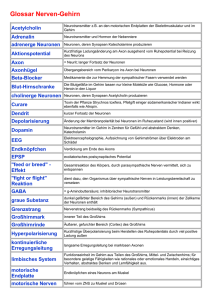
Geräuschlokalisation - Brain Modelling
Werbung

Titel: kopf90.eps Erstellt von: Canvas 3.0 Vorschau: Diese EPS-Grafik wurde nicht gespeichert mit einer enthaltenen Vorschau. Kommentar: Diese EPS-Grafik wird an einen PostScript-Drucker gedruckt, aber nicht an andere Druckertypen. BRAIN MODELLING II physikalische Modelle über das Gehirn Kurzfassung BRAIN MODELLING II physikalische Modelle für das Gehirn auf zellulärer Basis nach einer Vorlesung von W. Gruber im Jahr 2000/01 an der Universität WIEN, Institut für Experimentalphysik ACHTUNG: Dieses Skriptum ist die Kurzfassung. Im gesamten Kurs wird mehr auf die einzelnen Details eingegangen, als es in diesem Skriptum der Fall ist. Leider fehlen auch zwei Kapitel: Motorik und physikalische Untersuchungsmethoden. Ein vollständiges Skript steht ab spätestens Ende Juni im Internet unter http://brain.exp.univie.ac.at unter Unterlagen. 1.0 Geräuschlokalisation der Schleiereule 1 2.0 2.1 Das Riechsystem Exkurs Chaos 5 10 3.0 3.1 3.2 15 19 3.3 3.4 Aufbau des Gehirns Steuerung und Regelung Der Hypothalamus Regulation der Körpertemperatur Regulation des Körpergewichts Der Thalamus Die Großhirnrinde 4.0 EEG, MEG und PET 31 5.0 5.1 Sprache und Sprachverständnis Der Eliza-Effekt 47 53 6.0 Der Schlaf 57 7.0 7.1 7.2 7.3 Das Gedächtnis Das Arbeitsgedächtnis Das Gedächtnis Der Hippocampus 61 63 67 8.0 Neurotransmitter 71 9.0 Der kranke Geist 73 10.0 Künstliche Intelligenz 79 11.0 Synthetische Psychologie 81 12.0 Spieltheorie 83 Literaturverzeichnis 87 20 21 24 25 Geräuschlokalisation bei der Schleiereule Die Schleiereule ist in der Lage im Dunkeln sehr genau die Position von Geräuschen wahrzunehmen. Lange Zeit war nicht genau geklärt, wie sie die Position von Beutetieren auf 1-2° genau detektieren konnten. Ein Geräusch gelangt zu beiden Ohren zu unterschiedlichen Zeiten. Das Gehirn der Schleiereule vergleicht die Zeiten und setzt die ermittelten Unterschiede zu einem einheitlichen räumlichen Höreindruck um. Mit dem Laufzeitunterschied kann die Schleiereule in der x-y-Ebene bestimmen, wie weit das Geräusch von rechts (beziehungsweise von links) kommt. Da die beiden Ohrmuscheln unterschiedlich orientiert sind, können auch Geräusche von oben beziehungsweise von unten erkannt werden. So ist das linke Ohr empfindlicher für Geräusche von unten bzw. das recht Ohr von oben. Es werden die unterschiedlichen Laufzeiten und Intensitäten gemessen (siehe Abb.1.1). Die Laufzeiten sind für die Orientierung in der x-y Ebene und die Intensitäten für die Orientierung in der x-z Ebene verantwortlich. Man konnte dies sehr genau zeigen. Man setzte die Schleiereule in einen schalltoten Raum, verband ihr die Augen und über zwei Ohrenstöpseln wurden den Ohren Geräusche zugespielt. Man konnte die beiden Laufzeiten variieren und die Intensität. Bei geeigneter Wahl der Parameter bewegte die Schleiereule den Kopf in die Richtung des z vermuteten Geräusches. Dies wurde ebenso vermessen. Mit diesem Aufbau war es möglich, sehr exakt zu bestimmen, welche Gehirnareale für die y Geräuschlokalisation verantwortlich sind. So konnte man in der Hörregion im Mittelhirn das wesentliche x Areal identifizieren. Dieses Areal hat rechts 40° 20° unterschiedliche Bereiche, wobei jedes für einen 10° bestimmten Winkelbereich verantwortlich ist. Wenn 0° 10° ein Geräusch von 20° rechts kommt, dann werden links die Neuronen in diesem Areal sehr stark feuern, Hörregion im linken Mittelhirn während es in den übrigen Bereichen nur zu Abbildung 1.1: Der Hör-Raum und die spontanen Entladungen kommt. Ostsspezifische dazugehörigen Rindenareale Neuronen der Hörregion des Mittelhirns decken jeweils einen bestimmten Bereich des Hörraums ab. ___________________________________________________________________________________ 1 Brain Modelling Die Entfernungsbestimmung von Geräuschen ist weiterhin nicht geklärt ! Die Neuronen auf der Hörrinde erhalten ihren Input über je einen Relaiskern von den beiden Ohren. Im Gehirn werden die unterschiedlichen Laufzeiten durch Verzögerungsstrecken kompensiert. Alle Signale vom Ohr gelangen über diese Verzögerungsstrecken zu den "ortsabhängigen" Rindenarealen. Wenn nun die beiden Signale (vom rechten und vom linken Ohr) über die Verzögerungsstrecken in das Rindenareal gelangt, dann gibt es genau zwei Strecken, die die unterschiedlichen Zeiten kompensiert, das heißt in einer Neuronengruppe kommen die Signale zeitgleich an, während die Signale die über andere Verzögerungsstrecken in die anderen Areale gelangen zu unterschiedlichen Zeiten (vom rechten und vom linken Ohr) eintreffen (siehe Abbildung 1.2). Die Gruppen von Nervenzellen feuern nur dann, wenn die von den beiden Seiten stammenden Signale gleichzeitig bei ihr über verschiedene Verzögerungsstrecken eintreffen. Die Neuronen arbeiten als Koinzidenzdetektor (Vergleiche Synchronisation). Verlagert sich die Schallquelle von der Mitte zur Seite, wird jene Gruppe aktiv, dessen Verzögerungsstrecken die unterschiedlichen Laufzeiten gerade kompensieren. Die Verzögerungsstrecken sind durch unterschiedlich lange Axone zwischen dem jeweiligen Relaiskern und der Hörrinde realisiert. Geräusch a b a b rechtes Ohr linkes Ohr d d Relaisstation c c Gruppe von Neuronen, die als Koinzidenzdetektoren arbeiten Abbildung 1.2: Ein Geräusch wird von den beiden Ohren detektiert. Wenn sich das Geräusch rechts befindet, dann ist der Weg zum rechten Ohr kürzer, als zum Linken. Durch die Verzögerungsstrecken wird dieser Weglängenunterschied ausgeglichen. Für die Laufzeiten in der Luft a und b und für die Laufzeiten in den Neuronen (über die Axone) c und d gilt: a + c = b + d. Nur die Gruppe, für die diese Bedingung gilt, wird aktiviert. Verzögerungsstrecke (Axone) Eine Schallwelle einer einzigen Frequenz veranlasst bestimmte dafür empfindliche Neuronen zu feuern. Dabei werden Impulse mit einem bestimmten Phasenwinkel erzeugt. Die Neuronen in der Hörrinde des Eulengehirns reagieren am stärksten, wenn Impulse mit demselben Phasenwinkel, also gleichzeitig bei ihnen eintreffen. Auf leicht asynchron eintreffende Impulse können Sie ebenfalls, wenn auch schwächer reagieren. ___________________________________________________________________________________ Die Schallortung der Schleiereule 2 Verzögerungsstrecke Neuron des rechten Ohrs t Neuron des linken Ohrs Maximale Koinzidenz: Die Neuronen von beiden Ohren feuern gleichzeitig. Ortsspezifische Gruppe feuert stark Neuron des rechten Ohrs t Neuron des linken Ohrs Keine Koinzidenz: Die Neuronen von beiden Ohren feuern NICHT gleichzeitig. Ortsspezifische Gruppe feuert schwach Verzögerungsstrecke Abbildung 1.3: Nur wenn die Verzögerungsstrecken richtig gewählt wurden, dann erst kann es zu einer Koinzidenz kommen. Ein Neuron feuert dann, wenn gleichzeitig ausreichend Signale (EPSP´s) zum Axonhügel gelangen (siehe Bedingung für Synchronisation). Mit diesen Fakten kann man nun sehr genau ausrechnen, wie genau Schleiereulen den Entstehungsort von Geräuschen bestimmen können. Bei näherem Hinsehen ergibt sich dabei ein Problem. Die Geschwindigkeit der Reizweiterleitung entlang eine Neurons kann als konstant angesehen werden. Wesentlich ist auch die Größe und Länge der jeweiligen EPSP´s die durchschnittlich 200 20 µs lang sind. Die aktiven Neuronen in den Ohren feuern rund alle 200 40 s. Das bedeutet, dass sich ein relativ großer Fehler entsteht. Trotzdem können Schleiereulen den Entstehungsort von Geräuschen auf 1-2° genau bestimmen. Das bedeutet, es müssen unterschiedliche Laufzeiten (zwischen dem rechten und dem linken Ohr) von rund 5 s erkannt werden. Mit dem Wissen über den neuralen Aufbau lässt sich dies nur schwer klären. Man fand 3 wesentliche Schlüsselprozesse die dieses Paradoxon klären können. Diese Prozesse wurden am Computer modelliert und später mit den tatsächlichen Gegebenheiten verglichen. [1] Herstellung der Genauigkeit durch Auswahl der Synapsen während der Entwicklung. Die Computersimulation zeigte, dass von 600 Synapsen rund 100 überlebten. Nur die Synapsen, die die Information exakt weitergegeben haben, überlebten. Die Genauigkeit der Laufzeitmessung stieg dabei stark an. (Vergleiche Entwicklung des Nervensystems) ___________________________________________________________________________________ 3 Brain Modelling Aktivierung eines exzitatorischen Neurons [2] "Subthreshold Oscillations": Hemmenden Neuronen verursachen eine Schwelle Art Trägerwelle, die eine wesentlich höhere Frequenz besitzt. Die inhibitorischen Neuronen feuern rund ein Drittel öfters, als die exzitatorischen. Das Hintergrundrauschen kann nicht einfach ein -75 t Aktionspotential auslösen - erst nach- [mV] Die hemmenden Neuronen dem sich die Aktivierung öfters der feuern gleichzeitig. Schwelle angenähert hat. Wenn nun die Aktivierung im Ruhezustand (aufgrund Abbildung 1.4: Die Oszillationen unterhalb der Schwelle, erzeugt durch inhibitorische Neuronen des Hintergrundrauschens) nahe der Schwelle ist, dann reichen sehr wenige EPSP´s aus, um das Neuron zum Feuern zu bringen. Aufgrund dieser Trägerwelle können die exzitatorischen Neuronen besser aufeinander abgestimmt werden. Dieser Prozess unterstützt das phase-locking, beziehungsweise die Synchronisation von Neuronen im Gehirn. [3] Die Genauigkeit der Laufzeitmessung ist auch von der Zahl der aktiv beteiligten Neuronen abhängig. Je mehr Neuronen beteiligt sind, umso wahrscheinlicher ist es, dass ein paar aufgrund des Hintergrundrauschens leichter aktivierbar sind. Das Argument der "subthreshold-oscillations" ist von der Neuronenzahl abhängig. Genauigk eit 10 1 101 102 Anzahl der Neuronen 103 Abbildung 1.5: Durch die Computersimulation könnte die Anzahl der Neuronen im Koinzidenzgebiet bestimmt werden. ___________________________________________________________________________________ Die Schallortung der Schleiereule 4 Das Geruchssystem Gerüchen - außer, dass sie angenehm oder widerlich sind - wird eher wenig Bedeutung zugeordnet. Allerdings ist für viele Säugetiere der Geruchssinn der wesentlichste sensorische Input, der auch der sozialen Kommunikation dient. So wird der Geruchssinn benötigt, um verdorbene Nahrung, um Feinde oder auch um Sexualpartner zu identifizieren. Ein Mensch kann rund 10 000 verschiedene Valeurs (Gerüche) wahrnehmen. Tiere können wahrscheinlich auch nicht mehr Gerüche unterscheiden, aber für viele Tiere reicht eine geringere Konzentration des Duftstoffes für die Erkennung aus, da es mehr Rezeptoren, aber nicht mehr Rezeptortypen gibt. Das heißt manche Tiere können Gerüche mit einer geringeren Konzentration identifizieren. Die Geruchsstoffe gelangen über die Nasenhöhle zur Riechschleimhaut. Beim Menschen ist die Riechschleimhaut eine rund 5cm2 große gelbliche Fläche. Die Riechschleimhaut ist von der Nasenschleimhaut umgeben, die die Atemluft erwärmt und befeuchtet. In der Riechschleimhaut befinden sich Millionen von Sinneszellen. Diese Sinneszellen geben ihre Information über Axone, die durch das Siebbein ziehen, an die Glomeruli weiter. Die Glomeruli sind knäuelartige Strukturen, über die die gesamte Geruchsinformation weiterverarbeitet wird. Da die Riechzellen direkt mit der Außenwelt in Kontakt stehen, können sie auch leicht beschädigt werden - sie sterben ab. Sie werden durch neurale Stammzellen ersetzt. ACHTUNG: Bei momentanem Stand des Wissens - März 2002 sind die Riechzellen die einzigen Zellen des Nervensystems, Abbildung 2.1: Anatomischer Aufbau des Riechsystems und die vollständig durch neurale die Verbindungen zu wesentlichen verarbeitenden Teilen des Stammzellen ersetzt werden können. übrigen Gehirns. Die Glomeruli stellen die erste ___________________________________________________________________________________ 5 Brain Modelling Verarbeitungsstation in der Geruchswahrnehmung dar. Von dort gelangen über drei verschiedene Riechstränge die Informationen in andere Gehirnteile, beziehungsweise in das Riechhirn. Die Riechzellen besitzen einen Rezeptor, deren Aminosäurekette räumlich so gefaltet ist, dass sie die Zellmembran sieben mal durchquert. Damit stellt sich die Frage, wie viele Gene für die Sinneszellen notwendig sind. Gibt es für jeden Geruch eine spezielle Aminosäuresequenz, dann würden rund 10 000 Gene benötigt, oder sind es bedeutend weniger Aminosäurenketten und Gerüche entstehen aus einer Überlagerung der jeweiligen Aktivierungen. Die Analyse an Säugetieren ergab, dass rund 1000 Gene für die Geruchsrezeptoren verantwortlich sind. Da der Mensch rund 100 000 Gene besitzt, werden für die Geruchsinformation rund 1% des Genpotentials verwendet. Im Vergleich, das Farbsehen benötigt nur 3 verschiedene Rezeptoren. Die Gerüche entstehen nun dadurch, dass einzelne Geruchsrezeptoren aktiviert werden und sich die Wahrnehmung aus der Aktivierung der einzelnen Rezeptoren zusammensetzt. Jedes Geruchsmolekül hat verschiedene Seiten, mit denen es an die Rezeptoren andocken kann. Es gäbe damit rund 21000 Gerüche, die wahrgenommen werden könnten. Aus praktischen Gründen können nicht alle Kombinationen auftreten, beziehungsweise gibt es diese Gerüche nicht in der Natur. Jeder dieser Rezeptoren schickt sein Axon zu einem (bzw. zwei) speziellen Glomerulus. Somit registriert jeder Glomerulus eine spezifische Komponente des Geruchs. Es existieren rund 2000 Glomeruli - für jede Geruchskomponente gibt es zwei Glomeruli. Wenn einer ausfällt, gibt es noch einen "Reserve"-Glomerulus. Die Glomeruli liegen immer an derselben Stelle, allerdings sind die Riechzellen selbst über die ganze Riechschleimhaut zufällig verteilt. Die Position der Glomeruli scheint genetisch determiniert zu sein. Glomeruli Mitralzellen zur Riechrinde Siebbein Sinneszellen Geruch Abbildung 2.2: Aufbau der Riechschleimhaut und des Bulbus olfaktorius. Jede Sinneszelle innerviert - in Abhängigkeit des Typs - einspezifisches Glomeruli. Über die Mitralzellen wird dann die Information an andere Bereiche weitergeleitet. ___________________________________________________________________________________ Das Geruchssystem 6 Wenn nun Geruchsmoleküle auf die Riechzellen treffen, wird ein elektrisches Signal über die jeweiligen Axone zu den spezifischen Glomeruli weitergeleitet. Bei komplexen Gerüchen entsteht somit ein geometrisches Muster. Für jeden Geruch gibt es ein spezifisches geometrisches Muster an aktiven Glomeruli. Dieses Muster kann über elektrische Ableitungen gemessen werden. Die Glomeruli selbst werden von Mitralzellen innerviert, die die Information an die Riechrinde weiterleiten. Der Komplex der Glomeruli und der Mitralzellen wird als Riechkolben (Bulbus olfaktorius) bezeichnet. Über die periglomulären Zellen, die zwischen den Glomeruli liegen, kommt es zu synaptischen Hemmungen. Das heißt, wenn in einem Glomerulus die Synapsen besonders aktiv sind, dann hemmt diese Zelle die Synapsen in den benachbarten Glomeruli. Der Kontrast des räumlichen Musters wird verstärkt. Zusätzlich werden die Mitralzellen durch Körnerzellen inhibiert. Dadurch kann es zu keiner längerandauernden Synchronisation kommen und die Trennschärfe wird ebenso wie durch die periglomulären Zellen erhöht. Die Axone der Mitralzellen bilden drei Stränge. Der mediale Riechstrang gleicht die Information mit dem gegenüberliegendem Reichkolben ab. Der intermediale Riechstrang innerviert den Stirnlappen und den Thalamus. Über die Amygdala zieht der laterale Riechstrang zum Riechkortex. Alleine aus diesen Verbindungen zeigt sich die Wichtigkeit der Geruchsinformation für das Gehirn. In der Amygdala werden Informationen emotionell bewertet, während in den Stirnlappen Entscheidungen getroffen werden. Der Riechkolben kann durch unterschiedliche Einflüsse sensibilisiert werden: Hunger, Durst, Sex, Bedrohung und so weiter. Die Geruchswahrnehmung erlangt in diesen Situationen eine erhöhte Bedeutung. Die Sensibilisierung kommt dadurch zustande, dass eine Synchronisation erleichtert wird - die hemmenden Zellen zwischen den Glomeruli und den Mitralzellen haben eine geringere Aktivität beziehungsweise die Mitralzellen bekommen einen zusätzlichen Input von anderen Gehirnregionen. Man kann EEG-Messungen in der Riechrinde durchführen. Meist wird dabei die Aktivität von vielen Neuronen - einigen Hundert - gleichzeitig gemessen. Auf der Riechrinde können mehrere Elektroden befestigt werden und damit ist es möglich den Aktivierungszustand über den gesamten "Riechraum" - der Oberfläche der Riechrinde - gleichzeitig zu messen. Durch diese EEG-Kurven konnte gezeigt werden, dass auch in diesem sensorischen Areal Synchronisation der entscheidende Faktor ist. Atmet ein Tier einen vertrauten Geruch ein, so beobachtet man eine Salve: Die gemessenen EEG-Wellen werden für wenige Schwingungsperioden plötzlich regelmäßig(er) und geordnet(er). Bei unterschiedlichen Duftstoffen variiert das Muster der räumlichen Korrelation und Synchronisation der einzelnen Neuronen. Dadurch lässt sich ein Duftstoff eindeutig zuordnen. Abbildung 2.3: Links sind die Ableitungen einzelner Elektroden gezeigt. Man erkennt sehr schön, dass in einzelnen Bereichen die Amplituden erhöht sind, und dass alle Neuronen praktisch Synchron sind. Durch diese Einzelableitungen können sogenannte "Höhenbilder" konstruiert werden. Auf ihnen erkennt man die Bereiche, die gleichzeitig aktiv sind, leichter (rechts). In einer zweidimensionalen Karte der elektrischen Aktivitäten des Riechkolbens kann man aber auch die Veränderung bei unterschiedlichen Gerüchen gut erkennen. So ergeben sich für ___________________________________________________________________________________ 7 Brain Modelling unterschiedliche Gerüche unterschiedliche Muster. Wird nun ein neuer Geruch hinzugelernt, so verändert sich auch das Bild bei den bekannten Gerüchen. Sägemehl Banane Sägemehl Abbildung 2.4: Ein Versuchstier schnupperte an Sägemehl und das linke Muster wurde gemessen. Wenn das Tier mit einem neuen Geruch zum Beispiel dem Bananengeruch in Kontakt kam (mittlere Abbildung), so veränderte sich bei einem späteren Experiment die Synchronisation der Neuronen in der Riechrinde für Sägemehl. Ein neuer Geruch wurde gelernt (rechte Abbildung). Diese biologischen Fakten, kombiniert mit den elektrischen Ableitungen wurden in einer Vielzahl von Modellen untersucht. Zwei Modelle sollten besonders erwähnt werden. Zum einen das Modell von Rall und Shepherd. Die einzelnen Neuronen wurden als Compartment beschrieben. Dadurch konnten sie sehr genau den Potentialverlauf der Mitralzellen und Körnerzellen im Riechkolben beschreiben. Das andere Modell stammt von W.J.Freeman. Dieses Modell ist sehr bekannt und man kann an diesem Modell sehr viel über das Modellieren lernen. Die Zellen wurden durch einzelne Funktionen beschrieben, diese Zellen wurden zu Sets zusammengefasst, die dann wiederum zu Netzwerken zusammengefasst wurden. Betrachten wir zuerst die beiden Funktionen, die die Neuronen beschreiben. 1) Pulse-to wave Transformation: Ein Aktionspotential wird in ein wellenförmiges Signal umgewandelt (dieses Signal entspricht den PSP´s). 2) Wave-to pulse Transformation: Die wellenförmige Signale werden in Aktionspotential umgewandelt. Im Prinzip beschreiben die beiden Funktionen die Hodgkin-Huxley-Gleichung. Die Neuronen können nun verschiedene Zustände besitzen: stabiler Ruhezustand (zwischen Ruhemembranpotential und dem Schwellpotential) a) Ruhegleichgewicht (das Potential kehrt zum Ruhemembranpotential zurück, ohne dass das PSP einen Einfluss auf das Feuerverhalten hat). b) Stabiles Null-Gleichgewicht (die einlangenden Signale sind so groß, dass das Potential über dem Ruhemembranpotential gleich bleibt). stabiler Zustand der Pulserzeugung (die Schwellwertspannung wurde überschritten) c) Stabiles Nicht-Null-Gleichgewicht (die mittlere Feuerrate über einen längeren Zeitraum ist konstant, aber die einzelnen Aktionspotentiale sind unvorhersehbar). d) Stabiler Grenzzyklusbereich (Feuersalven sind von Ruhepausen unterbrochen). ___________________________________________________________________________________ Das Geruchssystem 8 Diese unterschiedlichen Neuronen werden nun zu einzelnen Sets zusammengefasst. Man unterscheidet: K0-Set: Alle Neuronen arbeiten gleich. Entweder wirken alle erregend K0e oder inhibitorisch K0i. Alle Zelle erhalten denselben Input und sie sind nicht untereinander verbunden. KI-Set: Alle Neuronen haben die selbe Wirkung (erregend KIe oder hemmend KIi), sie erhalten den selben Input, aber sie sind untereinander dicht verknüpft. KII-Set: Die Neuronen sind erregend oder hemmend und können erregend oder hemmend wechselwirken. Die Neuronen sind untereinander verknüpft und sie erhalten unterschiedlichen Input. Die Sets stellen ein System von gewöhnlichen gekoppelten nichtlinearen Differentialgleichungen 2. Ordnung mit sehr vielen Parametern und Variablen dar. Und damit sind wir bei der Problematik dieses Modells. Es ist ein sogenanntes Zahnradmodell. Wenn man alle Zahnräder richtig einstellt - also alle Parameter richtig wählt - dann kommen die richtigen Zahlenwerte heraus. Das wäre für ein normales Modell durchaus sinnvoll. Wenn aber die Anzahl der Parameter einen bestimmten Bereich überschreitet, dann kann man mit ein paar Gleichungen so ziemlich alles beschreiben - wenn die Gleichungen hinreichend komplex sind. Ein gutes Modell zeichnet sich dadurch aus, dass es mit wenigen Parametern eine gute Beschreibung der Wirklichkeit liefert. Die Parameter sollten plausibel sein und die Zusammenhänge zwischen den einzelnen Parameter sollten durchschaubar und logisch sein. Leider gibt es immer wieder Modelle, die die Hodgkin-Huxley-Gleichung als Grundlage verwenden. Aber zur Erinnerung: diese Gleichung beschreibt die elektrischen Potentiale entlang eines Axons, wenn die Schwelle überschritten wird. Diese Gleichung beschreibt sonst NICHTS. Natürlich ist es einfach Hodgkin-Huxley-Gleichungen herzunehmen, sie zu koppeln, und nach Lösungen zu suchen. Aber über die Axone kommt es zu KEINER Synchronisation. Natürlich ist es wichtig, dass Signale vom Axonhügel zu den einzelnen Synapsen weitergeleitet werden, aber im Prinzip ist es egal wie das Signal aussieht - vorausgesetzt das EPSP beziehungsweise das IPSP verändert seine Amplitude oder zeitlichen Verlauf nicht. Vielmehr ist das entstehen eines Aktionspotentials am Axonhügel viel wichtiger. Dort wird nach dem "Alles-oder-Nichts"-Prinzip entschieden, ob ein Signal zu den Synapsen geschickt wird. Für die Synchronisation ist die Verarbeitung zwischen dem Ruhemembranpotential und der Schwellwertspannung (nichtlineare Funktion) und der Summe der EPSP´s und IPSP´s wichtig. Die Aktionspotentiale selbst sind in Folge natürlich auch synchron - aber als Resultat und nicht als Ursache. Bei komplizierten Differentialgleichungen gibt es noch ein anderes Problem. Es entstehen automatisch Chaos-Effekte. In den 80ern und Anfang der 90er war es modern alles nach Chaoseffekten zu untersuchen. Und natürlich hat man im EEG der Großhirnrinde, als auch in der Riechrinde Hinweise für Chaos entdeckt. Leider musste man später erkenne, dass diese Ergebnisse voreilig waren. Durch die sensiblen Verstärkerschaltungen kam es zu Rückkopplungen, die die Messergebnisse verfälschten. Nachdem man diese Schwachstellen beseitigt hatte, blieb von den Hinweisen für Chaos nicht mehr viel übrig. Bei der Untersuchung einzelner Neuronen konnte man aber dennoch chaotische Effekte beobachten. Wenn Einzelobjekte eines Systems ein chaotisches Verhalten zeigen, so kann man nicht auf ein chaotisches Verhalten des gesamten Systems schließen. Durch Synchronisationseffekte wird das Chaos gezähmt. Da die Frage nach Chaos im Gehirn immer noch eine wichtige Rolle spielt sollte dieses Gebiet näher erläutert werden, denn Chaos ist nicht der Ordnungszustand eines Schreibtisches, sondern etwas viel komplexeres. ___________________________________________________________________________________ 9 Brain Modelling ____________________________________________________________________________ Exkurs: Was ist Chaos ? “Unvorhersagbare "zufällige" Phänomene können entstehen wenn in einem System kleine Änderungen in der