Kerntransfer und Reprogrammierung
Werbung

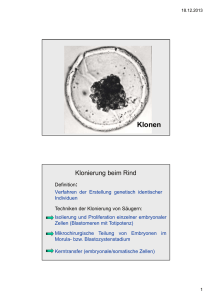
Fäden des Lebens LMU 50 Jahre DNA - Doppelhelix GENZENTRUM Lehrstuhl für Molekulare Tierzucht und Biotechnologie Münchner Wissenschaftstage 16. - 20. Juli 2003 Kerntransfer und Reprogrammierung – Chancen für eine individuelle Zelltherapie? Stammzellen – Heiler der Zukunft? Sobald eine Zelle eine Herzmuskelzelle geworden ist oder eine Hautzelle oder eine Nervenzelle, so bleibt sie dies bis zu ihrem Tod. Bei Stammzellen ist das nicht so – ihr Schicksal ist noch nicht festgelegt. Stammzellen können vermutlich viele Gewebe des Körpers bilden und eröffnen deshalb ein breites Spektrum von Anwendungsmöglichkeiten in der Medizin. Viele ihrer außerordentlichen Fähigkeiten wurden erst vor wenigen Jahren entdeckt. Der Begriff Stammzelle definiert Zelltypen, die Teilungs- und Differenzierungspotential besitzen. Nach der Vereinigung von Ei- und Samenzelle entstehen bei den ersten Zellteilungen zunächst Stammzellen, die als „totipotent“ bezeichnet werden. Jede einzelne dieser Zellen ist noch in der Lage, einen vollständigen Embryo mit Plazenta hervorzubringen. Nach gegenwärtigem Stand der Forschung verlieren die Zellen diese Totipotenz spätestens mit dem Eintritt des Embryos in das 16-Zell-Stadium und treten in den Zustand der Pluripotenz über. Im weiteren Verlauf kommt es zur Ausbildung der Blastozyste mit der inneren und äußeren Zellmasse - dem Embryoblast und dem Trophoblast. Aus den pluripotenten Zellen des Embryoblasten bilden sich nachfolgend die drei Keimblätter mit den Körperzellen. Aus dem Trophoblasten entsteht die Plazenta. Entnimmt man der Blastozyste eine Zelle aus dem Embryoblasten, so ist diese pluripotent, denn sie vermag zwar noch die Zellen des Körpers zu bilden, aber nicht mehr die Plazenta und somit keinen vollständigen, lebensfähigen Embryo. Pluripotente Stammzellen können auf verschiedenen Wegen erzeugt werden (Abb.1). Ihre Vorteile liegen in ihrer Fähigkeit zur Selbsterneuerung und in ihrem schnellen Wachstum. Dadurch kann vergleichsweise einfach die für eine therapeutische Anwendung benötigte große Zellzahl erzeugt werden: werden pluripotente ES-Zellen aus der Blastozyste isoliert, so vermehren sie sich unter Kulturbedingungen in Gegenwart eines bestimmten differenzierungshemmenden Faktors, ohne ihre Pluripotenz zu verlieren. Dabei bleibt ihr Potential erhalten, die meisten der circa 210 verschiedenen Zelltypen eines Säugers zu bilden. Nach Entzug des differenzierungshemmenden Faktors und Kultur der Zellen in Schalen, in denen sie nicht anheften können, bilden sich spontan Zellaggregate, die man auch als „embryoid bodies“ bezeichnet. Unter anderem findet man darin Herzmuskelzellen vor, erkennbar als Regionen spontan kontrahierender Aktivität. Pluripotente Stammzellen können in eine Vielzahl verschiedener Zelltypen differenzieren und so Basis für die Therapie eines breiten Spektrums von Erkrankungen sein. Die Differenzierung erfolgt unter verschiedenen Bedingungen spontan und kann durch Zusatz von Faktoren, die Wachstum und Zellteilung fördern, beeinflusst werden. In den meisten Fällen erhält man jedoch zunächst eine Mischpopulation verschiedener Zelltypen, aus der die für die Therapie benötigten Zellen erst selektiert werden müssen. Dafür stehen verschiedene Methoden wie beispielsweise das Einschleusen von zelltypspezifischen Markergenen zur Verfügung. In jedem Fall muss vor einer Transplantation erst eine homogene Population differenzierter Zellen des gewünschten Typs erzeugt werden. Eine Verunreinigung mit undifferenzierten Zellen birgt das Risiko der Entstehung eines Tumors. Innere Zellmasse So könnte ein Bild/eine Grafik eingefügt werden. Befruchtung Blastozyste Fetus ES ES = embryonale Stammzellen EG = embryonale Keimzellen EC = embryonale Karzinomzellen EG Teratokarzinom EC Pluripotente Stammzellen Abb. 1. Möglichkeiten