Der Farbschlüssel für Alpakas - ein Schna
Werbung

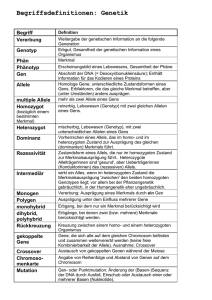
genetik Der Farbschlüssel für Alpakas stellt, die zu den beobachteten Phänotypen passen. Diese Artikel haben mich veranlasst, mein eigenes Material nochmals zu überprüfen und die Eigenschaften der in der Alpakafaser gefundener Pigmente zu untersuchen. Im Mai 2002 erhielt ich die Erlaubnis von Professor Peter Coloe, Leiter des Ministeriums für Biotechnologie und Umweltbiologie an der RMIT Universität in Melbourne, das Fluoreszenz-Phasenkontrastmikroskop des Ministeriums für Studien an den Pigmenten zu nutzen. Seitdem habe ich rund 500 Fotos von den Fasern von etwa 20 verschiedenen Tierarten gemacht, habe mich aber vorwiegend auf Alpakas konzentriert Foto: Heinz Kuhne-Pfaff Diese beiden Vorträge sind ein Teil meines Seminars mit dem Titel “Der Farbschlüssel für Alpakas”, in denen ich die wichtigsten Punkte der beiden neuesten Theorien mit den Pigmentsstudien, den Herdbuchdaten und den Abstammungsdaten zusammengeführt habe in dem Versuch, zu einer umfassenderen Sicht der Farbvererbung bei Alpakas zu gelangen. Die Farbe ist einer der wichtigsten Vorteile, den die Alpakas gegenüber anderen ebenfalls vliestragenden Tieren haben. Über die Vererbung von Farbe bei den Alpakas gibt es zahlreiche Theorien. Drei der wichtigsten aus den frühen Jahren sind die von Dr. George Davis aus Neuseeland, wonach braun dominant gegenüber schwarz ist und Farbe über weiß dominiert. Ron Wall und Bill Cole waren der Überzeugung, daß alle Farbgene miteinander verbunden sind. Dr. Julie Koenig aus den USA war der Meinung, dass schwarz dominant gegenüber braun ist, dass ein dominantes weißes Gen die Erklärung für die meisten weißen Alpakas ist und dass die meisten Grautöne auf rötlichgrauen Genen beruhen. Meine eigene Arbeit auf diesem Gebiet während der letzten vier bis fünf Jahre mündet, kurz gesagt, in der Aufstellung von Tabellen 40 über die Ergebnisse bei den Nachkommen aus Paarungen, die im A.A.A.-Herdbuch festgehalten sind und die ich im folgenden als „Herdbuch-Daten“ bezeichne. Allerdings haben viele dieser Daten zu keiner der Theorien (einschließlich meiner) gepasst, und einige der Ergebnisse passten auch nicht zu anerkannten Fakten der Genetik. Anfang 2002 erhielt ich Kopien von zwei anderen Theorien. Die erste war eine These von Ken Hart, einem Genetiker vom Landwirtschaftsministerium von Westaustralien, der auf dem Gebiet der Schafzucht arbeitet, die sich damit befasste, einen Beweis für ein dominantes weißes Gen bei Alpakas zu finden. Der zweite Artikel war von Dr. Phillip Sponenberg, Professor der Pathologie und Genetik am Virginia-Maryland Regional College der Veterinärmedizin, USA, in dem er über die verschiedenen Farbreihen sprach und eine Anzahl von Genotypen zur Diskussion GENETIK: Für jede Diskussion über Farbvererbung brauchen wir eine kurze Liste der wichtigsten Definitionen für einen grundlegenden Wortschatz der Genetik, damit wir die genetischen Wirkungsweisen, die hier am Werk sind, verstehen können. Die Zelle ist der Baustein aller lebenden Wesen, und der Zellkern ist das “Gehirn” oder Kontrollzentrum jeder lebenden Zelle. Er enthält die DNA, auch Desoxyribonukleinsäure genannt, eine Kette von chemischen Elementen, die die genetischen Informationen tragen, die für die Organisation der Aktivitäten der Zelle erforderlich sind. Wenn die Zelle mit ihrem Teilungsprozess beginnt, trennt sich die Masse der DNA in zwei Einzelstränge von DNA-Ketten, den Chromosomen. Die Gene sind die einzelnen Einheiten der genetischen Information, die in einem