Klassische Genetik2013
Werbung

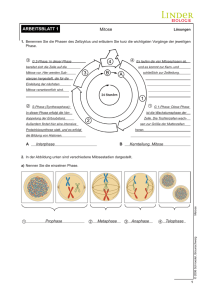
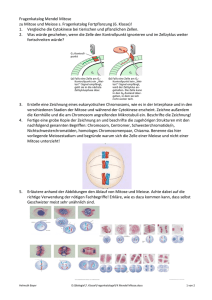
1 Klassische Genetik Historischer Rückblick • mit Diskussion um Abstammungslehre seit ~1800 reges Interesse an Genetik • 1865 Mendelsche Regeln • 1900 Wiederentdeckung: Beginn der klassischen Genetik; Verbindung mit Cytologie und Entdeckung der Chromosomen • ab 1953 Molekulare Genetik: Beginn durch Crick/Watson • 1960 Entzifferung des genetischen Codes • 80-er Jahre: Umsetzung in genetic engineering in großem Stil • 2000: human genome project 1.1 Mitose und Meiose 1.1.1 Allgemeines Aufgaben Zellteilung: Vermehrung, Erneuerung, Reparatur, Wachstum Bei Zellteilung muss gleichartiges genetisches Material (DNA) in beide Tochterzellen gelangen. Bei Eukaryoten sind DNA-Moleküle eng geknäuelt und auf Histone aufgespult, die Chromosomen sind nur kurz vor und während der Zellteilung im Mikroskop zu sehen. Bakterien: Vermehrung durch Spaltung. Die meisten Gene liegen auf 1 Chromosom („ringförmige“ DNA). Chromosom wird verdoppelt, Kopien bleiben an Membran gebunden. Zwischen den Kopien wächst Membran, wenn Zelle das Doppelte der Ausgangsgröße hat, wird sie nach innen eingeschnürt – Cytokinese (Zellteilung); Eukaryoten: größeres Genom als Prokaryoten. Gene auf mehrere Chromosomen (für jede Art charakteristische Anzahl - z.B. Mensch in somatischen Zellen 46, in Gameten (Keimzellen = Ei- und Samenzelle) 23). DNA mit verbundenen Proteinen ist Chromatin. Vor Zellteilung Verdoppelung der Chromosomen (Schwesterchromatiden). Das Centromer hält die Schwesterchromatiden zusammen. Anschließend werden sie in der Mitose getrennt und als neue Chromosomen auf 2 Zellkerne verteilt. Auf die Mitose folgt i.a. die Cytokinese (Teilung des Cytoplasma). Durch fortgesetzte Mitosen entstehen z.B. die Billionen Körperzellen des Menschen aus der Zygote. Die Gameten entstehen durch eine abgewandelte Zellteilung – Meiose = Reduktionsteilung. 1.1.2 Der Zellzyklus – Übersicht Die Mitose-Phase = M-Phase ist der kürzeste Teil des Zellzyklus. Zwischen den mitotischen Teilungen liegt die wesentlich längere Interphase (90% der Zeit). Während der Interphase wächst die Zelle, erfüllt ihre Aufgabe und verdoppelt ihre Chromosomen in Vorbereitung auf die nächste Mitose. Interphase besteht aus 3 Abschnitten: G1-Phase (Wachstum), S-Phase (Synthese-Phase, Replikation), G2-Phase (wachsen und Vorbereitung der Mitose). Eine Zelle, die sich nicht mehr vermehrt, tritt quasi in einen Ruhezustand ein = G0-Phase. Korrekter Ablauf des Zellzyklus ist entscheidend für das Überleben. Daher existieren Kontrollpunkte, an denen eine Qualitätskontrolle der Zelle stattfindet, bevor der Zyklus weiterläuft. Das entsch. Stadium ist die G1-Phase. Kurz vor Beginn der S-Phase liegt der Restriktionspunkt. Wenn die Zelle ihn überschreitet, verdoppelt sich die DNA und anschließend findet die Teilung statt. Bei Krebs sind solche Kontrollpunkte gestört. (Zellen, die den Zellzyklus nicht mehr oder nur noch langsam durchlaufen, müssen, wenn sie sich teilen sollen, durch Wachstumsfaktoren stimuliert werden. Bsp.: Blutplättchen geben PDGF (platelet derived growth factor) ab, der die an eine Verletzung