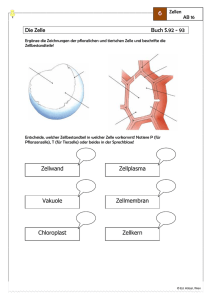
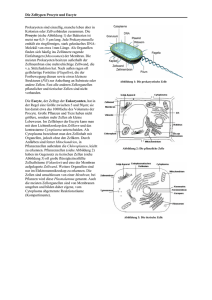
Teil: Cytologie - lern-soft
Werbung

Teil: A B E C D 1 BK_FOS_Biologie_FOH_EHW.doc 2 Seite - 1 - 3 (c,p)'98 lsp: dre (c,p)2007-2008 lsp: dre Legende: mit diesem Symbol werden zusätzliche Hinweise, Tips und weiterführende Ideen gekennzeichnet Nutzungsbestimmungen / Bemerkungen zur Verwendung durch Dritte: (1) Dieses Skript (Werk) ist zur freien Nutzung in der angebotenen Form durch den Anbieter (lern-soft-projekt) bereitgestellt. Es kann unter Angabe der Quelle und / oder des Verfassers gedruckt, vervielfältigt oder in elektronischer Form veröffentlicht werden. (2) Das Weglassen von Abschnitten oder Teilen (z.B. Aufgaben und Lösungen) in Teildrucken ist möglich und sinnvoll (Konzentration auf die eigenen Unterrichtsziele, -inhalte und -methoden). Bei angemessen großen Auszügen gehört das vollständige Inhaltsverzeichnis und die Angabe einer Bezugsquelle für das Originalwerk zum Pflichtteil. (3) Ein Verkauf in jedweder Form ist ausgeschlossen. Der Aufwand für Kopierleistungen, Datenträger oder den (einfachen) Download usw. ist davon unberührt. (4) Änderungswünsche werden gerne entgegen genommen. Ergänzungen, Arbeitsblätter, Aufgaben und Lösungen mit eigener Autorenschaft sind möglich und werden bei konzeptioneller Passung eingearbeitet. Die Teile sind entsprechend der Autorenschaft zu kennzeichnen. Jedes Teil behält die Urheberrechte seiner Autorenschaft bei. (5) Zusammenstellungen, die von diesem Skript - über Zitate hinausgehende - Bestandteile enthalten, müssen verpflichtend wieder gleichwertigen Nutzungsbestimmungen unterliegen. (6) Diese Nutzungsbestimmungen gehören zu diesem Werk. (7) Der Autor behält sich das Recht vor, diese Bestimmungen zu ändern. (8) Andere Urheberrechte bleiben von diesen Bestimmungen unberührt. Rechte Anderer: Viele der verwendeten Bilder unterliegen verschiedensten freien Lizenzen. Nach meinen Recherchen sollten alle genutzten Bilder zu einer der nachfolgenden freien Lizenzen gehören. Unabhängig von den Vorgaben der einzelnen Lizenzen sind zu jedem extern entstandenen Objekt die Quelle, und wenn bekannt, der Autor / Rechteinhaber angegeben. Zum Gemeingut erklärte Graphiken oder Fotos (u.a.). Viele der verwenpublic domain (pd) deten Bilder entstammen Webseiten / Quellen US-amerikanischer Einrichtungen, die im Regierungsauftrag mit öffentlichen Mitteln finanziert wurden und darüber rechtlich (USA) zum Gemeingut wurden. Andere kreative Leistungen wurden ohne Einschränkungen von den Urhebern freigegeben. gnu free document licence (GFDL; gnu fdl) creative commens (cc) od. neu … Namensnennung … nichtkommerziell … in der gleichen Form … unter gleichen Bedingungen Die meisten verwendeten Lizenzen schließen eine kommerzielle (Weiter-)Nutzung aus! Bemerkungen zur Rechtschreibung: Dieses Skript folgt nicht zwangsläufig der neuen ODER alten deutschen Rechtschreibung. Vielmehr wird vom Recht auf künstlerische Freiheit, der Freiheit der Sprache und von der Autokorrektur des Textverarbeitungsprogramms microsoft ® WORD ® Gebrauch gemacht. Für Hinweise auf echte Fehler ist der Autor immer dankbar. -2- (c,p) 2008 lsp: dre Inhaltsverzeichnis: Seite [ ! ] Vorbemerkungen............................................................................................................6 [ 0 ] Arbeitstechniken ...........................................................................................................8 1. intellektuelle Tätigkeiten / Operationen.............................................................................8 1.1. erfassende Tätigkeiten ..............................................................................................9 1.2. strukturierende / struktur-orientierte Tätigkeiten ......................................................11 1.3. didaktisch orientierte Tätigkeiten .............................................................................15 1.4. logisch orientierte Tätigkeiten ..................................................................................17 1.5. wertende Tätigkeiten ...............................................................................................20 1.6. mehr praktisch orientierte Tätigkeiten:.....................................................................20 1.7. moderne Tätigkeiten ................................................................................................22 1.8. Lesetechniken..........................................................................................................24 2. wissenschaftliche Tätigkeiten .........................................................................................25 3. die experimentelle Methode............................................................................................27 4. Umgang mit Medien (Medienkompetenz) .......................................................................28 4.2. Lesemethoden / Lesekompetenzen.........................................................................34 5. Aufgaben und Probleme, Arbeits- und Lerntechniken ....................................................36 5.1. Lösen von Aufgaben mittels Algorithmen ................................................................36 5.2. Problemlösestrategien .............................................................................................37 5.3. Lerntechniken ..........................................................................................................40 5.3.x. 20/80-Prozent-Regel / PARETO-Prinzip ...........................................................40 6. Beispiele / Arbeitmaterialien ...........................................................................................41 6.1. Analyse einer Anekdote ...........................................................................................41 6.2. Analyse und Bewertung eines Fachtextes...............................................................41 6.3. Interpretieren und Auswerten von Diagrammen ......................................................43 6.3.x. versteckte Daten ...............................................................................................43 [ A ] Wissenschaft Biologie ...............................................................................................45 1. die wichtigsten Zweige der Biologie................................................................................46 [ B ] Was ist eigentlich Leben?..........................................................................................47 2. Gibt es Leben auf anderen Planeten? ............................................................................49 [ C ] Einteilung der Organismen........................................................................................51 x.y. Taxonomie................................................................................................................51 x.y.z. weitere taxonomische Begriffe oder Ebenen.....................................................53 x.z. ein taxonomisches System.......................................................................................54 1. Bakterien und Blaualgen (Bacteria) ................................................................................55 2. Protoctisten (Protoctista) ................................................................................................55 3. Pilze ................................................................................................................................56 4. Tiere................................................................................................................................56 5. Pflanzen..........................................................................................................................56 [ D ] Die Zelle (Zytologie)....................................................................................................57 1. Bau der Zelle ..................................................................................................................57 1.1. Makroskopischer und lichtmikroskopischer Bau der Zellen.....................................57 1.2. elektronenmikroskopischer Bau der Zellen..............................................................60 2. Bau und Funktion der Zellbestandteile ...........................................................................63 2.1. Zellmembran, Plasmalemma ...................................................................................65 2.1.1. Transportvorgänge an Biomembranen .............................................................68 2.1.2. Rezeptionsvorgänge an Biomembranen...........................................................74 2.2. Zellwand ..................................................................................................................76 2.2.1. Mittellamelle ......................................................................................................76 2.3. Cytoplasma ..............................................................................................................77 -3- (c,p) 2008 lsp: dre 2.4. Kernäquivalent / Zellkern ........................................................................................ 79 2.5. Endoplasmatisches Retikulum, GOLGI-Apparat und Visikel .................................. 81 2.5.1. Endoplasmatisches Retikulum ......................................................................... 81 2.5.2. GOLGI-Apparat ................................................................................................ 81 2.5.3. weitere vesikuläre Strukturen........................................................................... 82 2.6. Tubuläre Strukturen ................................................................................................ 84 2.6.1. Zellskelett ......................................................................................................... 84 2.6.2. Mikrotubulli ....................................................................................................... 84 2.6.3. Centriolen und Spindelapparat......................................................................... 86 2.6.4. Cilien ................................................................................................................ 87 2.6.4. Geißeln............................................................................................................. 88 2.6.5. Actin-Filamente ................................................................................................ 