besonderen Locus oder Ort auf den Chromsomen gespeichert werden. Ein Genpaar führt dazu, dass ein Merkmal zum Ausdruck kommt oder eben nicht. Jedes Genpaar verfügt über einen “Ein/Aus”- genetik - ein Schnappschuss Mechanismus, fast wie ein Lichtschalter, aber die Mechanismen von mehreren Genpaaren können addiert werden, um eine abgestufte Wirkung zu erzeugen, wie dies bei einem „Dimmer“-Schalter geschieht. Die verschiedenen Formen eines Genpaares nennt man Allele. An dem einen Ort auf dem Chromosomenpaar können sich nur zwei Allele eines Gens befinden. Wenn die zwei Allele des Genpaars identisch sind, nennt man sie homozygot, sind sie verschieden, nennt man sie heterozygot. Wenn ein Allel eines Genpaars immer voll das Merkmal zum Ausdruck bringt, dann bezeichnet man dieses Allel als dominant gegenüber allen Allelen in dieser Kette. Das Genpaar kann homozygot oder heterozygot sein. Wenn das Genpaar für das dominante Allel homozygot ist, ist normalerweise nur eines der beiden Allele aktiv. Ein Allel wird als rezessiv bezeichnet, wenn zwei gleiche Allele erforderlich sind, damit dieses bestimmte Merkmal zum Ausdruck kommt, das Genpaar ist in diesem Fall immer homozygot. Mehr als zwei Allele für ein gegebenes Genpaar bezeichnet man als Reihe. Man beachte, dass sich der Ausdruck „dominant” und “rezessiv” nur auf Allele eines bestimmten Genpaars bezieht, und nicht auf Gene aus anderen Reihen. Eine Reihe von Genen kann als epistatisch bezeichnet werden, d.h. die Gene stehen über einer anderen Reihe von Genen. Zwei Allele eines Genpaars nennt man antagonistisch, d.h. sie haben eine genau entgegensetzte Wirkung aufeinander. Dies kann auch für unterschiedliche Genpaare und unterschiedliche Reihen zutreffen. Manchmal kommt das dominante Allel nur teilweise zum Ausdruck, wodurch das rezessive Allel ebenfalls zum Vorschein kommt, sodass das Tier als eine Zwischenform erscheint. Dies nennt man unvollständige Dominanz. Der Genotyp ist die gesamte Ausstattung mit Genpaaren auf all den Chromosomenpaaren innerhalb des Kerns der lebenden Zelle. Alle Zellen im Körper des Alpakas enthalten genau denselben Genotyp (mit Ausnahme der Spermien und der Eier). Der Genotyp wird zum Zeitpunkt der Befruchtung eines Eis durch ein Spermium bestimmt und kann nicht verändert werden. (Na ja, jedenfalls nicht von der Natur!) Der Phänotyp ist der körperliche Ausdruck des Genotyps des Alpakas. Wir können bestimmte Merkmale des Alpakas sehen oder messen. Der Phänotyp kann durch äußere Einflüsse wie Umwelt, Änderungen in der Nahrung, Stress, Krankheit, Medikamente etc. verändert werden. Nicht alle der in dem Genotyp vorhandenen Gene kommen zwingend im Phänotyp zum Ausdruck. Offensichtlich können ähnliche Phänotypen von ganz unterschiedlichen Genotypen hervorgerufen werden; dies kann insbesondere für ähnliche Phänotypen gelten, die bei verschiedenen Tierarten auftreten. Während der Produktion von Sperma und Phänotypen und Genotypen Eizellen halbiert sich die Anzahl der Chromosomen in jedem Kern. Bei der Befruchtung werden die einzelnen Chromosomen vom Sperma und dem Ei wieder zusammengeführt und ergeben die passenden Paare. Der neue Embryo hat exakt die Hälfte seines genetischen Materials von seiner Mutter und eine Hälfte von seinem Vater. Von nun an tragen alle seine Zellen einen eigenen und einzigartigen Genotyp. Zu beachten ist, dass auch jede Befruchtung einzigartig ist. Sie ist völlig unabhängig von der vorangehenden und der darauffolgenden Befruchtung. Zusammenfassend kann man sagen, dass das Alpaka nur zwei Allele eines beliebigen Genpaars erben kann, eines