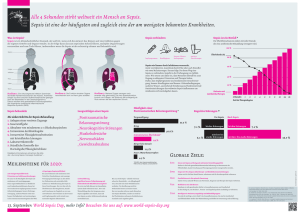
Untersuchungen zur T-Lymphopoese während polymikrobieller
Werbung

Universitätsklinikum Ulm Klinik für Anästhesiologie Ärztlicher Direktor: Prof. Dr. med. Dr. med. h. c. M. Georgieff Untersuchungen zur T-Lymphopoese während polymikrobieller muriner Sepsis Dissertation zur Erlangung des Doktorgrades der Medizin der Medizinischen Fakultät der Universität Ulm Vorgelegt von Richard Alexander Blothner aus Wangen im Allgäu 2011 Amtierender Dekan: Prof. Dr. Thomas Wirth 1. Berichterstatter: Prof. Dr. Uwe Senftleben 2. Berichterstatter: PD Dr. Mario Perl Tag der Promotion: 10.02.2012 Für meine lieben Eltern Inhaltsverzeichnis Inhaltsverzeichnis I Abkürzungsverzeichnis II 1 Einleitung ........................................................................................... 1 1.1 Sepsis ....................................................................................................... 1 1.2 Rolle der T-Lymphozyten bei der Pathogenese der Sepsis ...................... 4 1.3 Fragestellung und Zielsetzung ................................................................ 10 2 Material und Methoden ................................................................... 12 2.1 Chemikalien und Medikamente ................................................................... 12 2.2 Lösungen .................................................................................................... 12 2.3 Antikörper .................................................................................................... 12 2.4 Methoden .................................................................................................... 13 3 Ergebnisse ....................................................................................... 25 3.1 Überlebenszeitanalyse ................................................................................ 25 3.2 Auswertung der Narkoseprotokolle der Immunphänotypisierung ................ 27 3.3 T-Lymphozyten im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere .................................................................................................. 30 4 Diskussion ....................................................................................... 66 4.1 Einleitung .................................................................................................... 66 4.2 Überlebenszeitanalyse ................................................................................ 67 4.3 Auswertung der Narkoseprotokolle der Immunphänotypisierung ................ 68 4.4 T-Lymphozyten im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere .................................................................................................. 69 4.5 Schlussfolgerung und Ausblick ................................................................... 77 5 Zusammenfassung ......................................................................... 79 6 Literaturverzeichnis ........................................................................ 81 7 Danksagung .................................................................................... 86 8 Lebenslauf ....................................................................................... 87 I Abkürzungsverzeichnis Abb. Abbildung AD Außendurchmesser ANOVA Analysis of variance, Varianzanalyse APC Aktiviertes Protein C APZ Antigen-Präsentierende-Zellen Aqua dest. Destilliertes Wasser Bcl B-cell leukaemia/Lymphoma related protein BrdU Bromdesoxyuridin BSA Bovine Serum Albumin bzw. Beziehungsweise C5a Komplementfaktor 5a CD Cluster of differentiation CLP Cecal ligation and punction DN Doppel-negativ DP Doppel-positiv € Euro EDTA Ethylendiamintetraacetat EP Einfach-positiv FACS Fluorescense Activated Cell Sorting FITC Fluorescein-Isothiocyanat FL Fluoreszenz Foxp3 Forkhead-Box-Protein P3 FSC Forward Scatter Channel, Vorwärtsstreulicht G Gauge gE Erdbeschleunigung GITR Glucocorticoid-induced TNFR-related protein h Hour, Stunde IFN-γ Interferon-γ IL Interleukin IPEX Immune dysregulation, Polyendocrinopathy, Enteropathy X-linked syndrome kg Kilogram II l Liter m Milli (10¯3) M Molar max Maximal MHC Major Histocompatibility Complex min Minimal Min. Minute ml Milliliter mm Millimeter MODS Multi Organ Dysfunction Syndrome NF-κB Nukleärer Faktor kappa B NO Nitric oxid Nr. Nummer Op Operation PBS Phosphat Buffered Saline, Pufferlösung PerCP Peridin-Chlorophyll-a Proteinkomplex PFA Paraformaldehyd RAG Recombination activation gene rh APC rekombinantes humanes aktiviertes Protein C R-PE R-Phyoerythrin µ Mikro (10¯6) s.c. Subcutan sh. Siehe SIRS Systemic Inflammatory Response System SSC Sidewards Scatter Channel, Seitwärtsstreulicht Tab. Tabelle TGF-β Transforming Growth Factor beta T H1 T-Helferzelle Typ 1 T H2 T-Helferzelle Typ 2 TNF-α Tumor Nekrose Faktor α Tregs Regulatorische T-Zellen vs. Versus z.B. Zum Beispiel III IV 1 Einleitung 1.1 Sepsis Die Sepsis stellt in der Intensivmedizin eine der größten Herausforderungen dar. Trotz intensiver Forschungsbemühungen weist die Sepsis eine hohe Letalitätsrate und weltweit steigende Inzidenz auf [6, 9]. Neben dem Verständnis des Pathomechanismus, ist insbesondere die Entwicklung von Therapiestrategien zur Senkung der Letalität von großer Bedeutung. 1.1.1 Definition Die Sepsis wird als ein lebensbedrohliches, klinisches Syndrom angesehen, welches durch eine systemische inflammatorische Reaktion des Körpers auf eine Infektion verursacht wird [19]. Die aktuelle Definition der Sepsis wurde 1992 auf einer internationalen Konsensuskonferenz des American College of Chest Physicians und der Society of Critical Care Medicine festgelegt [8]. Durch das Fehlen beweisender Parameter zur Diagnosestellung, werden Sepsis, schwere Sepsis und septischer Schock als Krankheitskontinuum angesehen, das anhand der Kombination klinischer Symptome und Vitalparameter der SIRS (Systemic Inflammatory Response Syndrome) mit Laborparametern, Organfunktionen und der Hämodynamik definiert wird. 1.1.2 Epidemiologie und Kosten der Sepsis Trotz intensiver Forschungsbemühungen und großer Fortschritte bleibt die Sepsis ein häufiges und schwerwiegendes Krankheitsbild. In einer Studie aus dem Jahr 2007 konnten Engel et al. eine Prävalenz von 12,4 % für die Sepsis und 11 % für die schwere Sepsis und den septischen Schock in der Bundesrepublik Deutschland feststellen. Die geschätzte jährliche Neuerkrankungsrate lag für die Sepsis zwischen 85 und 116 Fällen und für die schwere Sepsis zwischen 76 und 110 Fällen pro 100.000 Einwohner. Mit einer Letalität von 55,2 % war die Sepsis die dritthäufigste Todesursache [9]. Da die Inzidenz und die Letalität der Sepsis mit dem Alter zunehmen, ist bei der aktuellen demographischen Entwicklung noch von einem weiteren Anstieg auszugehen [1]. 1 Dies stellt nicht nur die Intensivmedizin vor schwierige Aufgaben, sondern ist auch aus gesundheitsökonomischer Sicht relevant. Aufgrund einer langen Liegedauer der Patienten und einer teuren intensivmedizinischen Diagnostik und Behandlung sind die direkten anteiligen Kosten außerordentlich hoch. Zusammen mit den indirekten Kosten, dem volkswirtschaftlichen Produktivitätsausfall, entstehen durch die Sepsis jährlich zwischen 3,6 und 7,8 Milliarden € Kosten für die deutsche Gesellschaft [29]. Somit gehört die Sepsis zu den kostenintensivsten Krankheitsbildern der modernen Intensivmedizin. 1.1.3 Pathophysiologie der Sepsis Der Entstehung einer Sepsis liegt eine Dysregulation der induzierten Immunantworten auf eine Infektion zugrunde. Anfangs betrachtete man eine unkontrollierte inflammatorische Reaktion mit daraus resultierenden Gewebeschäden und Organdysfunktionen als Auslöser der Sepsis [19]. Die zu Beginn stattfindende Aktivierung von Entzündungszellen des angeborenen Immunsystems und die exzessive Produktion proinflammatorischer Mediatoren wurden dabei als ursächlich für die Entstehung des SIRS angesehen [39]. In mehreren klinischen Studien mit Substanzen, die die Entzündungskaskade blockieren, wie z.B. TNF-α-Antagonisten oder IL-1-Rezeptorantagonisten, erhöhte sich jedoch die Mortalität septischer Patienten. Durch den fehlenden Erfolg antiinflammatorischer Substanzen, wird die alleinige Rolle proinflammatorischer Zytokine in der Pathogenese der Sepsis in Frage gestellt [11, 12, 19, 39]. Heute wird ein zweiphasiger Verlauf der Sepsis favorisiert: In der Frühphase ist die Sepsis durch den starken Anstieg proinflammatorischer Mediatoren (TNF-α, IFN-γ, IL-1, IL-6, IL-12) charakterisiert. Mit Fortbestehen der Sepsis stellt sich in der Spätphase eine kompensatorische Verschiebung in Richtung antiinflammatorischen immunsuppressiven Status ein (Abbildung 1). 2 eines Auslöser Endothelzellen Infektion Trauma Schock Makrophagen Epithelzellen Intensität der Immunantwort Gefäßpermeabilität Intravasale Gerinnung Tachypnoe Leukozytose Fieber Tachykardie T-Helferzelle Typ 1: proinflammatorische Zytokine Reaktive Sauerstoffspezies Gewebsschäden Organversagen T-Helferzelle Typ 2: antiinflammatorische Zytokine Lymphozytenapoptose Infektanfälligkeit Hyperreaktive Immunantwort Hyporeaktive Immunantwort, Immunparalyse Dynamischer Zeitverlauf der Immunantwort während der Sepsis Abbildung 1: Unterschiedliche Stimuli können zur Aktivierung verschiedener Zelltypen und somit zur exzessiven Produktion proinflammatorischer Mediatoren führen. Gesteigerte Produktion proinflammatorischer Mediatoren, erhöhte Gefäßpermeabilität, Gewebsschäden und Organversagen sowie fehlerhafte Funktion des angeborenen Immunsystems verursachen eine Infektanfälligkeit in der späten hyporeaktiven Phase der Immunantwort, die mit einer Immunparalyse vergesellschaftet sein kann. Abbildung modifiziert nach Riedemann [31]. Dies erklärt sich zum einen durch die Verschiebung von TH1- zu TH2-CD4+ TZellen während proinflammatorischer der Sepsis. Zytokinen Dieser zu Wechsel Zytokinen mit von der Synthese antiinflammatorischen Eigenschaften wie IL-10, stellt ein möglicher Pathomechanismus bei der Entstehung der Immunparalyse in der Spätphase der Sepsis dar [19, 39, 50]. Des Weiteren deuten neuste Studien darauf hin, dass CD4+CD25+ regulatorische T-Zellen und CD8+ zytotoxische T-Zellen die Fähigkeit besitzen eine adaptive Immunantwort zu unterdrücken und somit an der Immunparalyse beteiligt sein könnten [51]. Wichtigster Aspekt der Immundysfunktion in der Spätphase der Sepsis ist die erhöhte Apoptoserate von Lymphozyten, die mit der Schwere der Erkrankung korreliert [39]. So findet man in murinen Lymphozyten des Knochenmarks, des 3 Thymus und der Milz 24 Stunden nach Sepsisinduktion mittels zökaler Ligatur und Punktion (CLP) erhöhte Apoptoseraten [2, 20]. Hotchkiss et al. zeigten in Autopsien an Sepsis Verstorbener, einen Abfall der B-Zellen, CD4+ T-Zellen und dendritischer Zellen, was zu einer Abnahme der Antikörperproduktion, der Aktivierung von Makrophagen und der Antigenpräsentation führte [19]. Die Hypothese, dass die Immundepression als Folge des Verlustes an Lymphozyten ein zentraler Pathomechanismus der Sepsis darstellt, wird durch die erhöhte Mortalität recombination activation gene RAG1-/- Lymphozyten-defizienter Mäuse in der Sepsis bekräftigt [22]. Als klinischer Hinweis auf eine gestörte Immunkompetenz im Sinne einer Immunparalyse, kann die Tatsache aufgefasst werden, dass bei protrahierter Sepsis zunehmend opportunistische bzw. nosokomiale Infektionen zu beobachten sind [36]. Neben inflammatorischen und antiinflammatorischen Reaktionen des Immunsystems, spielt die Gerinnung eine weitere Rolle in der Pathogenese der Sepsis. Eine Hyperkoagulabilität, die neben Mikrothromben in 30-50 % der Fälle zur disseminierten intravasalen Gerinnung führt, trägt zu Organdysfunktionen bei [7, 39]. Die Therapie mit rhAPC, das ein gerinnungs-, entzündungs- und apoptosehemmendes Wirkprofil aufweist, verdeutlicht diesen Zusammenhang [32]. Ziel der aktuellen Forschung bleibt, die Pathophysiologie der Sepsis besser zu verstehen und neue innovative Ansätze für eine mögliche Therapie zu definieren, um somit die Behandlungsqualität zu verbessern und die Gesamtkosten zu senken. 1.2 Rolle der T-Lymphozyten bei der Pathogenese der Sepsis 1.2.1 T-Helferzellen und zytotoxische T-Zellen Während die Funktionen der Zellen des angeborenen Immunsystems bei der Sepsis seit längerem untersucht werden, ist über eine mögliche Rolle der adaptiven Immunantwort in der Pathophysiologie weniger bekannt. T-Lymphozyten erkennen als Teil des adaptiven Immunsystems ihnen über Haupthistokompatibilitätskomplex(MHC)-Moleküle präsentierte Antigene. CD4+ THelferzellen differenzieren sich nach Aktivierung über den MHC-Klasse-II-Komplex 4 in TH1- oder TH2-Richtung und unterscheiden sich anhand ihres sezernierten Zytokinprofils. TH1-Zellen aktivieren durch Ausschüttung von Zytokinen wie IFN-γ eine zellvermittelte Immunantwort. Gleichzeitig wird eine T H2-Antwort durch die freigesetzten Zytokine inhibiert. T H2-Zellen stimulieren die humorale Immunantwort der B-Lymphozyten und hemmen durch Zytokine wie IL-4 und IL-10 die TH1Effektorzell-Entwicklung. Durch diese Kreuzhemmung wird eine einmal eingeschlagene Richtung für die Immunantwort beibehalten (Abbildung 2). Zytotoxische T-Zellen erkennen Peptid-MHC-I-Komplexe und lösen durch Granzyme, Perforine oder über Fas-FasL-Interaktion in der Zielzelle den programmierten Zelltod aus. APZ TH0 IFN-γ TH1 TH2 IL-4, IL-10 IL-4 IL-2 IFN-γ Zell- IL-5 Humorale vermittelte IL-6 Immunität IL-10 Immunität Abbildung 2: Differenzierung und Regulierung der T-Helferzellen. Antigen- Präsentierende-Zellen (APZ) stimulieren naive T-Helferzellen (TH0). Die weitere Ausdifferenzierung in T-Helferzellen Typ 1 (TH1) bzw. T-Helferzellen Typ 2 (TH2) wird durch die bei der Antigenstimulation bereitgestellten Zytokine mit beeinflusst. Die Polarisierung der Differenzierung zur Bildung von T H1-Zellen erfolgt hauptsächlich in Gegenwart von Interferon-γ (IFN-γ), während eine TH2-Polarisierung vornehmlich durch Interleukin(IL)-4 stimuliert wird. Die von beiden CD4-Populationen gebildeten Zytokinmuster sichern nicht nur die weitere Differenzierung zu den T H1- bzw. TH2Subpopulationen, sondern hemmen gleichzeitig Aktivierung und Proliferation des jeweils anderen T-Effektorzell-Typs. Abbildung modifiziert nach Miller [28]. 5 Es deutet vieles darauf hin, den T-Lymphozyten einen entscheidenden Beitrag in der Pathogenese der Sepsis zuzuschreiben. Als zentraler Mechanismus in der Entstehung der Immunparalyse während der Sepsis wird die erhöhte Apoptoserate von Lymphozyten angesehen. Im Thymus septischer Mäuse tritt dies bereits vier Stunden nach CLP auf und betrifft vor allem die unreife CD4+CD8+ T-Zellpopulation. Dabei führt die Abnahme CD4+CD8+ TLymphozyten zur Zunahme der CD4-CD8- Zellpopulation [2, 14, 23, 50]. T-Lymphozyten aus der Milz septischer Patienten weisen 12 Stunden nach Sepsisbeginn einen Anstieg der Apoptoserate auf, der direkt mit der Mortalität korreliert [23, 24, 50]. Die durchflusszytometrische Analyse CD4+ Splenozyten zeigte eine Abnahme um 45 %. Dagegen nahm der Anteil CD8+ zytotoxischer TZellen um 60 % zu, obwohl in Tierversuchen eine erhöhte Apoptoserate sowohl für CD4+ als auch für CD8+ T-Lymphozyten festgestellt wurde [23, 24]. Weiterhin konnte im Tiermodell gezeigt werden, dass durch den adoptiven Transfer apoptotischer Splenozyten die Mortalität der Sepsis IFN-γ-abhängig erhöht ist, während der Transfer nekrotischer Splenozyten zu einem Anstieg von IFN-γ führt und einen Überlebensvorteil bietet [18]. Dies bedeutet, dass Apoptose nicht nur durch den Funktionsausfall der toten Zellen die Immunkompetenz einschränkt, sondern dass die apoptotischen Zellen darüber hinaus immunsuppressiv wirken. Studien beweisen, dass dieser Verlust immunkompetenter Zellen eine zentrale Rolle in der Entstehung der Immundysfunktion während der Sepsis spielt [50]. Zum einen erhöhten sich durch die Gabe von Apoptosehemmstoffen die T H1Zytokinkonzentration und die Überlebenswahrscheinlichkeit CLP-behandelter Mäuse. Die Blockade der Lymphozytenapoptose bewirkte außerdem einen Abfall der TH2-Zytokine in der Spätphase der Sepsis [49, 50]. Die Tendenz in Richtung einer TH2-dominanten Immunantwort und damit verminderten Resistenz gegen eindringende Keime stellt ein möglicher Pathomechanismus der Sepsis dar [19, 28, 45]. Zum anderen führte die Überexpression des antiapoptotisch wirkenden Proteins Bcl-2 in transgenen Mäusen zu einem Schutz gegenüber Apoptose und somit zu einem Überlebensvorteil während der Sepsis [22]. 6 Diese Ergebnisse und die verminderte Überlebenswahrscheinlichkeit lymphozytendefizienter RAG1-/- - Mäuse im CLP-Modell, heben die Bedeutung der Lymphozyten in der Sepsis deutlich hervor [22]. In Tierversuchen zeigte die Depletion CD8+ T-Zellen eine Abnahme der IL-6Konzentration und ein verbessertes Überleben, was durch den Wegfall der suppressiven Wirkung zytotoxischer T-Zellen erklärbar ist [10, 38]. Für die Rolle der CD4+ T-Lymphozyten postulieren einige Studien eine frühzeitige Aktivierung während der Sepsis. Untersuchungen CD4-depletierter Mäuse im CLP-Modell zeigten aber unterschiedliche Ergebnisse. Während Enoh et al. keine Veränderungen in Hinblick auf Überleben und Zytokinproduktion CD4-depletierter Mäuse im Vergleich zu Wildtypmäusen feststellen konnten [10], zeigten Martignoni et al. die Bedeutung von CD4+ Lymphozyten in der Frühphase der Sepsis. Das Fehlen dieser Zellen führte zu einer erhöhten Sterblichkeit, und dies besonders innerhalb der ersten 30 Stunden nach Sepsisinduktion mittels CLP. Da durch die Gabe naiver CD4+ T-Zellen keine Verbesserung erzielt wurde, könnten für das Überleben nicht-naive oder regulatorische T-Zellen die entscheidende Rolle spielen [26]. 1.2.2 Thymozyten Die Vorläuferzellen der T-Lymphozyten entstehen im Knochenmark und wandern von dort über die Blutbahn in den Thymus ein, wo sie in verschiedenen Entwicklungsstadien reifen und differenzieren. Zu diesem Zeitpunkt weisen sie einen doppel-negativen (DN) CD-4, CD8- Phänotyp auf und entwickeln sich von CD44+CD25- (DN1) über CD44+CD25+ (DN2) zu CD44-CD25+ (DN3) und im Anschluss zu CD44-CD25- (DN4) [27]. DN4-Zellen proliferieren und entwickeln sich zu CD4+CD8+ doppel-positiven (DP) Thymozyten weiter, die 70 % der lymphatischen Zellen im Thymus ausmachen. Im letzen Schritt der Entwicklung, der positiven Selektion, reifen DP Thymozyten anhand ihrer Antigenspezifität entweder zu CD4+ T-Helferzellen oder zu CD8+ zytotoxischen T-Zellen (Abbildung 3). Autoreaktive Zellen werden im Rahmen der negativen Selektion durch Apoptose eliminiert [5, 25]. 