Die Reizleitung des menschlichen Nervensystems
Werbung

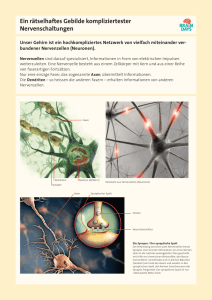
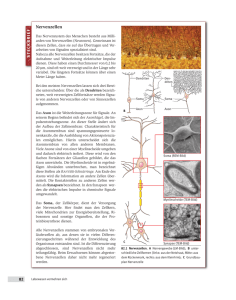
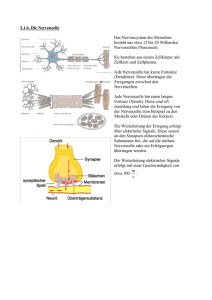
"Peter" <[email protected]> Die Reizleitung des menschlichen Nervensystems Aufbau des Nervensystems Die Reizleitung erfolgt über ein Nervensystem, das aus etwa hundert Milliarden Nervenzellen aufgebaut ist. Diese Zellen sind über den gesamten Körper (ungleichmäßig) verteilt und koordinieren die Aktivitäten sämtlicher Organe und Systeme. Das Nervengewebe dient zum Aufbau nervöser Zentren und peripherer Nerven, die die Aufgabe der Reizleitung von (efferent) und zu (afferent) diesen Zentren übernehmen. Die generell hohe Fortpflanzungsgeschwindigkeit der Impulse und der daraus resultierende hohe Energieverbrauch haben einerseits den hohen Spezialisierungsgrad der Zellen und andererseits die vornehmliche Eignung für kurzfristige Steuer- und Regelaufgaben zur Folge. Im allgemeinen besteht das Nervensystem aus den Neuronen und filamentartigen Fortsätzen, den Neuriten und Dendriten, die Impulse weiterleiten. Gestützt und versorgt wird dieses Informationssystem von den relativ unbekannten Gliazellen. Die Nervenzelle Bereits die ungewöhnliche Form der Zelle läßt Aufschlüsse über die Besonderheit der Zelle zu. Da die Reizleitung vor allem auf elektrischem Wege erfolgt, wird der innere Aufbau von dem Vorhandensein von Mitochondrien geprägt, um die potentialgenerierenden Systeme mit ausreichender Energie zu versorgen. Besagte Gliazellen versorgen in weiterer Folge die Zellen mit angemessen vielen Nährstoffen. Die Hauptmerkmale der Nervenzelle sind die Existenz der Dendriten oder Bäumchenfortsätze zur Aufnahme des Reizes und Verknüpfung mit vorhergehenden Nervenzellen, des Neurits oder Axons zur Weiterleitung des Impulses an weitere Nervenzellen oder verarbeitende Systeme und der Synapse zur Bildung einer interzellulären Verbindung zur Übermittlung des Reizes an andere Nervenzellen. Da Nervenzellen hochdifferenziert sind, ist eine Zellteilung und somit Vermehrung des Zellgewebes nur im embryonalen Stadium möglich. Nach Abschluß der embryonalen Entwicklung sind somit sämtliche Nerven vorhanden und eine Ersetzung abgestorbener Neuronen nicht mehr möglich. ! Der Zellkörper (Soma) Dem Zentrum jedes Neurons, dem Zellkörper, kommen vornehmlich Versorgungsaufgaben zu. Der grundlegende Aufbau entspricht selbstverständlich dem aller Körperzellen. Der zumeist kugel- oder pyramidenförmige Zellkörper beinhaltet den großen, ein klar erkennbares Kernkörperchen zeigenden, Zellkern. Im Zellplasma liegen die Nissl-Schollen (Tigroidsubstanz) und Neurofibrillen. Die Tigroidsubstanz, die sich bis in die Dendritenhügel erstreckt, stellt eine Art des endoplasmatischen Reticulums dar. Die Neurofibrillen sind kleinste impulsleitende Fäden, die die Information im Inneren des Zellkörpers weiterleiten. Weiters finden sich eine große Anzahl von Mitochondrien um die benötigte Energie bereitzustellen. Ferner sind auch Produktionsstellen für die Synthese von Enzymen anzutreffen. Eine wichtige Aufgabe kommt dem Zellkörper mit der Synthese von Membranproteinen zu: 2 Die Dicke der Membran beträgt so wie