angrenzenden Hautzellen zur Teilung und Wundheilung anregt. Weiteres Bsp. ist Erythropoetin (EPO): von Nieren produziert, regt Knochenmarkzellen zur Teilung an, stimuliert Erzeug. von Erythrocyten (auch Doping!!!)) Helmuth Bayer D:\Biologie\7. Klasse\2013\Klassische Genetik2013.docx 1 von 5 1.1.3 Mitose 5 Abschnitte: Prophase, Prometaphase, Metaphase, Anaphase, Telophase • Prophase: Spindel aus Mikrotubuli und and. Proteinen entsteht. Der Bau beginnt im Centrosom. Während der Interphase verdoppelt sich Centrosom und die beiden Tochtercentrosomen liegen dicht neben dem Zellkern. In Prophase und Prometaphase rücken sie weiter auseinander und von ihnen strahlen nach allen Seiten Spindelfasern aus. Die beiden Chromatiden eines Chromosoms besitzen je ein Kinetochor (Proteinkomplex im Bereich des Centromers), an dem einige vom Centrosom ausgehende Spindelfasern angeheftet sind (Kinetochorfasern). Andere Spindelfasern treten nicht mit Kinetochor in Kontakt sondern positionieren die Spindel in der Zelle (Polfasern). • Prometaphase Zerfall der Kernhülle (jetzt können die Spindelfasern in den Zellkern eindringen und mit den Chromosomen in Kontakt treten). • Metaphase: Aufreihung der Chromosomen in der Äquatorialplatte. (bester Zeitpunkt um Chromosomen zu erkennen, weil sie hier max. kondensiert sind) • Anaphase: beginnt mit der Teilung des Centromers in jedem der Chromosomen Schwesterchromatiden lösen sich voneinander (sie sind jetzt getrennte Tochter-Chromosomen) und wandern zu den entgegengesetzten Zellpolen (Tubuli werden depolymerisiert; 1µm/min). Die Polfasern sorgen für Streckung der Zelle (ATP!). • Telophase: Bildung neuer Zellkerne – Ende der Mitose („Kernteilung“). • Gleichzeitig teilt die Cytokinese (meist) die Zelle. 1.1.4 Cytokinese: • tierische Zellen: Furchung - innerhalb der Membran liegt ein Ring aus Aktinfasern (Muskel!) der sich in WW mit Myosin zusammenzieht und dadurch die Zelle einschnürt. Wenn die letzte Verbindung reißt sind Tochterzellen getrennt. • Pflanzenzellen: keine Furchung (starre Zellwand!)– stattdessen Bildung einer Zellplatte, die ca in der Ebene der Metaphaseplatte durch die Verschmelzung von Golgi-Vesikeln entsteht. Es entsteht im Zellinnern eine Doppelmembran, die schließlich mit der Plasmamembran zusammenwächst. Zwischen den Membranen der Zellplatte wird schließlich die neue Zellwand aufgebaut. Die Verteilung der restlichen Organellen erfolgt nicht genau. 1.1.5 Meiose 1.1.5.1 Allgemeines Vor Meiose Verdopplung der Chromosomen statt. Nach dieser Replikation ereignen sich 2 aufeinanderfolgende Zellteilungen – Meiose I und II. Dabei entstehen insgesamt 4 Tochterzellen, von denen jede nur halb so viele Chromosomen wie die Ausgangszelle besitzt (haploid). Während durch die Mitose Zellen entstehen, die genetisch identisch mit den elterlichen Zellen sind, gehen aus der Meiose Zellen hervor, die sich genetisch von den Elternzellen und auch untereinander unterscheiden. 