7 Abbildung 3: Darstellung der Thymopoese, von doppel-negativ (DN) über doppel-positiv (DP) zu einfach-positiv (EP) mit entsprechender CD-(Cluster of Determination) Zelloberflächenmarkierung. Abbildung modifiziert nach Kuo und Leiden [25]. Im Rahmen der Sepsis kann eine Atrophie des Thymus und eine vermehrte Thymozytenapoptose beobachtet werden. Die Thymusinvolution zeigt sich histologisch durch Auszehrung der kortikalen Schicht, Verlust der kortikomedullären Abgrenzung, erhöhte Apoptoseraten und verminderte Abgabe naiver T-Zellen an die Peripherie. Von der Apoptose sind überwiegend CD4+CD8+ DP Thymozyten betroffen [2, 15, 46, 48]. Als für die Involution verantwortliche molekulare Mechanismen werden Zytokine, wie TNFα, IL-6 oder Lymphotoxin, Stickstoffmonoxid (NO), Glukokortikoide und die direkte Wirkung von Pathogenen diskutiert [15, 46, 50]. Der Zusammenhang zwischen dem Verlust der Immunkompetenz während der Sepsis und der daraus resultierenden Mortalität konnte in einigen Studien verdeutlicht werden. Systemische Aktivierung des Komplementfaktors C5a scheint eine erhöhte Apoptoseneigung in Thymozyten zu verursachen. Riedemann et al. zeigten ein Überlebensvorteil septischer Mäuse bei Blockade des programmierten Zelltodes durch Einsatz von Komplement C5a-Antikörpern [13]. Desweiteren konnte während der Sepsis eine erhöhte Thymozytenapoptose durch Abnahme des antiapoptotisch wirkenden Transkriptionsfaktors NF-κB festgestellt werden. Die verstärkte Aktivierung von NF-κB in transgenen Mäusen führte zu einer geringeren Apoptose und somit zu einem Überlebensvorteil während der Sepsis [14]. Diese Ergebnisse belegen die Bedeutung einer funktionierenden Thymopoese für die Entwicklung und Aufrechterhaltung des Immunsystems während der Sepsis. 8 1.2.3 Regulatorische T-Zellen Als regulatorische T-Zellen (Tregs) wurden CD4+CD25+ T-Lymphozyten erstmals 1995 von Sakaguchi et al. beschrieben [33]. Als eine spezielle Subpopulation von T-Lymphozyten spielen sie eine wichtige Rolle bei der Kontrolle von Reaktionen des angeborenen und adaptiven Immunsystems. Zum einen limitieren Tregs durch Hemmung von CD4+ und CD8+ T-Zellen die Immunantwort gegenüber Fremdantigenen. Zum anderen verhindern Tregs Autoimmunität, indem sie Toleranz gegenüber Selbstantigenen vermitteln. Des Weiteren können Tregs durch Produktion immunregulatorischer Zytokine (IL-10, TGF- β) und Induktion von Apoptose in Zielzellen, immunregulatorisch wirksam werden [44]. Der Transkriptionsfaktor Foxp3, ein weiterer Marker für regulatorische T-Zellen, ist von entscheidender Bedeutung für deren Entwicklung. Durch die Entdeckung, dass die Mutation von Foxp3, zu der immunproliferativen Autoimmunerkrankung IPEX (Immune dysregulation, Polyendocrinopathy, Enteropathy X-linked syndrome) führt, verdeutlicht, dass Tregs eine entscheidende Rolle spielen ein Gleichgewicht im Immunsystem aufrecht zu erhalten [17]. In Bezug auf die Sepsis beschrieben Monneret et al. 2003 erstmals eine Zunahme von CD4+CD25+ T-Zellen im Blut septischer Patienten, die bereits bei Diagnosestellung festzustellen war und nur bei nicht-überlebenden Patienten persistierte. Dies wurde in einer anschließenden Studie als relative Zunahme der Tregs aufgrund einer Abnahme der CD4+CD25- T-Zellpopulation dargestellt und in weiteren Untersuchungen bestätigt [44]. Im CLP-Sepsismodell der Maus konnte eine IL-10 vermittelte Zunahme Foxp3exprimierender und damit immunsuppressiv wirkender Tregs in der Milz und im Blut beobachtet werden. Diese Zellen sind in der Lage die CD4 +CD25- TZellaktivierung und -proliferation zu unterdrücken [34, 51]. In einer aktuellen Studie wurde ein Zusammenhang zwischen dem Anstieg zirkulierender Tregs und der Anergie sowie herabgesetzten Proliferationskapazität von Lymphozyten im septischen Schock deutlich [43]. Somit könnten Tregs an der Immunparalyse in der Spätphase der Sepsis beteiligt sein. Dabei ist die immunologische Wirkung der Tregs folgend auf eine Infektion, von deren relativen Anzahl gegenüber TEffektorzellen abhängig [44]. Versuche, mit monoklonalen Antikörpern CD4+CD25+ bzw. CD25+ T-Zellen auszuschalten und somit das Überleben septischer Mäuse zu verbessern, zeigten jedoch keinen Erfolg [34, 51]. 9 Dagegen zeigten Scumpia et al. ein Überlebensvorteil CLP-behandelter Mäuse bei präoperativer Gabe von anti-GITR, welches die Treg-Funktion blockiert. Dies führte zu verbesserter T-Zellproliferation und verbesserter Zytokinantwort [35]. Trotz Widersprüche zur aktuellen Theorie der Pathophysiologie der Sepsis weist dagegen ein Transfer in-vitro stimulierter Tregs sechs Stunden vor bzw. nach CLP eine erhöhte Überlebenswahrscheinlichkeit, eine verbesserte bakterielle Beseitigung, eine erhöhte Rekrutierung peritonealer Mastzellen sowie eine erhöhte TNF-α Produktion auf. Dafür ist die Existenz funktionierender T-Zellen notwendig, da der Transfer von Tregs auf Lymphozyten-defiziente Mäuse keine Wirkung auf das Überleben zeigt [16]. 1.3 Fragestellung und Zielsetzung Die Sepsis stellt mit ihrer hohen Mortalitätsrate und den enormen Kosten für das Gesundheitssystem ein häufiges und gravierendes Problem auf Intensivstationen dar [9, 12]. In der Pathophysiologie der Sepsis kommt der überschießenden Immunantwort in der Frühphase mit dem kontinuierlichen Übergang in eine Immunparalyse in der Spätphase eine wesentliche Bedeutung zu. Dabei scheinen Lymphozyten bei der Entwicklung der Immunparalyse eine wichtige Rolle zu spielen [7, 19, 39, 50]. Die erhöhte Mortalitätsrate lymphozytendefizienter RAG1-/- Mäuse und die Verbesserung der Überlebenswahrscheinlichkeit durch die Gabe von Apoptosehemmstoffen beweisen den Einfluss der erhöhten Lymphozytenapoptose auf die Immunparalyse in der Spätphase der Sepsis [23, 26, 49, 50]. Des Weiteren scheinen Tregs und zytotoxische T-Zellen mit ihrer Fähigkeit, eine Immunantwort zu unterdrücken, an der Entstehung der Immunparalyse beteiligt zu sein [51]. Diese Überlegungen Mediatorenkaskaden legen und der nahe, durch Intervention inflammatorisch auf kompetenten Ebene der T-Zellen die pathophysiologischen Abläufe der Sepsis zu modulieren. Ziel dieser Arbeit war es, die intrathymale T-Zellentwicklung und Konzentrationsveränderungen von CD4 + T-Helferzellen, CD8+ zytotoxischen TZellen und Tregs im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere zu untersuchen. 10 Dazu wurde zu Beginn der Studie eine Überlebenszeitanalyse weiblicher Wildtypmäuse nach Sham-Operation, Laparotomie ohne weitere Manipulation, und nach Sepsisinduktion durch das Modell der zökalen Ligatur und Punktion (CLP) unterschiedlicher Nadelstärken durchgeführt. Anschließend wurden zur durchflusszytometrischen Untersuchung der T- Zellpopulationen während der Sepsis weibliche Wildtypmäuse einer CLP unterschiedlicher Nadelstärken unterzogen. Sham-operierte und unbehandelte Tiere dienten als Kontrollgruppen. Das Abtöten der Tiere erfolgte zu festgelegten Zeitpunkten. Aus Thymus, Milz und Lymphknoten wurden Lymphozyten gewonnen und mit Antikörpern gefärbt. Es folgte die Messung der jeweiligen T-Zellpopulation im Durchflusszytometer. Von Interesse war der Einfluss der Sepsis auf CD4+, CD8+ sowie CD4+CD8+ TLymphozyten aus dem Thymus. Dabei interessierte, ob im zeitlichen Verlauf der Sepsis und in Abhängigkeit der bei der CLP verwendeten Nadelstärken Konzentrationsveränderungen der Zellpopulationen festzustellen sind. Desweiteren war die intrathymale Entwicklung von T-Lymphozyten im doppelnegativen Stadium von Bedeutung. Es wurde der Frage nachgegangen, ob und zu welchem Zeitpunkt zusätzlich zur Apoptose von CD4+CD8+ Thymozyten eine durch die Sepsis verursachte Inhibition der T-Lymphopoese in früheren Entwicklungsstadien festzustellen ist. Dies könnte zur Thymusinvolution in der Sepsis und somit zu einer erhöhten Mortalität beitragen. Es wurden noch die Anteile CD4+ und CD8+ T-Lymphozyten aus Milz und Lymphknoten überprüft. Hierbei interessierte, ob die Sepsis zu Veränderungen der peripheren Zellpopulationen führt und ob dies zeitlich in Beziehung zu einer möglichen Unterbrechung der intrathymalen T-Zellentwicklung oder einem Anstieg von Tregs steht. Außerdem wurden Tregs aus Milz und Lymphknoten untersucht, da diese durch Unterdrückung einer adaptiven Immunantwort an der Entstehung einer Immunparalyse im Spätstadium der Sepsis beteiligt sein könnten. Hier interessierte insbesondere, wie sich eine mögliche Veränderung der TregPopulation im zeitlichen Verlauf der Sepsis verhält und ob diese mit Veränderungen der restlichen untersuchten T-Zellpopulationen in Zusammenhang steht. Ferner wurde eine eventuelle Abhängigkeit der Treg-Konzentration von der Sepsisschwere überprüft. 11 2 Material und Methoden 2.1 Chemikalien und Medikamente Produkt Hersteller Bestellnummer Bovine Serum Albumine PAA Laboratories GmbH K41-001-100 DPBS 1x GIBCO Invitrogen 14190-169 D-PBS(10x) GIBCO Invitrogen 14200-067 Aqua B. Braun Braun 75/12604052/0503 Ethanol absolut puriss. Sigma-Aldrich 32205 Natriumchloridlösung 0,9 % B. Braun Melsungen AG Forene Abbott GmbH B506 Temgesic ESSEX PHARMA 4800604 Fraction V 2.2 Lösungen FACS-Puffer: 0,1 % BSA; 0,1 % Natriumazid; 2 mM EDTA pH 8,0; in PBS PFA-Fixierlösung 4 %: Paraformaldehyd 16 %; dest. H2O 2.3 Antikörper 2.3.1 FACS-Antikörper Produkt Hersteller Bestellnummer Rat Anti Mouse CD4 FITC Serotec MCA1107F Rat Anti Mouse CD8 Alpha: Serotec MCA609PEB Serotec MCA609P647 RPE Rat Anti Mouse CD8 Alpha: RPE-Alexa Fluor 647 PerCP-Cy5.5-Conjugated Rat BD Biosciences 551071 Anti Mouse CD25 Mouse Anti Mouse CD44 Serotec MCA1014FB FITC 12 2.3.2 Negativkontrollen Produkt Hersteller Bestellnummer Rat IgG2a Negative Control Serotec MCA1212F Serotec MCA1212P647 BD Biosciences 551072 Serotec SFL928F FITC Rat IgG2a Negative Control RPE-Alexa Fluor 647 PerCP-Cy5.5 Rat IgG1, λ Isotype Control Mouse IgG1 Negative Control FITC 2.4 Methoden 2.4.1 Versuchstiere Für die Versuche wurden insgesamt 141 weibliche Wildtypmäuse des Stammes C57BL/6J im Alter zwischen 98 und 207 Tagen, durchschnittliches Alter 140 Tage, verwendet. Die Versuchstiere wurden im Tierforschungszentrum „Oberberghof“ der Universität Ulm gehalten und von angestellten Versuchstierpflegern betreut. Der Raum, in dem die Tiere untergebracht waren, war ohne Tageslicht und wurde zur Erhaltung eines Tag-Nacht-Rhythmus jeden Tag von 6 bis 18 Uhr künstlich beleuchtet. Pro Käfig waren fünf Mäuse untergebracht, die Zugang zu Wasser und Nahrung ad libitum hatten. Für die Durchführung der Versuche lag die Genehmigung der Tierschutzkommission des Regierungspräsidiums Tübingen vor (Reg.-Nr. 806). 2.4.2 Überlebenszeitanalyse Zur Untersuchung der Überlebenszeiten septischer Mäuse nach zökaler Ligatur und Punktion (CLP, sh. 2.4.4) standen insgesamt 32 weibliche Wildtypmäuse zur Verfügung. Die Versuche wurden in einer Gruppengröße von je acht Tieren durchgeführt. Ziel war, den optimalen Tötungszeitpunkt der Versuchstiere für die nachfolgende durchflusszytometrische Messung von T-Lymphozyten während der Sepsis festzustellen. Des Weiteren wurden die Überlebenszeiten in Abhängigkeit der bei der CLP verwendeten Nadelstärke untersucht, um somit Rückschluss auf verschiedene Schweregrade der Sepsis zu erlangen. 13 Dazu wurden für die CLP Nadelstärken in aufsteigendem Außendurchmesser (AD) ausgewählt. Je acht Tiere wurden mit der Nadelstärke 26G (AD 0,45 mm), 22G (AD 0,7 mm) und 18G (AD 1,2 mm) operiert. Als Kontrollgruppe dienten acht Tiere, die einer Sham-Operation, Laparotomie ohne weitere Manipulation, unterzogen wurden (Abbildung 4). CLP Sham-Op Laparotomie 26G 22G 18 G n=8 n=8 n=8 n=8 Abbildung 4: Überlebenszeitanalyse mit je acht Versuchstieren pro Gruppe (n=8) unterteilt nach Operationsverfahren: Sham-Op (Sham-Operation) bzw. CLP (cecal ligation and puncture) eingeteilt nach der verwendeten Nadelstärke in G (Gauge). Der Beobachtungszeitraum endete 20 Tage nach Op. Postoperativ wurden die Tiere über einen Zeitraum von 20 Tagen beobachtet. Tiere, die vor Ablauf der 20 Tage zunehmend folgende schwerere Krankheitssymptome aufwiesen, deutliche Aktivitätsabnahme nach Stimulation, trockene bzw. putride Augen, Atemdepression (unregelmäßige Atmung, Bradypnoe) bzw. Tachypnoe, verzögerten bzw. erloschenen Stellreflex und/oder Körpergewichtsabnahme von mehr als 15 %, wurden frühzeitig getötet und als Todesfall gewertet. Das Abtöten aller noch nach Ablauf des Beobachtungszeitraumes lebender Versuchstiere erfolgte durch zervikale Dislokation. 2.4.3 T-Lymphozyten während der Sepsis Für die durchflusszytometrischen Untersuchungen der verschiedenen T- Zellpopulationen während der Sepsis, standen insgesamt 109 Versuchstiere zur Verfügung. Messungen wurden an 98 Tieren durchgeführt, da 11 der 109 Tiere 14 während der Versuche noch vor dem geplanten Zeitpunkt des Abtötens verstarben. 2.4.3.1 CD4CD8-Immunphänotypisierung von T-Lymphozyten im Thymus Zur Untersuchung der CD4+ T-Helferzellen, CD8+ zytotoxischer T-Lymphozyten und CD4+CD8+ T-Lymphozyten im Thymus während der Sepsis, wurden an allen 98 weiblichen Wildtypmäusen Messungen durchgeführt. Die Einteilung der Versuchstiere erfolgte nach der Schwere der Sepsis. Innerhalb dieser Gruppen wurden Subgruppen gebildet, die die unterschiedlichen Zeitpunkte des Abtötens widerspiegeln. Gruppe 1 und Gruppe 2 dienten als Kontrollgruppen. An Tieren der Gruppe 1 wurde keinerlei Manipulation vorgenommen. An Tieren der Gruppe 2 wurde eine Sham-Operation (Sham-Op), Laparotomie ohne weitere Manipulation, durchgeführt. Die gemeinsame Haltung von Sham- und CLP-operierten Mäusen im Käfig erfolgte unter gleichen Bedingungen. Unbehandelte Tiere der Gruppe 1 dienten dazu eventuelle Veränderungen Sham-operierter Tiere der Gruppe 2 zu erfassen. Die Induktion einer Sepsis wurde mit dem Verfahren der CLP, Ligatur des Zökums und anschließende Punktion, durchgeführt. Die Schwere der Sepsis korrelierte mit der für die Punktion Versuchsgruppen 3, 4 des und Zökums 5 verwendeten wurden Nadelstärke. Nadelstärken in Für die aufsteigendem Außendurchmesser (AD) ausgewählt: 26G (AD 0,45 mm) für die Gruppe 3, 22G (AD 0,7 mm) für die Gruppe 4 und 18G (AD 1,2 mm) für die Gruppe 5. Desweiteren fand eine Aufteilung der Gruppen anhand der Sepsisdauer, Zeitpunkt der Operation bis zum Abtöten, statt. Der Zeitpunkt des Abtötens der unbehandelten Tiere der Gruppe 1 (n=4) wurde willkürlich gewählt. Die in der Kontrollgruppe 2 Sham-operierten Tiere wurden entsprechend den CLP-Gruppen 24 (n=6), 36 (n=4), 48 (n=6) bzw. 96 Stunden (n=6) nach Sham-Op abgetötet. Tiere der Gruppen 3 (26 G) sowie 4 (22 G) wurden 24 (n=11), 48 (n=11) bzw. 96 Stunden (n=11) nach CLP abgetötet. Um Veränderungen in möglichst schweren Sepsisstadien zu erfassen, wurden Tiere der Gruppe 5 (n=6) einer CLP mit der Nadelstärke 18G unterzogen. Da die Mehrzahl der Tiere innerhalb der ersten 48 Stunden verstarb, wurden sie 36 Stunden nach Sepsisinduktion getötet (Abbildung 5). 15 Unbehandelt CLP Sham-Op Ohne Laparotomie 26G 22G 18 G Operation 24h 36h 48h 96h 24h 48h 96h 24h 48h 96h 36h n=4 n=6 n=4 n=6 n=6 n=11 n=11 n=11 n=11 n=11 n=11 n=6 Gruppe 1 Gruppe 2 Gruppe 3 Gruppe 4 Gruppe 5 Abbildung 5: CD4CD8-Immunphänotypisierung von T-Lymphozyten im Thymus. Versuchsgruppen 1 bis 5 unterteilt nach Operationsverfahren: Gruppe 1 ohne Operation, Gruppe 2 Sham-Op (Sham-Operation), Gruppen 3, 4 und 5 CLP (cecal ligation and puncture) eingeteilt nach Nadelstärke in G (Gauge). Desweiteren sind in den Gruppen 2 bis 5 die Zeitpunkte des Abtötens in h (Stunden) nach Operation aufgeführt. 2.4.3.2 Immunphänotypisierung von doppel-negativen Thymozyten, peripheren T-Lymphozyten und regulatorischen T-Zellen Die Anteile doppel-negativer Thymozyten sowie die Anteile CD4+ T-Helferzellen, CD8+ zytotoxischer T-Lymphozyten und CD4+CD25+ Tregs aus Milz und Lymphknoten während der Sepsis wurden an 70 der insgesamt 98 Tiere untersucht. Da es sich hier um eine explorative Untersuchung handelte, wurden kleinere Gruppengrößen gewählt. Die Versuchstiere wurden wie in 2.4.3.1 beschrieben nach der Schwere der Sepsis und der Sepsisdauer eingeteilt. Gruppe 1 und Gruppe 2 dienten als Kontrollgruppen. An Tieren der Gruppe 1 wurde keinerlei Manipulation vorgenommen. Tiere der Gruppe 2 erhielten eine Sham-Op. Für die Versuchsgruppen 3, 4 und 5 wurden für die Sepsisinduktion mittels CLP Nadelstärken in aufsteigendem AD ausgewählt: 26G für die Gruppe 3, 22G für die Gruppe 4 und 18G für die Gruppe 5. Ebenso fand eine Aufteilung der Gruppen anhand der Sepsisdauer statt. Der Zeitpunkt des Abtötens der unbehandelten Tiere der Gruppe 1 (n=4) wurde willkürlich gewählt. Die in der Kontrollgruppe 2 Sham-operierten Tiere wurden entsprechend den CLP-Gruppen 24 (n=4), 36 (n=2), 48 (n=4) bzw. 96 Stunden 16 (n=4) nach Sham-Op abgetötet. Tiere der Gruppen 3 (26 G) sowie 4 (22 G) wurden 24 (n=8), 48 (n=8) bzw. 96 Stunden (n=8) nach CLP abgetötet. Tiere der Gruppe 5 (n=4) wurden einer CLP mit der Nadelstärke 18G unterzogen und 36 Stunden nach Sepsisinduktion getötet (Abbildung 6). Unbehandelt CLP Sham-Op Ohne Laparotomie 26G 22G 18 G Operation 24h 36h 48h 96h 24h 48h 96h 24h 48h 96h 36h n=4 n=4 n=2 n=4 n=4 n=8 n=8 n=8 n=8 n=8 n=8 n=4 Gruppe 1 Gruppe 2 Gruppe 3 Gruppe 4 Gruppe 5 Abbildung 6: Immunphänotypisierung von Thymozyten, peripheren T-Lymphozyten und regulatorischen T-Zellen. Versuchsgruppen 1 bis 5 unterteilt nach Operationsverfahren: Gruppe 1 ohne Operation, Gruppe 2 Sham-Op (Sham-Operation), Gruppen 3, 4 und 5 CLP (cecal ligation and puncture) eingeteilt nach Nadelstärke in G (Gauge). Desweiteren sind in den Gruppen 2 bis 5 die Zeitpunkte des Abtötens in h (Stunden) nach Operation aufgeführt. 2.4.4 Sepsisinduktion durch zökale Ligatur und Punktion (CLP) Zur Induktion einer polymikrobiellen Sepsis wurde das Modell der CLP angewandt. Die Inhalationsnarkose wurde über eine Gesichtsmaske mit 2 Vol.-% Isofluran eingeleitet. Anschließend wurden die Mäuse auf vorgeheizter Operationsfläche in Rückenlage fixiert. Die Anästhesie wurde mit einem Gemisch aus 50 % Sauerstoff, Stickstoff und 2 Vol.-% Isofluran fortgeführt. Nach Rasur und Desinfektion der Abdominalregion wurde unter sterilen Bedingungen der Bauchraum durch einen ein bis zwei cm langen medianen Schnitt mit einem Skalpell in kraniokaudaler Richtung eröffnet. Nach Identifikation des Zökums, wurde dieses distal der Ileozökalklappe mit einem Vicrylfaden ligiert ohne die Kontinuität des Magen-Darm-Traktes zu unterbrechen. Zur Sepsisinduktion wurde das Zökum mit Nadeln unterschiedlichen Durchmessers durchstochen (Abbildung 7). 