Thema61 Fettsaeuren 12
Werbung

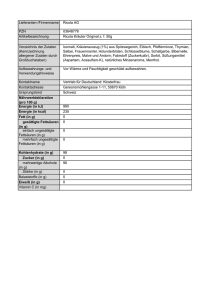
Biochemieseminar 6.1 Fettsäuren und Eikosanoide 6.1 erstellt von H. Wolfes und J. Alves Fettsäuren und Eikosanoide Definition: Gesättigte Fettsäuren haben die Summenformel CH3(CH2)n-2COOH. Meist ist n eine geradzahlige Zahl, weil die Fettsäuren aus C2-Einheiten (Acetyl-CoA) aufgebaut werden. Die der Carboxylgruppe benachbarte CH2-Gruppe wird mit α bezeichnet, die nächstfolgende mit β usw.. Der Methylgruppe am Ende der Fettsäure wird immer die Position ω zugewiesen. Abbau und Synthese der Fettsäuren laufen über die gleichen Zwischenstufen ab. Beim Abbau wird in der ß-Position eine Doppelbindung durch Elektronenübertragung auf FAD erzeugt. An diese wird Wasser angelagert, sodass die OH-Gruppe am ß-C-Atom steht. Diese wird dann durch Elektronenübertragung auf NAD+ zur Ketogruppe oxidiert. Eine solche ß-Ketoverbindung lässt sich leicht spalten. Im Stoffwechsel wird dabei Coenzym A eingeführt, sodass Acetyl-CoA und eine um 2 C-Atome verkürzte Fettsäure entstehen. Diese Oxidation geschieht in der Mitochondrienmatrix. Die Elektronen können so direkt in die Atmungskette eingeschleust werden. Die Synthese der Fettsäuren ist im Prinzip eine Umkehrung des Abbaus. Da sie im Cytosol statt- Vergleich von Abbau und Aufbau der Fettsäuren findet, nutzt sie NADPH + H+ als Elektronendonor für beide Reduktionen. Das NADP+/NADPHSystem wird in der Zelle im reduzierten Zustand gehalten (NADPH >> NADP+) während das NAD+/NADH-System immer oxidiert gehalten wird (NAD+>>NADH). So wirkt NADPH in 1 Biochemieseminar 6.1 Fettsäuren und Eikosanoide erstellt von H. Wolfes und J. Alves anabolen Reduktionen genauso wenig begrenzend wie NAD+ in katabolen Oxidationen. Ein weiterer kleiner Unterschied zwischen Auf- und Abbau liegt in der Stereochemie der ßHydroxysäure. Das spielt aber keine Rolle, da in den vor- und nachgeschalteten Strukturen das ßC-Atom jeweils in Doppelbindungen engagiert ist. Ein wesentlicher Unterschied besteht aber darin, dass in der Synthese nicht direkt Acetyl-CoA sondern Malonyl-CoA eingesetzt wird, das erst unter ATP-Verbrauch aus diesem gebildet werden muss. Der Grund für diese „Verschwendung“ liegt darin, dass die bei der Anlagerung der Acetyl-Gruppe erfolgende Freisetzung von CO2 die Reaktion irreversibel macht und so voran treibt. Bei Verwendung von Acetyl-CoA läge nämlich das Gleichgewicht nicht eindeutig auf einer Seite, wie man an der Bildung von Acetoacetyl-CoA durch die Thiolase bei der Ketonkörpersynthese sieht. Mit dem Malonyl-CoA hat man auch ein Zwischenprodukt, über dessen Bildung die Fettsäuresynthese reguliert werden kann und dessen Vorhandensein der Zelle signalisiert, dass Fettsäuren synthetisiert werden. Es kann so dafür genutzt werden, den Transport der gebildeten Acyl-CoA ins Mitochondrium zu hemmen, wo sie nur gleich wieder abgebaut würden. In unseren Zellen wird die Fettsäuresynthese von einem sehr großen multifunktionellen Enzym katalysiert, das neben den vier Reaktionen auch die Bindung der Intermediate an das Enzym gewährleistet, welches in einfacheren Organismen die Aufgabe eines eigenen Proteins ist: dem Acyl-Carrier-Protein (ACP). Damit geht auch einher, dass aus diesem Multienzym kaum kurzkettige Fettsäuren freigesetzt werden, sondern die Synthese in der Regel bis zur Palmitinsäure (C16) fortschreitet. In geringer Menge kann auch die Stearinsäure (C18) gebildet werden. Der Enzymbesatz der ß-Oxidation ist