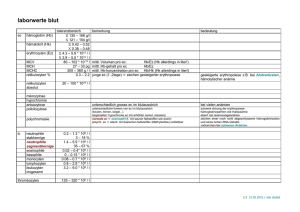
Immunotoxikologische Untersuchung zur Biokompatibilität von
Werbung

Tierärztliche Hochschule Hannover Immunotoxikologische Untersuchung zur Biokompatibilität von abbaubaren Implantaten aus Magnesiumbasislegierungen INAUGURAL-DISSERTATION zur Erlangung des Grades eines Doktors der Veterinärmedizin - Doctor medicinae veterinariae (Dr. med. vet.) vorgelegt von Klaas Feser Wilhelmshaven Hannover 2009 Wissenschaftliche Betreuung: Univ.-Prof. Dr. M. Kietzmann Institut für Pharmakologie, Toxikologie und Pharmazie 1. Gutachter: Prof. Dr. M. Kietzmann 2. Gutachter: Prof. Dr. A. Meyer-Lindenberg Tag der mündlichen Prüfung: 15.12.2009 Meiner lieben Familie Inhaltsverzeichnis Inhaltsverzeichnis 1. Einleitung 01 2. Literaturübersicht 03 2.1. Dendritische Zellen 03 2.1.1. Hämatopoese und Reifung der DC 03 2.1.2. Migration naiver DC und Antigen-Aufnahme 05 2.1.3. Migration reifender DC in lymphatische Organe 07 2.1.4. Antigenprozessierung und –präsentation durch DC 09 2.2. Rolle von Magnesium im Organismus 11 2.2.1. Magnesium: Resorption und Ausscheidung 12 2.2.2. Toxikologische Bedeutung von Magnesium 13 2.3. Magnesiumbasislegierung 2.3.1. Magnesium-Calcium-Legierungen 14 17 2.4. Capture-ELISA 19 2.5. FACS-Analyse 19 2.5.1. Annexin V-FITC und PI 20 2.6. XTT-Proliferationstest 21 2.7. MLR (Mixed Leukocyte Reaction) 21 3. Material und Methoden 3.1. Material 22 22 3.1.1. Geräte 22 3.1.2. Verbrauchsmaterial 23 3.1.3. Reagenzien 23 3.1.4. Magnesiumbasislegierungen 24 3.1.5. Puffer und Lösungen 25 3.1.6. Kits 26 3.1.7. Antikörper 27 3.1.8. Mäuse 27 3.1.9. Software 27 Inhaltsverzeichnis 3.2. Methoden 28 3.2.1. Generierung von DC 28 3.2.2. Gewinnung muriner T-Zellen 29 3.2.3. Herstellung der Degradationsmedien 29 3.2.3.1. Bestimmung der Magnesium- und Calciumkonzentration in den Degradationsmedien 30 3.2.4. In-vitro-Untersuchungen mit Magnesiumund Calcium-Salzen 3.2.4.1. Inkubation der DC mit MgCl2 und CaCl2 3.2.5. In-vitro-Untersuchungen mit Magnesiumbasislegierungen 3.2.5.1 32 33 36 Kultivierung der DC in den Degradationsmedien der Magnesiumbasislegierungen 37 3.2.6. Zytokinbestimmung 39 3.2.7. Migrationsassay 40 3.2.8. FACS-Analyse 41 3.2.8.1. Vitalitätstest (Annexin V-FITC und PI, XTT) 41 3.2.8.2. Untersuchung der Expression von CD11c und CD86 42 3.2.8.3. MLR (Mixed Leukocyte Reaction) 42 3.3. Statistische Auswertung 4. Ergebnisse 43 44 4.1. Magnesium und Calciumkonzentration in den Degradationsmedien 44 4.2. Vergleichende Darstellung der In-vitro-Ergebnisse zwischen MgCl2, CaCl2 und den Magnesiumbasislegierungen 45 4.2.1. DC-Zellzahl und MØ-Zellzahl im Vergleich zwischen MgCl2, CaCl2 und Degradationsmedien 45 4.2.2. Vitalität (Annexin V-FITC und PI, XTT) 52 4.2.3. Freisetzung von TNF-α 61 4.2.4. Migrationsassay 64 4.2.5. Expression der Oberflächenmoleküle CD11c und CD86 66 4.2.6. T-Zellproliferation in der MLR 71 Inhaltsverzeichnis 5. Diskussion 5.1. Degradationsversuche der Magnesium-Calcium-Legierungen 76 76 5.2. DC-Zellzahl, MØ-Zellzahl und Vitalität der DC nach MgCl2- und CaCl2Inkubation bzw. nach Kultivierung in den Degradationsmedien 78 5.3. TNF-α-Sekretion der DC nach MgCl2- und CaCl2Inkubation bzw. nach Kultivierung in den Degradationsmedien 79 5.4. Auswanderungsverhalten von DC nach MgCl2- und CaCl2-Inkubation bzw. nach Kultivierung in den Degradationsmedien 80 5.5. Expression von CD11c/CD86 auf DC nach MgCl2- und CaCl2Inkubation bzw. nach Kultivierung in den Degradationsmedien 81 5.6. Mixed Leukocyte Reaction mit DC, nach Präinkubation mit MgCl2 und CaCl2 bzw. nach Kultivierung in den Degradationsmedien 82 6. Zusammenfassung 85 7. Summary 87 8. Literaturverzeichnis 88 9. Anhang 108 Abkürzungsverzeichnis Abkürzungsverzeichnis Abb. Abbildung A. bidest Aqua bidestillat (zweifach destilliertes Wasser) Ag Antigen APC Antigenpräsentierende Zellen BC Boyden Chamber BSA Bovines Serum Albumin ca. circa Ca2+ Calcium CLP common lymphoid progenitor cells CMP common myeloid progenitor cells CCL Chemokin Ligand CCR Chemokin Rezeptor CD Cluster of Differentiation CFSE carboxyfluorescein diacetate succinimidylester CTL zytotoxische T-Zellen DC Dendritic Cell(s), dendritische Zelle(n) DC-SIGN DC-specific intercellular adhesion molecule-grabbing nonintegrin DMEM Dulbecco`s Modified Eagle`s Medium d.h. das heißt DMSO Dimethylsulfoxid DNCB 2,4-Dinitrochlorobenzol ELISA enzyme-linked immunosorbent assay et al. et alii (und andere) FACS Durchflusszytometer, fluorescensce-activated cell sorter FITC Fluorescein-Isothiocyanat FKS foetales Kälberserum FSC Vorwärtsstreulicht, forwardscatter g Gramm oder Erdbeschleunigung: 9,81m/s2 GM-CSF granulozyte-macrophage colony-stimulating factor Abkürzungsverzeichnis h Stunde iDC unreife DC, immature DC IFN Interferon IL Interleukin LAM Lipoarabinomannan LPS Lipopolysaccharid MØ Makrophagen MCP-1 monocyte chemoattractant protein-1 mDC reife DC, mature DC mg milli(10-3)gramm Mg2+ Magnesium MHC Haupthistokompatibilitätskomplex, major histcompatibility complex min Minute MIP-3β macrophage-inflammatory-Protein-3β ml milli(10-3)liter MLR mixed leukocyte reaction mmol milli(10-3)molar MMP Matrix-Metalloproteinasen MTT Zytotoxizitäts-Test (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenylTetrazoliumbromid) MW Molekulargewicht oder Mittelwert OD optische Dichte o.g. oben genannten NF-κB nuclear factor κB NK-Zellen natürliche Killerzellen PAMP Pathogen-assoziierte molekulare Muster PBS Phosphat-gepufferte Saline PE Phycoerythrin Pen/Strep Penicillin/Streptomycin PGE Prostaglandin E PI Propidium Iodid Abkürzungsverzeichnis pH negativer dekadischer Logarithmus der