die aller Zellen etwa fünf Nanometer. Sie ist aufgebaut aus Molekülen, die je einen lipophilen (fettfreundlichen), also hydrophoben (wasserfeindlichen) und hydrophilen (wasserfreundlichen), also lipophoben (fettfeindlichen) Teil besitzen. Sie lagern sich so zusammen, daß die hydrophilen Enden zum Plasma und zum Äußeren der Zelle weisen, die lipophilen Enden sich gegenseitig im Inneren der Membran berühren. Diese Membran ist von besagten Proteinen durchbrochen oder versehen, die der Zelle so bestimmte Funktionen zuweisen und erst überhaupt gewisse Prozesse ermöglichen. Sie werden in fünf Gruppen gemäß ihrer Aufgaben eingeteilt: I. Pumpenproteine: Mittels Energieverbrauch können sie den Durchgang von Ionen und anderen Molekülen durch die Membran entgegen einem Konzentrationsgefälle ermöglichen. II. Kanalproteine: Sie ermöglichen es Ionen und anderen Molekülen die sonst undurchlässige Membran einem Konzentrationsgefälle gemäß zu passieren. III. Rezeptorproteine reagieren mit Molekülen wie beispielsweise mit Hormonen oder Neurotransmittern, die sodann Zellfunktionen beeinflussen oder aktivieren. IV. Enzyme haben eine Katalysatorfunktion für chemische Reaktionen an oder in der Membran. V. Strukturproteine verbinden Zellen zu einem Organ und erhalten die Feinstruktur der Zellmembran intakt. Diese Proteine werden mittels Vesikel gelagert und bei Bedarf zu ihrem Bestimmungsort innerhalb der Zelle gebracht. Im eigentlichen Prozeß der Reizleitung kommt dem Zellkörper nur eine untergeordnete Rolle zu – er stellt zwar die Verbindung zwischen den Empfängern, den Dendriten, und der Weiterleitung durch das Axon dar, jedoch ist seine primäre Funktion in der Versorgung zu sehen. ! Die Bäumchenfortsätze (Dendriten) Die Dendriten sind dünne, röhrenförmige Fortsätze, die sich vielfach verästeln. Sie stellen den Kontakt zu Nachbarzellen her, übernehmen Impulse und leiten sie in Richtung des eigenen Zellkörpers weiter. Eine einzige Nervenzellen kann einen einzigen Dendriten besitzen, meist jedoch sind es einige Tausend. Gemäß der Anzahl ihrer Dendriten werden sie als unipolar, pyramidenförmig oder multipolar bezeichnet. Dendriten stellen im allgemeinen die postsynaptische Membran dar: Sie sind mit den Neuriten, den weiterleitenden Teilen anderer Nervenzellen, mit Synapsen verbunden. An dieser Verbindung erfolgt die Weitergabe des Impulses meist nicht auf elektrischem sondern auf chemischem Wege. Die Bäumchenfortsätze dienen aber nicht nur der Weiterleitung von Impulsen sondern auch der Registrierung eines Reizes. Sie befinden sich auf Rezeptoren, die mit Nervenzellen eng verwandt sind. Diese Sinneszellen haben die Aufgabe, auf eine gewisse einwirkende Energieform mit deren Umwandlung in elektrische Impulse und einer Verstärkung zu reagieren. Im allgemeinen fußt die Rezeption eines Reizes auf einer Veränderung des Ruhepotentials, dem Ionengefälle zwischen Innen und Außen der Zelle. Diese meist in Form einer Potentialverringerung auftretende Gradientenänderung erfolgt durch einen Anstieg der Na+-Permeabilität der Membran. Die Umwandlung der Reizenergie in das Rezeptorpotential (die Änderung der Ionengefälles) wird Transduktion, Transformation die Umwandlung in ein für Nervenzellen weiterleitbares Aktionspotential genannt. 