dagegen sehr viel komplexer. Da sich die Substrate durch die verschiedene Kettenlänge sehr stark in ihrer Löslichkeit und Hydrophobizität unterscheiden, haben die Zellen vier Enzymsysteme für kurzkettige (C4-C6), mittelkettige (C6-C10), langkettige (zwischen mittel- und sehr langkettig) und sehr langkettige (C12-C18) Substrate. Die Enzymaktivitäten für die Enoyl-CoA-Hydratase, die Hydroxyacyl-CoA-Dehydrogenase und die ß-Ketoacyl-CoA-Thiolase sind dabei in einem Enzymkomplex vereint. Pathobiochemie: Während keine Erbkrankheit der Fettsäuresynthese bekannt ist (die Fettsäuresynthase ist essentiell für Mäuse), gibt es einige seltene, rezessive Störungen der Fettsäureoxidation. Für alle Acyl-CoA-Dehydrogenasen sind Allele mit deutlich verminderter Aktivität bekannt. Ein MCAD-Mangel (Medium-Chain Acyl-CoA Dehydrogenase Mangel) ist aber bei weitem am häufigsten. Er wird in Deutschland in einem unter 15000 neugeborenen Kindern entdeckt. Seit in einem erweiterten Neugeborenenscreening massenspektrometrisch ein erhöhter Gehalt an mittel2 Biochemieseminar 6.1 Fettsäuren und Eikosanoide erstellt von H. Wolfes und J. Alves kettigen Acyl-Carnitinen im Blut nachgewiesen werden kann, stieg die Zahl der identifizierten Kinder, die homozygot für den MCAD-Mangel sind. Früher wurden sie hauptsächlich an den Symptomen erkannt, was häufig leider zu spät war. Allgemein sind die Kinder eher antriebslos und wenig ausdauernd. Es zeigt sich schon nach kurzem Fasten oder bei einer Infektion Erbrechen und Schlaffheit, die bis zum Koma gehen kann. Es kann auch zu einem Herzstillstand (Zusammenhang mit dem plötzlichen Kindstod) kommen und immer liegt eine Hypoglykämie vor. Im akuten Fall muss mit einer Glucosesubstitution gegengesteuert werden. Allgemein sollten die Patienten längere Intervalle ohne Essen vermeiden. Mit zunehmendem Alter können dabei längere Perioden (bei jungen Erwachsenen bis zu 12 Stunden) ohne Essen gut vertragen werden. Bei kleineren Kindern geht man von 4-6 Stunden zwischen den Mahlzeiten aus, die eher kohlenhydratreich und fettarm sein sollten. Einige Kinder profitieren auch von einer L-CarnitinGabe, das in Form von mittelkettigen Carnitinestern vermehrt mit dem Urin ausgeschieden wird. Ungesättigte Fettsäuren Gibt es im Kohlenstoffgerüst der Fettsäure eine Doppelbindung, spricht man von einer einfach ungesättigten Fettsäure. Wegen der Doppelbindung kann die Kette nicht mehr frei um die Achse zwischen den beiden Kohlenstoffen rotieren, damit wird die räumliche Anordnung fixiert. Liegen beide Substituenten der Kohlenstoffe auf der gleichen Seite der Doppelbindung, spricht man von einem cis-Isomer (Z wie zusammen), liegen die Substituenten gegenüber, liegt die trans-Konfiguration (E wie entgegen) vor. Die meisten in der Natur vorkommenden Fettsäuren liegen in der cis-Form vor. Die Lage der Doppelbindung wird durch den Buchstaben Δ angegeben. Δ3 bezeichnet daher eine Doppelbindung zwischen den Kohlenstoffen 3 und 4 einer Fettsäure. Mehrfach ungesättigte Fettsäuren enthalten die Doppelbindungen immer mit einer intermittierenden CH2-Gruppe. Doppelbindungen, die mehr als 9 Kohlenstoffatome von der Carboxylgruppe entfernt sind, können vom tierischen Organismus nicht eingeführt werden. Weil entsprechende Fettsäuren 3 Biochemieseminar 6.1 Fettsäuren und Eikosanoide erstellt von H. Wolfes und J. Alves aber wichtige Vorstufen für die Eikosanoidbildung darstellen, müssen sie mit der Nahrung zugeführt werden. Sie