90 2.6.6. Intermediär-Filamente ...................................................................................... 90 2.7. Zellorganellen.......................................................................................................... 91 2.7.1. Mitochondrien................................................................................................... 91 2.7.2. Chloroplasten ................................................................................................... 92 2.7.4. Leukoplasten.................................................................................................... 94 2.7.3. Chromoplasten................................................................................................. 94 2.8. Vakuole ................................................................................................................... 95 2.9. paraplasmatische (ergastische) Strukturen............................................................. 98 2.9.1. Lipid-Tröpfchen ................................................................................................ 98 2.9.2. Stärkekörner..................................................................................................... 98 2.9.3. Pigmentgranula ................................................................................................ 99 2.9.4. Sekretgranula................................................................................................... 99 2.10. kristalline und abiotische Zellbestandteile........................................................... 100 2.10.1. Fett-Tropfen ................................................................................................. 100 2.10.2. Kristalle ........................................................................................................ 100 [ E ] Stoffwechsel der Zelle (Zellphysiologie)................................................................ 101 0. Einteilung / Grundprinzipien der Stoffwechselvorgänge .............................................. 101 1. Biokatalyse und Metabolismus .................................................................................... 103 1.1. Enzyme und enzymatische Reaktionen ................................................................ 106 1.1.1. Abhängigkeit der Enzymaktivität .................................................................... 112 1.1.2. Regulation der Enzymaktivität (Modulation der Enzymaktivität) .................... 117 1.2. Transport von Energie und Reduktionsäquivalenten ............................................ 122 2. Dissimilations-Vorgänge .............................................................................................. 129 2.0. Geschichte der Dissimilation................................................................................. 131 2.1. anaerobe Dissimilation (Gärungen) ...................................................................... 132 2.1.1. Glycolyse........................................................................................................ 133 2.1.2. nach der Glycolyse ablaufende anaerobe Vorgänge ..................................... 139 2.2. aerobe Dissimilation (Zellatmung)......................................................................... 145 2.2.1. Zitrat-Zyklus ................................................................................................... 146 2.2.2. Atmungskette ................................................................................................. 151 3. Assimilations-Vorgänge ............................................................................................... 156 3.1. heterotrophe Assimilation...................................................................................... 157 3.1.1. heterotrophe Assimilation (auf zellulärer Ebene) ........................................... 158 3.1.2. heterotrophe Assimilation (auf Organismen-Ebene) ...................................... 159 3.1.3. heterotrophe Assimilation (auf Organ-Ebene)................................................ 165 3.2. autotrophe Assimilation......................................................................................... 166 3.2.1. Vorläufer der Photosynthese.......................................................................... 168 3.2.2. Photosynthese ............................................................................................... 169 3.2.3. Chemosynthese ............................................................................................. 193 [ F ] Physiologie der Nervenzelle (Neurophysiologie) .................................................. 195 [ G ] Verhalten von Organismen (Verhaltenslehre)....................................................... 198 [ H ] Organismen in der Umwelt (Ökologie)................................................................... 199 x.y. Die Gaia-Theorie ................................................................................................... 202 -4- (c,p) 2008 lsp: dre [ I ] Entwicklung der Organismen (Vererbung und Evolution) .....................................204 0. Vorbemerkungen ..........................................................................................................204 1. Individualentwicklung....................................................................................................205 2. Entwicklung von Populationen......................................................................................206 3. Entwicklung von (neuen) Arten.....................................................................................207 4. Entwicklung von Merkmalen .........................................................................................208 4.x. Das egoistische Gen ..............................................................................................208 4.x. Das Handicap-Prinzip ............................................................................................208 6. Historie der irdischen Evolution ....................................................................................210 6.1. Evolution vor der Entstehung der Erde ..................................................................211 6.2. Evolution vor der Entstehung des Lebens .............................................................214 6.3. Die Entstehung des Lebens...................................................................................214 6.4. Die Entwicklung des Lebens auf der Erde .............................................................214 6.4.1. Vom Einzeller zum Mehrzeller ........................................................................215 6.x. Die serielle Endosymbiontentheorie (SET) ............................................................215 6.z. Die Entstehung des Sex ........................................................................................217 6.x. Der Übergang vom Wasser zum Land...................................................................218 7. Vererbung und Genetik.................................................................................................220 7.1. Vererbung auf Organismen- und Zell-Ebene .........................................................221 7.2. Das Wirken MENDELs...........................................................................................224 Zusammenfassung (MENDELsche Regeln): ............................................................232 7.3. Die Weiterentwicklung der MENDELschen Vererbungslehre ................................234 7.4. Weitergabe und Verteilung der Erbinformation......................................................239 7.5. Die moderne klassische Genetik ...........................................................................243 7.5.1. Vererbung des Geschlechts beim Menschen .................................................248 7.6. Speicherung der Erbinformation ............................................................................250 7.7. Realisierung der Erbinformationen ........................................................................259 7.8. Veränderung der Erbinformation............................................................................271 7.3. moderne genetische Methoden, Theorien und Erkenntnisse ................................282 7.3.x. Klonierung .......................................................................................................282 7.3.x. Auf der Suche nach Adam und Eva ................................................................282 [ J ] .......................................................................................................................................283 [ K ].......................................................................................................................................284 [ L ] .......................................................................................................................................285 [ M ] Der Mensch ...............................................................................................................286 [ Z ] Literatur und Quellen:...............................................................................................287 -5- (c,p) 2008 lsp: dre [ D ] Die Zelle (Zytologie) Die Zelle als Struktur- und Funktionseinheit der Lebewesen 1. Bau der Zelle Die Zelle ist das Grundelement aller Lebewesen. Zellen können zwischen mehrere Meter lang bis wenige Mikrometer (µm = 10-6 m) groß sein. Typische Zellen werden mit 0,3 µm bis 0,1 mm ausgemessen. Der äußere Bau ist meist unspektakulär. Mit den Augen kann man direkt kaum genauere Strukturen ausmachen. Innere Strukturen sind mit bloßem Augen fast gar nicht zu erkennen. Erst mit der Erfindung von optischen Instrumenten (Lupen und Mikroskope) kam es zu einer stürmischen Entwicklung der Zellbiologie (Zellenlehre, Zytologie, Cytologie; cytos = Zelle; logos = Wissen, Lehre). Der Begriff Zelle leitet sich von cella und cellula ab, was Keller bzw. Kämmerchen bedeutet. Die ersten Zellen wurden 1665 von Robert HOOKE bei der Untersuchung von feinen Schnitten (Spänen) vom Flaschenkork entdeckt. 