5 Ökologie
Werbung

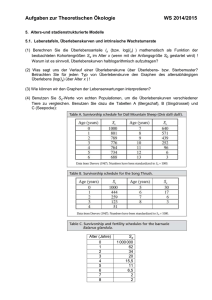
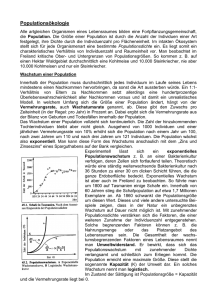
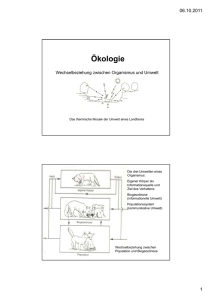
5 Ökologie 5.1 Aspekte der Ökologie 5.1.1 Grundbegriffe Ökologie ist eine Wissenschaftsdisziplin und die Umwelt eines Organismus oder einer Population ist Gegenstand der ökologischen Forschung. Sie ist aber vom wissenschaftlichen Naturschutz deutlich abgegrenzt! Einen Überblick über die Bandbreite der ökologischen Forschung gibt die Folie! Ökosystem: Es umfasst das Beziehungsgefüge zwischen den verschiedenen Organismenarten (Biozönose) in einem bestimmten Lebensraum (Biotop). Biozönose = Lebensgemeinschaft: Eine Gruppe von Populationen verschiedener Mikroorganismen, Pflanzen und Tiere, die sich an einem Ort infolge ähnlicher Ansprüche an ihre Umwelt einstellt. Population: Gesamtheit der Individuen einer Art, die in einem Siedlungsgebiet leben, sich dort in der Regel über mehrere Generationen fortpflanzen und so genetische Kontinuität zeigen. Benthal: Bodenzone von Gewässern Pelagial: freier Wasserbereich (nicht Ufer und nicht Boden eines Gewässers) Biomasse: Masse aller lebenden Organismen eines Ökosystems, einer Trophiestufe oder eines Gebiets bzw. organische Substanzen, die von autotrophen Organismen aus anorganischen Stoffen synthetisiert werden. Umweltkapazität K: Aufnahmekapazität eines Lebensraums, die mit einer maximalen Populationsgröße verbunden ist, die durch diesen Lebensraum noch ernährt werden kann. 5.1.2 Wachstum von Populationen Exponentielles Wachstum Bakterien können sich unter optimalen Bedingungen in 20min durch Teilung verdoppeln. Wenn sich die Geschwindigkeit fortsetzt, ohne dass Individuen absterben, wäre die Zahl der Bakterien nach 1,5d so groß, dass sie die Erde mit einer 30cm dicken Schicht bedecken würden. Dies gilt natürlich nur, wenn alle Individuen Zugang zu einer unbegrenzten Nahrungsressource haben und alle Möglichkeiten ihrer Reproduktionsfähigkeit ausschöpfen können. Die Pro-Kopf-Wachstumsrate ist unter solchen Bedingungen konstant. 𝒅𝑵 Es gilt: 𝒅𝒕 = 𝒓𝑵 dN = Veränderung der Populationsgröße dt = Zeitraum, in dem das Populationswachstum analysiert wird r = b-d, wobei b = Pro-Kopf-Geburtenrate und d = Pro-Kopf-Sterberate B gibt die Anzahl der Geburten an. Finden z.B. in einer Population von 1000 Individuen während eines Jahres 34 Geburten statt, liegt die jährliche Pro-Kopf-Geburtenrate bei 34/1000 oder 0,034. Wenn wir diese Rate kennen, können wir mithilfe der Formel B=bN berechnen, wie viele Geburten pro Jahr wieder in einer Population beliebiger Größe stattfinden werden. Wenn z.B. die jährliche Pro-Kopf-Geburtenrate 0,034 beträgt und die Population aus 500 Individuen besteht, ist B=bn – B=0,034x500=17 pro Jahr. Entsprechend kann man mit der Pro-Kopf-Sterberate verfahren. Der Wert r (b-d) gibt demnach an, ob eine Population wächst (r>0) oder schrumpft (r<0). Die Größe einer exponentiell wachsenden Population nimmt mit konstanter Geschwindigkeit zu (s. Diagramm). Die exponentielle Wachstumskurve ist für solche Populationen charakteristisch, die einen neuen Lebensraum erobert haben oder durch eine Katastrophe dezimiert wurden und sich nun wieder erholen können. Helmuth Bayer H:\Biologie\7. Klasse\2015\ÖkologieV15.docx 1 von 5 Das logistische Wachstumsmodell Man kann das exponentielle Wachstumsmodell dahingehend verändern, dass die Veränderung der Wachstumsrate mitberücksichtigt wird, wenn sich die Population der Wachstumsgrenze nähert. (Lotka-Volterra-Gleichungen). Zur Formulierung geht man vom exponentiellen Modell aus und fügt einen Ausdruck hinzu, durch den sich die Pro-Kopf-Zunahme mit zunehmender aktueller Populationsgröße N verringert. Wenn K die maximale Populationsgröße nach Erreichen der Umweltkapazität darstellt, entspricht K-N der Anzahl der zusätzlichen Individuen, die sich in einer Umwelt erhalten können, und (K-N)/K dem Anteil von K, der noch für ein weiteres Populationswachstum zur Verfügung steht. Multipliziert man die exponentielle Wachstumsrate rN mit (K-N)/K verändert sich mit zunehmendem N die Populationsgröße. 