werden daher als essentielle Fettsäuren bezeichnet. Beispiele hierfür sind Linolsäure und Linolensäure. Alternative Nomenklatur mehrfach ungesättigter Fettsäuren: bei der Linolensäure (18 Kohlenstoffe) liegen weitere Doppelbindungen 12 bzw. 15 C-Atome von der Carboxylgruppe entfernt und demzufolge 6 bzw. 3 C-Atome vor dem endständigen ω-C-Atom. Daher kann man die Linolensäure auch als ω3,6,9 Octadecatriensäure bezeichnen. Es existiert ein breites Spektrum von Omega-Fettsäuren mit jeweils unterschiedlichen Effekten nach ihrer Umwandlung in Eikosanoide (siehe unten), die mit der Position der ersten Doppelbindung, der Länge der Fettsäurekette und der Anzahl der Doppelbindungen in Zusammenhang stehen. In der Leber können gesättigte in ungesättigte Fettsäuren überführt werden. Das hierfür notwendige Enzym ist die Desaturase. Der aktive, membranständige, in den Mikrosomen lokalisierte Komplex besteht aus einer NADPH-Cytochrom b5 Reduktase, Cytochrom b5 und der Desaturase. Von NADPH+H+ fließen 2 Elektronen auf das Coenzym FAD, welches zweimal das Eisenion im Cytochrom b5 zum Fe2+ reduziert. Diese Elektronen werden auf ein Eisenion der Desaturase übertragen und reagieren mit molekularem Sauerstoff, der zusätzlich noch 2 Elektronen von dem gesättigten Acyl-CoA aufnimmt. Somit werden 2 H2O und eine Doppelbindung erzeugt. Desaturasen tierischer Zellen können nur Doppelbindungen zwischen der Carboxylgruppe und dem Kohlenstoffatom 9 erzeugen. Weiter entfernt gelegene Doppelbindungen können nur von pflanzlichen Organismen synthetisiert werden. Aus Linolsäure kann aber zum Beispiel die Arachidonsäure durch Einfügung einer Doppelbindung an C6, Kettenverlängerung um 2 C-Atome und Einfügung einer weiteren Doppelbindung an C5 im menschlichen Organismus hergestellt werden. Aus Linolensäure 4 Biochemieseminar 6.1 Fettsäuren und Eikosanoide erstellt von H. Wolfes und J. Alves entstünde in der gleichen Reaktionsfolge eine Fettsäure mit 20 C-Atomen und 5 Doppelbindungen, die Eicosapentaensäure. Prostaglandine, Thromboxane und Leukotriene Prostaglandine, Thromboxane und Leu- kotriene werden unter dem Begriff Eicosanoide zusammengefasst. Sie sind Mediatoren mit einem breiten Wirkungsspektrum und modulieren hormonelle und sonstige Signale. Die Ausgangssubstanz der Eicosanoide sind mehrfach ungesättigte C20 Fettsäuren. Der wichtigste Vertreter ist die Arachidonsäure (Δ5,8,11,14), die an Position 2 der Phospholipide der Plasmamembran gebunden ist. Allerdings kommen dort auch die Dihomo-γ-Linolensäure (Δ8,11,14-Eicosatriensäure) sowie die Eicosapentaensäure (Δ5,8,11,14,17) vor. Die Phospholipase A2 spaltet diese ungesättigten Fettsäuren von den Phospholipiden ab. Sie sind Substrate von zwei Enzymen: der Cyclooxygenase und der 5’-Lipoxygenase. Es existieren drei Isoformen der Cyclooxygenase: die konstitutive COX-1, die COX-2, die bei Entzündungen induziert wird, und eine neu gefundene COX-3. Die Cyclooxigenasen synthetisieren ein Endoperoxid unter Ausbildung eines alizyklischen Fünfrings (PGG2). Das PGG2 wird von einer Peroxidase zum Prostaglandin H2 (PGH2) umgewandelt, dies ist die Ausgangssubstanz zur Synthese der Prostaglandine PGD2, PGI2, PGE2, PGF2 sowie des Thromboxans TxA2 mit einem Oxanring. 