von seinem Vater und eines von seiner Mutter. Die Allele können homozygot oder heterozygot sein. Der Genotyp oder die Gesamtheit der Gene ist festgelegt. Der Phänotyp ist je nach dem Genotyp des Tieres und der gegenseitigen Beeinflussung dieses Genotyps und der Umwelt veränderbar. In der Genetik arbeitet man mit vielen Annahmen, wobei sich durch die von Elizabeth Paul Beobachtung der Zuchtergebnisse herausstellt, ob diese Annahmen zutreffen oder nicht. PIGMENT UND FASERSTRUKTUR Die Farbtöne in den Haaren, der Haut und den Augen von Tieren werden durch Melanin-Körnchen hervorgerufen. Dabei handelt es sich um ein großes, stabiles Molekül auf der Basis von Tyrosin, das verschiedene Wellenlängen von Licht absorbiert. Die speziellen Melanin-produzierenden Zellen nennt man Melanozyten. Sie entstammen dem Bereich des neuralen Kamms im Embryo. Nach ihrer Entstehung wandern sie über die Oberfläche des Embryos, bis sie die epidermische Schicht der Haut oder die Basen der Haarfollikel erreichen. Die Körnchen werden in die Epidermis oder in die Zellen des Haarschaftes abgegeben, wenn dieser aus dem Follikel herauswächst. Eine Haarfaser besteht aus einem inneren Kern aus Medullazellen, einer weiteren Schicht aus Cortexzellen sowie den Kutikulazellen oder Schuppen. Die verschiedenen Schritte auf biochemischer Ebene, die zur Melaninproduktion gehören, sind genetisch gesteuert. Es gibt zwei Pigmentarten, nämlich schwarz und gelb. Das schwarze Eumelanin, oder sein Abkömmling, das rezessive Braun (meist schwärzlich oder ein dunkles Schokobraun) erscheint als ovale Granula mit einer Größe von rund 0,7 x 0,3 Mikron. (1 Mikron = 1 millionster Teil eines Meters). Phaeomelanin oder Gelbpigment tritt in Form von runden, gelben Granula auf und ist der Grund für die meisten rötlichen und gelblichen Haartöne. (Searle, A.) Ein Melanozyt kann sowohl gelbes als auch schwarzes Pigment erzeugen, und manchmal auch beide in einer einzigen Haarfaser. Der endgültige Farbton ist abhängig von der Anzahl der Granula, ihrer Größe und Form, ihrer Anordnung und ihres Typs, wobei alle diese Punkte genetisch gesteuert werden. Eine Reihe von Veränderungen in der Genetik oder der Umwelt, wie etwa eine Verzögerung bei der Wanderung der Melanozyten oder Änderungen der Temperatur oder der Tageslänge können Auswirkungen auf jedes Stadium des Produktionsprozesses haben. Wenn sowohl die Kortex- als auch die Medul- 41 genetik lazellen viele Pigmentgranula in einer regelmäßigen Anordnung aufweisen, ist die Faser eindeutig intensiv gefärbt. Verringert sich die Menge der Pigmente oder ändert sich die Anordnung der Granula oder der Zellen, dann wird der Farbton der Faser sichtlich heller. In den Fasern der Haut von Falben und Grauschimmeln zum Beispiel sind die Pigmentgranula eindeutig auf einer Seite der Faser konzentriert. Auch die Probe von dem dunkel-cremefarbenen Pony weist Unterbrechungen in der Durchgängigkeit der Medullazellen auf, so dass nur das Pigment in den Cortexzellen den Fasern an diesen Stellen Farbe verleiht. MODELLE DER VERERBUNG DER VLIESFARBE Der wichtigste Punkt, den sowohl Dr. Sponenberg als auch Ken Harft in ihren Aufsätzen festgehalten haben, besagte, dass farbliche MUSTER bei der Festlegung des Genotyps ausschlaggebend sind. Zu diesen Mustern kann die Farbe auf nichtbewollten Flächen wie Kopf und Beine gehören, die bislang bei den Farbmusterkarten weitgehend ignoriert wurden. Es folgt eine kurze Zusammenfassung der Aufsätze; alle Fehler bei der Auslegung stammen von mir. Beide Autoren vertraten die Ansicht, dass es zwei antagonistische Gen-Hauptreihen