der Erstellung pluripotenter Stammzellen „Therapeutisches Klonen“ – Schlüssel zur individualspezifischen Zellersatztherapie? Ein Problem bei der Verwendung von pluripotenten Zellen beziehungsweise ihren differenzierten Abkömmlingen ist, dass sie eine Immunreaktion bei dem zu behandelnden Patienten auslösen können. Daher wäre für eine Transplantation eine Unterdrückung der Immunabwehr und/oder eine Toleranzinduktion erforderlich. Zudem wird eine genetische Modifikation der transplantierten Zellen diskutiert. Ein zumindest in der Theorie sehr attraktiver Ansatz ist die Erzeugung pluripotenter Stammzellen mit Hilfe der Kerntransfertechnik. Durch Pionierexperimente am Roslin-Institut in Schottland wurde erstmals am Schaf „Dolly“ gezeigt, dass die Kerne differenzierter Zellen nach Transfer in entkernte Eizellen reprogrammiert werden können und sich aus den erzeugten Kerntransferembryonen lebensfähige Nachkommen entwickeln können. Diese Reprogrammierung spezialisierter Zellen wurde unter dem Schlagwort „Therapeutisches Klonen“ auch als Möglichkeit diskutiert, um pluripotente Stammzellen zu generieren, die eine Zellersatztherapie ohne immunologische Komplikationen ermöglichen könnten (Abb. 2). So könnte ein Bild/eine Grafik eingefügt werden. Abb. 2. Prinzip des „Therapeutischen Klonens“ Der Kerntransfer mit differenzierten Kernspenderzellen gelang mittlerweile bei verschiedenen Tierarten. Allerdings ist die Erfolgsrate des Verfahrens bislang gering: der Anteil normaler Nachkommen bezogen auf die in Empfängertiere übertragenen Kerntransferembryonen bewegt sich bei den meisten Spezies in der Größenordnung von nur wenigen Prozent. Als Ursachen für zahlreiche Fehlschläge werden Fehler bei der Reprogrammierung diskutiert. Trotzdem gelang es bei der Maus, nach dem Prinzip des therapeutischen Klonens pluripotente Zellen aus Kerntransferblastozysten zu generieren und therapeutisch einzusetzen. Die Strategie „Therapeutisches Klonen“ ist beim Menschen jedoch problematisch. Neben den bislang nur unzureichend verstandenen Mechanismen der Reprogrammierung und häufig dabei entstehenden Fehlern stellt allein schon die Verfügbarkeit einer ausreichenden Zahl geeigneter Eizellen ein großes Problem dar. Möglicherweise ließe sich dieses Problem jedoch durch die kürzlich gelungene Herstellung von Eizell-ähnlichen Zellen aus embryonalen Stammzellen lösen. Ethisch problematisch ist, dass beim „Therapeutischen Klonen“ totipotente Stadien entstehen können, die nach Übertragung in eine Frau im geeigneten Zyklusstadium eventuell voll entwicklungsfähig sind. Insofern wäre die Grenze zwischen therapeutischem und reproduktivem Klonen nur durch die Intention, nicht aber den Vorgang per se definiert. Würde die Strategie „Therapeutisches Klonen“ tatsächlich möglich, so könnten männliche Patienten hinsichtlich der immunologischen Verträglichkeit benachteiligt sein: Mitochondrien, die energieerzeugenden Organellen in der Zelle, besitzen eine eigene DNA. Diese Erbinformation wird nur von der Mutter über deren Eizellen weitergegeben. Um Immunreaktionen auf Produkte dieser DNA zu vermeiden, sollte die für den Kerntransfer bestimmte Eizelle idealer Weise aus der erkrankten Person selbst gewonnen werden, was bei Männern ausgeschlossen ist. Als mögliche Quelle kämen weibliche Verwandte mütterlicherseits in Frage. Korrespondenz: Prof. Dr. Eckhard Wolf Lehrstuhl für Molekulare Tierzucht und Biotechnologie Genzentrum der Ludwig-Maximilians-Universität Feodor-Lynen-Strasse 25, 81377 München [email protected] Fäden des Lebens LMU 50 Jahre DNA - Doppelhelix GENZENTRUM Lehrstuhl für Molekulare Tierzucht und Biotechnologie Münchner Wissenschaftstage 16. - 20. Juli 2003 Epigenetische Mechanismen beim Kerntransfer Epigenetik – molekulares Gedächtnis zur Regelung spezifischer Zellfunktionen Die meisten der etwa 210 unterschiedlichen Zelltypen eines Säugerorganismus enthalten die