17 Abbildung 7: Durchstechen des Zökums im Rahmen einer CLP (cecal ligation and puncture) mit einer 22G (Gauge)-Nadel bei einer auf dem Rücken fixierten Maus in Isoflurannarkose. Die Ligatur des Zökums erfolgte bereits. Anschließend wurde das Zökum mit einer Pinzette leicht gequetscht, um eine geringe Menge Stuhl aus den Punktionslöchern zu drücken und eine Perforation der Darmwand sicher zu stellen. Nach Rückverlagerung des Zökums in den Bauchraum, wurde die Bauchwand in zwei Schichten mit nicht resorbierbaren Fäden wieder verschlossen. Kontrolltiere erhielten eine Sham-Op. Nach Narkoseeinleitung wurde der Bauchraum eröffnet und das Zökum freigelegt. Nach Identifikation des Zökums wurde dieses ohne Ligatur und ohne Punktion in den Bauchraum zurückverlagert. Der Wundverschluss und die postoperative Betreuung erfolgten wie bei den CLPTieren. Zur Kompensation des perioperativen Flüssigkeitsverlustes erhielten alle Tiere eine s.c. Injektion von 0,05 ml/g Körpergewicht isotonischer Kochsalzlösung 0,9 %. Die postoperative Analgesie erfolgte mit 25 µg/kg Buprenorphin s.c.. Nach durchgeführter CLP- bzw. Sham-Operation wurden die Mäuse bis zur Erholung aus der Narkose kontinuierlich überwacht. 18 2.4.5 Aufrechterhaltung der Analgesie und Flüssigkeitshomöostase Da die Trinkwasseraufnahme der Tiere post-CLP unzureichend war, erfolgte 12stündlich eine s.c.-Injektion von 0,05 ml/g Körpergewicht isotonischer Kochsalzlösung 0,9 %. Die Analgesie erfolgte alle 12 Stunden mit 25 µg/kg Buprenorphin s.c.. Zweimal täglich wurde der Zustand der Mäuse überprüft. 2.4.6 Blut- und Organentnahme Die Tötung der Tiere erfolgte durch zervikale Dislokation. Nach Eintauchen in Alkohol und Fixierung in Rückenlage wurde mit sterilem Präparierbesteck eine Relaparotomie durchgeführt und der Brustkorb eröffnet. Milz, Lymphknoten und Thymus wurden entfernt und bis zur Aufbereitung in PBS auf Eis gelagert. Die folgenden Schritte wurden immer mit auf Eis gekühlten Lösungen und Chemikalien durchgeführt. Während der Bearbeitung wurden die Proben ständig auf Eis gelagert. 2.4.7 Gewinnung von Lymphozyten Mit Hilfe eines Skalpells wurden die Organe auf einem Objektträger zerkleinert. Die dadurch gewonnene Single Cell Suspension wurde mit 10 ml PBS durch ein 40 µm Zellsieb in ein Falcon-Röhrchen überführt und für fünf Minuten bei 250 gE zentrifugiert. Anschließend wurden die Zellen mit 1xPBS gewaschen. Um die bei der Durchflusszytometrie störenden Erythrozyten zu lysieren, wurde das Pellet, der aus der Milz gewonnenen Proben, mit 9 ml dest. H2O resuspendiert. Sofort danach wurde 1 ml 10xPBS zugegeben und resuspendiert. Das Pellet aus Lymphkoten und Thymus wurde im gleichen Schritt mit 10 ml 1xPBS resuspendiert und zusammen mit den Milzproben erneut zentrifugiert. Im nächsten Schritt wurden die Proben in 1xPBS resuspendiert, gewaschen und zentrifugiert. Der Überstand wurde verworfen und das Pellet je nach Größe in 13 ml FACS-Puffer gelöst. Von den Proben wurden jeweils 100 µl pro FACSRöhrchen pipettiert. 19 2.4.8 Anfärben mit Antikörpern Nach Aufarbeitung der Organe wurden die gewonnenen Lymphozyten mit Antikörpern markiert. Die verwendeten monoklonalen Antikörper binden an für die Zellen spezifische CD-Oberflächenmoleküle. Sie sind an Farbstoffe konjungiert, die in der Durchflusszytometrie mit Laserlicht angeregt werden und Licht einer bestimmten Wellenlänge emittieren. Dadurch war eine Charakterisierung und Unterscheidung der untersuchten Immunzellen möglich. Als Negativkontrollen wurden weiteren Proben Isotypkontrollen entsprechend den Antikörperfärbungen zugegeben. Damit gelang es Autofluoreszenzen zu erkennen und durch Einstellungen am Durchflusszytometer zu minimieren. Die verwendeten Antikörper und Isotypkontrollen wurden entsprechend den Herstellerhinweisen zu den Proben in die FACS-Röhrchen hinzugegeben. Nach Zugabe der Antikörper und der Negativkontrollen wurden die Proben für 30 Min. im Dunkeln auf Eis inkubiert. Im Anschluss wurden die Proben zwei Mal mit FACS-Puffer gewaschen und dem Pellet am Ende 300 µl FACS-Puffer zugegeben. 2.4.9 Zellfixierung Die Fixierung der Zellen erfolgte durch Zugabe Paraformaldehydlösung 4 % für 30 Min. auf Eis. Danach wurden die Zellen zwei Mal mit 1xPBS gewaschen und in 300 µl FACS-Puffer resuspendiert. Durch die Fixierung mit Paraformaldehyd waren die Proben bei Aufbewahrung im Kühlschrank bei 4 °C für sieben Tage haltbar. 2.4.10 Durchführung der durchflusszytometrischen Analyse Die Durchflusszytometrie ist ein optisches Messsystem, das Streulicht- und Fluoreszenzsignale antikörpermarkierter Zellen analysiert. Dies ermöglicht das gleichzeitige Erfassen mehrerer physikalischer und biochemischer Parameter. Die Zellsuspension wird durch eine Kapillare aus den FACS-Röhrchen eingesogen und in Richtung Messkammer des Geräts geleitet. Dort befinden sich die Zellen durch hydrodynamische Fokussierung in einer linear angeordneten Reihe und werden nacheinander einzeln von einem Argonlaser mit einer Wellenlänge von 488 nm beleuchtet. Wenn Laserlicht auf eine Zelle trifft, wird ein Teil des Lichts 20 gestreut. Licht das von den Zellen in Richtung des ursprünglichen Strahls streut, Vorwärtsstreulicht (FSC = Forward Scatter Channel), spiegelt je nach Intensität die Größe des Partikels wider. Strahlen, die rechtwinkelig zur ursprünglichen Lichtquelle streuen, Seitwärtsstreulicht (SSC = Side Scatter Channel), entstehen durch intrazelluläre Bestandteile und Membraneigenschaften. Damit lassen sich die wichtigsten der in der Zellsuspension befindlichen Leukozytenpopulationen in einem Dot-Plot Diagramm (FSC gegen SSC) anhand ihrer charakteristischen Eigenschaften in Größe und Granularität darstellen. Aufgrund ihres kleinen Durchmessers und geringen Gehaltes zytoplasmatischer Granula besitzen Lymphozyten eine geringe Abschwächung und eine geringe Lichtstreuung. Bei der Datenauswertung wurde, mittels einer ungefärbten Zellprobe, für jedes Organ im Forward Scatter – Sideward Scatter Streudiagramm durch „Gating“, Eingrenzen und Auswählen der zu untersuchenden Zellen mit Hilfe eines Auswertefensters, die Analyse auf die Lymphozytenpopulation begrenzt (Abbildungen 8 und 9). SSC SSC R1 FSC FSC Abbildung 8: Repräsentative Abbildung 9: Repräsentative Zellverteilung von Lymphozyten aus der Zellverteilung von Lymphozyten aus der Milz im Forward Scatter (FSC) – Sideward Milz im Forward Scatter (FSC) - Sideward Scatter (SSC) Punktgraphen. Scatter (SSC) Punktgraphen. Die mit R1 abgegrenzte Zellfraktion wurde als Lymphozytenpopulation angesehen und die Messung auf diese eingegrenzt. Neben dem Vorwärts- und dem Seitwärtsstreulicht kann als dritter Parameter die Fluoreszenz der Zellen gemessen werden. Die bei den Antikörpern eingesetzten Fluoreszenzfarbstoffe emittieren bei Anregung durch den Argonlaser eine für sie bestimmte Wellenlänge Licht. Durch optische Filter wird von verschiedenen Detektoren nur Licht einer bestimmten Wellenlänge erfasst: 21 Kanal 1 (FL 1): Emissionsspektrum 515 – 545 nm (FITC) Kanal 2 (FL 2): Emissionsspektrum 564 – 606 nm (R-PE) Kanal 3 (FL 3): Emissionsspektrum > 670 nm (PerCP-Cy 5) Es folgte die Einstellung des Durchflusszytometers mittels ungefärbter Zellproben, einzeln- und doppelgefärbter Proben sowie Negativkontrollen. Die Verwendung von zu den Antikörpern identischen Isotypen, die an keinen Farbstoff konjungiert waren, minimierten das Auftreten falsch-positiver Ergebnisse durch die Autofluoreszenz der Antikörper (Abbildungen 10 und 11). CD 8 R 4: R1: 14 % 0,0 % CD 8 R3: R4: R1: 0,1 % 0,0 % R3: 85,7 % 98,8 % R2: R2: 0,4 % 1,1 % CD 4 CD 4 Abbildung 10: Repräsentative Abbildung 11: Repräsentative Punktgraph-Darstellung der CD4 Punktgraph-Darstellung der CD4 (Cluster of Differentiation) Intensität (Cluster of Differentiation) Intensität gegenüber der CD8 Intensität in gegenüber der CD8 Intensität in Lymphozyten der Milz in CD8 Einzelfärbung. Lymphozyten der Milz in Die mit R1 abgegrenzte Zellfraktion wurde Isotypenfärbung CD4 und CD8. als CD4 und CD8 positiv angesehen, R2 Die mit R1 abgegrenzte Zellfraktion wurde als allein CD 4 positiv, R3 als CD4 und als CD4 und CD8 positiv angesehen, R2 CD8 negativ, R4 als allein CD8 positiv. als allein CD4 positiv, R3 als CD4 und CD8 negativ, R4 als allein CD8 positiv. Die Emission der Fluoreszenzfarbstoffe sowie das SSC und das FSC werden von den Photodetektoren erfasst und in elektrische Signale umgewandelt. Bei der Mehrfarbenfluoreszenz können die Signale mehrerer Fluoreszenzfarbstoffe gleichzeitig gemessen werden. Für die Messungen wurde das FACSCalibur der Firma Becton & Dickinson verwendet. Pro Probe wurden mindestens 5 000 Zellen gemessen. 22 2.4.11 Auswertung und graphische Darstellung Zur Beschreibung der Überlebenszeitdaten weiblicher Wildtypmäuse nach CLP bzw. Sham-Op wurde die Darstellung der Kaplan-Meier-Kurven verwendet. Diese geben die bedingte Überlebenswahrscheinlichkeit im zeitlichen postoperativen Verlauf wider. Die Auswertung der im FACS gemessenen Daten erfolgte mit dem Programm Cell Quest Pro (Becton & Dicinson). Bei der Auswertung CD4+, CD8+ und CD4+CD8+ T-Lymphozyten aus dem Thymus während der Sepsis waren zum einen Konzentrationsveränderungen der Zellpopulationen in Abhängigkeit von der Sepsisschwere von Interesse. Dazu wurden zum jeweiligen Zeitpunkt Sham-Gruppe, 26G- und 22G-CLP-Gruppe, bei Normalverteilung und Varianzhomogenität in einer Varianzanalyse mittels One Way ANOVA, bei nicht normalverteilten Werten oder Varianzinhomogenität mittels Kruskal-Wallis One Way ANOVA on Ranks, miteinander verglichen. Die Überprüfung der Signifikanz nach One Way ANOVA erfolgte mit der StudentNewman-Keuls Methode, nach Kruskal-Wallis One Way ANOVA on Ranks mit der Dunn´s Methode. Der Nachweis signifikanter Veränderungen im zeitlichen Verlauf der Sepsis, wurde nach der gleichen statistischen Vorgehensweise durchgeführt. Hier wurden die in der Durchflusszytometrie gewonnenen Ergebnisse einer Gruppe zu den Zeitpunkten 24, 48 und 96 Stunden miteinander verglichen. Die Auswertung der Daten 36 Stunden nach Sham-Op bzw. 18G-CLP erfolgte getrennt. Bei normal verteilten Werten und Varianzhomogenität wurde zur Überprüfung der statistischen Signifikanz der t-Test, ansonsten der MannWhitney-U-Test herangezogen. Die statistische Auswertung der Daten erfolgte mit dem Programm SigmaStat 8.0. Das Signifikanzniveau wurde bei allen Tests mit p<0,05 festgelegt. Die Boxplot-Darstellungen wurden mit SigmaPlot 8.0 erstellt und geben Median, 25. und 75. Quartil sowie Whisker vom Minimum zum Maximum der Daten an. Bei der Untersuchung von doppel-negativen Thymozyten, peripheren TLymphozyten und regulatorischen T-Zellen handelte es sich um eine explorative Studie, die als Basis für Folgeexperimente dienen soll. Aufgrund der geringen Fallzahl der Versuchsgruppen (n=8, n=4 bzw. n=2) konnte hier kein statistischer Test durchgeführt werden. In einer deskriptiven Statistik sind die gemessenen Werte der Versuchsgruppen 1 (unbehandelte Tiere) und 2 bis 4 (Sham-Operation, 23 26G- und 22G-CLP) zu den Zeitpunkten 24, 48 und 96 Stunden, in Punktdiagrammen dargestellt. So ist ein Überblick über die Veränderungen der prozentualen Anteile der Immunzellen in den unterschiedlichen Gruppen im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere möglich. Durch einen vertikalen Balken getrennt wurden die Werte 36 Stunden nach 18GCLP sowie Scheinoperation aufgeführt. Um Ausreißer zu minimieren, wurde als Lagekennwert der Median angegeben. Desweiteren können in den Punktdiagrammen Minimal- und Maximalwert abgelesen werden. Für die graphische Darstellung wurde das Programm Microsoft Excel verwendet. 24 3 Ergebnisse Die vorliegende Arbeit beschäftigte sich mit der Untersuchung von T-Lymphozyten im zeitlichen Verlauf und in Abhängigkeit der Schwere der Sepsis. Zur Induktion einer polymikrobiellen Sepsis wurde das Modell der zökalen Ligatur und Punktion (CLP) angewandt. Dabei wurde weiblichen Wildtypmäusen unter sterilen Bedingungen das Zökum ligiert, ohne die Kontinuität des Magen-Darm-Traktes zu unterbrechen. Anschließend wurde das Zökum mit unterschiedlichen Nadelstärken durchstochen. Als Kontrollgruppen dienten Sham-Mäuse, die eine Scheinoperation, Eröffnung des Bauchraums ohne weitere Manipulation, erhielten. Unbehandelte Mäuse stellten eine weitere Kontrollgruppe dar. Anschließend wurde den Tieren zu festgelegten Zeitpunkten Thymus, Milz und Lymphknoten entnommen. Nach Aufarbeitung der Organe wurden die gewonnenen Lymphozyten mit Antikörpern markiert und durchflusszytometrisch gemessen. Die dabei gewonnenen Ergebnisse wurden unter Verwendung eines FACSCalibur und der CellQuest Software von Becton & Dicinson erzeugt. 3.1 Überlebenszeitanalyse Zur Untersuchung der Überlebenszeiten nach Sepsisinduktion durch zökale Ligatur und Punktion der Nadelstärken 26-, 22- bzw. 18G sowie nach ShamOperation, wurden zunächst Vorversuche in einer Gruppengröße von je acht Tieren durchgeführt. Der Beobachtungszeitraum begann mit der Op und endete nach 20 Tagen. Abbildung 12 zeigt nach Kaplan-Meier-Analyse die Überlebenswahrscheinlichkeiten innerhalb der unterschiedlichen Gruppen. In der Kontrollgruppe überlebten nach Sham-Op, Laparotomie ohne weitere Manipulation, alle acht Tiere den Beobachtungszeitraum. Bei den CLP-behandelten Tieren korrelierte die Überlebenswahrscheinlichkeit mit dem Außendurchmesser der verwendeten Nadelstärke. Alle Todesfälle traten zwei bis sechs Tage nach Sepsisinduktion auf. Nach CLP mit der Nadelstärke 26G verstarb am sechsten postoperativen Tag ein Versuchstier. Die Wahrscheinlichkeit den sechsten Tag zu überleben betrug 87,5 % und veränderte sich bis zum Ende der Beobachtung nicht. 25 Nach Sepsisinduktion mittels 22G-CLP verstarben drei Tiere innerhalb der beobachteten 20 Tage nach Op. Die Todeszeitpunkte waren am dritten, vierten und sechsten Tag nach CLP zu verzeichnen. Die Überlebenswahrscheinlichkeit lag an Tag 3 bei 87,5 %, an Tag 4 bei 75 % und an Tag 6 bei 62,5 %. Die Gesamtüberlebensrate für den Beobachtungszeitraum von 20 Tagen betrug 62,5 %. Die geringste Überlebensrate war nach Durchführung der CLP mit der Nadelstärke 18G festzustellen. Bereits 48 Stunden postoperativ lebten nur noch 50 % der Tiere. Am dritten Tag nach CLP verstarben weitere drei der insgesamt acht Tiere. Die Überlebenswahrscheinlichkeit betrug ab dem dritten postoperativen Tag Überlebenswahrscheinlichkeit in % 12,5 %. 100 80 60 40 Sham CLP 26G CLP 22G CLP 18G 20 0 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 Überlebenszeit (Tage) Abbildung 12: Kaplan-Meier-Kurven für die Überlebenswahrscheinlichkeit weiblicher Wildtypmäuse in Abhängigkeit von der Schwere der Sepsis und der Überlebenszeit bei einem Beobachtungszeitraum von 20 Tagen nach Sham-Operation bzw. Sepsisinduktion mittels 18-, 22- bzw. 26G (Gauge)-CLP(cecal ligation and puncture). 26 3.2 Auswertung der Narkoseprotokolle der Immunphänotypisierung 3.2.1 Vorzeitig verstorbene Tiere Für die durchflusszytometrische Untersuchung von T-Lymphozyten im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere wurden insgesamt 109 weibliche Wildtypmäuse verwendet. 11 Tiere verstarben vor dem Zeitpunkt des geplanten Abtötens und standen somit für die durchflusszytometrischen Untersuchungen nicht zur Verfügung. In der Gruppe 4, geplantes Abtöten 48 Stunden nach 22G-CLP, starben vier von 37 Tieren, eines am ersten, die restlichen drei am zweiten Tag nach CLP. Die Wahrscheinlichkeit 48 Stunden nach Sepsisinduktion mit der Nadelstärke 22G zu überleben betrug 89,2 %. Nach 18G-CLP verstarben sieben von 13 Tieren, davon zwei am ersten und fünf am zweiten Tag. Innerhalb der Gruppe 5 lag die Wahrscheinlichkeit 36 Stunden nach CLP mit der Nadelstärke 18G zu überleben bei 46,2 %. In Tabelle 1 sind durchgeführte Op, Alter, Narkose- und Op-Dauer sowie Todestag nach Op der vorzeitig verstorbenen Tiere aufgeführt. In den restlichen Gruppen ergab sich eine Überlebensrate von 100 % bis zum Zeitpunkt des Abtötens. Tabelle 1: Auflistung der Mäuse, die nach Sepsisinduktion mittels CLP (cecal ligation and puncture) der Nadelstärken 26, 22 und 18G (Gauge) vor Zeitpunkt des geplanten Abtötens, 24, 36, 48 bzw. 96 h (Stunden) nach Op (Operation) verstarben. Dargestellt sind die Versuchstiere mit entsprechend durchgeführter Op, Alter in Tagen, Narkose- und Op-Dauer in Minuten sowie Sterbezeitpunkt in Tagen nach Op. Narkosedauer Op-Dauer in Minuten in Minuten Todestag nach Op 130 14 10 2. „ 98 12 8 2. „ 98 28 16 2. „ 98 17 10 1. CLP 18G36h 131 12 9 1. „ 148 13 9 2. „ 148 13 10 2. „ 125 15 11 2. „ 125 14 9 2. „ 153 12 8 2. „ 174 15 10 1. Op-Art Alter in Tagen CLP 22G48h 27 3.2.2 CD4CD8-Immunphänotypisierung von T-Lymphozyten im Thymus Für die CD4CD8-Immunphänotypisierung im Thymus wurden alle 98 Versuchstiere verwendet, die bis zum Zeitpunkt des geplanten Abtötens überlebten. Tabelle 2 zeigt die einzelnen Subgruppen mit Angabe des durchschnittlichen Alters der Tiere sowie der durchschnittlichen Narkose- und Operations-Dauer. Es ergaben sich zwischen den einzelnen Subgruppen keine signifikanten Unterschiede in Alter, Narkose- bzw. Op-Dauer. Tabelle 2: Auflistung des durchschnittlichen Alters der Mäuse in Tagen sowie der durchschnittlichen Narkose- und Op(Operation)-Dauer in Minuten mit Angabe des Minimal- und Maximalwertes (min/max) eingeteilt nach durchgeführter Op-Art sowie Tötungszeitpunkt in h (Stunden) nach Op: unbehandelt sowie Scheinoperation (Sham) bzw. Sepsisinduktion mittels CLP (cecal ligation and puncture) der Nadelstärken 26, 22 und 18G (Gauge). Durchschnittliches Durchschnittliche Durchschnittliche Alter in Tagen Narkosedauer in Op-Dauer in (min/max) Minuten (min/max) Minuten (min/max) Unbehandelt (n=4) 182 (107/207) - - Sham 24h (n=6) 135 (107/189) 15,8 (13/19) 9 (8/11) Sham 36h (n=4) 148(125/189) 14,3 (13/17) 9,5 (9/10) Sham 48h (n=6) 127 (98/174) 12,8 (14/10) 7,5 (6/9) Sham 96h (n=6) 135 (113/167) 12,5 (11/13) 7,7 (7/8) CLP 26G24h (n=11) 141 (114/207) 14,5 (11/20) 9,3 (8/12) CLP 26G48h (n=11) 135 (98/174) 13,7 (12/16) 8,9 (7/10) CLP 26G96h (n=11) 137 (113/167) 16,2 (12/24) 9,7 (7/14) CLP 22G24h (n=11) 130 (107/189) 14,3 (13/17) 9,2 (8/11) CLP 22G48h (n=11) 160 (130/207) 14,4 (13/16) 9,5 (7/11) CLP 22G96h (n=11) 135 (113/160) 13,3 (11/16) 8,8 (7/11) CLP 18G36h (n=6) 143 (153/174) 13,9 (12/17) 9,7 (8/12) Op-Art 28 3.2.3 Immunphänotypisierung von doppel-negativen Thymozyten, peripheren T-Lymphozyten und regulatorischen T-Zellen Zur durchflusszytometrischen Messung doppel-negativer Thymozyten, peripherer T-Helferzellen und zytotoxischer T-Zellen sowie von Tregs wurden 70 der 98 Versuchstiere verwendet. In Tabelle 