gibt, die eine Erklärung für die meisten der beobachteten Phänotypen liefern könnten. Es sind dies die Agouti-Reihe und die Extension-Reihe, die in einem komplexen Verhältnis zueinander stehen. Der Extension-Locus steht für eine gleichmäßige Pigmentverteilung über das gesamte Fell. Die dominante Ausprägung ist vollständig schwarz, gilt aber als sehr selten. Die rezessive Ausprägung ist vollständig hell und wird “ee” oder “rezessives Rot” genannt. Ein kastanienbraunes Pferd ist ein Beispiel für ein rezessiv-rotes Tier und trägt auf dem ganzen Körper mehr oder weniger einen roten Farbton (wenn man die üblichen weißen Abzeichen außer Acht lässt), da das einzige im Fell produzierte Pigment das Phaeomelanin ist. Außerdem haben sie helle Hufe, keine dunklen. Die Extension-Allele sind alle epistatisch gegenüber dem Agouti, aber die Expression aller Agouti-Allele beruht auf dem Vorhandensein des“+” oder Wild-Typ-Allels am Erweiterungs-Locus. (Anm. d. Red.: epistatisch bedeutet Unterdrückung der Merkmalsausbil- 42 dung eines Gens durch ein anderes Genom) Die Agouti-Reihe wird mit dem Buchstaben A gekennzeichnet, bei dem das Tier ein Muster aus zwei Farben im Fell zeigt. Das dominanteste Agouti-Allel wird manchmal als dominantes Weiß oder Weiß/Braun bezeichnet und ist der hellste Ton für diese Art. Das rezessivste Allel “aa” ermöglicht ebenfalls die vollständige Expression von Eumelanin, aber nur in der Gegenwart von“+” oder dem WildTyp aus der Extension-Reihe. Dazwischen liegen Allele, die Muster mit unterschiedlichen Anteilen von Hell- und Dunkelpigmenten hervorbringen. Die helleren Muster, also diejenigen mit mehr Rot-Anteilen, dominieren über die Muster mit mehr Anteilen an dunkleren Farbtönen. Agouti selbst ist eine Farbverteilung, die als Tarnfarbe wirkt und die man bei kleineren Tieren häufiger sieht. Das südamerikanische Agouti, nach dem die Reihe benannt ist, ist ein kleines Nagetier mit dem typischen “graumelierten” Fell, bei dem sich in einem Haar sowohl Gelb- als auch Schwarzpigment findet. Die Deckhaare jedoch sind oft gleichmäßig schwarz. Bei den größeren Weidetieren tritt Agouti in symmetrischen, ziemlich klar abgegrenzten Mustern von roten und schwarzen Stellen im Fell auf. Hauspferde und –rinder mit einem derartigen Muster nennt man Rotbraune (engl.: bay). Sie haben im allgemeinen einen rotbraunen Körper und schwarze Abzeichen, d.h. eine beliebige Kombination aus Ohren/ Gesicht/Mähne/Schweif oder untere Extremitäten mit schwarzen Füßen. Der Rotton reicht von einem sehr hellen Rot über Gelb bis zum dunkelsten Mahagonirot, bei dem die schwarzen Abzeichen manchmal nicht zu erkennen sind. Der dritte Locus ist das Schwarz, wobei „B“Schwarz dominant gegenüber dem rezessiven “bb”-Braun ist, das man oft braun oder auch schokoladenbraun nennt. Bei keinem Tier können im Fell sowohl braune als auch schwarze Fasern vorkommen, da diese beiden Farbtöne zum selben Genpaar gehören. Das genetische Braun kommt bei kleineren Tieren wie Hunden und Katzen häufiger vor. Es tritt auch bei Ziegen auf, aber nach der Literatur gelten Pferde als homozygot für schwarz. Beide Autoren nahmen an, dass die Situation bei den Alpakas dieselbe sei. (Man beachte, dass der Ausdruck “braun” bei Pferden eine allgemeine Beschreibung der Farbe ist, die man an ihnen wahrnimmt und dass braune Hunde in der populärwissenschaftlichen Literatur zur Zucht als „rot“ bezeichnet werden. Vom Blickpunkt der Genetik aus ist keine dieser Beschreibungen korrekt.) Man schreibt die Reihen in der Form “A -/E -/B-”, was für Agouti / Extension / schwarz steht. Jedes