𝐝𝐍 𝐊−𝐍 = 𝐫𝐍 ∗ 𝐊 𝐝𝐭 Logistisches Modell und natürliche Populationen Das Wachstum von Laborpopulationen mancher kleinerer Tierarten entspricht bei begrenzten Ressourcen recht gut einer S-förmigen Wachstumskurve. Einige Grundannahmen, auf denen das logistische Modell beruht, treffen eindeutig nicht auf alle Populationen zu. In Wirklichkeit tritt häufig eine Verzögerung des Populationswachstums vor Erreichen der Kapazitätsgrenze ein, bevor die negativen Auswirkungen des Populationswachstums einsetzen. Wird z.B. die Nahrung für eine Population zum limitierenden Faktor, so geht die Reproduktion irgendwann zurück, aber die Weibchen können sich für kurze Zeit mithilfe ihrer Energiereserven noch weiter reproduzieren. Dies kann dazu führen, dass die Population vorübergehend über die Umweltkapazität hinaus anwächst. Verringert sich die Population dann unter die Grenze der Umweltkapazität, kommt es im Wachstum zu einer Verzögerung, bevor wieder eine größere Anzahl von Nachkommen geboren wird. Außerdem fällt Individuen das Überleben oder die Reproduktion schwerer, wenn die Population zu klein ist. Eine einzelne Pflanze wird z.B. durch starken Wind geschädigt, in einer ganzen Gruppe von Individuen dagegen ist sie gut geschützt; oder die Partnerfindung wird erschwert, wenn die Individuendichte zu gering ist. In Populationen, die sich an der Umweltkapazität befinden, sollten hohe Konkurrenzfähigkeit und effiziente Ressourcennutzung begünstigt werden. Bei niedriger Individuendichte dagegen fördert die Selektion eher Anpassungen, die eine schnelle Reproduktion ermöglichen. Die Selektion von Merkmalen, die auf die Individuendichte zielen, wird als K-Selektion oder dichteabhängige Selektion bezeichnet (Bsp.: ausgewachsene Bäume in einem alten Wald). Die Selektion auf Merkmale, die einen möglichst großen Reproduktionserfolg in einer ansonsten gering besiedelten Umwelt begünstigen, bezeichnet man dagegen als r-Selektion. Die Abkürzungen beziehen sich auf die Variablen in der Gleichung des logistischen Modells. Dichteabhängige Einflüsse auf das Populationswachstum Eine Geburten- oder Sterberate, die sich mit der Individuendichte einer Population nicht verändert, bezeichnet man als dichteunabhängig. Dagegen bezeichnet man eine Mortalitätsrate, die mit der Individuendichte ansteigt, als dichteabhängig. Für die Rückkopplung zwischen Individuendichte und der Geburten-/Sterberate sorgen Regulationsmechanismen, die zu einer Erhöhung der Mortalitätsrate und einer Erniedrigung der Natalitätsrate führen, bis das Populationswachstum zum Stillstand kommt, dies sind z.B.: Konkurrenz um Ressourcen, Revierverhalten, Krankheiten, Räuber, giftige Stoffwechselendprodukte (z.B. Kaulquappen) 5.1.3 Wechselbeziehungen zwischen Organismen Interspezifische Konkurrenz Sie hat Einfluss auf das Populationswachstum, Überleben und die Reproduktion der Konkurrenten. Helmuth Bayer H:\Biologie\7. Klasse\2015\ÖkologieV15.docx 2 von 5 Konkurrenzausschluss: Was geschieht in einer Lebensgemeinschaft über längere Zeiträume, wenn zwei Arten direkt miteinander um begrenzte Ressourcen konkurrieren? Das wurde u.a. an 2 Pantoffeltierarten (Paramecium aurelia und P. caudatum) untersucht. Wurden die Arten unter konstanten Bedingungen und getrennt gehalten, so nahm die