1.1.5.2 Ablauf Meiose I Prophase I: Paarung der replizierten (aus 2 Schwesterchromatiden bestehenden) Chromosomen mit ihren Homologen. Die 4 eng aneinander liegenden Chromatiden bilden eine Tetrade. Es kommt zu Überkreuzungen von Nicht-Schwesterchromatiden (Chiasmata). Chiasmata sind die sichtbare Folge eines genetischen Austauschs = Crossing-over. Metaphase I: Anordnung der homologen Chromosomenpaare in Metaphaseplatte. Anaphase I: Trennung der homologen Chromosomenpaare (Centromere teilen sich nicht) (in der Mitose Trennung der Schwesterchromatiden!) Durch Telophase I und Cytokinese (Teilung des Cytoplasma) entstehen 2 Tochterzellen Meiose II Helmuth Bayer D:\Biologie\7. Klasse\2013\Klassische Genetik2013.docx 2 von 5 trennt die Schwesterchromatiden und ihr Mechanismus entspricht völlig dem der Mitose. Da sich die Chromosomen jedoch zwischen Meiose I und II nicht replizieren, resultiert aus der Meiose eine Halbierung des Chromosomensatzes der Zelle. 1.1.5.3 genetische Variabilität der Nachkommen Freie Rekombination von Chromosomen Jedes homologe Chromosomenpaar kann sich in der Metaphase I unabhängig von anderen Chromosomenpaaren orientieren – daher führt Meiose I zu völlig willkürlicher Verteilung von mütterlichen und väterlichen Chromosomen. Jeder Gamet repräsentiert also eine von allen möglichen Rekombinationen (Neukombinationen) mütterlicher und väterlicher Chromosomen. Zahl der möglichen Chromosomenkombinationen in den Gameten beträgt 2n, wobei n die haploide Chromosomenzahl darstellt (beim Menschen ca. 8 Millionen, genau 8388608). Jeder von einem Menschen produzierte Gamet enthält also eine von acht Millionen möglichen Rekombinationen von väterlichen und mütterlichen Chromosomen. Crossing-over Das Crossing-over erzeugt Chromosomen, in denen Gene mütterlichen und väterlichen Ursprungs kombiniert sind (Prophase I). Crossing-over findet statt, wenn sich homologe Abschnitte zweier Nicht-Schwesterchromatiden berühren. Beim Menschen beobachtet man im Durchschnitt 2 oder 3 solcher Crossing-over Ereignisse pro Chromosomenpaar. Zufälligkeit der Befruchtung Ein menschliches Ei enthält eine von rund acht Millionen möglichen Kombinationen von Chromosomen und wird von einem Spermium befruchtet, das ebenso eine von etwa acht Millionen verschiedenen Kombinationen von Chromosomen enthält. Ohne das Crossing-over in Betracht zu ziehen, produzieren die beiden Eltern eine Zygote, die eine unter 64 Billionen möglichen diploiden Rekombinationen darstellt. Daher ist es nicht verwunderlich, dass Brüder und Schwestern so unterschiedlich sein können. 1.1.5.4 Fragen • Eine menschliche Zelle, die 22 Autosomen und ein Y-Chromosom enthält ist: eine Somazelle ♂, eine Zygote, eine Somazelle ♀, ein Spermium (r), ein Ei. • Homologe Chromosomen bewegen sich zu den entgegengesetzten Zellpolen einer sich teilenden Zelle während: der Mitose, der Meiose I (r), der Meiose II, der Befruchtung, der Querteilung • Die Meiose II ist einer Mitose ähnlich, weil: homologe Chromosomen eine Tetrade ausbilden, vor der Zellteilung DNA-Replikation stattfindet, die Tochterzellen diploid sind, sich die Schwesterchromatiden während der Anaphase trennen (r), die Chromosomenzahl reduziert wird. • Wieviele verschiedene Kombinationen mütterlicher und väterlicher Chromosomen können in Gameten von Organismen mit einer diploiden Chromosomenzahl von 8 auftreten: 2, 4, 8, 16 (r), 32, 64 • Crossing-over trägt zur genetischen Variabilität bei, da es zum Austausch von Chromosomenbereichen führt zwischen: Schwesterchromatiden eines Chromosoms, Chromatiden nicht-homologer Chromosomen, Nicht-Schwesterchromatiden homologer Chromosomen (r), nicht-homologen Loci des Genoms, Autosomen und Geschlechtschromosomen. 