3 sind, nach den Subgruppen aufgeteilt, durchschnittliches Alter sowie durchschnittliche Narkose- und Op-Dauer der 70 Versuchstiere aufgeführt. Es ergaben sich zwischen den einzelnen Subgruppen keine signifikanten Unterschiede in Alter, Narkose- bzw. Op-Dauer. Tabelle 3: Auflistung des durchschnittlichen Alters der Mäuse in Tagen sowie der durchschnittlichen Narkose- und Op(Operation)-Dauer in Minuten mit Angabe des Minimal- und Maximalwertes(min/max) eingeteilt nach durchgeführter Op-Art sowie Tötungszeitpunkt in h (Stunden) nach Op: unbehandelt sowie Scheinoperation (Sham) bzw. Sepsisinduktion mittels CLP (cecal ligation and puncture) der Nadelstärken 26, 22 und 18G (Gauge). Durchschnittliches Durchschnittliche Durchschnittliche Alter in Tagen Narkosedauer in Op-Dauer in (min/max) Minuten (min/max) Minuten (min/max) Unbehandelt (n=4) 182 (107/207) - - Sham 24h (n=4) 119 (107/128) 16,5 (15/19) 9 (8/11) Sham 36h (n=2) 157 (125/189) 15 (13/17) 9,5 (9/10) Sham 48h (n=4) 125 (98/174) 12,5 (10/14) 6,8 (6/7) Sham 96h (n=4) 138 (113/167) 12,8 (12/13) 7,3 (7/8) CLP 26G24h (n=8) 133 (114/207) 14,9 (11/20) 9,4 (8/12) CLP 26G48h (n=8) 136 (98/174) 14,1 (12/16) 8,8 (7/10) CLP 26G96h (n=8) 140 (113/167) 15,5 (12/24) 10,1 (7/14) CLP 22G24h (n=8) 118 (107/128) 14,5 (13/17) 9 (8/11) CLP 22G48h (n=8) 169 (131/207) 14,5 (13/16) 9,1 (7/11) CLP 22G96h (n=8) 137 (113/160) 14,5 (11/16) 8,8 (8/9) CLP 18G36h (n=4) 164 (153/174) 14,8 (13/17) 10,5 (10/12) Op-Art 29 3.3 T-Lymphozyten im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere 3.3.1 T-Helferzellen, zytotoxische T-Zellen und doppel-positive T- Lymphozyten aus dem Thymus Im Folgenden wurden die Veränderungen CD4+ T-Helferzellen, CD8+ zytotoxischer T-Zellen und CD4-CD8- doppel-negativer T-Zellen aus dem Thymus untersucht. Von weiterer Bedeutung waren CD4+CD8+ doppel-positive Lymphozyten, die eine Entwicklungsstufe unreifer Thymozyten vor der Maturation zu CD4+ oder CD8+ einfach-positiven T-Zellen darstellen. Dadurch sollten Veränderungen in der T-Lymphopoese während der Sepsis erfasst werden. Verglichen mit den entsprechenden Sham-Tieren als Kontrollgruppe konnte im Thymus septischer Mäuse eine starke Abnahme der CD4+CD8+ Zellpopulation im zeitlichen Verlauf der Sepsis festgestellt werden. Die niedrigsten Werte zeigten sich 96 Stunden nach CLP. Im Gegensatz dazu nahm bei septischen Tieren der Anteil CD4+, CD8+ und CD4CD8- T-Lymphozyten zu. Die höchsten Werte wurden 96 Stunden nach CLP gemessen. Abbildung 13 zeigt repräsentative Punktgraphdarstellungen der CD4- gegenüber der CD8-Intensität in Lymphozyten aus dem Thymus. Um Veränderungen der Zellkonzentrationen im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere zu zeigen sind Sham-Tiere als Kontrollgruppe und 22G- sowie 26G-CLP-Tiere 24 Stunden und 96 Stunden nach Operation dargestellt. 30 24h 3,7 % 96h 74 % 5,2 % [ [ G G e b 5,4 % 12,5 % e 68,5 % Sham 5,1 % 21,2 % b [ [ Sie e [Geben e [Geben Sie G G 6,1 % 83,5 % 7,8 % 25,2 % aus n ein Zitat n ein Zitat aus e e [ dem [[ CD8 26G dem b b G Dokument GG S 2,8 % e e 7,6 % S Dokument 46 % e ee 21 % oder die i oder die i n n b Zusammenf bb [ [ e e Zusammenf e assung ee G G assung 8,1 % 14,5 % 4,4 % S S75,6 % n eines n e e n eines e e i i [ 22G b b [i interessante interessante i e e G [S SS e e15,7 % n Punktes 4,3 % G n Punktes49,4 % n n e 28,1 % G i ein. Sie i n n i e ein. Sie e [e b e [ e ee das bZ können können das i i e b G G CD4Z S S an ei Textfeld in Textfeld an n en e e e einer i Punktgraph-Darstellung e Abbildung 13: Repräsentative der CD4 (Cluster of i e einer nt t b der b Differentiation)-Intensitätn in Lymphozyten aus dem Thymus. i gegenüber ii e e CD8-Intensität beliebigen beliebigen a a eZ Ergebnis eZ Dargestellt ist jeweils ein von bzw. 26G- und 22G(Gauge)n typisches nS n ShamStelle im S Stelle im tS t i ni puncture)-Mäusen ni behandelten CLP(cecal ligation and nach 24 und 96 h (Stunden) bei e e i Dokument Dokument t t voneinander unabhängigen mit jeweils vergleichbarem Resultat iZ Versuchsdurchführungen Ze i i Z positioniere ea Sham-Gruppen a S a n=6). a positioniere (CLP-Gruppen n=11 bzw. e S i ii n n u n. u n. ti it t Verwenden te t es Verwenden si e e e a Sie die a Z Z a Sie die i a a it tn i i t nd Registerkart d Registerkart n eu eu t t e e e e is is a 'Textfeldtool aZ a a a 'Textfeldtool Z m m i Z n n u s', wenn u t t u s', wenn Sie Sie i d d is sts tD das Format das Format e a t 31 D Ze Z ao des am a des o t a i im d Textfelds d u u d Textfelds tk k te t t ee s s Beim Betrachten der einzelnen Zeitpunkte der Sepsis, fiel im Vergleich zu ShamTieren eine Abnahme CD4+CD8+ T-Lymphozyten im Thymus septischer Mäuse auf (Abbildung 14). Signifikanter Unterschied und damit eine Abhängigkeit von der Sepsisschwere wurde zum Zeitpunkt 48 Stunden zwischen der Sham-Gruppe und Tieren nach 22G-CLP festgestellt (p=0,007). Eine signifikante Abnahme von CD4+CD8+ T-Lymphozyten war ebenfalls zum Zeitpunkt 96 Stunden zwischen Sham-operierten Tieren und 22G- sowie 26G-operierten CLP-Tieren nachzuweisen (p=0,001). Ebenso nahm im zeitlichen Verlauf der Sepsis die Konzentration CD4+CD8+ Thymozyten ab. Dies wurde beim Vergleich der Zeitpunkte 24 und 48 Stunden mit dem Zeitpunkt 96 Stunden nach 26G- und 22G-CLP statistisch signifikant (p<0,001). Thymus + Thymus CD4 CD8+ CD4+ CD8+ 100 + 60 + + CD4%CD8 in % 80 40 20 • * • + * Unbehandelt Sham CLP 26G CLP 22G + 0 Unbehandelt 24h 48h 96h Abbildung 14: Boxplot-Darstellung des prozentualen Anteils CD4⁺CD8⁺ (Cluster of Differentiation) T-Lymphozyten im Thymus. Dargestellt sind unbehandelte und Shambzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=6, CLP Gruppen n=11). Aufgeführt sind Median, 25. und 75. Quartil sowie Whisker vom Minimum zum Maximum der Daten. Legende: + = p<0,05 vs. Sham, * = p<0,05 vs. 24h, • = p<0,05 vs. 48h 32 Bei der Überprüfung doppelpositiver Thymozyten konnte zwischen Versuchstieren 36 Stunden nach Sham-Op und 18G-CLP kein statistisch signifikanter Unterschied der Zellkonzentration festgestellt werden (Abbildung 15). Thymus CD4+ CD8++ Thymus CD4 CD8+ 100 CD4+% CD8+ in % 80 60 40 20 Sham CLP 18G 0 36h Abbildung 15: Boxplot-Darstellung des prozentualen Anteils CD4⁺CD8⁺ (Cluster of Differentiation) T-Lymphozyten im Thymus. Dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=4, CLP Gruppe n=6). Aufgeführt sind Median, 25. und 75. Quartil sowie Whisker vom Minimum zum Maximum der Daten. Die durchflusszytometrischen Messungen CD4+ T-Lymphozyten ergaben bei CLPTieren eine von der Sepsisschwere abhängige Zunahme der Zellpopulation im zeitlichen Verlauf der Sepsis (Abbildung 16). In der statistischen Auswertung war zum Zeitpunkt 48 Stunden zwischen der Sham-Gruppe und 22G-CLP-Gruppe eine signifikante Zunahme der untersuchten Zellpopulation festzustellen (p=0,007). 96 Stunden nach Sepsisinduktion war dies zum einen beim Vergleich von ShamTieren mit 26G-operierten Tieren nachzuweisen (p=0,038). Sepsisinduktion mittels 22G-CLP führte nach 96 Stunden zur signifikanten Zunahme CD4+ T-Lymphozyten sowohl gegenüber Sham- (p<0,001) als auch gegenüber 26G-operierten Tieren (p=0,004). 33 Nach zökaler Ligatur und Punktion kam es auch im zeitlichen Verlauf der Sepsis zur Zunahme CD4+ Thymozyten. Dies wurde innerhalb der 26G-CLP-Gruppe zwischen den Zeitpunkten 24 und 48 Stunden (p=0,002) sowie 24 und 96 Stunden (p<0,001) signifikant. Nach Sepsisinduktion mit der Nadelstärke 22G nahm der Anteil CD4+ T-Lymphozyten im zeitlichen Verlauf der Sepsis beim Vergleich der Zeitpunkte 24 und 96 Stunden signifikant zu (p<0,001). Der Vergleich zwischen den Zeitpunkten 24 und 48 Stunden sowie 48 und 96 Stunden war mit einem pWert von 0,003 ebenfalls signifikant. Thymus + Thymus CD4 CD8⁻ CD4+ CD8- CD4+CD8⁻ in % % 60 ◊ • * Unbehandelt Sham CLP 26G CLP 22G * * + + + * 40 20 0 Unbehandelt 24h 48h 96h Abbildung 16: Boxplot-Darstellung des prozentualen Anteils CD4⁺CD8- (Cluster of Differentiation) T-Lymphozyten im Thymus. Dargestellt sind unbehandelte und Shambzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=6, CLP Gruppen n=11). Aufgeführt sind Median, 25. und 75. Quartil sowie Whisker vom Minimum zum Maximum der Daten. Legende: + = p<0,05 vs. Sham, * = p<0,05 vs. 24h, • = p<0,05 vs. 48h, ◊ = p<0,05 vs. CLP26G96h 34 Im Vergleich zur Sham-Gruppe wiesen Versuchstiere 36 Stunden nach 18G-CLP mit einem p-Wert von 0,01 signifikant höhere Werte CD4+ Thymozyten auf (Abbildung 17). Thymus CD4+ CD8- Thymus CD4+CD8⁻ 50 30 % CD4+CD8⁻ in % 40 + Sham CLP 18G 20 10 0 36h Abbildung 17: Boxplot-Darstellung des prozentualen Anteils CD4⁺CD8- (Cluster of Differentiation) T-Lymphozyten im Thymus. Dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=4, CLP Gruppe n=6). Aufgeführt sind Median, 25. und 75. Quartil sowie Whisker vom Minimum zum Maximum der Daten. Legende: + = p<0,05 vs. Sham 35 Abbildung 18 zeigt die Veränderungen CD8+ zytotoxischer T-Zellen aus dem Thymus während der Sepsis. Die Induktion einer Sepsis mittels 22G-CLP führte im Vergleich mit der entsprechenden Sham-Gruppe zum Zeitpunkt 48 Stunden zu signifikant höheren Konzentrationen CD8 + Thymozyten (p=0,006). Im zeitlichen Verlauf der Sepsis war nach 26G-CLP zwischen den Zeitpunkten 24 und 96 Stunden eine signifikante Zunahme CD8+ T-Lymphozyten festzustellen (p<0,001). Nach 22G-CLP war dies sowohl beim Vergleich der Zeitpunkte 24 Stunden mit 48 Stunden (p=0,024) sowie 96 Stunden (p<0,001), als auch beim Vergleich der Zeitpunkte 48 und 96 Stunden (p=0,009) zu verzeichnen. Thymus CD4- CD8+ Thymus CD4⁻CD8+ 40 + CD4⁻CD8 in % % 30 Unbehandelt Sham CLP 26G CLP 22G * • * * + 20 10 0 Unbehandelt 24h 48h 96h Abbildung 18: Boxplot-Darstellung des prozentualen Anteils CD4 -CD8⁺ (Cluster of Differentiation) T-Lymphozyten im Thymus. Dargestellt sind unbehandelte und Shambzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=6, CLP Gruppen n=11). Aufgeführt sind Median, 25. und 75. Quartil sowie Whisker vom Minimum zum Maximum der Daten. Legende: + = p<0,05 vs. Sham, * = p<0,05 vs. 24h, • = p<0,05 vs. 48h 36 Sepsisinduktion durch 18G-CLP führte zum Zeitpunkt 36 Stunden zu einer signifikanten Erhöhung CD8+ Thymozyten gegenüber Sham-operierten Tieren: p=0,003 (Abbildung 19). Thymus CD4- CD8+ Thymus CD4⁻CD8+ 16 14 + Sham CLP 18G 10 % + CD4⁻CD8 in % 12 8 6 4 2 0 36h Abbildung 19: Boxplot-Darstellung des prozentualen Anteils CD4 -CD8⁺ (Cluster of Differentiation) T-Lymphozyten im Thymus. Dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=4, CLP Gruppe n=6). Aufgeführt sind Median, 25. und 75. Quartil sowie Whisker vom Minimum zum Maximum der Daten. Legende: + = p<0,05 vs. Sham 37 Bei Überprüfung CD4-CD8- Thymozyten während der Sepsis (Abbildung 20) fiel im Vergleich zur entsprechenden Kontrollgruppe 96 Stunden nach 26G-CLP eine signifikante Zunahme der Zellpopulation auf (p=0,009). Im zeitlichen Verlauf wurde die Zunahme doppel-negativer Thymozyten innerhalb der CLP-Gruppen deutlich. Ein signifikanter Unterschied konnte mit einem p-Wert von <0,001 nach 26G-CLP zwischen den Zeitpunkten 24 und 96 Stunden sowie zwischen den Zeitpunkten 48 und 96 Stunden nachgewiesen werden. Diese Zunahme war auch nach Sepsisinduktion mit der Nadelstärke 22G festzustellen. Im Vergleich zwischen den Zeitpunkten 24 und 96 Stunden sowie 48 und 96 Stunden nach CLP war eine signifikante Zunahme CD4-CD8- Thymozyten nachzuweisen (p<0,001). Thymus Thymus CD4⁻CD8⁻ CD4- CD8- • * + CD4⁻CD8⁻ % in % 60 Unbehandelt Sham CLP 26G CLP 22G • * 40 20 0 Unbehandelt 24h 48h 96h Abbildung 20: Boxplot-Darstellung des prozentualen Anteils CD4-CD8- (Cluster of Differentiation) T-Lymphozyten im Thymus. Dargestellt sind unbehandelte und Shambzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=6, CLP Gruppen n=11). Aufgeführt sind Median, 25. und 75. Quartil sowie Whisker vom Minimum zum Maximum der Daten. Legende: + = p<0,05 vs. Sham, * = p<0,05 vs. 24h, • = p<0,05 vs. 48h 38 Zum Zeitpunkt 36 Stunden konnte zwischen Sham-Op und 18G-CLP kein statistisch signifikanter Unterschied der Konzentration CD4-CD8- T-Lymphozyten festgestellt werden. Abbildung 21 zeigt die Boxplots beider Gruppen. Thymus CD4- CD8- Thymus CD4⁻CD8⁻ 70 60 Sham CLP 18G CD4⁻CD8⁻ in % % 50 40 30 20 10 0 36h Abbildung 21: Boxplot-Darstellung des prozentualen Anteils CD4-CD8- (Cluster of Differentiation) T-Lymphozyten im Thymus. Dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=4, CLP Gruppe n=6). Aufgeführt sind Median, 25. und 75. Quartil sowie Whisker vom Minimum zum Maximum der Daten. 3.3.2 Thymozyten Zur vollständigen Untersuchung der T-Lymphopoese während der Sepsis wurden aus dem Thymus gewonnene Zellen mit Antikörper für CD44 und CD25 gefärbt. In den Experimenten konnte, verglichen mit den Kontrollgruppen, bei septischen Tieren eine Zunahme CD44+CD25- (DN1), CD44+CD25+ (DN2) und CD44-CD25+ (DN3) Thymozyten im zeitlichen Verlauf ermittelt werden. Die höchsten Werte, jedoch mit breiter Streuung der Einzelwerte, fanden sich in allen Färbungen zum Zeitpunkt 96 Stunden. Eine Korrelation des Anstieges DN1-, DN2- und DN3Thymozyten mit der Sepsisschwere war zu den Zeitpunkten 24 und 96 Stunden festzustellen. 39 Dagegen nahm die Konzentration CD44-CD25- (DN4) Thymozyten im zeitlichen Verlauf der Sepsis ab. Dies war zu den Zeitpunkten 24 und 96 Stunden nach CLP von der Schwere der Sepsis abhängig. Abbildung 22 zeigt repräsentative Punktgraphdarstellungen der CD44- gegenüber der CD25-Intensität in Lymphozyten aus dem Thymus. Um Veränderungen der Zellkonzentrationen im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere zu zeigen, sind Sham-Tiere als Kontrollgruppe und 22G- sowie 26G-CLP-Tiere 24 Stunden und 96 Stunden nach Operation dargestellt. 24h 2,8 % 96h 1,4 % 2,9 % 1,9 % Sham 74 % 21,8 % 77,9 % 3,1 % 1,1 % 3,4 % 17,3 % 3,7 % CD25 26G 78, 8% 17,1% 70,2 % 22,7 % 2,1 % 5,9 % % 1,5 % 1,1 % %% 22G 69,1 % 28,3 % 47,3 % 44,7 % CD44 Abbildung 22: Repräsentative Punktgraph-Darstellung 4 der CD44(Cluster of Differentiation)-Intensität gegenüber der CD25-Intensität in Lymphozyten aus dem Thymus. Dargestellt ist jeweils ein typisches Ergebnis von Sham- bzw. 26G- und 22G(Gauge) CLP(cecal ligation and puncture)-Mäusen nach 24 und 96 h (Stunden) bei voneinander unabhängigen Versuchsdurchführungen mit jeweils vergleichbarem Resultat (CLP-Gruppen n=8 bzw. Sham-Gruppe n=4). 40 In der CD44-Einzelfärbung (Abbildung 23) war innerhalb der Kontrollgruppen eine Schwankung der Mediane zu erkennen, in der Sham-Gruppe zusätzlich Ausreißer zu den Zeitpunkten 48 und 96 Stunden. Für die unbehandelten Tiere errechnete sich für CD44+CD25- ein Median von 8,4 %. Sham-Tiere wiesen 24, 48, und 96 Stunden nach Operation für CD44+CD25- einen Median von 14,3 %, 11,4 % und 18,2 % auf. Septische Thymi wiesen im zeitlichen Verlauf der Sepsis eine tendenzielle Zunahme der CD44+CD25- (DN1) T-Zellpopulation auf. Dies war zu den Zeitpunkten 24 und 96 Stunden von der Sepsisschwere abhängig. 24 Stunden nach 26G-CLP befand sich der Median im Bereich der entsprechenden Kontrollgruppen. Dagegen führte die Sepsisinduktion mittels 22G-CLP bereits nach 24 Stunden mit einem Median von 27,2 % zu einem Anstieg CD44+CD25Thymozyten. 48 und 96 Stunden nach Sepsisinduktion konnten im Vergleich zu den entsprechenden Kontrollgruppen in der 26G- und 22G-CLP-Gruppe deutlich höhere Mediane ermittelt werden. Der Median CD44+CD25- Lymphozyten lag zum Zeitpunkt 48 Stunden nach 26G-CLP bei 24,4 %, nach 22G-CLP bei 19,9 %. Die höchsten Werte, aber mit breiter Streuung der Einzelwerte, zeigten sich nach 96 Stunden. Sepsisinduktion mit der Nadelstärke 26G führte zu einem Median von 29,3 %, mit der Nadelstärke 22G zu einem Median von 46,2 %. Im Vergleich zu unbehandelten Tieren führte die CLP mittels 18G-Nadel nach 36 Stunden zu einer leichten Erhöhung des Median (M 14,7 %). Nach Scheinoperation errechnete man einen der CLP-Gruppe entsprechenden Median, wobei die Einzelwerte breit streuten. 41 Thymus CD44⁺CD25⁻ CD44⁺CD25¯ in % 60 50 40 30 20 10 0 Unbehandelt Unbehandelt Abbildung 23: Sham 24h 48h CLP 26G CLP 22G 96h CLP 18G 36h Median Darstellung des prozentualen Anteils CD44⁺CD25¯ (Cluster of Differentiation) T-Lymphozyten im Thymus. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22GCLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. Doppel-positive CD44+CD25+ (DN2) Thymozyten (Abbildung 24) zeigten innerhalb der Kontrollgruppen vergleichbare Ergebnisse. Zum Zeitpunkt 48 und 96 Stunden nach Scheinoperation waren Ausreißer zu erkennen. Durch Induktion einer Sepsis mittels CLP nahm der Anteil CD44+CD25+ Thymozyten im zeitlichen Verlauf der Sepsis zu. Zu den Zeitpunkten 24 und 96 Stunden war dies von der Sepsisschwere abhängig. 24 Stunden nach 26G-CLP lag der Median noch im Bereich der entsprechenden Kontrollgruppen. CLP mit der Nadelstärke 22G führte bereits nach 24 Stunden zu einem leicht erhöhten Median von 2,1 %. Der Median zum Zeitpunkt 48 Stunden lag nach 26G-CLP bei 2,9 %, nach 22G-CLP bei 2,7 %. Die zunehmende Sepsisdauer führte 96 Stunden nach CLP zu einem deutlichen Anstieg CD44+CD25+ Thymozyten. Für 26G-operierte Tiere errechnete man einen Median von 6 %, für 22G-operierte Tiere von 8,3 %. Es war eine breite Streuung der Einzelwerte 96 Stunden nach CLP zu erkennen. 42 Verglichen mit unbehandelten Tieren (M 1 %) zeigte die Sepsisinduktion mittels 18G-Nadelstärke mit einem Median von 2,4 % eine leichte Zunahme der CD44+CD25+ Lymphozyten-Konzentration. Thymus CD44⁺CD25⁺ CD44⁺CD25⁺ in % 14 12 10 8 6 4 2 0 Unbehandelt Unbehandelt Abbildung 24: 24h Sham 48h CLP 26G CLP 22G 96h CLP 18G 36h Median Darstellung des prozentualen Anteils CD44⁺CD25⁺ (Cluster of Differentiation) T-Lymphozyten im Thymus. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22GCLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. Messungen CD44-CD25+ (DN3) T-Lymphozyten in den Kontrollgruppen, unbehandelte Tiere mit einem Median von 2,3 % und Sham-Tiere 24, 48 bzw. 96 Stunden nach Operation mit einem Median von 1,5 %, 0,8 % bzw. 1,8 % machten nur geringe Schwankungen der Ergebnisse deutlich. Zu den Zeitpunkten 48 und 96 Stunden waren Ausreißer in der Sham-Gruppe zu erkennen (Abbildung 25). Bei septischen Tieren war, im Vergleich zu den Kontrollgruppen, in der CD25Färbung eine von der Sepsisdauer abhängige Zunahme CD44 -CD25+ Thymozyten zu erkennen, die zum Zeitpunkt 96 Stunden von der bei der CLP verwendeten Nadelstärke abhing. Zum Zeitpunkt 48 Stunden waren im Vergleich zur ShamGruppe nach 26G-CLP mit einem Median von 2,9 % und 48 Stunden nach 22GCLP mit einem Median von 2,7 % erhöhte CD25+-Anteile festzustellen. 