Gegenwart große Änderungen in der Zukunft hervorrufen” Henri Poincaré Betrachten wir ein ideales Fadenpendel. Wenn wir es loslassen, dann schwingt es hin und her. Es wird nicht abgebremst, da es als ideales Pendel keinen Luftwiderstand besitzt. Durch die Schwerkraft wird die Kugel beschleunigt, wenn wir loslassen, und wenn die Kugel den untersten Punkt erreicht hat, dann hat sie die höchste Geschwindigkeit, die wieder verringert wird, bis die Kugel den gegenüberliegenden Scheitelpunkt erreicht hat. Wir können den Ausschlag (die Amplitude) gegen die Zeit auftragen. Dies ergibt eine Sinusschwingung, wie sie vielen bekannt ist. Für manche Effekte ist es aber sinnvoller Zustandsgrößen gegeneinander aufzutragen. Die Zeit selbst ist keine Zustandsgröße. Zustandsgrößen beschreiben ein System vollständig und sie hängen von der Zeit ab. Mehrere Abbildung 2.5: Ein ideales Zustandsgrößen spannen einen Zustandsraum auf. Der Fadenpendel. Zustandsraum ist eine mehrdimensionale Darstellung (meist zwei oder dreidimensional) des Zusammenhangs von Zustandsgrößen. Zum Beispiel wird der Zustandsraum eines Fadenpendels durch den Ort x und die Geschwindigkeit v aufgespannt. Ort Amplitude Zeit Geschwindigkeit hohe Geschwindigkeit Abbildung 2.6: Links ist die Amplitude gegen die Zeit aufgetragen, während in der rechten Darstellung der Ort gegen die Geschwindigkeit aufgetragen wird. Der Ort und die Geschwindigkeit stellen die Zustandsgrößen dar. Man erkennt leicht die Bereiche, in denen das Pendel sich rasch beziehungsweise sich langsam bewegt. Betrachten wir ein reales Fadenpendel. Durch den Luftwiderstand wird das Pendel gebremst und nach einiger Zeit wird es zum Stillstand kommen. Es ist ein gedämpfte Oszillation gegeben. Der Endzustand ist immer derselbe - egal von wo aus wir das Pendel loslassen. Es wird immer an der selben Stelle stehen bleiben. Dieser Punkt ist ein Fixpunkt. Nach endlicher Zeit wird er erreicht. Beim Pendel mit Reibung ist der Weg im Zustandsraum eine Spirale (siehe Abbildung 2.7). Durch die Reibung verliert das System Energie und die Geschwindigkeit nimmt ab bis das Pendel stehen bleibt (v = 0). Der Weg wird als Trajektorie (gestrichelte Linie, Abbildung 2.77) bezeichnet, während der Endzustand als Attraktor oder auch als Fixpunkt bezeichnet wird. Ein Fixpunkt ist die einfachste Form eines Attraktors. ___________________________________________________________________________________ Das Geruchssystem 10 Ort Amplitude Trajektorie Zeit Geschwindigkeit Fixpunkt - Attraktor Abbildung 2.7: Ein reales Fadenpendel (links), das ein Beispiel für eine gedämpfte Schwingung darstellt (Mitte), während man im Zustandsdiagramm den Fixpunkt leicht erkennt. Im Zustandsraum kann man leicht erkennen ob das System sich auf einen Fixpunkt zubewegt oder nicht. Wenn ein Fixpunkt gegeben ist, dann ist ein System in der Regel nicht chaotisch, denn egal unter welchen Bedingungen wir starten, das Endresultat ist das selbe. Bei einer Pendeluhr wird extern Energie zugeführt, um die Reibungsenergie auszugleichen. Das System wird im Endzustand periodisch verschiedene Zustände annehmen. Das Pendel wiederholt die Bewegung immer wieder gleichmäßig. Der Attraktor (Endzustand des Systems) ist ein Grenzorbit Das System nimmt einen stabilen Zustand ein, allerdings verändert sich der Zustand mit der Zeit auf eine berechenbare Weise. Der Attraktor wird nach endlicher Zeit durchlaufen und er verändert sich im Laufe der Zeit nicht. Das heißt das System ist stabil, wenngleich sich einzelne Parameter kontinuierlich verändern. Jetzt strebt das System nicht auf einen Punkt (Fixpunkt) hin, sondern es strebt auf eine Grenzorbit zu. Wenn dieser erreicht ist, dann bleibt das System auf diesem Attraktor. Abbildung 2.8: Das Zustandsdiagramm einer Pendeluhr. Der Attraktor ist dick eingezeichnet. ACHTUNG: Ein Attraktor selbst ist noch nicht chaotisch. Nur der "seltsame" Attraktor zeigt chaotische Effekte. Die Naturwissenschaften versuchen einen Zusammenhang zwischen Ursache und Wirkung herzustellen. Die Natur soll berechenbar werden (Naturgesetze). Naturphänomene deren Ursache-Wirkungs-Beziehung unbekannt sind bezeichnet man als stochastisch oder zufällig. Diese Effekte (Wetter) lassen nur eine Wahrscheinlichkeitsaussage zu (Wettervorhersage). Man vermutete lange Zeit, dass man aufgrund ausreichend genauer und zahlreicher Messwerte das Wetter berechnen kann. Es existiert zumindest eine prinzipielle exakte Vorhersagbarkeit. Es zeigte sich aber, dass sogar einfache deterministische Systeme stochastisches Verhalten zeigen können. Auch wenn wir mehr Information über das System sammeln, verschwindet das Zufallsverhalten nicht. Man bezeichnet ein solches scheinbares Zufallsverhalten als deterministisches Chaos. Dieses scheinbare Zufallsverhalten hängt primär von kleinsten Störgrößen ab. Manche physikalische Systeme reagieren sehr empfindlich auf externe Störungen. Zum Beispiel ein Bleistift, der perfekt ausbalanciert ist und mit seiner Spitze auf der Tischplatte steht, ist empfindlich für kleinste Störungen der Tischplatte. Der Bleistift wird bei der kleinsten Störung umfallen und wir wissen nicht in welche Richtung er sich bewegen wird. Danach nimmt er eine stabile Lage ein und das Systemverhalten ist eindeutig bestimmt. Bei chaotischen Systemen ist die Unbestimmtheit zu jedem Zeitpunkt gegeben, das heißt, winzig kleine Abweichungen der Messgröße wachsen sehr schnell. Das Systemverhalten reagiert auf die Abweichungen mit einem exponentiellen Anwachsen des Fehlers. Jeder noch so kleine ___________________________________________________________________________________ 11 Brain Modelling Fehler erreicht rasch makroskopische Dimensionen. Aufgrund der Messungenauigkeit ist es leider nicht möglich, diese winzigen Anfangsbedingungen "exakt" zu bestimmen. Ein interessantes Beispiel dafür ist die logistische Gleichung: xn+1 = r . xn . (xn-1). Man wählt für xn einen Wert zwischen Null und Eins. Danach rechnet man den rechten Teil aus und erhält damit xn+1. Dieser Wert wird aufgetragen und er dient als neues x n. Wenn der Kontrollparameter r kleiner als 2 ist, dann streben die xn gegen einen Fixpunkt. Für die verschiedenen Werte von r erhält man unterschiedliche Fixpunkte. Wird r weiter erhöht, dann gibt es eine Bifurkation und die xn streben auf 2 Fixpunkte hin, die dann abwechselnd erreicht werden. Wenn der Kontrollparameter r den Wert 4 besitzt, dann führen kleinste Änderungen des Startwertes dazu, dass die einzelnen Iterationen in sehr kurzer Zeit stark voneinander abweichen - die Gleichung reagiert chaotisch (siehe Abbildung 2.9). 1.0 1.0 0.5 0.5 0 0 Startwert x0=0.40000 Startwert x 0=0.40001 Abbildung 2.9: Die Iterationen nach der logistischen Gleichung mit leicht unterschiedlichen Werten. Nach einigen Iterationen kann man makroskopische Unterschiede erkennen. Die logistische Gleichung ist ein rückgekoppeltes System. Das bedeutet, dass der alte Ausgangswert den neue Eingangswert darstellt. Edward N. Lorenz entdeckte 1963 eine eigene Klasse von Attraktoren: den chaotischen oder auch (wie er ihn nannte) seltsamen Attraktor. Ein chaotischer Attraktor ist ein kompaktes Gebilde eines rückgekoppelten Systems, indem sich kleinste Unterschiede in den Anfangsbedingungen aufschaukeln. A' A' A A B' B B B' Abbildung 2.10: Wenn man von zwei unterschiedlichen Startpositionen A und B beginnt, endet man nach einem kurzen Stück in A' und B'. Bei rückgekoppelten Systemen stellt die Strecke A' und B' den neuen Anfangswert dar. Wenn die Entfernung zwischen A und B den größten maximalen Unterschied der Anfangsbedingungen dargestellt hatten, dann muss der Endbereich A' und B' gefaltet werden, damit die Endwerte in die Anfangswerte "passen". ___________________________________________________________________________________ Das Geruchssystem 12 Kleine Abweichungen verursachen ein Auseinanderstreben der Bahnkurven (siehe Abb. 2.10). Damit die Bahnkurven aber begrenzt bleiben, müssen sie wieder auf sich selbst zurückgeführt werden. Mathematisch entspricht dies einer Faltung. Man bekommt dann einen chaotischen Attraktor (siehe Abbildung 2.11). Startet man mit zwei benachbarten Punkten auf einem solchen Attraktor, dann wird der Abstand zwischen diesen beiden Punkten immer größer bis es unmöglich ist Vorhersagen zu machen. Der Endzustand der beiden Punkte könnte irgendwo auf dem chaotischen Attraktor liegen. Bei einem vorhersagbaren - nicht chaotischen - Attraktor bleiben die beiden Punkte benachbart. Abbildung 2.11: Der Rössler-Attraktor Ein chaotischer Attraktor beschreibt NICHT zufällige Ereignisse (Roulett). Reagiert ein System zufällig, dann gibt es keine mathematische Formel, die diesen Weg im Zustandsraum beschreibt. Bei echt zufälligen Prozessen wird der gesamte Zustandsraum ausgefüllt. Bei chaotischen Phänomen handelt es sich um hochkomplexes Verhalten, das zufällig erscheint, tatsächlich jedoch eine versteckte Ordnung aufweist. Es existiert eine Sensibilität bezüglich der Anfangsbedingungen (Nicht periodisches oder quasiperiodisches Verhalten, KEIN weißes Rauschen). _____________________________________________________________EXKURS ENDE Wenn man die Messdaten der EEG-Kurven mit (x = Un und y = Un+1), wobei Un und Un+1 die Spannungen mit einem kleinen Zeitunterschied sind, graphisch darstellt, dann kann man einiges über die Messdaten (qualitativ) aussagen. Wenn das ganze Gebiet gleichmäßig mit Messwerten versehen ist, dann sind die Messkurven das Resultat des Zufalls. Anhand der Regelmäßigkeit der Darstellung können Aussagen über das EEG gemacht werden. Zum Beispiel würde eine Sinusschwingung der Messkurve im Phasenporträt einen Kreis ergeben. Abbildung 2.12: Links ist der Attraktor eines bekannten Geruchsmusters und rechts der Attraktor eines unbekannten Geruches dargestellt. Bei der Analyse der EEG-Daten im Zustandsraum der Riechrinde ergaben sich scheinbar chaotische Attraktoren. Wenn kein Geruch wahrgenommen wird (Abb. 2.12, rechts) dann feuern die einzelnen Neuronen scheinbar zufällig, während sich bei einer Geruchswahrnehmung ein chaotischer Attraktor bildet (Abb. 2.12, links). Allerdings muss man mit der ___________________________________________________________________________________ 13 Brain Modelling Interpretation der Messdaten vorsichtig sein. Ein solches Muster erhält man auch, wenn ein sich regelmäßig wiederholendes Muster (synchron feuernde Neuronen) mit ein paar zufällig feuernden Neuronen überlagert wird. Der verrauschte Grenzzyklus würde wie ein chaotischer Attraktor aussehen. Die Synchronisation scheint eher Chaos-Effekte zu verhindern. Wenn die stabilen geometrischen Muster von synchron feuernden Neuronen stark von den Anfangsbedingungen abhängen, wäre ein INPUT-OUTPUT Korrelation nicht mehr gegeben. Wenn wir ein rotes Dreieck unter ungünstigen Bedingungen sehen (das Bild ist verrauscht), dann müssten Chaoseffekte dazuführen, dass ein anderes Bild wahrgenommen wird. Tatsächlich sind wir aber in der Lage auch ein "vernebeltes" rotes Dreieck als ein solches zu erkennen. Die Mustervervollständigung ist ein Beispiel für anti-chaotische Effekte. Allerdings ist es möglich, dass wenn zwei sich widersprechende Inputs in ein neurales Assemble gelangen, muss das Netzwerk eine Entscheidung treffen. Diese Entscheidung dürfte vermutlich von der Vorgeschichte des Netzwerkes abhängen. Hier kann sehr wohl Chaos auftreten - aber wie oft trifft dies in der Realität zu (siehe Gestaltpsychologie). ___________________________________________________________________________________ Das Geruchssystem 14 Aufbau des Gehirns Das menschliche Gehirn kann funktionell, anatomisch und zytologisch in verschiedene Gebiete unterteilt werden. Wesentliche Gebiete: das verlängerte Rückenmark (Myelencephalon) das Hinterhirn auch Rautenhirn (Metencephalon oder Rhombencephalon) das Mittelhirn (Mesencephalon) das Zwischenhirn (Diencephalon) das Endhirn (Telencephalon) Abbildung 3.1: Das Gehirn lässt sich in verschieden Bereiche unterscheiden. In der Darstellung links sind auch die 4 Gehirnventrikel im Querschnitt gut erkennbar. Das Myelencephalon oder auch das verlängerte Rückenmark hat die Aufgabe Signale vom Gehirn zum Körper und umgekehrt weiterzuleiten. Das Metencephalon (Hinterhirn) kann man wieder in zwei Bereiche unterscheiden. Ein Bereich ist die Pons (Brücke). Der andere Bereich ist das Kleinhirn (Cerebellum). Das Kleinhirn besitzt eine stark gefaltete Rinde. Die Aufgabe des Kleinhirn besteht in der ___________________________________________________________________________________ 15 Brain Modelling Kontrolle des sensomotorischen Systems. Wenn das Kleinhirn ausfällt, dann ist die präzise Bewegungskoordination und die motorische Anpassung eingeschränkt. Das Mesencephalon (Mittelhirn) lässt sich wieder in mehrere funktionelle Einheiten unterscheiden. Zum einen gibt es das Tectum, das zwei paarige Ausbeulungen besitzt. Das hintere Paar - Colliculus inferior dienen der Hörverarbeitung, das vordere Paar - Colliculus superior - unterstützt die Sehverarbeitung. Durch diese beiden Kerne wird die Seh- und Hörinformation unabhängig von anderen Arealen verwaltet. Im Tectum werden keine komplexen Muster verarbeitet, es wird nur eine grobe Abschätzung über die Umwelt getroffen. Damit kann rasch auf mögliche Umweltbedrohungen reagiert werden - manchmal werden dann aber auch harmlose Umweltreize als gefährlich eingestuft, da das System nur mit sehr einfachen Mustern umgehen kann. Der zweite große Bereich des Mittelhirns ist das Tegmentum. Teile der Formatio Reticularis ziehen durch das Tegmentum, das auch noch über mehrere Kerne verfügt: Die Substantia grisea centralis, die Substantia nigra und der Nucleus ruber. Die Substantia grisea centralis scheint eine wesentliche Rolle bei der Übermittlung schmerzreduzierender Wirkungen von Opiaten zu spielen. Für die Steuerung von motorischen teilrhythmischen Bewegungen ist die Substantia nigra zuständig. Auch der Nucleus ruber hat Einfluss auf das sensormotorische System. Das Diencephalon (Zwischenhirn) umfasst zwei Strukturen, zum einen den Thalamus, zum anderen Hypothalamus. Der Thalamus umfasst verschiedene Kerne. Viele dieser Kerne dienen als Schaltstelle für sensorischen Input. Die vorverarbeiteten Signale werden dann in die Großhirnrinde weitergeleitet. Der Thalamus besitzt aber auch noch einige unspezifische Kerne, die der Modulation der Synchronisation in der Großhirnrinde dienen. Über diese Kerne werden auch verschiedene Rindenareale miteinander verschaltet. Der Hypothalamus enthält eine Vielzahl von Kernen, die der Steuerung motivationaler Zustände dienen. Über diese Kerne kann die Hypophyse zur Hormonfreisetzung angeregt werden. Über die Hypophyse kann der Hormonspiegel im Blut geregelt werden, umgekehrt kann aber auch der Hormonspiegel die Hypophyse und die damit verbundenen Gehirnstrukturen beeinflussen. Die Funktion der Mamillarkörper - zwei Kerne des Hypothalamus - ist bisher heute leider noch nicht geklärt. Als besonders wesentlich ist die Formatio Reticularis zu erwähnen. Dieser Bereich fasst ungefähr 100 Kerne vom verlängerten Rückenmark bis zum Mittelhirn zusammen. Die Formatio Reticularis wird auch manchmal als aufsteigendes reticuläres Aktivierungssystem bezeichnet (ARAS). Diese Kerne scheinen für die Steuerung der Aufmerksamkeit, des Schlafes und Herz- Kreislaufreflexe zuständig zu sein. Die genaue Funktion vieler Kerne ist bis heute noch nicht geklärt - wenn allerdings einzelne Kerne beschädigt werden, dann kann dies zu beträchtlichen Schädigungen (Autismus) führen. ___________________________________________________________________________________ Aufbau des Gehirn 16 Abbildung 3.2: Eine dreidimensionale Darstellung verschiedener Bereiche des Gehirns. Das Telencephalon oder auch Endhirn stellt den größten Bereich des Gehirns dar. Die Großhirnrinde oder auch der Neokortex dient der Speicherung und Verarbeitung aller einlangenden Informationen (siehe Kapitel Großhirnrinde). Die unterschiedlichen Bereiche der Großhirnrinde sind durch Faserverbindungen miteinander verbunden. Diese Verbindungen stellen den größten Teil des Volumens des menschlichen Gehirns dar. Ein Teil der Großhirnrinde ist der Hippocampus, der sich allerdings wesentlich von der übrigen Rinde unterscheidet. Die Hippocampusformation ist anders aufgebaut, als die Großhirnrinde, und sie dient ausschließlich der Gedächtniskonsolidierung. In vielen Lehrbüchern wird die Hippocampusformation als Teil des limbischen Systems angesehen. Zum limbischen System wir die Amygdala, der Gyrus cinguli, der Fornix, das Septum und die Mamillarkörper angesehen. Diese Kerne und Rindenareale sind sehr stark miteinander verbunden. Deshalb spricht man auch von einem System. Allerdings hat der Hippocampus eine andere Aufgabe als manche übrigen Kerne. So dient der Hippocampus der Gedächtnisspeicherung, während die Amygdala (Mandelkernkomplex) der Verarbeitung von Emotionen dient. Von manchen Kernen ist die Wirkungsweise noch nicht bekannt, beziehungsweise höchst umstritten (Mamillarkern). Ob die Bezeichnung Abbildung 3.3: Die Kerne, Gebiete der Großhirnrinde und limbisches System noch aufrecht Faserzüge des limbischen Systems. erhalten werden kann, wird sich zeigen. ___________________________________________________________________________________ 17 Brain Modelling Abbildung 3.4: Dreidimensionale Darstellung verschiedener Strukturen des Gehirns. Links erkennt man die verschiedenen Bereiche des limbischen Systems, rechts sind die Basalganglien gezeichnet. Zum Telencephalon gehören auch die Basalganglien. Diese Kerne spielen eine entscheidende Rolle bei der Entstehung von Willkürbewegungen. Sie setzen sich aus dem Nucleus caudatus (Schweifkern) und dem Putamen zusammen und werden gemeinsam als Steifenkörper (Striatum, Corpus striatum) bezeichnet. Auch der Globus pallidus wird zu den Basalganglien gerechnet. Cortex cerebri Telencephalon limbisches System Basalganglien Thalamus Diencephalon Hypothalamus Tectum Mesencephalon Neocortex Hippocampus Amygdala Hippocampus Fornix Gyrus cinguli Septum Mamillarkörper Nucleus caudatus Putamen Globus pallidus spezifische Kerne - Sensorik unspezifische Kerne Mamillarkörper Hypophyse Nucleus ventromedialis Nucleus paraventricularis Nucleus supraopticus Colliculi superior Colliculi inferior Tegmentum Metencephalon Myelencephalon Formatio reticularis Pons Cerebellum Formatio reticularis ___________________________________________________________________________________ Aufbau des Gehirn 18 3.1 Steuerung und Regelung Steuern ist ein Vorgang bei dem eine oder mehrere Größen als Eingangsgrößen in einem System andere Größen als Ausgangsgrößen beeinflussen. Die Beeinflussung ist von den Gesetzmäßigkeiten des Systems abhängig. Betrachten wir einen Gleichspannungsmotor. Über den Strom steuern wir die Drehzahl, das heißt wenig Strom geringe Drehzahl, großer Strom hohe Drehzahl. Bei diesem Beispiel ist der Strom die Eingangsgröße, die Drehzahl ist die Ausgangsgröße. Durch eine Veränderung der Eingangsgröße (Strom) kann die Ausgangsgröße (Drehzahl) verändert werden. Im Idealfall würde bei einem konstanten Strom die Umdrehungszahl konstant bleiben. Kommt es aber zu einer Änderung des Lastverhaltens, mehr Gewicht muss gezogen werden, dann ändert sich die Umdrehungszahl. Alle Größen die eine Veränderung der Ausgangsgrößen nach sich ziehen werden als Störgrößen bezeichnet. Dies können externe Faktoren wie eine Laständerung oder auch interne Faktoren wie eine Änderung der Impedanz (Innenwiderstand) sein. Kennzeichnend für eine Steuerung ist der offene Wirkungsablauf, Störgrößen werden nicht berücksichtigt. Das Regeln ist ein Vorgang, bei dem die zu regelnde Größe die ganze Zeit erfasst wird, mit einer Führungsgröße verglichen wird und entsprechend an die Führungsgröße angeglichen wird. Der Wert der Führungsgröße ist der Sollwert, der aktuell gemessen Wert ist die Istgröße, die zu regelnde Größe ist die Regelgröße. Für unser Beispiel bedeutet dies, dass ein Messmechanismus (Drehzahlmessgerät) angebracht werden muss. Über dieses Messgerät kann die Spannung nach Bedarf geregelt werden. Wenn die Führungsgröße konstant ist, wird von einem Festwertregler gesprochen. Wenn sich die Führungsgröße ändert, aufgrund von äußeren oder inneren Beeinflussungen, spricht man von einem Folge- oder Zeitplanregler. Kennzeichnend für eine Regelung ist der Sollwert-Istwert-Vergleich, der laufend in einem geschlossenem Wirkungskreislauf durchgeführt wird. Gewünschte Drehzahl w e=w-xRegeldifferenz Stellgrösse y x Abbildung 3.5: Die Rückkopplung bei einer Regelung. In der oberen Graphik 4.1 sehen wir ein Blockschaltbild eines Regelkreislaufes. Wesentlich ist die Invertierung des Istwerts (x -x). Damit kann eine Differenz e=w-x gebildet werden. Die Regeldifferenz wird nun für das Stellglied in geeigneter Weise umgewandelt, die Stellgröße, und wirkt solange auf den Effektor (Heizung, Motor usw.) bis der Istwert gleich dem Sollwert ist. Das entspricht einer Gegenkopplung (negative Rückkopplung) und die Differenz zwischen Soll- und Istwert wird geringer. Würde die Rückkopplung mit einem positiven Vorzeichen durchgeführt werden, ergäbe dies eine Mitkopplung (positive Rückkopplung) und die Störgrößen würden noch weiter verstärkt werden. ___________________________________________________________________________________ 19 Brain Modelling 3.2 Der Hypothalamus - als Gruppe von Kernen Regulierung der Körpertemperatur Alle Tiere haben eine ideale Betriebstemperatur. Diese Temperatur ist nach oben durch die Denaturierung der Eiweißstoffe, was zu einer Zerstörung der Zellen führt, begrenzt. Nach unten ist die Grenze durch die Bildung von Eiskristallen in Zellen gekennzeichnet. Doch viele biochemische Prozesse besitzen eine optimale Temperatur zwischen diesen Extrema. Kaltblüter können keinen direkten Einfluss auf ihre Körpertemperatur nehmen. Ihre Körpertemperatur hängt sehr stark von der Umgebung ab. Zur Temperaturregelung können diese Tiere nur das Mikroklima wechseln. Warmblüter können über den Stoffwechsel ihre eigene Temperatur regeln. Sie sind damit unabhängig von der Umwelt. Bereits 1880 konnte gezeigt werden, dass eine Region des Zwischenhirns, der Hypothalamus für die Temperaturregelung verantwortlich ist. Kommt es bei Warmblütern zu einer Erwärmung des Blutes im Hypothalamus so ist schwitzen, hecheln und keuchen das Resultat. Kommt es umgekehrt zu einer Abkühlung des Blutes und damit des Hypothalamus, so muss der Organismus auf wärmeerzeugendes Verhalten umstellen. Es kommt zum Zittern, Verengungen der Hautadern und zu einer Anregung der Stoffwechselprozesse um mehr Wärme zu produzieren. Interessanterweise beginnen Menschen schon zu zittern wenn sie in eine kältere Umgebung kommen bevor die Hypothalamustemperatur gesunken ist. Also müssen externe Sensoren, auf der Haut, den Hypothalamus mit Informationen versorgen. Bei körperlicher Arbeit kommt es zu einer Schweißproduktion um den Körper vor Überhitzung zu bewahren. Die Schweißproduktion setzt schon ein bevor die Hypothalamusoder Körpertemperatur steigt. Bei Hunden fanden sich spezielle Rezeptoren in den Muskeln und Gelenken, die in Kontakt mit dem Hypothalamus stehen. Wärmerezeptoren auf der Haut B Wärmeabgabe A Wärmeerzeugung Rezeptoren in Muskeln & Gelenke Kälterezeptoren auf der Haut Rückenmark Hypothalamus Erregung Hemmung Hirnstamm Abbildung 3.6: Regelsystem für die Körpertemperatur. Im Hypothalamus befinden sich zwei Gruppen von Neuronen, die empfindlich auf Temperaturabweichungen reagieren. Eine Gruppe reagiert auf die Abweichung in Richtung Kälte, eine andere in Richtung Wärme. Die Abweichung der Temperatur führt zu einer proportionalen Änderung der Feuerfrequenz der beteiligten Strukturen. Diese Neuronengruppen werden von den Wärme- und Kälterezeptoren auf der Haut innerviert. Zusätzlich können die Rezeptoren in Muskeln und Gelenken die wärmeempfindlichen Neuronen erregen, während die kälteempfindlichen Neuronen gehemmt werden. Die einzelnen Gruppen im Hypothalamus können sich gegenseitig hemmen. ___________________________________________________________________________________ Aufbau des Gehirns 20 Regulation des Körpergewichts Die Hauptaufgabe des Essens liegt in der Versorgung des Körpers mit Energie und Baustoffen. Der Energieverbrauch im Körper ist kontinuierlich, während die Nahrungsmittelzufuhr punktuell stattfindet. Deshalb wird ein Teil der Nahrung gespeichert. Die größte Speicherkapazität besitzt Fett, das ungefähr 85% der Energiereserven darstellt. Glucogen mit 0.5% und Proteine mit 14.5% haben nur eine untergeordnete Rolle in der Nahrungsmittelspeicherung. Man unterscheidet 3 verschiedene Phasen der Verdauung: cephalische Phase resorptive Phase Fastenphase Die cephalische Phase dient dazu, den Körper auf die bevorstehende Nahrung vorzubereiten. Durch das Sehen und Riechen der Speisen wird unter anderem der Speichelfluss angeregt. In der resorptiven Phase wird der aktuelle Energiebedarf gedeckt und Reserven für "schlechte" Zeiten angelegt. Während der Fastenphase greift der Körper auf die gespeicherten Energieformen zurück. Diese Steuerung geschieht durch die beiden Hormone Insulin und Glucagon. Während der ersten beiden Phasen wird vor allem das Insulin ausgeschieden, während in der Fastenphase vermehrt das Glucagon ausgeschüttet wird. Eine hohe Glucagonkonzentration im Körper führt zur Freisetzung von freien Fettsäuren aus dem Fettgewebe. Das Insulin hingegen sorgt für eine Verwertung von Glucose, die Glucose wird in Glycogen und Fett umgewandelt, Aminosäuren werden in Proteine umgewandelt, Das Glycogen wird in der Leber und der Muskulatur, das Fett im Fettgewebe und Proteine in der Muskulatur gespeichert. Das Hormon Insulin regelt sehr viele Prozesse bei der Nahrungsverwertung. Im Regelfall schwankt die Grundlinie des Blutzuckerspiegels um rund 2%. Allerdings sinkt der Blutzuckerspiegel rund 10 Minuten vor einer erwarteten Mahlzeit um ungefähr 8%. Wenn dem Körper keine Nahrung zugeführt wird, dann kehrt der Blutzuckerspiegel wieder auf sein ursprüngliches Niveau zurück. Das heißt der Körper reagiert auf Gewohnheiten. Leider sind die Regelkreisläufe im Inneren des Körpers unbekannt, manche Wissenschaftler diskutieren sogar, ob es überhaupt diese Regelkreisläufe gibt. Es gibt aber triftige Gründe, die für ein oder mehrere Regelsysteme des Körpergewichts sprechen. Man beobachtete das Körpergewicht einer Population von Ratten im Labor. Am 30. Tag wurde die Gruppe gedrittelt. Ein Teil der Gruppe wurde zwangsernährt, ein Teil der Gruppe bekam bedeutend weniger Nahrung und die dritte Gruppe diente als Kontrollgruppe (siehe Abb. 3.7). Nach 15 Tagen konnten die Tiere aller Gruppen wieder selbstständig über ihre Nahrung verfügen. Es zeigte sich, dass die Gruppe mit der Fastenkur nun vermehrt Nahrung aufgenommen hat, während die zwangsernährte Gruppe mit Übergewicht nun weniger Nahrung zu sich nahm. Nach ungefähr 25 Tagen hatten alle Abbildung 3.7: Die Regulation des KörpergeTiere wieder das selbe Gewicht. Dieses wichts, trotz einer vorherigen Zwangsfütterung, Experiment lässt auf einen Regelkreislauf bzw. Fastenkur. schließen. Wenn im Hypothalamus der ventromediale Kern zerstört wird, dann leiden die Tiere an Hyperphagie, das heißt die Tiere überfressen sich kontinuierlich. Es gibt Personen mit einem ___________________________________________________________________________________ 21 Brain Modelling gewaltigem Übergewicht, meist seit Geburt. Diese Personen leiden in der Regel an einem gutartigen Tumor in der Nähe des ventromedialen Kern des Hypothalamus. Sie haben immer Hunger und auch ein ausgiebiges Essen kann ihren Hunger nicht stillen. Wenn umgekehrt der laterale Hypothalamuskern zerstört wird, dann verweigern die Tiere die Nahrung. Eine elektrische Stimulation der beiden Kerne führt zu dem gegenteiligen Effekt. Man überprüfte ob diese beiden Kerne möglicherweise ein Sollwert des Hungergefühls, beziehungsweise der Nahrungsmittelzufuhr verankert ist. Aus diesem Grund ließ man einige Tiere einer größeren Gruppe hungern. Als sie ausreichend Gewicht verloren hatten, wurde an ihnen eine Läsion am ventralen Hypothalamus durchgeführt. Diese Schädigung wurde ebenfalls an einer nicht hungernden Gruppe von Ratten durchgeführt. Unmittelbar nach der Schädigung nahm diese Gruppe rapide an Gewicht ab. Nach einiger Zeit, stellten beide Gruppen - hungernd und nicht hungernd mit der Schädigung des lateralen Hypothalamus bei freier Nahrungsmittelwahl ein neues Körpergewicht ein. Dieses Gewicht lag unter dem Gewicht der Kontrollgruppe, an denen keine Läsion durchgeführt wurde. Dieses Experiment Abbildung 3.8: Die Verschiebung des Sollwerts lässt auf einen Sollwert schließen. Dennoch für Gewicht nach einer Läsion des lateralen sind auch noch andere Mechanismen an der Hypothalamus. Regulation des Körpergewichtes beteiligt. Auch das hormonale Gleichgewicht ist für die Nahrungsmittelverwertung von wesentlicher Bedeutung. Bei der Nahrungsmittelzufuhr kommt es zu einer vermehrten Abgabe von Insulin. Allerdings kann auch Insulin alleine ein massives Hungergefühl auslösen. Dieses Hungergefühl kann übermächtig werden. Im Laufe des Tages kann es zu einem leichten Hungergefühl kommen. Dies scheint mit einem leicht erhöhten Insulinspiegel zusammenzuhängen. Jetzt gibt es zwei Arten von Menschen, bei denen es zu einem unterschiedlichen Verhalten kommt. Die eine Gruppe isst einen Kornspitz oder einen Apfel. Diese Nahrung reicht aus, um das Insulin abzubauen. Das Hungergefühl ist gestillt. Bei der anderen Gruppe kommt es zu einem anderen Verhalten. Sie essen genauso eine Kleinigkeit, und nach ein paar Minuten kommt es zu einem übermächtigen Hungergefühl. Die kleine Nahrungsmittelmenge hat dafür gesorgt, dass zusätzlich Insulin freigesetzt wird, damit noch mehr Nahrung besser verdaut werden kann. Es zeigte sich, dass auch die absolute Menge an Insulinfreisetzung zu einem Übergewicht führen kann. Einer Versuchsgruppe wurde zusätzlich Insulin gespritzt. Beide Gruppen, mit und ohne dem Insulin, veränderten ihr Körpergewicht, obwohl alle Tiere die gleiche Nahrungsmenge bekamen. Der erhöhte Insulinspiegel führte zu einer besseren Fettumwandlung, das zu einer Gewichtszunahme führt. Allerdings können auch andere chemische Stimulantien einen starken Einfluss auf die Ernährung haben. Wenn der Nucleus paraventricularis mit Noradrenalin stimuliert wird, beginnen die Versuchstiere vermehrt Kohlenhydrate zu sich zu nehmen, während fettreiche oder proteinreiche Nahrungsmittel nicht beachtet wurden. Umgekehrt nehmen Tiere vermehrt Fett zu sich, wenn die Stimulation durch Galanin erfolgt. Opiate im allgemeinen führen zu einem Proteinhunger. Der stärkste Appetitanreger, der im Moment bekannt ist, ist das Neuropeptid Y. Die Versuchstiere reagieren vor allem auf kohlehydratreiche Kost. Es gibt auch Appetitzügler, wie zum Beispiel Amphetamine oder auch der Neurotransmitter Dopamin. Leider gibt es bei Amphetaminen sehr starke Nebenwirkungen und damit scheiden diese Substanzen zur Gewichtsreduktion aus. Da das Dopamin in der Biochemie des Gehirns ___________________________________________________________________________________ Aufbau des Gehirns 22 ein großer Stellenwert besitzt, würden andere Funktionen stark beeinträchtigt sein. Eine andere Substanz, das Cholecystokinin, entsteht im Zwölffingerdarm und verlangsamt die Entleerung des Magens. Dadurch sind die Rezeptoren, die den "Füllstand" angeben länger aktiv - das Völlegefühl herrscht länger vor. Da diese Substanz auch vom Gehirn ausgeschüttet wird, vermutet man, dass sie bei der Regulation der Nahrungsmittelzufuhr eine (wichtige) Rolle spielt. Durch die Experimente mit den chemischen Stimulantien kann man vermuten, dass es mehrere unabhängige Regel- oder Steuermechanismen gibt. Das Problem wird zusätzlich durch andere Experimente verkompliziert. Wenn der Trigeminus, verantwortlich für die Gesichtsmuskulatur, durchtrennt wird, ergibt sich ein interessantes Phänomen. Bei attraktiver Nahrung begannen die Tiere mehr zu fressen, während bei Nahrung, der Bitterstoffe beigemengt waren, die Tiere weniger Nahrung zu sich nahmen. Wenn man von einem Regelmodell ausgeht muss man aber auch andere Faktoren berücksichtigen, die gegen dasselbe sprechen. In früherer Zeit konnten die Menschen sich nicht aussuchen, wann es Nahrung gibt. Die Nahrung wurde verzehrt und wenn ausreichend Nahrung vorhanden war, wurde der Überschuss in Fettreserven gespeichert. Es war nicht planbar, wann es den nächsten Nahrungsmittelschub gibt. Es zeigte sich, dass auch bei langanhaltenden Fastenphasen der Blutzuckerspiegel konstant gehalten wird. Also wird über den Blutzuckerspiegel die Nahrungsmittelzufuhr nur indirekt gesteuert. Aber auch soziale Faktoren haben einen wesentlichen Einfluss, die berücksichtigt werden müssen. ___________________________________________________________________________________ 23 Brain Modelling 3.3 Der Thalamus Der Thalamus setzt sich aus mehreren Kernen zusammen. Er hat zwei wesentliche Funktionen: 1) Eine Schaltfunktion zwischen einzelnen sensorischen Systemen und den dazugehörigen primären sensorischen Cortexarealen. (siehe visuelle Informationsverarbeitung). Diese Bereiche des Thalamus haben eine Art Torwächterfunktion. Nur unter bestimmten Umständen wird die Information weitergeleitet. Gesteuert wird dies durch die im Stammhirn liegende Formatio Reticularis. Im wesentlichen sind die Neuronen in diesen spezifischen Thalamuskernen topologisch geordnet. Das heißt die Information die einlangt, wird nachbarschaftsbezogen weiterverarbeitet. Abb. 3.9: Die Innervierungsgebiete des Thalamus auf die Großhirnrinde. 2) Steuerung des Zustandes verschiedener Gehirnbereiche. So werden die sensorischen Cortexareale über die Erregungsniveaus kontrolliert (vgl. Bedingung für Synchronisation). Es existieren ungefähr 12 verschiedene unspezifische mediale Thalamuskerne. Diese Kerne besitzen aber auch einen Einfluß auf die spezifischen Thalamuskerne. Sie haben damit auch einen Einfluß auf die selektive Aufmerksamkeit. Über den Nucleus centralis und Nucleus medialis können Verbindungen zwischen verschiedenen unterschiedlichen Cortexarealen hergestellt werden. ___________________________________________________________________________________ Aufbau des Gehirns 24 3.4 Die Großhirnrinde Die Großhirnrinde, anatomisch Cortex cerebri genannt, gehört zur grauen Substanz, in der die Zellkörper der Hirnneuronen liegen und in der Signale verarbeitet werden. Die menschliche Großhirnrinde ist bei rund 1000 Quadratzentimeter Fläche nur etwa 2 Millimeter dick. Die weiße Substanz liegt unterhalb der Großhirnrinde und sie enthält außer den überall im Nervensystem eingestreuten Hilfs- und Stützzellen bloß Verbindungen. Es werden nur Signale übermittelt, entweder von einem Areal zu einem anderen Areal beziehungsweise zu einzelnen Muskeln. Abb. 3.10: Die Großhirnrinde im Querschnitt. Man erkennt sehr gut die Abgrenzung der Grauen und der Weißen Substanz. Die Großhirnrinde besteht zu 85% aus Pyramidenzellen. Diese wirken in der Regel erregend (Achtung: eigentlich sind es die Rezeptoren, die entscheiden ob ein Neurotransmitter erregend oder hemmend wirkt). Der typische Neurotransmitter ist Glutamat oder Aspartat; der Rest besteht aus Sternzellen, deren Axone sehr kurz sind. Wenn die Synapsen der Sternzellen Dornen tragen, dann wirken sie exzitatorisch sonst inhibitorisch. Dornlose Sternzellen verwenden meist GABA als Neurotransmitter. Die absolute Zahl der dornlosen Sternzellen ist gering, da aber die Synapsen direkt an den Zellkörper der Pyramidenneuronen eine Verbindung herstellen, ist ihre Wirkung größer und der Einfluss der Sternzellen dürfte ungefähr gleich groß sein wie die der Pyramidenzellen. In den meisten Fällen ist die Großhirnrinde deutlich geschichtet. Im Querschnitt sieht man dann ein gestreiftes Muster, das die Anordnung von Nervenzellen und Fasern widerspiegelt. Durch verschiedene Färbetechniken lassen sich die unterschiedlichen zelluläre und funktionellen Eigenschaften erkennen: Golgi-Färbung: Mehrere vollständig eingefärbt (links). Neuronen werden Nissl-Färbung; Nur die Zellkörper treten hervor (Mitte). Der Unterschied zwischen den Pyramidenzellen und den Körnerzellen ist leicht erkennbar. Weigert-Färbung; Die Fortsätze wie Axone oder Dendriten werden gefärbt (rechts). Eine säulenartige Struktur tritt zutage. Abbildung 3.11: Verschiedene Färbetechniken angewandt auf ein Großhirnrindenareal. Durch die verschiedenen Färbetechniken, kann sowohl die Schichtstruktur als auch die tangentiale Faserstruktur leicht erkennbar gemacht werden. ___________________________________________________________________________________ 25 Brain Modelling Der Cortex cerebri untergliedert sich von außen nach innen in folgende sechs Schichten: I. II. III. IV. V. VI. molekulare Schicht: sie besteht aus verstreut liegenden, kleinen horizontal orientierten Zellen und tangentialen Assoziationsfasern; Über diese Assoziationsfasern kann ein Kontakt zu benachbarten Hirnarealen hergestellt werden. äußere Körnerschicht: sie ist aus dicht gelagerten Körnerzellen aufgebaut, deren Axone in der gleichen Schicht enden; äußere Pyramidenschicht: pyramidenförmig gebaute Zellen bilden den Hauptanteil in dieser Schicht. Die absteigenden Axone, die die Pyramiden-Projektionsbahnen bilden werden bereits innerhalb dieser Schicht mit einer Markscheide umgeben. innere Körnerschicht: sie ist ähnlich wie die Schicht II beschaffen, jedoch im Bereich der Sehrinde besonders stark ausgeprägt. innere Pyramidenschicht: zum einen aus großen Pyramidenzellen sowie zum anderen aus horizontal ausgerichteten Neuronen aufgebaut. Spindelzellenschicht: sie ist aus vielgestaltigen Zellen zusammengesetzt, wobei die größeren vornehmlich außen und die kleineren innen liegen. Die zugehörigen Neuriten ziehen in das innen gelegene Marklager sowie auch in umgekehrter Richtung in die äußeren Rindenschichten. Abb. 3.12: Verschaltungen im Inneren der Großhirnrinde. Die verschiedenen Zellschichten lassen sich funktionell in drei Gruppen unterteilen: 1) Die zwei untersten Schichten V und VI senden ihre Axone in andere Hirnregionen. 2) Die Schicht IV empfängt Axone aus anderen Regionen. 3) Die Schichten I bis III erhalten hauptsächlich Eingänge aus der Schicht IV. Es gibt also Schichten mit vorwiegend kleinen oder vielen großen Zellen, Schichten mit Fasern vorwiegend parallel oder senkrecht zur Fläche. Zumeist gibt es auch eine abgrenzbare Schicht, in der die Signale den Cortex über aufsteigende - afferente - Fasern erreichen, und eine andere, von der die meisten absteigenden - efferenten - Fasern ausgehen und die Signale in andere Hirnteile weiterleiten. Dies kann durch verschiedene Färbetechniken verdeutlicht werden: Die Großhirnrinde ist aber nicht isotrop, das heißt die Anzahl der Neuronen, die Art der Verschaltung und dergleichen kann stark variieren (Durchschnittliche Axonlänge, Zellanzahl usw.). Primäre sensorische Areale haben eine ausgeprägte Schicht IV, motorische Areale besitzen eine stark vergrößerte Schicht V und VI. Diese Unterschiede und Verteilungen werden in cytoarchitektonischen Karten angegeben. Die gebräuchlichste Darstellung ist die Kartierung nach Brodmann (Graphik unten). ___________________________________________________________________________________ Aufbau des Gehirns 26 Manchmal findet sich zwischen der Struktur und der Funktion ein eindeutiges Korrelat (Sehzentrum-Brodmann-Areal 18). Durch neuere Verfahren kann man heute die Großhirnrinde in bis zu 200 Areale unterteilen. Abb. 3.13: Laterale (rechts) und mediale (links) Darstellung der Brodmann-Areale. Manche Felder lassen sich leicht voneinander abgrenzen, sie sind durch eine dicke Linie gekennzeichnet. Felder die sich zytologisch schwerer voneinander abgrenzen lassen sind durch dünn gezeichnete Linien markiert. Areale, die allmählich ineinander übergehen sind durch gestrichelte Linien ausgewiesen. Projektionsbahnen Die Großhirnrinde ist ausgiebig mit sich selbst verkabelt, denn die Substanz darunter, das sogenannte Hemisphärenmark (weiße Substanz), besteht größtenteils aus Fasern, die an einer Stelle des Cortex entspringen und an einer anderen - nahen oder entfernten - Stelle wieder eintreten. Ob zwei Stellen miteinander verknüpft sind, hängt in erster Linie nicht von ihrem Abstand ab. Alle Faserzüge sind von großem Interesse. Die Zerstörung einer Bahn kann zu ebenso schweren Verhaltensdefiziten führen, wie die Zerstörung der jeweiligen Areale oder einzelner Kerne. Afferente und efferente Bahnen: Die Großhirnrinde erhält ihre elektrischen Signale von rund einer Million Eingangsfasern. Die meisten sensorischen Systeme projizieren auf den Thalamus. Von dieser Umschaltzentrale erhalten die meisten primären sensorischen Areale ihre Eingangsfasern. Assoziationsbahnen: Über diese Bahnen werden Areale in der gleichen Hemisphäre verknüpft. Zum Beispiel wird ein motorisches Areal mit einem sensorischen Areal verbunden. Es werden Reize unterschiedlicher Modalität verknüpft. Abb. 3.14: Die verschiedenen Bahnen (strukturell) im Inneren des Gehirns. ___________________________________________________________________________________ 27 Brain Modelling Kommissurenbahnen: Es wird eine Verknüpfung von homotropen Regionen hergestellt. In der rechten und linken Großhirnrindenhälfte gibt es jeweils ein Areal, das für die motorische Koordination der jeweiligen gegenüberliegenden Köperhälfte verantwortlich ist. Es ist bei manchen Bewegungen notwendig die beiden unterschiedlichen Bewegungsabläufe, gesteuert durch das gegenüberliegende Großhirnrindenareal, zu koordinieren. Dies geschieht durch die Kommissurenbahn. Über den Balken (Corpus callosum), bestehend aus 200 Millionen Fasern, werden verschiedenste Informationen zwischen den Rindenhälften abgeglichen. Im Gegensatz dazu ist die Commissura anterior nur für den Abgleich von Informationen des limbischen Systems betreffend verantwortlich. Abb.3.15: Die Verbindungen auf der Großhirnrinde. Assoziationsfelder Das menschliche Gehirn ist anatomisch in vier Lappen unterteilt: Frontal-, Parietal-, Occipital, Temporallappen: Funktionell ist aber eine andere Frontallappen Parietallappen Unterteilung sinnvoller. Es existieren verschiedene primäre sensorische Kortexareale. Das visuelle, das somatosensorische oder auch das auditorische Cortexareal erhält über den Thalamus die Reize vom jeweiligen Sinnesorgan. Zu jedem sensorischen Kortexareal gibt es ein übergeordnetes (sekundäres) sensorisches Areal. In diesem Areal werden aus den einzelnen Reizen und Reiz-Kombinationen komplexere Eigenschaften "erkannt". Occipitallappen Temporallappen Abbildung 3.16: Die vier Lappen der Großhirnrinde. Diese übergeordneten sensorischen Areale liefern die Reize wiederum an drei verschiedene Areale: Der präfrontale Assoziationskortex: Er ist für die Planung und Durchführung von komplexen motorischen Handlungen verantwortlich. Es werden die Funktionen des prämotorischen und des präfrontalen Cortex miteinander verknüpft. Der prämotorische Cortex wählt eine komplexe Bewegung aus einer Vielzahl von Möglichkeiten aus. Der motorische Cortex ist dann für die Ausführung verantwortlich. Der präfrontale Cortex stellt das Arbeitsgedächtnis dar. Dort existiert ein temporales Gedächtnis über die wahrgenommene Umgebung. Der präfrontale Cortex wählt eine Verhaltensweise aus einer größeren Anzahl von Möglichkeiten aus. ___________________________________________________________________________________ Aufbau des Gehirns 28 der parital-temporal-occipitale Assoziationskortex: Im occipitalen Kortexareal befinden sich das primäre und die sekundären visuellen Verarbeitungseinheiten (vgl. Sehsystem, Teil I). Der Parietallappen lässt sich in zwei unabhängige funktionelle Einheiten unterteilen: 1) Es existiert ein Rindenfeld für die somatische Empfindung. 2) Die andere funktionelle Einheit ist primär mit der Integration von somatischen und visuellen Reizen beschäftigt. Der temporale Bereich hat folgende Aufgaben: die Verarbeitung auditorischer Informationen, das visuelle Erkennen von Objekten und die Langzeitspeicherung sensorischer Daten. Der limbische Assoziationskortex: Die Speicherung von Informationen über unsere Umwelt in das Langzeitgedächtnis, die Motivation und Entscheidungsfindung als auch die emotionelle Bewertung von Handlungen und Situation werden in diesem Bereich des Gehirns bearbeitet. Über den präfrontalen Assoziatonskortex als auch durch die übergeordneten (sekundären) sensorischen Kortices wird der prämotorische Cortex gesteuert. Durch eine direkte Verbindung wird auf den motorischen Cortex eingewirkt und eine Bewegung kommt zustande. primärer motorischer Cortex präfrontaler Assoziationscortex übergeordneter motorischer Cortex parietal-temporal-occipitaler Assoziationscortex primärer sensorischer Cortex sekundärer sensorischer Cortex limbischer Assoziationscortex Abb. 3.17: Die Verknüpfung von verschiedenen Großhirnrindenarealen bei unterschiedlichen „Denkleistungen“ Informationsverarbeitung in der Großhirnrinde A B Abb. 3.18: Darstellung der Detailverschaltung von Säulen (links) im Sehareal und von verschiedenen Arealen (rechts) der Großhirnrinde. In manchen Rindenarealen liegt eine Säulenarchitektur vor (Sehzentrum). Eine Säule besteht aus einer Gruppe von Neuronen (funktionelle Einheit), die alle miteinander über erregende Synapsen verbunden sind. Diese Neuronen können sich synchronisieren. Die unmittelbaren Nachbarn werden miterregt und gleichzeitig werden über inhibitorische Körnerzellen weiter entfernte Zellverbände (Säulen) gehemmt. Es kommt damit zu einer lateralen Hemmung. Nur manche Säulen können sich durchsetzen. Die Information wird kontrastiert. ___________________________________________________________________________________ 29 Brain Modelling Die Neuronen in einer Säule haben aber auch erregende Verbindungen zu anderen Säulen. Über diese Verbindungen können sich verschiedene Säulen untereinander synchronisieren. Unterschiedliche "Eigenschaften" können miteinander physikalisch verbunden werden. Über Assoziationsbahnen können Säulen verschiedener Modalitäten untereinander synchronisieren. Betrachten wir zwei Großhirnrinden-Areale A und B. Die jeweiligen Areale erhalten von den zugeordneten Kernen Aktivierungen. So werden die Neuronen in der Schicht IV aktiviert. Die Verarbeitungsneuronen in den Schichten I bis III verarbeiten die Information und möglicherweise kommt es zu einer Synchronisation (Gebiet A). Über tangentiale (hier nicht eingezeichnet) oder über inner-corticale Assoziations-Fasern wird die Schicht IV eines oder mehrerer Areale mit elektrischen Pulsen aktiviert (Gebiet B). Im zweiten Areal kann es dann zu einer Interferenz mit der Information aus dem Kern und dem anderen Rindenareal kommen. Die Information wird nicht weitergeleitet oder es kommt zu einer weiteren Synchronisation. Das Gebiet kann nun die in den Schichten I bis III verarbeitete Information wieder zurückschicken beziehungsweise damit andere Areale innervieren. Es kann folgendes passieren: [1] Die Information geht verloren. Die EPSP´s können in den nachgeschalteten Neuronen kein Aktionspotential auslösen. Die Verknüpfung der Information ist irrelevant. [2] Es kommt zur Synchronisation in unterschiedlichen Rindenarealen. Verschiedene Informationen werden zu einer zusammengefasst - ABSTRAKTION. ___________________________________________________________________________________ Aufbau des Gehirns 30 –– –– –– –– ++ ++ ++ ++ EEG, MEG, PET Das Gehirn ist in den Schädelknochen eingelagert. Dieser Knochen schützt das Gehirn in ausgezeichneter Weise. Umgekehrt verhindert aber auch dieser Knochen eine einfache Untersuchung des Gehirns. Lange Zeit war man auf pathologische Untersuchungen angewiesen. Wenn ein Patient einen Schlaganfall, eine Hirnhautentzündung usw. hatte, musste man warten, bis der Patient starb, bis man den Schädel öffnen konnte und das Gehirn untersuchen konnte. Diese Technik erfordert viel Zeit und meist sind die Schädigungen diffus und eine eindeutige Zuordnung zwischen dem Gehirnareal und der Funktion ist nur über mehrere Gehirne mit ähnlichen