Schutzengel oder Beelzebub?
Werbung

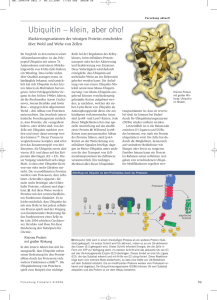
Special 498 Schutzengel oder Beelzebub? Die zwei Seiten des Tumorsuppressors p53 Wolfgang Deppert Heinrich-Pette-Institut für Experimentelle Virologie und Immunologie, Universität Hamburg Die Tumorinduktion ist ein komplexes Geschehen, an dem Mutationen in zwei verschiedenen Gen-Klassen beteiligt sind: Die Aktivierung von Proto-Onkogenen zu Onkogenen führt zu vermehrten Zellteilungen und damit zu unkontrolliertem Wachstum, die Inaktivierung von Tumorsuppressorgenen führt zum Ausschalten wichtiger Kontrollpunkte der Zellproliferation. Sowohl die Aktivierung von Onkogenen als auch die Inaktivierung von Tumorsuppressorgenen sind in der Regel Folge von Mutationen als Ergebnis vielfältiger endogener oder exogener DNA-Schädigungen. Dennoch gibt es einen wichtigen Unterschied: Da die Aktivierung eines Proto-Onkogens einen Funktionsgewinn darstellt, genügt schon die Mutation eines Allels der beiden vom Vater oder der Mutter stammenden Gene, um eine onkogene Wirkung, z.B. die Beschleunigung der Zellproliferation, zu erreichen. Anders bei den Tumorsuppressorgenen. Wird bei diesen ein Allel ausgeschaltet, so bleibt das zweite Allel aktiv. Die funktionelle Inakti- vierung eines Tumorsuppressorgens setzt somit voraus, dass beide Allele des Gens mutiert sind. Im Prozess der Tumorentwicklung spielt der Tumorsuppressor p53 eine zentrale Rolle. p53 hat vielfältige Aufgaben, die alle dazu dienen, die genetische Integrität einer Zelle bzw. des gesamten Organismus zu schützen. p53 - Schlüsselprotein der Tumorentstehung Die zentrale Rolle des Tumorsuppressor p53 in der Tumorentstehung wird schon dadurch belegt, dass das p53-Gen das am häufigsten mutierte Gen in menschlichen Tumoren darstellt (ca. 50 Prozent aller Tumoren). Der enge Zusammenhang zwischen dem Verlust der Tumorsuppressorfunktion von p53 und der Tumorentstehung wird bei einer sehr seltenen Erbkrankheit des Menschen, dem LiFraumeni Syndrom, deutlich. Die Betroffenen haben ein ererbtes mutiertes Allel des Abb. 1: Domänenstruktur von p53. Römische Ziffern kennzeichnen die fünf Regionen, die in allen Vertrebraten hoch konserviert sind. Phosphorylierungs- (P) und Azetylierungsstellen (Az) sind markiert. Die senkrechten Balken markieren häufige Punktmutationen, wobei die relative Länge der Balken ihre relative Häufigkeit widerspiegelt. Horizontale Balken unter dem Molekül kennzeichnen die im Text beschriebenen biochemischen Domänen von p53, darunter liegende Balken Interaktionsdomänen von p53 mit verschiedenen viralen und zellulären Proteinen. CSB: Cockayne Syndrom Protein B; MDM2: Mouse double minute protein 2; NES: nukleäres Exportsignal; NLS: nukleäres Importsignal; RP-A: Replikationsprotein A; SV40: Simian Virus 40; TAF: transkriptions-akvierender Faktor; TBP: TATA-Box bindendes Protein; TF: Transkriptionsfaktor; XPB: Xeroderma pigmentosum Protein B; XPD: Xeroderma pigmentosum Protein D. p53-Gens, das andere Allel ist intakt. Kommt es im Laufe des Lebens auch zum Verlust des bis dato noch intakten p53-Allels in einzelnen Körperzellen, wird die Tumorentstehung begünstigt. Li-Fraumeni Patienten entwickeln daher schon in relativ jungen Jahren Karzinome und ihre mittlere Lebenserwartung ist gegenüber der Normalbevölkerung deutlich reduziert. Multiple biochemische Funktionen p53 ist ein multifunktionelles Protein. Es besteht aus diversen funktionellen Domänen (Abb. 1). Die zentrale Domäne des Moleküls (AS 102–292) vermittelt die sequenz-spezifische Bindung von p53 an DNA, und hat außerdem intrinsische 3’-5’ ExonukleaseAktivität. Die Bedeutung der zentralen Domäne für die Tumorsuppressorfunktion von p53 wird auch daran deutlich, dass die Mehrzahl der Mutationen im p53-Gen im Bereich dieser Domäne liegt (siehe Abb. 1). Im extremen N-Terminus enthält p53 eine saure Transaktivator-Domäne (AS 1–42). Zusammen