1.2 Mendelsche Regeln Arbeit mit Erbsen; Augustiner Chorherr • 1. Mendelsche Regel (Uniformitätsregel) Linder3 S 36 Helmuth Bayer D:\Biologie\7. Klasse\2013\Klassische Genetik2013.docx 3 von 5 Kreuzt man zwei reine Linien miteinander so sind deren F1-Nachkommen (Tochtergeneration) alle gleichartig (uniform). Sie können in der Merkmalsausprägung in der Mitte zwischen den Eltern liegen (intermediäre Vererbung) oder vollkommen dem einen Elter, der dann als dominant bezeichnet wird, gleichen. Die Eigenschaften des anderen Elters bezeichnet man als rezessiv. • 2. Mendelsche Regel (Spaltungsregel) Kreuzt man Individuen der F1-Generation untereinander, so findet in der F2-Generation eine Aufspaltung der Merkmale nach bestimmten Gesetzen statt. Beim monohybriden Erbgang (Eltern unterscheiden sich in einem Merkmal) lautet das Verhältnis dominant:rezessiver Elterntyp 3:1, beim intermediären Erbgang 1:2:1. • 3. Mendelsche Regel (Kombinationsregel) Unterscheiden sich die Eltern in mindestens zwei Merkmalen, so ergibt sich nach Kreuzung der F1-Individuen in der F2-Generation eine Zufallskombination der unabhängigen Merkmale (⇒ es treten auch Neukombinationen auf). Die Häufigkeit der Kombinationstypen verhalten sich beim dominant-rezessiven Erbgang wie 9:3:3:1. (Erbgang mit dominanten Merkmalen ist am häufigsten). Darstellung im Punnett-Quadrat. Erklärung Mendels In Zellen befinden sich diskrete, materielle Einheiten = Erbfaktoren = Gene, die das Vererbungsphänomen bedingen. Jedes Gen ist in jeder Zelle zweifach vorhanden, außer in Geschlechtszellen (einfach). Wichtig: Mendelsche Regeln lassen keine verbindlichen Prognosen sondern nur Wahrscheinlichkeitsaussagen zu! (s. Stammbaumanalyse – gehört zu diesem und zum Kapitel Humangenetik!!!!) Einige andere wichtige Grundbegriffe Unterschiedliche Anlagen für dasselbe Merkmal (z.B. Blütenfarbe rot und weiß) nennt man Allele. Allele können reinerbig (= homozygot = AA oder aa) vorkommen oder mischerbig (= heterozygot = Aa) (meist ist ein Allel dominant, das andere rezessiv!) Phänotyp = äußeres Erscheinungsbild Genotyp = Erbkonstitution Die Mendelschen Regeln sind oft unzureichend, um beobachtete Erbgänge zu erklären. Gründe dafür sind z.B. Abstufungen der Dominanz (Allele können unterschiedliche Abstufungen der Dominanz bzw. Rezessivität untereinander aufweisen), Kodominanz (Bsp. Blutgruppe AB; 2 Allele wirken sich unabhängig voneinander auf den Phänotyp aus, die Merkmalsausprägung liegt nicht zwischen den beiden Zuständen), multiple Allele (die meisten Gene haben mehr als 2 verschiedene Allele) und Pleiotropie (die meisten Gene können sich auf mehr als einen Phänotyp auswirken, z.B. für die unterschiedlichen Symptome von Mukoviszidose). Außerdem können auch zwei oder mehr Gene an der Festlegung eines bestimmten Phänotyps beteiligt sein. So bezeichnet Epistasie die übergeordnete Wirkung eines Gens und liegt dann vor, wenn ein Gen die phänotypische Expression eines anderen Gens verändert. So ist bei vielen Säugern die schwarze Fellfarbe dominant über eine braune. Allerdings ist hier noch etwas zu bedenken. Ein zweites Gen bestimmt nämlich, ob das verantwortliche Pigment überhaupt erst in den Haaren abgelagert wird oder nicht. Ist eine Maus an diesem zweiten Genort homozygot rezessiv, kann sich kein Pigment ablagern und die Maus ist ein Albino – egal, welches Allel am Genort für Farbe vorliegt! 