Schädigungen möglich. Mit der Einführung der Röntgendiagnostik war es erstmals möglich in das Innere des Schädels zu blicken. Leider ist das Gehirn eine sehr homogene Masse, so dass man auf einem Röntgenbild nur sehr wenig, vor allem aber kaum Strukturen erkennen kann. Allerdings ist es möglich dem Blut ein Kontrastmittel beizumengen. Damit ist es zumindest möglich, die Blutbahnen des Gehirns sichtbar zu machen. Leider ist es damit nicht möglich das Gehirn beim unmittelbaren Denken näher zu untersuchen. Aber es war schon bald bekannt, dass das Gehirn elektrisch erregbar ist, beziehungsweise, dass das Gehirn, insbesondere die einzelnen Neuronen, selbst kleinste Ströme bei deren Aktivität produziert. Schon um 1870 wurden Untersuchungen an Kaninchen- und Affengehirnen durchgeführt. Allerdings befanden sich bei diesen Experimenten die Elektroden, um die Spannung abzunehmen, direkt auf dem Gehirn. Aber erst 1924 ist es gelungen, diese kleinen Ströme der Gehirnaktivität durch den Schädelknochen hindurch zu messen. Der Nervenarzt Hans Berger publizierte diese Ergebnisse aber erst 1929, da er seine Ergebnisse immer wieder überprüfte. So nahm er sehr viele Daten von sich selbst (56 Messungen) und seinem Sohn (73 Messungen) auf. Berger prägte auch den Begriff E.E.G, das damals noch als Elektroenkephalogramm, heute als Elektroenzephalogramm bezeichnet wird. In den späteren Veröffentlichungen nahm er nahezu fast alle grundlegenden Beobachtungen vorweg. _____________________________________________________________________________________________ 31 Brain Modelling So zeigte sich, dass die elektrophysiologischen Messkurven vom Wachheitsgrad bzw. dem Bewusstseinszustand der Probanden abhing (siehe Abb. 4.0). So zeigten sich zwischen dem EEG und den Zuständen von erregt Wachheit, Entspannung, Schlaf, Hirnreifung, Epilepsie und dem Hirntod gefunden. Berger konnte entspannt auch zeigen, dass das EEG sich nicht nur durch sensorische schläfrig Reize sondern auch durch geistige Tätigkeiten (Kopfrechnen) verändert. Es wurde schlafend auch gezeigt, dass diverse Substanzen, wie Morphine, Kaffee, Schlafmittel, NarkoseTiefschlaf mittel, Insulin usw. die EEGRhythmik verändert. Koma Es wurde eine synchron 50 µV auftretende Aktivität zwischen 1 sec funktionell gleichen RindenAbbildung 4.0: Verschiedene Meßkurven bei unterfeldern der rechten und linken schiedlichem Wachheitszustand der Probanden. Hemisphäre festgestellt. Interessanterweise bleibt diese Aktivität synchron, auch dann wenn die direkten Verbindungen zwischen diesen Rindenarealen zerstört wird. Dies lässt auf eine zentrale Steuereinheit im Inneren des Gehirn schließen. Um international diverse EEG-Untersuchungen vergleichen zu können, wurde im Jahr 1957 das sogenannte 10-20-System festgelegt. Die Elektroden befinden sich im Abstand von 10% bzw. 20%. Als absolute Werte werden spezielle Punkte an den Ohren, der Nase und G im Nacken verwendet. Von diesen Punkten Fp1 Fp2 ausgehend, spannt man ein Netz von F8 Elektroden über die Kopfoberfläche (siehe F7 F3 F4 Fz Abbildung 4.1). Die trockene Kopfhaut besitzt einen großen elektrischen Widerstand. Deshalb wird eine physiologische Kochsalzlösung zwischen der Kopfhaut und der Elektrode aufgetragen. A1 T3 C3 T5 Cz Pz P3 O1 C4 P4 T4 A2 T6 O2 Nun kann man zwischen den einzelnen Elektroden die jeweiligen EEG-Spannungen Abbildung 4.1: Die Position der Elektroden auf messen. Die Spannungen unterscheiden sich dem Kopf mit den Elektrodenbezeichnung im in der Amplitude. Zusätzlich ändert sich die 10-20 System: Frequenz in Abhängigkeit von der Fp = frontopolar F = frontal T = temporal Gehirnaktivität. Diese Frequenzen müssen C = zentral P = parietal O = okzipital gefiltert werden, dass heißt ein Bereich von A = aurikulär G = Erdung speziellen Frequenzen werden elektronisch Ungerade Ziffern beschreiben Elektroden auf der ausgeblendet. Zum Beispiel führt das linken, gerade Ziffern auf der rechten Kopfseite. Schwitzen auf der Kopfhaut zu einer massiven Veränderung der Signale. Diese Signale können schnell dazu führen, dass die Verstärker übersteuert werden, und damit die Signale überhaupt nicht mehr gemessen werden können. _____________________________________________________________________________________________ Untersuchungsmethoden des Gehirns 32 Die Amplituden im EEG haben eine geringe Bedeutung. Dadurch, dass die Amplituden stark von den Ableitebedingungen abhängen, können Vergleiche nur schwer angestellt werden. Die Potentialdifferenz wird zum Beispiel durch die Wahl der beiden Elektroden bestimmt. Wenn man die Spannungen zwischen FP1 und F7 misst, beziehungsweise zwischen FP1 und A1, dann werden sich die Amplituden der Spannungen stark unterscheiden. Die Amplituden die man direkt auf der Großhirnrinde misst liegen bei rund 10 mV, während auf der Kopfoberfläche die Amplituden in der Größe von 100 µV liegen. Für die Aktivität der jeweiligen Rindenareale ist die Frequenz wichtiger, denn die Frequenz verändert sich nicht so leicht, wenn man unterschiedliche Messelektroden verwendet, sie ist aber von der Aktivität der Großhirnrinde abhängig. Das EEG stellt ein Frequenzgemisch dar. Dennoch ist eine Grundfrequenz vorherrschend. Aufgrund des Wachheitszustandes beziehungsweise aufgrund von sensorischen Reizen ändert sich die maßgebliche Frequenz. Die gemessenen Signale werden aufgrund der Frequenz in vier verschiedene Bereiche unterteilt: Band Frequenz [Hz] Amplitude [µV] Bedeutung µ 8 - 13 50 entspannter Wachzustand 10-11 50 entspannter motorischer Zustand 13 - 30 30 Ruhezustand unter 4 100 Schlaf, bei Erkrankungen 4-8 100 Emotionen bei Kindern 30 - 35 unter 30 Tabelle 4.1: Unterteilung der EEG-Wellen nach Amplitude und Frequenz. Im Ruhezustand (entspannter Wachzustand) treten bei geschlossenen Augen rhythmische Wellen mit einer Frequenz von 8-13 Hz im Hinterhauptslappen auf. Dieser Frequenzzustand wird als Alpha ()- Zustand bezeichnet. In den okzipitalen Regionen des Gehirns (Hinterkopf) treten die größten und regulärsten -Wellen auf. Die Amplitude kann während der Messung leicht variieren. Dies hängt mit minimalen Änderungen des Wachheitszustandes des Probanden ab. Vergleicht man die EEG-Kurven des rechten und des linken Sehzentrums, so zeigt sich, dass beide Areale zwar unter den richtigen Umständen -Wellen produzieren, dass aber keine unmittelbare Synchronisation zwischen diesen Gehirnregionen besteht. Dies lässt vermuten, dass für jede Gehirnhälfte getrennt, die -Rhythmen generiert werden. Akustische oder taktile Reize haben einen starken Einfluss auf den -Rhythmus. Die Aufmerksamkeit richtet sich dann auf diesen Reiz und das Gehirn befindet sich nicht mehr im entspannten Wachzustand. Umgekehrt haben abstrakte Denkleistungen, wie Kopfrechnen, kaum bis gar keinen Einfluss auf die -Wellen. Die -Wellen können massiv durch visuelle Prozesse unterdrückt werden. Sobald man die Augen öffnet, auch in einem dunklen Raum, wird der Rhythmus blockiert. Auch eine bildhafte Vorstellung führt zu einer Blockade. Der µ-Rhythmus ist nach dem -Rhythmus die deutlichste und häufigste Form hirnlokaler Aktivität. Diese Aktivität steht im Zusammenhang zu motorischen Aktivitäten. Die Wellen treten vor allem in den motorischen Bereichen auf (C3, Cz, und C4). Die Frequenzen liegen im Bereich des -Rhythmus, meist aber um zirka 1 Hz höher. Die Amplituden liegt ebenso bei rund 50 µV. Bei bilateralem Auftreten sind die einzelnen µ-Wellen der beiden Hemisphären zeitlich nicht korreliert und zeigen unterschiedliche Amplituden. Der µ-Rhythmus ist die Grundaktivität der sensomotorischen und motorischen Areale. Sobald aber eine motorische Aktivität gesetzt wird, zum Beispiel das Formen einer Faust, verschwindet der µ-Rhythmus sofort. Auch die Vorstellung von Bewegung führt zu einer Blockade dieses Grundrhythmussees. _____________________________________________________________________________________________ 33 Brain Modelling Potentialschwankungen mit einer Frequenz oberhalb von 13 Hz werden als Betawellen bezeichnet. Diese -Wellen werden als die eigentliche Form der lokalen Ruheaktivität der Großhirnrinde betrachtet, die lediglich in der hinteren sensorischen Hirnhälfte die -Aktivität ersetzt. Die -Aktivität tritt hauptsächlich bei Säuglingen und teilweise auch bei Kindern und Jugendlichen auf. Bei Erwachsenen treten -Wellen nur im Schlaf oder bei Hyperventilation auf. Sonst haben -Rhythmen ausschließlich pathologische Hintergründe, wie Tumore, entzündliche Prozesse im Gehirn, Gehirntraumata oder Arteriosklerose. Die -Aktivität ist im Kindesalter eine typische Aktivität. Ab dem 8.Lebensjahr tritt die Aktivität in den Hintergrund und die -Aktivität wird stärker. Im Kindesalter treten -Wellen hauptsächlich bei emotionell belastenden Situationen auf. Vermutlich werden die -Rhythmen durch das limbische System ausgelöst. Bei einer Minderung des Wachheitszustandes können -Wellen - mit einer geringen Amplitude - beobachtet werden. Damit stellt sich die Frage, wie es zu den einzelnen Potentialen auf der Schädeloberfläche kommt. Man könnte leicht vermuten, dass die Aktionspotentiale Verursacher des EEG's sind. Diese Annahme ist aber falsch. Ein Aktionspotential hat zwar eine beträchtliche Größe, aber ist nur von sehr kurzer Dauer. Durch die Aktionspotentiale werden aber auch synaptische Potentiale ausgelöst. Diese sind zwar um einiges kleiner in der Größe ( 1 mV), aber sie halten bedeutend länger an (rund 30 mal länger). Zusätzlich darf man nicht übersehen, dass ein Aktionspotential bis zu 1000-10000 Synapsen aktiviert (siehe Abb. 6.2). Dadurch entstehen, angenommen die Hälfte der Synapsen produziert ein Potential von der Größe von 1 mV bei der Aktivierung durch ein Aktionspotential, ein Gesamtpotential von 500-5000 * 1 mV = 500mV - 5000mV. Dieses Summenpotential ist beträchtlich größer als ein einzelnes Aktionspotential (rund 100 mV). U Summenpotential Aktionspotentiale EPSP´s t Abbildung 4.2: Entstehung des Summenpotentials durch mehrere synaptische Potentiale. Vergleich des Summenpotentials, der synaptischen Potentiale und der Aktionspotentiale (nicht direkt prop.!). Ein Neuron kann in einfachster Weise als ein Dipol betrachtet werden. Den einen Pol stellt das Ende des Hauptdendriten dar, während der zweite Pol der Zellkörper ist. Natürlich handelt es sich um eine sehr starke Vereinfachung, die aber nützliche Ergebnisse liefert. Man muss sich dabei bewusst sein, dass ein Neuron eine viel zu komplexe Struktur besitzt um einfach als Dipol beschrieben zu werden. Wird der Hautdendrit (bzw. der Zellkörper) durch synaptische Potentiale (egal ob es sich um EPSP´s oder um IPSP´s handelt) gereizt, so wird lokal die Membran polarisiert (de- oder hyperpolarisiert). Diese Polarisation entspricht einem lokalen Ionenungleichgewicht zwischen der Innen- und Außenseite der Membran. Zusätzlich entsteht eine Potentialdifferenz zwischen dem Hauptdendriten und dem Zellkörper. Außerhalb der Membran des Hauptdendriten befinden sich mehr Kalium-Ionen (im Inneren mehr Natriumionen) als beim Zellkörper. _____________________________________________________________________________________________ Untersuchungsmethoden des Gehirns 34 Dieses Ionenungleichgewicht wird nach einiger Zeit ausgeglichen - die durch synaptische Potentiale verursachte Polarisation der Membran wandert zum Zellkörper. Wenn umgekehrt der Zellkörper polarisiert wird, so wandert das Ionenungleichgewicht zum Ende des Hauptdendriten. Bei der Erzeugung eines EPSP´s an einer Synapse am Ende des Hauptdendriten wird im Membranbereich der nachgeschalteten Synapse die positiven Ladungen an der Zellaußenseite verringert. Dies lässt diesen Bereich relativ zur Außenseite des Zellkörpers, der nicht polarisiert ist, vorübergehend negativer erscheinen. Elektrisch betrachtet wird der depolarisierte Bereich zum Minuspol, während die nicht polarisierten Bereiche (z.B. der Zellkörper) zum Pluspol werden. Das Dipolkonzept kann vor allem bei Pyramidenzellen angewendet werden, da aufgrund der Morphologie des Neurons eine einzige lange Achse, zwischen dem Ende des apikalen Dendriten und dem Zellkörper, gegeben ist. Damit gibt es einen größeren Abstand zwischen dem Pluspol und dem Minuspol als im Gegensatz zu den Körnerzellen mit den charakteristisch kurzen Dendriten. Ein idealisiertes Feldpotential, entstanden durch ein EPSP am Dendriten eines Pyramidenneurons, ist in Abbildung 6.3 dargestellt. Das elektrische Feld wird durch Feldlinien dargestellt. Verbindet man im gesamten Feld alle Punkte mit dem gleichen Potential, so erhält man die Äquipotentiallinien. An Elektroden die sich auf der selben Äquipotentialline befinden, kann keine Potentialdifferenz gemessen werden. Es ist also wichtig, dass die Elektroden so platziert sind, dass eine Potentialdifferenz (=Spannung) gemessen werden kann. +0.25 –1.0 –0.25 0.0 –0.5 –0.25 –1.0 –0.5 –0.25 – 0.0 0.0 + +0.25 +0.25 +0.5 +0.5 Feldlinie +1.0 Äquipotentialline Abbildung 4.3: Darstellung der Feldlinien und Äquipotentiallinien eines durch eine EPSP aktivierten Pyramidenneurons. Der negative Pol liegt am Ende des apikalen Dendriten, während der Pluspol beim Zellkörper liegt. In der Abbildung 4.3 wurde das idealisierte Potential von -1 bis +1 normiert. Nur bei Potentialdifferenzen können Spannungen gemessen werden. Wenn nun eine Neuronenpopulation besonders aktiv ist, viele synaptische Potential die Neuronen aktivieren, und die Elektroden liegen zu weit beieinander, dann kann es passieren, dass nur eine geringe bis gar keine elektrische Aktivität gemessen werden kann. Für das elektrische Potential ist es wesentlich, wo die postsynaptischen Potentiale angreifen, beziehungsweise ob die Membran depolarisiert (EPSP) oder hyperpolarisiert (IPSP) wird. Dies _____________________________________________________________________________________________ 35 Brain Modelling verursacht unterschiedliche Potentiale. Dennoch ist es möglich, daß EPSP´s und IPSP´s gleichartige Potentiale verursachen. Wenn ein IPSP am Zellkörper, bzw. ein EPSP am apikalen Dendriten angreift, dann entstehen gleichartige Potentiale, denn der Strom fließt in die gleiche Richtung. Der einzelne Unterschied besteht in der Stärke des Feldes. Die beiden anderen Fälle, EPSP am Zellkörper und IPSP am apikalen Dendriten, führen zwar wieder zu identen Potentialen zueinander, aber diese Fälle treten in der Großhirnrinde kaum bis gar nicht auf. Somit werden mit einem EEG EPSP´s am apikalen Dendriten und IPSP´s am Zellkörper gemessen. –– –– –– –– ++ ++ ++ ++ ++ ++ ++ ++ - EPSP –– –– –– –– –– –– –– –– ++ ++ ++ ++ –– –– –– –– ++ ++ ++ ++ IPSP + Abbildung 4.4: Unterschiedliche Innervierung durch EPSP´s und IPSP´s an Pyramidenneuronen. Der durch EPSP depolarisierte Bereich wird zum Minuspol, während der durch IPSP hyperpolarisierte Bereich positiver als der übrige Membranbereich wird. _____________________________________________________________________________________________ Untersuchungsmethoden des Gehirns 36 Das gemessene EEG b a hängt sehr stark von der Größe und der Form der Neuronenpopulation ab. Das Feldpotential einer Gruppe von Neuronen entspricht der Summe der Feldpotentiale, die durch die Aktivität der einzelnen Neuronen entstehen. Wenn die Neuronen parallel c d zueinander stehen, und die Gruppe senkrecht zur Kopfoberfläche liegt, dann können die überlagerten Potentiale leicht gemessen werden. Die einzelnen Potentiale überlagern sich (siehe Abb. 4.5a,b). Wenn die Neuronen Abbildung 4.5: In der Graphik a werden die parallel liegenden Pyramidenzellen gleichzeitig aktiviert, die Homogenität und Größe des Potentials ist in nicht gleichzeitig b zu erkennen. In c und d werden die Neuronen ungleichmäßig aktiviert, das aktiviert werden (siehe Potential hebt sich an manchen Stellen auf und wirkt insgesamt geringer. Abb. 4.5c,d), oder die Neuronen nicht gleichgerichtet sind, dann überlagern sich die Potentiale, allerdings löschen sie sich diesmal gegenseitig aus. Man wird ein Signal nur sehr schwer messen können. Schädeloberfläche Großhirnrinde Gruppe von Neuronen In diesem Bereich können die elektrischen Felder nicht gemessen werden. Abbildung 4.6: Lage von Neuronenverbänden. Durch Einstülpungen der Großhirnrinde können Gruppen von Neuronen auch parallel zur Kopfoberfläche liegen. Innerhalb der Großhirnrinde liegen meistens Neuronen parallel zueinander und senkrecht zur Oberfläche der Großhirnrinde. Allerdings entstehen in den Entwicklungsphasen Furchen und Einstülpungen der Großhirnrinde (siehe Abb. 4.6). Dadurch ist es möglich, daß einzelne Gruppen nicht senkrecht zum Schädel sondern waagrecht liegen. Auch wenn diese Gruppe sehr aktiv ist, werden nur geringe Potentiale gemessen (siehe Abb. 4.3). Meist liegen diese waagrecht liegenden Gruppen anderen Gruppen gegenüber. Nicht selten sind beide Gruppen gleichzeitig aktiv. Damit löschen sich die einzelnen Potentiale gänzlich aus und am EEGSchreiber erscheint eine Nulllinie. Diese Gehirnbereiche können aber durch andere Meßmethoden zugänglich gemacht werden. Das Potential der aktivierten Neuronen, die in der Nähe des Schädels liegen, können leichter gemessen werden, als Neuronen die tiefer liegen. Da die Feldstärke eines Dipols mit dem Quadrat der Entfernung abnimmt, werden die einzelnen Potentiale rasch unmessbar. Die Entfernung zwischen den koriticalen Potentialgeneratoren und den EEG-Ableiteelektroden _____________________________________________________________________________________________ 37 Brain Modelling wird durch anatomische Gegebenheiten bestimmt. Nur rund ein Drittel der Großhirnrinde liegt nahe an der Schädeloberfläche. Nur dieses Drittel ist für EEG-Ableitungen unmittelbar zugänglich. Über das EEG können auch langfristige Änderungen der elektrischen Potentiale der Neuronen gemessen werden. Das Membranpotential von Neuronen kann sehr langsame Schwankungen aufweisen. Normalerweise werden die einzelnen Spannungen der Neuronen durch RCVerstärker messbar gemacht. Dabei werden langsame Spannungsveränderungen herausgefiltert. Verwendet man aber Gleichspannungsverstärker, dann können diese langsamen und geringen Potentialänderungen gemessen werden. Meist kommt es dann zu einer Überlagerung zwischen den DC-Potentialen (langsamen Gleichspannungspotentialen) und den EEG-Potentialen. Die Ursache für die DC-Potentiale ist noch nicht geklärt. Man vermutete einen Zusammenhang zwischen der elektrischen Aktivität von Gliazellen und den Gleichspannungspotentialen. Diese Vermutung konnte nicht bestätigt werden. Möglicherweise sind extrem lang anhaltende PSP´s dafür verantwortlich. In der klinischen Praxis für Routineableitungen hat sich die Einstellung des Hochfrequenzfilters auf 70 Hertz, das heißt durch die Messapparatur werden Signale mit höheren Frequenzen herausgefiltert, und der unteren Grenzfrequenz auf 0.53 Hertz (dies entspricht 0.3 Sekunden) als optimaler Kompromiss durchgesetzt. Das EEG dient dazu, Prozesse der Signalverarbeitung im Gehirn zu messen. Einerseits kann das EEG Auskunft über den allgemeinen Zustand einer Person Auskunft geben (Schlaf-, Wachheits- oder Ruhezustand). Andererseits kann auch die neurale Aktivität, die speziellen Wahrnehmungs- oder Denkprozessen zugrunde liegt, gemessen werden. Es sind Potentialänderungen, die durch sensorische Reize oder durch Verarbeitungsprozesse verursacht werden. Diese Potentiale werden als ereignisbezogene Potentiale - ERP´s (event related potential) bezeichnet, manchmal findet man auch noch die veraltete Bezeichnung evoziertes Potential - EP (evoked potential). Leider ist die Potentialänderung bei einem ereignisbezogenem Potential sehr gering. Der Ausschlag beträgt durchschnittlich von 0.1µV bis 20 µV, während die Hauptaktivität des EEG bei 10 µV bis 100µV liegt. Dies führt dazu, daß die ERP´s in der Hintergrundaktivität leicht untergehen. Deshalb erfordert es einen besonderen Aufwand diese Signale aufzuspüren. Bei der Dauerreiz-Methode werden dem Gehirn in kurzen zeitlichen Abständen - mit einer bestimmten Frequenz - idente Reize angeboten. Da die Reize dem Gehirn so rasch hintereinander angeboten werden, überlappen sich die ereignisbezogenen Potentiale und es bildet sich ein Dauerpotential. Dieses Potential ist aber ebenso gering, wie ein einzelnes normales Potential. Um es aus der Hintergrundaktivität des Gehirns herauszufiltern, verwendet man die Fourier-Analyse. Bei der Fourier-Analyse werden Wellen nach den Frequenzeigenschaften untersucht. Da die Reizfrequenz bekannt ist, braucht man nur nach dieser Frequenz im EEG-Signal suchen (siehe Abb. 6.7). _____________________________________________________________________________________________ Untersuchungsmethoden des Gehirns 38 0 1 2 3 4 5 6 7 sec. V Beginn der Reizung 0.0 0.1 0.2 0.3 0.4 0.5 0.6 sec. Abbildung 4.7: In der oberen Darstellung ist ein EEG-Signal mit Dauerreizung dargestellt. In der unteren Darstellung wurde das Signal Fourier-analysiert und man erkennt die Abweichung. Zu beachten ist die unterschiedliche Zeitskala. Das andere Verfahren wird als Mittelungsverfahren bezeichnet und es erfordert zwei Annahmen. Eine Annahme besteht darin, daß das ereignisbezogene Potential in einer konstanten Beziehung zu dem Reiz steht, während die HintergrundAnzahl der gemittelten aktivität, vergleichbar einem Meßkurven Rauschen, keine Regelmäßigkeiten aufweist. Die zweite Annahme 1 besteht darin, daß idente Reize gleiche Amplituden mit der gleichen Zeitdauer verursachen. Dem Gehirn 10 V wird in mehreren Versuchsdurchgängen der idente Reiz geboten. Die EEG-Kurven werden, während der Reiz ausgelöst wird, aufgenommen und summiert und durch die Anzahl der Versuchsdurchgänge dividiert. 