1.1. Makroskopischer und lichtmikroskopischer Bau der Zellen Die ersten Licht-Mikroskope waren eher gute Lupen. Bei Vergrößerungen um das 50–fache konnte man gerade größere Zellen und Mikroorganismen (z.B. Pantoffeltierchen (A ) Parameceum spec.) beobachten. Mit heutigen Licht-Mikroskopen werden Auflösungen bis zum 1000fachen erzielt. Objekte bis zu einer Kleine von 0,4 µm sind dann noch scharf abbildbar. Der typische Aufbau eines Mikroskops ist in der nebenstehenden Abbildung ersichtlich. Das notwendige Licht wird über Spiegel (F) oder eine Lampe an der gleichen Stelle über die Beleutungsoptik (D) geleitet. Auf dem Objekttisch befindet sich das Objekt (C), welches bei der Durchlichtmikroskopie durchsichtig sein muss. Das Bild wird über Objektiv (B) und Okular (A) vergrößert. Bei Auflichtmikroskopen erfolgt die Beleuchtung von schräg oben. Mit solchen Geräten lassen sich dann vorrangig Oberflächen beobachten. Q: de.wikipedia.org (Tomia) - 57 - (c,p) 2008 lsp: dre Mit Licht-Mikroskopen beobachtbare Teile in Zellen lassen sich in folgenden schematischen Abbildungen zusammenfassen. Heute unterscheidet man zwei grundsätzlich verschiedene Grund-Zelltypen, die sich deutlich im Bau unterscheiden: Prokarionten-Zelle, Prokaryoten-Zelle, Procyte (ohne Zellkern; (r+) Procaryota; (r ) Bacteria (Bakterien + Blaualgen)) Q: de.wikipedia.org (LadyofHats) Eukarionten-Zelle, Eukaryoten-Zelle, Eucyte (mit Zellkern; (r+) Eukaryota) Q: www.zum.de (mallig) - 58 - (c,p) 2008 lsp: dre Eukarionten-Zellen (Eucyten) lassen sich weiter unterscheiden. Die Unterscheidung korrelliert mit den großen Gruppen (Reichen), die auf Eucyten basieren. Pflanzen-Zelle ((R ) Pflanzen; (r ) regnum plantae) Tier-Zelle ((R ) Tiere; (r ) regnum animalia) Auch die zelluläre Grundeinheit des vierten Organismen-Reiches (drittes eucytisches Reich) unterscheidet sich von den Tier- und Pflanzen-Zellen: Pilz-Zelle (Mycel) ((R ) Pilze, (r ) regnum fungi) - 59 - (c,p) 2008 lsp: dre 1.2. elektronenmikroskopischer Bau der Zellen Das Problem der Licht-Mikroskope ist die relativ lange Wellenlänge (normal 380 – 780 nm) des verwendeten Lichts für die Untersuchung. Nur Objekte mit einer Größe bis ungefähr der Hälfte der Wellenlänge können damit abgebildet werden. Diese Gesetzmäßigkeit gilt auch für die Elektronen-Mikroskope (EM). Nur ist hier die Wellenlänge der verwendeten Elektronenstrahlen wesentlich kleiner (runter bis 1 nm). Damit lassen sich Objekte bis zur Größe von 0,05 nm beobachten. Mit den neuesten TunnelElektronen-Mikroskopen kann man sogar die Atome selbst darstellen. Diese Mikroskope funktionieren aber nicht über Strahlung, sondern es wird eine feinste Spitze über das Material bewegt und der zwischen der Spitze und dem Untersuchungsmaterial fließende (Tunnel-)Strom gemessen und graphisch umgesetzt. Q: dk.wikipedia.org (KristianMolhave) [zum Vergleich: CRT .. Fernsehbildröhre] Nach dem Bauprinzip unterscheidet man z.B. Transmissions(TEM) und Raster-Elektronenmikroskope (REM, auch: SEM Q: de.wikipedia.org (Stahlkocher) Scanning electron microscope). Durch die gute Auflösung moderner Elektronen-Mikroskope sind viele neue Erkenntnisse über den Bau der Zelle und seiner Bestandteile bekannt geworden. Praktisch wird bei der Betrachtung von Bau und Funktion der einzelnen Bestandteile nicht mehr zwischen licht- oder elektronenmikroskopischer Erkennbarkeit unterschieden. Alle Beobachtungsmöglichkeiten werden genutzt, um ein möglichst umfassendes Bild zu erhalten. Der Bau der Zelle (für die Schul-Biologie) erweitert sich um: • • • • • Endoplasmatisches Retikulum (ER) GOLGI-Apparat (Dictyosom) Lipidkörperchen (Oleosomen) Lysosomen … - 60 - (c,p) 2008 lsp: dre Pflanzen-Zelle ((R ) Pflanzen; (r ) regnum plantae) Q: de.wikipedia.org () Tier-Zelle ((R ) Tiere; (r ) regnum animalia) Q: de.wikipedia.org () Pilz-Zelle (Mycel) ((R ) Pilze, (r ) regnum fungi) - 61 - (c,p) 2008 lsp: dre Exkurs: erweiterter Vergleich (Unterschiede) zwischen Procyte und Eucyte Merkmal Prokaryont / Procyte normale Größe Tendenz zur Vielzelligkeit Zelldifferenzierung Generationsdauer und Eukaryont / Eucyte 0,3 – 2,5 µm 2 – 20 (- 300) µm keine ausgeprägt 20 min Zellzyklus mehrere Stunden G1, S, G2, M Zellteilung Septenbildung / Spaltung Mitose und Cytokinese Organisation des Genoms 1 zirkuläres Molekül DNA-Menge 7*10-4 – 1*10-2 pg mehrere lineare (Chromosomen) 2*10-2 – 100 pg kaum überwiegend selten vorhanden genetische Rekombination durch Konjugation durch Meiose und Syngamie Nucleosomen (Histone) nein ja nein ja 70 S (30 S + 50 S) 80 S (40 S + 60 S) ja nein wenig, selten nein ja vielseitig Membranfluss, Exo- u. Endozytose semiautonome Organellen nein ja nein Mitochondrien, Chloroplasten Gasvakuolen Halobakterien, Cyanobakterien nein nein Mikrotubuli, Dynein-System, Geißeln (Cilien) Extrazelluläre rotierende Flagellen nein ja ja nein Fettsäure-Synthase-Komplex meist als Einzelenzyme 3fach ungesättigte Fettsäuren selten als 1 – 2 multifunktionale Polypeptide ja als Membranlipide - Sterole - Cardiolipin Peptidoglykan als Wandsubstanz selten ja häufig häufig nur in innerer Mitoch.-mem. nein Anaerobiose häufig nur Hefe N-Fixierung über Nitrogenase häufig nein Chemolithotrophie vielfältig nein nichtkodierende der DANN Introns Abschnitte separate RNA-Polymerasen mRNA, rRNA u. tRNA Größe der Ribosomen auf für Inhibition der Translation - mit Chloramphinicol - mit Cycloheximid intrazelluläre Kompartmentierung Actomyosinsystem Moleküle ja nach /4/ - 62 - (c,p) 2008 lsp: dre Grundplasma Chromatin Kernkörperchen (Nucleolus) Kerngrundplasma Kernplasma (Karyoplasma) Membransysteme Zytoplasma Zellkern (Nucleus) Protoplasma 2. Bau und Funktion der Zellbestandteile - 63 - (c,p) 2008 lsp: dre - 64 - (c,p) 2008 lsp: dre 2.1. Zellmembran, Plasmalemma Die Abgrenzung der lebenden Einheit (Cytoplasma, Protoplasma) von der Umgebung ist eine elementare Notwendigkeit. Diese Aufgabe übernehmen die Zellmembranen. Ihre Aufgaben und Merkmale sind sehr vielgestaltig und zum Teil sogar scheinbar gegensätzlich: • • • • • Abgrenzung, Schutz Zusammenhalt des Zellinneren, Widerstand gegen Zellinnendruck (Tugor) Nahrungsaufnahme, Schadstoffabgabe Informationsaufnahme (Reizbarkeit, Signalaufnahme) Beweglichkeit / Formveränderung Die stoffliche Zusammensetzung der Zellmembran konnte schon frühzeitig mit chemischen Methoden geklärt werden. So sind neben fettähnlichen Stoffen (Lipoide) vor allem verschiedenste Proteine enthalten. Weiterhin wurden Polysaccharide und Kombinationen zwischen den genannten Stoffen (Glycoside, Glykolipide, Glykoproteine) gefunden. Das Grundelement der Biomembranen sind verschiedenste Phospholipide. Sie bestehen – ähnlich wie die Fette (Lipide) – aus dem zentralgelagerten Glycerol (Glyzerin) sowie meist zwei angeesterten Fettsäuren und einem (ebenfalls angeesterten) Phosphatrest. Dadurch ergeben sich in einem Molekül extrem unterschiedliche Stoffeigenschaften. Die Seite mit dem Phosphat-Rest und auch der Glycerol-Rest sind wasserlöslich (hydrophil, wasserfreundlich, lipophob, fettfeindlich). Dagegen ist die Fettsäure-Seite fettlöslich (lipophil, fettfreundlich) und nicht wasserlöslich (hydrophob, wasserfeindlich). Die beiden Fettsäuren lagern sich wegen der starken VAN-DER-WAALS-Kräfte zu einer Seite hin. Aus den bekannten Stoffeigenschaften und den elektronenmikroskopischen Bildern wurden verschiedene Modelle entwickelt. Diese müssen vor allem die oben genannten Membraneigenschaften und –funktionen erklären können. Die Grundstruktur der Membranen ist aus den Lösungseigenschaften schnell abgeleitet. Beim Zusammenlagern von mehreren Molekülen ordnen sich diese immer so an, dass sich gleichlösliche Teile zueinander gesellen. Es bilden sich vor allem an Phasengrenzen Schichten / Ebenen. Zwischen den Fettsäure-Resten sind starke VAN-DER-WAALS-Kräfte wirksam. An Glycerol- und Phosphat-Rest wirken recht starke polare Kräfte. Ein Verschieben aus der Ebene ist nur mit sehr großen Kraftaufwendungen möglich. In der Ebene selbst ist die Beweglichkeit der Lipoide wesentlich besser, da keine Kraftsprünge (polar - unpolar) überwunden werden müssen. Der Effekt wird noch stärker, wenn sich die Phospholipide in wässrigen (polar) oder gemischten (polar und unpolar) Umgebungen befinden. - 65 - (c,p) 2008 lsp: dre Andere – in der Membran vorkommende – Lipoide sind den Phospholipiden sehr ähnlich. Statt der Phosphorsäure ist ein anderer Rest angeestert. Allen Resten gemeinsam ist ihre gute Wasserlöslichkeit. Sie können einzelne Phospholipide dementsprechend auch jederzeit in der Membran ersetzen. Das Cholesterol (Phosphatidylcholin, Cholesterin) ist ein solcher – vom Namen recht bekannter – Membranbaustein. In den Biomembranen hat Cholesterol vor allem eine Kit-Funktion. Gib man bei einem Experiment Phospholipide auf eine wässrige Lösung, dann bilden die Phospholipide eine geordnete Schicht. Bei einer Durchmischung entstehen Doppelschichten (Bilayer) und kugelförmige Objekte (Bläschen), die auch Micellen (Mizellen) genannt werden. Sind Fette oder fettähnliche (unpolare) Stoffe in Lösung, dann ordnen sich diese innerhalb der Micelle an. Prinzipiell können Micellen auch doppelwandig sein. Die Doppelschichtigkeit der Membranen konnten GORTER und GRENDEL schon 1925 nachweisen. Sie stellten fest, dass rote Blutkörperchen ungefähr doppelt so viel Phospholipide enthielten, wie für die Oberfläche eigentlich notwendig wären. In diesem Grundmodell fehlt noch der beobachtete Proteinanteil. Die ersten Membranmodelle hatten noch große Probleme bei der Erklärung von Membraneigenschaften. DARNIELLI und DAVSON entwickelten 1935 das erste Modell, welches auch den Proteinanteil berücksichtigte. Ihr Sandwich-Modell konnte aber kaum den Stofftransport erklären, noch konnte später die Schichtdicke mittels der Elektronenmikroskopie nachgewiesen werden. - 66 - (c,p) 2008 lsp: dre Erst durch elektronenmikroskopische Aufnahmen erkannte man den genauen Bau der Biomembranen und konnte darauf passende Modelle entwickeln. Die gesamte Struktur ist rund 8 nm dick. Im Elektronenmikroskop sind drei abgegrenzte Schichten (trilaminarer Bau) zu erkennen. Manche Elemente durchdringen die Zellmembran, andere liegen in einer der drei Schichten. Die großen "Klumpen" überragen das dreischichtige Gebilde oft sehr weit. Im Jahre 1972 stellten NICOLSON und SINGER ein wesentlich weitergefasstes Modell vor. Ihr Flüssig-Mosaik-Modell (fluid mosaic model) geht davon aus, dass Proteine sich auch in der Membran befinden können. Je nach ihren Oberflächeneigenschaften (polar und / oder unpolar) schwimmen sie in oder auf der Membran (wie Eisberge in einem See). Das gesamte Gebilde sieht aus der Fläche betrachtet, wie ein Fleckenteppich oder ein Mosaik. Die gesamte Struktur ist gut beweglich und sehr dynamisch. Man spricht von einem Membranfluss. Aus aktuellen hochaufgelösten elektronenmikroskopischen Aufnahmen und biochemischen Markierungen (mit metallorganischen, radioaktiven od. fluoressierenden Verbindungen) wissen wir, dass neben den Phospholipiden, eine Vielzahl weiterer Moleküle am Aufbau der Zellmembran beteiligt sind. So