Alpaka hat Allele aus allen drei Reihen in seinem Genotyp, die für die drei Grundmuster mit dem rezessiven Rot (kastanienbraun), Agouti oder Rotbraun und Schwarz verantwortlich sind. Meine Auslegung der Genotypen ist die folgende: --/ee/-- für rezessives Rot; A-/+-/B- für Rotbraun sowie aa/+-/B- für Schwarz. Für die praktische Zuchtarbeit bedeutet dies, dass zweimal rezessives Rot rein vererbt wird, da selbst die Gene der rezessiven Extension-Reihe dominant gegenüber der Agouti-Reihe sind. Zweimal Schwarz ergibt Schwarz und auch ein gelegentliches rezessives Rot, falls beide zufällig auch ein “e”-Allel tragen. Sie befinden sich im unteren Bereich der Agouti-Reihe, deshalb sind sie wohl kaum in der Lage, die helleren Braun-Phänotypen hervorzubringen. Zwei Rotbraune ergeben überwiegend Rotbraun, ein paar Mal rezessives Rot (falls beide ein “e”-Allel tragen) und einige Male Schwarz (falls beide ein „a“-Allel tragen). PIGMENTSTUDIE: Ursprünglich habe ich meine Pigmentstudie begonnen um herauszufinden, ob das Pigment in einer Palette von „braunen“ Alpakafasern tatsächlich braun oder gelb ist. Ferner habe ich Faserproben vom Kamel, dem Vikunja und dem Guanako untersucht. Bei allen Proben, angefangen vom hellen Fawn bis zum dunkelsten Mahagonirot, fand ich Phaeomelanin. Unter dem Mikroskop hatten die Phaeomelanin-Granula einen reinen Gelbton. Bei den meisten Arten waren sie recht groß und traten in vielen Fällen wie Stränge von goldenen Perlen auf. Unter fluoreszierendem Licht leuchteten sie in einem wunderschönen Blau. DATENBANKSTUDIE: Mittlerweile habe ich die Ergebnisse von mehr als 40.000 im A.A.A.-Herdbuch festgehaltenen Paarungen zusammengetragen. Tabelle 1 zeigt die Ergebnisse aller Paa- genetik rungen zwischen nicht-grauen Elterntieren aus Herdbuch 2-11. (Die Ergebnisse von Paarungen mit mindestens einem grauen Elterntier sind in Tabelle 2 erfasst; diese Ergebnisse werden im zweiten Teil behandelt). Nach Auswertung der obigen Informationen habe ich alle “braun-“Paarungen (engl.: brown) in meinen Daten in Paarungen von „Rotbraunen“ (engl.: „bay“) umbenannt. Diese einfache Änderung ergab für die Ergebnisse der Paarungen plötzlich viel mehr Sinn. Insbesondere die Paarungsergebnisse für „braun x braun“, die 67 % „braune“, 11 % schwarz und 17 % fawn-farbige ergaben. Zwei genetisch braune (brown) Tiere können nach einer Paarung nur genetisch braune Nachkommen haben oder manchmal auch rezessiv-rote, niemals aber schwarze Nachkommen. Bei der Paarung von zwei Rotbraunen entstehen meistens Rotbraune, aber ab und zu auch rezessiv-rote oder schwarze Nachkommen, was sehr viel besser zu den Paarungsergebnissen passt. Man beachte, dass der Ausdruck “fawn” eine weitere Beschreibung ist, die aus der Sicht der Genetik kein bisschen exakter ist als der Ausdruck „braun“. Die meisten „fawn-farbigen“ Alpakas, insbesondere die dunkleren, sind wahrscheinlich eher helle Rotbraune mit schwarzen Füßen als rezessiv-rote Tiere. Für diese Diskussion habe ich jedoch nur den Ausdruck „braun“ ersetzt. Die Zucht in Australien ist weitgehend auf die Steigerung der Erzeugung von weißen Vliesen konzentriert. Bei 53 % aller Paarungen ist mindestens ein Elterntier weiß. Die Paarung von weißen mit farbigen Tieren ergibt 30 % weiße und 70 % farbige Nachkommen. Ich bin ferner der Ansicht, dass weiße Alpakas von ihrem Äußeren her anhand der Farbe ihrer Augen, der Haut und der Ballenpolster in drei unterschiedliche Gruppen eingeteilt werden können. Weiße Alpakas mit rosa Ballen und dunklen Augen sind wahrscheinlich sehr helle rezessiv-rote Tiere, und weiße Alpakas