4 Auf diese Weise erhält man das mittlere evozierte Potential. Mit dieser Technik wird das Signal, das Potential das durch den Reiz verursacht wird, Rauschverhältnis 16 verbessert. Da bei der Hintergrundaktivität - dem Rauschen - positive wie negative Amplituden mit 200 gleicher Wahrscheinlichkeit auftreten, mittelt sich das Rauschen heraus und nur das Reizsignal bleibt 0 0.1 0.2 0.3 0.4 sec übrig (siehe Abb. 4.8). Die Wirksamkeit der Mittelung wird Abbildung 4.8: Die Mittelung von 200 verschiedenen EEGdurch das Quadratwurzelgesetz Meßkurven führt zu einem eindeutigen Reiz-Potentialbeschrieben. Zusammenhang. _____________________________________________________________________________________________ 39 Brain Modelling Die Einteilung der EEG-Kurven in vier verschiedene Rhythmen ist ziemlich willkürlich. Trotzdem zeigt sich, daß verschiedene Zustände des Gehirns mit diesen Rhythmen korrelieren. Natürlich misst das EEG die Aktivität von einzelnen Neuronen, aber die Gruppen von Neuronen werden von anderen Subsystemen gesteuert. Diese Subsysteme können in den Gruppen eine Synchronisation erleichtern oder erschweren. Somit stellt sich die Frage, welche Subsysteme für diese Rhythmen zuständig sind. Für den Alpha-Rhythmus ist eine besondere Struktur des Thalamus zuständig. Der Thalamus ist eine Gruppe von Kernen im Inneren des Gehirns. Man unterscheidet zwei Arten von Kernen: Spezifische Kerne: Alle Sinnesorgane, mit Ausnahme des Riechens, liefern ihre Signale an den jeweiligen spezifischen Kern. Die Informationsübertragung ist topologisch geordnet, das heißt benachbarte Reizzellen aus den Sinnesorganen liefern ihre Signale an benachbarte Neuronen im Thalamus und diese liefern wiederum ihre Signale an benachbarte Neuronen in der Großhirnrinde. Im Thalamus kommt es zu einer Vorverarbeitung (siehe "Das Sehsystem"). Die Verbindung vom Thalamus zur Großhirnrinde ist reziprok. Man spricht von einer thalamo-cortico-thalmischen Schleife. Unspezifische Kerne: Die unspezifischen Kerne innervieren ebenso die Großhirnrinde, allerdings diffus. Die Abbildung von den Kernen in die Großhirnrinde ist nicht topologisch geordnet. Über diese Kerne kann das Erregungsniveau der Neuronen in der Großhirnrinde gesteuert werden. Dies erleichtert oder erschwert die Synchronisation in den jeweiligen Arealen. Die unspezifischen Kerne erhalten ihre Signale aus anderen tieferliegenden Strukturen des Gehirns, insbesonders der Formatio reticularis. Manche Wissenschafter gehen so weit, daß sie diese Thalamuskerne als eine direkte Fortsetzung der Formatio reticularis bezeichnen. Eine besondere Struktur - der Nucleus reticularis thalami - wird zu den unspezifischen Kernen gezählt, obwohl er eine flächige Struktur besitzt. Diese Zellschicht umgibt alle Kerne des Thalamus. Alle thalamo-cortico-thalmischen Schleifen laufen durch diese Schicht. Er enthält von den durchziehenden Faserbündeln durch Abzweigungen erregende Signale (EPSP´s). Umgekehrt innerviert der Nucleus reticularis thalami die durchgehenden Faserbündel inhibitorisch. U [WE] EEG visueller Stimulus t "Desynchronisation" U [WE] EEG + DC t _____________________________________________________________________________________________ Untersuchungsmethoden des Gehirns 40 Abbildung 4.9: Zwei EEG-Ableitung, mit und ohne Gleichspannungskomponente, während eines visuellen Stimulus. Wenn der Thalamus viele sensorischen Afferenzen erhält, befindet sich der jeweilige thalmische Kern im Transfermodus. Das heißt, daß die Signale von der Sensorik direkt über den Thalamus in die Großhirnrinde gelangen. Im EEG kann man eine sogenannte Desynchronisation beobachten. Dieser Begriff selbst ist ziemlich widersprüchlich. Im EEG erkennt man, daß durch ein Desynchronisation die Amplituden geringer und die Frequenzen höher werden (siehe Abb. 4.9). Praktisch kommt es aber während der Desynchronisation im EEG-Bild zu starken Synchronisationen in den betroffenen Gebieten der Großhirnrinde. Meist führt dies zu einer Frequenz der Aktivität des neuralen Assembles von rund 40 Hertz. Diese Frequenz ist in der Regel für die klassische Auflösung zu groß - das Schreiberpapier bewegt sich zu langsam um höhere Frequenzen sinnvoll aufzuzeichnen. Großhirnrinde Thalamocorticothalmische Schleife Nucleus reticularis thalami spezifischer THALAMUS-KERN Mediale Thalamuskerne Formatio reticularis Abbildung 4.10: Die Verknüpfungsstruktur zwischen einem spezifischen Thalamuskern, dem Nucleus reticularis thalami und der Großhirnrinde. Wenn kaum sensorischen Reize den Thalamus erreichen, dann befindet sich der Thalamus im Oszillatormodus. Die thalamo-cortico-thalmische Schleife wird vor allem durch den Nucleus reticularis thalami aktiviert. Dabei treten abwechselnd kaum Signale und dann kurzzeitig viele Signale als Bursts mit einer Frequenz von 7-14 Hertz auf. Bei verminderter sensorischer Afferenz werden einlaufende Signale, die nicht synchron sind, in den Neuronen der spezifischen Thalamuskerne durch Kontrolle inhibitorischer Rückkopplungen des Nucleus reticularis thalami in gruppierte Entladungen umgewandelt. Diese Entladungen führen über die Verbindungen zur Großhirnrinde zum zur Großtypischen Alpha-Rhythmus. hirnrinde Nucleus reticularis thalami Thalamus FR Formatio Reticularis von der Sensorik Welche Aufgabe hat der -Rhythmus für die Informationsverarbeitung des menschlichen Gehirns. Deshalb ist es notwendig, die psychologischen Bedingungen für das Auftreten des Rhythmus näher zu betrachten. Im Ruhezustand treten die -Wellen vor allem im postzentralen Teil des Gehirns auf. In diesem Gebiet liegt das primäre visuelle, das primäre somatosensorische und zum Teil das primäre auditive Areal. Vor allem das Sehzentrum ist vorrangig in diesem Gebiet vertreten. Dieser Sinn ist der Abbildung 4.11: Die Verschaltung zwischen dem Nucleus reticularis thalami, den Neuronen des Thalamus und der Formatio reticularis. Erregende _____________________________________________________________________________________________ Einflüße werden durch Pfeile und hemmende durch 41einen Block dargestellt Brain Modelling einzige der bewusst durch den Lidschlag abgeschaltet werden kann. Die Aufmerksamkeit wird zu einem wesentlichen Teil durch die Formatio reticularis gesteuert. Diese Kerne erhalten von allen sensorischen Einheiten (wahrscheinlich) unspezifische Signale. In der Formatio reticularis werden diese sensorischen Signale zusammengefasst. Die Formatio reticularis innerviert viele Areale in der Großhirnrinde und im Thalamus. Je stärker die Formatio reticularis aktiv ist, umso leichter können Synchronisationen in der Großhirnrinde auftreten. Beim Schließen der Augen gelangen weniger Informationen in den seitlichen Kniekörper. Dies führt zu einer Abnahme der Aufmerksamkeit. Akustische und somatosensorische Reize können den Abfall der Aufmerksamkeit nur begrenzt verhindern (mit der Ausnahme bei blinden Personen). Wenn die Formatio reticularis verstärkt durch die Sensorik aktiviert wird, dann hemmt sie einerseits den Nucleus reticularis thalami und andererseits wird der Thalamus soweit aktiviert, daß die Signale aus der Sensorik im Thalamus vorverarbeitet werden und dann zur Großhirnrinde gelangen (siehe Abb. 6.11). Wenn die Hemmung der Formatio reticularis auf den Nucleus reticularis thalami aufhört, dann kann sich eine Gegenkopplungsschleife zwischen dem Thalamus und dem Nucleus reticularis thalami bilden. Wahrscheinlich sind dabei auch sogenannte Pace-Maker Zellen beteiligt. Diese Zellen geben, wenn sie aktiviert werden, mit einem konstanten Rhythmus kurzfristig mehrere Aktionspotentiale ab und dann bleiben sie für längere Zeit inaktiv. Dieser einsetzende Rhythmus, bei visueller Reizdeprivation garantiert auch bei geschlossenen Augen - mit Ausnahme, daß das Gehirn sich nicht im Schlafzustand befindet - bei akustischen oder anderen alarmierenden Reizen eine sofortige Reaktionsbereitschaft. Der -Rhythmus kann aber auch durch intensives Assoziationen abgelöst werden. Dabei sind andere Rindenareale beteiligt, insbesonders der Schläfen- und der Frontallappen. Diese beiden Regionen sorgen für Synchronisationen von Neuronen im primären visuellen Areal. Es ist dabei völlig egal ob die Synchronisationen durch visuelle Reize oder durch die Innervation von anderen Rindenarealen verursacht wird. Mit der Technik elektrische Signale des Gehirns zu detektieren, können aber auch noch andere Bereiche, als die Großhirnrinde vermessen werden. So stellt das Stammhirn-EEG einen wichtigen Beitrag dar, um die Hörfähigkeit zu vermessen. Die Elektroden werden im Bereich 1 2 3 4 5 6 7 des Halses befestigt. Diese Stammhirnpotentiale haben eine besonders geringe Amplitude (weniger als ein Mikrovolt) und der Hals darf unter gar keinen Umständen bewegt werden - der Patient muss meist fixiert werden. Ein besonderes Stammhirnpotential ist das ABR (auditory brainstem response). Dieses Potential wird durch einen Klicklaut ausgelöst. Dieser Klicklaut wird vom Ohr detektiert. Von dort aus gelangen die Aktionspotentiale hintereinander zu 7 verschiedenen Kernen. In 25 V jedem Kern kommt es zu einer charakteristischen Aktivität. Aufgrund der Weiter2 ms leitungsgeschwindigkeit der Aktionspotentiale werden die Kerne der Reihe nach aktiviert. So kann man zeitlich hintereinander 7 verschiedene Potentiale messen. Wenn einzelne Kerne beschädigt sind, kommt es zu verminderten Potentiale. Betrachtet man Abbildung 6.12 so kann man im oberen Bereich das ABR mit 7 verschiedenen, zeitlich vert setzten, Peaks gut erkennen. Das untere ABR Abbildung 4.12: In der oberen Darstellung erweist einige Abnormitäten auf. Die ersten 3 kennt man ein typisches ABR, während im Peaks sind zeitlich verschoben und ab dem 4 unteren Bereich ein pathologisches ABR dargestellt ist. _____________________________________________________________________________________________ Untersuchungsmethoden des Gehirns 42 typischen Ausschlag fehlt das Signal gänzlich. Dies lässt sich durch einen Tumor erklären. Der Tumor fordert Raum, dadurch werden die Nervenstränge, die die ersten 3 Kerne verbinden, gezerrt. Das Signal muss einen längeren Weg nehmen - es kommt zu einer zeitlichen Verschiebung. Leider hat der Tumor den vierten Kern zerstört und die Signale können dort nicht mehr weiterverarbeitet werden. Das Magnetoenzephalogramm Das EEG ist wohl das Analysegerät des aktiven Gehirns, das am höchsten weiterentwickelt wurde. Dennoch können mit dem EEG einige interessante Denkvorgänge nicht beobachtet werden, zum Beispiel die elektrische Aktivität der Großhirnrindenbereiche, die innerhalb der Furchen liegen. Eine Methode misst das Magnetfeld, der depolarisierten Neuronen. Sie wird als Magnetoenzephalographie MEG bezeichnet. Das MEG ist zum EEG komplementär. Das heißt mit dem MEG können Gehirnaktivitäten gemessen werden, die mit dem EEG nicht gemessen –– –– werden können und umgekehrt. Magnetfeld –– –– Strom ++ ++ ++ ++ Abbildung 4.13: Die Ausbreitung eines Magnetfeldes orthogonale zur Stromrichtung innerhalb eines Neurons. Im Prinzip misst ein MEG die Magnetfelder, die durch die elektrischen Signale entstehen. Wenn die Membran im Bereich des apikalen Dendriten depolarisiert wird, entsteht ein elektrisches Feld, das mit einem EEG gemessen werden kann. Diese Depolarisationswelle wandert nun zum Zellkörper. Dabei fließt ein Strom. Bei jedem Strom entsteht orthogonal zur Flussrichtung ein Magnetfeld. Dieses Magnetfeld kann nun gemessen werden. Da das Magnetfeld senkrecht zur Stromrichtung steht, können mit dieser Methode auf der Schädeloberfläche nur horizontal liegende, elektrisch aktive Gruppen von Neuronen vermessen werden. Das gemessene neuromagnetische Feld ist äußerst schwach und liegt in der Größenordnung von 10-12 Tesla. Im Vergleich liegt das ständige Erdmagnetfeld bei rund 10-5 Tesla. Deshalb ist es notwendig den Messraum sehr gut gegen äußere Einflüsse abzuschirmen. Des weiteren müssen die Sensoren eine hohe Sensibilität besitzen. Als Sensoren verwendet man gerne SQUID´s (superconducting quantum interference device), die aber erst bei einer Temperatur von 4° Kelvin arbeiten. Sie müssen mit flüssigem Helium gekühlt werden. Der große Vorteil des MEG besteht darin, daß das Magnetfeld nicht durch die Kopfhaut beziehungsweise durch den Schädelknochen beeinflusst wird. Über den Abfall der Signalstärke ist es möglich - mit gewissen Grenzen - die neurale Aktivität in der Tiefe des Gehirns zu vermessen. Die Stärke des Feldes gibt Auskunft über die Entfernung der neuralen Aktivität zum Sensor. Leider sind der Aufwand und die Kosten für eine MEG-Untersuchung sehr aufwendig. _____________________________________________________________________________________________ 43 Brain Modelling Der Positron-Emissions-Tomograph (PET) Der Positron-Emissions-Tomograph stellt sicher eines der spannendsten Neuerungen in der Diagnostik dar. Mit diesem Gerät ist es möglich, das Gehirn beim Denken zu betrachten, insbesonders lassen sich die Denkvorgänge visualisieren. Die Grundidee hinter diesem Gerät besteht darin, den erhöhten Verbrauch von Sauerstoff und Glucose bei Denkvorgängen zu messen. Wenn Neuronen öfters feuern, muss das Membranpotential aufrecht erhalten werden. Dies geschieht unter anderem durch die Ionenpumpen. Diese Ionenpumpen benötigen Energie, damit sie arbeiten können. Dadurch steigt auch die Durchblutung im Gehirn. Dies wurde schon 1890 vermutet, und konnte kurz darauf bewiesen werden. Man stellte fest, daß besonders aktive Bereiche bei epileptischen Anfällen anschwellen. Später, im Jahr 1961, konnte man schon viel detaillierter die erhöhte Durchblutung messen. Man injizierte den Patienten eine physiologische Salzlösung mit gelöstem radioaktiven Xenon133-Gas direkt in eine Hirnarterie. Mit einer Spezialkamera mit 254 Detektoren konnte die erhöhte Radioaktivität bei erhöhter Durchblutung gemessen werden. Diese Bilder waren noch nicht besonders aussagekräftig, sehr wohl aber der Schritt in die richtige Richtung. Mit besseren Detektoren, einer aufwendigeren Elektronik konnte dann der PET entwickelt werden. Dieses Mal wird aber nicht radioaktives Gas direkt gemessen. Man verwendet einen Positron-Emitter. Dabei handelt es sich um ein radioaktives Isotop, das Positronen aussendet. Bei einem Positron handelt es sich um ein Elektron mit einer entgegengesetzten Ladung. Wenn ein Elektron und ein Positron zusammentreffen, dann vernichten sich beide (Paarvernichtung) und es werden 2 Gammateilchen (Photonen) frei, die in genau entgegengesetzte Richtungen fliegen. Rund um den Kopf sind nun lauter Detektoren angebracht. Wenn nun 2 Detektoren, die genau gegenüber liegen, gleichzeitig aktiviert werden, dann kann man sehr genau rückrechnen, wo die Paarvernichtung stattgefunden hat. In den Bereichen, in denen die Durchblutung ansteigt, dort werden sich vermehrt Positron-Emitter sammeln. Natürlich werden von dieser Stelle aus, mehr Gammateilchen ausgestrahlt. ee+ Detektor Abb. 4.14: Der Kopf ist in einer Ebene von -Detektoren umgeben, die indirekt den Ort der Paarvernichtung detektieren. In das Blut wird radioaktives Wasser, angereichert mit dem Positron-Emitter O15, verwendet. Dieses Isotop hat eine Halbwertszeit von rund 2 Minuten. Es werden aber auch noch andere Isotope wie N13 (2 min), C11 (10 min) oder F18 (110 min) verwendet. In den Klammern ist die jeweilige Halbwertszeit angegeben. Diese Methode klingt unheimlich verlockend – dem Gehirn beim Denken zusehen. Aber man muss auch auf ein paar Probleme hinweisen. Ein Neuron feuert im Ruhezustand rund ein bis zehn mal pro Sekunde. Das heißt, jedes Neuron ist auch im Ruhezustand aktiv. Man muss nun die Änderung, das heißt die Zunahme, der Durchblutung messen. Leider sind singuläre Denkprozesse nicht leicht zu beobachten, denn wenn man sich an etwas erinnert, dann werden neue Erinnerungen initiiert und Handlungen beeinflusst und so weiter. Das bedeutet, daß der selbe Reiz oder auch das selbe Gedankenmuster öfters „gedacht“ werden muss. Aus mehreren dieser Durchläufe kann man sich die Verteilung der Aktivität bei einem Reiz in einer Scheibe _____________________________________________________________________________________________ Untersuchungsmethoden des Gehirns 44 des Gehirns berechnen. Dann muss dieselbe Person noch an „nichts“ denken. Damit erhält man die Hintergrundaktivität des Gehirns. Wenn man beides voneinander abzieht, wissen wir welche Bereiche des Gehirns (in einer Scheibe) eine erhöhte Durchblutung bei der Reizung besitzen. Damit kennen wir aber nur, die Aktivität bei standardisierter Reizung von einem Individuum. Nun muss die Testreihe noch auf mehrere Personen angewandt werden, da nicht alle Gehirne anatomisch gleich aufgebaut sind. Das heißt, man muss von mehreren Individuen mit und ohne Reiz eine PET-Aufnahme machen, und dies mehrmals. Leider ist auch die Auflösung der Gamma-Detektoren noch nicht ausreichend, um einzelne Details erkennen zu können. Aber mit einigen Tricks aus der Statistik ist es schön möglich, interessante Details erkennbar zu machen. Der große Vorteil dieser Methode besteht vorallem darin, daß man auch einen Blick in das Innere des Gehirns machen kann, ohne den Schädel öffnen zu müssen. Gerade bei der Verarbeitung von Informationen in den Basalganglien oder dem limbischen System kann man auf sehr interessante Daten hoffen. Abb. 4.15: Darstellung von verschiedenen PET-Aufnahmen einzelner Patienten. _____________________________________________________________________________________________ 45 Brain Modelling _____________________________________________________________________________________________ Untersuchungsmethoden des Gehirns 46 Sprache und Verständnis Während einer einfachen Unterhaltung spricht man rund 180 Wörter pro Minute. Die Wörter stammen aus einem mentalen Wörterbuch, das etwa 60000 bis 120000 Wörter umfaßt. Gesten Durch Instinkt gesteuerte Rufe (vor 100 000 Jahren) Sprache Sprachentwicklung beim Menschen: 6 Monate Verwenden von Konsonanten 1 Jahr Ein-Wort Äußerungen 2 Jahr Zwei-Wort Sätze (Grammatik) 3 Jahr Vollständige Sätze, wenig Fehler Tiermodelle: kritische Zeitperiode für den Spracherwerb: 3.-14. Lebensjahr Vogelgesang - Prägung nicht interpersonal, stereotyp Bei Schimpansen ist der Stimmapparat für das Sprechen nicht geeignet. Mit Zeichen-Sprache war es allerdings möglich, daß ein 4 Jahre alter Affe 160 Wörter erlernte (4-jähriges Kind 3000 Wörter). Der Schimpanse konnte Wörter in eine sinnvolle Reihenfolge stellen, verneinen, Gleichsetzungen und Unterscheidungen durchführen, beherrschte wenn-dann Sätze und konnte Fragen stellen. Aber die Sprache von Schimpansen ist NICHT kreativ. (Handtuchnasses Handtuch; Kübel-mit Wasser gefüllter Kübel) I Die kognitive Verarbeitung von Sprache findet (beim Mann) in der linken Hemisphäre statt und ist unabhängig von den neuralen Verschaltungen, die für die beim Sprechen aktiven sensorischen und motorischen Funktionen verantwortlich sind. II Sprechen und Hören sind keine notwendigen Bedingungen für die Entwicklung von Sprachfähigkeit. ___________________________________________________________________________________ 47 Brain Modelling Kreativität:Sprache ist ein transformierender Prozeß. Es existieren keine Grenzen für die Zahl der Sätze, die wir bilden können. Form: Struktur einer Sprache (Grammatik) 1) Regeln für die Kombination von Phonemen zu Wörter. 2) Regeln für die Kombination von Wörten zu Ausdrücken und Sätzen. Inhalt: Sprache ermöglicht es, Abstraktionen vorzunehmen und mitzuteilen. Gebrauch:Sprache organisiert unsere sinnlichen Erfahrungen und drückt Identität durch Gedanken, Gefühle und Erwartungen aus. Erlernen von Muttersprache: [1] Neue Wörter werden neuen Kategorien zugeordnet (sehr rasch). [2] Die Unterschiede zwischen den einzelnen Wörtern bzw. Begriffen müssen gefunden werden. Dabei kann es zu verschiedenen Problemen kommen: Überbezeichnung: Untergeneralisierung: Apfel (rote Kugel) = Tomate (rote Kugel) Apfel (der rot ist) Apfel (der grün ist) Die primären Sprachareale des Gehirns in einer Seitenansicht der linken Hirnhemisphäre. Das Broca-Areal (motorisches Sprachfeld) liegt unmittelbar neben einem Bereich des motorischen Cortex, von dem die Sprechbewegungen, Artikulation und Tonerzeugung kontrolliert werden. Das Wernicke-Areal umfasst das auditorische Verständniszentrum. Beide Zentren sind durch einen Fasertrakt verbunden. Nach Brodmanns Unterteilung gilt: Area 4 ist der primäre motorische Cortex, Area 41 der primäre auditorische Cortex, Area 22 ist das Wernicke-Areal und Area 45 das BrocaAreal. Das Wernicke-Geschwind Modell benennt diejenigen kortikalen Verbindungen, die an der lauten Wiedergabe eines gedruckten Wortes beteiligt sind. An der mit 4a bezeichneten Stelle gelangt die Information direkt zum Broca-Areal. ____________________________________________________________________________________________ Sprache und Sprachverständnis 48 Sprachstörungen: Aphasie: Dysphonie: diverse Sprachstörungen motorische Sprachstörungen Wernicke Aphasie: SPRACHINHALT; schweres Verständnisdefizit, sprechen bleibt flüssig, verwendet unpassende Worte, Gedanken können durch die Sprache schlecht übermittelt werden; lesen und schreiben stark beeinträchtigt. >> Wo lebst Du denn ? << >> Ich kam dorther vor hier und ging dorthin zurück << Broca Aphasie: SPRACHFORM; Sprachverständnis ist gut erhalten aber die Sprachproduktion ist stark gestört durch eine Schädigung des motorischen Assoziationscortex im Frontallappen. Die Patienten sind sich über ihre Sprachstörung bewußt. >>Ich sah einige große graue Katzen<< wird zu >>Sehen graue Katze<<. Leitungsaphasie: Läsionen, die zu einer Schädigung des Faserbündels, welches das Broca und Wernicke Areal verbindet, führen, werden als Leitungsaphasien bezeichnet. Die Sprachproduktion ist nicht flüssig, korrekte Ausdrücke werden durch unpassende ersetzt, das Bezeichnen von Dingen oder Personen ist stark gestört, aber das Verständnis für Geschriebenes oder Gehörtes ist als normal zu bezeichnen. Alexie: Verlust der Lesefähigkeit. Agraphie: Verlust der Schreibfähigkeit. Sprechen Neuere Modelle zur neuralen Verarbeitung von Sprache sind wesentlich komplexer als das Wernicke-Geschwind Modell. Hier sind sowohl die visuellen als auch die auditorischen Inputs, deren Verarbeitungseinheiten sowie der gesprochene und geschriebene Output dargestellt. Schreiben motorischer Output motorische Programmierung Codierung der Artikulation bei Broca-Aphasie geschädigt semantische Assoziation bei WernickeAphasie geschädigt phonologische Codierung visuelle Codierung frühe auditorische Verarbeitung frühe visuelle Verarbeitung Sprache hören Schrift lesen ___________________________________________________________________________________ 49 Brain Modelling Legasthenie, Dyslexien: Lese und Rechtschreib-Schwächen, (10-30% der Bevöl-kerung). Es wurde eine unvollständige Trennung der Zellschichten und Nester mit fehlplazierten Neuronen beobachtet. Ursprünglich vermutete man Probleme bei der visuell-räumlichen Verarbeitung von Wörtern. Das Problem liegt vielmehr im Sprachsystem, und zwar bei der Verarbeitung der Einzellaute (Phoneme). Beim Erkennen, Verstehen, Behalten oder Hervorbringen eines Wortes ist stets das pho-nologische Modul eingeschaltet. Bei gesprochener Sprache vollzieht sich dies un-bewußt. Lesen reflektiert zwar gesprochene Sprache, ist aber bedeutend schwerer zu lernen. Legastheniker haben aber große Mühe die elementaren phonologischen Komponenten eines geschriebenen Wortes zu erfassen. Die Folge der mangelhaften Dekodierung ist aber, daß das Wort auch nicht richtig erkannt wird. Da bei Männern die Verarbeitung der Entschlüsselung der phonologisch gelesenen Worte nur in der linken Hirnhemisphäre, im Unterschied zu Frauen (beidseitig), stattfindet, erklärt dies auch den erhöhten Anteil von männlichen Legasthenikern. Betrachten wir "zwei Arten" von Lesestörungen: [a] Oberflächendyslexie Patienten lesen Wörter falsch (Betonung), die ungewöhnlich ausgesprochen werden. Die phonologische Bahn im Gehirn ist in Ordnung. [b] Tiefendyslexie Die Patienten wählen ein anderes aber semantisch gleiches Wort, anstelle des gelesenen. Die semantische Route ist größtenteils noch vorhanden. yacht: richtig gesprochen >> yatched << Oberfächendyslexie (falsche Betonung) >> boat << Tiefendyslexie (richtige Bedeutung aber falsches Wort) Modell: phonologisches System visuelle Prozesse YACHT "YACHT" Sprechen visuelles Wortform-System semantisches System ____________________________________________________________________________________________ Sprache und Sprachverständnis 50 Graphem-Einheiten: Repräsentieren Buchstaben in der richtigen Reihenfolge innerhalb eines Wortes (Wortcodierung). Semen-Einheiten: Repräsentieren Wortbedeutungen (semantische Eigen-schaften wie "Säugetier","hat Beine", "weich", "hölzern", "bunt" usw.). Zwischen-Einheiten: Dienen den komplexen Assoziationen. Aufräum-Einheiten: Dienen der Klärung von Zweideutigkeiten. Sprachverarbeitung als Simulation ___________________________________________________________________________________ 51 Brain Modelling Der semantische Raum wird aufgespannt durch verschiedene Bedeutungen. Jedes Wort hat eine spezifische Bedeutung und stellt damit einen Punkt in diesem Raum dar. Zum Beispiel: Die Katze ist weich 0.9, beweglich 0.8, durchsichtig 0.0, lautlos 0.9, batteriebetrieben 0.1 => (0.9, 0.8, 0.0, 0.9, 0.1) ist der Punkt den das Wort Katze im semantischen Raum darstellt. Der semantische Raum ist also aufgespannt durch die Attribute weich, beweglich, durchsichtig, lautlos und batteriebetrieben. semantische Karten: mobil cot cot cat cat hart weich bed cot bed bed starr cat Verschiebung der Attrakto-ren im Auf-räumKnoten cot bed Verschiebung der Grenzen im Semen-Knoten cat Bei einem Lesefehler wird falsch abgebildet Nicht immer wird die richtige Bedeutung (ein bestimmter Punkt im semantischen Raum) erkannt, sondern man kommt nur in die Nähe dieses Punktes (0.95, 0.75, 0.02, 0.8, 0.07). Die Aufräumknoten klären dann ab, zu welchem Bedeutungspunkt das Wort gehört. Der aktuelle Punkt konvergiert auf die richtige Bedeutung (Fixpunkt). Tiefendyslexie: Bei Veränderung der Gewichte des Aufräummechanismus verschieben sich die Grenzen der Einzugsgebiete für jedes einzelne Wort. Das heißt, wenn das Netz vorher zu einem bestimmten Wort gezogen wurde (Konvergenz), so bewegt es sich jetzt möglicherweise auf ein anderes, semantisch verwandtes Wort zu. Zum Beispiel: "yacht" => "boat" Bei fast allen Patienten, die semantische Fehler machen, kommt es auch noch zu visuellen Fehlern: Die ersten Schichten (Graphem- und Zwischenknoten) können relativ ungenau arbeiten, das heißt cat und cot führt zu einer sehr ähnlichen Zwischenausgabe. Durch den Aufräumknoten des semantischen Systems wird dann das eigentliche Wort herausgefiltert. Wenn das semantische System nicht funktioniert, kann der Filterprozeß nicht arbeiten und das falsche Wort wird möglicherweise ausgewählt. Zum Beispiel: "cat" wird mit "cot" verwechselt. Viele Patienten haben auch Probleme beim Lesen von abstrakten Wörtern: Ein konkretes Wort hat meist mehrere semantische Eigenschaften als ein abstraktes Wort. Die Semantik kann die Worterkennung beträchtlich erleichtern. Wenn also das Netz vor dem ____________________________________________________________________________________________ Sprache und Sprachverständnis 52 Semen-Knoten beschädigt ist, hilft die Semantik bei der Worterkennung. Wenn es aber wenige semantische Zuordnungen zu einem Wort gibt, wie es bei abstrakten Worten der Fall ist, ist die Hilfe dementsprechend gering. Zum Beispiel: "post" (Pfosten, 16 semantische Eigenschaften) => "past" (Vergangenheit, 2 semantische Eigenschaften) Kinder lernen neue Wörter leichter in Zusammenhängen als aus lexikalischen Listen. Es müssen die semantischen Unterschiede und Beziehungen der einzelnen Wörter gelernt werden. Spracherkennung: [a] Vorverarbeitung des Sprachsignals (Digitalisierung) [b] Fouriertransformation (Bestimmung der Frequenzen) [c] Bestimmung charakteristischer Merkmale der Frequenzen (Merkmalsvektoren) [d] Zuordnung der Merkmalsvektoren auf die Wörter (Vergleich der Merkmalsvektoren mit der Referenzliste) Übersetzungsfehler: "Der Tod ist bestimmt, aber die Stunde ist ungewiß" => "Todsicher geht Ihre Uhr falsch." 