Größe beider Populationen schnell zu. Mit steigender Individuendichte näherten sich die Populationen ihrer Kapazität an und die Wachstumskurve flachte ab. Brachte man die beiden Kulturen dagegen zusammen in ein Gefäß, starb nach einer gewissen Zeitspanne P. caudatum aus. Wenn keine weiteren Einflüsse hinzukommen, nutzt eine Art die Ressourcen also effizienter als die andere und reproduziert sich entsprechend schneller. Schon ein geringfügiger Reproduktionsvorteil führt schließlich dazu, dass der unterlegene Konkurrent lokal ausstirbt – Konkurrenzausschluss-Prinzip! Ökologische Nische: Wenn der Lebensraum einer Art ihre Adresse ist, dann ist die Nische ihr ausgeübter Beruf. Sie ist demnach durch einen Raum gekennzeichnet, in dem die Individuen der Art leben und der sich aus Raum, Zeit, Nahrung, Temperatur und anderen Umweltfaktoren zusammensetzt. Mit der ökologischen Nische kann das Konkurrenzausschluss-Prinzip neu formuliert werden: Zwei Arten können in einer Lebensgemeinschaft nicht auf Dauer nebeneinander existieren, wenn sie dieselben ökologischen Nischen besitzen. Eine Ressourcenaufteilung zwischen Arten mit ähnlichen Umweltansprüchen führt zur Koexistenz dieser Arten in einer Lebensgemeinschaft (Nischenaufteilung). Prädation Ist eine Wechselwirkung zwischen 2 Arten, wobei der Räuber die Beute tötet und ganz oder teilweise frisst. Räuber-Beute-Beziehungen wirken sich bei beiden Partnern immer auch auf den Reproduktionserfolg ihrer Populationen aus. Innerhalb der Prädatoren werden je nach Jagdstrategie verschiedene funktionelle Gruppen unterschieden: Jäger (Katze), Fallensteller (Netzspinnen), Lauerer (Gottesanbeterin), Sammler (z.B. Watvögel), Filtrierer (Flamingo). Während Räuber spezifische Anpassungen besitzen, die ihnen das Fangen der Beute erleichtern, können Beutetiere ebenfalls über verschiedene Adaptationen dem Gefressenwerden entgehen. Aktive Selbstverteidigung kommt seltener vor, andere suchen Verstecke auf, flüchten oder bilden Herden, die es dem Angreifer erschweren, sich auf ein bestimmtes Beutetier zu konzentrieren (Sardinen). Darüber hinaus besitzen Tiere eine Vielzahl morphologisch-anatomischer und physiologischer Anpassungen, die der Verteidigung dienen (Mimese (Tarnung), Mimikry (Nachahmung)). Parasitismus Ist eine Interaktion, bei der ein Organismus, der Parasit, von einem anderen, dem Wirt, ernährt wird, wobei der Wirt immer geschädigt wird. Parasitoide töten ihren Wirt (Schlupfwespen, Raupenfliegen). Man unterscheidet Endo- und Ektoparasiten. Herbivorie Ist eine Interaktion, bei der der Herbivore Teile von Pflanzen verzehrt. Bei den meisten Herbivoren handelt es sich um Wirbellose (Insekten, Schnecken) Mutualismus = Symbiose Ist eine Wechselbeziehung, die beiden Arten einen Vorteil bringt. Je nach der Stärke der Bindung der beiden Partner unterscheidet man zwischen einem fakultativen Mutualismus (Allianz) und einem für beide Partner lebensnotwendigen obligaten Mutualismus (Eusymbiose). Ein Beispiel für fak. Mutualismus ist die Beziehung zwischen der Büffelhornakazie und einer Knotenameise, die in deren Dornen wohnt. Weitere Beispiele: Bestäubungsanpassungen, Samenausbreitung durch Vögel, Putzsymbiosen (Fische, Vögel), Mykorrhiza, Flechten Helmuth Bayer H:\Biologie\7. Klasse\2015\ÖkologieV15.docx 3 von 5 Parabiose und Kommensalismus Ist eine Wechselbeziehung zwischen 2 Arten, die der einen Art von Vorteil ist, der anderen aber weder nutzt noch schadet. Handelt es sich dabei um eine Nahrungsbeziehung, so spricht man auch von Kommensalismus. Bei der Parökie nutzt eine Art den Schutz einer anderen. Vögel suchen oft Schutz in der Nähe anderer Tiere. Schwarzhalstaucher brüten in Kolonien