Neurologische Grundlagen - Goethe
Werbung

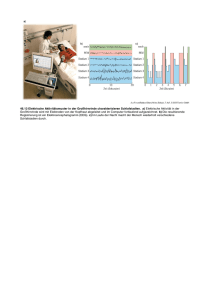
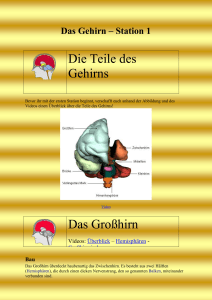
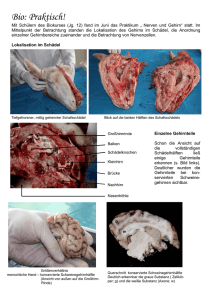
Prof. Dr.med. Wolfgang Schlote Neurologisches Institut (Edinger Institut) Deutschordenstr. 46 der Johann Wolfgang Goethe-Universität e-Mail: [email protected] Titelbild Human Brain Mapping 1993, zeigt eines der ersten PET-Aufnahmen (Positron Emission Tomographie) zur funktionellen Aktivierung lokaler Hirnregionen Aktuelle Publikation in Brain and Language 93, 2005, 32-45 über die differente Lokalisation von belebten und unbelebten Objekten mit dem fMRI (funktionelles Magnet Resonanz Imaging), dem gegenwärtig besten in vivo anwendbaren Verfahren Abbildung aus der vorangehenden Publikation, zeigt die erheblich verbesserte topographische Auflösung gegenüber dem PET von 1993 Gehirn, Schnitt durch die Mittellinie, zeigt Großhirn und Kleinhirn (gelb) und Hirnstamm (weiß) sowie Hirnkammersystem (grün) und Hirnhäute (blau) aus dem Buch „Nervensystem“ von Mennel, Gebert und Bewermeyer, Schattauer Verlag Stuttgart 1992. Die phylogenetische internationale Gliederung des Gehirns enthält jedoch 5 Teile (Endhirn, Zwischenhirn, Mittelhirn, Nachhirn und verlängertes Mark). Zentralnervensystem (= Gehirn und Rückenmark) und peripheres Nervensystem (hier: Hirnnerven), aus didaktischen Gründen ist links das sensible System, rechts das motorische System dargestellt, in Wirklichkeit sind beide Systeme beidseits vorhanden. Die römischen Ziffern beziehen sich auf die 12 Hirnnerven. IC bedeutet Inselcortex (Sprache!) Historische Darstellung hypothetischer Funktionen der Großhirnrinde (Cortex cerebri) aus dem Jahr 1864. Simplifizierte Einteilung der Großhirnrinde in funktionelle Sphären durch Karl Kleist, Frankfurt (früherer Direktor der Psychiatrischen Universitätsklinik) aus dem Jahr 1921. Außenseite der menschlichen Hirnhemisphäre. Die Zahlen beziehen sich auf die international gültigen morphologisch differenzierten Brodmann-Felder (Areae), die noch heute Bezugspunkte für die Verständigung über Hirnlokalisationen sind. Innenseite der Hirnhemisphäre, nach Karl Kleist Differenzierte funktionelle Gliederung des menschlichen Gehirns nach Karl Kleist, aufgrund seiner Erfahrungen an Hirnverletzten (aus dem 1. Weltkrieg!). Die Lokalisationen stimmen im wesentlichen, die Grenzen sind aber zu scharf gezogen, sie variieren individuell! Sehr genau festgelegte Lokalisation in den Projektionszentren (Motorik und Sinneszentren), hier am Beispiel der vorderen Zentralwindung (Willkürmotorik, rechts) und der hinteren Zentralwindung (Tastzentrum, links) mit dem kopfstehenden Humunculus. In der Realität gibt es beide Zentren in beiden Hirnhälften, mit gekreuztem Körperbezug. Sprachregion mit grüner Linie umfahren (kein Zentrum! Assoziationsbereich, sog. sekundäre Leistung). Zeichenerklärung: B Broca-Bereich, W Wernickebereich, F Fasciculus arcuatus, L Lexikon, S Gyrus supramarginalis, Feinmotorik, Praxie, A Gyrus angularis , transferiert visuelle Signale zum Wernicke-Bereich, „Carrefour visuo-acustique“). Außerhalt der Sprachregion die Projektionszentren M Willkürmotorik, T Tastzentrum, E Extrapyramidale Motorik, H Hörzentrum, V Sehzentrum Blutversorgung des Gehirns durch die 3 großen Hirnarterien A. cerebri anterior, media und posterior (unerstrichen). Nach Klaus Poeck (Aachen) ist die BrocaAphasie Folge einer Blutversorgungsstörung in der A. praerolandica, die Wernicke-Aphasie Folge einer Blutversorgungsstörung in der A. temporalis posterior, beide entspringen aus der A. cerebri media. Gehirn von unten gesehen (Basalfläche) mit Augäpfeln, Hirnstamm, Kleinhirn (eine Hälfte entfernt) und Hirnnerven (einseitig dargestellt) Gleiche Sicht wie vorangehendes Bild, die Zahlen der Hirnnerven IXII sind eingetragen. Sie gehören zum peripheren