3 ! Die Nervenfaser (Axon, Neurit) Das Axon ist das impulsweiterleitende Element der Nervenzelle. Es empfängt den Impuls über die Dendriten und das Soma und überträgt ihn über seine beachtliche Länge an weitere Zellen. Entsprechend dem empfangenden Gegenstück unterscheidet man axo-somatische, axo-axonische und axo-dendritische Übermittlung. Diese Übertragungsstelle ist die Synapse. Durchschnittlich besitzt jede Nervenzelle etwa tausend bis zehntausend dieser Verbindungsstellen. Der genaue Vorgang der Impulsübertragung an der Synapse wird später behandelt. Sollten Axone nicht auf eine andere Nervenzelle, sondern auf einen Muskel treffen, um dort Aktionen einzuleiten, spricht man von einer motorischen Endplatte. Jedes Neuron besitzt genau ein Axon, jedoch kann sich dieses in einiger Entfernung vom Zellkörper in etliche Verzweigungen aufspalten. Diese Kollaterale genannten Endigungen ermöglichen eine Impulsweitergabe in großem Umfang. Aufgrund seiner Bestimmung für die Impulsleitung enthält das Axon keine Tigroidsubstanz, jedoch Neurofibrillen. Ein weiterer Unterschied zu den Bäumchenfortsätzen besteht in seinen Ausmaßen: Es ist um Vieles länger und etwas dünner. Das Axon, das aus dem Soma am Axonhügel entspringt, kann eine Länge von bis zu einem Meter erreichen. Ein bedeutender Bestandteil einiger Nervenzellen ist die Myelinscheide. Das ist eine lipidhaltige Schicht, die von den umgebenden Schwannschen Scheidezellen um die Längsachse des Axons aufgebaut wird. Die Scheidezellen sind eine spezielle Art der Gliazellen. Ihr Zweck ist die Beschleunigung der Impulsleitung in den Axonen. Axone, die einzeln von Schwannschen Zellen mit einer Myelinhülle umgeben sind, werden markhältig genannt. Als marklos bezeichnet werden Nervenbahnen, deren Axone nur von einer Schicht Schwannscher Zellen umgeben sind und deren Bündelung erst von einer Myelinscheide umgeben ist. Markhaltige Fasern stellen den Großteil der Gehirn- und Rückenmarkssubstanz. Zumal sie eine erhöhte optische Reflektivität aufweisen, wird diese Substanz auch als weiß bezeichnet. Marklose Fasern hingegen erscheinen eher grau und sind im besonderen die postganglionären Fasern des autonomen (vegetativen) Nervensystems. Periphere Nervenfasern beiderlei Typs werden durch Bindegewebshüllen (Perineurum) zu dickeren Kabeln gebunden. Diese wiederum formen größere Nerven, welche ebenso durch Bindegewebe (Epineurium) zusammengehalten werden. Im Nerv befinden sich des weiteren Versorgungselemente, zum Beispiel Blutgefäße. Das Epineurium schützt sämtliche beteiligten Nervenzellen vor mechanischen Beschädigungen wie beispielsweise Überstreckung. Die Myelinscheide des Axons wird periodisch (all 1-2 mm) von den Ranvierschen Schnürringen unterbrochen. Sie stellen die Enden der Schwannschen Scheidezellen dar. Eine ihrer Funktionen ist die Versorgung der Zellmembran mit Nährstoffen. Die Hauptaufgabe besteht jedoch darin, die Leitung des elektrischen Impulses zu beschleunigen: Der Impuls pflanzt sich nicht kontinuierlich durch das ganze Axon fort, sondern springt von Schnürring zu Schnürring. Da die lipoide Myelinschicht kaum ionendurchlässig