selbe genetische Information und unterscheiden sich trotzdem stark in ihrer Morphologie und Funktion. Diese Unterschiedlichkeit hat ihre Ursache in der selektiven Expression beziehungsweise Repression von Genen. Epigenetische Mechanismen stellen eine Art molekulares Gedächtnis dar. So binden beispielsweise spezifische Proteinkomplexe an die DNA, wodurch stabile und an die Tochterzellen vererbbare Chromatinstrukturen gebildet werden, die eine zelltypspezifische Expression von Genen zulassen, während die Expression anderer Gene, die in einem spezifischen Zelltyp nicht aktiv sein dürfen, unterdrückt wird. Trotz dieser komplexen Mechanismen, die den Differenzierungsgrad einer Zelle regeln, zeigen die erfolgreich durchgeführten Kerntransferexperimente bei verschiedenen Spezies, dass Zellkerne – auch wenn sie aus hochspezialisierten Zellen kommen – durch den Transfer in eine entkernte Eizelle in den Status der Totipotenz versetzt werden können. Die Mechanismen, die diese außergewöhnliche Plastizität des Genoms ermöglichen, sind bislang nur wenig bekannt, ihre Erforschung wird aber maßgeblich dazu beitragen, Stammzellen für therapeutische Zwecke in einen geeigneten Differenzierungs- und Funktionszustand zu versetzen. Der Kerntransfer mit differenzierten Spenderzellen ist mittlerweile für die Spezies Schaf, Maus, Rind, Ziege, Schwein, Katze, Kaninchen und Pferd mehr oder weniger gut etabliert [1]. Um eine erfolgreiche Entwicklung der Kerntransferembryonen zu gewährleisten, muss eine weitgehende epigenetische Reprogrammierung des Spenderzellkerns ablaufen. Die dabei involvierten Mechanismen sind in Abb. 1 zusammengefasst. Veränderungen der globalen DNA-Methylierung in der Embryonalentwicklung sind bei Maus und Rind ähnlich Nach der Befruchtung kommt es zu einer schnellen Demethylierung der SpermienDNA, während die DNA der Eizelle langsamer demethyliert wird. Der niedrigste Methylierungsgrad ist bei der Maus im Morulastadium, beim Rind schon etwas früher erreicht. Im Stadium der Blastozyste sind dann wieder starke Methylierungssignale erkennbar (Abb. 3) (2). Maus Rind Abb. 3. Globale DNA-Methylierung in der frühen Embryonalentwicklung Bei Embryonen aus dem Kerntransfer kommt es zu Störungen der DNA- und Histon H3-Methylierung Bei vielen Kerntransferembryonen kommt es zu einer vorzeitigen Remethylierung der DNA (Abb. 4) (2), die häufig mit einer verstärkten Methylierung des Lysin 9 an Histonprotein H3 assoziiert ist (Abb. 5) (3). Somit treten bei zwei entscheidenden Mechanismen der epigenetischen Genregulation nach Kerntransfer regelmäßig Störungen auf. Kerntransfer Abb. 1. Epigenetische Mechanismen nach dem Kerntransfer Die Effizienz der Kerntransfertechnologie ist beim Rind am höchsten. Etwa 10-25% der auf Empfängertiere übertragenen Kerntransferembryonen können sich zu lebenden Nachkommen entwickeln. Abb. 2 zeigt einen klinisch gesunden Rinderklon, der in Kooperation mit der Biotechnologiefirma Agrobiogen GmbH, Hilgertshausen erzeugt wurde. Nichtsdestoweniger gibt es auch beim Rind relativ häufig Probleme bei Trächtigkeiten und Nachkommen aus dem Kerntransfer. Um die Ursachen dieser Störungen zu klären, haben wir in Kooperation mit der Arbeitsgruppe Wolf Reik in Cambridge Störungen der epigenetischen Reprogrammierung nach Kerntransfer in frühen Embryonalstadien untersucht. Abb. 4. Vorzeitige DNA-Remethylierung in Kerntransferembryonen 5-MeC IVP α-methH3-K9 IVP NT 5-MeC NT Abb. 5. Abnormale DNA- und K9-H3-Methylierung bei Kerntransferembryonen Abb. 2. Klinisch gesunder Rinderklon „Lara“ Literatur: (1) Shi et al., Differentiation 71, 91-113, 2003; (2) Dean et al., PNAS 98, 13734-13738, 2001; (3) Santos et al., Curr Biol 13, 1116-1121, 2003 Korrespondenz: Prof. Dr. Eckhard Wolf Lehrstuhl für Molekulare Tierzucht und Biotechnologie Genzentrum der Ludwig-Maximilians-Universität Feodor-Lynen-Strasse 25, 81377 München [email protected]