mit schwarzen Füßen und dunklen Augen sind vermutlich sehr helle Agouti- oder Braune-(bay)-Typen. Falls dies zutrifft, dann sollten sich zwei weiße Alpakas mit rosa Füßen genauso fortpflanzen und es wäre zu erwarten, dass sie weißer erscheinen als weiße Alpakas mit schwarzen Füßen. Es ist nicht überraschend, dass die meisten importierten Alpakas des “Peru-Typs”, die vermut- lich teilweise wegen des Weißgrades ihres Vlieses ausgesucht worden waren, ebenfalls rosa Füße haben. Ein dominant-weißes Tier wäre ein weißes Tier mit dunklen Augen und schwarzen Füßen, und es müsste durch die Zuchtergebnisse bewiesen werden, dass es mindestens 50 % weiße Nachkommen mit dunklen Partnern hervorbringt. System der Beschreibung, das die wichtigen Einzelheiten der Farbmuster umfasst. Dies würde es den Züchtern ermöglichen, Entscheidungen auf der Basis des Genotyps ihrer Alpakas zu treffen und nicht anhand der Farbe, die sie im Vlies auf dem Rücken der Tiere wahrnehmen. Jeder der beiden Typen könnte natürlich auch jede erdenkliche Kombination aus weiß-markierenden Genen tragen. Die dritte Gruppe sind weiße Tiere mit blauen Augen; über diese werden wir im zweiten Teil ausführlicher sprechen. Elizabeth Paul Erewhon Alpacas [email protected] Ganz offensichtlich sind die Herdbuchdaten mit Fehlern behaftet, da die zur Beschreibung der Farben verwendeten Parameter wenig Bezug zu den tatsächlichen Genotypen von Alpakas haben. Zum jetzigen Zeitpunkt haben wir keine Möglichkeit, das Verhältnis von weißen Tieren mit rosa Füßen zu weißen Tieren mit schwarzen Füßen in der nationalen Herde zu bestimmen, wir wissen auch nicht, wie viele „schwarze“ Tiere in Wirklichkeit dunkle Rotbraune sind. Trotzdem können wir den Daten viele nützliche Informationen und interessante Trends entnehmen. Während des erfassten Zeitraums der Zucht waren nahezu 80 % der Nachkommen farbig, wobei der Großteil wahrscheinlich ein “Rotbraun”- Farbton war. SCHLUSSFOLGERUNG: Kombiniert man die Informationen in den Vererbungsmodellen mit den Pigmentstudien, dem phänotypischen Erscheinungsbild sowie den Zuchtergebnissen, kommt man zu der Annahme, dass bei Alpakas im allgemeinen dasselbe System bei der Farbvererbung vorliegt wie bei anderen Großtieren. Ein “dunkel-fawn-farbiges” Alpaka könnte beim Scheren dasselbe dunkelbraune Vlies wie ein “hellbraunes” Alpaka zeigen, da das Pigment bei beiden Vliesen dasselbe ist. Das wirtschaftliche Ergebnis wird das gleiche sein. Vom Standpunkt der Genetik aus könnten beide entweder rezessiv-rote Tiere oder „Rotbraune“ oder je eins von beiden sein. Die Paarungsergebnisse werden sich entsprechend unterscheiden. Dies ist der Grund für die Verwirrung bei der Zucht auf Farbe bei Alpakas. Also brauchen wir eindeutig ein genaueres Übersetzt von Karin Roßmann LITERATURANGABEN: Australian Alpaca Association Herdbuch Band 1-11 Gower, J. “Horse Colour Explained – A Breeder’s Perspective” 1999. Kangaroo Press Hart, Ken. “ The Dominant White allele is the Top Dominant allele in the Agouti Series”, presented to the University of Western Australia in November 2001. Little, C.L. “The Inheritance of Coat Colour in Dogs.” 5th Ed 1973. Howell Book House, New York. Paul, Elizabeth: ‘The Alpaca Colour Key” privately published November 2002. Searle, A.G. “Comparative Genetics of Coat Colour in Mammals”. Logos Press Limited, London 1968. Sponenberg, Dr. Phillip “Some Educated Guesses on Colour Genetics of Alpacas,” The Alpaca Registry Journal, Vol 1V, No1, Spring 2001. Sponenberg, D. Phillip “Equine Colour Genetics” 2nd Ed. 2003 Iowa State Press Blackwell Pub. Co. 43