ergibt sich heute ein vielgestaltiges Bild der Biomembranen: Der Stofftransport kann z.B. über die Membranporen, die Tunnel- und Carrier-Proteine erfolgen. Die Glycolax wird für die rezeptiven Funktionen verantwortlich gemacht. Biomembranen sind beim Aufbau vieler Zellkompartmente beteiligt. Beispielhaft sei hier auf GOLGI-Apparat / Dictyosomen und Endoplasmatisches Retikulum hingewiesen. Bei allen größen Gebilden (Plastiden, Vakuole usw.) dienen sie zur äußeren Abgrenzung. Die äußere Biomembran der Zelle wird auch als Plasmalemma (Plasmamembran, Zellmembran) bezeichnet. Ein räumlichen Eindruck und einige weitere Bauelemente des Plasmalemma einer tierischen Zelle vermittelt die nachfolgende Abbildung: - 67 - (c,p) 2008 lsp: dre Q: de.wikipedia.org () 2.1.1. Transportvorgänge an Biomembranen Wie wir schon besprochen haben, ist eine der wichtigsten Aufgaben der Biomembran im Stofftransport zu suchen. Natürlich geht es nicht um die ungerichtete und freie Bewegung von irgendwelchen Stoffen. Das würde ohne Membranen viel unkomplizierter und schneller ablaufen. Beim Stofftransport an einer Biomembran geht es um zielgerichtetes, selektives und aktives Bewegen von Stoffen. Für Transportbewegungen stehen an Biomembranen prinzipiell folgende Möglichkeiten zur Verfügung: • Diffusion, Osmose (A) • Tunnelproteine (B) • passive Transportproteine (C) • aktive Transportproteine (D) • aktiver Transport an Carrier-Proteinen (E) • Endocytose (F) • Exocytose (G) Die Möglichkeiten A bis E verlaufen ohne Veränderungen der Membran – nur durch sie hindurch. Dies sind TransmemQ: de.wikipedia.org (Zoph) bran-Transporte. Bei E und F werden auch Membranabschnitte bewegt – man spricht hier von Membranverlagendem Transport. Solche Transportvorgänge sind auch mikroskopisch beobachtbar. Schauen wir uns die einzelnen Vorgänge etwas genauer an. - 68 - (c,p) 2008 lsp: dre Diffusion: Diffusion ist der freie, ungehinderte Konzentrationsausgleich eines oder mehrerer Stoffe. Sie basiert auf der BROWNschen Molekularbewegung und der allgemeinen Tendenz im Universum eine maximale Entropie (Maß für die Unordnung) zu erreichen. Wird z.B. ein Kristall einer Substanz in einem abgeschlossen Gefäß mit einem Lösungsmittel (z.B. Wasser) gebracht, dann löst sich dieser auf. In ungelöster Form (Kristall) hat die Substanz eine sehr hohe Konzentration (am Ort). Am Ende sind die Teilchen im Lösungsmittel zufällig verteilt. Die Lösung ist gleichmäßig konzentriert – es hat eine Konzentrationsausgleich statt1 gefunden. 2 3 (Eine Zusammenlagerung (Kristall) wie in der ersten Abbildung ist zwar auch möglich, aber extrem unwahrscheinlich. Dies entspricht einer sehr geringen Entropie.) Nun kann der Lösungsmittelraum durch eine Membran (od. ein ähnliches Gebilde) geteilt sein. Die Poren sein so groß, dass die gelösten Teilchen der Substanz diese passieren können. Unabhängig, ob die Substanz in fester Form (Kristall) oder in gelöster Form auf nur einer Seite bereitgestellt wird, ist es offensichtlich, dass der Konzentrationsausgleich langsamer abläuft. Hier sprechen wir von Permeation. Permeation ist eine behinderte, verlangsamte Diffusion durch eine Membran. Je weniger störenden die Membran bzw. umso größer die Poren, umso mehr nähert sich die 1 2 3 Permeation einer "normalen" Diffusion an. - 69 - (c,p) 2008 lsp: dre Osmose: Voraussetzung für eine Osmose ist eine Membran, die bestimmte Teilchen z.B. wegen ihrer Größe nicht hindurchläßt. Andersher- um können natürlich auch die Poren zu klein für bestimmte Teilchen sein. 1 2 3 Das Lösungsmittel und alle anderen (kleineren) Teilchen können die Membran frei passieren und es kommt zum Konzentrationsausgleich. Da die größeren Teilchen auf der einen Seite verbleiben, entsteht hieraus auf dieser Seite ein erhöhter Druck. Dieser entsteht dadurch, dass sich eben mehr Teilchen das gleiche Volumen teilen müssen. Es kommt zu mehr Zusammenstößen u.a. auch mit der Wand – was eben Druck ist. Der osmotische Druck ist beobachtbar und messbar. U.U. kann er so stark sein, dass Zellen usw. zerplatzen. Kann sich das Volumen verändern, dann bewirkt das Mehr an Teilchen natürlich zuerst eine Volumenzunahme. Exakterweise spricht man statt von einem Konzentrationsausgleich (bei Diffusion, Permeation und Osmose) besser von Gradientenausgleich. Gradienten sind allgemeine Unterschiede. In den besprochenen Fällen war dies immer die Konzentration. Es können aber z.B. auch Temperatur-, Dichte- oder Ladungsunterschiede in Lösungen auftreten. Auch für diese ergeben sich Gradienten-abbauende Tendenzen / Bewegungungen. Die Osmose wird gerne als biologischer Vorgang beschrieben. Dies ist nicht richtig, da die Osmose nicht an lebende Membranen oder Zellen oder ähnliches gebunden ist. Sie tritt an jeder semipermeablen Membran (lebend oder tot; natürlich oder künstlich) auf. Grundlage sind auch hier die elementaren Teilchenbewegungen (BROWNsche Molekularbewegung; Wärmebewegung). Zumeist wird in der Schule die Osmose zuerst und ausschließlich bei biologischen Sachverhalten besprochen. So entsteht der Eindruck eines biologischen Vorgangs. Seiner Natur nach ist die Osmose – wie die Diffusion auch – ein zutiefst physikalischer Vorgang. - 70 - (c,p) 2008 lsp: dre & ' ! " ) , "" "( * " + * " # " " - 1 - $ % . /0 " % 2 3. 4 , " . Exp. Nr. 1 + . 0 " 5 Lösung A Wasser Membran vollpermeabel 2 Natriumchlorid-Lösung 3 5 Cupfersulfat-Lösung (hellblau) KaliumpermanganatLösung destilliertes Wasser 6 7 10 M Lösung Glucose Glucose-Lösung (farblos) 8 9 3 M Lösung Saccarose Wasser 10 1 M Lösung Saccarose 4 6 0 + 8 7 4* " " Lösung B Natriumchlorid-Lösung (Kochsalz) permeabel für Natrium- Kaliumpermanganatchlorid und Kaliumper- Lösung (violett) manganat permeabel für A und B Magnesiumsulfat-Lösung (farblos) vollpermeabel Magnesiumsulfat-Lösung nicht permeabel für Glucose (semipermeabel) semipermeabel permeabel für Natriumchlorid undurchlässig für Zucker nicht permeabel für Glycerol undurchlässig für Zucker " (7 9 " ( +) " % 0 + . "1 ! 7 3+ ". ; . ( 8 5: " 3 5 5: " 0 + < 5: " 3 5 5: " 3 5 = ( 0 + ! 7 " % 1 ( <0 + < < <0 + < <0 + < " Glucose-Lösung 1 M Lösung Glucose Natriumchlorid-Lösung 3 M Glucose-Lösung Glycerol 3 M Glucose-Lösung 7 8 : " 2 * ) 5 %: % < < + ( + (7 0 > - 71 - (c,p) 2008 lsp: dre Transport an Tunnelproteinen: Viele Moleküle sind viel zu groß, um einfach durch die Zellmembran durchzudiffundieren. Außerdem würden sie zumeist etweder im polaren Teil oder noch wahrscheinlicher im unpolaren Teil nicht gelöst werden können und damit dort "hängen" bleiben. Ein weiteres "Problem" der Zelle ist, dass sie natürlich nicht alle Stoffe braucht. Sie "möchte" die Stoffe selektieren. Mittels Tunnenproteien hat die Natur eine sehr effektive Lösung für die erwähnten Probleme gefunden. Tunnenproteine sind integrale Eiweiße mit einer zentralen "Röhre". Durch diese räumliche Struktur (Tertiär- und Quartärstruktur-Elemente des Proteins) wird der Stoff geleitet. Der Transport erfolgt zumeist wesentlich schneller, als durch normale Teilchenbewegung. Deshalb spricht man auch von erleichterter Diffusion. Viele Tunnelproteine besitzen an der "Einlaßstelle" zumeist eine Stelle, die den zu transportierenden Stoff "erkennt". Andere Tunnenproteine lassen alle Stoffe mit bestimmten Eingenschaften (z.B. Größe, Ladung) durch. Bei Untersuchungen hat man auch Tunnelproteine gefunden, deren Funktion durch bestimmte Moleküle ein- und ausgeschaltet werden kann. Transportproteine: Andere Proteine verfügen über keine Tunnel oder Kanäle. Sie transportieren Stoffe z.B. durch innermolkulare Bewegungen (Veränderung der Raumstruktur (meist Tertiärstruktur)) oder durch Bewegungen des Protein-Molekül-Komplexes in der Membran (Membranfluss). Wird für den Transport Energie verbraucht, dann ist dies ein aktiver Transport. Solche Transporte machen für die Zelle nur Sinn, wenn z.B. ein Transport entgegen dem Gradienten (entgegen dem Konzentrationsgefälle) erfolgen oder der Transport beschleunigt werden soll. Passive Transporte (z.B. Diffusion, Permeation und Osmose) erfolgen mit dem Grandienten ohne Energieverbrauch. Transportproteine werden nach der Anzahl der transportierten Stoffe und Richtungen unterschieden. Transportiert ein Protein nur einen Stoff, dann spricht man von einem Uniport (I). Werden zwei Stoffe gleichzeitig in die gleiche Richtung transportiert, nennen wir sie Symport (II). Beim Antiport werden die Stoffe in entgegengesetzte Richtungen bewegt. Beim Transport von zwei Stoffen ist die Q: de.wikipedia.org (Zoph) Anwesenheit beider Stoffe notwendige Voraussetzung. aktiver Transport an Carrier-Proteinen / Substanzpumpen: 1957 entdeckte der dänische Mediziner Jens Christian SKOU ein Enzym (Protein), das unter ATP-Verbrauch Na+-Ionen ins Zelläußere und K+-Ionen nach innen transportiert (1997 gab's dafür den NOBEL-Preis für Chemie). Wir werden diese K+-Na+-Ionen-Pumpe in der Neurophysiologie ausführlicher darstellen. Hier nur kurz das Arbeitsprinzip. Die beiden Teile Teile des Proteiens (Enzym-Nr. 3.6.3.9.) bilden einen scherenartigen Umklappmechanismus. Zuerst sind die beiden Teile zum Zellinneren geöffnet. Insgesamt drei Na+-Ionen müssen sich zuerst an den zugehörigen Bindungsorten anlagern, damit im nächsten Schritt mit ATP eine Phosphorilierung des einen Proteinteils erfolgen kann. Der Proteinkomplex erfährt eine Konformationsänderung und die "Schere" klappt zur anderen Seite um. Nun können die Na+-Ionen abwandern. An einer anderen Bindungsregion können nun zwei K+-Ionen andocken. Dies bewirkt ein Zurückklappen der Proteinstrukturen und das Abspalten von Phosphat. Solange genug Na+- und K+-Ionen sowie ATP vorhanden ist, solange kann sich der Vorgang wiederholen. - 72 - (c,p) 2008 lsp: dre ( ++ , ? -2 2 : 13 & . ( ( ++ -! "0- ( % 7 % * " " " * ) ! "05 0 6 ) @ 0 5 "" 2 " + " A ( ( ! A + (7 3 ( - % Endocytose: Die Endocytose ist der erste Transportprozess, den wir auch direkt mikroskopisch beobachten können. Besonders gut beobachtbar ist die Endocytose größerer Objekte – wie z.B. Nahrungspartikel (z.B. Bakterien). Diese können für ein noch besseren Sichtbarkeit mit sogenannten Vital-Farbstoffen (z.B. ) angefärbt werden. Kommt es zum Kontakt von Bakterium und Zellmembran, dann stellen Membranrezeptoren (Glycocalyx) Verbindungen her. Die fressende Zelle (Phagocyt) erkennt die Nahrung über die Oberfläche (Schlüssel-Schloß-Prinzip). Nach und nach wird immer mehr BakteriumsOberfläche von der "Fresser"-Membran umschlossen. Am Schluss ist es dann nur noch eine Frage der Oberflächenspannung und es bildet sich ein Bläschen mit einem Bakterium als Inhalt. Die Zelloberfläche verschließt sich wieder und steht für eine