ist, ist in diesem Bereich eine Erregung des Potentials nahezu unmöglich. Daher überspringt der Impuls diese Gebiete, was diesem Vorgang die Bezeichnung der saltatorischen Erregungsleitung einbringt. Marklose Fasern leiten Impulse mit einer Geschwindigkeit von 0,5 m/s bis 30m/s. Dickere Fasern ermöglichen eine schnellere Leitung, jedoch ist dem eine praktische Grenze von einer Dicke von 1 mm gesetzt, was 30 m/s entspricht. Daß die saltatorische Erregungsleitung eine große Rolle bei genauen und raschen Koordinationsprozessen spielt, beweist die typische Geschwindigkeit von 120 m/s. 4 Die Gliazellen Die Stütz-, Schutz- und Ernährungsfunktion wird von den Gliazellen erfüllt. Das Verhältnis von Nervenzellen zu Gliazellen beträgt etwa 1:50. Gliazellen können auch Heilungsprozesse beschädigter Nerven durch Vernarbung begünstigen. Der Raum zwischen den verletzlichen Neuronen wird von ihnen erfüllt, sodaß sie als direktes Bindeglied zwischen Neuronen und Versorgungselementen fungieren. Die Nervenzellen des menschlichen Gehirns verbrauchen pro Tag etwa 80 g Traubenzucker, was der 16fachen Menge des Blutzuckergehaltes unseres Blutes entspricht. Das macht eine effiziente Nährstoffversorgung unabdingbar. Die Kapillaren des Gehirns werden von Gliazellen umfaßt, die diese Nährstoffe direkt und wohldosiert an die Neuronen abgeben. Dieser Mechanismus sieht folgendermaßen aus: Durch die Durchblutung der Haargefäße des Gehirns nehmen die anliegenden Gliazellen Traubenzucker und Glykogen aus dem Blut auf. Sobald sich ein gewisser Grad der Sättigung eingestellt hat, wird die Blutzufuhr langsam durch die anschwellende Gliazelle unterbunden. Somit erhält die Gliazelle weniger Nährstoffe, wird aber gleichzeitig von der aktiven Nervenzelle vorhandener Zucker- und Glykogenmoleküle beraubt. Dadurch schwillt die Gliazelle wieder ab, erhöht die Blutversorgung und reichert sich mit Nährstoffen an. Eine weitere Art der Gliazellen ist die Schwannsche Scheidezelle: Sie wickelt sich im Laufe der embryonalen Entwicklung um das Axon und baut die fettartige Myelinhülle auf. Sie ermöglicht eine weitaus schnellere Leitung des Impulses. Im allgemeinen läßt sich jedoch sagen, daß über die speziellen Funktionen der Gliazellen weniger bekannt ist als über Nervenzellen. Die Impulsleitung Die Leitung vom Empfänger zum Zentrum wird von empfindungsleitenden (afferenten, zentripetalen, sensiblen) Fasern durchgeführt. Die Leitung von Zentren zu ausführenden Organen wird von bewegungsauslösenden (efferenten, zentrifugalen, motorischen) Fasern hergestellt. Signale, die in Nervenfasern geleitet werden, sind elektrische Impulse. Zur Übermittlung an andere Zellen, wie beispielsweise an den Synapsen, bedient sich das Nervensystem auch chemischer Transmissionstechniken. Das Ruhepotential Das Vorhandensein des Ruhepotentials ist die Grundlage für eine rasche Impulsleitung. Es ermöglicht die Leitung eines Impulses ohne eine unmittelbar auf den Reiz folgende Generierung eines elektrischen Feldes; somit wird eine extrem rasche Reaktion auf den einwirkenden Reiz möglich. Das Ruhepotential stellt sozusagen einen leicht anzapfbaren Energiespeicher dar. Die Ausgangssituation ist sowohl im