Kognitive Neurowissenschaft des Gedächtnisses
Werbung

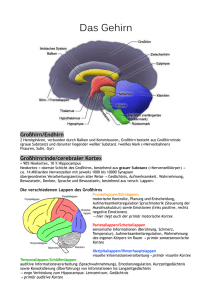
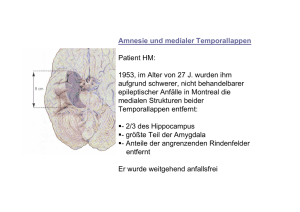
Fachrichtung Psychologie Vorlesung WS 2014/15 Kognitive Neurowissenschaft des Gedächtnisses: Multiple Gedächtnissysteme Thomas Goschke 1 Literaturempfehlungen Gluck, M.A., Mercado, E. & Myers, C.E. (2010). Lernen und Gedächtnis. Vom Gehirn zum Verhalten. Heidelberg: Spektrum Verlag. Kapitel 3. Purves et al. (2013). Principles of cognitive neuroscience. (2nd ed.). Sinauer. Chapter 8 + 9. Gazzaniga, M., Ivry, R. & Mangun, R. (2013). Cognitive neuroscience. The biology of the mind (4th ed.). Norton. Chapter 9. Forschungsansätze und Paradigmen Neurowissenschaftliche Perspektive Grundlage aller psychischen Prozesse ist die Signalübertragung zwischen Nervenzellen Gedächtnisleistungen beruhen auf erfahrungsabhängigen Veränderungen der neuronalen Signalübertragung 4 „Any activity-dependent process that modifies, in a sufficiently stable and longlasting way, the excitatory or inhibitory interactions between pairs of neurons could serve as a mechanism of learning, and any long-lasting alteration of intercellular communication can be considered an engram." (Singer, 1990, S.211) 6 Fragen zur Neuropsychologie des Gedächtnisses Welche Hirnstrukturen sind an Gedächtnisleistungen beteiligt? Sind Gedächtnisfunktionen an einem bestimmten Ort im Gehirn lokalisiert? Gibt es unterschiedliche Gedächtnissysteme (und wenn ja: welche und wie viele)? Was sind die Funktionsprinzipien unterschiedlicher Gedächtnissysteme? Karl Lashley (1950): „In search of the engram“ Lashley trainierte Ratten, Futter in einem Labyrinth finden Nach dem Lernen zerstörte er unterschiedliche Teile der Hirnrinde Gedächtnisfehler hingen von Menge zerstörten kortikalen Gewebes ab; Ort der Läsion spielte geringere Rolle Lashley schloss daraus: • Verschiedene kortikale Areale sind gleichermaßen für das Lernen geeignet (Äquipotentialitätsthese) • Gedächtnisspuren sind nicht an einem Ort lokalisiert, sondern über den Kortex verteilt Kritik: • Die Ratten nutzten u.U. Informationen aus unterschiedlichen Sinnesmodalitäten • Keine Untersuchung subkortikaler Strukturen Fehler beim Labyrinth-Lernen als Funktion des Ausmaßes kortikaler Läsionen bei Ratten Der Fall H.M. Patient mit schwerer Epilepsie (seit dem 10. Lebensjahr) 1953 im Alter von 27 Jahren operative bilaterale Entfernung großer Teile des medialen Temporallappens (inkl. des anterioren Hippocampus), als ultima ratio zur Eindämmung der Epilepsie Nach der OP: • Normale Intelligenz, Intakte Wahrnehmung, Sprache • Intaktes Kurzzeit- und Arbeitsgedächtnis (normale Gedächtnisspanne) Massive anterograde Amnesie • Verlust der Fähigkeit, neue Ereignisse oder Fakten zu behalten • Kann normale Unterhaltung führten, aber erinnert sich nach wenigen Minuten der Ablenkung nicht mehr an das Gespräch Retrograde Amnesie • Beeinträchtigte Erinnerung an Ereignisse, die vor der Operation stattfanden • Aber: Erinnerung an weit zurückliegende Ereignisse (> 11 Jahre vor der Operation) ist erhalten (Milner, Corkin & Teuber, 1968; Corkin, 2002) H.M. (Henry Gustav Molaison, 1926-2008) “Every day is alone in itself, whatever enjoyment I’ve had, and whatever sorrow I’ve had…. Right now I’m wondering, have I done or said something amiss? You see, at this moment everything looks unclear to me, but what happened just before? That’s what worries me. It’s like waking from a dream; I just don’t remember.” Gabrieli, Cohen & Corkin, 1988 • H.M. und neurologisch gesunde Kontrollprobanden sollten Definitionen von Worten lernen, die sie vorher nicht kannten No. of errors • Auswahl der korrekten Definition, eines Synonyms oder passenden Satzrahmens No. of trials until learning criterion Beispiel für H.M.