neue Nahrungsaufnahme wieder bereit. Im Falle der Aufnahme fester Objekte spricht man als Spezialfall der Endocytose von einer Phagocytose (griech.: phagein = essen). Bei flüssigen Stoffen nennt man es demgegenüber von Pinocytose (griech.: pinein = trinken). Die Bildung von nach innen gestülpten Bläschen wird durch Proteine (Clathrin) verstärkt, die muskelfaserähnliche Funktionen haben. Wenn außen an den Rezeptoren (Glycocalyx) bestimmte Stoffe andocken, dann bewirken die aktivierten Rezeptoren eine Kontraktion dieser Proteine. Duch das Zusammenziehen entsteht eine Eindellung der Zellmembran. Die Nahrungsbläschen verschmelzen mit Lysosomen ( GOLGI-Apparat). Die Lysosomen beinhalten "Verdauungs"-Enzyme. Die Enzyme sorgen für eine Zerlegungung des Bläscheninhalts (z.B. Bakterien, Hefen usw.). Die monomeren Moleküle werden dann durch die schon beschriebenen Transportvorgänge "ins Zellinnere transportiert", wo sie für weitere assimilatorische oder dissimilatorische Vorgänge genutzt werden. B 4 B + + B 7 & . + " " A 0 B " A 2 " + + 2 - 1 7 1 Exocytose: Die unverdaulichen Reste der Nahrungsbläschen, aber auch andere Visikel (mit Stoffwechselabfallprodukten), müssen irgendwann entsorgt werden. Zellen nutzen dazu einfach die Umgebung. Die Bläschen wandern an die Zellmembran und verschmelzen mit dieser. Man kann sich das so vorstellen, wie Luftblasen, die im Wasser aufsteigen und dann an der Oberfläche zerplatzen. Der Inhalt der Bläschen ergießt sich in die Umgebung. Die Exocytose wird auch auch Ptyocytose oder Extrusion genannt. - 73 - (c,p) 2008 lsp: dre 2.1.2. Rezeptionsvorgänge an Biomembranen Zellen müssen irgendwie Informationen (Reize) aus ihrer Umgebung aufnehmen können. Auf der Ebene einer Zelle sind dies vor allem chemische Informationen, die wichtig sind. Ist Nahrung in der Nähe? In welcher Richtung befindet sich die Nahrungsgsquelle? Gibt es chemische Informationen von anderen Zellen in der Umgebung? Ist der Nachbar Freund, Feind oder Nahrung? Ein (Chemo-)Rezeptor (entspricht sozusagen unseren Sinneszellen / - organen) besteht aus mehreren funktionellen Teilen. Diese werden oft Domänen genannt. Zumeist ist ein Rezeptor ein sehr komplexes Protein. Nach Außen (in den periplasmatischen Raum) auf der Zellmembran befindet sich die Rezeptor-Domäne. Sie ist für die Erkennung eines speziellen Stoffes (Reiz; Reizstoff; z.B. Lockund Schreckstoffe, Nahrung, Zellgifte) vorgesehen. Der Stoff (- auf den der Rezeptor reagieren soll -) und die Rezeptor-Domäne passen wie Schlüssel und Schloss zusammen. Mit mehreren Peptidketten ist der Rezeptor in der Biomembran verankert (MembranDomäne). In das Zellplasma (Cytosol) reicht die auslösende Domäne. An ihr ist ein Stoff (Botenstoff) angekoppelt, der bestimmte biochemische Prozesse in der Zelle steuert. Zumeist sind dies Aktivatoren oder Inhibitoren (Hemmstoffe) für bestimmte Enzyme ( E 1.1. Enzyme und enzymatische Reaktionen). Die meisten Rezeptionsvorgänge (Informations-aufnehmenden Vorgänge) laufen nach folgendem Schema ab. - 74 - (c,p) 2008 lsp: dre Dockt an der Rezeptor-Domäne nun der passende Stoff für den Rezeptor an, dann kommt es durch innermolekulare Veränderungen an der auslösenden Domäne zum Abspalten des Botenstoffs. Dieser bewirkt dann charakteristische Veränderungen im Stoffwechsel der Zelle. Nach der Abkopplung des Botenstoffs kann auch der Reizstoff wieder von der Rezeptor-Domäne abwandern. Unter ATP-Aufwändung wird nun wieder der Botenstoff (ein neues Molekül natürlich) an der auslösenden Domäne angebunden. Damit ist der Rezeptor wieder sensibel (arbeitsfähig). Andere Rezeptoren sind geregelte IonenKanäle. Ein gut untersuchtes Beispiel ist der Acetylcholin-Rezeptor (AcCh-Rezeptor) an den Synapsen (Nervenendköpfchen) von Nervenzellen. Hier dienen sie zur chemischen Informationsweitergabe von Nervenzelle zu Nervenzelle. Der Rezeptor ist ein integrales Protein mit einem Ionen-Kanal. Bei den Kanal-Rezeptoren verläuft die Informationsaufnahme ungefähr so. Normalerweise (z.B. beim AcCh-Rezeptor) ist der Kanal verschlossen. An der Außenseite hat der Rezeptor Andockstellen für das Acetylcholin. Dockt Acetylcholin an diese Stellen an, verändert sich die Raumstruktur des Kanals. Er öffnet sich und bestimmte Stoffe können den Kanal passieren. Beim AcCh-Rezeptor sind dies Na+-Ionen, die nun massiv auströmen können und das elektrische Potential an der Membran ändern – die Nervenzelle wird erregt. Wandern die Reizstoffe / Transmitter ab, dann verschliesst sich der Kanal wieder. - 75 - (c,p) 2008 lsp: dre 2.2. Zellwand Bei Pflanzenzellen ist die Zellwand die eigentlich äußerste Schicht einer Zelle. Die Zellwand besteht hier vorrangig aus Zellulose-Fasern und diversen Einlagerungen. Die bekannteste ist das Lignin – der sogenannte Holzstoff. Auch Pilze verfügen über eine Zellwand. Statt der Zellulose fungiert Chitin (bekannt von den Außenskeletten der Insekten) als Trägermaterial. Die Zellwand wird bei Pflanzenzellen erst nach dem Abschluss des Größenwachstums (Volumenwachstum) angelegt. Vorher gebildete Zellwände würden das Wachstum behindern. Nach und nach werden Zellulosefasern auf der Zellmembran abgelagert. Die Fasern bilden wechselnde Texturen (Primärwand: verflochten (Streuungstextur); Sekundärwand: ausgerichtet, parallel (Paralleltextur)). Mit Lignin verklebt bilden sie sehr stabile Außenhüllen der Zellen. Die verholzten Zellwände können auch nach Ableben der Zelle noch lange erhalten bleiben. Das Lignin verhindert einen schnellen Abbau der ZelluloseGerüste. In einigen Fällen folgt beim Zellwandaufbau noch eine Tertiärwand. Sie stellt den Abschluß zur Zellmembran dar. Die Zellulosefassern haben hier wieder eine Streuungstextur. Eingelagert werden wieder Lignin EM-Aufnahme: (Verholzung), Suberin (Verkorkung) oder auch Farbstoffe, Wachse, SalStreuungstextur ze (SiO2, CaCO3) und Gerbstoffe. Q: en.wikipedia.org (LadyofHats) + geänd. Drews 2.2.1. Mittellamelle Nach der Zellteilung wird zuerst eine junge Zellwand (Primodialwand) angelegt. Sie liegt sozusagen zwischen den beiden neuen Zellen. Die Bildung der Primordialwand vollendet die Trennung der Tochterzellen bei der Zellteilung. Im Wesentlichen besteht sie aus Pektinen (Pectine) und anderen Kohlenhydraten (Polyglucaronsäure) einschließlich Derivaten (z.B.: Polyglucaronsäure, Pectinsäure). Von Innen werden dann zuerst Membranabschnitte angelagert, welche die neue Zellmembran darstellen. Zwischen Mittellamelle und Zellmembran wird später (nach dem Zellwachstum) die Zellwand gebildet. - 76 - (c,p) 2008 lsp: dre 2.3. Cytoplasma Bei der Suche nach dem eigentlichen Lebensort einer Zelle sind wir beim Cytoplasma an der richtigen Stelle. Von der Zellmembran zusammengehalten und abgegrenzt beinhaltet es die verschiedenen Zellbestandteile. Der Stoff- und Komponenten-Mix des Cytoplasma's ist der Raum für die zig-Millionen Reaktionen und Vorgänge, die das eigentliche Leben ausmachen. Das Cytoplasma liegt in einem fließenden Übergang zwischen Gel und Sol vor. Die Konzentration und die Art der gelösten Stoffe ist so beschaffen, dass das Cytoplasma in einem Zustand zwischen flüssig bzw. eher leimartig (Sol) und fest (Gel) ist. Bei reichlichem Wasserangebot (auch innerhalb abgegrenzter Bereiche (Kompartmente)) geht das Cytoplasma in den Sol-Zustand über. Gelöste Stoffe sind gut beweglich. Bei geringerem Wasseranteil ist das Cytoplasma gelartig. Große (mehr oder weniger gelöste oder gequollene) Moleküle binden das Restwasser recht fest an sich. Die Wasser- und die kleineren gelösten Moleküle können sich – wenn überhaupt – nur langsam und kurzstreckig bewegen. Im Gel-Zustand bestehen auch für fettähnliche (lipophile, hydrophobe) gute Möglichkeiten an passende Reaktionspartner und zugehörige Enzyme zu kommen. - 77 - (c,p) 2008 lsp: dre Exkurs: Sol- und Gel-Zustand Internet-Links: Q: de.wikipedia.org () - 78 - (c,p) 2008 lsp: dre 2.4. Kernäquivalent / Zellkern Durch spezielle Färbungen (z.B. FEULGENFärbung (fuchsinschweflige Säure + Chlorwasserstoffsäure)) kann man im zentralen Bereich von Zellen ein mehr oder weniger kugelförmiges Objekt sichtbar machen. Die Färbung basiert auf dem vorhandenen genetischen Material (DNA). Bei Pflanzen, wo die Vakuolen meist den Zentralraum belegen, ist der Zellkern mit dem anderen Cytoplasma an den Randbereich gedrängt. Das Kernäquivalent von Procyten ist weniger scharf abgegrenzt als der echte Zellkern von Eucyten. Weiterhin fehlt eine membranöse Abgrenzung. Das Chromatin ist während des gesamten Q: de.wikipedia.org (zituba) Zellzyklus gleichmäßig undifferenziert (es bilden sich keine mit Chromosomen vergleichbaren Strukturen). Zellkerne haben meist einen Durchmesser um die 0,5 (Pilze) bis 500 µm (Eizellen). Ein echter Zellkern ist von einer Doppelmembran umgeben, die in regelmäßigen Abständen von Poren durchzogen sind. Die Poren und die Kernmembran gehen fließend in das Endoplasmatische Retikulum über. Das innere Plasma (Kryoplasma, Kernplasma) hat von Cytoplasma abweichende Eigenschaften. Es erscheint dickflüssiger bzw. dichter. Deshalb ist bei vielen Zellen der Zellkern auch schon lichtmikroskopisch ohne spezielle Färbungen zu erkennen. Im Inneren des Zellkerns liegen die Chromationfäden. Mit Beginn der Kernteilung (Mitose) kommt es zur Spiralisierung des Chromatins zu Chromosomen. Weiterhin befindet sich im Kernplasma noch das Kernkörperchen (NucQ: en.wikipedia.org (LadyofHats); geändert Drews leolus), der Aufgaben bei der Zellteilung (Mitose) übernimmt. Der Vorgang der Mitose ist im Abschnitt Genetik dieses Skripts ausführlich beschrieben ( I 7.4. Weitergabe und Verteilung der Erbinformation). Normalerweise finden wir in einer Zelle nur einen Zellkern. In einigen Zellzusammenschlüssen (Syncytien) bleiben die Zellkerne enthalten, so dass der Eindruck entsteht, eine Muskelzelle enthielte mehrere Zellkerne. Pilz-Hyphen (Pilz-Fäden), aber auch anderer sehr große Zellen (Milchröhren oder Bastfasern bei Pflanzen) halten oft ebenfalls größere Mengen an Zellkernen, da hier die trennenden Zellmembranen zwischen den "Einzelzellen" nicht mehr vorhanden sind. Selten sind ausgewachsenen Zellen kernlos. Bei diesen Zellen ist dann kei- - 79 - (c,p) 2008 lsp: dre ne Zellteilung mehr möglich und der Zelltod tritt in absehbarer Zeit ein. Ein typisches Beispiel sind die roten Blutkörperchen (Erythrocyten) beim Menschen (!). Zellkern bzw. Kernäquivalent stellen die Informations- und Steuerzentralen der Zellen dar. Der Hauptteil der Informations- und Steuerungsaufgaben wird über das genetische Material realisiert ( I 7.6. Speicherung der Erbinformation). Trotz intensiver Forschung sind viele der Vorgänge und Phänomene in ihren