Nervensystem (außer Nr. I und II) und enthalten afferente (zum Hirn führende) und efferente (vom Hirn wegführende) Nervenfasern („gemischte Nerven“). Modell einer Nervenzelle im ZNS (Typ Pyramidenzelle) mit dem Empfangsteil (multiple apikale und basale Dendriten, Zellkörper= Soma) und dem Sendeteil (der Neurit = Axon), feine Linie an der Basis des Zellkörpers entspringend, Ende von mir rot markiert. Rechts vergrößerte Darstellung von axodendritischen Synapsen, die mit sog. Dornfortsätzen der Dendriten verknüpft sind, und einer axosomatischen Synapse. From: Hamlyn Horizontale Gliederung der Großhirnrinde in 6 Schichten (layers) I-VI rechts und Verknüpfung dieser Schichten mit Bildung von Netzwerken (neuronal networks), rechts. Vertikale Gliederung der Großhirnrinde in Säulen (Columns), links oben. Cave: diese Säulen entsprechen nicht den funktionellen hypothetischen Modulen von Fodor!! Diese sind weit ausgedehnter und können sich über mehrere Hirnareale erstrecken. Nervenzellen in corticalen Säulen (schwarz oder weiß dargestellt) können kontinuierlich (A) oder diskontinuierlich (B) miteinander via Neuriten und Synapsen verbunden sein. From: Amir, Harel and Malach 1993. Entwicklungsperioden des menschlichen Gehirns, dargestellt aufgrund der Zunahme des Hirngewichts ab Konzeption. A Periode der embryonalen Hirnanlage, B Periode der embryonalen Gestaltbildung des Gehirns, C Periode der stürmischen Hirnentwicklung, bedingt vor allem durch die Bildung der (gewichtsrelevanten) Markscheiden an den Neuriten (Myelogenese) und der Dendritenbäume.Mitten in dieser Periode liegt der Zeitpunkt der Geburt (*). Periode D weitere Entwicklung des Gehirns bis zum Erreichen des des Hirngewichts des Erwachsenen (1200-1400 g) im 12.-13. Lebensjahr. E Definitive strukturelle Ausreifung des Gehirns (nicht mehr gewichtsmäßig relevant) mit Bildung der letzten Neuritenausläufer und Synapsen innerhalb der Großhirnrinde. Entwicklung der Dendritenbäume (und damit der Anzahl der Synapsen) von der Geburt bis zum Ende des zweiten Lebensjahres in der Großhirnrinde (hier: Sehzentrum) bei Anwendung der GolgiMethode (Silberimprägnation) nach Conel 1959 Die Bildung der Dendriten in der Großhirnrinde erfolgt zum größeren Teil (ca. 70%) aufgrund genetischer Programmierung, weitere Dendriten (3035%) aufgrund von Stimuli aus der Umwelt (Sinnesorgane). Dieser Anteil führt zur maximalen Leistungsfähigkeit des Systems, hier dargestellt durch Vergleich des Sehzentrums von Mäusen bei normaler Aufzucht (obere Kurve) und bei Dunkelaufzucht (unten). Plastizität des Gehirns nach Schädigungen (schematisch). Nach Ausfall einer Nervenzelle geht der Neurit zugrunde, die Zielzelle wird nicht mehr innerviert. Transplantierte (unter günstigen Bedingungen auch benachbarte) Nervenzellen bilden neue Neuriten aus und können das deafferenzierte Neuron erreichen und dort neue Synapsen herstellen. Dies funktioniert nur über kurze Entfernungen (Größenordnung: Millimeter). Spracherwerb. Die notwendigen Vorbedingungen und die verschiedenartigen Faktoren, die beim Spracherwerb mitwirken, sind anschaulich von Wendlandt in seinem Buch „Sprachstörungen im Kindesalter“ dargestellt. Unabdingbare Quelle ist die Kommunikation (Gießkanne). Die Ursachen von Sprech- und Sprachentwicklungsstörungen bei Kindern und Jugendlichen unterscheiden sich von den Ursachen der Sprach- und Sprechstörungen (Aphasien und Dysarthrien) bei Erwachsenen nach Abschluss des Spracherwerbs, sie überlappen sich nur teilweise. Hören und Sprache. Akustische Signale treffen auf beiden Seiten des Gehirns in den primären Hörzentren (Heschl’sche Querwindungen) ein (oben links). Handelt es sich um linguistische Informationen, werden sie einseitig in der sprachdominanten Hemisphäre in der Wernicke-Region verarbeitet. Diese ist nach Messungen am Planum temporale (Bild unten links) durch Geschwind und Levitzky (1968) bei 65% der Erwachsenen links voluminöser als rechts, bei 21% rechts voluminöser, bei 14% seitengleich ausgebildet. Eigene Untersuchungen haben diese Asymmetrie an einer Stichprobe bestätigt. Horizontalschnitt durch das Gehirn eines Erwachsenen. Das Planum temporale (Wernicke-Region) ist beidseits rot markiert, vor diesem liegen die Heschl’schen Querwindungen (primäres Hörzentrum). Das größere Planum temporale der sprachdominanten Hemisphäre enthält nicht mehr Nervenzellen, sondern die Nervenzellen haben größere Abstände, zwischen ihnen befindet sich somit ein reicher ausgebildetes neuronales Netzwerk (morphologisch: Neuropil). Die Überlappung der Dendritenbäume ist geringer (rote Markierung), damit ist die Trennschärfe bei der Verarbeitung linguistischer Informationen besser. Witelson and Palay (Brain 96, 641-646 (1973) konnten zeigen, dass die Asymmetrie des Planum temporale bereits bei Neugeborenen zu finden ist. Seit dieser bedeutenden Entdeckung ist klar, dass damit ein phylogenetisch erworbenes Angebot der Natur vorliegt, die Sprachregion auf der besser ausgestatteten Seite zu etablieren. Offenbar ist es ein Selektionsvorteil, Sprachkompetenz in einer Hirnhemisphäre zu konzentrieren. Die nicht-sprachdominante Hemisphäre nimmt in jedem Fall in begrenztem Maß an sprachlichen Leistungen teil, wie aus den Erfahrungen bei Aphasie bekannt ist. Lange vor Erfindung der bildgebenden Verfahren (Imaging Methods) konnte an der freigelegten Großhirnrinde bereits die Sprachlokalisation geprüft werden, und zwar durch Unterdrückung einer Sprachantwort (speech arrest) nach Reizung mit schwachen Strömen. Die ersten Ergebnisse wurden 1952 von Penfield und Rasmussen publiziert. Umgekehrt hat man viel später in den gleichen Regionen durch Reizung von Hirnarealen sprachliche Äußerungen provoziert, ebenfalls bei Hirnoperationen, zuerst durch Zusammenarbeit des Neurophysiologen Creutzfeldt mit dem Neurochirurgen Ojemann Eric Lenneberg hat als erster darauf hingewiesen, dass bei Hirnschädigungen während des Spracherwerbs und Sprachverlust eine neue Sprachkompetenz erworben werden kann, wenn die Schädigung bis zum 5. Lebensjahr eingetreten ist. Dies wurde bisher mehrfach bestätigt, wie hier von Vargha-Kadem und Mitarbeitern. Bei einer nach dem 5. Lebensjahr eintretenden Schädigung ist dies nur noch teilweise möglich. glich Es gibt sogar Berichte über neue, durch Überkompensation besonders gute wenn auch stark verlangsamt erworbene Sprachkompetenz wie hier nach neurochirurgischer, wegen eines Tumors und unstillbarer epileptischer Anfälle notwendig gewordener Entfernung der linken Großhirnhemisphäre. Wie sind die Beziehungen zwischen Denken und Sprache? Gibt es kognitive Leistungen ohne Sprache? Die Erfahrungen mit Aphasikern sind geeignet, diese Frage zu bejahen. Aus ganz anderem Blickwinkel hat sich Einstein zu dieser Frage geäußert, als er gefragt wurde, wie er denke. Frühe wissenschaftliche Untersuchungen (Piaget, Wygotsky, Whorf) kommen zu verschiedenen Resultaten. Moderne linguistische Ansätze von Baldo und Mitarbeitern (Brain and Language 2005, 240-250) zur Untersuchung der inneren Sprache sind in ihren Schlussfolgerungen vorsichtig: „Recent evidence suggests that such internal verbalisation do indeed support normal cognition“ – „Such findings imply that language supports cognition“, vor allem bei Problemlösungsaufgaben. Auch transcorticale Aphasien sprechen hierfür. denn bei ihnen ist der Einbezug kognitiver Leistungen in die Sprachkompetenz gestört. Im Lauf der Phylogenese wurde der frei verfügbare, nicht durch vitale Funktionen besetzte Bereich der Großhirnrinde, der „non committed cortex“, hier weiß dargestellt, immer größer. So konnte er in Sprachperzeption und –produktion einbezogen werden. Literaturverzeichnis