Inneren der Zelle als auch im Äußeren eine Flüssigkeit, in der K+- und Na+-Ionen gelöst sind, wobei das Äußere etwa zehnmal so viele Na+-Ionen wie das Plasma 5 enthält; das Konzentrationsgefälle der Kaliumionen ist umgekehrt und beträgt etwa 40:1. Um diesen Gradienten zu erhalten, ist die Zellmembran mit sogenannten Pumpen durchsetzt, die Ionen durch die Membran selektiv durchlassen. Dieser Ionenflux erfolgt in fixierter Stöchiometrie, d. h. es werden gleichzeitig drei Natriumionen aus der Zelle und zwei Kaliumionen in die Zelle transferiert. Ein stöchiometrisch fixierter Transport von Ionen wird als Cotransport bezeichnet, im speziellen, dem gegengerichteten Transport gleichsinnig geladener Ionen, als Antiport. Sollten bei diesem Vorgang Ionen in gleichem stöchiometrischem Anteil bewegt werden, so würde er als elektroneutral bezeichnet werden. Im vorliegenden Fall jedoch wird ein elektrischer Gradient erreicht, weswegen der Pumpvorgang rheogen genannt wird. Hier erfolgt der Ionentransport weiters noch aktiv: Der Transportprozeß wird durch energieliefernde Stoffwechselvorgänge ermöglicht. Die Kalium-Natrium-Pumpen sind Transport-ATPasen – Energie wird aus der Hydrolyse von ATP zu ADP bezogen. Aktive rheogene Pumpen werden des weiteren auch als elektrogen bezeichnet, da sie ein Transmembranpotential durch gekoppelten Flux generieren. Die Kalium-Natrium-Pumpe, die eine relative Molekülmasse von 275000 aufweist, kann rund 100 Na+-Ionen und 130 K+-Ionen in der Sekunde transportieren. Eine kleine Nervenzelle erreicht, da sie etwa eine Million dergestalter Pumpen aufweist, eine Transportleistung von bis zu 200 Millionen Na+Ionen pro Sekunde. Nun ist aber die Permeabilität der Membran für die K+-Ionen, die ein recht starkes Konzentrationsgefälle aufweisen, besonders hoch. Da zu jedem Kation aber auch ein entsprechendes Anion gehört, das jedoch nicht die Zellmembran durchdringen kann, entsteht kontinuierlich ein Anionenüberschuß in der Zelle. Ein elektrisches Feld entsteht, das bei Vorhandensein eines gewissen Gradienten keinen weiteren Kationen-Ausstrom gestattet. Dieses Feld wird als Membranpotential bezeichnet, welches im Ruhezustand etwa -70 mV beträgt. Dieses im Inneren der Zelle stets vorhandene negative Ruhepotential ist der Ausgangspunkt für eine sofort verfügbare Energie zu Weiterleitung des eingehenden Impulses. Das Aktionspotential Ein eingehender Reiz kann nun bewirken, daß das Membranpotential einen positiveren Wert als –50 mV erreicht. Ab dieser Spannung nämlich öffnen sich Kanäle in der Membran und eine plötzlicher Ionenstrom gemäß dem vorhandenen Gefälle setzt ein. Ändert sich an einer Nervenfaser zunächst örtlich begrenzt die Spannungsdifferenz auf besagte –50 mV, so aktivieren sich erst die Natriumkanäle ein Stück weiter in Richtung der vorgesehenen Leitrichtung. Derartige Kanäle sind proteingesteuerte Passagen in der Membran, in denen freier Durchlaß speziellen Molekülen gewährt wird. Daraufhin strömen in großem Umfang Na+-Ionen in das Zellinnere, das somit positiv auf 30 mV geladen wird. Sogleich erfolgt die Schließung dieser Kanäle, und andere Kanäle öffnen sich. Diese lassen nun K+-Ionen nach außen ausströmen. Der Ausstrom