Protonen-Konzentration PLA Polylaktid PRR Mustererkennungs-Rezeptoren, pattern recognition receptors PS Phosphatidylserin PTH Parathormon RDH recommended daily allowance RNA Ribonukleinsäure XTT Tetrazoliumsalz (Natrium3`-[1-(phenylaminocarbonyl)-3,4tetrazolium]-bis(4-methoxy-6-nitro)-benzolsulfonsäure-hydrat) RANTES regulation and activated normal T-cell expressed and secreted RT Raumtemperatur S.D. Standartabweichung SE Seltene Erden s.o. siehe oben (im selben Abschnitt) SSC Seitwärtsstreulicht, sidescatter Tab. Tabelle TCR T-cell receptor TH-Zelle T-Helfer-Zelle TLR Toll-like receptor TNF Tumornekrosefaktor TM Warenzeichen u.a. unter anderem wt% Gewichtsprozent, weight percent z.B. zum Beispiel % Prozent ® eingetragenes Warenzeichen °C Grad Celsius µ mikro, 10-6 * schwach signifikant (Irrtumswahrscheinlichkeit 5 %) ** signifikant (Irrtumswahrscheinlichkeit 1 %) *** hoch signifikant (Irrtumswahrscheinlichkeit 0,1 %) Einleitung 1. Einleitung Der Einsatz von abbaubaren Implantaten aus Magnesiumbasislegierungen in der Chirurgie setzt bei vollständiger Degradation im Körper eine uneingeschränkte Biokompatibilität voraus. Das Immunsystem spielt dabei eine wichtige Rolle. Fremdstoffe, die in den Organismus gelangen, werden vom Immunsystem erkannt und bewertet. Als Reaktion des Immunsystems kann es zur Ausbildung einer Toleranz oder einer Immunität kommen (JANEWAY et al., 2002). Das Immunsystem besteht aus einer unspezifischen (angeborenen) Immunabwehr und einer spezifischen (erworbenen) Immunabwehr, denen verschiedene Funktionsweisen zugrunde liegen und die sich bei einer Immunantwort gegenseitig ergänzen. Dendritische Zellen (DC) werden der unspezifischen Immunabwehr zugeordnet und spielen, als wichtige Gruppe der antigenpräsentierenden Zellen (APC), eine große Rolle für die Aktivierung und Einbeziehung der spezifischen Immunabwehr, da sie in der Lage sind Antigene aufzunehmen und diese den T-Lymphozyten der regionalen Lymphknoten zu präsentieren. Durch Stimulation der T-Lymphozyten kann somit eine Immunantwort initiiert werden (JANEWAY et al., 2002). Ziel der vorliegenden Arbeit ist es, Erkenntnisse über die Beeinflussung der Zellfunktionen von murinen DC durch abbaubare Magnesiumbasislegierungen zu gewinnen. In der Literatur gibt es lediglich Angaben zur Allergietestung von degradablen metallischen Magnesiumlegierungen (ABELN, 2009). Die Beeinflussung der Funktion von DC durch abbaubare Magnesiumbasislegierungen wurde bisher nicht untersucht. Zum Einsatz kommen reines Magnesium und Legierungen auf Magnesium-Calcium-Basis. In vitro wird zuerst der Einfluss von Magnesium- und Calcium-Salzen auf die unterschiedlichen Phasen der DC-Maturation untersucht. Im Mittelpunkt stehen zum einen die Vitalität der DC und die Expression diverser Oberflächenmoleküle, sowie das Auswanderverhalten der DC in einem Migrationsassay. Die T-Zellstimulation durch DC in der Mixed Leukocyte Reaction (MLR) ist ein weiterer Schwerpunkt dieser Arbeit. -1- Einleitung Einleitung Anschließend werden Degradationsmedien aus verschiedenen abbaubaren Magnesiumbasislegierungen hergestellt. Untersucht wird das Degradationsverhalten sowie die Magnesium- und Calciumkonzentration in den Degradationsmedien mit Hilfe photometrischer Degradationsmedien auf Messungen. die Weiterhin unterschiedlichen wird Phasen der der Einfluss der DC-Maturation hinsichtlich der Vitalität, der Expression diverser Oberflächenmoleküle, sowie des Auswanderverhaltens untersucht. Außerdem wird die T-Zellstimulation durch DC in der Mixed Leukocyte Reaction (MLR) untersucht. -2- Literaturübersicht 2. Literaturübersicht 2.1. Dendritische Zellen Die dendritischen Zellen (DC) sind leukozytären Ursprungs und spielen eine wichtige Rolle in der Überwachung und Steuerung der Immunabwehr. Ihre hauptsächliche Funktion ist die Präsentation von Antigenen an T-Lymphozyten, um diese zu stimulieren. Daher bezeichnet man die DC als professionelle antigenpräsentierende Zellen (APC) (CAUX et al., 2000). DC sind für die Sensitivierung MHC-restringierender T-Zellen, die Entwicklung der T-Zell abhängigen Antikörperproduktion sowie für die Induktion der immunologischen Toleranz verantwortlich (STEINMAN, 1991; STINGL und BERGSTRESSER, 1995). Erstmals wurden die DC 1868 von LANGERHANS in der basalen Epidermis menschlicher Haut entdeckt. Aufgrund ihrer vielen Ausläufer (Dendriten) missdeutete er jedoch ihre Form und hielt sie für Nervenzellen (LANGERHANS, 1868). Erst in der Mitte der siebziger Jahre des letzten Jahrhunderts wurden die DC als solche von STEINMAN und COHN (1973) als neuer Zelltyp in peripheren lymphatischen Organen der Maus entdeckt und beschrieben. Aufgrund ihrer baumartig verzweigten Zytoplasmaausläufer wurden sie als Dendritsche Zellen (dendron, griechisch = Baum) bezeichnet. In den folgenden Jahrzehnten wurden DC zusätzlich im Blut (VAN VOORHIS et al., 1982), im Knochenmark (EGNER et al., 1993) und in verschieden Organen beschrieben, wie zum Beispiel in der Leber (PRICKETT et al., 1988), in der Lunge (SERTL et al., 1986) und im Darm (PAVIL et al., 1996). 