1.3 Geschlechtsbestimmung Linder3 S 48 1.3.1 Genotypische Geschlechtsbestimmung = Festlegung des Geschlechts durch Erbfaktoren • Haplogenotypische Geschlechtsbestimmung Helmuth Bayer D:\Biologie\7. Klasse\2013\Klassische Genetik2013.docx 4 von 5 • • Geschlechtsdifferenzierung in der haploiden Phase. Bei Niederen Pflanzen, Moosen, Farnen. Aus der Zygote entstehen 2 Männchen und 2 Weibchen. Diplogenotypische Geschlechtsbestimmung Geschlechtsdifferenzierung erfolgt in diploider Phase Häufigster Fall ist der X-Y-Typus: im Chromosomensatz befinden sich neben normalen Chromosomen = Autosomen auch spezielle Chromosomen, in denen sich die Chromosomensätze der Geschlechter unterscheiden = Heterosomen → 2 Formen: X-, YChromosom (Jede Geschlechtszelle enthält 1 Heterosom und haploiden Autosomensatz). XXWeibchen können nur X-Eier, die XY-Männchen X- oder Y-Spermien produzieren ⇒ Geschlechterverhältnis 1:1. In manchen Tiergruppen (Schmetterlinge, einige Fische, Amphibien, Reptilien, Vögel) sind die Weibchen heterogametisch und die Männchen homogametisch → W-Z-Typus. Bei anderen Tieren (z.B. Wanzen) haben die Weibchen ein Heterosom mehr als die Männchen (X-0-Typ der Geschlechtsbestimmung) Welche genetischen Faktoren bestimmen Geschlecht o Drosophila-Typ (Taufliege): Geschlecht wird durch Verhältnis der Autosomen zu XChromosomen bestimmt. A:X ≤ 0,5 ⇒ Männchen; A:X ≥ 1 ⇒ Weibchen; Bei dazwischenliegendem Verhältnis entstehen Intersexe (Y-Chromosom nicht entscheidend!) o Melandrium-Typ: (Lichtnelke) Y-Chromosom ist männlich determinierend (auch gegenüber mehreren X-Chromosomen), z.B. beim Menschen. o Lymantria-Typ (Schwammspinner): W-Z-Typ; Es gibt starke und schwache Rassen; Intersexualität tritt in jenem Geschlecht auf, das bei der Kreuzung der schwachen Rasse angehört, z.B. Weibchen (schwach) gekreuzt mit Männchen (stark) ergibt starke Männchen und intersexuelle Weibchen. Parthenogenetische Geschlechtsbestimmung Weibchen entstehen i.a. aus befruchteten Eiern, Männchen aus unbefruchteten Eier, z.B. Bienen 1.3.2 Phänotypische Geschlechtsbestimmung = Geschlechtsbestimmung durch Umweltfaktoren; relativ selten! • Bonellia (Echiuride): Junge werden in Anwesenheit eines geschlechtsreifen Weibchens zu Männchen, sonst zu Weibchen. • Stabheuschrecken: tiefe Temperatur führt zur Entstehung von Weibchen, Temperaturen ab ~30°C zur Entstehung von Männchen • Ophryotrocha (Ringelwurm): kleine Tiere sind Männchen und schlagen ab gewisser Größe ins weibliche Geschlecht um. Durch Zurückschneiden kann der männliche Zustand wieder hergestellt werden. • Geschlechtsumwandlungen jedoch auch bei höheren Tieren (Fische, Amphibien). Helmuth Bayer D:\Biologie\7. Klasse\2013\Klassische Genetik2013.docx 5 von 5