5 Biochemieseminar 6.1 Fettsäuren und Eikosanoide erstellt von H. Wolfes und J. Alves Der Index bezeichnet die Anzahl der noch vorhandenen Doppelbindungen und gibt so Aufschluss über die Ausgangsfettsäure; bei Index 2 war dies die Arachidonsäure während Index 3 auf die Eicosapentaensäure hinweisen würde. Leukotriene Leukotriene werden ebenfalls durch Modifikation der Arachidonsäure in wenigen Zelltypen wie Mastzellen, Granulozyten und Makrophagen synthetisiert. Aus Arachidonat bildet die 5-Lipoxygenase das 5-Hydroperoxyeicosatetraenoat (5-HPTE). Eine Umlagerung zum Peroxid führt zum Leukotrien A4, das weiter zum Leukotrien B4 umgewandelt wird. Wird Leukotrien A4 mit Glutathion gekoppelt, entsteht das Leukotrien C4, aus dem extrazellulär Leukotrien D4 und E4 erzeugt werden können. Auch bei den Leukotrienen bezeichnet der Index die Anzahl der Doppelbindungen und korrespondiert mit der Ausgangsfettsäure (4 ~ Arachidonsäure, 5 ~ Eicosapentaensäure). In einigen Geweben werden auch eine 12- und eine 15-Lipoxygenase gebildet. Funktionen der Eicosanoide Die Synthese und Ausschüttung der Eicosanoide kann durch Verletzungen, neurale Reize, Mediatoren wie Histamin oder durch gastrointestinale Hormone (Gastrin) ausgelöst werden. Die biologische Wirkung der Eicosanoide ist komplex und noch nicht vollständig verstanden. Sie sind in geringen Konzentrationen von 10-8 - 10-10 mol/L wirksam, die Effekte werden entweder auf parakrinem Wege auf benachbarte Zellen ausgeübt oder auf autokrinem Wege auf die 6 Biochemieseminar 6.1 Fettsäuren und Eikosanoide erstellt von H. Wolfes und J. Alves produzierende Zelle selbst. Wegen der kurzen Halbwertszeit der Mediatoren (Sekunden bis Minuten) ist die Reichweite begrenzt. Die Abkömmlinge der Arachidonsäure zeigen die stärksten Wirkungen während diejenigen der Eicosapentaensäure eher milder oder sogar entgegengesetzt wirken. Deswegen wird Patienten mit entzündlichen Gelenkerkrankungen empfohlen, Seefisch oder Fischöl zu sich zu nehmen, da darin ein hoher Anteil an ω3-Fettsäuren enthalten ist, die Entzündungen und Schmerzen lindern können. Auch die entzündungshemmende Wirkung des Kurkuma (Gelbwurz) wird darauf zurückgeführt, dass es die Bildung der Arachidonsäure aus Dihomogammalinolensäure hemmt. Letztere ersetzt dann jeweils die Arachidonsäure und ist auch Vorläufer von Prostaglandinen mit schwächerer Wirkung. Die Prostaglandine binden an spezifische, membranständige Rezeptoren, die mit einer Ausnahme das Signal an heterotrimere G-Proteine weitergeben. Einige Prostaglandinrezeptoren und ihre Effekte sind in der Tabelle zusammengefasst. Es ist bekannt, das PGD, PGF2 und TXA2 als die potentesten Konstriktoren der bronchialen Muskulatur wirken und für asthmatische Anfälle mitverantwortlich sind. PGE wirkt hier aber relaxierend. Ferner sind sie maßgeblich an der Entstehung entzündlicher Vorgänge und der Schmerzempfindung beteiligt. Sie bewirken Vasodilatation und Fieber und steigern die Kapillarpermeabilität und die Schmerzempfindlichkeit ohne direkt Schmerzen hervorzurufen. Außerdem spielen sie eine wichtige Rolle bei der Blutstillung, wobei das von den Thrombozyten gebildete Thromboxan A2 die Plättchenaggregation induziert, während das im Gefäßendothel gebildete Prostaglandin I2 (Prostacyclin) ihm direkt entgegenwirkt. Im Magen hat Prostaglandin E2 einen protektiven Effekt, indem es die HClSekretion hemmt. Die Wirkungen der Prostaglandine sind also sehr vielfältig, heterogen und gewebespezifisch. Rezeptor PGD2 PGE2 Subtyp EP1 Subtyp EP2 Subtyp EP3 PGF2α TxA2 PGI2 Effekt cAMP ↑ Organ Ileum IP3 ↑ cAMP ↑ cAMP ↓ IP3 ↑ cAMP↓ cAMP ↑ Nieren Thymus, Lunge, Myokard, Milz, Ileum, Uterus Fettgewebe, Magen, Nieren, Uterus