2.1.1. Hämatopoese und Reifung der DC Die dendritischen Zellen entwickeln sich über Zwischenstufen der DCVorläuferzellen aus pluripotenten häematopoetischen CD34+-Stammzellen aus dem Knochenmark. Bei den DC-Vorläuferzellen können zwei Typen unterschieden werden: die myeloiden Vorläuferzellen (common myeloid progenitor cells, CMP) und die lymphoiden Vorläuferzellen (common lymphoid progenitor cells, CLP) (LIU, 2001). Die Stimulation von linienspezifischen Vorläuferzellen mit granulocyte- -3- Literaturübersicht macrophage colony-stimulating factor (GM-CSF) (PULENDRAN et al., 2001) und TNF-α führt zur Bildung von vier unterschiedlichen DC-Subtypen. Es entstehen epitheliale DC (Dermis), interstitielle DC (Herz, Niere, Darm, Lunge), myeloide DC und plasmazytoide DC (VERMAELEN und PAUWLS, 2005). Unterschieden werden die DC-Subpopulationen aufgrund ihrer Verteilung im Gewebe, ihrer Morphologie und ihrer Funktion (WU und DAKIC, 2004), wobei jedes Mitglied die Kontrolle über einen anderen Teil des Immunsystems besitzt. Für die Funktion als antigenpräsentierende Zellen durchlaufen myeloide DC verschiedene Stadien der Reifung (Maturation). Man unterscheidet immature, semi-maturierte und mature DC. Am Anfang besiedeln immature gewebeständige DC die peripheren Organe. Diese sind unreife DC mit einer geringen Anzahl costimulatorischer Oberflächenmoleküle und einer Heraufregulation endozytotischer Rezeptoren. In den peripheren Organen verharren sie in einer Art Ruhestadium, um Antigene phagozytotisch und makropinozytotisch aus ihrer Umgebung aufzunehmen (JANEWAY et al., 2002). Nachdem immature DC Antigene aufgenommen haben, werden sie durch inflammatorische Signale (wie z.B. TNF-α, IL-1β) aktiviert und wandern hauptsächlich durch das lymphatische System zu den sekundären Lymphorganen. Bei ihrer Wanderung kommt es zur Reifung maturer DC, die die Antigene (Ag) für die AgPräsentation aufbereiten. Mature DC sind reife DC. Sie unterscheiden sich von den unreifen DC in zwei wesentlichen Punkten: Zum einen können sie keine Antigene mehr aufnehmen, dafür besitzen sie jedoch eine starke costimulatorische Aktivität sowie eine hohe Expression von MHC-Klasse II-Strukturen (CAUX et al., 2000). Über die CD-Moleküle (cluster of differentiation) können immature und mature DC voneinander unterschieden werden (Tabelle 1). Charakteristisch für reife DC ist die Expression des Zelloberflächenproteins CD11c, welches in der Lage ist, eine Adhäsion zwischen Zellen bei der Immunantwort zu ermöglichen. Weiterhin wichtig für die DC-Entwicklung ist CD40. Durch Bindung des Liganden CD40L an aktivierte T-Zellen kommt es zur terminalen Reifung der DC durch Hochregulation von CD80, CD86 (LAMBRECHT, 2001). Diese Hochregulation sorgt zusätzlich für eine Aktivierung naiver T-Zellen (JANEWAY und TRAVERS, 1997). Von immaturen und maturen DC sind zusätzlich die semi-maturierten DC zu unterscheiden. Dieses sind -4- Literaturübersicht DC, die nicht vollständig gereift sind. Sie unterscheiden sich von maturierten DC durch eine geringe Expression von kostimulatorischen Molekülen. Außerdem fehlt den semi-maturierten DC die Fähigkeit, proinflammatorische Zytokine auszuschütten. Aufgrund dieser Tatsache sind sie in der Lage, Selbstantigen und apoptotisches Zellmaterial zu den sekundären Lymphorganen zu transportieren und über dort ansässige T-Zellen eine Toleranz zu vermitteln (LUTZ und SCHULER, 2002; STEINMAN et al., 2003a). Dies geschieht durch Reifung unreifer T-Zellen zu IL-10 produzierenden Supressor-Zellen (JONULEIT et al., 2000; DHODAPKAR et al., 2001). Sie spielen also eine wichtige Rolle in der Immuntoleranz. Tabelle 1: Einteilung der DC nach den Cluster of Differentiation (CD) (JANEWAY und TRAVERS, 1997) CD31/34 MHCII CD11b CD11c CD14 CD25 CD40 CD68 CD80/86 Stammzelle + - - - - - - - - Unreife DC - - + -/+ + - - + - Reife DC - + - + - + + - + 2.1.2. Migration naiver DC und Antigen-Aufnahme Neu generierte DC migrieren aus dem Blut in die nicht lymphatischen Organe. Sie orientieren sich bei ihrer Wanderung anhand von Oberflächenstrukturen auf ortständigen Zellen und Chemokinreizen an den Orten lokaler Infektionen, in die sie verstärkt einwandern. Je nach Reifungsgrad weisen die DC unterschiedliche Rezeptormuster auf und variieren dementsprechend unterschiedlich in ihrem Verhalten gegenüber Chemokinen. Für die Migration von unreifen DC spielen vor allem inflammatorische Chemokine wie z.B. CCL2/MCP-1, CCL3/MIP-1α, CCL4/MIP1β und CCL5/RANTES eine große Rolle (CAUX et al., 2000). Diese Chemokine werden u.a. von unreifen DC nach Antigenkontakt für die Rekrutierung immaturer DC und T-Zellen sezerniert. Zur Unterstützung der Migration sind unreife DC außerdem in der Lage, Matrix-Metalloproteinasen (MMP) zu synthetisieren, welche in den Geweben eine elastolytische Kapazität aufweisen (CAUX et al., 2000). Am Ort der -5- Literaturübersicht Entzündung stehen den DC verschieden Mechanismen zur Antigenaufnahme zur Verfügung: die rezeptorvermittelte Endozytose und Phagozytose und die rezeptorunabhängige Mikro- und Makropinozytose (SALLUSTO et al., 1995). Bei der rezeptorvermittelten Endozytose werden verschiedene Mustererkennungs- Rezeptoren (pattern recognition receptors, PRR) durch die Erkennung von Pathogen-assoziierten molekularen Mustern (pathogen-associated molecular pattern, PAMP) aktiviert (JANEWAY und MEDZHITOV, 2002). Zu den wichtigsten PRR gehören u.a. die C-Typ-Lektin-Rezeptoren (Mannoserezeptor) (ENGERING et al., 1997), DEC-205 und DC-SIGN (DC-specific intercellular adhesion molecule- grabbing nonintegrin) (MAHNKE et al., 2000; GEIJTENBEEK et al., 2000). Außerdem können über Fc-y Rezeptor Typ 1 (CD64) und Typ 2 (CD32) und C3b Komplement-Rezeptor (CD11b) optionierte Partikel und Immunkomplexe aufgenommen werden (FANGER et al., 1996). Die Phagozytose dient der Inkorporation von apoptotischen und nekrotischen Zellfragmenten (CD36 und avβ3 oder avβ5 Integrinen) (ALBERT et al., 1998), Viren und Bakterien (INABA et al., 1993). Bei der rezeptorunabhängigen Makropinozytose wird von den DC permanent extrazelluläre Flüssigkeit mit eventuell vorhandenen Pathogen-Strukturen in Bläschen aufgenommen (JANEWAY et al., 