‘s Beeinträchtigung beim Lernen und Abrufen neuer Fakten 13 © 2011 Cengage Learning Dissoziation von Arbeitsgedächtnis und deklarativem Gedächtnis Patient H.M.: Läsion im bilateralen anterioren medialen Temporallappen Patient K.F.: Läsion im linken temporoparietalen Kortex 14 Medial-temporales Gedächtnissystem Temporallappen Hippokampus Amygdala Gazzaniga et al. (2010). © W.W.Norton Medial-temporales Gedächtnissystem Gluck, Mercado and Myers: Learning and Memory, Copyright © 2008 by Worth Publishers Medial-temporales Gedächtnissystem 17 H.M.‘s Läsion Strukturen im medialen Temporallappen, von denen man aufgrund des Berichts des Chirurgen annahm, dass sie bei H.M. entfernt worden waren 21 Späterer MRT Scan zeigte, dass Teile von H.M.’s posteriorem Hippocampus nicht entfernt worden waren. Allerdings zeigte Gewebe Anzeichen von Atrophie. Corkin et al. (2007). H.M.’s medial temporal lobe lesion: Findings from magnetic resonance imaging. Journal of Neuroscience 17: 3964–3979. MRT-Scans von H.M.’s Gehirnläsion Anteriorer Hippocampus wurde vollständig entfernt 22 Posteriorer Hippocampus ist in beiden Hemisphären noch vorhanden Widerlegte die 40 Jahre lang vorherrschende Meinung, dass H.M. keinen Hippocampus mehr hat! Amnesien Hirnorganisch bedingte Gedächtnisstörungen als Folge von chirurgischen Eingriffen, Schädel-Hirn-Traumata, Schlaganfällen, Infektionen (Enzephalitis), Neurodegenerativen Erkrankungen (Alzheimer- u. Korsakoff-Krankheit) • • • medialer Temporallappen (Hippokampus u. angrenzende Gebiete) Operation (H.M.) Sauerstoffmangel (R.B.) © 2013 by Worth Publishers • Diencephalon (dorsomedialer Kern des Thalamus; Mammilarkörper) • Pat. N.A. (Fechtverletzung) • Korsakoffkrankheit (Vitamin-B1-Mangel als Folge von Unterernähung bei Alkoholsucht) Beeinträchtigte freie Reproduktion bei Patient N.A. und Korsakoffpatienten Evidenz für die Rolle des Hippokampus für die Enkodierung ins deklarative Gedächtnis • Enkodierung (im MRT-Scanner): • Probanden kategorisierten Worte als belebt/unbelebt oder groß/klein • Farbe signalisiert die jeweils auszuführende Kategorisierung • Test (nach der MRT-Session): • Probanden beurteilten alte und neue Worte danach, • (a) wie sicher sie diese wiedererkannten • (b) ob sie zuvor in rot oder grün dargeboten worden waren Subsequent-memory effect Aktivierung im posterioren Hippocampus und posterioren parahippocampalen Kortex sowie im Frontalkortex während der Enkodierung korrelierte mit der späteren Erinnerungsleistung Konsolidierung und Interaktionen zwischen Hippokampus und Neocortex 31 Hirnschädigung Zeit Retrograde Amnesie Anterograde Amnesie Beeinträchtigte Gedächtnis für Ereignisse, die vor der der Hirnverletzung stattfanden Beeinträchtigtes Gedächtnis für Ereignisse oder neue Fakten, die nach der Hirnverletzung enkodiert werden Zeitlicher Gradient bei der retrograden Amnesie Ribot (1882): Retrograde Amnesie nach Kopfverletzungen betrifft ältere Erinnerungen weniger stark als jüngere Erinnerungen Gluck, Mercado and Myers: Learning and Memory, Copyright © 2008 by Worth Publishers Gedächtnisleistung Zeitlicher Gradient bei der retrograden Amnesie Normale Vergessenskurve Kontrollgruppe Hippokampusläsion 0,5 Zeit zwischen Lernen und Zeitpunkt der Läsion Weiter zurückliegende Ereignisse sind weniger von der Läsion betroffen! Zeitlicher Gradient bei retrograder Amnesie (a) Ratten; Futterpräferenz (b) Ratten; kontextuelle Furchtkonditionierung (c) Affen; Objektdiskrimination Kreise: Hippokampale Läsion Quadrate: Kontrollgruppe Days between prior learning and lesion McClelland et al., 1995, Psychol Rev Zeitlicher Gradient bei zeitweiser retrograder Amnesie nach einer elektrokonvulsiven Behandlung 41 Konsolidierungshypothese Abruf neuer Gedächtnisinhalte hängt nur für gewisse Zeit