Zusammenhängen und Abläufen noch ungeklärt. Die Nucleoli (Kernkörperchen) sind sehr dichte Bereiche im Zellkern. In diploiden Zellen befinden sich im Zellkern meist zwei Nucleoli. Es wurden aber auch schon keine bis insgesamt sieben Stück beobachtet. In den Kernkörperchen findet die Synthese der rRNA und der Ribosomen statt. Während der Kernteilung verschwinden die Nucleoli und tauchen nach der Bildung der neuen Q: de.wikipedia.org EM-Aufnahme: Zellkern Kernhüllen wieder auf. Im elektronenmikroskopischen Bild kann man sehr gut den unmittelbaren Übergang von Kernmembran und Endoplasmatischen Retikulum (parallele streifige Strukturen) erkennen. Die Erbinformationen aus dem Zellkern werden hier zu Eiweißen umgesetzt ( I 7.7. Realisierung der Erbinformationen). - 80 - (c,p) 2008 lsp: dre 2.5. Endoplasmatisches Retikulum, GOLGI-Apparat und Visikel Im Cytoplasma laufen gleichzeit mehrere einhunderttausend Reaktionen zur gleichen Zeit ab. Damit diese sich nicht behindern und gebildete Zwischenprodukte nicht gleich wegdiffundieren, verfügen eucytische Zellen über eine ausgeprägte Kompartmentierung (räumliche Strukturierung, Unterteilung). Kleine Bereiche sind durch verschiedenste Abgrenzungen (Gel-Sol-Phasengrenzen, Membranen) voneinander abgeteilt. Die entstehenden Räume nennt man Kompartmente. Membranen als Kompartmentgrenzen bieten Enzymen und Ribosomen Halt. 2.5.1. Endoplasmatisches Retikulum Das größte Kompartmentierungssystem ist das Endoplasmatische Retikulum (ER). Dieses ist ein weit verzweigtes (labyrintartiges) Membransystem, dass fast die gesamt Zelle durchzieht. Das Membransystem beginnt schon an den Zellkernporen (2). Sind auf den Membranen (3) Ribosomen (5) angelagert, entsteht im Elektronenmikroskop ein pickliger Eindruck. Dieses wird rauhes ER (3) genannt. Glattes ER (4) enthält kaum Ribosomen. Das rauhe ER ist der Ort der Biosynthese der Proteine und von Membranabschnitten. Mit diesen kann dann nach einer Kernteilung die neue trennende Zellmembran gebildet werden. Innerhalb des glatten ER finden Unmengen weiter biochemischer Vorgänge statt. Die gebildeten Stoffe (6) werden in der gesamten Zelle weiterverwendet. Q: de.wikipedia.org (Magnus Manske) 2.5.2. GOLGI-Apparat Dictosomen (GOLGI-Körper) sind charakteristische Gebilde in den Zellen. Sie sehen aus wie Stapel von immer größer werdenden doppelschichtigen Membranscheiben. Die Dictosomen des GOLGI-Apparates (Gesamtheit aller Dictyosomen einschließlich der GOLGI-Vesikel (7)) sind ein Ort sehr intensiver Stoffproduktion. Hier – in den Zisternen (11) – entstehen z.B. Enzyme für die "Verdauung" aufgenommener Nahrungspartikel. Zwischen ER und Dictyosomen existiert ein intensiver Stofftransport. Abgeschnürrte Vesikel des ER enthalten frisch produzierte Proteine (Enzyme). Die Vesikel wandern in Richtung cisEnde (9) des Dictyosoms und verschmelzen mit den Membranstapeln. Die Membranstapel werden zum trans-Ende langsam immer ausgedehnter (10). In der Zwischenzeit werden die enthaltenen Proteine immer weiter gewandelt und durch neue Stoffe (Hormone, Sekrete) ergänzt. - 81 - (c,p) 2008 lsp: dre Die Dictyosomen schnüren an den Enden der Membranstapel immer wieder neue GOLGI-Vesikel ab. Später verschmelzen diese mit den endocytotisch gebildeten Nahrungsbläschen. Desweiteren bilden Dictyosomen sekretorische Vesikel, die vor allem Hormone, Transmitter usw. enthalten können. Diese Vesikel werden in Richtung Zellmembran transportiert und der Inhalt (Sekrete) nach außen ausgeschüttet (Exocytose, Sekretion). Q: micro.magnet.fsu.edu (geändert Drews) 2.5.3. weitere vesikuläre Strukturen Neben den großen Vesikel gibt es im Cytoplasma noch verschiedene andere kleinere Vesikel, die sich primär in den enthaltenen Enzymen und Stoffen unterscheiden. 2.5.3.1. Lysosomen Lysosomen dienen der Verdauung. Sie enthalten Phosphatase (Leitenzym). Mit diesem Enzym werden die Nahrungs-Partikel zersetzt. In Hungersituationen kann es bei Pflanzen zur sogenannten Autophagie kommen. Nicht mehr dringend benötigte Zellbestandteile oder auch ältere Mitochondrien werden dann abgebaut, um einen elementaren Stoffwechsel aufrechtzuerhalten. Q: biology.unm.edu 2.5.3.2. Microbodies Microbodies sind mit 0,2 bis 1,5 µm wirklich sehr kleine Vesikel. Sie haben nur eine kurze "Lebenszeit" von 2 bis 3 Tagen. Microbodies sind in der Lage aus Kohlenhydraten diverse andere organische Stoffe herzustellen. Dabei entsteht als Nebenprodukt oft das gefährliche Wasserstoffperoxid. Microbodies enthalten als Leitstoff (Leitenzym) die Katalase, das genau dieses Wasserstoffperoxid schnell umwandeln kann (Entgiftungsenzym). Wasserstoffperoxid ist chemisch sehr aggressiv und reagiert mit sehr vielen anderen Stoffen, die dabei oxidiert werden. Unter biochemischen Verhältnissen bedeutet dies meist die Zerstörung des Stoffes selbst oder dessen Funktion (weil dieser dann ein anderer ist). - 82 - (c,p) 2008 lsp: dre Ursache ist die Bildung von äußerst reaktiven Sauerstoff-Radikalen (O ) während des Zerfalls des Wasserstoffperoxids. H2O H2O + O Die Radikale (mit ihren ungpaarten Elektronen) reagieren praktisch mit jedem Stoff in der Zelle und verändern dabei Bau und Eigenschaften der zelleigenen Stoffe. In den meisten Fällen können die oxidierten Stoffe nicht mehr die Funktion der ursprünglichen Verbindungen nachkommen. Wahrscheinlich kommt die Katalase nur in Microbodies vor. Die von ihr geförderte Reaktion: H2O2 H2O + ½ O2 produziert auch freie Energie. Im Gegensatz zu den Mitochondrien kann diese aber nicht in Form von ATP gebunden werden, sondern wird als Wärme frei. Zu den Microbodies gehören auch die Peroxisomen (Peroxysomen), die zur Glucogenese fähig sind. Glucogenese ist die Bildung von Kohlenhydraten z.B. aus Aminosäuren. Peroxysomen kommen bei höhreren Tieren in der Leber und auch in der Niere vor. In Pflanzen sind Peroxysomen bei der Photorespiration (Lichtatmung) tätig. Dabei werden mit Hilfe von Licht direkt Aminosäuren (Glutaminsäure, Glycin, Serin) produziert. Die Lichtatmung ist ein alternativer Nebenweg zur Photosynthese ( E 3.2.2. Photosynthese). Eine weitere Art sind die Glyoxysomen. Sie sind zum direkten Abbau von Fettsäuren zu Acetyl-Coenzym A fähig. Glyoxysomen kommen vor allem in fettspeichern Geweben von Pflanzen vor. Microbodies sind also hochentwickelte Stoffwechselspezialsten, die auf abgegrenzten Raum alle Werkzeuge und Hilfsmittel für ihren Arbeitsauftrag (vorrangig Entgiftungen) zusammenhalten. Microbodies sind wahrscheinlich evolutionär wesentlich älter als Mitochondrien. In den heutigen Eucyten kooperieren Microbodies und Mitochondrien biochemisch sehr intensiv. - 83 - (c,p) 2008 lsp: dre 2.6. Tubuläre Strukturen Tubuläre oder fibrilläre (faserförmige) Strukturen sind in der Zelle für Formgebung und Bewegung verantwortlich. 2.6.1. Zellskelett Zellen ohne Zellwand müssten eigentlich auf Grund der Oberflächenspannung und der Druckverhältnisse im Cytoplasma mehr oder weniger kugelförmig sein. Vor allem bei tierischen Zellen wird die abweichende Zellform durch faserförmige bis netzartige Innen-Strukturen erzeugt. Spezielle Anfärbungen und Mikroskopiertechniken (Fluoressenz-Mikroskopie) machen die Moleküle der Innenstrukturen (z.B. Tubulin) sichtbar. Im nebenstehenden Bild sind sie grün fluoreszierend. Die blauen Regionen sind die Zellkerne und die Zellmembran wird durch rot leuchtende Moleküle (auch Actin) markiert. Viele Zellskelette sind nicht nur starr – sie ermöglichen oft auch einfachste Bewegungen. Durch molekülinterne Konfigurationsänderungen (Actin) oder Ab- bzw. Aufbau (Tubulin) sind Längenveränderungen möglich. Letztendlich kann dann bei koordinierten Vorgängen eine Formveränderung oder Bewegung der Zelle erreicht werden. Q: de.wikipedia.org (rsb.info.nih.gov) 2.6.2. Mikrotubulli Mikrotubuli sind die Grundbauelemente für größere Einheiten, wie z.B. Spindelapparat und Geißeln. Der Grundbaustoff ist das Eiweiß Tubulin. In der Praxis unterscheiden wir u.a. α- und β-Tubulin. Von der Raumform kann man sie sich wie Maiskörner vorstellen. Jeweils ein Molekül α- und β-Tubulin bilden zusammen eine Baueinheit (Hetero-Dimer). Die Baueinheiten können weiter polymerisieren. Dabei ist die Polymerisierung außer in die Längsrichtung auch in die Breite möglich. (Wachstum 8 µm/s (Abbau 17 µm/s)) Durch die Molekülform des Tubulins kommt es nicht zur Ausbildung einer Fläche, sondern nach 13 bis 14 Molekülen zum Ringschluss mit einem leichten Versatz der Monomere. So entsteht eine Helix (Schrauben-Struktur). - 84 - Tubulin-Hetero-Dimer Q: de.wikipedia.org (Toreau) (c,p) 2008 lsp: dre Das helikale Gebilde erinnert dann auch wieder an einen Maiskolben. Die Hohlstruktur hat einen Außendurchmesser von 18 bis 30 nm. Die Bildung polymerisierter Strukturen erfolgt nicht spontan sondern an sogenannten Mikrotubuli-Organisationszentren (MTOC). Fertige Mikrotubuli werden dann vom MTOC abgelöst.Weitere sehr langestreckte Proteine (mikrotubulusassoziierte Proteine, MAPs) setzen sich in die Furchen und stabilieren den gesamten Komplex noch weiter. Zum anderen bieten diese Proteine und das Tubulin selbst auch wieder Ansatzstellen für weitere Eiweiße (z.B. Dynein, Kinesin, …) und andere Moleküle. Bewegungen in Längsrichtungen (Verkürzung bzw. Verlängerung) stellt man sich heute volgendermaßen vor. Die Mikrotubuli sind an den Enden angbunden. In der Mitte sitzen Riesenenzyme, die aus dem Tubulus nach und nach Tubulin entfernen oder hinzufügen. Dabei wir dann der Faserschluß wieder hergestellt, damit der Zusammenhalt nicht gefährdet ist. Die Mikrotubuli können aber auch als Schienensystem fungirien. Das Kinesin (schraubesförmiges Molekül mit "Füßchen") sitzt auf dem Mikrotubuli. Unter ATP-Verbrauch macht das Kinesin einen "Schritt" (8 nm) um ein Hetero-Dimer (je eine rote u. weiße Kugel). Am langen Ende des Kinesin können größere Objekte (z.B. Visikel) andocken, die so langsam durch die Zelle gezogen (Kraft 5 pN) werden. Die Wanderung ist immer in AufbauQ: de.wikipedia.org (Moez) richtung des Tubulus (Minus-nachPlus, vom Centrosom weg). Das etwas anders gebaute Dynein (Protein mit "kurzen gespreizten Beinchen") wandert in die Gegenrichtung. Mikrotubuli sind in Nervenzellen auch am axonalen Transport von Neurotransmittern beteiligt. In der Zelle finden wir auch höher organisierte Mikrotubuli. Bei diesen bilden zwei oder drei Röhren eine Einheit. Die erste Röhre (A-Tubulus, Subfaser A) wird um einen Dreiviertel-Ring (B-Tubulus, Subfaser B) von (9 –)10 Hetero-Dimeren ergänzt. Drei (bis vier) Tubulin-Dimere werden in der Dublette (auch: Duplette, Doppeltubuli) gemeinsam benutzt. Eine eventuelle dritte Röhre (C-Tubulus, Subfaser C) ist