Nieren, Uterus Thrombozyten, Thymus, Lunge, Nieren, Myokard Thrombozyten, Thymus, Myokard, Milz 7 Biochemieseminar 6.1 Fettsäuren und Eikosanoide erstellt von H. Wolfes und J. Alves Die Leukotriene A4, C4, D4 und E4 gehören zu den stärksten Konstriktoren der bronchialen Muskulatur. Leukotrien C4 ist um den Faktor 104 wirksamer als Histamin und spielt ebenfalls bei der Entstehung von Asthmaanfällen eine entscheidende Rolle. Die physiologische Funktion der Leukotriene ist weitgehend unverstanden. Sicher ist, dass sie an entzündlichen Prozessen beteiligt sind, sie erhöhen die Kapillarpermeabilität und führen zu Ödemen. Leukotrien B4 verursacht bei Leukozyten Chemotaxis, daher wird vermutet, dass es bei der Wanderung von weißen Blutkörperchen beteiligt ist. Oxidationen an Position 12 oder 15, die auch durch entsprechende Lipoxigenasen durchgeführt werden, führen zu den Lipoxinen, die gegenteilige, z.B. antiinflammatorische Wirkungen haben. Bei der vielfältigen Wirkung der Eikosanoide ist es nicht verwunderlich, dass eine Reihe von Therapeutika gibt, die zu ihrer Hemmung eingesetzt werden. Glucocorticoide induzieren die Synthese des Proteins Lipocortin, das spezifisch die Phopholipase A2 hemmt und seine Neusynthese senkt. Daher führt eine hohe Corticosteroidausschüttung zu einer Drosselung der Eicosanoidsynthese allgemein, betrifft also Prostaglandine und Leukotriene gleichermaßen. Derivate der Salicylsäure (Acetylsalicylsäure, Aspirin) hemmen die Cyclooxygenase und damit speziell die Synthese der Prostaglandine und des Thromboxans. Diese Hemmung ist irreversibel, da die Acetylsalicylsäure im Enzym kovalent gebunden wird, sodass es den Zugang zum aktiven Zentrum für die ungesättigten Fettsäuren aus der Membran blockiert. So erklärt sich der protektive Effekt bei drohender Bildung von Blutgerinnseln durch eine lang andauernde Gabe von Aspirin in sehr geringen Dosen. Die Thromboxanbildung wird inhibiert, da Plättchen irreversibel gehemmte Cyclooxygenase nicht mehr neu bilden können, während Endothelzellen immer wieder neues Enzym bilden, sodass eine Ausschüttung von Prostacyclin erhalten bleibt. Natürlich ist aus dem gleichen Grund eine schmerzlindernde Wirkung bei diesen Dosen ebenso wenig zu erwarten wie die für die Schmerztherapie typische Nebenwirkung einer Magenübersäuerung. Letztere findet man auch nicht bei COX-2 spezifischen 8 Biochemieseminar 6.1 Fettsäuren und Eikosanoide erstellt von H. Wolfes und J. Alves Inhibitoren. Ein weiteres Zielenzym des Wirkstoffs Acetaminophen (Paracetamol), der schmerzlindernd und fiebersenkend wirkt aber nicht an COX-1 oder COX-2 bindet, wurde postuliert. Ein Gen für eine COX-3 aber bisher nicht gefunden. Es scheint eine Splice-Variante von COX-1 zu sein. Ob die im Menschen zu einem funktionstüchtigen Enzym führt, ist noch nicht klar. Grundlegende Literatur Löffler, Basiswissen Biochemie, 7. Auflage S. 91-93, 99-113 Löffler Petrides Heinrich, Biochemie & Pathobiochemie, 8. Auflage S. 33-34, 403-425 Rassow Hauser Netzker Deutzmann, Biochemie, 3. Auflage S. 122-126, 225-231, 627633 Themen, die im Vortrag angesprochen werden sollten: • Vergleich von Fettsäureabbau und –synthese • MCAD-Mangel • Mehrfach ungesättigte Fettsäuren und Nomenklatur • Essentielle Fettsäuren • Synthese von Prostaglandinen aus Arachidonsäure • Synthese von Leukotrienen aus Arachidonsäure • Funktionen der Eikosanoide • Hemmstoffe - Medikamente • Bedeutung der ω3-Fettsäuren in der Nahrung 9