2002). Durch Ausleiten der Flüssigkeit durch Wasserkanäle (Aquaporine) werden die in den Bläschen befindlichen Antigene konzentriert. Durch die Fähigkeit der DC, Antigene zu erkennen, wird schon bei der Initiierung einer Immunantwort die Wichtigkeit der aufgenommenen Strukturen erkannt und somit die DC mit der Information einer Erregerinvasion in Alarmzustand versetzt und aktiviert. Beispielsweise erkennen DC LPS (Lipopolysaccharid) aus Zellwänden gramnegativer Bakterien mit der Kombination der Toll-like receptors (TLRs) TLR2 und TLR4 und CD14 (RESCIGNO et al., 1999). Über die TLRs wird der Transkriptionsfaktor NF-KB (nuclear factor kB) aktiviert. Es kommt zur Reifung der DC und Regulierung verschiedener Effektorfunktionen, einschließlich der Expression des major histocompatibility complex (MHC) und der Zytokinausschüttung (WALLET et al., 2005). Aber auch andere Antigene, wie zum Beispiel bakterielle DNS (HACKER et al., 1998; AKBARI et al., 1999), Doppelstrang RNS aus Influenzaviren (CELLA et al., 1999) und LAM (Lipoarabinomannan) (KRUTZIG und MODLIN, 2004), können -6- Literaturübersicht eine Reaktion der DC in Form eines Reifungsschrittes bei Kontakt verursachen. Neben den exogenen Faktoren, also von außen auf den Körper einwirkenden Faktoren, können auch endogene körpereigene immunologische Mechanismen in ruhenden DC einen Ausreifungsschritt induzieren. So kann z.B. eine lokale Produktion von TNF-α oder IL-1 (CUMBERBATCH und KIMBER, 1995), eine Änderung des Gleichgewichts zwischen pro- und inflammatorischen Signalen oder die Aufnahme von Immunkomplexen über Fc-Rezeptoren ruhende DC dazu veranlassen, einen Reifungsschritt in Richtung maturer DC durchzuführen. Die zahlreichen Möglichkeiten, DC zu aktivieren, dienen alle der Rekrutierung immunkompetenter Zellen in inflammatorische Gebiete und somit der Steigerung der angeborenen Immunität. Nach der Aktivierung der DC kommt es zu zahlreichen morphologischen Veränderungen auf Seiten der DC. Durch die Neuorganisation des Zytoskeletts, die Modifikation der Adhäsionsstrukturen, den Verlust der Endozytoseund Phagozytoserezeptoren und die Heraufregulation von MHC-Molekülen bereitet sich die DC auf ihre Funktion als antigenpräsentierende Zelle vor (WINZLER et al., 1997). 2.1.3. Migration reifender DC in lymphatische Organe Nach der Aktivierung der DC kommt es zur Reifung und zur Migration der DC über die Lymphflüssigkeit zu den lymphatischen Organen. Bei der Reifung kommt es zu einer Desensibilisierung der DC gegenüber inflammatorischen Chemokinen (CCL3/MIP-1α, CCL4/MIP-1β und CCL5/RANTES) durch Herunterregulierung der entsprechenden Rezeptoren (SALLUSTO et al., 1998; SOZZANI et al., 1999). Durch diese Herunterregulierung können sich maturierende DC den lokalen Gradienten der inflammatorischen Chemokine entziehen. Parallel wird der Chemokinrezeptor CCR7 hochreguliert. Dieser Rezeptor ist für die Wanderung der DC sehr wichtig, da er die von lymphatischen Organen ausgehenden Chemokine CCL19 und CCL21 erkennt (STEINMAN, 2003b). Somit können die DC das inflammatorische Gewebe über den Lymphstrom entlang eines chemotaktischen Gradienten in Richtung der lymphatischen Organe verlassen. Nach Ankunft in den T- und B-Zell-Zonen -7- Literaturübersicht sekundärer lymphoider Organe können die DC selbst CCL19 und CCL21 sezernieren um zusätzlich für eine Verstärkung der chemotaktischen Signale zu sorgen (DIEU et al., 1998, SALLUSTO et al., 1998). Darüber hinaus werden durch die Chemokine CCL19, CCL18 und CCL22 zusätzlich naive T-Zellen und TGedächtnis-Zellen von den maturierenden DC angelockt. Die Abbildung 2.1.3.-a zeigt eine schematische Darstellung des Zusammenhanges zwischen Chemokinen und der DC-Reifung in Anlehnung an MCCOLL (2002) und LEBRE et al. (2005). -8- Literaturübersicht Ag CCR7 maturierende DC CCL2 /3/5 drainierendes Lymphgefäß CCR1/2/5/6 Blutgefäß CCL19/21 Immature DC naive TZellen mDC MHCII CD40 Immunantwort CD80/86 (Immunität, Toleranz) sekundäres Lymphorgan (Lymphknoten/Milz etc.) Chemokine Chemokinrezeptoren Oberflächenmoleküle Abb. 2.1.3.-a: Schematische Darstellung der Zusammenhänge zwischen Chemokinen und der DC-Reifung (MCCOLL, 2002; LEBRE et al., 2005). Immature DC gelangen über die Blutgefäße entlang des Chemokingradienten der inflammatorischen Chemokine CCL2/3/5 in die inflammatorischen Gewebe. Nach Ag-Aufnahme kommt es zur Expression des Chemokinrezeptors CCR7. Durch diesen Rezeptor ist es den nun reifenden (maturierenden) DC möglich, entlang des Chemokingradienten von CCL19 und CCL20 zu den sekundären Lymphorganen zu wandern. Dort kommt es zur Antigenpräsentation und zur Einleitung einer Immunantwort. -9- Literaturübersicht 2.1.4. Antigenprozessierung und –präsentation durch DC Die Migration der DC in die lymphatischen Organe dient der Präsentation der aufgenommenen Antigene an immunkompetente Zellen, um eine spezifische Immunantwort zu induzieren. In den lymphatischen Organen können die DC mit TZellen, B-Zellen und natürlichen Killerzellen (NK-Zellen) interagieren. Für die T-ZellAntigenpräsentation werden die von den DC aufgenommenen Pathogene durch intrazelluläre Enzyme in immunogene Peptide abgebaut und an MHC-Moleküle gebunden. Es existieren zwei verschiedene Sorten von MHC-Molekülen (MHCKlasse I-Moleküle und MHC-Klasse II-Moleküle), die bei der Beladung mit Peptiden aus jeweils unterschiedlichen Zellkompartimenten (dem Zytosol und dem vesikulären System) bedient werden (PAMER und CRESSWELL, 1998). Peptide, die an MHCKlasse I-Moleküle gebunden werden, interagieren mit dem T-cell receptor (TCR) von CD8+ zytotoxischen T-Zellen (CTL). Peptide aus dem vesikulären