine (0 = 7), und bei 15 (entspricht 68%) eine Verbesserung der Symptomatik festgestellt werden. männl. 5 4 3 ``0 2 `+ 1 0 0-5 Jahre 10-15 Jahre 15-20 Jahre 20-30 Jahre `-50 Jahre `-70 Jahre über 70 Jahre Alter Abb. 37 : Auswertung der männlichen Probanden (n = 12) nach Altersklassen (heller Balken = keine Veränderung, dunkler Balken = Verbesserung) weibl. 5 4 3 ``0 2 `+ 1 0 0-5 Jahre 10-15 Jahre 15-20 Jahre 20-30 Jahre `-50 Jahre `-70 Jahre über 70 Jahre Alter Abb. 38 : Auswertung der weiblichen Probanden (n = 22) nach Altersklassen (heller Balken = keine Veränderung, dunkler Balken = Verbesserung) 72 Bei der Gesamtbetrachtung ergibt sich daraus, dass bei insgesamt 10 Probanden keine Verbesserung, aber auch keine Verschlechterung der Symptomatik auftrat und bei insgesamt 24 Probanden eine Verbesserung zu verzeichnen war. Prozentual bedeutet das, dass bei 71% der Probanden eine Verbesserung eingetreten ist. 100 90 80 70 60 männl. weibl. 50 insges 40 30 20 10 0 verschl. gleich verbess. Abb. 39: Gesamtbetrachtung der prozentualen Verteilung mit n = 34 Diese kleine Untersuchung sagt natürlich nichts darüber aus, wie nachhaltig die Behandlung mit den Bioresonanzsystemen ist. Da es noch keine groß angelegten Studien darüber existiert, sind hierüber keinerlei Aussagen zulässig. Auch über die Häufigkeit der Behandlung gibt es nur die Angaben der jeweiligen Hersteller, dass in den meisten Fällen, mindestens eine Woche zwischen den Behandlungen liegen und das Regime einer Behandlung maximal 1 - 1,5 Stunden dauern sollte. Trotzdem bieten die biophysikalischen Regulationssysteme, wie die Bioresonanzverfahren neue Möglichkeiten in der Funktionsdiagnostik und in der Regulation von Funktionsstörungen. Dabei muss lediglich bedacht werden, dass der Organismus als energetisch offenes System reagiert und dementsprechend muss die Analyse der Verfahren durchgeführt werden. Wünschenswert für zukünftige Untersuchungen wäre sicher die enge Zusammenarbeit mit der schulmedizinischen Zustandsdiagnostik, um Ergebnisse der Bioresonanztherapien zu verifizieren und zu falsifizieren. Auch ist es unumgänglich für die Anwender dieser Verfahren, die Ergebnisse zu protokollieren damit eine Möglichkeit der statistischen Auswertung zu schaffen. 73 4.2. Vergleichbare Verfahren: Akupunktur und Homöopathie Während bei Bioresonanzverfahren versucht wird über die Applikation von elektromagnetischen Feldern unterschiedlicher Frequenzen Regulationsprozesse eines Organismus zu unterstützen oder in Gang zu setzen, wird bei der Akupunktur über die Stimulation bestimmter Punkte regulierend eingegriffen. Wie in Kapitel 2.3.2.2. schon dargestellt, können die Akupunkturpunkte als "Fenster" zur Grundsubstanz bzw. zur extrazellulären Matrix (nach Heine) verstanden werden. Durch die Anregung der jeweiligen Punkte, kann regulierend in den Organismus eingegriffen werden. Besonders geeignet ist die Akupunktur zur Linderung von Schmerzen sowie zur Behandlung von gestörten Organfunktionen, wie Verstopfung, Magenkrämpfen, Schlafstörungen, Kopfschmerzen oder Schwindel. Zerstörtes Gewebe, durch Verletzungen oder Krebs, lässt sich durch Akupunktur nicht heilen. Jedoch können Begleitsymptome wie Schmerzen oder Übelkeit durch Akupunktur gelindert werden und damit dem betroffenen Organismus bessere Möglichkeiten zur Selbstregulation liefern. Auch in der Homöopathie wird versucht, die Selbstheilungskräfte zu stärken und zu aktivieren. Mit dem Grundsatz der Homöopathie "Similia similibus curantur" - Ähnliches wird mit Ähnlichem geheilt, setzte ihr Begründer Samuel Hahnemann (1755- 1843) homöopathische Mittel ein, die bei gesunden Versuchspersonen die gleichen Krankheitszeichen erzeugen konnten, wie bei Patienten mit den jeweiligen Beschwerden. Selbst wenn die Präparate extrem hoch verdünnt sind, erfolgt eine Reaktion nach der Einnahme. Demnach löst ein extrem schwacher, aber spezifischer biochemischer Reiz eine starke Reaktion im Organismus aus, regt so die Körperfunktionen an und aktiviert damit die Selbstheilungskräfte. Im Gegensatz zu der Bioresonanztherapie, wo mit elektromagnetischen Feldern ein biophysikalischer Informationsinput erfolgt, setzt die Homöopathie auf den biochemischen Informationsinput um regulierend einzugreifen. Die Regulation und die Betrachtung des Organismus als energetisch offene Funktionseinheit haben die Homöopathie, die Akupunktur und auch die Bioresonanzverfahren gemeinsam. Auch wenn diese Verfahren mit unterschiedlichen Inputsystemen arbeiten (Homöopathie = biochemisch, Akupunktur = Schmerzreiz und Bioresonanz = biophysikalisch) sind sie doch von der Grundüberlegung vergleichbar, die einen Organismus als individuell eingestellte Funktionseinheit verstehen, dessen Krankheit oder Symptomatik aus einer Störung von regulatorischen Vorgängen hervorgeht. 5. Bioresonanz- Systeme 5.1. Life System 5.2. Magnetmatten 74 6. Anhang Quellennachweis der im Skript verwendeten Abbildungen: Biologischer Teil: Der Körper des Menschen. Einführung in Bau und Funktion, von Adolf Faller (Neu bearbeitet von Michael Schünke) 1999 Georg Thieme Verlag Stuttgart, New York (13. Auflage) Zoologie von Rüdiger Wehner und Walter Gehring 1990 Georg Thieme Verlag Stuttgart, New York (22. Auflage). Natur. Das große Buch des Allgemeinwissens. 1996 Verlag Das Beste Stuttgart, Zürich, Wien. Physiologie des Menschen von R.F. Schmidt und G. Thews 1980 Springer-Verlag Berlin, Heidelberg, New York (20. Auflage). Die unbekannte Seite des Gehirns von R. Douglas Fields aus Spektrum der Wissenschaft September 2004 (S. 46 - 56). Das System der Grundregulation von Alfred Pischinger (Neu bearbeitet von Hartmut Heine) 1998 Karl F. Haug Verlag Heidelberg (9. Auflage). Lehrbuch der biologischen Medizin von Hartmut Heine 1997 Hippokrates Verlag Stuttgart (2. Auflage). Physikalischer Teil: Strahlen, Wellen, Felder von Norbert Leitgeb 1990 Georg Thieme Verlag Stuttgart, New York dtv- Atlas zur Physik Band 2 (Elektrizität, Magnetismus, Festkörper) von Hans Breuer 1988 Deutscher Taschenbuch Verlag Physik für Mediziner von Harten (unter Mitarbeit von H. Nägerl, J. Schmidt, H.-D. Schulte) 1980 Springer Verlag, Berlin, Heidelberg, New York (4. Auflage) 75