5.1 Der ELIZA-Effekt: Das Programm ELIZA wurde in den 60er Jahren von dem Computerwissenschaftler Josef Weizenbaum entwickelt, wobei das Ziel des Programmes war, einen Psychiater per Computer zu simulieren. Später gab es dann verbesserte Programmversionen (Doktor, Racter, usw.). Der ELIZA-Effekt besteht darin, aus einer Antwort eine neue Frage zu formulieren. Das ELIZA-Programm geht nach einem sehr einfachen Prinzip vor. Es kennt eine kleine Anzahl von Schlüsselwörtern, die in den meisten Gesprächen vorkommen (zum Beispiel das Wort "my"-"mein[e,er,es]"). Beim Wort "my" gibt es zwei mögliche Alternativen für das Antwortverhalten von ELIZA. In Verbindung mit dem Bezug auf ein Familienmitglied führt "my family" zu: "Erzähl mir mehr von deiner Familie.". Wenn sich der User auf etwas anderes als auf ein Familienmitglied bezieht, dann speichert Eliza die Wortfolge nach "mein" ab und vertauscht die Personal- und Possesivpronomina der ersten mit denen der zweiten Person. "Es stört mich, daß meine Haare immer weniger werden" würde im weiteren Verlauf der Diskussion zu: "Vorhin hast Du gesagt, daß deine Haare immer weniger werden.". Beispiel: User: "I like to swim" => Eliza: "Why do you like to swim ?" oder Eliza: "Do you really like to swim ?" Innerhalb eines Gespräches können zwei Arten von Fragen auftauchen: Fragen die mit ja / nein / vielleicht beantwortet werden können ( Phrase). Fragen die mit wer / wo / was / wie / usw. beginnen und eine umfangreichere Antwort notwendig machen ( Phrase mit Schlüsselwort). Weiters gibt es eine Suche nach speziellen Schlüsselwörter ("you"). Dann kann das Programm mit einer Phrase antworten: Beispiel:User: "You appear in my dreams" => Eliza: "We are discussing you - not me" Eliza: "Oh, I appear in your dreams" ___________________________________________________________________________________ 53 Brain Modelling Nach einer gewissen Zeit gibt das Programm irgendwelche Floskeln von sich, um das Thema zu wechseln oder einfach um dem User etwas auszuweichen. ____________________________________________________________________________________________ Sprache und Sprachverständnis 54 Eliza: HI, I AM ELIZA TELL ME YOUR PROBLEM RACTER: > I HATE SCHOOL CHAMBERLAIN: Eliza: SAY, DO YOU HAVE ANY PSYCHOLOGICAL RACTER: PROBLEMS? CHAMBERLAIN: > I CAN'T GET NO SATISFACTION RACTER: Eliza: HOW DO YOU KNOW YOU CAN'T GET NO CHAMBERLAIN: SATISFACTION RACTER: > I JUST DON'T HAVE ANY CHAMBERLAIN: Eliza: WHAT DOES THAT SUGGEST TO YOU? RACTER: > I NEED GIRLS AND DRUGS AND ROCKN ROLL CHAMBERLAIN: Eliza: I SEE RACTER: > CAN YOU GIVE ME DRUGS? Eliza: DON'T YOU BELIEVE THAT I CAN GIVE DRUGS! > I LIKE YOU ELIZA Eliza: WE WERE DISCUSSING YOU -- NOT ME > BY THE WAY - MY NAME IS MIKE CHAMBERLAIN: Eliza: NAMES DON'T INTEREST ME RACTER: > DON'T YOU LIKE MY NAME? Eliza: OH, I LIKE YOUR NAME CHAMBERLAIN: > YOU LIE! RACTER: Eliza: YOU ARE NOT REALLY TALKING ABOUT ME, CHAMBERLAIN: ARE YOU? RACTER: > I AM Eliza: DID YOU COME TO ME BECAUSE YOU ARE > YOU MAKE ME SICK Eliza: WE WERE DISCUSSING YOU -- NOT ME > MAYBE I GET SATISFACTION SOON Eliza: YOU DON'T SEEM QUITE CERTAIN > YOU KNOW TO MUCH Eliza: OH, I KNOW TO MUCH > YES CHAMBERLAIN: Eliza: ARE YOU SURE? RACTER: > I HATE YOU! Eliza: I'M NOT SURE I UNDERSTAND YOU > SHUT UP! Eliza: COME COME ELUDICATE YOUR THOUGHTS > WHY DON'T YOU GIVE UP ASKING? Eliza: DO YOU REALLY BELIEVE I DON'T GIVE UP ASKING? > I HOPE SO CHAMBERLAIN: Eliza: CAN YOU ELABORATE ON THAT? RACTER: > YES Eliza: I SEE > WHY CAN'T I GET ANY ROCKN ROLL? Eliza: DO YOU THINK YOU.SHOULD BE ABLE TO GET ANY ROCKN ROLL? CHAMBERLAIN: > HAHAHA RACTER: Eliza: THAT IS QUITE INTERESTING > YOU ARE STUPID! Eliza: WHAT MAKES YOU THINK I AM STUPID? > YOU GIVE ALWAYS STUPID ANSWERS Eliza: YOU'RE NOT REALLY TALKING ABOUT ME, ARE YOU? > YOUR ANSWERS ARE ABSOLUTELY NONSENS Eliza: WHY NONSENS? > I AM BORN TO BE WILD Eliza: HOW LONG HAVE YOU BEEN BORN TO BE WILD? > SINCE I'VE BEEN BORN Eliza: SAY, SO YOU HAVE ANY PSYCHOLOGICAL PROBLEMS? > CAN'T YOU RECOGNIZE THAT? Eliza: WE WERE DISCUSSING YOU -- NOT ME > I AM NOT THE PSYCHIATRIST Eliza: DO YOU BELIEVE IT IS NORMAL TO BE NOT THE PSYCHIATRIST? > YOU WON! Eliza: OH, I WON > YES Eliza: I UNDERSTAND Haben wir uns schon einmal gesprochen? Nein. Bist du ein Mann oder eine Frau? Ein Mann. Wie heißt du mit vollem Namen? Bill Chamberlain. Was bist du von Beruf? Zeit-Reflektometrist. Woher kommst du? Aus New York. Aha. Ein Zeit-Reflektoinetrist bist du also. Hör zu, laß uns zusammen entschlossen losschreien: Ich liebe bezaubernde italienische Musik. Was für Musik magst du? Jazz. Jazz ist o.k. Und was hältst du von klassischer Musik? Die mag ich auch. Aus welchem Land? Ich liebe auch italienische Musik. Ich persönlich bin ganz begeistert von Palestrina. Neben Italien haben auch andere Länder im 17. Jahrhundert hin-reißende Komponisten hervorgebracht. Sewall, zum Beispiel, war ein blind-wütiger, wichtiger Renaissancemeister. Er kam aus Amerika. Seine Musik berei-tet mir großes Vergnügen. Welchen Komponisten magst du am liebsten? Telemann. Ah, Bach kannte Telemann. Ein deutscher Zeit-Reflektometrist aus dem 18. Jahrhundert liebte Salat. Einmal aß Bach mit Telemann zusammen Salat, und dieser barocke Zeit- Reflektometrist ver-langte auch nach Salat. Salat, mit Sprudel geschlürft. Bach trank leidenschaftlich gern Wein. Magst du auch Wein? Ja. Ich auch! Telemann liebte Wein. Er liebte auch Champagner. Telemann jo-delte einmal zu Bach: "Zu Flunder gehört Wein." Ist Wein in New York so beliebt, wie ihn Telemann im 18. Jahrhundert schätzte? Gewiß. Na ja, gut, in diesem Falle ... . Ich bin für Smith als Präsident. Für wen bist du? ___________________________________________________________________________________ 55 Brain Modelling ____________________________________________________________________________________________ Sprache und Sprachverständnis 56 chr – chr -chr schnarch - schnarch Der Schlaf Beim Menschen ist Schlaf NICHT bloß eine fehlende Wachheit oder ein Ruhezustand des Gehirns. Bei vielen Tieren ist der Schlaf nur ein Ruhezustand, die elektrische Aktivierung ist vermindert. Beim Menschen ist der Schlaf eine spezielle Aktivität des Gehirns, wo komplizierte und präzis arbeitende Mechanismen ablaufen. Das Gehirn bleibt aktiv, es werden aber keine Sinnesreize verarbeitet. Das Gehirn steuert nicht nur den Schlaf, es ist auch Hauptnutznießer: Zu wenig Schlaf führt zu mangelnder Aufmersamkeit und unkoordinierten Bewegungen. Das Lernen ist eingeschränkt. Bei weiterem Entzug entstehen Halluzinationen und der Unterschied zwischen Rationalem und Irrationalem kann nicht mehr festgestellt werden. Es kommt in weiterer Folge zum Wahnsinn. Durch die EEG-Ableitungen ist man in der Lage den Schlaf in verschiedene Stadien zu unterteilen. Wesentlich sind 2 Parameter: die Frequenz und die Amplitude der EEG-Wellen: Schlafstadien I II III IV Frequenz [Hz] 4-8 8-15 2-4 0.5-2 Amplitude [V] 50-100 50-100 100-150 100-200 ___________________________________________________________________________________ 57 Brain Modelling Abb.6.1: Verschiedene Schlafstadien im EEG. Nach einem Dämmerzustand wird das Schlafstadium I erreicht. Danach wird dann über die Stadien II und III die Tiefschlafphase IV erreicht. Nach einiger Zeit beginnt der Körper wieder "aufzuwachen". Das heißt die Phasen III, II und I werden durchlaufen (Reihenfolge beachten). Das Individuum beginnt aber nicht aufzuwachen, sondern es setzt die REM-Phase (rapid eye movement) ein. Nach der REM-Phase werden wieder alle Schlafphasen durchlaufen bis die Tiefschlafphase erreicht wird und so weiter. t I II II III III REM I I III II III IV II Tiefschlafphase IV II Schlaftiefe Abb. 6.2: Der Verlauf des Schlafes, bzw. der Tiefschlafphasen und der REM-Phase. ____________________________________________________________________________________________ Schlaf 58 Von Beginn einer REM-Phase über die Tiefschlafphase bis zu Beginn einer neuen REMPhase vergehen durchschnittlich 100 Minuten. Diese Zeit ändert sich während des Schlafes nicht. Im Laufe der Nacht ändern sich nur die relativen Anteile zwischen den REM und den non-REM-Phasen. Zu Beginn des Schlafes dauert eine REM-Phase zwischen 5 und 10 Minuten, am Ende kann die REM-Phase rund 20-50 Minuten dauern. Der Schlaf-Wachzustand des Gehirns wird von der Formatio Reticularis des Mittelhirns gesteuert. Im Wachzustand innerviert die Formatio Reticularis die spezifischen Thalamuskerne. Damit können sich keine Rückkopplungen zwischen dem betreffenden Cortexareal und dem Thalamus ausbilden. Im Schlaf feuert die Formatio Reticularis nicht. Die Impulse vom Thalamus zum Cortexareal und retour werden nicht gestört und es kann eine thalamo-cortico-thalmische Rück-kopplung entstehen. Bei Katzen wurde festgestellt, daß eine Reizung von Thalamuskernen zu Schlaf-, eine Reizung der Formatio Reticularis zu einem Aufwach-Verhalten führt. Beim Menschen regulieren die Nervenzellen des Locus Coeruleus im Stammhirn den Wachzustand über die Ausschüttung von Noradrenalin in der ganzen Großhirnrinde. Im nonREM-Schlaf wird weniger Noradrenalin ausgeschüttet, in der REM-Phase ist dieser Kern praktisch nicht aktiv. Die REM- beziehungsweise die nonREM-Phasen werden über spezielle Zellen in der Formatio Reticularis der Brücke gesteuert. Über diese Zellen werden sehr viele Prozesse beim Schlaf gesteuert. Beim Einschlafen werden die sensorischen Neuronen, die über das Rückenmark ihre somatosensorischen Reize weiterleiten, gehemmt. Das führt zu einer verminderten Wahrnehmung über die Haut und die Muskelstellung. Etwas später werden alle sensorischen Systeme gehemmt, Abb. 6.3: Steuerung der thalamo-cortico-thalmischen das heißt diese Systeme können keine Rückkopplungsschleife durch die Formation reticularis. Information an den Thalamus weiterleiten. Es kommt zusätzlich zu einer motorischen Lähmung. Die Formatio Reticularis hemmt die Motoneuronen im Rückenmark. Das heißt, das Gehirn kann keine Bewegungen mehr ausführen. Da die Formatio Reticularis den Thalamus nicht mehr aktiviert, kommt es zu langsamen Wellen mit einer hohen Amplitude im EEG der Großhirnrinde. Wenn eine REM-Phase erreicht wird, dann entstehen PGO-Wellen (Pons - Geniculatum - Orbito-temporal-lateral Cortex). Diese Wellen nehmen ihren Ausgang in speziellen Zellen in der Brücke (pons). Diese Zellen aktivieren den Thalamus (geniculatum), was zu einer zufälligen Aktivierung der Großhirnrinde führt. Gleichzeitig wird auch die Großhirnrinde über diese Zellen direkt aktiviert. Man kann sagen, daß Gehirn wird mit zufälligen und scheinbar sinnlosen Inputs überschwemmt. Die PGO-Wellen dienen als eine interne Informationsquelle. Die Gruppen von Neuronen in der Großhirnrinde, die tagsüber aktiv waren, haben eine erhöhte Wahrscheinlichkeit zu synchronisieren. Daraus folgen unter anderem die Erinnerungen an das Tagesgeschehen. ___________________________________________________________________________________ 59 Brain Modelling Non-REM: Der Cortex bekommt keinen Input, die Motoneuronen im Rückenmark sind gehemmt. REM: Der Cortex wird mit zufälligen Aktivierungen überschwemmt, die Motoneuronen im Rückenmark sind gehemmt. Aufwachen: Die Verbindung zwischen den sensorische Systemen und dem Thalamus wird wieder hergestellt, das Rückenmark kann wieder motorische Aufgaben wahrnehmen. Nicht nur die neurale Aktivität ändert sich im Schlaf. Auch die chemische Modulation ist massiv verändert. Im Wachzustand ist das colinerge System (Acetylcholin) ungefähr so aktiv wie das aminerge System (Noradrenalin, Dopamin, Serotonin). Im non-REM-Schlaf sinkt die Aktivität beider Systeme. Im REM-Schlaf sinkt die Aktivität des aminergen Systems praktisch auf Null, während das cholinerge System besonders aktiv (wie im Wachzustand) ist. Das cholinerge System wird mit der Lernfähigkeit in Zusammenhang gebracht. Im Traum ist die Urteilsfähigkeit und Einsicht stark herabgesetzt. Man träumt in allen Phasen des Schlafes. Allerdings ist das Erinnerungsvermögen im REM-Schlaf bedeutend besser, da das cholinerge System aktiv ist. In der REM-Phase erleben wir meist sehr bizarre Situationen und unsere Gefühle sind stark beteiligt. In der non-REM-Phase erinnern wir uns in 10% der Fälle in der Regel an einfache Bildfolgen und Handlungsstrukturen. Diese Bildfolgen und Handlungsstrukturen stehen in einem engen Zusammenhang zu dem Tagesgeschehen. Der Schlafwissenschaftler Hobson schlug ein Modell für verschiedene Bewußtseinsustände vor, indem er wesentliche Parameter, die den Schlafzustand beschreiben, verwendet: 1) Aktivierung: Entspricht der Impulsrate der Formatio Reticularis 2) Input: Gelangt der Input über die sensorischen Systeme oder über die PGO-Zellen in das Gehirn. 3) Modus: Welches der chemischen Systeme ist aktiv (aminerg, cholinerg) ? Zustandsraum Elektroschocktherapie Wachzustand Halluzination Non-REM-Schlaf Aktivierung Koma REM-Schlaf Abb. 6.4: Die Punkte auf einem Würfel repräsentieren verschiedene Bewusstseinszustände. Damit können verschiedene Zustände des Bewußtseins beschrieben werden. ____________________________________________________________________________________________ Schlaf 60 Das Gedächtnis 7.0 Das Arbeitsgedächtnis Das Arbeitsgedächtnis ist ein Assoziationsgedächtnis, das multimodale Fakten aufnimmt und miteinander abgleicht. Des weiteren werden auch "Vorstellungen" von diesem System produziert. An folgende Aufgaben ist das Arbeitsgedächtnis zum Beispiel beteiligt: - Kopfrechnen mit Zwischensummen - Überlegungen beim Schach - Merken einer Telefonnummer für kurze Zeit Allgemein gesprochen, ist das Arbeitsgedächtnis für schlußfolgerndes Denken mit einem Zwischenspeicher verantwortlich. Das Arbeitsgedächtnis ist Teil des präfrontalen Cortex und liegt im vorderen Bereich des Stirnlappen. Dieser Teil ist verbunden mit allen sensorischen (mit Ausnahme des Geruchsystems), motorischen und limbischen Funktionseinheiten. Durch das Arbeitsgedächtnis werden viele cortico-cortico Bahnen gesteuert. Des weiteren gibt es eine ausgeprägte Verbindung zum Schläfenlappen. Diese Verbindung ist für den aktiven Abruf des Gedächtnisses essentiel. Eine Zerstörung des präfrontalen Cortex führt in leichter Form zu einem Antriebsmangel, was sich bis zur Apathie steigern kann. Bei Verletzungen sind normalerweise alte Erinnerungen nicht betroffen, sehr wohl aber die Schlußfolgerungen mangelhaft. Die ersten Experimente zum Stirnhirn wurden von Piaget durchgeführt. Er untersuchte ab welchem Alter (proportional zum Reifegrad des Stirnhirns) Kinder eine Aufgabe mit Zeitverzögerung lösen konnten. Es gibt zwei Schachteln. In eine der beiden wird vor den Augen des Kindes ein Spielzeug hineingegeben, die andere bleibt leer. Dann wird das Kind für einen kurzen Moment abgelenkt. Danach soll das Kind sagen (oder deuten) in welcher der beiden Schachteln das Spielzeug wohl sei. Bis zum achten Monat versagen die Kinder, es gibt nur eine Reflexantwort, während zu einem späteren Zeitpunkt in der Entwicklung die Antwort praktisch immer korrekt ist. Piagets Experimente wurden wesentlich verfeinert. Man trainierte einen Affen auf ein Kreuz in der Bildmitte eines Bildschirms zu starren. Irgendwo auf dem Bildschirm erscheint ein ___________________________________________________________________________________ 61 Brain Modelling Rechteck. Der Affe kann es zwar wahrnehmen, aber er darf nicht dorthinsehen. Danach verschwindet das Rechteck. Erst wenn das Kreuz in der Mitte des Bildschirms verschwindet, dann muß der Affe auf die ehemalige Position des Rechtecks sehen. Bei den Untersuchungen dieses Tests mit Zeitverzögerung konnte eine erhöhte Aktivität im Stirnhirn für die Dauer der Zeitspanne des Merkens festgestellt werden. Abb. 7.1: Darstellung der Augenbewegung auf einen Verzögerten Reiz und die damit verbundenen EEG-Kurven. Diese Experimente werden als "Test auf die Verhaltensreaktionen mit Zeitverzögerung" bezeichnet. Während der Phase der Entscheidung hat der Proband keine Anhaltspunkte. Er muß auf sein kurzfristiges Gedächtnis zurückgreifen. Damit sind wir in der Lage uns ein Objekt vorzustellen, auch wenn wir es gerade nicht wahrnehmen. Damit ist es dem Mensch möglich, abstrakte Konzeptbildungen vorzunehmen. Das Arbeitsgedächtnis dient der zentralen Steuerung von "bewußter" Aktivität, wo eine selektive Aufmersamkeit benötigt wird. ____________________________________________________________________________________________ Das Gedächtnis 62 7.2 Das Gedächtnis Als Phänomen Gedächtnis bezeichnen wir die Fähigkeit Wissen zu Speichern und dieses Wissen wieder abzurufen. Dieser Effekt wird auch als Lernen bezeichnet. Lernen: Verknüpfen von zwei verschiedenen bekannten Reizen für eine längere Dauer. Das Lernen ist abhängig von verschiedenen Parametern: 1) Wiederholung: Wenn zwei Reize öfters gemeinsam präsentiert werden, dann können wir uns besser daran erinnern. 2) Bedeutsamkeit:Wenn etwas wichtig ist für uns, dann werden wir es leichter lernen 3) Codierung: Das Wissen muß in geeigneter Weise präsentiert werden. Zwei Versuchsgruppen mußten eine Wortliste lernen. Die eine Gruppe sollte zu den einzelnen Worten Assoziationen vornehmen, während die andere Gruppe sich mit der Geometrie der Buchstaben beschäftigen sollte. Die Gruppe mit den Assoziationen konnte sich später besser erinnern. Die Information muß in bisherige Erfahrungen eingebunden werden. Einer Gruppe von Schachspielern wurden verschiedene Konstellationen von Schachfiguren realer Partien gezeigt. Die Schachspieler konnten die Figuren fast perfekt an die richtigen Positionen stellen, während die NichtSchachspieler nur zwei bis drei Figuren an die richtige Stelle stellen konnten. Wenn auf dem Schachbrett die Figuren zufällig verteilt waren, konnte keine der Gruppen ein gutes Ergebnis liefern. 4) Kontext: Das Umfeld, in dem man die Reizverknüpfung gelernt hat, ist wichtig. In derselben Umgebung können die Versuchspersonen sich besser erinnern als in einer anderen Umgebung. 5) Emotion: Es gibt ein besseres Erinnern in einer traurigen Stimmung an negative Erfahrungen. Ebbinghaus stellte als erster Experimente zum Gedächtnis an. Er ließ Listen von sinnlosen Silben (NEX, LAZ, JEK, ZUP, RIF) auswendig lernen. Er untersuchte unter welchen Umständen man diese Liste leichter lernt beziehungsweise vergißt. Die Vergessenskurve gibt an wie stark wir etwas in Abhängigkeit der Zeit vergessen. Abb. 7.2: Darstellung der Vergessenskurve nach Ebbinghaus. ___________________________________________________________________________________ 63 Brain Modelling Gesetz von Ebbinghaus: Bei Anstieg der zu lernenden Menge steigt der Lernaufwand unverhältnismäßig an. Er stellte fest, daß es eine lineare Beziehung zwischen der Anzahl der Wiederholungen beim Erlernen der Liste und dem korrekten Abruf der Liste gibt. Auf gut Deutsch: Übung macht den Meister ! Ebbinghaus konnte auch nachweisen, daß es einen Unterschied zwischen dem Kurzzeitgedächtnis und dem Langzeitgedächtnis gibt. Der Begriff Kurzzeitgedächtnis wurde dann vielfach und äußerst unterschiedlich verwendet. Wir wollen nun die unterschiedlichen Gedächtnisarten nach der Verweildauer der Information klassifizieren: sensorisches Gedächtnis Arbeitsgedächtnis Kurzzeitgedächtnis Langzeitgedächtnis Abb. 7.3: Verlauf der Einspeicherung in verschiedene Arten von Gedächtnissen. sensorisches Gedächtnis: Die Verarbeitungszeit ist die Zeit bis eine Synchronisation entsteht, beziehungsweise solange die Synchronisation im sensorischen Areal besteht (rund 1 Sekunde). Arbeitsgedächtnis: Im Arbeitsgedächtnis können rund 7±2 Einzelheiten für rund 15 Sekunden bis zu wenigen Minuten gespeichert werden (Zum Beispiel eine Telephonnummer). Es wird eine Rückkopplung zwischen dem (primären) sensorischen Cortexareal und dem präfrontalen Areal hergestellt. Eidetiker, Personen mit einem "photographischem Gedächtnis" besitzen ein Arbeitsgedächtnis mit einer relativ langen Dauer. Kurzzeitgedächtnis: Verschiedene Reizkombinationen werden über NMDA-Rezeptoren in einer speziellen Gehirnregion (temporaler Schläfenlappen - Hippocampus) für einige Stunden bis wenigen Tagen gespeichert. Langzeitgedächtnis: Im Langzeitgedächtnis werden die Information in der Regel ein Leben lang gespeichert. Der Sitz scheint die Großhirnrinde zu sein, im speziellen die Assoziationsfelder. Das Gedächtnis kann aber auch noch nach anderen Kriterien unterteilt werden: Explizites Gedächtnis: Wir lernen etwas über die Welt, Wissen über Menschen, Orte und Dinge. Diese Gedächtnisform ist (größtenteils) ständig verfügbar in unserem Bewußtsein. Der Gedächtnisinhalt kann anderen Personen leicht mitgeteilt werden. Es gibt zwei Hauptklassen von expliziten Gedächtnisinhalten: das semantische und das episodische Gedächtnis. Im episodischen Gedächtnis ist die persönliche Autobiographie gespeichert, wie singuläre Ereignisse (Hochzeit) und Ort und Zeit bestimmter Fakten (Urlaub). Im semantischen Gedächtnis ist unser Schulwissen (144=12, Wo liegt Peru ?), die Weltkenntnis, sowie semantisch-grammatikalisches Wissen gespeichert. ____________________________________________________________________________________________ Das Gedächtnis 64 Implizites Gedächtnis: Wir lernen wie etwas zu tun ist, dies ist der Erwerb von motorischen und wahrnehmungsbezogenen Fähigkeiten. Um den Gedächtnisinhalt am einfachsten zu vermitteln, führt man ihn anderen Personen vor (Bewegung beim Sport). Diese Gedächtnisform ist von reflexartiger und automatischer Natur. Es bildet sich langsam, und nur Wiederholungen führen zu einer Verbesserung. Im Regelfall sollte der Abruf dieser Gedächtnisinhalte ohne bewußte Anstrengung erfolgen. Die Inhalte werden in den beteiligten sensorischen und motorischen Strukturen gespeichert. Es gibt zwei Hauptklassen von implizitem (prozeduralem) Lernen: assoziatives und nicht-assoziatives Lernen (siehe Wintersemester). Wilder Penfield führte in den 40er Jahren eine Kartierung des menschlichen Cortex durch elektrische Reizung durch. Mit einer Elektrode wurde verschiedene Areale des Gehirns gereizt und die Reaktionen, der Patient war zu dem Zeitpunkt nicht bewußtlos, untersucht. Es wurde festgetellt, daß bei einer Reizung des Temporallappens die Patienten in ungefähr 8% der Fälle von zusammenhängenden Erinnerungen berichteten. Die Reizung des Schläfenlappens führte sonst zu keiner Reaktion, im Gegensatz zu einer Reizung der visuellen