von Lachmöwen. Sie verlassen sich auf das Alarmsystem der Möwen und genießen Schutz gegen Greifvögel und Krähen als Eiräuber. Zwergtaucher schwimmen hinter Blässhühnern her und verspeisen die von diesen aufgejagten Wasserinsekten. Viele Jungfische suchen Schutz bei Medusen. Gegen das Nesselgift sind sie offenbar immun oder durch einen Stoff geschützt. Beliebte Vergesellschaftungspartner sind die Diademseeigel, die im Seichtwasser an der Basis der Korallenriffe leben. Zwischen ihren langen Stacheln suchen vorwiegend Fische und Garnelen mechanischen und optischen Schutz. Unter Synökie versteht man die Einmietung von Organismen in anderen Nestern. An den deutschen Küsten brütet die Brandgans in Dachs-, Kaninchen- und Fuchsbauten (ohne dass sie der Fuchs frisst!). In Bauten von Säugetieren leben Insekten und Milben als Gäste. Epökie ist die Besiedelung eines anderen Organismus. Der Aufsiedler oder Symphoriont kann dabei beweglich sein oder festgeheftet auf seinem Partner leben. Beispiele dafür bieten Haarund Federlinge in der Körperbedeckung von Säugern und Vögeln. Sie ernähren sich meistens von Keratin. Entökie ist der Aufenthalt in nach außen offenen Körperhöhlen anderer Tierarten. In die Mantelhöhle der Teichmuschel legt das Weibchen des europäischen Bitterlings seine Eier, aus denen dort die Fischlarven schlüpfen. An ein entökes Leben sind die Nadelfische deutlich angepasst. Das Hinterende der schuppenlosen Fische hat keine Schwanzflosse, sondern läuft spitz aus. Die Brustflossen sind rückgebildet oder fehlen ganz, und die Afteröffnung kann so weit nach vorn verlagert sein, dass die Fische ihren Wirt bei der Kotabgabe nicht zu verlassen brauchen. Hauptaufenthaltsort sind Seegurken. Phoresie ist die aktive Benutzung eines anderen Tieres für Transportzwecke, und wird am besten durch die Situation des blinden Passagiers gekennzeichnet. In faulenden Substraten lebende Milben und Nematoden besteigen bei Verschlechterung der Milieuverhältnisse als Larven flugfähige Insekten und lassen sich in eine frische Lebensstätte bringen. Beispiele sind die Schiffshalter, die sich an andere Fische ansaugen und von ihnen transportieren lassen. Metabiose Ist ein auf Folgenutzung beruhendes Bisystem. Viele Vogelarten, z.B. Hohltaube, Kleiber, Fledermäuse nisten in Höhlen, die sie nicht selbst bauen (können). Sie sind auf Spechte angewiesen, die diese einst für sich selbst hergestellt haben. 5.1.4 Der Einfluss von dominanten Arten und Schlüsselarten auf die Struktur von Lebensgemeinschaften Artendiversität Sie umfasst zwei Parameter, den Artenreichtum (Anzahl verschiedener Arten in einer Biozönose) und die Häufigkeit = Abundanz der einzelnen Arten (Anteil der jeweiligen Art an der Gesamtzahl der Individuen) (s. Folie). Trophische Strukturen Struktur und Dynamik einer Lebensgemeinschaft hängen zu einem großen Teil von den Nahrungsbeziehungen zwischen den Organismenarten ab, das heißt von der trophischen Struktur. Diese beschreibt die Übertragung der Nahrungsenergien durch die verschiedenen Trophieebenen von ihrer Ausgangsbasis, den grünen Pflanzen und anderen autotrophen Organismen (Primärproduzenten) über Herbivoren (Primärkonsumenten) zu den Carnivoren (Sekundär-, Helmuth Bayer H:\Biologie\7. Klasse\2015\ÖkologieV15.docx 4 von 5 Tertiär- und Quartärkonsumenten) und schließlich Destruenten. Hinzu kommen Omnivoren. Diese sind Teil einer Nahrungskette, die oft als ein kompliziertes Nahrungsnetz aufgebaut ist (s. Folie). Arten mit großer Bedeutung für die Lebensgemeinschaft Einige Arten haben für Struktur und Erhaltung einer Lebensgemeinschaft eine besonders große Bedeutung – Schlüsselarten. Innerhalb der Schlüsselarten gibt