96 43 Stunden nach CLP mittels 26G-Nadel erhöhte sich der Median deutlich auf 6 %, nach CLP mittels 22G-Nadel auf 8,3 %. Die gemessenen Einzelwerte wiesen in diesen Gruppen eine breite Streuung auf. 36 Stunden nach 18G-CLP zeigten sich keine Veränderungen des Anteils CD44CD25+ Thymozyten gegenüber den Kontrollgruppen. Thymus CD44⁻CD25⁺ CD44⁻CD25⁺ in % 14 12 10 8 6 4 2 0 Unbehandelt Unbehandelt Abbildung 25: 24h Sham 48h CLP 26G CLP 22G 96h CLP 18G 36h Median Darstellung des prozentualen Anteils CD44¯CD25⁺ (Cluster of Differentiation) T-Lymphozyten im Thymus. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22GCLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. Innerhalb der Kontrollgruppen zeigten sich bei der Messung CD44 -CD25- (DN4) Thymozyten Abweichungen der Mediane. In der unbehandelten Gruppe lag der Median bei 88,3 %, in den Sham-Gruppen 24, 48 bzw. 96 Stunden nach Operation bei 83,6 %, 84,2 % bzw. 77,5 %. 48 und 96 Stunden nach Scheinoperation wurden Ausreißer deutlich (Abbildung 26). Im Gegensatz zu der Zunahme CD44 +CD25- (DN1), CD44+CD25+ (DN2) und CD44-CD25+ (DN3) Thymozyten führte die Induktion einer Sepsis zu einer Verringerung doppelnegativer Zellen. Zu den Zeitpunkten 24 und 96 Stunden war dies von der Sepsisschwere abhängig. Gegenüber scheinoperierten und 26G44 behandelten Tieren konnten 24 Stunden nach Sepsisinduktion mittels 22G-CLP mit einem Median von 69,4 % geringere Prozentsätze der CD44+CD25+Lymphozyten ermittelt werden. 48 Stunden postoperativ betrug der Median nach 26G-CLP 65,5 %, nach 22G-CLP 74,9 %. Die niedrigsten Werte und eine breite Streuung der Einzelwerte, wurden 96 Stunden nach CLP gemessen. Zu diesem Zeitpunkt betrug der Median nach 26G-CLP 56,8 %, nach 22G-CLP 43,7 %. Sepsisinduktion mittels 18G-CLP führte nach 36 Stunden mit 79,2 % zu einer Abnahme des Median gegenüber unbehandelter Tiere (M 88,3 %). Verglichen mit Tieren 36 Stunden nach Scheinoperation zeigten sich bei einem Median von 80,5 %, aber einer großen Spannbreite der Einzelwerte, keine Veränderungen der CD44-CD25--T-Zellpopulation. Thymus CD44⁻CD25⁻ CD44¯CD25¯ in % 100 80 60 40 20 0 Unbehandelt Unbehandelt Abbildung Sham 24h 48h CLP 26G CLP 22G 96h CLP 18G 36h Median 26: Darstellung des prozentualen Anteils CD44¯CD25¯ (Cluster of Differentiation) T-Lymphozyten im Thymus. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22GCLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. 45 3.3.3 T-Helferzellen und zytotoxische T-Zellen aus der Peripherie In Milz und Lymphknoten wurde die durchflusszytometrische Analyse CD4 +, CD8+ und CD4-CD8- T-Lymphozyten durchgeführt, um Konzentrationsveränderungen von T-Lymphozyten in sekundär lymphatischen Organen während der Sepsis zu erfassen. 3.3.3.1 Milz Die Messungen von Splenozyten septischer Mäuse zeigten im Vergleich zu den Kontrolltieren eine tendenzielle Abnahme CD4+ und CD8+ T-Lymphozyten. Dies wurde zu den Zeitpunkten 48 und 96 Stunden nach CLP deutlich. Analog zu diesen Ergebnissen konnte eine Zunahme CD4-CD8- T-Lymphozyten während der Sepsis festgestellt werden. Eine Abhängigkeit von der Sepsisschwere war bei allen Zellpopulationen zum Zeitpunkt 48 Stunden festzustellen. Die bei Messungen CD4+ Splenozyten gewonnenen Ergebnisse sind in Abbildung 27 dargestellt. Innerhalb der Kontrollgruppen fiel eine große Streubreite der Einzelwerte auf, bei Sham-Tieren eine tendenzielle Zunahme CD4 + T- Lymphozyten im zeitlichen Verlauf. Für unbehandelte Tiere errechnete man einen Median von 19,5 %, für Sham-Tiere 24, 48 bzw. 96 Stunden nach Operation einen Median von 17,5 %, 18,4 % bzw. 21,5 %. Verglichen mit den Kontrollgruppen nahm in der Milz septischer Tiere der Anteil CD4+ Splenozyten tendenziell ab und war zu den Zeitpunkten 24 und 48 Stunden von der Schwere der Sepsis abhängig. Sepsisinduktion mit einer Nadelstärke von 22G führte zum Zeitpunkt 24 Stunden zu einem Median von 15,8 %. 48 Stunden nach CLP wurde die Abnahme der untersuchten Zellkonzentration sowohl nach 26G-CLP mit einem Median von 15,4 % als auch nach 22G-CLP mit einem Median von 12,9 % deutlich. Die Abhängigkeit der CD4+ T-Lymphozyten-Abnahme von der Nadelstärke war 96 Stunden nach Sepsisinduktion nicht mehr zu erkennen. Für 26G-operierte Tiere ergaben die Messungen einen Median von 14,4 %, für 22G-operierte Tiere einen Median von 15,2 %. Die Induktion einer Sepsis mit der Nadelstärke 18G führte zu einem mit den Kontrollgruppen vergleichbaren Median. 46 Milz CD4⁺CD8⁻ 30 CD4⁺CD8¯ in % 25 20 15 10 5 0 Unbhandelt Unbehandelt 24h Sham 48h CLP 26G CLP 22G 96h CLP 18G 36h Median Abbildung 27: Darstellung des prozentualen Anteils CD4⁺CD8¯ (Cluster of Differentiation) T-Lymphozyten in der Milz. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLPbehandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. In den Kontrollgruppen fiel der Anteil CD8+ T-Lymphozyten (Abbildung 28) annäherungsweise gleich aus, mit einem Median von 12,7 % für unbehandelte Tiere und 12,4 % bzw. 12,7 % 24 bzw. 48 Stunden nach Scheinoperation. Einzelne Ausreißer waren in diesen Gruppen zu erkennen. Lediglich 96 Stunden nach Scheinoperation errechnete man einen leicht höheren Median von 14,8 %. Durch Sepsisinduktion mittels CLP stellte sich zum Zeitpunkt 48 und 96 Stunden eine tendenzielle Abnahme CD8+ Splenozyten dar. Dies war mit einem Median von 10,5 % in der 26G-Gruppe und 9,7 % in der 22G-Gruppe 48 Stunden postoperativ am deutlichsten. Die Werte 96 Stunden nach CLP betrugen 11 % nach 26G-CLP und 11,3 % nach 22G-CLP. Auch CLP mittels 18G-Nadel führte zu einer Abnahme CD8+ Splenozyten. Der errechnete Median lag zum Zeitpunkt 36 Stunden bei 10,4 % im Vergleich zur Sham-Gruppe mit einem Median von 13,3 % und der unbehandelten Gruppe mit einem Median von 12,7 %. 47 Milz CD4⁻CD8⁺ CD4¯CD8⁺ in % 20 15 10 5 0 Unbehandelt Unbehandelt Sham 24h 48h CLP 26G CLP 22G 96h CLP 18G 36h Median Abbildung 28: Darstellung des prozentualen Anteils CD4¯CD8⁺ (Cluster of Differentiation) T-Lymphozyten in der Milz. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLPbehandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. Die Messungen CD4-CD8- Zellen in der Milz (Abbildung 29) ergaben innerhalb der Kontrollgruppen bis zum Zeitpunkt 48 Stunden vergleichbare Ergebnisse. Für unbehandelte Tiere errechnete man einen Median von 68 %, für scheinoperierte Tiere 24 bzw. 48 Stunden postoperativ einen Median von 68,1 % bzw. 69,8 %. 96 Stunden nach Sham-Op berechnete man einen niedrigeren Median von 63,9 %. Bei septischen Tieren zeigte sich zu den Zeitpunkten 48 und 96 Stunden eine tendenzielle Zunahme doppelnegativer Splenozyten. Die höchsten Werte und eine Abhängigkeit von der Sepsisschwere zeigten sich zum Zeitpunkt 48 Stunden mit einem Median von 74 % nach 26G-CLP und einem Median von 77,3 % nach 22G-CLP. 96 Stunden nach Induktion einer Sepsis mit einer Nadelstärke von 26G betrug der Median 74 %, mit einer Nadelstärke von 22G 73,1 % Im Vergleich zu unbehandelten Tieren (M 68 %) und scheinoperierten Tieren zum Zeitpunkt 36 Stunden (M 67 %), führte eine CLP mittels 18G-Nadel mit einem Median von 71,4 % zu einer tendenziellen Zunahme der CD4-CD8- Konzentration. 48 Milz CD4⁻CD8⁻ CD4¯CD8¯ in % 100 80 60 40 20 0 Unbehandelt Unbehandelt Abbildung 29: 24h Sham Darstellung des 48h CLP 26G CLP 22G prozentualen 96h 36h CLP 18G Median Anteils CD4¯CD8¯ (Cluster of Differentiation) T-Lymphozyten in der Milz. Links dargestellt sind unbehandelte und Shambzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLPbehandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. 3.3.3.2 Lymphknoten Verglichen mit den Kontrollgruppen zeigten die Messungen aus septischen Lymphknoten eine tendenzielle Abnahme CD4+ T-Helferzellen ab 36 Stunden nach Sepsisinduktion sowie eine tendenzielle Abnahme CD8+ zytotoxischer TLymphozyten zum Zeitpunkt 96 Stunden. Bei doppelnegativen Zellen konnte eine durch die Sepsis verursachte Zunahme zu den Zeitpunkten 48 und 96 Stunden festgestellt werden. Die Überprüfung CD4+ T-Lymphozyten (Abbildung 30) ergab innerhalb der Kontrollgruppen vergleichbare Werte. In allen Gruppen zeigten sich große Varianzen. Für unbehandelte Tiere errechnete man einen Median von 35,6 %, für Sham-Tiere 24, 48 bzw. 96 Stunden nach Operation einen Median von 30,6 %, 34,8 % bzw. 33,5 %. Die Induktion einer Sepsis führte im Lymphknoten zu einer tendenziellen Abnahme CD4+ T-Lymphozyten. 22G- und 26G-CLP bewirkten zum Zeitpunkt 49 24 Stunden keine Veränderungen der CD4+ T-Zellkonzentration gegenüber der Sham-Gruppe, jedoch gegenüber der unbehandelten Gruppe. Für 22G-operierte Tiere ergaben die Messungen einen Median von 31,1 %, für 26G-operierte Tiere einen Median von 31,2 %. Ab dem Zeitpunkt 48 Stunden war sowohl im Vergleich mit unbehandelten als auch Sham-operierten Tieren eine tendenzielle Abnahme CD4+ T-Lymphozyten festzustellen. Diese war von der Sepsisschwere abhängig: Sepsisinduktion mittels 26G-CLP führte zu einem Median von 28,6 %, mittels 22GCLP zu einem Median von 25,2 %. 96 Stunden nach CLP mit der Nadelstärke 26G betrug der Median 28 %, mit der Nadelstärke 22G 30,1 %. Zu beiden Zeitpunkten lag ein Großteil der gemessenen Einzelwerte septischer Tiere im Bereich der Werte der Kontrollgruppen. Auch nach Induktion einer Sepsis mit der Nadelstärke 18G zeigte sich bei einem Median von 30 % eine tendenzielle Abnahme CD4+ T-Lymphozyten gegenüber den Kontrollgruppen. 36 Stunden nach Scheinoperation errechnete man einen Median von 33 %. Lymphknoten CD4⁺CD8⁻ CD4⁺CD8¯ in % 50 40 30 20 10 0 Unbehandelt Unbehandelt 24h Sham 48h CLP 26G CLP 22G 96h CLP 18G 36h Median Abbildung 30: Darstellung des prozentualen Anteils CD4⁺CD8¯ (Cluster of Differentiation) T-Lymphozyten im Lymphknoten. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLPbehandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. 50 Die Messungen CD8+ T-Lymphozyten aus dem Lymphknoten (Abbildung 31) lieferten bei unbehandelten Tieren mit einem Median von 28,7 % und bei ShamTieren 24, 48 bzw. 96 Stunden nach Operation mit einem Median von 30,1 %, 31,7 % bzw. 29,1 % vergleichbare Ergebnisse. Die Induktion einer Sepsis führte lediglich 96 Stunden nach 22G-CLP zu einer tendenziellen Abnahme CD8+ T-Lymphozyten, wobei ein Großteil der Ergebnisse im Wertebereich der entsprechenden Sham-Gruppe lag. Der Median betrug 25 %. Im Vergleich mit den Kontrolltieren, führte Sepsisinduktion mit der Nadelstärke 18G zu keinen Veränderungen der untersuchten Zellpopulation. Lymphknoten CD4⁻CD8⁺ 40 CD4¯CD8⁺ in % 35 30 25 20 15 10 5 0 Unbehandelt Unbehandelt 24h Sham 48h CLP 26G CLP 22G 96h CLP 18G 36h Median Abbildung 31: Darstellung des prozentualen Anteils CD4¯CD8⁺ (Cluster of Differentiation) T-Lymphozyten im Lymphknoten. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLPbehandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. Bei der Untersuchung CD4-CD8- Zellen (Abbildung 32) fiel in allen Gruppen, außer zum Zeitpunkt 36 Stunden, eine große Streubreite der Einzelwerte auf. Innerhalb der Kontrollgruppen zeigten sich bis auf den Zeitpunkt 24 Stunden (M 37,3 %) vergleichbare Werte. Für unbehandelte Tiere errechnete man einen 51 Median von 32,8 %, für Sham-Tiere 48 bzw. 96 Stunden nach Scheinoperation einen Median von 31,9 % bzw. 31,4 %. Septische Tiere wiesen, verglichen mit den Medianen der Kontrollgruppen, tendenziell einen erhöhten Anteil doppel-negativer Zellen im Lymphknoten auf. 24 Stunden nach 26G- bzw. 22G-CLP war dies mit einem Median von 39,7 % bzw. 39 % gegenüber unbehandelten Tieren festzustellen. Sepsisinduktion mittels 26Gbzw. 22G-Nadel führte nach 48 Stunden mit einem Median von 40,6 % bzw. 43 % zu einer Zunahme doppel-negativer Zellen gegenüber beiden Kontrollgruppen. 96 Stunden nach 26G-CLP errechnete man einen Median von 41,7 %. Nach Induktion einer Sepsis mittels 18G-CLP war bei einem Median von 38,3 % keine Veränderung gegenüber der Sham-Gruppe (M 38,4 %), aber eine Zunahme CD4-CD8- Zellen gegenüber unbehandelten Tieren festzustellen (M 32,8 %). Lymphknoten CD4⁻CD8⁻ 60 CD4ˉCD8¯ in % 50 40 30 20 10 0 Unbehandelt Unbehandelt Abbildung 32: 24h Sham Darstellung des 48h CLP 26G 96h CLP 22G prozentualen 36h CLP 18G Anteils Median CD4¯CD8¯ (Cluster of Differentiation) T-Lymphozyten im Lymphknoten. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22GCLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. 52 3.3.4 Regulatorische T-Zellen Zur Untersuchung der Konzentration regulatorischer T-Zellen (Tregs) während der Sepsis wurden Lymphozyten aus Milz und Lymphknoten mit CD4- und CD25Antikörpern markiert. 3.3.4.1 Milz Bei Kontrolle der Tregs in der septischen Milz, konnte, wie bereits in 3.3.3.1 beschrieben, eine Abnahme CD4⁺ T-Zellen ermittelt werden. Dies war 36, 48 und 96 Stunden nach Sepsisinduktion zu erkennen. Bei der Untersuchung CD4+CD25+ Tregs und CD4-CD25+ T-Lymphozyten aus der Milz wurden keine durch die Sepsis verursachten Konzentrationsveränderungen festgestellt. Bezogen auf die gesamte CD4+ T-Zellpopulation erhöhte sich aber mit zunehmender Sepsisdauer der Anteil der Tregs. Bei Messungen CD4-CD25- T-Zellen fiel bei septischen Tieren eine tendenzielle Zunahme doppelnegativer Zellen auf. Abbildung 33 zeigt repräsentative Punktgraphdarstellungen der CD4- gegenüber der CD25-Intensität in Splenozyten. Um Veränderungen der Zellkonzentrationen im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere zu zeigen sind Sham-Tiere als Kontrollgruppe und 22G- sowie 26G-CLP-Tiere 24 Stunden und 96 Stunden nach Operation dargestellt. 53 24h 3,1 % 96h 4,4 % 2,1 % 3,6% Sham 75,8 % 16,7 % 1,9 % 3,8 % 75,8 % 3,1 % 18,6 % 3,1 % CD25 26G 75, 3% 19 % [ [[Geben SieGein 2,5 % 2,1 % Zitat aus dem G e eDokument oder b die b e 82,2 % 11,6 % [Ge [Geben Sie ein 3 % ben 4,8 % Zitat aus dem Sie Dokument oder 22G ein die 14,2 % 79,1 % Zitat 13,1 % 81,3 % Zusammenfass eZusammenfass n aus ung eines nung eines [Geb [Ge dem interessanten interessanten en S CD4 ben Dok Abbildung 33: Repräsentative Punktgraph-Darstellung ein. Punktes SPunktes Sie Sieein.der CD4(Cluster of i ume Differentiation)-Intensität gegenüber der das CD25-Intensität inein Lymphozyten aus der Milz. Sie können das i Sie können ein e nt Dargestellt ist jeweils ein typisches Ergebnis von Shambzw. 26Gan Textfeld an und 22G(Gauge)-CLP eTextfeldZitat Zitat (cecal ligation and puncture)-Mäusen nach 24 und 96hode (Stunden) bei voneinander einer beliebigen einer aus aus e r die unabhängigen Versuchsdurchführungen mit jeweils vergleichbarem Resultat (CLPbeliebigen eStelle im demi dem Zus Gruppen n=8 bzw. Sham-Gruppe n=4). Stelle im i Dokument Dok n Dok am Dokument npositionieren. ume ume men Verwenden positionieren. nt ZSie nt fass Verwenden Zdie oderi oder Sie ung die die i Registerkarte die t eine Registerkarte t'Textfeldtools', Zusaa Zus s das 'Textfeldtools', awenn Sie mme am t inter des wenn Sie tFormatnfas mendas ess Textfelds Formatfass des sunga ante Textfelds a'Textzitat' eineu ung n 54 'Textzitat' uänderns eine s Pun änderns smöchten.] inter ktes möchten.] essa inter Bei der CD4-Einzelfärbung (Abbildung 34) konnten innerhalb der Kontrollgruppen Schwankungen der Mediane sowie Ausreißer zum Zeitpunkt 48 und 96 Stunden festgestellt werden. In der unbehandelten Gruppe lag der Median bei 16,5 %. Bei der Sham-Gruppe wurde im zeitlichen Verlauf eine Zunahme des Medians deutlich. 24, 48 bzw. 96 Stunden nach Sham-Op betrug der Median 14,4 %, 17,2 % bzw. 19,9 %. Sepsisinduktion mittels CLP bewirkte im zeitlichen Verlauf der Sepsis gegenüber Kontrolltieren eine Verringerung des Anteils CD4+ Splenozyten. 48 Stunden nach CLP errechnete man für 26G-operierte Tiere einen Median von 11,6 %, für 22Goperierte Tiere einen Median von 9 %. 96 Stunden nach 26G- bzw. 22G-CLP betrug der Median 11,5 % bzw. 11,2 %. Verglichen mit unbehandelten Tieren (M 16,5 %) und scheinoperierten Tieren zum Zeitpunkt 36 Stunden (M 15,1 %), verringerte sich nach Sepsisinduktion mit der Nadelstärke 18G mit einem Median von 12,8 % der Anteil CD4+ T-Lymphozyten. Milz CD4⁺CD25⁻ 35 CD4⁺CD25¯ in % 30 25 20 15 10 5 0 Unbehandelt Unbehandelt Abbildung 34: 24h Sham Darstellung des 48h CLP 26G 96h CLP 22G prozentualen 36h CLP 18G Anteils Median CD4⁺CD25¯ (Cluster of Differentiation) T-Lymphozyten in der Milz. Links dargestellt sind unbehandelte und Shambzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLPbehandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. 55 In der CD25-Einzelfärbung (Abbildung 35) ergaben die Messungen in den Kontrollgruppen vergleichbare Ergebnisse. Beim Vergleich der Mediane der CLP-Gruppen mit den Medianen der Kontrollgruppen konnten keine wesentlichen durch die Sepsisinduktion verursachten Konzentrationsveränderungen festgestellt werden. Milz CD4⁻CD25⁺ CD4¯CD25⁺ in % 6 5 4 3 2 1 0 Unbehandelt Unbehandelt Abbildung 35: 24h Sham Darstellung des 48h CLP 26G 96h CLP 22G prozentualen 36h CLP 18G Anteils Median CD4¯CD25⁺ (Cluster of Differentiation) T-Lymphozyten in der Milz. Links dargestellt sind unbehandelte und Shambzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLPbehandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. 56 Die Untersuchung der Tregs aus der Milz (Abbildung 36) zeigte keine deutlichen Unterschiede zwischen Kontrolltieren und septischen Tieren. Der Großteil aller gemessenen Einzelwerte lag zwischen 2 und 4 % anteiliger CD4+CD25+ Lymphozyten. Milz CD4⁺CD25⁺ CD4⁺CD25⁺ in % 6 5 4 3 2 1 0 Unbehandelt Unbehandelt Abbildung 36: 24h Sham Darstellung des 48h CLP 26G 96h CLP 22G prozentualen 36h CLP 18G Anteils Median CD4⁺CD25⁺ (Cluster of Differentiation) T-Lymphozyten in der Milz. Links dargestellt sind unbehandelte und Shambzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLPbehandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. Weder bei der durchflusszytometrischen Untersuchung CD25+ T-Lymphozyten noch bei CD4+CD25+ Tregs aus der Milz kam es, verglichen mit den Kontrollgruppen, zu deutlichen Zu- bzw. Abnahmen im Verlauf der Sepsis. Rechnete man dagegen den Anteil der Tregs bezogen auf die gesamte CD4+ TZellpopulation aus (CD4+/CD4+CD25+ Ratio), erhöhte sich dieser mit zunehmender Sepsisdauer (Abbildung 37). Innerhalb der Kontrollgruppen wurden mit einem Median von 13,5 % in der unbehandelten Gruppe und Medianen von 13 %, 13,5 % bzw. 13 % in der ShamGruppe 24, 48 bzw. 96 Stunden nach Scheinoperation annäherungsweise gleiche Mediane der CD4+/CD4+CD25+ Ratio ermittelt. 57 Im Vergleich mit den Kontrollgruppen zeigte sich bei septischen Tieren mit zunehmender Sepsisdauer eine Erhöhung der CD4 +/CD4+CD25+ Ratio. 