mit der zentralen Domäne wird p53 dadurch zu einem sequenz-spezifischen Transaktivator, der nach seiner Aktivierung die Transkription verschiedener p53-Zielgene einleiten kann. Der sauren Transaktivatordomäne folgt eine hydrophobe Domäne mit vielen Prolinresten (AS 63–97), die für die Auslösung der Apoptose durch p53 wichtig ist. Auch der C-terminale Bereich des p53 Moleküls besitzt mehrere funktionelle Domänen. Er enthält die Signale für den Transport von p53 in und aus dem Zellkern, eine Oligomerisierungsdomäne (AS 323–356), die dafür verantwortlich ist, dass p53 in Zellen hauptsächlich als Tetramer vorliegt. Der extreme C-Terminus (AS 363–393) besteht aus basischen Aminosäuren. Er kann Einzelstrang und Doppelstrang-DNA oder RNA binden und reguliert die Aktivitäten der zentralen Domäne. Über diese Domänen interagiert p53 mit einer Vielzahl von zellulären und viralen Proteinen (s. Abb. 1). Die physiologisch relevanteste Interaktion ist die zwischen p53 und dem MDM2 Protein. MDM2 bindet an die N-terminale Transkativator-Domäne von p53 und inhibiert so seine Transaktivatorfunktion. Weiterhin wird p53 durch MDM2 ubiquitiniert und in den proteasomalen Abbauweg eingeschleust. Da das mdm2-Gen durch p53 transkriptionell aktiviert wird, sind p53 und MDM2 über einen negativen Feedback-Loop miteinander verbunden. Genomische Integrität Die best-untersuchteste TumorsuppressorFunktion von p53 ist die eines sequenz-speBIOspektrum · Sonderausgabe · 10. Jahrgang Special 499 Abb. 2: Duale Funktion von p53 als Wächter des Genoms. Beschreibung siehe Text. zifischen Transaktivators von Genen, die entweder Kontrollpunkte des Zellzyklus aktivieren oder zur Apoptose führen. In Abwesenheit von zellulärem Stress wird p53 durch seine Interaktion mit MDM2 sehr schnell abgebaut und liegt nur in geringer Molekülzahl vor. Unterschiedliche zelluläre Stresssituationen, vor allem aber DNA-Schädigungen, aktivieren verschiedene Signalkaskaden, die zur Phosphorylierung und Azetylierung von p53 führen (s. Abb. 1). Dadurch wird die Interaktion zwischen p53 und MDM2 inhibiert, was zu einer Stabilisierung von p53 und damit zu seiner Anreicherung in der Zelle führt. p53 wirkt dann als sequenz-spezifischer Transaktivator, der die Expression von Genen induziert, die für Zellzyklus-Inhibitoren wie p21waf1 oder 14.3.30 kodieren und einen transienten Wachstumsarrest in verschiedenen Phasen des Zellzyklus bewirken. Weiterhin induziert p53 reparatur-assoziierte Gene, wie gadd45. Die p53-vermittelte Attenuation des Zellzyklus erlaubt die Reparatur von DNASchäden, bevor diese während der nächsten Runde der DNA-Replikation als Mutationen fixiert werden. p53 induziert weiterhin eine Vielzahl von Apoptose-induzierenden Genen, darunter bax, fas-R, PUMA, sowie einige redox-assoziierte PIG Gene. Das Apoptose-auslösende Gen konnte aber noch nicht identifiziert werden. Die p53-vermittelte Apoptose wird weiterhin dadurch kompliziert, dass p53 Apoptose auch über nichttranskriptionelle Mechanismen direkt aktivieren kann. Die Lösung der Frage, welche Parameter die Selektivität der p53-Antwort auf DNA-Schäden determinieren, und wie diese Antwort in Richtung auf Apoptose beeinflusst werden kann, ist gegenwärtig eines der wichtigsten Themen der p53-Forschung. p53 interagiert mit einer Reihe von Reparatur-assoziierten Proteinen (s. Abb.1) und ist neben den oben beschriebenen Aktivitäten auch direkt an Reparaturprozessen und der Überprüfung ihrer exakten Durchführung beteiligt. In diesen Prozessen spielt auch die intrinsische 3’-5’ Exonukleaseaktivität von p53 eine wichtige Rolle. Bislang konnte gezeigt werden, dass die p53-Exonukleaseaktivität die Genauigkeit der homologen DNA-Rekombination kontrolliert. Custom Monoclonal Antibodies in Just 8 Weeks! Tel: +49 89 899 27 234 Antibodies by Design A Division of MorphoSys Fax: +49 89 899 27 5321 Email: [email protected] Special 