System werden von den DC an MHC-Klasse II-Molküle gebunden. Diese interagieren mit den TCR der CD4+ T-Helferzellen. Das alleinige Erkennen eines präsentierten Antigens durch einen TCR einer T-Zelle reicht jedoch noch nicht für eine adäquate Aktivierung der TZellen aus. Aus diesem Grund werden von reifen DC neben den MHC-Molekülen zusätzlich Korezeptoren auf ihrer Oberfläche exprimiert. Für die Modulation einer TZell Antwort wichtige Korezeptoren sind CD80, CD83, CD86 und CD40 (LATCHUMANAN et al., 2002; FLORIDO et al., 2004). Erst durch die Kombination der Verbindungen MHC-Moleküle zu TCRs und der Korezeptoren CD80/CD86 zu CD28 (T-Zellen) kommt es zur Bildung immunologischer Synapsen (LAMBRECHT, 2001). Zusätzlich bewirkt die Bindung zwischen CD40 zu CD40L (aktivierte T-Zellen) eine Steigerung der Expression von stabilen MHC-Molekülen, eine Hochregulation von CD86 (LAMBRECHT, 2001) und eine vermehrte Produktion von IL-12, IL-6 und TNF-α (LUTZ und SCHULER, 2002; GAD et al., 2003) mit der Folge einer zusätzlichen Aktivierung weiterer T-Zellen. Aber nicht nur T-Zellen können von DC aktiviert werden. Sie sind außerdem in der Lage, die B-Zell-Proliferation und Differenzierung zu beeinflussen (DUBOIS et al., 1999) sowie Zellen des angeborenen Immunsystems, wie zum Beispiel NK-Zellen (ZITVOGEL, 2002), - 10 - Literaturübersicht Eosinophile und Makrophagen über Chemokine, Zytokine oder direkten Kontakt zu aktivieren (PICCIOLI et al., 2002). 2.2. Rolle von Magnesium im Organismus Magnesium ist ein essentielles Mineral im Organismus und ist an einer Vielzahl von physiologischen Funktionen im Organismus beteiligt. Besonders für intrazelluläre, energieabhängige (metabolische) Reaktionen spielt Magnesium eine große Rolle, da es praktisch an allen ATP-abhängigen Reaktionen beteiligt ist (WOLF und CITTADINI, 2003). Wichtige Enzyme sind u.a. die Adenylatzyklase, ATPasen in Membranen (Substrat der Enzyme: ATP-Mg2+-Komplex) oder die alkalische Phosphatase, bei der Magnesium das Enzym bindet und aktiviert (WILLIAMS, 1993). Außerdem ist Magnesium beteiligt an der Aufrechterhaltung des elektrischen Potentials von Nervenzellen, der Muskelkontraktion (physiologischer Calcium-Antagonist) (BIESALSKI et und al., der Regulation 1999). von Schließlich Kaliumist und Calciumkanälen Magnesium bei der Nukleinsäurebiosynthese wichtig, da es zum einen der Stabilisierung ihrer tertiären Struktur, und zum anderen als Co-Faktor im Metabolismus der Nukleinsäuren dient (WOLF und CITTADINI, 2003). Im Großen und Ganzen ist Magnesium an über 300 enzymatischen Reaktionen als Co-Faktor beteiligt, so dass eine ständige Regulation des Magnesiumspiegels im Organismus gewährleistet sein muss. Im Körper eines Erwachsenen befindet sich ca. 20-28 g Magnesium wobei 60-65 % im Knochen gespeichert sind. Dort ist Magnesium ein wichtiger Bestandteil und übt in Form des Hydroxylapatits in Knochen und Zähnen Stützfunktion aus. Der Rest befindet sich hauptsächlich in der Muskulatur (27 %) und in anderen Organen (6-7 %) (SHILS, 1997). Im Plasma befindet sich nur etwa 1 % Magnesium. Ein Großteil des intrazellulären Magnesiums ist im Endoplasmatischen Retikulum, gefolgt von den Mitochondrien und im Zellkern lokalisiert (HEATON, 1993). Dabei liegt etwa 90 % in gebundener Form vor, wobei es hauptsächlich an Nukleinsäuren, ATP (als ChelatKomplex), negativ geladene Phospolipide und Proteine gebunden ist (WOLF und CITTADINI, 2003). Je höher die metabolische Aktivität einer Zelle desto höher ist - 11 - Literaturübersicht auch dessen intrazelluläre Magnesiumkonzentration, so dass gerade bei Skelett- und Herzmuskelzellen intrazelluläre Magnesiumkonzentrationen von 5 bis zu 20 mmol/l erreicht werden können (MCCARTHY und KUMAR, 1999). 2.2.1. Magnesium: Resorption und Ausscheidung Die freie extrazelluläre Magnesiumkonzentration wird mit Hilfe eines komplexen Regelsystems konstant gehalten. Die Resorption von Magnesium erfolgt über den Dünndarm (Jejunum und Ileum) parazellulär durch passive Diffusion (SCHWEIGEL und MARTENS, 2000) als auch transzellulär durch einen carriervermittelten Prozess (TRPM6-Kanal) (FINE et al., 1991; QUAMME, 1997; WÄRING, 2008). Durch diesen aktiven Transport ist es möglich, Magnesium entgegen ungünstigen Membranspannungsverhältnissen und Magnesiumgradienten in den interstitiellen Raum zu transportieren (FINE et al., 1991; QUAMME, 1997). Unter normalen Bedingungen liegt die Resorptionsrate zwischen 30-50 % des durch die Nahrung aufgenommenen Magnesiums. Bei einer geringen Magnesiumaufnahme kann die Resorptionsrate allerdings über den aktiven Transporter bis zu 70 % gesteigert werden, um die extrazelluläre Magnesiumkonzentration konstant zu halten (BRANNAN et al., 1976; WÄRING, 2008). Außerdem werden bei einem geringen Magnesium-Serumspiegel u.a. Calcitriol (aktive Form von Vit. D) und Parathormon (PTH) freigesetzt, so dass die Magnesiumaufnahme aus dem Dünndarm und der Transport in den Extrazellularraum erhöht werden (BIESALSKI et al., 1999). Die Ausscheidung von Magnesium erfolgt überwiegend über die Niere. Nach glomerulärer Filtration wird Magnesium jedoch zum größten Teil wieder rückresorbiert, so dass nur etwa 3-5 % der glomerulär filtrierten Magnesiummenge tatsächlich mit dem Endharn ausgeschieden werden (QUAMME, 1993; SCHAAFSMA, 1997). Als Hauptregulationsorgan ist die Niere in der Lage, über spezielle Sensoren Magnesiumkonzentration Veränderungen wahrzunehmen. in der Bei einem extrazellulären geringen freien Magnesium- Serumspiegel kommt es im Organismus zu einer vermehrten Bildung von PTH und folglich zur Erhöhung von Calcitriol mit der Folge einer Erhöhung der tubulären - 12 - Literaturübersicht Magnesium-Rückresorption und einer Hemmung der renalen Magnesium- Ausscheidung (BIESALSKIE, 2002; FLEET et al., 2001). Bei einem MagnesiumÜberschuss im Organismus wird vermehrt Calcitonin (Antagonist von PTH) synthetisiert, welches die renale Magnesium-Ausscheidung stimuliert und somit die extrazelluläre Magnesium-Konzentration senkt (QUAMME, 1993; SARIS et al., 2000; FLEET und CASHMAN, 2001). 