24 Stunden nach Sepsisinduktion errechnete man für 26G-operierte Tiere einen leicht erhöhten Median von 16,5 %, für 22G-operierte Tiere einen Median von 14 %, der im Bereich der Kontrollgruppen lag. 48 und 96 Stunden nach CLP wurde eine von der Sepsisschwere abhängige Zunahme der CD4 +/CD4+CD25+ Ratio deutlich. Zum Zeitpunkt 48 Stunden nach CLP mittels 26G-Nadel betrug der Median 21,3 %, mittels 22G-Nadel 25 %. 96 Stunden nach Sepsisinduktion mit der Nadelstärke 26G berechnete man einen Median von 21 %, nach Sepsisinduktion mit der Nadelstärke 22G einen Median von 24,5 %. Die Durchführung der 18G-CLP führte bei einem Median von 21,1 % ebenfalls zu einer tendenziellen Zunahme der CD4+/CD4+CD25+ Ratio verglichen mit der unbehandelten Gruppe (M 13,5 %) und der Sham-Gruppe (M 17,7 %). CD4⁺/CD4⁺CD25⁺ Ratio in % Milz CD4⁺/CD4⁺CD25⁺ Ratio 35 30 25 20 15 10 5 0 Unbehandelt Unbehandelt Sham 24h 48h CLP 26G CLP 22G 96h CLP 18G 36h Median Abbildung 37: Darstellung des prozentualen Anteils der CD4⁺/CD4⁺CD25⁺ (Cluster of Differentiation) Ratio von T-Lymphozyten in der Milz. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22GCLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die errechneten Einzelwerte mit entsprechendem Median pro Versuchsgruppe. 58 Die Messungen doppelnegativer Zellen in der Milz (Abbildung 38) ergaben innerhalb der Kontrollgruppen ähnliche Ergebnisse. Für unbehandelte Tiere errechnete man einen Median von 78,5 %, für scheinoperierte Tiere 24, 48 bzw. 96 Stunden postoperativ einen Median von 79,5 %, 79,2 % bzw. 78,4 %. Septische Tiere zeigten, verglichen mit den Kontrollgruppen, eine tendenzielle Zunahme doppelnegativer Splenozyten, die jedoch beim Vergleich der Mediane sehr gering ausfiel. Für 26G-operierte Tiere errechnete man 24, 48 bzw. 96 Stunden nach CLP einen Median von 79,6 %, 82,6 % bzw. 82,3 %. Die 22G-CLP führte nach 24, 48 bzw. 96 Stunden zu einem Median von 81,1 %, 84,4 % bzw. 82,3 %. Auch nach Induktion einer Sepsis mit der Nadelstärke 18G zeigte sich im Vergleich zu den Kontrollgruppen eine geringe Zunahme der CD4-CD25Zellfraktion. Der Median der CLP-Tiere betrug 82,1 %, der Median der ShamGruppe zum Zeitpunkt 36 Stunden 79,6 % und der unbehandelten Tiere 78,5 %. Milz CD4⁻CD25⁻ CD4¯CD25¯ in % 100 80 60 40 20 0 Unbehandelt Unbehandelt Abbildung 38: 24h Sham Darstellung des 48h CLP 26G CLP 22G prozentualen 96h 36h CLP 18G Median Anteils CD4¯CD25¯ (Cluster of Differentiation) T-Lymphozyten in der Milz. Links dargestellt sind unbehandelte und Shambzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLPbehandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. 59 3.3.4.2 Lymphknoten Zur Untersuchung von Tregs aus Lymphknoten im Verlauf der Sepsis wurde eine Antikörperfärbung mit CD4 und CD25 durchgeführt. Es zeigte sich im zeitlichen Verlauf der Sepsis, verglichen mit unbehandelten und Sham-operierten Tieren, in septischen Lymphknoten eine tendenzielle Abnahme des Anteils CD4+ T-Helferzellen nach 36, 48 und 96 Stunden. Bei Messungen der Treg-Konzentration konnte bei septischen Mäusen eine geringe Abnahme gegenüber den entsprechenden Sham-Gruppen beobachtet werden. Der Anteil CD4-CD25- Zellen war verglichen mit den Kontrollgruppen vor allem zu den Zeitpunkten 36 und 48 Stunden, aber auch leicht zum Zeitpunkt 96 Stunden erhöht. Die Anteile CD4+ T-Lymphozyten (Abbildung 39) wiesen innerhalb der Kontrollgruppen Unterschiede auf. Für unbehandelte Tiere errechnete man mit 35,9 % einen höheren Median als für Sham-operierte Tiere. Innerhalb der ShamGruppe war eine tendenzielle Zunahme CD4 + T-Lymphozyten zu beobachten. 24 Stunden nach Scheinoperation betrug der Median 28,4 %, nach 48 Stunden 29,8 % und nach 96 Stunden 31,7 %. Es fiel in allen Kontrollgruppen eine breite Streuung der Einzelwerte auf. Verglichen mit unbehandelten Tieren war in allen CLP-Gruppen geringere CD4+ Konzentrationen festzustellen. Beim Vergleich der CLP-Gruppen mit entsprechender Sham-Gruppe zeigte sich zum Zeitpunkt 48 Stunden nach Sepsisinduktion mit der Nadelstärke 26- bzw. 22G mit einem Median von 24,1% bzw. 22,7 % eine tendenziellen Abnahme des Anteils CD4+ T-Zellen. Gegenüber scheinoperierten Tieren wurde auch 96 Stunden nach 26G- bzw. 22G-CLP mit einem Median von 27,2 % bzw. 27,6 % eine leichte Abnahme der untersuchten Zellpopulation deutlich. Gegenüber CLP-Tieren zum Zeitpunkt 48 Stunden war eine erneute Zunahme CD4+ T-Zellen festzustellen. Die Induktion einer Sepsis mit der Nadelstärke 18G führte ebenfalls zu einer Abnahme CD4+ Zellen. Im Vergleich zu scheinoperierten Tieren zum Zeitpunkt 36 Stunden mit einem Median von 27,7% und unbehandelten Tieren (M 35,9 %), errechnete man für die CLP-Gruppe einen geringeren Median von 24,5 %. 60 Lymphknoten CD4⁺CD25⁻ CD4⁺CD25¯ in % 60 50 40 30 20 10 0 Unbehandelt Unbehandelt Abbildung 39: 24h Sham Darstellung 48h CLP 26G des 96h CLP 22G prozentualen 36h CLP 18G Anteils Median CD4⁺CD25¯ (Cluster of Differentiation) T-Lymphozyten im Lymphknoten. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22GCLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. Bei der Überprüfung CD25+ Lymphozyten (Abbildung 40) lag innerhalb der Kontrollgruppen der Median zwischen 2,3 % und 3 %. Zum Zeitpunkt 48 Stunden fielen Ausreißer in der Sham-Gruppe auf. Weder Sepsisinduktion mittels 26G- noch mittels 22G- oder 18G-CLP führte zu Veränderungen des Anteils CD25+ T-Lymphozyten verglichen mit den entsprechenden Kontrollgruppen. Zu den Zeitpunkten 48 und 96 Stunden waren Ausreißer zu erkennen. 61 Lymphknoten CD4⁻CD25⁺ 7 CD4¯CD25⁺ in % 6 5 4 3 2 1 0 Unbehandelt Unbehandelt Abbildung 40: 24h Sham Darstellung des 48h CLP 26G 96h CLP 22G prozentualen 36h CLP 18G Anteils Median CD4¯CD25⁺ (Cluster of Differentiation) T-Lymphozyten im Lymphknoten. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22GCLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. Der Anteil CD4+CD25+ Tregs (Abbildung 41) lag bei unbehandelten Tieren im Median bei 7,4 %. In der Sham-Gruppe war eine Zunahme im zeitlichen Verlauf festzustellen. 24, 48 bzw. 96 Stunden nach Scheinoperation betrug der Median 6,6 %, 9,8 % bzw. 10,3 %. Zum Zeitpunkt 96 Stunden fielen Ausreißer auf. Die Untersuchung der Treg-Konzentration in septischen Lymphknoten zeigte keine Veränderungen zwischen unbehandelten Tieren und CLP-Tieren. Beim Vergleich der Sham-Gruppen mit den entsprechenden CLP-Gruppen fielen Abweichungen der Mediane auf. 24 Stunden nach Sepsisinduktion mittels 22G-CLP errechnete sich ein Median von 5,6 %. Verglichen mit der Sham-Gruppe führten sowohl eine 26G- als auch eine 22G-CLP nach 48 Stunden zu einer tendenziellen Abnahme der Treg-Konzentration. Der Median nach 26G-CLP lag bei 6,4 %, nach 22G-CLP bei 7 %. 96 Stunden nach 22G-CLP betrug der Median 8,1 %, nach 22G-CLP 8 %. Die Induktion einer Sepsis mit der Nadelstärke 18G hatte verglichen mit den Kontrollgruppen keinen wesentlichen Einfluss auf die Konzentration der Tregs. 62 Lymphknoten CD4⁺CD25⁺ CD4⁺CD25⁺ in % 20 15 10 5 0 Unbehandelt Unbehandelt Abbildung 41: 24h Sham Darstellung des 48h CLP 26G 96h CLP 22G prozentualen 36h CLP 18G Anteils Median CD4⁺CD25⁺ (Cluster of Differentiation) T-Lymphozyten im Lymphknoten. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22GCLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. Bei der durchflusszytometrischen Untersuchung von Tregs aus dem Lymphknoten kam es, verglichen mit den entsprechenden Sham-Gruppen, zu einer tendenziellen Abnahme im Verlauf der Sepsis. Errechnete man, wie bei der Milz geschehen, den Anteil der Tregs bezogen auf die gesamte CD4+ T-Zellpopulation, konnten im Gegensatz zu der Milz, in den Lymphknoten keine deutlichen Veränderungen der CD4+/CD4+CD25+ Ratio festgestellt werden. Es fiel sowohl in den Kontrollgruppen als auch in den CLP-Gruppen eine breite Streuung der Einzelwerte auf (Abbildung 42). Innerhalb der Kontrollgruppen wurden mit einem Median von 16,9 % in der unbehandelten Gruppe und Medianen von 19,5 %, 23,5 % bzw. 22,2 % in der Sham-Gruppe 24, 48 bzw. 96 Stunden nach Scheinoperation unterschiedliche Mediane der CD4+/CD4+CD25+ Ratio ermittelt. Während die Induktion einer Sepsis mit der Nadelstärke 26G im Vergleich zu den Sham-Gruppen keine Veränderungen bewirkte, nahmen die Mediane der 26G- 63 CLP-Gruppen im Vergleich zur unbehandelten Gruppe zu. 24, 48 bzw. 96 Stunden nach 26G-CLP errechnete man einen Median von 19,5 %, 21,4 % bzw. 22,8 %. 24 Stunden nach Sepsisinduktion mittels 22G-CLP errechnete man einen von der Sham-Gruppe erniedrigten Median von 15,6 %. Mit 24,5 % bzw. 21,8 % lagen die Mediane 48 bzw. 96 Stunden nach 22G-CLP im Bereich der entsprechenden Sham-Gruppen. Gegenüber der unbehandelten Gruppe nahm die CD4+/CD4+CD25+ Ratio zu. Die Induktion einer Sepsis mit der Nadelstärke 18G führte verglichen mit den Kontrollgruppen zu keinen Veränderungen der CD4+/CD4+CD25+ Ratio. CD4⁺/CD4⁺CD25⁺ Ratio in % Lymphknoten CD4⁺/CD4⁺CD25⁺ Ratio 35 30 25 20 15 10 5 0 Unbehandelt Unbehandelt 24h Sham 48h CLP 26G CLP 22G 96h CLP 18G 36h Median Abbildung 42: Darstellung des prozentualen Anteils der CD4⁺/CD4⁺CD25⁺ (Cluster of Differentiation) Ratio von T-Lymphozyten im Lymphknoten. Links dargestellt sind unbehandelte und Sham- bzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLP-behandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. Die Messungen doppel-negativer Zellen (Abbildung 43) zeigten innerhalb der Kontrollgruppen, außer 24 Stunden nach Sham-Op, vergleichbare Mediane. Für unbehandelte Tiere betrug der Median CD4-CD25- Zellen 55,3 %, für Sham-Tiere 24, 48 bzw. 96 Stunden nach Scheinoperation 62,2 %, 57,3 % bzw. 56,6 %. In der unbehandelten Gruppe fiel eine große Streubreite der Einzelwerte auf. 64 Die Induktion einer Sepsis führte zur tendenziellen Zunahme doppel-negativer Zellen. 24 Stunden nach Sepsisinduktion lag der Median nach 26G-CLP mit 64,4 % und nach 22G-CLP mit 63 % im Bereich des Median der Sham-Gruppe. Verglichen mit der unbehandelten Gruppe stellte man eine Zunahme der untersuchten Zellpopulation fest. Eine Erhöhung des Median gegenüber beiden Kontrollgruppen war mit einem Median von 65,7 % bzw. 66,9 % nach Induktion einer Sepsis mit der Nadelstärke 26- bzw. 22G zum Zeitpunkt 48 Stunden zu erkennen. Auch 96 Stunden nach 26G- bzw. 22G-CLP zeigten sich gegenüber den Kontrollgruppen leicht erhöhte Mediane von 61,9 % bzw. 60,8 %. 36 Stunden nach Sepsisinduktion mit einer 18G-Nadel stellte man eine Zunahme CD4-CD25- Zellen im Lymphknoten fest. Im Vergleich zur unbehandelten Gruppe (M 55,3 %) und Sham-operierten Tieren 36 Stunden postoperativ (M 61,8 %), errechnete man nach 18G-CLP einen Median von 67,2 %. Lymphknoten CD4⁻CD25⁻ 80 CD4¯CD25¯ in % 70 60 50 40 30 20 10 0 Unbehandelt Unbehandelt Abbildung 43: 24h Sham Darstellung des 48h CLP 26G 96h CLP 22G prozentualen 36h CLP 18G Anteils Median CD4 -CD25- (Cluster of Differentiation) T-Lymphozyten im Lymphknoten, eingeteilt in unbehandelte und Shambzw. CLP(cecal ligation and puncture)-behandelte Mäuse 24, 48 und 96h (Stunden) nach Scheinoperation bzw. Sepsisinduktion mittels 26G(Gauge)- bzw. 22G-CLP (unbehandelt n=4, Sham-Gruppen n=4, CLP Gruppen n=8). Rechts dargestellt sind Sham- bzw. CLPbehandelte Mäuse 36h nach Scheinoperation bzw. Sepsisinduktion mittels 18G-CLP (Sham-Gruppe n=2, CLP Gruppe n=4). Aufgeführt sind die gemessen Einzelwerte mit entsprechendem Median pro Versuchsgruppe. 65 4 Diskussion 4.1 Einleitung Die Sepsis stellt ein schwerwiegendes und für das Gesundheitswesen kostenintensives Krankheitsbild dar. Trotz intensiver Forschung konnten bislang keine durchschlagenden Erfolge in der Entwicklung neuer Therapien erzielt werden. In der Pathophysiologie der Sepsis gehen aktuelle Theorien von einem zweiphasigen Verlauf der Erkrankung aus. Zu Beginn reagiert der Wirt auf eine Infektion durch exzessive Produktion proinflammatorischer Mediatoren mit einer überschießenden Immunantwort im Sinne eines SIRS (Systemic Inflammatory Response Syndrome). Im Anschluss stellt sich eine kompensatorische antiinflammatorische Immunparalyse ein. Ein genauer zeitlicher und kausaler Zusammenhang konnte bislang im Detail nicht aufgeklärt werden. Ziel der vorliegenden Arbeit war, die Veränderungen des Immunsystems, im Speziellen der T-Lymphozyten, im Verlauf der Sepsis besser zu verstehen, da sie durch ihre immunregulatorische Bedeutung in der Pathophysiologie der Sepsis eine wichtige Rolle zu spielen scheinen. Als mögliche Ursache eines immunsuppressiven Zustandes in der Spätphase der Sepsis wurde schon früh eine erhöhte Apoptoserate von Thymozyten und peripheren Lymphozyten festgestellt [2, 48]. Die veränderte Mediatorenfreisetzung, durch eine Verschiebung von TH1- zu TH2T-Zellen, wird als weiterer möglicher Pathomechanismus des zweiphasigen Verlaufs der Sepsis angesehen. Nach erhöhten Konzentrationen von TH1Zytokinen in der Frühphase der Sepsis ist im weiteren Verlauf eine Verschiebung in Richtung einer antiinflammatorisch wirkenden TH2-Zytokin-Dominanz festzustellen [19, 39, 49, 50]. Des Weiteren belegen Studien, dass T-lymphozytäre Subpopulationen die Fähigkeit aufweisen, eine adaptive Immunantwort zu unterdrücken [51]. Eine Zunahme dieser regulatorischer Zellen könnte in der Pathophysiologie der Sepsis eine Rolle spielen. Um die immunologischen Veränderungen und deren Auswirkungen auf den Wirt im Verlauf der Sepsis besser zu verstehen, beschäftigte sich die vorliegende Arbeit mit der durchflusszytometrischen Charakterisierung von Thymozyten, T66 Helferzellen, zytotoxischen T-Zellen und Tregs weiblicher Wildtypmäuse im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere. Die Induktion einer Sepsis erfolgte durch Ligatur des Zökums und anschließender Punktion (CLP) mit unterschiedlichen Nadelstärken. Das Sepsismodell der CLP bei der Maus wurde erstmals 1983 von Baker et al. beschrieben und stellt das vorherrschende Tiermodell einer polymikrobiellen intraabdominellen Sepsis dar, um die klinischen Stadien der Sepsis zu imitieren [3, 37]. 4.2 Überlebenszeitanalyse Zu Beginn der Arbeit wurden Versuche zur Evaluation der Überlebenszeiten weiblicher Wildtypmäuse nach zökaler Ligatur und Punktion durchgeführt. Dabei konnte ein von der Nadelstärke abhängiges Überleben festgestellt werden. Die geringste Überlebenswahrscheinlichkeit war nach CLP der Nadelstärke 18 Gauge (G) mit einem Außendurchmesser (AD) von 1,2 mm zu verzeichnen und betrug für den Beobachtungszeitraum von 20 Tagen 12,5 %. Sepsisinduktion mittels 22GCLP (AD 0,7 mm) führte ebenfalls zu einer deutlichen Abnahme der Überlebenswahrscheinlichkeit auf 62,5 %. In der 26G-Gruppe (AD 0,45 mm) lag die Überlebensrate mit 87,5 % innerhalb der CLP-Gruppen am höchsten. Die für die Punktion verwendete Nadelstärke spiegelte somit den Schweregrad der Sepsis wider. Dies wird durch eine Studie von Otero-Antón et al. bekräftigt, die eine Zunahme der Mortalität bei steigendem Durchmesser der Punktionsnadel bei der CLP beschreibt [30]. Um Veränderungen in verschiedenen Schweregraden der Sepsis zu untersuchen, wurden die nachfolgenden Versuche mit den Nadelstärken 26-, 22- und 18G durchgeführt. Ferner konnten durch die Ergebnisse der Überlebenszeitanalyse die Tötungszeitpunkte für die anschließenden durchflusszytometrischen Messungen von T-Lymphozyten während der Sepsis festgelegt werden. Da am zweiten Tag nach 18G-CLP bereits 50 % der Versuchstiere verstorben waren, wurde für diese Gruppe als Zeitpunkt des Abtötens 36 Stunden nach CLP gewählt, um dennoch möglichst aussagekräftige Veränderungen der Immunzellen in schweren Sepsisstadien zu erhalten. Für 26G- und 22G-operierte Tiere wurden als Tötungszeitpunkte 24, 48 und 96 Stunden nach CLP festgelegt, um Aussagen 67 über mögliche Veränderungen der Immunzellen sowohl in der Früh- als auch in der Spätphase der Sepsis treffen zu können. Sham-operierte Versuchstiere wiesen bei der Überlebenszeitanalyse eine Überlebenswahrscheinlichkeit von 100 % für den Beobachtungszeitraum auf. Dadurch konnte eine schwerwiegende Erkrankung der Kontrolltiere, z.B. durch perioperative Infektion oder Stress, ausgeschlossen werden. 4.3 Auswertung der Narkoseprotokolle der Immunphänotypisierung Die bei der Überlebenszeitanalyse gewonnenen Ergebnisse wurden, beim Vergleich mit den Daten der für die durchflusszytometrische Untersuchung von TLymphozyten verwendeten Überlebensrate mit Versuchstiere, bekräftigt. zunehmendem Außendurchmesser Demnach der sank bei der die CLP verwendeten Nadelstärke. Nach Sepsisinduktion starben 11 der insgesamt 109 Versuchstiere vor dem Zeitpunkt des geplanten Abtötens, davon vier Tiere der Gruppe 4 (22G-CLP, geplantes Abtöten nach 48 Stunden) und sieben Tiere der Gruppe 5 (18G-CLP, geplantes Abtöten nach 36 Stunden). In den übrigen CLPGruppen betrug die Überlebensrate 100 %. Das Ableben der sieben von insgesamt 13 Tieren der Gruppe 5, was einer Überlebensrate von 46,2 % entspricht, ist durch den großen Defekt des Darmes, der durch den Außendurchmesser von 1,2 mm der verwendeten Nadelstärke 18G entsteht, zu erklären. Wie in der Überlebenszeitanalyse verstarben die meisten Tiere am zweiten Tag nach Sepsisinduktion mittels 18G-CLP. Narkose- und Operationsdauer sowie das Alter der verstorbenen Tiere lagen in Nähe des Durchschnitts der restlichen CLP-Gruppen und lieferten somit keine weitere Erklärungsmöglichkeiten für das geringe Überleben. Vier von insgesamt 37 Tieren der Gruppe 4 verstarben innerhalb von 48 Stunden, ein Tier am ersten und drei am zweiten Tag nach CLP mit der Nadelstärke 22G. Die Wahrscheinlichkeit, den zweiten postoperativen Tag zu überleben, lag bei 89,2 % und veränderte sich bis 96 Stunden nach CLP nicht. Im Gegensatz zur Überlebenszeitanalyse traten die Todesfälle nach Sepsisinduktion mit der Nadelstärke 22G früher auf. Obwohl jungen Mäusen ein Überlebensvorteil nach Durchführung einer CLP bescheinigt wird [42], wiesen drei der vier verstorbenen Tiere mit einem Alter von 98 Tagen das geringste Alter aller Versuchstiere auf. Mit 68 einer überdurchschnittlich langen Narkosedauer von 28 Minuten und einer OpDauer von 16 Minuten lag ein Versuchstier weit außerhalb der durchschnittlichen Zeiten, was eine Ursache für dessen frühzeitiges Versterben sein könnte. Weiterhin kann die Mortalität von der Durchführung der Ligatur des Zökums abhängen, wie Singleton et al. [40] bei Versuchen an der Ratte beschreiben. Beträgt der ligierte Abschnitt mehr als 30 % der Zökumlänge liegt die Mortalität vier Tage nach CLP mit der Nadelstärke 20G zwischen 90 und 100 %. Liegt dieser Abschnitt unter 5 %, ist keine Mortalität zu verzeichnen. Deshalb wurde bei der Ligatur auf eine konstante Länge ligierten Zökums von ungefähr 20 bis 30 % geachtet. Sham-operierte Tiere überlebten alle den Versuch bis zur Organentnahme 24, 36, 48 bzw. 96 Stunden nach Laparotomie. Demzufolge konnte eine schwerwiegende Erkrankung der Sham-Tiere während des Versuchsablaufs ausgeschlossen werden. 