es solche, die in besonders hoher Anzahl und Biomasse vertreten sind (Schlüsseldominanten) oder solche, die auch ohne Ausbildung einer großen Biomasse eine entscheidende Schlüsselrolle in der Lebensgemeinschaft haben (Schlusssteinarten). Ein Beispiel dafür ist der Meerotter, der im Nordpazifik als Räuber die Rolle einer Schlusssteinart einnimmt. Seine Hauptnahrung sind Seeigel, die von Braunalgen leben. Fallen die Meerotter aus, so werden von den Seeigeln die Braunalgen stark dezimiert und die besonders artenreiche Lebensgemeinschaft benthischer und pelagischer Wirbelloser sowie Fischen verschwindet. 5.1.5 Der Einfluss von Störungen auf Artendiversität und Artenzusammensetzung Noch vor einigen Jahrzehnten herrschte in der Ökologie die Ansicht, Lebensgemeinschaften müssten sich in einem mehr oder weniger stabilen Gleichgewichtszustand befinden, solange sie durch den menschlichen Einfluss nicht ernsthaft gestört werden Veränderungen in der Struktur und Zusammensetzung terrestrischer Lebensgemeinschaften lassen sich gut verfolgen, wenn nach einem größeren Störungsereignis, z.B. nach einem Vulkanausbruch, die dortigen Flächen zunächst völlig vegetationsfrei sind. Solche Flächen werden dann im Laufe der Zeit von verschiedenen Arten besiedelt, die nach und nach von anderen Arten abgelöst werden. Diesen Ablauf bezeichnet man als ökologische Sukzession. Von Primärsukzession spricht man, wenn dieser Vorgang in einem zuvor unbelebten Gebiet beginnt und eine Neulandbesiedlung sattfindet (Vulkaninseln, Gletschervorfeld, Flussufer mit starker Dynamik). Zunächst sind die Besiedler häufig nur autotrophe und heterotrophe Prokaryonten und Protisten. Die ersten photosynthetisch aktiven Organismen sind in der Regel Flechten und Moose. Wenn einmal Boden entstanden ist, werden Moose und Flechten von Kräutern und Gräsern überwachsen und es folgen Sträucher und Bäume. Am Ende wird die Fläche von einer Pflanzengesellschaft besiedelt, die als Schlussgesellschaft das Endstadium der Sukzession darstellt. Bis dorthin vergehen häufig Jahrhunderte bis Jahrtausende! Sekundärsukzession findet statt, wenn bereits entwickelte Organismengemeinschaften einer Störung unterliegen und die Entwicklung von Neuem wieder beginnt. Ein Beispiel ist die Regeneration des Waldes im Yellowstone-Nationalpark nach den Waldbränden des Jahres 1988. Sekundärsukzessionen folgen häufig auf einen menschlichen Eingriff. Bei Sukzessionen besteht ein Zusammenhang zwischen dem Sukzessionsgeschehen und den Umweltveränderungen, die durch den Vegetationswechsel verursacht werden (Bsp. Besiedelung eines Gletschervorfelds von der Glacier Bay (Alaska)). Der nackte Boden hat zunächst einen pHWert von 8 bis 8,4 (Karbonatgestein). Mit der beginnenden Vegetationsentwicklung geht er schnell zurück. Insbesondere die Zersetzung der sauren Fichtennadeln lässt den pH-Wert bis auf 4,0 sinken. Auch die Konzentration der mineralischen Nährstoffe im Boden ändert sich im Lauf der Zeit. Da der offene und organismenfreie Boden nach dem Rückzug des Gletschers nur einen geringen Stickstoffgehalt hat, können fast alle Pionierpflanzen zu Beginn der Sukzession wegen der unzureichenden Stickstoffversorgung nur schlecht wachsen. Frei lebende Cyanobakterien, die Luftstickstoff fixieren können, reichern Stickstoff an. Nachdem sich Silberwurz und Erlen etabliert haben (Symbiose mit N2-fixierenden Bakterien!), nimmt der N2-Gehalt des Bodens rapide zu. Durch die kontinuierliche Veränderung der Bodeneigenschaften schaffen die jeweiligen Besiedler die Voraussetzungen, damit sich auch neue Pflanzenarten etablieren können (Metabiose). Helmuth Bayer H:\Biologie\7. Klasse\2015\ÖkologieV15.docx 5 von 5