auch wieder so eine Erweiterung um (9 –)10 Hetero-Dimere. - 85 - (c,p) 2008 lsp: dre 2.6.3. Centriolen und Spindelapparat Die verschiedenen Mikrotubuli-Strukturen sind die Bauelemente für Centriolen, Cilien und Geißeln. Im Centrosom liegen die rund 150 nm dicken (Durchmesser) und 300 bis 500 nm langen Centriolen. Sie haben eine röhrenförmige Struktur. Eine Röhre selbst ist aus 9 ringförmig angeordneten Dubletten oder Tripletten aufgebaut (selten nur Singulette). Die Vermehrung von Centrosom und Centriolen erfolgt in der Interphase. Centrosomen enthalten wahrscheinlich ihre eigene DNA. Ein Tochterzwei Centriolen aus einem Centrosom Centriolen bildet sich neben dem Mutter-Centriol (scheibar aus dem Nichts). Der neue Centriol steht senkrecht zum alten. Wie genau diese Vorgänge ablaufen, ist ungeklärt. Centriolen sind an der Bildung von Geißeln und des Spindelapparates beteiligt. Zwischen den Centriolen spannen sich die Zentralfasern durch die ganze Zelle. Die Zentralfasern sind sehr stabil. Sie bilden sozusagen das feste Rückrat der Zellen. Zur Ausbildung des Spindelapparates wandern die Centrosomen in Richtung der Zellpole. Zwischen den Centrosomen werden dabei die Spindelapparat Spindelfasern ausgebildet. Diese bestehen aus (Anaphase der Mitose) einem bis mehreren – z.T. verdrillten – MikrotuQ: de.wikipedia.org buli. Die Mikrotubuli werden in der Metaphase der Mitose am Centromer des Chromosoms (Kinetochore) verankert. Während der Anaphase wandern die Centrosomen weiter zu den Zellpolen. Die gebunden Spindelfasern gleiten an freien Fasern entlang, so dass die Chromatiden zu den Zellpolen gezogen werden. Mikrotubuli des Spindelapparates an einem Chromsosom Q: de.wikipedia.org (Ron de Leeuw (geänd. Drews)) - 86 - (c,p) 2008 lsp: dre 2.6.4. Cilien Cilien und Geißeln (Flagellen) haben einen ähnlichen Bau und werden unter dem Begriff Undulipodien zusamengefasst. Von Cilien (Wimpern) sprechen wir, wenn es sehr viele an der Zelloberfläche sind. Bei wenigen spricht man eher von Geißeln (Flagellen). Geißeln sind zudem größer und länger. Das Flimmer-Epithel in der Luftröhre des Menschen (s. Abb.) ist ein gutes Beispiel für einen Cilienbesatz der Zelloberfläche. Die Cilien- od. Flagellen-Faser wird auch Axonem genannt. Q: de.wikipedia.org (Charles Daghlian) In einer Axonem () ordnen sich 9 Dubletten um zwei zentrale Einzel-Tubuli an. Vom BTubulus stellen Dynein-Moleküle eine Verbindung zum A-Tubulus der nächsten Dublette im Kreis her. Der Abstand zwischen den Dubletten wird durch Nexin– u. Dynein-Moleküle aufrechterhalten. Die Gesamtstruktur hat einen Durchmesser von rund 150 nm. Die Bewegung der Cilien wird durch eine Gleitbewegung zwischen den MikrotubuliStrukturen erreicht. Das zwischen den Dubletten liegende Dynein macht unter ATPVerbrauch eine Formveränderung durch. Diese verschiebt die Mikrotubuli gegeneinander – die Mikrotubuli gleiten gewissermaßen aneinander vorbei. Das Protein Nexin sorgt für einen gleichbleibenden Abstand zwischen die Mikrotubuli in der Axonem-Struktur. - 87 - (c,p) 2008 lsp: dre 2.6.4. Geißeln Wie schon besprochen, haben Geißeln einen ähnlich Aufbau wie Centriolen und Cilien. Der Basalkörper (Kinetosomen) – sozusagen die Wurzel einer Geißel – entspricht einem Centriol. Kommt es am Dynein unter ATPVerbrauch zu einer Konformationsänderung, so schieben sich die Tubuli aneinander vorbei. Man spricht von einer Gleitbewegung. Auf die gesamte Länge einer Geißel kann so eine weitausladende mikrokopisch sichtbare Bewegung entstehen. Durch koordinierte Muster der DyneinAktivierung entstehen verschiedene Bewegungsmuster. Wie diese Musterbildung und die Koordinierung zwischen benachbarten Cilien erfolgt, ist noch unklar. Q: de.wikipedia.org (Brudersohn) Q: de.wikipedia.org (Brudersohn) Bakterien, Spermien z.B. (A ) Homo sapiens sapiens, (A ) Euglena Bakterien-Geißel (auch: Flagelle), starr 10 µm lang, d= 10 – 20 nm (dünner als ein Mikrotulus aus Tubulin), aus Flagellin , ähnliche Grundaufbau wie Mikrotubuli (8 – 11 Moleküle im Röhrenring), "Kugellager"-verankert in der Bakterien-Zellwand (bzw. Zellmembran); Bewegung über Ionen-Transport, Prinzip eines Elektromotor, 40 – 50 Hz - 88 - Q: de.wikipedia.org () (c,p) 2008 lsp: dre 1 .. Geißel 2 .. periplasmatischer Raum 3 .. Winkelstück 4 .. Koppelstück 5 .. Lager (L-Ring) 6 .. Rotor 7 .. Lager (P-Ring) 8 .. Zellwand 9 .. Stator 10 .. MS-Ring 11 .. C-Ring 12 .. Typ III-Sekretionssystem (Drehrichtungsumsteller 13 .. äußere Membran 14 .. Cytoplasmamembran (innere Membran) 15 .. Geißeldeckel "Technische" Daten: Arbeitsspannung: 25 – 125 mV; bis 20000 Umdrehungen min-1; Wirkungsgrad bei 80 % Q: de.wikipedia.org (LadyofHats) Verwunderlich ist der extrem spezialisierte und abgestimmte Aufbau der Geißel. Am Bau sind ungefähr 40 verschiedene Proteine beteiligt. Dazu kommen noch 8 Proteine zur Steuerung der Bewegung. Wie diese evolutionär entstanden sein sollen, ist völlig ungeklärt. Jedes einzelne Protein ist für die Gesamt-Funktion unabdingbar. Eine schrittweise Entstehung ist kaum denkbar. (Da kann man schon mal den intelligenten Designer (Kreatinismus) auf die Tagesordnung rufen. Aber auch für ihn gibt es keinen Beweis, so dass die Forschung hier riesigen Nachholebedarf hat!) interessante(r) Internet-Link(s): www.nanonet.go.jp/english/mailmag/2004/011a.html (Bilder + Movie (Erforschung; Aufbau u. Funktionsweise (~ 18 u. 23 min.)) 34 min lang) - 89 - (c,p) 2008 lsp: dre 2.6.5. Actin-Filamente =Mikrofilamente + Myosin-Filamente (d=6-8 nm), bewirken Cytoplasmabewegung , Myosin kann ATP spalten, Energiefreisetzung bewirkt Konformationsänderung des Actins, Kontraktion des gesamten Actin-Myosin-Filaments d=6 nm Q: de.wikipedia.org () 2.6.6. Intermediär-Filamente Durchmesser zwischen Mikrofilamente und Mikrotubuli verschiedene Proteine u.a. Kreatin d=10 nm Verankerung über spezielle Proteine an der Zellmembran abgestorbene Zellen mit viel Kreatin bilden Hornhaut, Haare langsamer Aufbau, sehr stabil, unbeweglich Q: de.wikipedia.org () - 90 - (c,p) 2008 lsp: dre 2.7. Zellorganellen Zellorganellen sind durch Doppelmebranen abgegrenzte – recht große (lichtmikroskopisch sichtbar) Objekte in der Zelle. Bei der Sichtbarkeit stellen die Mitochandrien eine Ausnahme dar. Sie werden in den besten Lichtmikroskopen gerade mal als kleiner Fleck (Punkt) sichtbar. 2.7.1. Mitochondrien In den Mitochondrien findet die aerobe Dissimilation statt. Sie sind die ultimativen Kraftwerke aller eucytischen Zellen. In einer Zelle kommen einhundert bis einige hunderttausend Mitochondien vor. Ihre äußere Form ist zumeist zylindrisch mit halbkugelförmigen Enden. Sie können aber auch kugel- oder fadenförmig sein. Die Größe variert in der Länge zwischen 1 bis 5 µm und in den schmalen Ausdehnungen (Breite) zwischen 0,5 und 1 µm. Im Elektronenmikroskop kann man eine doppelschichtige Umhüllung beobachten. Die innere Membran stülbt sich zuQ: de.wikipedia.org (Louisa Howard) EM-Aufnahme dem weiter nach innen ein. So entstehen verschiedene Typen (Tubuli-, Cristae- u. Sacculi-Typ), die sich aber funktionell wenig unterscheiden. Die blattartigen Einstülpungen (Cristae-Typ) stellen wohl den häufigsten Fall dar. Zwischen der Außenmembran und der inneren liegt der Intermediärraum. In ihm befindet sich die äußere Matrix, od. auch das äußere Mitochondienplasma. Die (innere) Matrix füllt den gesamten Innenraum aus. In der Innenmembran befinden sich die Enzyme / Redoxsysteme der Atmungskette. Die vorlaufenden und zusätzlichen dissimilatorischen Vorgänge finden in der Matrix statt. Dazu gehören Glycolyse, Citratcyclus sowie die Fettsäureoxidation. Mitochondrien verfügen über ein eigenes genetisches Material. Sie vermehren sich durch Teilung / Spaltung. Mitochondrien können nicht spontan oder direkt von der Zelle gebildet werden. Sie können nur aus anderen Mitochondrien hervorgehen. - 91 - (c,p) 2008 lsp: dre 2.7.2. Chloroplasten Nur bei Pflanzen kommen Chloroplasten vor. In den Chloroplasten ist der grüne Blattfarbstoff Chlorophyll konzentiert. Bei violetten Laubblättern überdecken Xanthophylle das grün. Die Konzentrationen und die Mengen der Blattfarbstoffe bestimmen die natürliche Blattfarbe. Die Laubfarben entstehen beim Abbau der verschiedenen Blattfarbstoffe. Chloroplasten sind zwischen 2 – 8 µm groß und damit schon lichtmikroskopisch sichtbar. Pflanzenteile, die nur wenig oder gar nicht mit Licht in Kontakt kommen, haben keine ausgebildeten Chloroplasten. Aber auch in belichteten Pflanzenteilen sind sie nicht immer beobachtbar. So fehlen sie z.B. in vielen Blüten, Früchten und z.B. auch in den SpaltQ: rsb.info.nih.gov öffnungszellen der sonst grünen Blätter. Einfache Pflanzen verfügen über netzörmige, schraubenfärmige, gelappte oder sternförmige Chloroplasten. Linseförmige Chloroplasten kommen mehr bei höheren Pflanzen vor. Sie bewegen sich mit dem Cytoplasma durch die Zelle. Die Anzahl in einer Zelle kann stark variieren. Auch die Chloroplasten besitzen eine doppelte Umhüllung ((1) und (3)). Die äußere (1) ähnelt sehr den Membranen der restlichen Zelle. Die innere Membran (3) ist eher bakterienähnlich. Im Innenraum (4) befindet sich die Chloroplasten-Matrix – meist Stroma genannt. Auch die Intermembranzone (2) ist mit Plasma ausgefüllt. Die innere Membran faltet sich im Innenraum Q: de.wikipedia.org (Kristian Peters) weiter. Dabei entstehen blattartige Strukturen – die Thylakoide. Im Stroma einzeln liegende Membranschichten werden als Stromathyllakoide (8) bezeichnet. An bestimmten Stellen bilden die Thyllakoide Stapel. In guten Lichtmikroskopen sind diese als Grana (7) (dt.: Flecken) sichtbar. Die Thyllakoide hier heißen deshalb Granathyllakoide (6). Der Innenbereich zwischen den Thyllakoidmembranen wird Lumen (5) genannt. Im Stroma finden wir noch Stärkekörner (9) und Globuli (10) (enthalten Fette, Glycolipide, Chinone, Carotinoide und andere Farbstoffe). Chloroplasten verfügen ebenfalls über eigene genetische Informationsspeicher (11), die sehr einer Bak- - 92 - (c,p) 2008 lsp: dre Q: en.wikipedia.org (SuperManu) terien-DNA ähnelt. Chloroplasten können nur aus ihresgleichen heraus gebildet werden oder aber durch verschiedene Umwandlungen auch aus anderen Plastiden. Eine Urzeugung in der Zelle ist nicht möglich. Zum Nachweis dienen Pflanzen mit panachierten Blättern (hellgefleckt, s.a. Abb. weiter oben). Bei ihnen wurden – meist durch künstliche Maßnahmen – Chloroplasten-Verluste erzielt. Bei vegetativer Fortpflanzung bleiben diese an der gleichen oder einer abgeleiteten Stelle erhalten. Ein anderer Beleg ist bei Kakteen möglich. Sicher haben Sie in einem Pflanzenladen schon einmal farbige (gelb oder rötlich) Kakteen gesehen. Diese haben einen Chlorophyll-Verlust. Überleben können solche Exemplare nur, wenn sie auf einem anderen (grünen) Kaktus augepfropft werden. Dieser versorgt die Aufsitzer mit den notwendigen Stoffen. In den Chloroplasten findet die vollständige Photosynthese ( E 3.2.2. Photosynthese) statt. Aus der Lage des Chlorophylls innerhalb der Thyllakoidmembranen, kann man ableiten, dass hier die Lichtreaktionen ablaufen. Die Dunkelreaktionen finden vorrangig im Stroma statt. - 93 - (c,p) 2008 lsp: dre 2.7.4. Leukoplasten In Speicherorganen und unbelichteten Pflanzenteilen findet man Leukoplasten. Sie sind oval, ei- bis kugelförmig gebaut. Leukoplasten sind üblicherweise farblos. Mit Iod-Kaliumiodid-Lösung kann eine Blau- bis Schwarzfärbung erzeugt werden. Die typische Funktion von Leukoplasten ist die Einspeicherung von Glucose in Form von Stärke. Im Bedarfsfall kann die Stärke von den Leukoplasten auch wieder zerlegt werden und die freiwerdende Glucose anderen Zellen zur Dissimilation (und heterotrophen Assimilation) bereitgestellt werden. Stärkespeichernde Leukoplasten werden auch als AmyQ: de.wikipedia.org (Mnolf) loplasten (Amylose: eine Stärkeart) geführt. Die Speicherorgane dienen natürlich vornehmlich der eigenen Versorgung in schlechten Zeiten oder als Initialstoffreserve (Speicherorgan zweijähriger Pflanzen). 