4.4 T-Lymphozyten im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisschwere Beim Betrachten der Einzelwerte innerhalb der Kontrollgruppen fielen bei einigen zytometrischen Analysen Ausreißer auf. Dies könnte durch eine Infektion während der Sham-Operation oder durch Stress im Käfig verursacht sein, da Kontrolltiere und CLP-Tiere zusammen im Käfig gehalten wurden. Umweltstressoren wie langanhaltender physischer oder emotionaler Stress, können die HypothalamusHypophysen-Nebennierenrinden-Achse aktivieren, was zur vermehrten Produktion des Stresshormons Kortisol führt und wiederum eine Thymusinvolution und eine Abnahme der Thymopoese verursachen kann [15]. In zukünftigen Studien soll evaluiert werden, ob Sham-Tiere stressbedingte Veränderungen der Lymphozytenkonzentration aufweisen und ob dies durch getrennte Haltung im Käfig vermieden werden kann. Von einer Infektion Sham-operierter Tiere ist nicht auszugehen, da auf steriles Arbeiten geachtet wurde und Ausreißer auch bei unbehandelten Tieren auftraten. 69 4.4.1 T-Helferzellen, zytotoxische T-Zellen und doppelpositive T- Lymphozyten aus dem Thymus T-Lymphozyten spielen eine wichtige Rolle als Träger der zellvermittelten Immunität im adaptiven Immunsystem [41]. Bei der Entstehung der Immunparalyse in der Spätphase der Sepsis stellen T-Lymphozyten aufgrund erhöhter Apoptoseraten und vermindertem lymphoproliferativen Ansprechen auf Fremdantigene wohl einen zentralen pathogenetischen Mechanismus dar. Um den Einfluss der Sepsis auf die Konzentration CD4+, CD8+ und CD4+CD8+ Thymozyten während der Sepsis zu untersuchen, wurden aus dem Thymus gewonnene Lymphozyten mit CD4- und CD8-Antikörper markiert. Dabei konnte im Thymus septischer Mäuse, im Vergleich mit Sham-operierten Tieren, eine signifikante Abnahme der CD4⁺CD8⁺ Zellpopulation festgestellt werden. 48 Stunden nach Sepsisinduktion war dies von der Sepsisschwere abhängig und nur nach 22G-CLP zu beobachten. Zum Zeitpunkt 96 Stunden wurden sowohl nach 26G- als auch nach 22G-CLP deutlich geringere Konzentrationen CD4⁺CD8⁺ Thymozyten gemessen. Desweiteren wurde beim Vergleich der einzelnen Zeitpunkte eine Abnahme der untersuchten Zellpopulation im zeitlichen Verlauf der Sepsis deutlich. Die durch erhöhte Apoptose bedingte Abnahme unreifer CD4⁺CD8⁺ Thymozyten während der Sepsis stellt nach aktuellem Wissensstand einen bedeutenden Pathomechanismus bei der Entstehung der Immunparalyse dar und konnte in der vorliegenden Studie bestätigt werden [2, 14, 15, 48]. Bei der Untersuchung des Anteils CD4⁺ T-Helferzellen im Thymus fiel eine von der Sepsisschwere abhängige Zunahme der Zellpopulation auf. Im Vergleich mit Sham-Tieren wurden 36 Stunden nach Sepsisinduktion mittels 18G-CLP signifikant höhere Konzentrationen CD4+ Thymozyten gemessen. Zum Zeitpunkt 48 Stunden war zwischen Sham-Tieren und 22G-operierten Tieren ebenfalls eine Zunahme von CD4+ T-Helferzellen festzustellen. 96 Stunden nach 22G-CLP wurde der Anstieg sowohl gegenüber Sham-Tieren als auch 26G-operierten Tieren deutlich. Dabei nahm der Anstieg der CD4⁺ T-Zellen mit zunehmender Sepsisdauer zu. Diese Ergebnisse werden durch eine Studie von Ayala et al. [2] bestätigt, welche bei septischen Mäusen einen Anstieg CD4+ Thymozyten bei unveränderter Apoptoserate beschreibt. Dies steht im Widerspruch zu einer Studie 70 von Hotchkiss et al. [23], die eine erhöhte Apoptoserate von CD4⁺ Thymozyten während der Sepsis darstellt. Dies war allerdings bei Bcl-2-Immunglobulintransgenen Mäusen nachzuweisen, Wildtypmäuse wurden nicht untersucht. Ebenso spricht die Abnahme der CD4+CD8+ Zellpopulation, aus der sich CD4+ TZellen entwickeln, gegen eine Zunahme CD4+ T-Helferzellen. Mögliche Ursachen für die Zunahme CD4+ T-Zellen könnten zum einen geringere Apoptoseraten oder eine Aktivierung im Rahmen der Sepsis sein, die zur vermehrten Bildung dieser Zellen führt und durch erhöhte Proliferation nachzuweisen wäre. Auch für zytotoxische T-Zellen wurde während der Sepsis ein Anstieg der Konzentration festgestellt. 36 Stunden nach 18G-CLP sowie 48 Stunden nach 22G-CLP nahm im Vergleich zu den entsprechenden Sham-Gruppen der Anteil CD8⁺ T-Lymphozyten im Thymus signifikant zu und war somit nur in schweren Sepsisstadien nachzuweisen. Außerdem fiel, beim Vergleich zwischen den einzelnen Zeitpunkten innerhalb der jeweiligen CLP-Gruppe, im zeitlichen Verlauf der Sepsis eine Zunahme der CD8+ Thymozyten auf. Diese Ergebnisse werden durch die bereits erwähnte Studie von Ayala et al. [2] bekräftigt, in der, im Vergleich zu Sham-operierten Kontrolltieren, bei septischen Mäusen eine Zunahme CD8+ Thymozyten beschrieben ist. Gegen die Zunahme + CD8 Thymozyten spricht der in der vorliegenden Arbeit erbrachte Nachweis einer gestörten T-Lymphopoese. Für ein besseres Verständnis sind hier weitere Studien notwendig, die prüfen, welche Subpopulationen von CD8+ T-Zellen während der Sepsis zu- bzw. abnehmen. Ein Anstieg immunsuppressiv wirkender CD8+ regulatorischer T-Zellen könnte an der Immunparalyse der Sepsis beteiligt sein [26]. Diese Theorie wird durch Studien bekräftigt, die CD8-depletierten Tieren erhöhte Überlebenswahrscheinlichkeiten nach CLP bescheinigen [10, 38]. Von weiterem Interesse wäre, den Zellumsatz sowie das „Homing“ von TLymphozyten, also die Rückkehr in sekundär lymphatische Organe nach Antigenkontakt zur spezifischen Proliferation, während der Sepsis zu untersuchen. Beide Faktoren könnten an den Konzentrationsveränderungen von CD4 + und CD8+ Thymozyten während der Sepsis beteiligt sein. Desweiteren nahm die CD4⁻CD8⁻ Zellpopulation, im Vergleich zur entsprechenden Sham-Gruppe, 96 Stunden nach Sepsisinduktion mittels 26GCLP signifikant zu. Auch im zeitlichen Verlauf der Sepsis wurde innerhalb der einzelnen CLP-Gruppen die Zunahme doppel-negativer Thymozyten signifikant. 71 Dies könnte durch eine Störung der Entwicklung von Thymozyten im doppelnegativen Stadium verursacht sein und wurde nachfolgend untersucht (sh. 4.4.2). 4.4.2 Doppel-negative Thymozyten Die durchgeführten FACS-Analysen zeigten, dass mit zunehmender Sepsisdauer der prozentuale Anteil CD4⁺CD8⁺ T-Zellen im Thymus deutlich abnimmt, während die relativen Anteile der CD4⁻CD8⁻ Zellen zunehmen. Dies stimmt mit Untersuchungen von Ayala et al. an septischen Mäusen überein [2]. Dieses Ergebnis lässt die Schlussfolgerung zu, dass neben einer erhöhten Apoptose der CD4⁺CD8⁺ Thymozyten zusätzlich die intrathymale T-Zellentwicklung im CD4⁻CD8⁻ DN-Vorläufer-T-Zellstadium durch die Sepsis inhibiert sein könnte. Um festzustellen, in welcher der vier CD4⁻CD8⁻ DN-Entwicklungsphasen dies auftritt, wurden die relativen Anteile doppel-negativer Thymozyten während der Sepsis mittels FACS-Analysen bestimmt. Dabei konnte im zeitlichen Verlauf der Sepsis eine tendenzielle Zunahme von CD44⁺CD25⁻ (DN1), CD44⁺CD25⁺ (DN2) und CD44⁻CD25⁺ (DN3) Thymozyten ermittelt werden. Eine Abhängigkeit von der Sepsisschwere war zu den Zeitpunkten 24 und 96 Stunden nach CLP festzustellen. Bei den Messungen CD44⁻CD25⁻ (DN4) Thymozyten wurde dementsprechend eine Abnahme beobachtet. Beim Betrachten der Veränderungen im zeitlichen Verlauf der Sepsis wurde deutlich, dass sich die Zu- bzw. Abnahme doppel-negativer ThymozytenUntergruppen, wie die Abnahme von CD4⁺CD8⁺ Thymozyten, mit der Dauer der Sepsis verstärkt. Dies ist ein Hinweis darauf, dass die Thymopoese mit der Dauer der Sepsis in bestimmten Stadien der Lymphopoese zunehmend beeinträchtigt wird. Diese Ergebnisse gehen mit der Auswanderung naiver T-Zellen in die Peripherie, der Thymusatrophie und der Auszehrung des Kortex während der Sepsis einher [15] und stellen neben der Apoptose der CD4⁺CD8⁺ Thymozyten eine weitere mögliche Ursache bei der Entstehung der Immunparalyse in der Spätphase der Sepsis dar. Als Pathomechanismus der beschriebenen Veränderungen ist neben einer erhöhten Apoptose von CD44⁻CD25⁻ (DN4) Thymozyten auch eine Störung der T72 Lymphopoese in den früheren Stadien möglich. Um dies zu klären, ist die Bestimmung der Apoptoseraten in den Entwicklungsstadien der Thymopoese notwendig. In einer Studie von Baseta et al. induziert TNF (Tumor Nekrose Faktor) die Apoptose der CD44⁻CD25⁺ (DN3) und CD44⁻CD25⁻ (DN4) Thymozyten, und in höheren Dosen auch eine Proliferation der DN3-Population [4]. Die bei Sepsis erhöhten Werte des proinflammatorisch wirkenden TNF könnten so als Initiator einer erhöhten Apoptose von CD44⁻CD25⁻ (DN4) Thymozyten wirken. Zum anderen könnte eine Blockade in der Entwicklung der Thymozyten Ursache für die beobachteten Veränderungen sein. Dies ist durch Messung der Proliferation von Thymozyten in vivo, mittels Markierung mit Bromdesoxyuridin (BrdU) zum Nachweis stattgefundener DNA-Synthese, zu klären. Für die Zukunft sind hier weitere Untersuchungen notwendig, um den genauen Pathomechanismus der gestörten T-Lymphopoese im doppel-negativen Stadium und einen eventuellen Zusammenhang mit Mediatoren aufzudecken. 4.4.3 T-Helferzellen und zytotoxische T-Zellen aus der Peripherie Erhöhte Apoptoseraten und verminderte Proliferationskapazität von Lymphozyten sind während der Sepsis neben dem Thymus auch in sekundär lymphatischen Organen beschrieben [2, 20, 23, 43]. Um Veränderungen der Anteile der CD4+ und CD8+ T-Zellen aus der Peripherie während der Sepsis zu untersuchen, wurden Lymphozyten aus Milz und Lymphknoten mit CD4- und CD8-Antikörper markiert. 4.4.3.1 Milz In der Milz konnte während der Sepsis eine tendenzielle Abnahme CD4⁺ TLymphozyten beobachtet werden. Dies wurde insbesondere zu den Zeitpunkten 48 und 96 Stunden nach 26G- und 22G-CLP deutlich. Die Abnahme CD4⁺ THelferzellen in der septischen Milz wird durch zahlreiche Studien bestätigt und ist durch Sepsis-induzierte Apoptose von Lymphozyten verursacht, die einen zentralen Pathomechanismus in der Entstehung der Immunparalyse in der Spätphase der Sepsis darstellt [23, 24, 50]. Des Weiteren können der in der vorliegenden Studie nachgewiesene Verlust an CD4+CD8+ Thymozyten sowie die gestörte Entwicklung doppel-negativer Thymozyten während der Sepsis eine geringere Abgabe naiver T-Zellen an die Peripherie verursachen, und somit an der 73 Abnahme CD4+ Splenozyten beteiligt sein. Ein weiterer möglicher Pathomechanismus stellt eine Inhibition der Proliferation CD4⁺ T-Helferzellen durch Aktivierung von Tregs während der Sepsis dar. Dies wird durch den in der vorliegenden Arbeit nachgewiesenen zeitlichen Zusammenhang zwischen der relativen Zunahme von Tregs und der Abnahme CD4+ Splenozyten bekräftigt (sh 4.4.4). Um dies zu belegen, sollte die Proliferation CD4+ T-Zellen während der Sepsis mittels in vivo Untersuchungsmethode BrdU-Applikation ermöglicht auch das untersucht „Homing“, werden. Diese Auswandern der Lymphozyten in sekundär lymphatische Organe zur spezifischen Proliferation nach Antigenkontakt, als weitere mögliche Ursache von Konzentrationsveränderung im Rahmen einer Sepsis zu überprüfen. Die Abnahme der CD4+ T-Zellpopulation steht im Widerspruch zur massiven klonalen Expansion, die im Rahmen einer Immunantwort physiologischerweise zu erwarten wäre [24]. Für CD8⁺ Splenozyten konnte in den durchgeführten Versuchen eine tendenzielle Abnahme während der Sepsis nachgewiesen werden. 36 Stunden nach 18G-CLP sowie 48 und 96 Stunden nach 26G- sowie 22G-CLP wurden im Vergleich mit den Kontrollgruppen geringere Mediane errechnet. Dies wird durch die gestörte Thymopoese sowie durch eine Studie von Hotchkiss et al. bekräftigt, in der erhöhte Apoptoseraten CD8+ T-Lymphozyten in der Milz CLP-behandelter Mäuse beschrieben werden [23]. Auch bei der Untersuchung der Milz an Sepsis verstorbener Patienten wurden eine Abnahme der Lymphozytendichte sowie erhöhte Apoptose von Lymphozyten nachgewiesen [21]. Für die CD4⁻CD8⁻ Zellpopulation wurde während der Sepsis eine tendenzielle Zunahme festgestellt. Dies ist durch die beschriebene Abnahme der CD4⁺ und CD8⁺ T-Lymphozyten zu erklären. 4.4.3.2 Lymphknoten Bei der Untersuchung von CD4⁺ und CD8⁺ T-Lymphozyten aus den Lymphknoten konnte, im Vergleich mit den entsprechenden Sham-Gruppen 48 und 96 Stunden nach Sepsisinduktion, eine tendenzielle Abnahme der CD4⁺ T-Helferzellen festgestellt werden. Wie die Abnahme der CD4⁺ Splenozyten ist diese Reduktion durch erhöhte Apoptoseraten und möglicherweise durch geringere Abgabe naiver 74 T-Zellen aus dem Thymus verursacht und widerspricht einer bei systemischen Infektion erwarteten klonalen Proliferation CD4+ T-Helferzellen [24]. Die geringe Abnahme CD8⁺ T-Lymphozyten 96 Stunden nach 22G-CLP ist kritisch zu bewerten, da der Großteil der Werte im Bereich der entsprechenden ShamGruppe liegt. Dennoch ist aufgrund einer nachgewiesenen Lymphozyten-Apoptose während der Sepsis im theoretischen Gesamtzusammenhang ein Abfall CD8+ TZellen im Lymphknoten plausibel. Wie bereits im Thymus beschrieben, könnte in zukünftigen Studien auch im Lymphknoten durch in-vivo Markierung von Lymphozyten mittels BrdU-Applikation der Zellumsatz sowie die Migration während der Sepsis untersucht werden. 4.4.4 Regulatorische T-Zellen Die Untersuchung regulatorischer T-Zellen (Tregs) aus Milz und Lymphknoten wurde durchgeführt, um zu überprüfen, ob im Rahmen der Sepsis Veränderungen des relativen Anteils von Tregs auftreten. Durch ihre Fähigkeit inhibierend auf das adaptive Immunsystem einzuwirken, könnte eine Zunahme von Tregs mitverantwortlich für die Immunparalyse in der Spätphase der Sepsis sein. 4.4.4.1 Milz Die Ergebnisse durchflusszytometrischer Messungen in der septischen Milz zeigten eine Abnahme von CD4⁺ T-Helferzellen (sh. 4.4.3.1). Dies war 36, 48 und 96 Stunden nach Sepsisinduktion zu erkennen. Bei der Untersuchung des Anteils der CD25⁺ T-Lymphozyten und des Anteils von CD4⁺CD25⁺ Tregs aus der Milz waren im Vergleich mit den Kontrollgruppen keine durch die Sepsis verursachten Veränderungen festzustellen. Errechnete man jedoch den Anteil der Tregs bezogen auf die CD4⁺ T-Zellpopulation, zeigte sich durch die Abnahme CD4⁺ Splenozyten eine indirekte Zunahme der Tregs im zeitlichen Verlauf der Sepsis. Zu den Zeitpunkten 48 und 96 Stunden war dies von der Sepsisschwere abhängig. Diese Ergebnisse stimmen mit Studien überein, die während der Sepsis ebenfalls eine relative Zunahme von Tregs in der Milz aufgrund einer Abnahme von CD4⁺ T-Zellen beschreiben [34, 44]. Die Ursache dafür könnte sein, dass Tregs, im Gegensatz zu CD4+ T-Helferzellen, aufgrund einer Resistenz gegenüber Apoptose nicht von dem durch Sepsis verstärkt induzierten Zelltod betroffen sind [47]. 75 Die indirekte Zunahme der Tregs kann zu der hypodynamen Spätphase der Sepsis beitragen. Tregs führen zur erhöhten Freisetzung von IL-10, welches von TH2-T-Zellen freigesetzt wird [34, 51], und können Immunantworten auf T-ZellAktivierungs-Ebene nach unten regulieren [33]. So spielt die Zunahme der Tregs eine Rolle in der Anergie und verminderten proliferativen Kapazität von Lymphozyten während der Sepsis. Die Bestimmung des Phänotyps CD4⁺CD25⁺ allein beweist nicht die Aktivität von Tregs. Vorangegangene Studien belegten aber, durch Bestimmung der Expression des Treg-spezifischen Forkhead-BoxProteins P3 (Foxp3), die Funktionsfähigkeit der Tregs septischer Mäuse nach CLP und darüber hinaus eine erhöhte immunsuppressive Wirkung im Vergleich zu Sham-behandelten Tieren [34, 51]. Die relative Zunahme von Tregs nach Sepsisinduktion und deren Fähigkeit, die Aktivierung und Proliferation von CD4⁺ T-Zellen zu inhibieren, zeigen die mögliche Beteiligung von Tregs an der Immunparalyse der Sepsis. Dies ist jedoch eher im Zusammenspiel mit anderen Faktoren, vor allen Dingen der Apoptose, zu sehen, da die Depletion von Tregs bei Mäusen keinen entscheidenden Einfluss auf das Überleben hat [34, 51]. Somit ist die immunsuppressive Wirkung der Tregs nur ein weiterer Faktor der zur Immundysfunktion in der Sepsis beiträgt. Im Widerspruch dazu steht eine erhöhte Überlebenswahrscheinlichkeit nach Transfer von Tregs vor bzw. kurz nach CLP, wobei eine mögliche Erklärung nicht aufgezeigt werden konnte. Bei Lymphozyten-defizienten Mäusen war dies nicht nachzuweisen, wodurch die Wechselwirkung zwischen Tregs und Lymphozyten sowie die Bedeutsamkeit funktionierender Lymphozyten in der Sepsis belegt wird [16]. Als kritisch sind die gewonnen Ergebnisse zum Verhalten von Tregs während der Sepsis insofern zu bewerten, als dass die Verwendung weiterer Marker, z.B. Foxp3 oder CD127, die Bestimmung auf Tregs allein beschränkt hätte. Durch Oberflächenmarkierung von CD4 und CD25 werden neben Tregs auch aktivierte CD4+ T-Zellen erfasst, da diese ebenfalls CD25 exprimieren können. Von weiterem Interesse für zukünftige Studien ist die Entwicklung von Tregs im Thymus während der Sepsis. Tregs entwickeln sich im Thymus aus CD4⁺ TZellen. Aufgrund erhöhter Apoptoseraten von CD4⁺CD8⁺ Thymozyten und einer Atrophie des Thymus wäre, trotz Resistenz gegenüber Apoptose, eine Abnahme von Tregs in der Spätphase der Sepsis zu erwarten. 76 4.4.4.2 Lymphknoten In Lymphknoten führte die Induktion einer polymikrobiellen Sepsis zu einer tendenziellen Abnahme CD4⁺ T-Zellen, insbesondere zum Zeitpunkt 48 Stunden. Beim Vergleich der Mediane mit den jeweiligen Sham-Gruppen 36, 48 und 96 Stunden nach Sepsisinduktion, fand sich eine tendenzielle Abnahme der TregFraktion. Dies ist jedoch als kritisch zu bewerten, da dies gegenüber unbehandelten Tieren nicht nachzuweisen war und der Großteil aller gemessenen Einzelwerte im Bereich von 5 bis 10 % lag und somit keine deutlichen Abweichungen festgestellt werden konnten. Im Gegensatz zur Milz konnten während der Sepsis keine Veränderungen der CD4⁺/CD4⁺CD25⁺ Ratio festgestellt werden. Dies kann dadurch bedingt sein, dass im Lymphknoten der Abfall CD4+ T-Lymphozyten während der Sepsis nicht so deutlich wie in der Milz auftritt. Da auch im Lymphknoten von einer Resistenz der Tregs gegenüber Apoptose auszugehen ist, ist aufgrund einer eher konstanten Konzentration CD4+ T-Lymphozyten keine Veränderung der CD4⁺/CD4⁺CD25⁺ Ratio zu beobachten. Über das Verhalten von Tregs in septischen Lymphknoten ist bislang wenig bekannt. Insbesondere in mesenterialen Lymphknoten wäre im Rahmen einer CLP-induzierten Sepsis Veränderungen der relativen Anteile zu erwarten. Bei Beurteilung der vorliegenden Ergebnisse scheinen Tregs in septischen Lymphknoten aber keine bedeutende Rolle zu spielen. 