von medialtemporalen Strukturen ab Medial-temporale Strukturen können nicht der anatomische Ort sein, an dem Gedächtnisspuren langfristig gespeichert werden Konsolidierungshypothese (vgl. bereits Müller & Pilzecker, 1900) • Neue Gedächtnisspuren sind zunächst labil und störanfällig • Langzeitgedächtnis beruht auf strukturellen Änderungen neuronaler Verknüpfungen • Diese Veränderungen benötigen Zeit („Konsolidierung“) Neues Ereignis Konsolidierungshypothese Evidenz für Konsolidierungsprozesse • Ereignisse nach der Enkodierung können die Stabilität neuer Gedächtnisspuren beeinflussen z.B. Elektroschocks; elektrische Reizung von Nervenzellen; Hemmung der Proteinsynthese; Blockierung von Neurotransmittern; Ausschüttung von Hormonen oder Peptiden; Stress und Emotionen; • Effekte werden kleiner, je größer der zeitliche Abstand zur Lernepisode ist Zwei Arten von Konsolidierungsprozessen (Dudai, 2011) 1. Bildung neuer Gedächtnisspuren als Folge synaptischer Veränderungen (Minuten – Stunden) 2. Stabilisierung und Reorganisation von Gedächtnisspuren (Wochen – Jahre) Zwei Formen der Konsolidierung Mögliche Funktion des hippokampalen Systems Einzelaspekte eines Ereignisses (Ort, Form, Farbe, Bedeutung etc.) werden in verteilten Regionen des Neokortex repräsentiert Vermutete Funktion des HS = Schnelle Bindung von Einzelaspekten einer neuen Episode zu einer Gesamtrepräsentation Beim Abruf ist HS für gewisse Zeit notwendig, um Einzelaspekte der Episode in verteilten neokortikalen Arealen zu reaktivieren Wiederholte Reaktivation Bildung direkter Assoziationen zwischen den im Neokortex gespeicherten Teilaspekten Sind diese Assoziationen hinreichend stark Aktivierung eines Teilaspekts führt zum Abruf der gesamten Episode HS ist nicht länger notwendig für den Abruf Standardmodell der Interaktion von Hippokampus und Neokortex während der Konsolidierung © 2008 Worth Publishers © 2013 Sinauer Ass. Standardmodell der Interaktion von Hippokampus und Neokortex während der Konsolidierung Einzelaspekte eines Ereignisses werden in verteilten Kortexregionen gespeichert, die an der Verarbeitung des Ereignisses beteiligt sind Während der Konsolidierung werden die Gedächtnisspuren stabilisiert Im Lauf der Zeit werden Im Hippokampus werden direkte Assoziationen „kondensierte“ Repräsentationen zwischen den Aspekten gespeichert, die wie „Zeiger“ des Ereignisses in oder „Indizes“ auf die kortikalen neokortikalen Regionen Repräsentationen verweisen etabliert 47 Abrufhinweise aktivieren die hippokampale „Indizes“, was zur Reaktivierung der kortikalen Gedächtnisspuren und zum Abruf des Ereignisses führt Abrufhinweise können externe Reize sein oder intern generiert werden (unter Beteiligung des PFC) Wenn sich direkte Assoziationen zwischen den Aspekten des Ereignisses in neokortikalen Regionen gebildet haben, kann ein Einzelaspekt zur Reaktivierung der gesamten Gedächtnisspur führen (inhaltsadressierter Abruf) Erklärung der Folgen von Hirnschädigungen durch das Standardmodell Kleine kortikale Läsionen nur schwache Effekte auf deklaratives Gedächtnis, da Gedächtnisspuren distribuiert in mehreren Regionen gespeichert sind selektive Gedächtnisausfälle, je nachdem welche Region geschädigt ist (z.B. Ausfall kategorienspezifischen Wissens über belebte Dinge) Hippokampusläsionen anterograde Amnesie, da keine neuen „Indexspuren“ angelegt werden können retrograde Amnesie für rezente Erinnerungen, da Zerstörung der „Indexspuren“ Zugriff auf die kortikalen Gedächtnisspuren blockiert Weiter zurückliegende (konsolidierte) Erinnerungen sind weniger stark betroffen, da diese unabhängig vom Hippokampus durch Abrufhinweise reaktiviert werden können 49 Simulationsmodell der Interaktion von Hippokampus und Neokortex (Alvarez & Squire, 1994) Simuliertes neuronales