Bakterien Genetik der Bakterien
Werbung

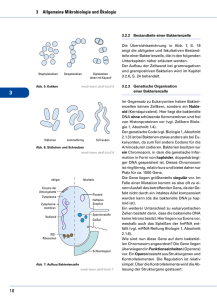
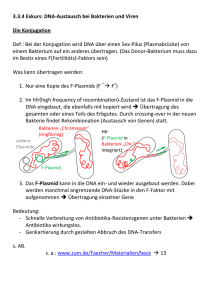
Bakterien Genetik der Bakterien Bakterien sind asexuelle haploide Organisma, und es war fraglich, ob der Austausch der genetischen Information kommt überhaupt bei Bakterien vor. J. Lederberg und E.L. Tatum führte die erste „Kreuzung” der Bakterien durch, welche zur Bildung Rekombinanten führte. Sie nahmen die auxotrophen Stämme „A” und „B” von Escherichia coli, die gemeinsam in Flüssigkultur gewachsen wurden. Auxotrophe Bakterien tragen Genmutationen, die ihr Wachstum verhindern, und wachsen nur in dem Fall, wenn die Stoffe, die die Bakterien selbst produzieren können, ins Kulturmedium (Minimalmedium) zugegeben werden. Das Minimalmedium besteht aus Wasser, Kohlenstoff- und Stickstoffquellen (Glycerin, Ammonia) und anorganische Sälze. Die prototrophen (oder wildtyp) Bakterien wachsen auf das Minimalmedium, wie die Rekombinanten aus „A”x”B” „Kreuzung” Das Auftreten der prototrophen Bakterien kann nur durch eine Veränderung ihrer genetischen Information erklärt werden, die als Konjugation genannt wurde. Die Richtung der Konjugation wurde durch Streptomycin-resistent Stämme bestimmt . Die Donorzelle (F+) DNA abgibt, die Emphängerzelle die DNA Aufnimmt. Die F+-Zellen von E. coli besitzen zwei genetische Elemente, das Chromosom und das F (Fertilitäts)-Plasmid/Faktor. Ein direkter Kontakt mit der F- Zelle über der Konjugationsbrücke, oder Sex-Pili (Protein-Rohre, kodiert das F+Plasmid) ist für den Gentransfer unbedingt benötigt. Die F+ Donorzelle überträgt den Fertilitätsfaktor (F-Plasmid) in die F- Emphängerzelle als Einzelsträngige DNA, und der Emphänger synthetisiert den zweiten Strang. Während der Konjugation wandelt sich die F- Stamm in F+. Der Transfer beginnt immer an der oriT (transfer) Stelle des F-Plasmids. Dieser Mechanismus erklärt es nicht, wie Gene des Chromosoms konjugiert/mobilisiert werden können. Das F-Plasmid kann ins Chromosom integrieren, damit eine sog. Hfr-Zelle (high frequency of recombination), mit einer 10-3 Wahrscheinlichkeit entsteht. Während der Paarung/Konjugation überträgt die Hfr-Zelle ihre einzelsträngige DNA in die F- -Zelle so, dass die chromosomale DNA auch mobilisiert wird. Die Übertragung beginnt, genau wie bei der F+-Zelle, bei der oriT-Sequenz des integrierten F-Faktors. Die Übertragung des ganzen Chromosoms dauert ~100 Minuten, aber wird spontan unterbrochen als die Konjugationsbrücke brechen. Nun die Emphängerzelle enthält ihr vollkommenes, ringförmiges Chromosom, und zusätzlich einen mehr oder weniger lange lineare DNA-Abschnitt aus der Donorzelle. Weshalb, ist die Zelle Merozygot, nur teilweise Diploid (Abb. 8). Die übertragene DNA kann jetzt mit der DNA der Emphängerzelle rekombinieren. Ein einzellcrossover zwischen der linearen und ringförmigen DNA ergibt ein lineares Chromosom, welches nicht lebensfähig ist. Nur eine Doppelrekombination sorgt für eine lebensfähige Zelle, und einer der Rekombinationspartner, das Ringförmige Chromosom überlebt und trägt teilweise die Gene der Donorzelle. Da die Konjugation ein in der Zeit abgelaufener Vorgang ist, und die Konjugation bei einer bestimmten Hfr-Zelle immer bei der oriT-Stelle iniziiert wird, werden die Gene der Donorzelle immer in der selben Reihenfolge übertragen. Je weiter ein Gen von oriT entfernt liegt, desto später gelangt es in die F- Zelle. Während der unterbrochenen Kartierung messen wir die Zeit, benötigt für die Übertragung eines Gens, als die Einheit der genetischen Karte. Mithilfe Streptomycin-resistent Emphängerzellen und sensitiv Donorzellen töten wir sie in der Anwesenheit des Antibiotikums, nur die resistenten und rekombinanten Emphängerzellen überleben. Die Unterbrochene Konjugation erlaubt nicht die Kartierung eng gekoppelter Gene. Rekombinationskartierung aber ermöglicht es, eine detaillierte Karte zu erzeugen. siehe noch Bakterien im Buch! Grundanforderung 12. Vorlesung 1