4.5 Schlussfolgerung und Ausblick Die in der vorliegenden Arbeit gewonnenen Ergebnisse bestätigen den quantitativen Einfluss der Sepsis auf T-Lymphozyten und deren mögliche Beteiligung an der Entstehung der Immundysfunktion in der Spätphase der Sepsis. Neben einer statistisch signifikanten Abnahme von CD4+CD8+ Thymozyten konnten Konzentrationsveränderungen von Thymozyten im CD4-CD8- doppelnegativen Stadium festgestellt werden. Die Ergebnisse zeigen eine Störung der TLymphopoese während der Sepsis. Während die Abnahme von CD4+CD8+ doppel-positiver Thymozyten am ehesten durch erhöhte Apoptoseraten bedingt ist, muss die Ursache für die nachgewiesenen Konzentrationsveränderungen im 77 doppel-negativen Stadium noch erforscht werden. Hierbei sollten vor allem die Apoptoseraten sowie eine eventuelle Blockade in den einzelnen Entwicklungsstadien doppel-negativer Thymozyten überprüft werden. Im Widerspruch zur gestörten T-Lymphopoese, wurde während der Sepsis eine Zunahme von CD4+ und CD8+ Thymozyten festgestellt. Bei Überprüfung bisher durchgeführten Studien zum Verhalten dieser Zellpopulationen während der Sepsis war keine klare Tendenz erkennbar. Zur genaueren Einschätzung der gewonnenen Daten sind hier weitere Experimente zur Untersuchung der Proliferation von CD4+ und CD8+ Thymozyten sowie der Konzentrationen von CD8+ T-Zellsubpopulationen in der Sepsis notwendig. Des Weiteren konnte in der vorliegenden Arbeit während der Sepsis eine Abnahme CD4+ und CD8+ Splenozyten sowie CD4+ T-Lymphozyten im Lymphknoten beobachtet werden. Dies ist nach aktuellem Wissensstand, wie bei doppel-positiven Thymozyten, durch erhöhte Apoptoseraten bedingt. Eine mögliche Beteiligung an der Abnahme peripherer CD4+ T-Lymphozyten hat die in der Arbeit nachgewiesene relative Zunahme von Tregs in der Milz, die inhibierend auf die Gesamtzusammenhang Proliferation betrachtet, ist CD4 + Splenozyten die Rolle der wirken. Tregs bei Im der Immundysfunktion während der Sepsis allerdings als gering einzuschätzen. Auch Homing, Rückkehr von Lymphozyten in sekundär lymphatische Organe nach Antigenkontakt zur spezifischen Proliferation, kann Veränderungen der Konzentration von T-Lymphozyten in lymphatischen Organen bewirken. Dies sollte in folgenden Studien mittels in vivo BrdU-Applikation untersucht werden. Durch die beschriebenen Veränderungen von T-Lymphozyten ist während der Sepsis eine adäquate adaptive Immunantwort auf eindringende Pathogene nicht gewährleistet. Für die Zukunft ist, neben einem genaueren Verständnis des Pathomechanismus der Sepsis, die Entwicklung von immunmodulierenden Therapeutika zur Senkung der Kosten und der Mortalität der Sepsis von großer Bedeutung. Therapeutika, die während der Sepsis eine regelrechte Entwicklung von T-Lymphozyten im Thymus ermöglichen und die Abnahme von Lymphozyten in der Peripherie verhindern, könnten die Entstehung der Immunparalyse in der Spätphase der Sepsis abschwächen. Dies würde wahrscheinlich dem Immunsystem ermöglichen, adäquat auf eine Infektion im Rahmen der Sepsis zu reagieren. 78 5 Zusammenfassung Mit ihrer hohen Mortalität stellt die Sepsis trotz intensiver Forschungsbemühungen immer noch ein schwerwiegendes Problem auf Intensivstationen dar. In der Pathophysiologie der Sepsis spielen sowohl eine überschießende Entzündungsreaktion als auch eine Immunparalyse eine wichtige Rolle. Deren kausale und zeitliche Zusammenhänge sind allerdings nicht vollständig geklärt. TLymphozyten kommt in der Pathogenese der Sepsis wohl eine große Bedeutung zu. Durch erhöhte Apoptoseraten, Anergie und einen Übergang von T-Helferzellen Typ 1 zu Typ 2 sind T-Lymphozyten an der Entstehung der Immunparalyse in der Spätphase der Sepsis entscheidend beteiligt. Einen möglichen Beitrag wird auch regulatorischen T-Zellen zugeschrieben, da sie in der Lage sind, eine adaptive Immunantwort zu unterdrücken. Ziel der vorliegenden Arbeit war, nach Sepsisinduktion mit dem Modell der zökalen Ligatur und Punktion, Veränderungen der Anteile von T-Zellpopulationen im zeitlichen Verlauf der Sepsis und in Abhängigkeit der Sepsisstärke zu untersuchen. Hierzu wurden zu Beginn der Arbeit im Rahmen einer Überlebenszeitanalyse die Überlebensraten weiblicher Wildtypmäuse nach zökaler Ligatur und Punktion unterschiedlicher Nadelstärken evaluiert. Dabei konnte ein von der Nadelstärke abhängiges Überleben festgestellt werden. Mit Zunahme des Außendurchmessers der Nadel sank die Überlebenswahrscheinlichkeit der Versuchstiere. Dies erlaubte, einen Rückschluss auf verschiedene Schweregrade der Sepsis zu ziehen, und diente zur Festlegung der Nadelstärken und der Zeitpunkte des Abtötens nach Sepsisinduktion für die nachfolgenden Untersuchungen. Zur durchflusszytometrischen Analyse von Thymozyten, T-Helferzellen, zytotoxischen T-Zellen und Tregs im zeitlichen Verlauf und in Abhängigkeit der Sepsisschwere wurde bei weiblichen Wildtypmäusen mittels zökaler Ligatur und Punktion verschiedener Nadelstärken eine Sepsis eingeleitet. Die Tötung der Tiere erfolgte in festgelegten zeitlichen Abständen zur Operation. Aus Thymus, Milz und Lymphknoten extrahierte Lymphozyten wurden mit Antikörpern markiert und anschließend durchflusszytometrisch analysiert. Aufgrund einer statistisch signifikanten Abnahme unreifer CD4⁺CD8⁺ (Cluster of Differentiation) Thymozyten und Veränderungen der Konzentrationen von 79 Thymozyten im CD4-CD8- doppel-negativen Stadium konnte im Thymus septischer Mäuse eine Störung der T-Lymphopoese während der Sepsis festgestellt werden. Diese verstärkte sich mit zunehmender Sepsisdauer und trägt wohl zur Entstehung der Immunparalyse in der Spätphase der Sepsis entscheidend bei. Im Gegensatz dazu nahm die Konzentration CD4⁺ T-Helferzellen und CD8⁺ zytotoxischer T-Zellen aus dem Thymus in Abhängigkeit von der Sepsisschwere während der Sepsis zu. Dies steht im Widerspruch zu in anderen Studien nachgewiesenen erhöhten Apoptoseraten dieser Zellpopulationen und einer gestörten Entwicklung von T-Lymphozyten während der Sepsis. Für periphere T-Lymphozyten wurde eine Abnahme CD4⁺ und CD8⁺ Splenozyten sowie CD4+ T-Lymphozyten in Lymphknoten nachgewiesen. Es ist bekannt, dass dies durch eine erhöhte Apoptose von septischen Lymphozyten verursacht wird. Möglicherweise ist die relative Zunahme regulatorischer T-Zellen (Tregs) in der Milz an der Abnahme peripherer CD4 + T-Lymphozyten beteiligt. Während der Sepsis führen Tregs zu Anergie und herabgesetzter Proliferationskapazität von Lymphozyten. Dies stellt einen weiteren möglichen Pathomechanismus in der Entwicklung einer Immunparalyse während der Sepsis dar, wobei im Gesamtzusammenhang, die Rolle der Tregs als gering einzuschätzen ist. Die Ergebnisse zeigen, dass die Sepsis einen entscheidenden Einfluss auf die Konzentration von T-Lymphozyten hat und bestätigen die Bedeutung eines funktionierenden adaptiven Immunsystems in der Sepsis. Die beschriebenen Veränderungen führen zu herabgesetzter Fähigkeit, auf eine Infektion angemessen zu reagieren, und stellen wohl einen zentralen Pathomechanismus in der Entwicklung der Immundysfunktion in der Spätphase der Sepsis dar. Von herausragender Bedeutung für die Zukunft ist es, die Immunantwort während der Sepsis im Detail weiter aufzuklären sowie immunmodulierende Therapeutika zu entwickeln, um Mortalität und Kosten der Sepsis zu senken. 80 6 Literaturverzeichnis 1. Angus D, Linde-Zwirble W, Lidicker J, Clermont G, Carcillo J, Pinsky M: Epidemiology of severe sepsis in the United States: Analysis of incidence, outcome, and associated costs of care. Crit Care Med 29: 1303-1310 (2001) 2. Ayala A, Herdon C, Lehman D, Ayala C, Chaudry I: Differential Induction of Apoptosis in Lymphoid Tissues During Sepsis: Variation in Onset, Frequency, and the Nature of the Mediators. Blood 87: 4261-4275 (1996) 3. Baker C, Chaudry I, Gains H, Baue A: Evaluation of factors affecting mortality rate after sepsis in a murine cecal ligation and puncture model. Surgery 94: 331-335 (1983) 4. Baseta J, Stutman O: TNF Regulates Thymocyte Production by Apoptosis and Proliferation of the Triple Negative (CD3CD4CD8) Subset. J Immunol 165: 5621-5630 (2000) 5. Baumann S, Krueger A, Kirchhoff S, Krammer P: Regulation of T cell Apoptosis during the Immune Response. Curr Mol Med 2: 257-272 (2002) 6. Brunkhorst F, Reinhart K: Sepsistherapie. Chirurg 79: 306-314 (2008) 7. Cohen J: The Immunopathogenesis of sepsis. Nature 420: 885-891 (2002) 8. Dellinger P, Levy M, Carlet J, Bion J, Parker M, Laeschke R, Reinhart K, Angus D, Brun-Buisson C, Beale R, Calandra T, Dhainaut J-F, Gerlach H, Hervey M, Marini J, Marshall J, Ranieri M, Ramsay G, Sevransky J, Thompson T, Townsend S, Vender J, Zimmermann J, Vinvent JL: Surviving Sepsis Campaign: International guidelines for management of severe sepsis and septic shock. Crit Care Med 36: 296-327 (2008) 9. Engel C, Brunkhorst F, Bone H-G, Brunkhorst R, Gerlach H, Grond S, Gruendling M, Huhle G, Jaschinski U, John S, Mayer K, Oppert M, Olthoff D, Quintel M, Ragaller M, Rossaint R, Stuber F, Weiler N, Welte T, Bogatsch H, Hartog C, Loeffler M, Reinhart K: Epidemiology of sepsis in Germany: results from a national prospective multicenter study. Intensive Care Med 33: 606-618 (2007) 10. Enoh V, Lin S, Etogo A, Lin C, Sherwood E: CD4⁺ T-cell depletion is not associated with alterations in survival, bacterial clearance, and inflammation after cecal ligation and puncture. Shock 29: 56-64 (2008) 81 11. Fisher C, Agosti J, Opal S, Lowry S, Balk R, Sadoff J, Abraham E, Schein R, Benjamin E: Treatment of septic shock with the tumor necrosis factor receptor: Fc fusion protein. N Engl J Med 334: 1697-1702 (1996) 12. Fisher C, Slotman G, Opal S, Pribble J, Bone R, Emmanuel G, Ng D, Bloedow D, Catalano M: Initial evaluation of human recombinant interleukin-1 receptor antagonist in the treatment of sepsis syndrome: a rondomized, open-label, placebo-controlled multicenter trial. Crit Care Med 22:12-21 (1994) 13. Gerard C: Complement C5s in the Sepsis Syndrome – Too Much of a Good Thing? N Engl J Med 348: 167-169 (2003) 14. Groesdonk H, Wagner F, Hoffarth B, Georgieff M, Senftleben U: Enhancement of NF-κB Activation in Lymphocytes Prevents T Cell Apoptosis and Improves Survival in Murine Sepsis. J Immunol 179: 80838089 (2007) 15. Gruver A, Sempowski G: Cytokines, leptin, and stress-induced thymic atrophy. J Leukoc Biol 84: 915-923 (2008) 16. Heuer J, Zhang T, Zhao J, Ding C, Cramer M, Justen K, Vonderfecht S, Na S: Adoptive Transfer of In Vitro-Stimulated CD4⁺CD25⁺ Regulatory T Cells Increases Bacterial Clearence and Improves Survival in Polymicrbial Sepsis. J Immunol 174: 7141-7146 (2005) 17. Hori S, Nomura T, Sakaguchi S: Control of regulatory T cell development by the transcription factor Foxp3. Science 299: 1057-1061 (2003) 18. Hotchkiss R, Chang K, Grayson M, Tinsley K, Dunne B, Davis C, Osborne D, Karl I: Adoptive transfer of apoptotic splenocytes worsens survival, whereas adoptive transfer of necrotic splenocytes improves survival in sepsis. Proc Natl Acad Sci U S A 100: 6724-6729 (2003) 19. Hotchkiss R, Karl I: The Pathophysiology and Treatment of Sepsis. N Engl J Med 348: 138-150 (2003) 20. Hotchkiss R, Swanson P, Cobb P, Jacobsen A, Buchman T, Karl I: Apoptosis in lymphoid and parenchymal cells during sepsis: Findings in normal and T- and B-cell-deficient mice. Crit Care Med 25: 1298-1307 (1997) 82 21. Hotchkiss R, Swanson P, Freeman B, Tinsley K, Cobb P, Matuschak G, Buchman T, Karl I: Apoptotic cell death in patients with sepsis, shock, and multiple organ dysfunction. Crit Care Med 27:1230-1251 (1999) 22. Hotchkiss R, Swanson P, Knudson M, Chang K, Cobb P, Osborne D, Zollner K, Buchman T, Korsmeyer S, Karl I: Overexpression of Bcl-2 in Transgenic Mice Decreases Apoptosis and Improves Survival in Sepsis. J Immunol 162: 4148-4156 (1999) 23. Hotchkiss R, Tinsley K, Swanson P, Chang K, Cobb P, Buchman T, Korsmeyer S, Karl I: Prevention of lymphocyte cell death in sepsis improves survival in mice. Proc Natl Acad Sci U S A 96: 14541-14546 (1999) 24. Hotchkiss R, Tinsley K, Swanson P, Schmieg R, Hui J, Chang K, Osborne D, Freeman B, Cobb P, Buchman T, Karl I:Sepsis-induced Apoptosis Causes Progressive Profound Depletion of B and CD4⁺ T-Lymphocytes in Humans. J Immunol 166: 6952-6963 (2001) 25. Kuo C, Leiden J: Transcriptional Regulation of T Lymphocyte Development and Function. Annu Rev Immunol 17: 149-187 (1999) 26. Martignoni A, Tschöp J, Goetzmann H, Choi L, Reid M, Johannigman J, Lentsch A, Caldwell C: CD4-expressing cells are early mediators of the innate immune system during sepsis. Shock 29: 591-597 (2008) 27. Matsutani T, Ohmori T, Ogata M, Soga H, Yoshioka T, Suzuki R, Itoh T: Alteration of T-cell receptor repertoires during thymic T-cell development. Scand J Immunol 64: 53-60 (2006) 28. Miller A, Rashid R, Elamin E: The „T“ in trauma: the Helper T-cell Response and the Role of Immunomodulation in Trauma and Burn Patients. J Trauma 63: 1407-1417 (2007) 29. Moerer O, Buchardi H: Kosten der Sepsis. Anaesthesist 55: 36-42 (2006) 30. Otero-Antón E, González-Quintela A, López-Soto A, López-Ben S, Llovo J, Pérez L: Cecal Ligation and puncture as a Model of Sepsis in the Rat: Influence of the Puncture Size on Mortality, Bacteremia, Endotoxemia and Tumor Necrosis Factor Alpha Levels. Eur Surg Res 33: 77-79 (2001) 31. Riedemann N, Guo R-F, Ward P: Novel strategies for the treatment of sepsis. Nature 9: 517-524 (2003) 32. Russell J: Management of Sepsis. N Engl J Med 355: 1699-1713 (2006) 83 33. Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M: Immunologic SelfTolerance Maintained by Activated T Cells Expressing IL-2 Receptor αChains (CD25). J Immunol 155: 1151-1164 (1995) 34. Scumpia P, Delano M, Kelly K, Malley K, Efron P, McAuliffe P, Brusko T, Ungaro R, Barker T, Wynn J, Atkinson M, Reeves W, Salzer M, Moldawer L: Increased Natural CD4⁺CD25⁺ Regularory T Cells and Their Suppressor Activity Do Not Contribute to Mortality in Murine Polymicrobial Sepsis. J Immunol 77: 7943-7949 (2006) 35. Scumpia P, Delano M, Kelly-Scumpia K, Weinstein J, Wynn J, Winfield R, Xia C, Chung C, Ayala A, Atkinson M, Reeves W, Clare-Salzer M, Moldawer L: Treatment with GITR agonistic antibody corrects adaptive immune dysfunction in sepsis. Blood 110: 3673-3681 (2007) 36. Seeger W, Grimminger F, Walmrath D: . In Werdan K, Schuster H-P, Müller-Werdan U (Hrsg) Sepsis und MODS. Springer, Heidelberg, 4. Auflage, S. 249-274 (2005) 37. Shelley O, Murphy T, Paterson H, Mannick J, Lederer J: Interaction between the innate and adaptive immune systems is required to survive sepsis and control inflammation after injury. Shock 20: 123-129 (2003) 38. Sherwood E, Enoh V, Murphey E, Lin C: Mice depleted of CD8⁺ T and NK cells are resistant to injury caused by cecal ligation and puncture. Lab Invest 84: 1655-1665 (2004) 39. Shimaoka M, Park E: Advances in understanding sepsis. Eur J Anaesthesiol 25: 146-153 (2008) 40. Singleton K, Wischmeyer P: Distance of Cecum Ligated Influences Mortality, Tumor Necrosis Factor-Alpha and Interleukin-6 Expression following Cecal Ligation and Puncture in the Rat. Eur Surg Res 35: 486-491 (2003) 41. Spellberg B, Edwards JE Jr.: Type1/Type2 immunity in infectious diseases. Clin Infect Dis 32: 76-102 (2001) 42. Turnbull I, Buchman T, Javadi P, Woolsey C, Hotchkiss R, Karl I, Coopersmith C: Age disproprtionately increases sepsis-induced apoptosis in the spleen and gut epithelium. Shock 22: 364-368 (2004) 84 43. Venet F, Chung C, Kherouf H, Geeraert A, Malcus C, Poitevin F, Bohé J, Lepape A, Ayala A, Monneret G: Increased circulating regulatory T cells (CD4(+)CD25(+)CD127(-)) contribute to lymphocyte anergy in septic shock patients. Intensive Care Med 35: 678-686 (2009) 44. Venet F, Chung C, Monneret G, Huang X, Horner B, Garber M, Ayala A: Regulatory T cell populations in sepsis and trauma. J Leukoc Biol 83: 523535 (2008) 45. Venet F, Pachot A, Debard A, Bohé J, Bienvenu J, Lepape A, Monneret G: Increased percentage of CD4+CD25+ regulatory T cells during septic chock is due to the decrease of CD4+CD25- lymphocytes. Crit Care Med 32: 2329-2331 (2004) 46. Verinaud L, de Souza Souto P, Brito V: Thymic Atrophy In Infectious Diseases. Braz J morphol Sci 21: 111-116 (2004) 47. Wang H, Bloom O, Zhang M, Vishnubhakat JM, Ombrellino M, Che J, Frazier A, Yang H, Ivanova S, Borovikova L, Manogue K-R, Faist E, Abraham E, Andersson U, Molina P-E, Abumrad N-N, Sama A, Tracey KJ: HMG-1 as a Late Mediator of Endotoxin Lethality in Mice. Science 285: 248-51 (1999) 48. Wang S, Huang K, Lin Y, Lei H: Sepsis-Induced Apoptosis of the Thymocytes in Mice. J Immunol 152: 5014-5021 (1994) 49. Weaver J, Rouse M, Steckelberg J, Badley A: Improved survival in experimental sepsis with an orally administered inhibitor of apoptosis. FASEB J 18: 1185-1191 (2004) 50. Wesche D, Lomas-Neira J, Perl M, Chung C, Ayala A: Leukocyte apoptosis and ist significance in sepsis and shock. J Leukoc Biol 78: 325-337 (2005) 51. Wisnoski N, Chung C, Chen Y, Huang X, Ayala Y: The contribution of CD4⁺CD25⁺ T-regulatory cells to immune suppression in sepsis. Shock 27: 251-257 (2007) 85 7 Danksagung Herrn Prof. Dr. U. Senftleben danke ich für die Bereitstellung des interessanten Themas und sein Interesse am Gelingen dieser Arbeit. Ein ganz besonderer Dank gilt Bettina Stahl für die tatkräftige Unterstützung und freundliche Zusammenarbeit sowie für viele wertvolle Ideen und Ratschläge. Ebenso gilt mein Dank Herrn Dr. Florian Wagner und Herrn Dr. Jörg Thomas für die sehr gute Betreuung und ihre große Hilfsbereitschaft. 86 8 Lebenslauf Persönliche Angaben Name Geburtsdatum / -ort Anschrift Richard Alexander Blothner 18. Juli 1981 / Wangen im Allgäu Arlacher Str. 4 88459 Tannheim Schulbildung 1991 -2000 Gymnasium Marianum Buxheim 2000 – 2001 Rettungshelfer Zivildienst im Krankentransport und Rettungsdienst, BRK Memmingen Berufsausbildung 2002 – 2003 Ausbildung zum staatlich Rettungsassistenten, geprüften Medizinisches Trainings- und Bildungszentrum Kempten Akademische Ausbildung 2003 – 2009 Studium der Humanmedizin, Universität Ulm August 2005 Abschluss der ersten ärztlichen Prüfung Seit 2007 Anfertigung der Dissertation zum Thema „Untersuchungen zur T-Lymphopoese während polymikrobieller muriner Sepsis“ August 2008 – Juli 2009 Praktisches Jahr an der Oberschwabenklinik in Ravensburg und an der Klinik Hirslanden in Zürich/Schweiz November 2009 Seit Oktober 2010 Abschluss der zweiten ärztlichen Prüfung Assistenzarzt der Anästhesie, Klinikum Westallgäu / Wangen 87 88