Netzwerk wurde mit Mustern trainiert Verbindungen zwischen kortikalen Regionen ändern sich nur langsam Verbindungen zwischen medial-temporalem System (MTL) und Kortex ändern sich schnell Nachdem Muster im Kortex enkodiert ist, wird es wiederholt durch das MTLSystem reaktiviert inkrementelle Verstärkung der Verbindungen zwischen den kortikalen Repräsentationen Simulationsmodell der Interaktion von Hippokampus und Neokortex (Alvarez & Squire, 1994) Nach dem Lernen: „Läsion“ des medial-temporalen Systems Test: Aktivierung eines Teilmusters in Kortexregion 1 Wie gut wird assoziiertes Muster in Kortexregion 2 reaktiviert? Lädiertes Modell produziert zeitlich abgestufte retrograde Amnesie Verbindungen zwischen Hippokampus und Neokortex Neokortikale Assoziationsareale projizieren zum parahippokampalen + perirhinalen Kortex Von dort gelangt die Information zum entorhinalen Kortex = Konvergenzzone, in der hoch verarbeitete Informationen aus neokortikalen Systemen zu einer Gesamtrepräsentation integriert werden Diese Information gelangt über den Gyrus Dentatus zur CA3- und CA1-Region des Hippokampus und wird von dort über das Subiculum zurück zum entorhinalen Kortex geleitet, der zurück in die neokortikalen Regionen projiziert, aus denen die Informationen stammten 53 Famous faces test Probanden sahen berühmte und nicht berühmte Gesichter und sollten versuchen, sich an die Namen der Personen zu erinnern Hirnaktivität während des Famous faces test Aktivierung im rechten enthorinalen Kortex nahm umso mehr ab, je weiter zurück die abzurufenden Erinnerungen lagen (enthorinaler Kortex: Region, durch die Information vom Kortex zum Hippokampus und umgekehrt geleitet wird) Haist et al., 2001 Rekonsolidierung Standard-Konsolidierungs-Modell: • Neue Gedächtnisspuren sind für eine gewisse Zeit nach der Enkodierung labil und interferenzanfällig Rekonsolidierungshypothese • Auch ältere (konsolidierte) Gedächtnisspuren können wieder anfällig für Störungen werden, sobald sie abgerufen werden 58 Rekonsolidierung Furchtkonditonierung: Ton + Schock Bedingung 1: • 24 Std. später wurde der CS präsentiert um die Furcht-Gedächtnisspur zu reaktivieren • Direkt danach erhielten einige Tiere einen electrokonvulsiven Schock (ECS) und andere nicht Bedingung 2: • Keine Reaktivierung der Gedächtnisspur • ECS vs. kein ECS A.V.: Konditionierte Furchtreaktion Resultat: • nach Reaktivierung der Gedächtnispur störte ECS den Abruf der gelernten Assoziation • ECS hatte keinen Effekt, wenn die Gedächtnisspur nicht reaktiviert wurde 59 Rekonsolidierung Gedächtnisinhalte können in aktivem (Kurzzeitgedächtnis) oder inaktivem (Langzeitgedächtnis) Zustand sein Zwei Möglichkeiten, Gedächtnisspuren in aktiven Zustand zu versetzen: • Neue Erfahrungen erzeugen aktive Gedächtnisspur • Abruf / Reaktivierung gespeicherter Gedächtnisspuren transformiert diese wieder in aktiven Zustand Aktivierte Gedächtnispuren sind anfällig für Störungen 60 Rekonsolidierung Experiment von Nader et al 2000) Ratten wurden auf einen auditorischen CS konditioniert, der mit einem Schock (US) gepaart wurde Nach der Reaktivierung der konditionierten Furchtreaktion wurde die Proteinsynthese im basolateralen Kern der Amygdala gehemmt Anisomycin beeinträchtigte das Langzeitgedächtnis für die konditionierte Furchtassoziation, aber hatte keinen Effekt auf die kurzzeitige Speicherung 61 Rekonsolidierungshypothese • Abruf einer Gedächtnisspur hat zwei Effekte: • (A) Die synaptischen Verbindungen zwischen den Neuronen, die die Gedächtnisspur enkodieren, werden aufgelöst • (B) Ein neuerlicher Prozess der Proteinsynthese wird ausgelöst, der zur Rekonsolidierung der Gedächtnisspur führt. • Wird dieser Prozess gestört / unterbunden, wird die Gedächtnisspur geschwächt oder geht verloren. 62