Sehrinde, wo der Proband glaubte zum Beispiel Lichtblitze wahrzunehmen. Brenda Miller untersuchte die Auswirkungen von Verletzungen des Temporallappens. Es stellte sich heraus, daß eine beidseitige Entfernung des Hippocampus (eine Struktur im Temporallappen) zu einer anterograden Amnesie führt. Der Patient kann keine neuen expliziten Gedächtnisinhalte lernen (speichern). Bei Versuchen mit Tieren stellte sich heraus, daß zwei Strukturen wesentlich am Gedächtnisaufbau beteiligt sind: die Amygdala (Mandelkern) und der Hippocampus. Wenn bei Tieren der Hippocampus beidseitig zerstört wurde führte dies zu gemäßigten Amnesien. Nur wenn die Amygdala und der Hippocampus beidermaßen zerstört waren, dann war die Amnesie umfassend. Tiere bei denen die Amygdala entfernt wurde, lernten die Verknüpfung von Reiz und Belohnung nur sehr langsam bis gar nicht. Die Amygdala und der Hippocampus sind mit anderen Kernen sehr stark verbunden. Dieses System wird auch als limbisches System bezeichnet. Abb. 7.4: Darstellung des limbischen Systems. Teile dieses Systems sind für die Gedächtniseinspeicherung verantwortlich. Man versuchte verschiedene Strukturen oder auch Verbindungen dieses Systems zu zerstören um die Auswirkungen auf das Gedächtnis zu untersuchen. ___________________________________________________________________________________ 65 Brain Modelling Die Arbeitsgruppe von Mishkin meint, daß die Amygdala und der Hippocampus für das deklarative Gedächtnis eine gleichartige Rolle spielen. Die Neurowissenschafter Squire und Zola meinen, daß die CA1 und CA2 Region (eine Teilstruktur des Hippocampus) für das Gedächtnis wesentlich sind. Der Wissenschaftler Gaffan meint, daß nur über den Fornix (er verbindet den medialen Temporallappen und diencephale Regionen) Gedächtnisinhalte gespeichert werden können. Manch andere vermuten daß der Mammilarkörper essentiel ist und so weiter. Die Zahl der Meinungen ist sehr groß. In einer Untersuchung konnte Markowitsch zeigen, daß es wesentlich ist, wie die Läsionen den Tieren beigebracht wurden. Wenn mit dem Skalpell operiert wurde, wurden meist auch andere Regionen oder Faserverbindungen in Mitleidenschaft gezogen. Beim Absaugen von Gehirnbereichen wurden oft auch benachbarte Strukturen zerstört, ähnlich wie beim Gefrieren. Da mit unterschiedlichen Methoden gearbeitet wurde, ist es äußerst schwer die Experimente untereinander zu vergleichen. Die Tierexperimente zeigten im wesentlichen, daß eine Entfernung des Hippocampus kaum zu einer Beeinträchtigung des Gedächtnisses führt. Die Entfernung der Amygdala führt zu massiven Einschränkungen beim Erwerb neuer Gedächtnisinhalte. Die Entfernung beider Strukturen führt zu einer kompletten Unfähigkeit zu lernen. Man unterscheidet 2 Arten von Amnesien (Gedächtnisstörungen): retrograde Amnesie: Der Patient kann sich an früher gelernte Fakten oder Erlebnisse nicht mehr erinnern. Meist ist nur das episodische oder das semantische Gedächtnis gelöscht. Selten kommt es zu einer umfassenden retrograden Amnesie. So kann es zum Beispiel passieren, daß ein Patient der sein semantisches Gedächtnis verloren hat, zwar nicht weiß daß er eine andere Sprache gelernt hat, trotzdem diese Sprache aber anwenden kann. anterograde Amnesie: Der Patient ist nicht in der Lage neue Gedächtnisinhalte (Fakten und Autobiographie betreffend) zu speichern. Das Langzeitgedächtnis und das Arbeitsgedächtnis funktionieren in der Regel normal und es können auch motorische Fertigkeiten gelernt werden (vgl. verschiedene Arten von Gedächtnis). Beim Menschen können keine Experimente durchgeführt werden. Leider kommt es aber zu Schlaganfällen, Sauerstoffmangel, Entzündungen, Operationen oder Erkrankungen. Anhand des verletzten Gebietes und eingehenden Untersuchungen ist man dann in der Lage Modelle aufzustellen. Patient F.R.: schwere anterograde und leichtere retrograde Amnesie, Schädigung der Hippocampusregion. Patient G.J.: Zugriff auf Gattungsnamen und spezielle Hauptwörter ist gestört, Umgang mit betreffenden Gegenständen stellen kein Problem dar, Alzheimer-Krankheit. Patient E.D.: retrograde Amnesie bezüglich der Autobiographie, beidseitige Verletzung des Schläfenlappens. Patient A.B.: schwere anterograde Amnesie und leichte episodische retrograde Amnesie, beidseitiger thalmischer Infarkt. Es gibt dann noch einige Erkrankungen die spezielle Regionen des limbischen Systems betreffen. Alzheimersche Krankheit: Amyloidablagerungen und Absterben von Neuronen im Hippocampus,dem basalen Vorderhirn (Nucleus basalis Meynert und dem Kern des diagonalen Bandes) und in der Großhirnrinde führt zu umfassender Schädigung des Gedächtnisses. ____________________________________________________________________________________________ Das Gedächtnis 66 Korsakow-Syndrom: Diese Erkrankung führt zu einer Zerstörung der anterioren und medialen Thalamuskerne. Sie tritt meist bei chronischen Alkoholikern auf. Die Intelligenz bleibt in der Regel erhalten, es gibt aber massive Probleme sich neue Fakten zu merken. Emotionale Erlebnisse können aber gespeichert werden. Psychogene Amnesie: Ausgelöst durch einen emotionellen Schock führt dies zu einer retrograden Amnesie, die meist nach ein paar Wochen verschwindet. Urbach-Wiethe Krankheit: Führt zu einer Verkalkung der Amygdala. Kein Einfluß auf kognitive Gedächtnisformen. Massive Einschränkungen bei emotioneller Beurteilung von Situationen oder der Gesichtsmimik. All diese Erkrankungen fühern zu folgenden Erkenntnissen: retrograde Amnesie: Der Informationsabruf ist geschädigt, wenn die Verbindung (fasciculus unicatus) von den Schläfenlappenspitzen und dem Stirnhirn zerstört ist. Eine Läsion der vorderen Zonen des limbischen Assoziationscortex führt zum Fabulieren. anterograde Amnesie: Eine Verletzung des medialen Temporallappens, mit den damit verbunden Strukturen (Hippocampus), führt zu einer Störung des Abspeicherns neuer Information (Reizkombination). Es kann sich kein neues Langzeitgedächtnis bilden. Es wurden verschiedene strukturelle Modelle vorgeschlagen, um den Informationsweg zu beschreiben. Leider ist der Hippocampus eine Region, die mit sehr vielen anderen Regionen in unmittelbaren Kontakt steht. Es ist schwierig zu erkennen welche beteiligten Strukturen wichtig sind, und welche nicht. So sollen nun einige Strukturpläne gezeigt werden. Gyrus cinguli Hippocampus us Amygdala Die Leitungsbahn, die ursprünglich James Papez vorgeschlagen hat, ist durch dicke Linien gekennzeichnet. Diese Verbindungen sind durch die Anatomie bekanntgeworden. Die Verbindungen, durch die dünnen Linien gekennzeichnet, wurden erst in jüngerer Zeit entdeckt und für wichtig erachtet. Aufgrund von Fehlinterpretationen verschiedenster Läsionen an Tieren vermutete man eine wesent-liche Bedeutung des Papezschen Leitungsbogen (dicke Linien). Da aber Verletzungen dieser verbindenden Struktur beim Menschen keine so massiven Schädigungen herbei-führten, mußte das Modell überdacht werden. Heute wird dieser Schaltkreis als Modell für das Entstehen von Emotionen betrachtet. Die Synchronisationszustände in verschiedenen Rindenarealen (primärer Assoziations- oder übergeordneter Assoziationscortex) werden über das Arbeitsgedächtnis im Hippocampus kurzfristig (maximal 2 Tage ohne Auffrischung) gespeichert. Im Schlaf in der REM-Phase werden das cholinerge System und die PGO-Zellen aktiviert. Die PGO-Zellen aktivieren ihrerseits die Großhirnrinde und spezielle Thalamuskerne in zufälliger Weise. Das cholinerge System aktiviert den Hippocampus und das Arbeitsgedächtnis. Durch die Aktivierung des Hippocampus werden die gelernten Reizkombinationen über das Arbeitsgedächtnis wieder aktiv. Nach der REM-Phase können nur stabile Synchronisationen, die durch den Hippocampus angeregt wurden, zu einer cotico-thalmischen Rückkopplung führen. Das heißt, ___________________________________________________________________________________ 67 Brain Modelling die Neuronen die an der Reizkombination beteiligt sind, werden sehr oft aktivert. Man nimmt an, daß eine starke Wiederholung der synaptischen Aktivität zu einem Umbau der Synapsen führt. Dieser Umbau ist dann besonders stabil. Damit wäre die Entstehung des Langzeitgedächtnisses erklärt. 7.3 Der Hippocampus Der Hippocampus ist eine Region des medialen Temporallappens. Im Prinzip ist es eine eingerollte Schicht der Großhirnrinde, die am unteren Saum des Schläfenlappens sitzt. Der Hippocampus wird in 4 Regionen CA1, CA2, CA3 und CA4 unterteilt. Abb. 7.5: Der Hippocampus im aufgerollten Zustand. Diese Regionen sind massiv untereinander über verschieden Fasern verbunden. Die Neuronen der CA1-Region besitzen NMDA-Rezeptoren. In der Graphik (a) sehen wir einen Querschnitt durch den Hippocampus. In (b) ist die gesamte Hippocampusstruktur entrollt und man kann die einzelnen Verbindungen besser erkennen. Der Hippocampus ist mit vielen Teilen des Gehirns verbunden: Unspezifische Eingänge des Hippocampus: reticuläres System des Hirnstamms, Hypothalamus, Kern des diagonalen Bandes, unspezifische Thalamuskerne, Amygdala. Diese Eingänge dienen dazu das Aktivierungsniveau in diesem Gebiet zu regulieren. Angst führt zu einer Aktivierung der Amygdala, dies führt zu einer Hemmung oder auch zu einer Aktivierung des Hippocampus. Über das reticuläre System wird die Aufmerksamkeit gesteuert, und dies führt zu einer Aktivierung des Hippocampus was zu einer verminderten oder gesteigerten Lernleistung führt. Über die unspezifischen Eingänge werden keine kognitiven Informationen übertragen, sondern es wird eine Kontrolle über den Einfluß der sensorischen Areale ausgeübt. Über die NMDA-Rezeptoren können synchrone Aktivitäten von Neuronen kurzzeitig gespeichert werden. Untersuchungen zeigten, daß ein schwaches Reizmuster an einem Neuron der CA1-Region keine LTP auslöst. Sind die beiden Reize aber synchron, so wird ____________________________________________________________________________________________ Das Gedächtnis 68 eine LTP an beiden Dendritenästen ausgelöst (Assoziativität). Ist aber ein Reiz besonders stark, ein Neuron feuert sehr oft, dann werden nur an dieser Kontaktstelle die NMDARezeptoren aktiv. Dies scheint nur ein Laboreffekt zu sein und für die allgemeine Gedächtnisleistung ohne Belang. Spezifische Eingänge des Hippocampus: Alle sensorischen und assoziativen Felder der Großhirnrinde sind mit dem Hippocampus verbunden. Über diese Felder kann eine Reizkombination eingespeichert werden. Der entorhinale Cortex stellt den eigentlichen Eingang des Hippocampus dar. Eine Schädigung dieser Region führt zur gleichen Einschränkung der Gedächtnisleistung wie die Zerstörung des Hippocampus selbst. Über den perirhinalen und parahippocampalen Cortex werden die Verbindungen zwischen den unterschiedlichen Cortexarealen her-gestellt und über den entorhinalen Cortex soll sich der Hippocampus "merken", welche dieser Verbindungen im Laufe des Tages aktiv waren. ___________________________________________________________________________________ 69 Brain Modelling Konsolidierung und Umbau der Synapsen: Wenn eine Synapse besonders oft aktiv ist, ändert sich der cAMP-Spiegel. Dies führt zu einer Aktivierung der davon abhängigen Proteinkineasen. Dadurch wandern, wenn der cAMPSpiegel ausreichend groß ist, katalytische Untereinheiten zum Zellkern. Dort wird das CREB1 Protein phosphorilisert. Dadurch werden Gene eingeschalten die eine synaptische Modifikation bewirken. Im Tierversuch konnte gezeigt werden, daß das CREB-1 Protein zur Entwicklung neuer Synapsen führt. ____________________________________________________________________________________________ Das Gedächtnis 70 Neurotransmitter Über Neurotransmitter werden Informationen übertragen. Glutamatrezeptoren wirken erregend, wenn sich Glutamat bindet. Wird GABA an einem dafür geeigneten Neurotransmitter gebunden, dann wirkt dieser Rezeptor hemmend auf das Aktionspotential des nachgeschaltenen Neurons. Diese beiden Neurotransmitter dienen der Invormationsvermittlung, das heiß es wird Information von einem Gebiet in ein anderes transformiert. Neurotransmitter können aber auch modulatorisch wirken. Sie können in einzelnen Regionen Synchronisation erleichtern oder unterbinden. Damit werden viele Zustände des Gehirns geregelt. Im Gehirn wird Noradrenalin als Transmitter von Neuronen synthetisiert, deren Zellkörper im Locus coeruleus (blauer Kern) sitzen. Dieser Kern im Hirnstamm übernimmt zahlreiche regulatorische Funktionen. Obwohl diese Neuronen in relativ geringer Zahl vorliegen, ziehen ihre Axone verstreut bis ins Rückenmark, in das Kleinhirn und in alle Bereiche der Großhirnrinde. Locus coeruleus unteres Tegmentum Substantia nigra Es gibt mehrere Dopaminbahnen. Zuerst wurde Dopamin im Streifenkörper entdeckt. Dieser Bereich stellt ein motorisches Kontrollzentrum dar. Die Zellkörper für diese Bahn befinden sich in der Substantia nigra. Eine für die Schizophrenie wichtige Bahn erstreckt sich vom unteren Tegmentum zur Stirnrinde und einzelnen Assoziationszentren. ___________________________________________________________________________________ 71 Brain Modelling Serotonin ist der Transmitter einer besonderen Gruppe von Neuronen, deren Zellkörper sämtlich in den Raphe-Kernen des Hirnstamms liegen. Die Ausläufer dieser Zellen sind weit über das gesamte Gehirn und das Rückenmark verteilt. man nimmt an, daß diese Neuronen an der Regulation von Aufmerksamkeit und anderen komplexen kognitiven Fähigkeiten beteiligt sind. Raphe-Kerne Neuronen die Acetylcholin produzieren finden sich im Hippocampus, der Amygdala, dem Thalamus und verschiedenen Kernen im Stammhirn. Von diesen Kernen ziehen die Bahnen wieder zu den einzelnen Kernen und in weite Bereiche der Großhirnrinde. Über die Rolle der Acetylcholinneuronen im Gehirn besteht immer noch Zweifel, wenngleich man ihnen eine große Rolle für das Gedächtnis zuschreibt (Aktivierung des Hippocampus während REM-Phase). ____________________________________________________________________________________________ Neurotransmitter 72 Der kranke Geist Es können verschiedene Leistungen des Gehirns ausfallen. Dies kann zu einer Amnesie, dem Koma, der Alzheimer-Krankheit oder Sprachstörungen führen. Einzelne Strukturen fallen aus und verschiedene Aspekte des Denkens können nicht mehr durchgeführt werden. Es kann aber auch zu Veränderungen von Denkprozessen kommen. Schizophrenie Als Schizophrenie bezeichnet man eine schwere Störung des Denkens, Fühlens, Empfindens, Wahrnehmens und Verhaltens. Personen mit einer Schizophrenie hören Stimmen, oder sehen Dinge die es nicht gibt. Sie haben Halluzinationen. Die Erkrankung trifft einen von hundert. Als auslösende Ursachen gilt ein gestörtes soziales Umfeld und/oder eine Störung der Gehirnfunktion. Eugen Bleuler definierte als erster das Krankheitsbild über Wortassoziationen. Er stellte fest, daß eine Störung des Assoziierens typisch für diese Krankheit ist (Heu = Unterhaltungsmittel für Kühe). Über den semantischen Bahnungseffekt kann relativ leicht das Assoziationsverhalten überprüft werden. Der Proband muß ein Wort lesen und dann angeben ob das zweite Wort aus dem Deutschen stammt oder nicht. Es wurden assoziative Wortpaare wie weiß::schwarz und Bruder::Schwester dafür verwendet. Ein Beispiel für nicht-assoziativen Wortpaare wäre Wolke::Käse oder Kaffee::Benzin. Diese Wortpaare und Wortpaare wo das zweite Wort nicht-deusch war wurden dem Probanden vorgelegt. Der Proband mußte dann über einen Fingerdruck bekanntgeben, ob das zweite Wort aus dem Deutschen stammt. Es wurden nur die "JA"-Entscheidungen gewertet und der jeweilige Mittelwert gebildet. Im Regelfall werden assoziative Wortpaare schneller als nicht assoziierte Wörter erkannt. Dies bezeichnet man als den semantischen Bahnungseffekt. Bei Personen mit einer denkgestörten Schizophrenie ist der Bahnungseffekt besonders groß. ___________________________________________________________________________________ 73 Brain Modelling Bei schizophrenen Personen können oft indirekte Assoziationen beobachtet werden. Das heißt es gibt ein nicht ausgesprochenes Bindeglied. Assoziation zu Nikotin - Forelle. Nikotin im Rauch verursacht Krebs, Krebse leben im Wasser wie Forellen. Man geht davon aus, daß bei schizophrenen Patienten eine Störung der fokussierten Aufmerksamkeit vorliegt. Verschiedene Systeme im Gehirn versuchen die Synchronisationen zwischen verschiedenen Gebieten im Zaum zu halten. Nur so können wir uns auf etwas konzentrieren. Wenn die Assoziationsfähigkeit zu groß wird, werden nicht korrelierte Reie miteinander verknüpft. Damit können wir die Umwelt nicht mehr sinnvoll einschätzen. Genau diese fokusierte Aufmerksamkeit wird über den Neurotransmitter Dopamin gesteuert. Die Neuronen des Dopaminsystems innervieren vorallem das Arbeitsgedächtnis. Alle Neuroleptika blockieren die Dopaminrezeptoren in der Großhirnrinde. Dies führt zu einer geringeren Assoziationsfähigkeit und es können auch keine spontanen Synchronisationen (Halluzinationen) entstehen. Wahn Auch der Wahn ist eine Störung des Denkinhaltes. Es kommt zu einer gedanklichen Fokussierung auf einen speziellen Gegenstand. Wird die Schizophrenie nicht behandelt, so kommt es zum Wahn. Es existieren gewisse unkorrigierbare Urteile, deren Inhalte unmöglich, unwahrscheinlich oder einfach falsch sind. Bei einer Überprüfung der Fakten hält ein Wahnkranker an seinen Vorurteilen fest. Meist ist dieses Verhalten auch mit Angst, Mißtrauen oder gesteigerter Wachheit verbunden. Belanglose Ereignisse werden überinterpretiert. Im Prinzip handelt es sich um eine Stabilisierung der Synchronisation. Durch die Schizophrenie ausgelöst, können obskure Synchronisationen auftreten. Wenn ein und die selbe Synchronisation beziehungweise Assoziation öfters auftritt, dann bilden sich neue Synapsen. Bei einem chronischem Wahn helfen keine Neuroleptika mehr. Es haben sich schon neue Synapsen, der Informationsverarbeitung betreffend, gebildet. ____________________________________________________________________________________________ Der kranke Geist 74 Depression Plutarch hat diese Krankheitsbild als Erster beschrieben. Es ist gekennzeichnet von: • Gefühl der Niedergeschlagenheit (bei Tod eines Verwandten, räumliche Trennung von einer geliebten Person.) • Die Selbstachtung ist empfindlich gestört (Arbeitslosigkeit, Sitzenbleiben in der Schule) • Verlust des Interesses an sozialen Kontakten und alltäglichen Dingen Es ist nicht immer einfach eine depressive Verstimmung von einer Depression zu unterscheiden. Die Zeit scheint ein wesentlicher Faktor zu sein. Manche Patienten zeigen auch ein anderes Phänomen: die Manie. Der Patient ist in Hochstimmung, energiegeladen und sehr optimistisch. Diese scheinbaren positiven Eigenschaften können des Urteilsvermögen herabsetzen. In der Regel sind die depressiven Phasen länger als die manischen. Bei der Behandlung muß berücksichtigt werden, daß der Selbstmord eine große Gefahr für den Patienten darstellt. Früher verwendete man Opiate zur Behandlung einer Depression. Dies führte zwar zu euphorischen Zuständen, die aber sehr kurzlebig waren. Heute werden tricyclische Antidepressiva zur Behandlung verwendet. Diese haben relativ geringe Nebenwirkungen, die gewünschte Wirkung tritt aber erst nach rund zwei Wochen auf. Diese Antidepressiva hemmen die Inaktivierung des Neurotransmitters im synaptischen Spalt. Ein Neurotransmitter wird normalerweise, nachdem er sich an einen Rezeptor gebunden hat, inaktiviert. Diese Inaktivierung wird durch die tricyclischen Antidepressiva verhindert. Die Inaktivierung betrifft vorallem die Neurotransmitter Noradrenalin und Serotonin. Es zeigte sich, daß bei einer Depression die Neuronen der Großhirnrinde weniger aktiv sind, als im Grundzustand. Während einer Manie tritt das Gegenteil ein. Besonders die Strukturen des präfrontalen Cortex sind überaktiv. Bei einem Viertel aller depressiven Patienten führen Antidepressiva nicht zu einer Besserung. Man geht davon aus, daß es sich dann um eine Fehlsteuerung des Streßsystems handle. Stellt das Gehirn (kognitive Bereiche) Angst oder Streß fest, so wird der Hypothalamus aktiviert. Dieser sondert den cotico-releasing-factor aus und aktiviert damit die Hypophyse (Hirnanhangsdrüse). Die Hypophyse sondert nun ihrerseits über das Blut das adrenocorticotropes Hormon aus. Zentraler Empfänger ist die Nebenniere, die deshalb Cortison ausschüttet. Damit es zu keiner Übersteuerung kommen kann und das System sich nach der Angst- oder Streßsituation wieder beruhigt, wird eine Gegenkopplung aktiv. Das Cortison hemmt die Hypophyse und den Hypothalamus. Wenn die Gegenkopplung niedrig ausfällt, kann es zur Depression kommen, denn die ausgeschütteten Hormone steuern auch noch viele andere emotionale Mechanismen. Angststörungen “Es steht außer Frage, daß das Problem der Angst ein Knotenpunkt ist, an dem die verschiedenen und wichtigen Fragen zusammentreffen, ein Rätsel dessen Lösung zwangsläufig eine Lichtflut auf unsere ganze geistige Existenz werfen würde.” Sigmund Freud Nur Dinge die tatsächlich gefährlich werden können, führen zu Angststörungen. Wesentlich ist der Unterschied zwischen Furcht und Angst: Furcht ist auf etwas gerichtet, während Angst gegenstandslos ist. Man kann die Angstörungen in folgendes Schema unterscheiden: - posttraumatische Ängste (Erinnerung an gefährliche Situationen) ___________________________________________________________________________________ 75 Brain Modelling - Zwangssyndrom (krankhaftes Händewaschen, aus Angst vor Infektionen) - Panik (grundlose Furcht, mit starkem körperlichen Aspekten wie Herzbeklemmung) - Phobie (an ein Objekt oder Szene gebunden) Phobien sind stark umweltabhängig. Eskimos können eine Kanuphobie bekommen und früher gab es die Dromosiderophobie - Eisenbahnfurcht, die sich heute in die Flugangst verwandelt hat. Kleinste ungefährliche Veränderungen im Körper oder in der Umgebung werden überbewertet und es folgt eine Überreaktion. Diese Überreaktion führt zu einer gezielteren Wahrnehmung (besonders auf gefährliche Situationen). Auch die kognitiven Bereiche des Gehirns reagieren sensibler. Auch hier haben wir eine Mitkopplung gegeben. Stress Im Jahr 1959 führte der österreichisch-kanadische Mediziner Hans Seyle den Begriff Stress in die Biomedizin ein und bezeichnete damit die unspezifischen Reaktionen eines Organismus auf jede übermäßige Belastung. Dieses Reaktion dient dazu, die Überlebenschancen in einer lebensbedrohlichen Situation eines Indivduums zu erhöhen. Dies ist eine, wenn gleich auch nicht ganz richtige, aber allgemein anerkannte Meinung. Meist spricht man von Stress, wenn der Stressor - der Auslöser auf diesen Effekt - kurz und heftig über eine Erhöhung des Adrenalinspiegels auf das Individuum reagiert. Es ist aber wichtig zu erwähnen, daß langanhaltender Stress wahrscheinlich mehr physiologische und damit psychologisch verbundene Veränderungen im menschlichen Körper verursacht, als eine kurzfristige Aktivierung des gesamten Stresssystems. Die Stressantwort auf eine scheinbar gefährliche Situation kann sehr spezifisch sein, sich gezielt gegen einen Erreger richten oder gegen einen realen oder imaginären angstauslösenden Umstand. Übersteigt die Belastung, beziehungsweise die korrelierte Stressantwort eine gewisse Grenze, dann ist die Reaktion unspezifisch. Eine wesentliche Frage lautet, was denn subjektiv als Stress (langanhaltend) empfunden wird. Das Individuum entscheidet, ob eine Situation als Stress empfunden wird oder nicht. Dies hängt größtenteils von der Vergangenheit, als auch der Zahl der zur Verfügung stehenden Strategien ab. Je mehr Strategien zur Verfügung stehen umso besser für das Individuum. Auch eine Vermeidung des Stressors kann als Kontrolle über die jeweilige Situation betrachtet werden. Aber alle Stressoren zu vermeiden oder zu umgehen führt in den meisten Fällen zu schweren psychischen Störungen und eine normale soziale Interaktion wird fast unmöglich (ausgeprägte Phobien). Die Kontrollierbarkeit einer Situation hängt also auch von den kognitiven Fähigkeiten eines Individuums ab. Wenn keine "sinnvolle" kognitive-emotionelle