2.7.3. Chromoplasten Chromoplasten finden wir in Blütenblättern, Früchten und verschiedenen Speicherorganen. Die typische Färbung geht ins rotorange-violette. Die enthaltenen Carotine und Xanthophylle (zusammen Carotinoide) bestimmen die Farbe. Der Bau entpricht den Leukoplasten. Funktionell unterscheiden sie sich dagegen stark. Ihre primäre Funktion ist Speicherung (und Präsentation) von Farbstoffen (statt Stärke). Mit diesen sollen Insekten oder andere Tiere angelockt werden, um die Verbreitung der Samen oder Pollen zu forcieren. Für die meisten Plastiden sind sogenannte Proplastiden die Vorform. Diese können sich durch Teilung / Spaltung vermehren. Proplastide sind weitesgehend undifferenziert. Je nach Lage in der Pflanze bzw. abhängig von den Bedingungen (z.B. Licht) entwickeln sie sich in die eine oder andere Plastidenart. - 94 - Q: de.wikipedia.org () (c,p) 2008 lsp: dre 2.8. Vakuole Im Zentrum vieler Zellen befinden sich ein bis wenige membramumhüllte Flüssigkeitsansammlungen. Diese werden als Vakuole bezeichnet. Sie nehmen dort mit zunehmenden Zellalter einen immer größer werdenden Raum ein. Bei sehr alten Zellen oder Zellen in Früchten oder Speicherorganen nehmen Vakuolen oft einen sehr großen Raum (bis ungefähr 95%) ein. Vakuolen haben vorrangig Speicher- und Sammelfunktionen. Desweiteren sind sie entscheidend an der Regulation des Wasserhaushalts und der damit zusammenhängenden Aufrechterhaltung des Zellinnendrucks (Tugor) beteiligt. Vakuolen bilden sich nur aus ihresgleichen oder aus Ausstülpungen des endoplasmatischen Retikulums bzw. des GOLGI-Apparates. Die umgebende (einfache) Membran einer Vakuole heißt Tonoplast. Der Zellsaft enthält neben anorganischen Salzen auch organische Säuren, lösliche Kohlenhydrate, Zuckeralkohole, Aminosäuren, Alkaloide, Glykoside und Farbstoffe. (Farbstoff-haltigen Vakuolen (z.B. aus Q: de.wikipedia.org (Mnolf) Linguster-Beeren, untere Epidermis von Alpenveilchenblättern, alle farbigen Zellen der (A ) Roten Rübe, Rotkohl-Blätter) lassen sich sehr gut für Beobachtungen verwenden.) Bei wechselnden osmotischen Verhältnissen verändert sich die Größe (Innenvolumen) der Vakuole. Umgibt man eine pflanzliche Zelle (mit Zellwand und Vakuole) mit einer konzentrierten (hypertoniQ: de.wikipedia.org (Mnolf) schen) Lösung, dann kommt es zu osmotischen Vorgängen. Wasser tritt verstärkt in das Außenmedium aus. Das Volumen der Vakuole reduziert sich dabei. Da die Zellwand als starre Einheit ein fester Raster darstellt, kann man diesen Effekt gut beobachten (siehe Abb. rechts). Das Plasmalemma löst sich von der Zellwand und Umgebungsflüssigkeit strömt in den freiwerdenden Raum (Zell-Lumen). Das Cytoplasma ist von den Vorgängen ebenfalls betroffen, nur wird der Effekt wegen der Volumenverhältnisse nicht so deutlich sichtbar. Wird das Umgebungsmedium nun wieder durch normales Wasser oder eine isotonische Lösung (gleichkonzentriert; bezogen auf die Ausgangsbedingungen in der Vakuole) ersetzt, kehren sich die osmotischen Verhältnisse um. Wasser wandert nun wieder verstärkt in die Vakuole. Sie dehnt sich aus und mit steigendem Volumen wird das Umgebungsmedium aus dem Zwischenraum zwischen Zellwand und Plasmalemma herausgedrückt. Die Zelle nimmt letztendlich wieder den ursprünglichen Raum ein. Die Umkehrung der Plasmolyse nennt man Deplasmolyse. Setzt man die Zelle einer hyptonischen Lösung (sehr schwach konzentrierte Lösung oder reines Lösungsmittel; z.B. dest. Wasser) aus, dann wird der Wassereinstrom in die Vakuole weiter begünstigt. Da wegen der begrenzenden Zellwand kaum zusätzliches Volumen zur Verfügung steht, steigt der Druck in der Vakuole. Unter bestimmten Bedingungen kann es dann auch zum Platzen der Zelle kommen. - 95 - (c,p) 2008 lsp: dre 7 ! ) " C 0 " C % & A ) 43 A ( 5< . + - &DE 3 & 5 0 ( " 7 Q: botit.botany.wisc.edu % 3 G 6 0 ( ++ ( $ 0 7 9 . AC 0 " 7F "" 5 * -A 3 ! " "" " % 7 + ( ++ ! 0 " + (4 " " H 3 " /0 " % ! (5 " C A - 96 - DE G C 0 $ % " " A A ! " C % A % , + "" 1 ! 4 + (c,p) 2008 lsp: dre Eine besondere Ausprägung einer Vakuole ist die sogenannte pulsierende Vakuole einiger tierischer Einzeller (z.B. (a ) Paramecium spec. (Pantoffeltierchen)). Diese ist nicht direkt mit den Vakuolen pflanzlicher Zellen vergleichbar. Gleichwohl sind sie auch für die Regulation des Wasserhaushaltes verantwortlich. Pantoffeltierchen leben im Süßwasser. Bedingt durch einen hohen Anteil an gelösten Stoffen im Cytoplasma (dadurch relativ weniger Wasser) kommt es zu einer ständigen Wasseraufnahme durch die Zelle. Die pulEM-Aufnahme Q: www.ebiomedia.com sierende Vakuole "sammelt" das überschüssige Wasser aus dem Cytoplasma. Die Vakuole ist durch ein feines Kanälchen mit der Außenwelt verbunden. Mittels einer Kontraktion wird der Inhalt der Vakuole in den extrazellulären Raum abgeleitet. Diese Kontraktionen finden in regelmäßigen Abständen statt. Deshalb spricht man von einer pulsierenden / kontraktilen Vakuole. (7 % 7 " ! 0 . . ( 4- % 0 & F 0 ( ( 0 # H C0 6 + +1 . ( " (4 1 + " % 0 - 97 - # - (c,p) 2008 lsp: dre 2.9. paraplasmatische (ergastische) Strukturen Membranumhüllte Strukturen, die hauptsächlich der Speicherung von Stoffwechselendprodukten dienen, werden paraplasmatische oder ergastische Strukturen genannt. Sie kommen nicht in allen Zellformen und je nach Organismengruppe sehr unterschiedlich vor. 2.9.1. Lipid-Tröpfchen Lipid-Tröpfchen sind membranumhüllte Mikropartikel. Sie sind auf Grund der Oberflächenspannung kugelförmig und zwischen 50 und 500 nm groß. Sie könnten Abschnürrungen des ER oder des GOLGI-Apparates sein. Lipid-Tröpfchen werden in den meisten Eucyten beobachtet. Q: de.wikipedia.org () 2.9.2. Stärkekörner In Speicherorganen (z.B. Kartoffelknollen) finden wir in den Zellen mit Iod-Kaliumiodid-anfärbbare rundliche Strukturen. Ähnliche Strukturen sind auch in Chloroplasten oder Leukoplasten nachweisbar. Hierbei handelt es sich um Stärkekörner. Im mikroskopischen Bild kann man oft sogar ringförmige Innenstrukturen erkennen. Diese sind – wie Baumringe – das Ergebnis unterschiedlich starker Speichervorgänge. Die Glucose aus der Photosynthese wird über die Leitbündel zu den Speicherorganen (auch zu Früchten) transportiert. Hier (sekundär) oder eben gleich (primär) in den Plastiden wird die Glucose hauptsächlich zu Amylose polymerisiert. Dieser eignet sich als Speicherstoff besser als Glucose, da sie wesentlich weniger wasserlöslich ist und Stärkekörner (Kartoffelknolle); gefärbt Q: de.wikipedia.org (Mnolf) damit nicht die osmotischen Verhältnisse verändert. Die primäre Stärke in Chroplasten wird auch Assimilationsstärke genannt. In Leukoplasten nennen wir die Stärke Speicherstärke. Stärkekörner in Speicherorganen sind von Leukoplasten-Membranen umgeben. - 98 - (c,p) 2008 lsp: dre 2.9.3. Pigmentgranula Pigmentgranula sind ebenfalls membranumschlossene Speicher-Partikel. Im Innenraum befinden sich verschiedenste Farbstoff – z.T. in kristalliner Form. In den relativ seltenen gelb bis rot gefärbten Lipophoren befinden sich Carotin-ähnliche Farbstoffe. Wesentlich häufiger kommen Melanophoren vor, die braune bis schwarze Farbstoffe enthalten. Hier ist besonders das Melanin zu nennen. Melanophoren werden vom GOLGI-Apparat abgeschnürt und sind zu Anfang nur mit wenig Melanin gefüllt. In den reifen Melanophoren befinden sich dann recht große Mengen Melanin. Mit Hilfe dieser Pigmentgranula können einige Tiere die Färbung ihrer Haut verändern. Aktiv tun dies z.B. die Kalmare. Bei ihnen wird die Hautverfärbung zur innerartlichen Kommunikation genutzt. Beim Menschen ist die Melaninfärbung eher passiv und genetisch bedingt. Bei erhöhter Sonneneinstrahlung (z.B. beim Sonnenbaden) wird die Melaninbildung und – einlagerung aktiviert. Q: de.wikipedia.org () 2.9.4. Sekretgranula Vom GOLGI-Apparat gebildete Sekrete können u.U. auskristallisieren und bilden mit der abgeschnürten Dyctosomen-Membran feste Sekretgranula. Sie werden zum Plasmalemma transportiert und hier in die Zellumgebung (z.B. Körperflüssigkeit, Blut, Drüsenflüssigkeit, …) abgegeben. Q: de.wikipedia.org () interessante(r) Internet-Link(s): http://multimedia.mcb.harvard.edu/anim_innerlife_Hi.html (Video über Zellbestandteile, … (engl.)) - 99 - (c,p) 2008 lsp: dre 2.10. kristalline und abiotische Zellbestandteile 2.10.1. Fett-Tropfen In Zellen gebildete Fette lösen sich fast nicht im vorwiegend wässrigen Mileu des Cytoplasmas. In feiner Form verteilt stellen sie eine Fett-in-Wasser-Emulsion dar. Bei Kontakt der kleinen Micellen verschmelzen diese langsam zu immer größerern Fettropfen. In tierischen Zellen können diese Tropfen beachtliche Ausmaße annehmen. Fetttröpfchen befinden sich meist im Zentrum der Zelle. Q: de.wikipedia.org () 2.10.2. Kristalle Bestimmte Stoffwechselprodukte bilden schwerlösliche Salze. Ein Beispiel ist das Calciumoxalat (ein Salz der Oxalsäure (Diessigsäure)). Das Calciumoxalt bildet große – z.T. kreuz- od. morgensternförmige – Kristalle. Den Kristallen konnte bis jetzt noch keine praktische Funktion zugeordnet werden. Andere Salze, die sich ebenfalls in Zellen niederschlagen, sind Siliciumdioxid und Calciumcarbonat. Beide Salze werden u.a. in der Zellwand abgelagert und leisten dabei einen beachtlichen Beitrag zur festigkeit der Zellwand. - 100 - Q: de.wikipedia.org () (c,p) 2008 lsp: dre