von Kationen bewirkt somit eine neuerliche negative Ladung der Nervenzelle. Diese Depolarisierung des Membranpotentials pflanzt sich äußerst schnell durch das ganze Axon fort. Der Overshoot von –70 mV auf +30 mV erfolgt in etwa 0,5-1 ms; dem ebenso raschen Abfall folgt eine Refraktärphase, in der die Nervenzelle noch nicht wieder erregbar ist. Sie gliedert sich in die absolute Refraktärphase, in der keine weitere Leitung möglich ist, da die Zelle erst im Begriff ist, das Ruhepotential zu regenerieren, und die relative, in der die Reizschwelle gehoben ist, d. h. ein stärkerer Reiz einwirken muß, um eine Fortpflanzung einzuleiten. Die Refraktärphase verhindert somit auch die Möglichkeit, daß der Impuls in der Nervenzelle rückwärts laufen könnte. Eine weitere Einschränkung des Alles-oder-Nichts-Prinzips ist die Tatsache, daß auch unterschwellige Reize ein Aktionspotential auslösen können, wenn mindestens zwei in einem Zeitraum von maximal 100 ms die Nervenzelle erregen. Dieses Phänomen wird als Summation der Reizwirkung bezeichnet. 6 Das Rezeptorpotential, das durch den Reiz generiert wird, ist proportional zur physikalischen Reizstärke. Das Aktionspotential hingegen weist stets die gleiche Amplitude auf. Je stärker der Reiz ist, desto höher ist das Rezeptorpotential und desto öfter wird ein Aktionspotential ausgelöst. Die ursprüngliche Information der Reizstärke ist nun als Frequenz (Aktionspotentiale/Zeiteinheit) verschlüsselt. Wäre die Höhe des Aktionspotentials der Informationsträger (Amplitudenmodulation), würde die Potentialhöhe bei langen Leitstrecken (beim Menschen bis zu 1 m Länge) viel leichter verändert und die Information verfälscht. Daher ist die Frequenzmodulation ein sichereres Übertragungsmittel. An der nächsten Synapse wird die frequenzmodulierte Information wieder demoduliert. Je höher diese Frequenz, desto mehr Neurotransmitter werden freigesetzt. In marklosen Fasern ist die Geschwindigkeit des Impulses proportional zur Quadratwurzel des Durchmessers. Eine Vervierfachung des Querschnitts führt also zur Verdoppelung der Impulsgeschwindigkeit. Die saltatorische Leitung jedoch ermöglicht eine weitaus schnellere Leitung aufgrund der sprunghaften Fortpflanzungsart, wenn auch damit ein erhöhter Energieverbrauch einhergeht. Vorgänge an der Synapse Die Übertragung von Impulsen von Zelle zu Zelle findet an speziellen Strukturen, den Synapsen, statt. Neben elektrischen Synapsen, die Verbindungen durch Ionendiffusionen herstellen, sind auch und vor allem die chemischen bekannt. Grundsätzlich bestehen Synapsen aus dem präsynaptischen Element (zumeist ein Axon), dem synaptischen Spalt, und dem postsynaptischen Element (meist ein Dendrit). An den Synapsen erfolgt die Übertragung des Impulses meist auf chemischem Wege: Der ankommende Impuls veranlaßt das synaptische Endknöpfchen, sogenannte Transmittermoleküle, allen voran Acetylcholin und auch Noradrenalin, auszuschütten. Mit dem Eintreffen des Aktionspotentials öffnen sich im synaptischen Knöpfchen durch die sofort stattfindende Depolarisation Ca++-Kanäle. Die einströmenden Ionen veranlassen die Ausschüttung von Neurotransmittern; die Ca++-Ionen werden sofort chemisch