500 Abb. 3: p53 in der Vielschritt-Karzinogenese. Beschreibung siehe Text. Ob die p53-Exonuklease auch noch an anderen DNA Reparatur- und Synthesevorgängen beteiligt ist, z.B. als „Korrektor“ (proof reader) der proof reader defizienten DNA-Polymerasen α und β, ist derzeit noch in der Diskussion. Abbildung 2 fasst die vielfältigen Aufgaben von p53 in der Erhaltung der genomischen Integrität zusammen. Nach dem Modell der „dualen Funktion“ übernimmt p53 eine Basisfunktion in der Kontrolle der genomischen Integrität und schützt durch seine direkte Beteiligung an Reparaturprozessen vor Mutationen aus endogen entstehenden DNA-Schäden. Die Aktivierung von p53 als Folge genotoxischen Stresses führt zu seiner Funktion als übergeordnetes Kontroll- und Steuerungselement der zellulären Schadensantwort. Basisfunktion und SignalIntegratorfunktion von p53 überlappen sich, da auch aktiviertes p53 direkt an Reparaturvorgängen beteiligt sein kann. Vom Schutzengel zum Beelzebub Mutiertes p53 weist ein für einen Tumorsuppressor ungewöhnliches Mutationsspektrum auf: Während alle anderen bislang bekannten Tumorsuppressoren durch Deletion, Trunkierung oder Abschalten ihres Gens inaktiviert werden, stehen beim p53Gen Punktmutationen im Vordergrund, wodurch eine Aminosäure im p53 Molekül ausgetauscht wird (s. Abb. 1). Bei der Tumorentstehung wird offensichtlich auf diese Mutationen selektioniert, d.h. Tumore, die ein solchermaßen mutiertes p53 exprimieren, müssen einen selektiven Wachstumsvorteil aufweisen. Die onkogene(n) Funktion(en) von p53 sind auf molekularer Basis noch wenig charakterisiert, phänotypisch zeigen sie sich in erhöhter Tumorigenität und einem erhöhten Metastasierunspotenzial des Tumors. Weiterhin zeigen Tumore mit mutiertem p53 häufig Resistenz gegen Strahlenund Chemotherapie. Das ist zum einen Folge des Verlusts von Wildtyp p53 und damit dessen Fähigkeit, nach DNA-Schädigung Apoptose zu induzieren. Zum anderen aber liegt es auch an der von mutiertem p53 induzierten Resistenz dieser Tumorzellen gegen p53-unabhängige Apoptosewege. präkanzeröse Zelle in ihrer Evolution zur Tumorzelle. Dieser Schritt bedingt immer die Ausschaltung von p53, entweder durch Mutation, wie es in ca. 50% aller Tumoren der Fall ist, oder durch Inaktivierung bzw. Umgehung von p53-Signalwegen. Ist dieser Schritt erfolgt, beginnt nach der Phase der Tumorinitiation die Phase der Tumorprogression, die zum Primärtumor bis hin zum metastasierenden Tumor führen kann. Die Tumorprogression kann auch in Gegenwart von Wildtyp p53 erfolgen (allerdings müssen dann einige p53-Signalwege ausgeschaltet sein!), wird aber in vielen Tumoren durch die Gegenwart von mutiertem p53 begünstigt, mit den oben beschriebenen negativen Folgen für den Tumorpatienten. Multiple Funktionen von p53 in der Vielschritt-Karzinogenese Literatur Abbildung 3 fasst die Rolle von p53 in der Tumorentstehung zusammen. Wildtyp p53 schützt nach DNA-Schädigungen vor Mutationen, indem es die Reparatur der geschädigten Zellen einleitet, oder diese, bei zu starker Schädigung, durch Apoptose eliminiert. Dies ist – glücklicherweise – der häufigste Fall. Wird dennoch durch eine Mutation als Folge einer unvollständigen Reparatur ein Onkogen aktiviert, führt dies zu einer DNA-schadens-unabhängigen Aktivierung von p53, und die präkanzeröse Zelle kann ebenfalls noch durch Apoptose oder terminalen Wachstumsarrest (Seneszenz) entfernt werden. Die Überwindung dieses Blocks ist die schwierigste Hürde für eine Oncogene Reviewheft „p53“. Oncogene 18, no.53 (1999) Human Mutation Sonderheft: „Focus on p53 and cancer“Human mutation 21, no.3 (2003) Korrespondenzadresse: Prof. Dr. Wolfgang Deppert Heinrich-Pette-Institut für Experimentelle Virologie und Immunologie Universität Hamburg Martinistr. 52 D-20251 Hamburg Tel.: 040-48051 261 Fax: 040-48051 117 [email protected] BIOspektrum · Sonderausgabe · 10. Jahrgang