2.2.2. Toxikologische Bedeutung von Magnesium Magnesium ist relativ untoxisch. Die empfohlene Recommended Daily Allowance (RDA) für Magnesium liegt bei Erwachsenen zwischen 310-420 mg (NATIONAL ACADEMY OF SCIENCES, 1999). Die physiologische MagnesiumKonzentration im Serum liegt zwischen 0,75-1 mmol/l (WEISINGER und BELLORINFONT, 1998). Da der Magnesiumhaushalt durch ein komplexes Regelsystem über Aufnahme und Abgabe konstant gehalten wird, ist bei gesunden Menschen eine Hypermagnesiämie in der Regel relativ selten, da zu viel aufgenommenes Magnesium über die Niere wieder ausgeschieden wird. Bei exzessiver Einnahme von Magnesiumsalzen oder magnesiumhaltiger Arzneimitteln (z.B. Antacida) kann es gerade bei Patienten mit einer gestörten Nierenfunktion zu Symptomen einer Hypermagnesiämie kommen. Leitsymptome sind vor allem zentralnervöse Störungen mit curareähnlichen Lähmungserscheinungen, Bradykardie, Hypotonus und Hypoventilation. Die ersten Symptome können ab einer Magnesium-Konzentration im Serum von 2,0 mmol/l auftreten. Lähmung, Herzstillstand und Koma treten bei Konzentrationen über 7 mmol/l auf (SWAMINATHAN, 2003). Die Hypomagnesiämie, also eine Unterversorgung mit Magnesium, kommt im Vergleich zur Hypermagnesiämie häufiger vor. Ursachen können zum einen eine Umverteilung des Magnesiums von der Extrazellulärflüssigkeit in die Zellen oder Knochen sein oder mangelhafte nutritive Aufnahme (AL-GHAMDI et al., 1994; FAWCETT et al., 1999). Weitere Ursachen sind u.a. gastro-intestinale (z.B. chronische Diarrhoe), renale (z.B. erhöhte Diurese, Nephropathien), Alkoholismus und medikamentöse Ursachen (z.B. - 13 - Diuretika, elektrolytfreie Infusionen) Literaturübersicht (SWAMINATHAN, 2003). Eine Hypomagnesiämie tritt häufig in Kombination mit anderen Elektrolytstörungen auf. So kann zusätzlich eine Hypokaliämie (RYAN, 1993), Hypophosphatämie oder Hypocalcämie (FATEMI et al., 1991) vorliegen. An Symptomen treten Schwindel, Ataxie, vermehrte Erregbarkeit, Krämpfe, Tetanie und schwerwiegende Organstörungen wie Herzarrhythmien auf (SWAMINATHAN, 2003). Außerdem wurde bei Patienten mit Osteoporose ein Magnesiummangel in den trabekulären Knochen festgestellt (SOJKA, 1995). 2.3. Magnesiumbasislegierungen Der Einsatz von Magnesiumbasislegierungen als resorbierbares metallisches Osteosynthesematerial und in der Weichteilchirurgie zeigte sich in den letzten Jahren als sehr viel versprechend (MEYER-LINDENBERG et al., 2003; SWITZER, 2005; WITTE et al., 2005). Gegenüber herkömmlich eingesetzten nichtresorbierbaren Implantaten aus Titan oder Stahl haben sie den Vorteil, dass sie sich mit der Zeit im Organismus auflösen. Dadurch kann eine zweite Operation zur Entfernung der Implantate und somit ein zweites Narkoserisiko umgangen werden (STAIGER et al., 2006). Auch im Vergleich zu anderen degradablen Implantatmaterialien wie zum Beispiel synthetische Polymere (resorbierbare Implantate an nicht tragenden Knochen) zeigen die Magnesiumbasislegierungen bei frei Resorptionszeit eine deutlich höhere Festigkeit (CLAES, 1992; OKUNO, 1999; KAESE, Magnesiumbasislegierungen 2002). die Somit guten kombinieren mechanischen einstellbarer SHIKINAMI und Implantate aus Eigenschaften der herkömmlichen Implantate mit den Vorteilen der resorbierbaren Materialien. Der Gedanke, Implantate aus Magnesium in der Osteosynthese einzusetzen, ist jedoch nicht neu. Bereits 1907 setzte LAMBOTTE eine Magnesium-Platte ein, um eine Unterschenkelfraktur zu versorgen. Da das Implantatmaterial aus reinem Magnesium jedoch unter starker Gasbildung zu schnell korrodierte, schlug der Versuch fehl (LAMBOTTE, 1932). Daraufhin wurden in den folgenden Jahrzehnten Implantate aus verschiedenen Korrosionsbeständigkeit sowie Magnesiumlegierungen die entwickelt, In-vivo-Eigenschaften - 14 - zu um die verbessern. Literaturübersicht VERBRUGGE (1934) und MCBRIDE (1938) setzten in den 30er Jahren Implantate aus Magnesium-Aluminium-Legierung zur Versorgung verschiedener Frakturen beim Menschen erfolgreich ein. Vorteilhaft für die Magnesiumlegierungen war eine komplette Auflösung im Organismus ohne Irritationen und toxische Nebenwirkungen. Allerdings stellte VERBRUGGE (1934) ebenfalls bei der Korrosion der Implantate eine Gasbildung fest, jedoch ohne Schädigung des Organismus. Die Vermutung von VERBRUGGE (1934), dass das entstandene Gas Wasserstoff sei, widerlegte MCBRIDE (1938) indem er die Gasblasen punktierte und analysierte. Er fand ein Gemisch aus 81 % Stickstoff, 7 % Wasserstoff, 6 % Sauerstoff und 5 % Kohlendioxid. TROITSKII und TSITRIN untersuchten 1944 die Auswirkung von MagnesiumCadmium-Legierungen als Implantatmaterial für verschiedene Frakturen. Im Bereich des Implantats wurden keine Entzündungsreaktionen festgestellt. Auch trat kein Anstieg der Magnesiumkonzentration im Serum auf. Die Korrosionsbeständigkeit variierte bei den Versuchen zwischen fünf Wochen und 12 Monaten (TROITSKII und TSITRIN, 1944). 