gebunden. Diese Stoffe befinden sich in Vesikeln, mit deren Hilfe sie sowohl gespeichert, als auch rasch in den synaptischen Spalt entleert werden können. Auf ein Zeichen hin bewegt sich die membranartige Hülle in Richtung des synaptischen Spaltes. Dort verschmilzt die Hülle des Transportvesikels mit der Zellmembran und der Inhalt ergießt sich in den synaptischen Spalt. Dieser Vorgang der Verschmelzung und Entleerung nennt man Exozytose. Die Neurotransmitter diffundieren in weniger als 100 µs durch den 0.2 nm breiten Spalt und reagieren mit der postsynaptischen Membran. Rezeptoren an der postsynaptischen Membran registrieren die ausgeschütteten Neurotransmitter und öffnen Ionenkanäle. Diese komplexen Proteine werden deswegen neurotransmitter-kontrollierte Ionenkanäle genannt, weil ihnen die Reaktion mit Neurotransmittern und das Vorhandensein von Ionenkanälen gemein ist. Durch die Andockung des Transmittermoleküls ändert das Molekül seine Konformation, wodurch die Kanalkomponente eine Passage des ausgewählten Ions ermöglicht. Das Molekül des Acetylcholinrezeptors ist aus fünf weiter unterteilbaren Untereinheiten aufgebaut, von denen zwei mit Acetylcholin besetzt werden müssen, um den Kanal zu öffnen. Ein weiterer interessanter Aspekt des Moleküls ist seine offenbare Lernfähigkeit: Das Molekül ist in der Lage, seine Konformation so abzuändern, daß es auf den Botenstoff verschieden stark anspricht. Einerseits ist diese Fähigkeit ein Schutz vor Übererregung, andererseits kann das Molekül wahrscheinlich die Signalübertragung an der Synapse steigern oder drosseln, was einem Lernprozeß gleichkäme. Das freigesetzte Acetylcholin bewirkt zumeist eine Öffnung des Ionenkanals für eine Dauer von etwa 1 ms. Der Inhalt eines präsynaptischen Transportvesikels öffnet ungefähr 2000 Kanäle; im Falle einer motorischen Endplatte strömen je etwa zwanzigtausend Na+-Ionen in die Muskelzelle. 7 Man unterscheidet ferner zwei Arten von chemischen Synapsen: Erregende und hemmende: Wenn Kationen die Ionenkanäle passieren und eine positive Ladung im postsynaptischen Element initiieren, handelt es sich um erregende Synapsen. Hier wird ein Aktionspotential durch die Veränderung des Ruhepotentials in positiver Richtung erzeugt. Hemmende Synapsen hingegen schleusen positive Ionen aus dem Zellinneren heraus oder bringen negative Ionen ins Innere. Da das einer Initialisierung eines Aktionspotentials entgegenwirkt, ist hier von hemmenden Synapsen die Rede. Durch die eindeutige Unterscheidung von prä- und postsynaptischem Element läßt sich eine klare Richtung der Impulsleitung bestimmen. 8 Verwendete Literatur AULICH Dieter – DECKERS Heinrich, Elektrophysiologisches Praktikum. Wiesbaden 1995. CHANGEUX Jean-Pierre, Der Acetylcholin-Rezeptor. In: GOODY Roger S. (Hg.), Proteine. Beiträge aus Spektrum der Wissenschaft. Heidelberg – Berlin – Oxford 1995. FALLER Adolf, GLASER Roland, Biophysik. 4., völlig überarb. Aufl. Jena – Stuttgart 1996. MANDL Lothar, Organismus und Umwelt 2. Wien 1996. ROTHMAN James E. – ORCI Lelio, Knospung von Transportvesikeln in Zellen. In: Spektrum der Wissenschaft 5/1996. Heidelberg 1996. STEVENS Charles F., Die Nervenzelle. In: Spektrum der Wissenschaft