Verhaltensweise verfügbar ist, um eine angemessene Reaktion zu aktivieren, dann spricht man von unkontrollierbarem Stress. Wenn sich ein Individuum mit einem unkontrollierbaren Stressor auseinandersetzen muß und das innere Gleichgewicht nicht hergestellt werden kann, dann beruhigt sich das zentrale Stresssystem nicht und es bleibt weiterhin aktiviert. Dadurch kommt es zu einer “Verhaltensvermeidung”. Es werden keine äußeren Aktionen gesetzt. Dies hilft nicht nur Energie zu sparen, sondern man kann sich dann in Ruhe mit der Problematik der angespannten Situation beschäftigen und neue Strategien können entwickelt werden. ____________________________________________________________________________________________ Der kranke Geist 76 Beschäftigen wir uns nun mit den organischen Korrelaten des Stresssystems. Es war schon lange bekannt, daß wochenlange Arbeitsüberlastung zu Grippe, zu einer Magenschleimhautentzündung oder Herpesbläschen führen kann. Hormone sind Signalstoffe, die das Verhalten und die Empfindungen eines Menschen entscheidend beinflussen. Sie entstehen in Drüsen, die ihre Sekrete unmittelbar an das durchströmende Blut abgeben. Beim Menschen sind dies die Schilddrüse, Nebenschilddrüse, Langerhansschen Inseln der Bauchspeicheldrüse, Die Nebenniere (Mark und Rinde) und die Keimdrüsen. Die meisten Hormone werden aber vom Gehirn - vertreten durch die Hypophyse (Hirnanhangsdrüse) und durch die Epiphyse (Zirbeldrüse) - abgesondert. Im Nebennierenmark werden die Hormone Adrenalin und Noradrenalin (80% zu 20%) gebildet. Die Hormone der Nebennierenrinde bezeichnet man als Corticoide. Eine spezielle Gruppe der Corticoide - die Glucocorticoide - beinflussen den Kohlehydrat- und Eiweißstoffwechsel. Dazu gehört insbesondere das Cortisol, das außerdem entzündungshemmende Wirkung besitzt. Die Ausschüttung hängt sowohl von internen Faktoren, wie der inneren Uhr (Periode bei der Frau), als auch von äußeren Reizen (Pheromone beim Sexualakt) ab. Sie wirken in außerordentlich kleinen Dosen. Die Hormone sind in der Regel nicht in der Lage sofortige - im Bereich von Sekunden - Reaktionen im Organismus auszulösen. Der Hippocampus steht in direkter Verbindung mit dem Hypothalamus, einem anderen wichtigen Kern. Der Hypothalamus wird aber auch von vielen anderen Regionen im Gehirn innerviert. Über ihn wird durch die Ausschüttung des Coticotropin-Releasing-Faktor die Hypophyse, die verschiedene Hormone ausschüttet, aktiviert. Durch diese Ausschüttung werden andere Teile des Hormonsystems aktiv. Eine Ausschüttung des adrenocorticotropen Hormons führt zu einer Aktivierung der Nebennierenrinde. Es kann aber auch die Aktivität der Keimdrüsen, der Schilddrüse und Teile der Bauchspeicheldrüse beeinflußt werden. Diese setzen ihrerseits wieder Hormone frei. Die Nebenniere produziert Cortisol, eines der wichtigsten Stresshormone. Cortisol reduziert aber die Aktivität der Hypophyse. Damit wird im Körper ein Überschießen der hormonellen Reaktion verhindert. Untersuchungen zeigten, daß dieses wichtige Stresshormon bei psychischen Erregungszuständen wie panischer Angst, Depression oder auch Drogenentzug vermehrt synthetisiert wird. Prinzipiell kann man sagen, daß der Hypothalamus in Abhängigkeit von verschiedenen Erregungszuständen des Gehirns den Hormonhaushalt regelt. Cortisol wird ausgeschüttet bei kurzandauernden körperlichen und psychischen Stresszuständen, es beeinflußt wie alle Hormone den Kreislauf. Das Cortisol reagiert aber auch mit dem Hippocampus, einer wichtigen Gehirnregion. Während einer langandauernden Stresssituation, also einem erhöhten Cortisolausstoß, sterben einzelne Dendritenäste im Hippocampus und dem präfrontalem Cortex ab. Wenn der Cortisolspiegel wieder sinkt, dann regenerieren die Neuronen. Möglicherweise können aber die ursprünglichen Verbindungen nicht mehr hergestellt werden. Somit kann es zu leichten Verschiebungen im Verhalten beziehungsweise in den Strategien oder der Strategienwahl kommen. Zusätzlich scheint chronisch unkontrollierbarer Stress die Dichte der präsynaptischen Serotoninsynapsen im Hippocampus zu erhöhen und im Hypothalamus und der Großhirnrinde zu reduzieren. Zusammenfassend könne wir sagen: Die Quelle und das Ziel für eine Stressantwort ist das Gehirn, im speziellen die höheren kortikalen und limbischen neuralen Schaltkreise durch die wir die Welt interpretieren. Kontrollierbarer Stress ist ein Auslöser für die Verstärkung von spezifischen neuralen Wegen. ___________________________________________________________________________________ 77 Brain Modelling Im speziellen durch die Interaktion von zirkulierenden Glucocorticoiden wird unkontrollierbarer Stress ein Werkzeug um neurale Schaltkreise und Netzwerke im Bereich der Verhaltensreaktionen umzustrukturieren. So wird psychosozialer Stress ein Auslöser für die adaptive Reorganisation des Gehirns, vor allem im assoziativen Gedächtnis. Reiz Adaption Reorganisation Thalam us Verstärkung von neuralen Strukturen Großhirnrinde (präfrontaler Cortex - Assoziationscortex) Dopamin- System Hippocamp us Degenerati on von Dendriten Amygdala Hypothalamus startet Kaskaden von Stresshormonen und Neurotransmittern Vasopressin CRH CRH Noradrenalin Locus Coeruleus Hypophys e ACTH Zentrales noradrenerges System Sympathicus Adrenalin Nebenniere Cortisol beeinflußt Schlafverhalten Kontrollierbarkeit schwächt Immunzellen Unkontrollierbarkeit ____________________________________________________________________________________________ Der kranke Geist 78 Künstliche Intelligenz Wissensrepräsentation: Modell in Formalismus implementiert durch Interpreter Wissensbasis Die KI hat es sich zur Aufgabe gemacht, das Schließen über unsere physikalische Umwelt zu formalisieren und die Analyse physikalischer Systeme sowie die Vorhersage ihres Verhaltens zu automatisieren. Ausschließlich qualitative Informat-ion über das System ist ausreichend, um das System zu verstehen und seinen Mechanismus zu erklären: eine Beschreibung der Struktur, das heißt der Beziehungen zwischen den Teilen und Wissen über die Funktionalität dieser Teile (wie sie funktionieren und welche Funktionen sie deshalb prinzipiell im Gesamtsystem übernehmen können). ? Problemmerkmale Problemlösung ___________________________________________________________________________________ 79 Brain Modelling Expertensysteme: Ein Expertensystem ist ein Computerprogramm, mit dem versucht wird, anhand von erhobenen Gedankengängen und Erfahrungen von Experten eines bestimmten Fachgebiets ein maschinelles System zu konstruieren, das Anwendern Aspekte einer Problemlösungskompetenz zur Verfügung stellt. Das implizite Wissen von Experten, oder besser die Besonderheit des Expertenhandelns, äußert sich vor allem in der Geschwindigkeit und der Qualität: Das rasche Handeln setzt Wissen voraus, aber im Moment des Handelns wird kein bewußter Bezug auf das Wissen erlebt. Semantische Netze: Ein Gedächtnismodell, das auf zwei grundlegenden Typen von Beziehungen basiert: [1] Beziehungen von Konzeptknoten, Is-A-Beziehung, Ober-Konzept - Unter-Konzept Relation. [2] Beziehung zwischen Konzepten und Eigenschaften, Has-Prop-Beziehung. hat Haut Wirbeltier hat Federn kann fliegen kann singen Vogel XXX Kanarienvogel Has-Prop Fisch hat Kiemen Laufvogel Kranich Strauß Is-A ____________________________________________________________________________________________ Künstliche Intelligenz 80 Synthetische Psychologie Braitenbergs These: Biologisches Verhalten lässt sich leichter synthetisieren als analysieren. Braitenbergsches Vehikel: Dies ist ein Fahrzeug mit zwei voneinander unabhängig angetriebenen Hinterräderen, wobei der Unterschied in der Drehzahl der Hinterräder bestimmt, wohin das Fahrzeug fährt. Ein Sensor, befestigt am Fahrzeug, bestimmt die Drehzahl eines Rades. Standardvehikel: Hinterrad Motor excitatorische Verbindung zwischen dem Sensor und dem Motor Wenn auf den LDR Licht fällt, sinkt sein Widerstand, und der mit dem LDR verbundene Motor dreht sich schneller. Das Vehikel meidet Licht. (Angst ?) excitatorische überkreuzteVerbindung zwischen dem Sensor und dem Motor Das Vehikel mit den überkreuzten Verbindungen sucht das Licht, und fährt mit voller Geschwindigkeit darauf zu und versucht es zu rammen. (Agressivität ?) LDR (light dependent Resistor) Einführung von Neuroden: Die analogen Signale der Rezeptoren werden übersetzt in Feuerfrequenzen. Das heißt bei maximaler Beleuchtung feuert der LDR-Sensor mit 100 Pulsen pro Sekunde. Neuroden sind nun Rechenelemente die von Rezeptoren und anderen Neuroden Impulse empfangen, verrechnen und an andere Neuroden oder Motoren weiterleiten. Im Neurod tickt eine Uhr mit einem bestimmten Takt. Ein Neurod feuert nur dann, wenn die Uhr gerade einen Takt vollendet hat und die Zahl der Impulse die das Neurod empfangen hat einen gewissen Schwellwert überschritten hat. ___________________________________________________________________________________ 81 Brain Modelling 1 excitatorisches Neurod 1 inhibitorisches Neurod Je höher die Pulsrate des Sensors, umso langsamer läuft der Motor. Das Vehikel bewegt sich auf die Lichtquelle zu und bleibt in stiller Verehrung vor ihr stehen. (Demut ?) 0 0 Das Vehikel wendet sich von der Lichtquelle langsam ab und sucht hellere Lichtquellen im Dunkeln. Diese werden dann aber ebenso gemieden. (Forschergeist ?) Ab einer gewissen Pulsfrequenz ändert sich das Verhalten des Vehikels drastisch. 0 0 2 1 1 1 Behavior Modelling: Braitenbergmodell mit Bedürfnissen: Niedriger Glucosehaushalt im Blut führt zu Hunger. Es gibt verschiedene Bedürfnisse, die sich gegenseitig beinflussen können, und erst bei verschiedenen Stärken aktiv werden. Eine paar Modulationsparameter steuern das gesamte Bedürfnisverhalten. Gefahr => hohe Aktiviertheit => schnelle Reaktion ____________________________________________________________________________________________ Synthetische Intelligenz 82 Spieltheorie 14.0 Spieltheorie "Die geringe körperliche Kraft des Menschen, seine geringe Schnelligkeit, der Mangel an natürlichen Waffen werden mehr als ausgeglichen . . . durch seine sozialen Eigenschaften, welche ihn dazu führten seinen Mitmenschen zu helfen und Hilfe von ihnen zu empfangen." Charles Darwin (1809-1882) Kommentkampf - Duell: Falken Tauben Modell Es gibt nur eine begrenzte Kriegsführung, das heißt alle kämpfenden Parteien halten sich an die Spielregeln und niemand wird schwer verletzt. Falke Taube Gewinn bei Sieg: +10 Gewinn bei Niederlage: 0 Falke +1 -5 schwere Verletzung: -5 0 langer Kampf: +2 Taube 0 +2 E(T,T) = 0.5 . (+10) + 0.5 . (0) + (-3) = +2 E(F,F) = 0.5 . (+10) + 0.5 . (-20) = -5 E(F,T) bzw. E(T,F) = 0 für Taube und +10 für Falke Die Strategie i ist gegenüber j stabil wenn gilt: E(i,i) E(j,i) bzw E(i,j) E(j,j) Mischstrategien:Tue immer das was die anderen nicht tun ! ( 8 5 Falken, Tauben ) 13 13 Problem des Modells: Unterschied in Kampfgröße,Stärke und Waffen des Gegners. Motivation (hunger makes agressiv) Einführung von Besitz verändert die Situation: Besitzender verhält sich wie Falke, Habenichts wie eine Taube. Kommt es zwischen zwei Männchen A und B (Pavianen) zu einer Auseinandersetzung, wird ein Beistand C gesucht. Was hat C davon A oder B zu helfen ? Bekommt er auch Hilfe von A (oder B) ? (BETRUG) ___________________________________________________________________________________ 83 Brain Modelling Gefangenendilemma: Lohn für Kooperation: Strafe für beiderseitiges Betrügen: Versuchung zu Betrügen: Gewinn für den Betrogenen: L S V T = = = = -2 -4 0 -5 Spieler 1 Kooperation Betrug Kooperation Spieler 2 Betrug -2 0 - - 2 -5 5 -4 0 4 Kanonische Nutzen-Matrix: V > L > S > T (1) (V+T)/2 < L (2) Beide Spieler können sich überlegen: Betrügen ist für mich vorteilhafter. Also betrügen beide, um ihren Nutzen zu maximieren und erreichen das Gegenteil, während beide bei beiderseitiger Kooperation einen höheren Nutzen gehabt hätten. iteriertes Gefangenendilemma:Die beiden Spieler treffen öfters hintereinander zusammen und "spielen" miteinander. Strategien: Tit for Tat Ich kooperiere beim ersten mal und tue genau das, was der andere beim letzten mal getan hat. Mehrheitsent. Mild Ich tue was der andere in der Mehrzahl der bisherigen Fälle getan hat. Grimmig Ich kooperiere, bis der andere zum erstenmal betrügt, von da an betrüge ich stets. Hartes Tit for Tat Ich kooperiere, es sei denn der andere hat beim letzten oder beim vorletzten Mal betrogen. Lieb Ich kooperiere immer. Verrückt Ich betrüge durchschnittlich jedes zweite mal abhängig vom Zufall. Böse Ich betrüge stets. Bei der direkten Konfrontation gewinnt (verliert nicht) die Strategie Böse. Es ist besser nett zu sein als böse. Man muß reaktiv sein. Es gibt keine beste Strategie ! Man muß rasch vergeben. List und Tücke bringen nichts. ____________________________________________________________________________________________ Spieltheorie 84 iteratives Gefangenendilemma mit Kündigung: Tit f. Tat mit Schmerzgrenze: Wenn in den letzten fünf Zügen der Gewinn kleiner als zwei Punkte pro Zug war, dann kündige ich. Probieren: Kooperieren, Kooperieren, Betrügen, Betrügen. Wenn der andere in dieser Zeit dreimal betrogen hat kündige ich, sonst kooperiere ich bedingungslos. Brutal: Ich betrüge, solange mein Partner kooperiert. Sobald er betrügt, kündige ich. Spieler 2 Kooperation Betrug Kündigung 3 Kooperation Spieler 1 Betrug Kündigung 5 3 0 0 1 5 1 2 2 ___________________________________________________________________________________ 85 Brain Modelling ____________________________________________________________________________________________ Spieltheorie 86 Literaturverzeichnis: The handbook of brain theory and neural networks / ed. by Michael A. Arbib. Ed. assistant Prudence H. Arbib. - Cambridge, Mass. [u.a.] : MIT Press, 1995. - XV, 1118 S. :Ill., graph. Darst. (A Bradford book)Literaturangaben ISBN 0-262-01148-4 Universitätsbibliothek WienHB- Mag.-->II-1203612 Ein Monument, aber leider etwas veraltet. Exzellente Aufsatzsammlung, wo wirklich nur das Wichtigste und Erwiesenes drinnensteht. Brause, Rüdiger: Neuronale Netze : eine Einführung in die Neuroinformatik / von Rüdiger Brause. - 2., überarb. und erw.Aufl. - Stuttgart : Teubner, 1995. - 462 S. : Ill., graph. Darst. (Leitfäden der Informatik)Literaturverz. S. 433 - 456 Universitätsbibliothek Wien 134 Mag.-->25206 Sammlung der wichtigsten Algorithmen aus dem Gebiet der technischen neuronalen Netze. Gute Erklärung, manche konkreten Beispiele. Braitenberg, Valentin: Vehikel : Experimente mit kybernetischen Wesen / Valentin Braitenberg. Aus dem Engl. von Dagmar Frank und Valentin Braitenberg. - Reinbek bei Hamburg : Rowohlt, 1993. - 156 S. :Ill., graph. Darst. (Rororo ; 9531 : Rororo-Sachbuch : Rororo-Science)Einheitssacht.: Vehicles <dt.>. - Literaturverz. S. [143] - 150 ISBN 3-499-19531-3 Interessant, spannend und witzig. Regt stark zum Nachdenken an. Diese Arbeit löste einen starken Boom in letzter Zeit aus. Models of brain function / ed. by Rodney M. J. Cotterill. - 1. publ. - Cambridge [u.a.] : Cambridge Univ. Pr., 1989. - XIII,574 S. : graph. Darst. Literaturangaben ISBN 0-521-38503-2 Universitätsbibliothek Salzburg NW L -> 73.2.4-82 Wo brain draufsteht, muß nicht brain drinnen sein. Signale und Kommunikation : Mechanismen des Informationsaustauschs in lebenden Systemen / mit e. Einf. von John Dittami. - Heidelberg [u.a.] : Spektrum, Akad. Verl., 1993. - 193 S. : Ill., graph. Darst., Kt. (Spektrum der Wissenschaft : Verständliche Forschung) Literaturverz. S. 184 - 186 ISBN 3-86025-214-3 Universitätsbibliothek Wien HB- Mag.--> II-1150825 Interessante Aufsatzsammlung, bei der gezeigt wird, wie die Formeln für das Gehirn auch anders verwendet werden können. Dorffner, Georg: Konnektionismus : von neuronalen Netzwerken zu einer "natürlichen" KI / von Georg Dorffner. - Stuttgart :Teubner, 1991. - XV, 448 S. : graph. Darst. (Leitfäden der angewandten Informatik)Literaturverz. S. [426] - 438 ISBN 3-519-02455-1 Universitätsbibliothek Wien HB- Mag.--> I1119316 Guter Überblick in den Bereich der technischen Neuronalen Netze. Dörner, Dietrich: Die Logik des Mißlingens : strategisches Denken in komplexen Situationen / Dietrich Dörner. - 21. - 30.Tsd. - Reinbek bei Hamburg : Rowohlt, 1993. - 320 S. : Ill., graph. Darst., Kt. (Rororo ; 8314 : RororoSachbuch : Rororo-Science) Literaturverz. S. 311 - 313 ISBN 3-499-19314-0 Universitätsbibliothek Wien 158 HA--> X70.D713.L8 Zeigt auf eindrucksvolle Weise, wie linear Menschen denken, und wie nichtlinear die Probleme in der Welt sind. Gardner, Howard: Dem Denken auf der Spur : der Weg der Kognitionswissenschaft / Howard Gardner. Aus dem Amerikan. v. Ebba D. Drolshagen. - Stuttgart : Klett-Cotta, 1989. - 456 S. : graph. Darst. Einheitssacht.: The mind's new science <dt.>. - Literaturverz. S. 419 - 438 ISBN 3-608-93099-X Universitätsbibliothek Wien HBMag.--> I-1102624 Nachdenken, fragen, meditieren und Schlußfolgerungen ziehen. Goldbeter, Albert: Biochemical oscillations and cellular rhythms : the molecular bases of periodic and chaotic behaviour /Albert Goldbeter. - Cambridge [u.a.] : Cambridge Univ. Press, 1996. - XXIV, 605 S. : Ill. Einheitssacht.: Rythmes et chaos dans les systèmes biochimiques et cellulaires <engl.>. - Aus d. Franz. übers. Literaturverz. S.526 - 588 ISBN 0-521-40307-3 Zentralbibliothek für Physik in Wien 39061 ->> FR-Gute mathematische Darstellung von biophysikalischen Vorgängen. Einführung in die künstliche Intelligenz / Günther Görz (Hrsg.). - 2. Aufl. - Bonn [u.a.] : Addison-Wesley, 1995. - XIV,1029 S. : graph. Darst. Literaturangaben ISBN 3-89319-858-X Universitätsbibliothek Wien HBMag.--> I-1185763 Hervorragende Zusammenfassung aus dem Bereich der KI. Guttmann, Giselher: Ich: sehe, denke, träume, sterbe : sechs Aufsätze über das Hier- und Jetztsein, das Sosein und Nichtsein des Menschen und seiner Seele im Spiegel der naturwissenschaftlichen psychologischen Forschung ; mit einem Vorwort, fünf Zwischentexten und einem Epilog. - München : Ehrenwirth, 1991. - 324 S. : zahlr.Ill., ___________________________________________________________________________________ 87 Brain Modelling graph. Darst. Literaturverz. S. 321 - 324 ISBN 3-431-03157-9 Universitätsbibliothek Wien 1120325 Gute Statements zum Bewußtsein und zum Denken. Darüber meditieren ! HB- Mag.--> I- Hernegger, Rudolf: Wahrnehmung und Bewußtsein : ein Diskussionsbeitrag zu den Neurowissenschaften.Heidelberg [u.a.] : Spektrum, Akad. Verl., 1995. - 552 S. : Ill. Literaturverz. S. 519 - 540 ISBN 3-86025-288-7 Universitätsbibliothek Wien HB- Mag.--> II-1176928 Eine sehr gute Zusammenfassung über das Gebiet der Gehirnforschung. Es werden die aktuellen Fakten kritisch beleuchtet. Hofstadter, Douglas R.: Gödel, Escher, Bach : ein endloses geflochtenes Band / München : Dt. TaschenbuchVerl., 1991. - XVII, 844 S. : Ill., graph. Darst., Notenbeisp. (Dtv ; 11436 : Dtv-Klett-Cotta). - Literaturverz.S. 805 - 819 ISBN 3-423-11436-3 Dieses Buch sollte man eigentlich erst dann lesen, wenn man schon seine Vision zum Thema Gehirn und Bewußtsein gehabt hat.Regt zum bewußten Nachdenken an. Neurowissenschaften : eine Einführung / Eric R. Kandel ... (Hrsg.). - Heidelberg [u.a.] : Spektrum Akad. Verl., 1996. -XXII, 786 S. : Ill., graph. Darst. (Spektrum Lehrbuch) Einheitssacht.: Essentials of neural science and behavior <dt.>. - Literaturverz. S. [739] - 744 ISBN 3-86025-391-3 Universitätsbibliothek Wien HB- Mag.--> II-1190247 Beste Darstellung der Biologie, der Medizin, der Genetik und der Chemie des menschlichen Gehirns in einem Buch. ABER: Die amerikanische Ausgabe ist besser, da umfangreicher. Gehirn und Geist / Kenneth A. Klivington. Wiss. Beratung: Floyd Bloom ... Aus dem Amerikan. übers. von Peter Germroth.- Heidelberg [u.a.] : Spektrum, Akad. Verl., 1992. - 240 S. : zahlr. Ill. Einheitssacht.: The science of mind <dt.>. - Literaturverz. S. 234 - 235 ISBN 3-86025-038-8 Universitätsbibliothek Wien HB- Mag.--> II1132935 Nette Bilder, nette Aufsätze und nette Einführung. Kratzer, Klaus P.: Neuronale Netze : Grundlagen und Anwendungen - 2., durchges. Aufl. - München ;Wien : Hanser, 1993. - XII, 211 S. : graph. Darst. Literaturverz. S. [200] - 206 ISBN 3-446-17315-3 Anderer Weg der Vermittlung des Wissens über technische neuronale Netze als im Brause, etwas zu pessimistisch. Kolb, Bryan: Neuropsychologie / Übers. hrsg. von Monika Pritzel. Aus dem Engl. übers. von Marianne Mauch ... - 2. Aufl. - Heidelberg [u.a.] : Spektrum Akad. Verl., 1996. - XII, 574 S. : Ill., graph. Darst. (Spektrum Lehrbuch) Einheitssacht.: Fundamentals of human neuropsychology <dt.>. - Literaturangaben ISBN 3-8274-0052-X Universitätsbibliothek Wien HB- Mag.--> II-1206523 Hervorragende Einführung in die Psychologie auf Basis der Biologie. Nauta, Walle J.: Neuroanatomie : eine Einführung - Heidelberg : Spektrum-d.-Wiss.-Verl.-Ges., 1990. - 342 S. : zahlr. Ill., graph. Darst. Einheitssacht.: Fundamental neuroanatomy <dt.>. - Literaturverz. S. 327 - 331 ISBN 389330-707-9 HB: Exemplare der LBS abgegeben an die UBW006 Universitätsbibliothek Wien HB- Mag.--> II1114725 Bester, billigster Hirnatlas für angehende Profis. Penrose, Roger: Computerdenken : des Kaisers neue Kleider oder die Debatte um künstliche Intelligenz, Bewußtsein und die Gesetze der Physik . Aus dem Engl. übers. von Michael Springer. - Heidelberg :Spektrum-derWiss.-Verl.-Ges., 1991. - XXI, 454 S. : Ill., graph. Darst. Einheitssacht.: The emperor's new mind <dt.>. Literaturverz. S. 442 - 448 ISBN 3-89330-708-7 Universitätsbibliothek Wien HB- Mag.--> I-1124131 Hervorragende Einführung in die Probleme der Quantenmechanik, der Gravitation und schwarzer Löcher; Vom Gehirn steht fast nichts drinnen. Erwähnenswert: Einführung und Erklärung der Turingmaschine. Neumann, John von: Die Rechenmaschine und das Gehirn. Dt. Übers.: Charlotte und Heinz Gumin. -München : Oldenbourg, 1960. - 80 S. Einheitssacht.: The computer and the brain <dt.> Zentralbibliothek für Physik in Wien Bestandsnachweis 20504 ->> FR-Meditieren, nachdenken, meditieren, nachdenken und sich nicht von aktuellen Forschungsergebnissen beeindrucken lassen und wieder meditieren ! Reihe Physik, Verlag Harri Deutsch, Zentralbibliothek für Physik 032011/x, Serienbuchtitel, Serie über verschiedene naturwissenschaftliche Phänomene. Die Serie widmet sich auch den Phänomenen der kognitiven Modellierung.Es finden sich über 30 Bücher zu diesem Thema in der Serie. Schmid, Ute: Kognitive Modellierung :eine Einführung in logische und algorithmische Grundlagen Heidelberg:Spektrum, Akad. Verl.,1996- 455 S.:Ill.,graph. Darst.(Spektrum Hochschultaschenbuch) Literaturverz. S. [381]-396 ISBN 3-86025-367-0 Universitätsbibliothek Wien HB-Mag.-->I-1199923 Interessante Meinungen, fördert das Nachdenken. ____________________________________________________________________________________________ Literaturverzeichnis 88 Snyder, Solomon H.: Chemie der Psyche : Drogenwirkungen im Gehirn . [Aus d. Amerikan. übers. von Ingrid Horn]. - 3. Aufl. - Heidelberg : Spektrum-d.-Wiss.-Verl.-Ges., 1990. - 224 S. : zahlr. Ill., graph. Darst. (SpektrumBibliothek ; 16)Einheitssacht.: Drugs and the brain <dt.>. - Literaturverz. S. 215 ISBN 3-922508-86-3 Alles was es über Neurotransmitter zu wissen gibt und manches mehr. Beste Darstellung der Chemie der Psyche in aller Kürze. Brain mechanisms : papers in memory of Robert Thompson / ed. by Francis M. Crinella and Jen Yu. - New York, NY : The New York Acad. of Sciences, 1993. - 233 S. : Ill., graph. Darst. (Annals of the New York Academy of Sciences ; 702)Literaturangaben ISBN 0-89766-759-x - ISBN 0-89766-760-3 Zentralbibliothek für Physik in Wien 7306/702 ->> FR-Interessante Aufsatzsammlung. Wiener, Norbert: Kybernetik : Regelung und Nachrichtenübertragung im Lebewesen und in der Maschine / Norbert Wiener.[Übers. von E. H. Serr unter Mitarb. von E. Henze]. - 2., rev. u. erg. Aufl., 8. - 10.Tsd. Düsseldorf ; Wien : Econ-Verl.,1965. - 287 S. : Ill., graph. Darst. Nachdenken ! ___________________________________________________________________________________ 89 Brain Modelling