1972 wurden erstmals Multikomponenten-Legierungen als Implantatmaterial eingesetzt, die neben Calcium oder Aluminium zusätzlich 4 wt% Seltene Erden (SE) und geringe Mengen an Silber, Mangan, Zirkonium oder Silizium enthielten (STROGANOV et al., 1972) mit dem Resultat einer verlangsamten Korrosion. Die Implantate in Form von Pins (Ø 3mm) konnten im Organismus bis zu fünf Monate nachgewiesen werden. Der Gedanke, Magnesiumlegierungen mit geringen Anteilen an Seltenen Erden (SE) als Implantatmaterial zu verwenden, wurde in neueren Arbeiten weiter untersucht (SWITZER, 2005; WITTE et al., 2005; WITTE et al., 2006a; ABELN, 2006). Die Tabelle 2 soll einen Überblick über die eingesetzten Magnesiumlegierungen geben. Tabelle 2: Übersicht der eingesetzten Magnesiumlegierungen (ABELN, 2006) Magnesium-Aluminium-Zink-Legierungen Magnesium-Seltene Erden-Legierungen AZ 31 (3 wt% Aluminium, 1 wt% Zink) LAE 442 (4 wt% Lithium, 4 wt% Aluminium, 2 wt% Seltene Erden (SE) AZ 91 (9 wt% Aluminium, 1 wt% Zink) WE 43 (4 wt% Yttrium, 3 wt% SE) - 15 - Literaturübersicht SWITZER (2005) untersuchte die Implantationseigenschaften der o.g. Magnesiumlegierungen im Vergleich zu herkömmlichen Implantatmaterialien wie PLA (Polylaktid) und Titan. Die Legierungen wurden als Magnesiumstifte intramedullär in die Femora von Meerschweinchen implantiert. Bei den Versuchen konnte gezeigt werden, dass bei der Korrosion der Implantate im Organismus ebenfalls Gas gebildet wurde, allerdings war die Gasbildung bei WE43 und LAE442 deutlich geringer als bei AZ31 und AZ91 (SWITZER, 2005). Wie auch bei MCBRIDE (1938) konnte bei der Untersuchung des gebildeten Gases kein Wasserstoff nachgewiesen werden (WITTE et al., 2005). Aus immunologischer Sicht untersuchten WITTE et al. (2006b), ob biodegradable Magnesiumlegierungen bei ihrer Degradation zu einer Aktivierung des Immunsystems führen. Hierfür wurden Zylinder der Magnesiumlegierung AZ91 in die rechten distalen Femurkondylen von Kaninchen implantiert. Als Kontrolle dienten autogene Knochenzylinder, die in die linken distalen Femurkondylen implantiert wurden. Drei bzw. sechs Monate nach Implantation konnte keine signifikant erhöhte Immunantwort festgestellt werden. Als Indikator für eine Immunantwort wurde die Anwesenheit von neutrophilen Granulozyten und CD3+ T-Zellen gemessen (WITTE et al., 2006b). ABELN (2006) führte prädiktive Allergietestungen mit den Legierungen AZ91, AZ31, WE43 und LAE442 (Tabelle 2) durch, um das allergene Potential der Magnesiumlegierungen zu ermitteln. Hierfür wurden die Testsubstanzen zunächst in gelöster Form und anschließend als Späne im Epikutantest bei Mäusen untersucht. Ein allergenes Potential konnte für die getesteten Legierungen nicht festgestellt werden. Als Kriterium für eine allergene Wirkung wurden Biopsien der Haut auf Spongiosis, Ödem, mononukleäres Infiltrat in der Dermis und Epidermis und perivaskuläres Infiltrat in der Dermis untersucht (ABELN, 2006). WITTE et al. (2006a) verglich das Korrosionsverhalten der Magnesiumlegierung AZ91 und LAE442 zwischen In-vitroKorrosion und In-vivo-Korrosion. Er stellte fest, dass die Korrosionsraten der Legierungen im In-vivo-Tiermodell vier mal niedriger waren als die im In-vitroVersuch. Außerdem zeigten LAE442 und AZ91 im Vergleich In-vitro- zu In-vivoUntersuchungen gegensätzliche Korrosionsraten. Im In-vivo-Tiermodell erwies sich - 16 - Literaturübersicht die Legierung LAE442 als korrosionsresistenter als die Legierung AZ91. Im In-vitroVersuch zeigte jedoch die Legierung AZ91 höhere Korrosionsresistenz als LAE442. 2.3.1. Magnesium-Calcium-Legierungen Mit dem Hintergrund eine absolute Bioverträglichkeit der Magnesiumbasislegierungen zu garantieren, wurden in den letzten Jahren zusätzlich Untersuchungen mit Magnesium-Calcium-Legierungen durchgeführt (BACH et al., 2005; KRAUSE et al., 2005, 2006; MEYER-LINDENBERG et al., 2007; VON DER HÖH, 2008; LI et al., 2008). Trotz Allergietestung (ABELN, 2006) scheinen Aluminium und Seltene Erden, die in den Magnesiumbasislegierungen AZ31, AZ91 bzw. WE43 und LAE442 verwendet wurden, aus toxikologischer Sicht als bedenklich (LI et al., 2008). Aluminium ist bekannt als neurotoxisch bei parenteraler Gabe und wird in Verbindung gebracht mit Alzheimischer Krankheit (EL-RAHMAN, 2003). Seltene Erden wie zum Beispiel Yttrium, Cer(ium) und Praseodymium haben bei parenteraler Gabe lebertoxische Wirkung (YUMIKO et al., 1997). Dagegen erscheint Calcium zur Herstellung binärer Magnesiumlegierungen als resorbierbares Osteosynthesematerial vielversprechend, da Calcium wie auch Magnesium ein wichtiger Bestandteil im Knochengewebe ist (LI et al., 2008). Außerdem erhofft man sich einen positiven Effekt für die Frakturheilung bei der Freisetzung von Mg2+ und Ca2+ während der Degradation, da Magnesium essentiell für die Aufnahme des Calciums in den Knochen ist (SERRE et al., 1998). Erste In-vitro-Versuche mit Magnesium-Calcium-Legierungen zeigten ausreichende mechanische Eigenschaften für den Einsatz als Knochenimplantate (BACH et al., 2005). KRAUSE et al. (2005) und MEYER-LINDENBERG et al. (2007) untersuchten eine Calcium-haltige Magnesiumlegierung (MgCa0.8 mit 0,8 wt% Ca) und zwei Magnesiumlegierungen mit Beimengungen von Seltenen Erden (LAE442 und WE43) im Tiermodell. Dafür wurden Magnesiumstifte der drei Legierungen intramedullär in die Tibiamarkhöhle von Kaninchen implantiert. MgCa0.8 zeigte eine gute Biokompatibilität bei moderater Degradationsrate in den ersten drei Monaten nach - 17 - Literaturübersicht Implantation (KRAUSE et al., 2005). Im direkten Vergleich zu WE43 und LAE442 zeigte MgCa0.8 geringere Festigkeitswerte (KRAUSE et al., 2005). In einer Pilotstudie Degradationsverhalten Kaninchenmodell. Für untersuchte verschiedener den Versuch VON (2008) das Magnesium-Calcium-Legierungen im wurden DER aus vier HÖH Magnesium-Calcium- Legierungen Implantate mit drei verschiedenen Oberflächen (glatt, rau und gestrahlt) hergestellt. Die Tabelle 3 soll einen Überblick über die verwendeten MagnesiumCalcium-Legierungen geben. Tabelle 3: Übersicht verwendeter Magnesium-Calcium-Legierungen (VON DER HÖH, 2008) Magnesium-Calcium-Legierung wt% Calcium MgCa0.4 0,4 MgCa0.8 0,8 MgCa1.2 1,2 MgCa2.0 2,0 Zur Herstellung einer rauen Implantatoberfläche wurden die Implantate durch einen nachgelagerten Schleifprozess bearbeitet. Die gestrahlten Implantatoberflächen wurden durch Partikelbestrahlung von glatten Implantaten erzeugt (Partikeldurchmesser 300-400 µm, Strahlzeit 30 s). Anschließend wurden die Implantate für sechs Wochen in die medialen Femurkondylen beider Hintergliedmaßen eingesetzt. Unabhängig von den Calciumkonzentrationen in den Implantaten oder der Oberflächenstruktur zeigten die Implantate eine gute Verträglichkeit. Untersuchungen auf das Degradationsverhalten zeigten, dass Implantate mit glatter Oberfläche stabiler degradierten als Implantate mit rauer oder gestrahlter Oberfläche. Die Degradation in Bezug auf die Calciumkonzentration war bei MgCa1.2 am schwächsten, bei MgCa0.4 am stärksten (VON DER HÖH, 2008). Diese Ergebnisse stehen im Gegensatz zu In-vitro-Degradationsversuchen von DENKENA et al. (2005, 2006). Für den Degradationsversuch wurden die Magnesium-Calcium-Legierungen MgCa0.4, MgCa0.8 und MgCa2.0 für 72 Stunden - 18 - Literaturübersicht einer 5%igen NaCl2-Lösung ausgesetzt. Die Auswertung zeigte, dass bis zu einem Calciumanteil von 0,8 wt% Calcium die Legierungen gleichmäßig korrodierten. Bei den Legierungen mit höheren wt% Calcium verlief die Degradation stärker und inhomogener (DENKENA et al., 2005; DENKENA et al., 2006). LI et al. (2008) führten Zellkulturversuche mit verschiedenen MagnesiumCalcium-Legierungen durch. Unter anderem untersuchten sie den Einfluss von MgCa1 (1 wt% Calcium) auf die Vitalität und Morphologie von murinen Fibroblasten (L929- Zellen). Dafür wurden die L929-Zellen in Dulbecco`s Modified Eagle`s Medium (DMEM) kultiviert. Ab Tag 1 der Kultivierung wurden die L929-Zellen mit unterschiedlichen Verdünnungsstufen (0 %, 50 %, 90 %) des Degradationsmediums der Legierung MgCa1 inkubiert. Am Tag 2, 4 und 7 wurde die Vitalität der Zellen mittels MTT bestimmt. Die Auswertung zeigte, dass MgCa1 keinen zytotoxischen Effekt auf L929-Zellen hat. Darüber hinaus schien die Vitalität der in unverdünnten Degradationsmedium kultivierten L929-Zellen besser zu sein als die Vitalität von L929-Zellen in der Kontrolle (LI et al., 2008). 2.4. Capture-ELISA Beim Capture-ELISA beschichtet man eine Mikrotiterplatte mit Antikörper gegen das nachzuweisende Antigen. Durch Zugabe der zu untersuchenden Probe wird das Antigen von den Antikörpern gebunden (Antigen-Antikörper-Komplex). Durch Waschen kann anschließend nicht gebundenes Material entfernt werden. Anschließend wird durch Zugabe eines zweiten Antikörpers, an dem ein Enzym gebunden ist, eine Farbreaktion ausgelöst. Die entstehende Farbreaktion kann mit Hilfe eines Photometers gemessen werden. Sie ist proportional zur Enzymaktivität und somit zu der vorhandenen Menge an Antigen (BROCK et al., 2001; LUTTMANN et al., 2006). 2.5. FACS-Analyse Die FACS-Analyse (Fluorescence activated cell sorting) beruht auf dem Prinzip der Durchflusszytometrie. Dabei werden Zellen über eine Kapillare an einen Laserstrahl - 19 - Literaturübersicht vorbei gesaugt. An den Zellen kommt es zu einer Lichtstreuung, die von einem Photodetektor gemessen wird. Für die Beurteilung der Lichtstreuung dienen zwei Parameter. Die Zellgröße wird durch die Streuung des Lichtes in Richtung des Laserstrahl bestimmt (Vorwärtsstreulicht, forwardscatter, FSC). Die Größe und Struktur des Zellkerns (Granularität) wird durch das in einem 90° Winkel an inneren Zellstrukturen reflektierte Licht bestimmt (Seitwärtsstreulicht, sidescatter, SSC). Neben diesen Parametern können die Zellen zusätzlich mit einem Fluoreszenzfarbstoff markiert werden. Dabei kommt es beim Passieren des Lasers zur Anregung des Farbstoffes, der daraufhin Licht einer bestimmten Wellenlänge emittiert. Man unterscheidet bei der Markierung der Zellen mit Fluoreszenzfarbstoff zwischen der Oberflächenmarkierung und der intrazellulären Markierung. Außerdem wird zwischen einer direkten Markierung (Antikörper mit Fluoreszenzfarbstoff) und einer indirekten Markierung (unmarkierter Primärantikörper und markierter Sekundärantikörper) unterschieden (JANEWAY et al., 2002; LUTTMANN et al., 2006). 2.5.1. Annexin V-FITC und PI In apoptotischen Zellen wird das Membranphospholipid Phosphatidylserin (PS) durch Veränderung der Zytoplasmamembran nach außen gekehrt. Der markierte Antikörper Annexin V-Fluorescein-Isothiocyanat (Annexin V-FITC) bindet mit hoher Affinität an PS. Der Farbstoff Propidiumiodid (PI) dient der intrazellulären Markierung von Nukleinsäuren. In lebenden Zellen mit intakter Zytoplasmamembran kann PI nicht in das Zytoplasma gelangen. Aufgrund dieser Eigenschaften lassen sich die Zellen bezüglich ihrer Vitalität einteilen in vitale Zellen (Annexin V-FITC negativ; PI negativ), Zellen in frühem Apoptose-Stadium (Annexin V-FITC positiv, PI negativ) und sterbende oder tote Zellen (Annexin V-FITC positiv, PI positiv) (BD PharmingenTM Technical Data Sheet, Product Information). - 20 - Literaturübersicht 2.6. XTT-Proliferationstest Zum Nachweis zytotoxischer Effekte kann die metabolische Aktivität einer Zelle Aufschluss über ihre Vitalität geben. Mit Hilfe des XTT-Proliferationstest kann diese metabolische Aktivität vitaler Zelle gemessen werden. Unter Oxidation von NAD(P)H +H+ zu NAD(P)+ wird das schwach gefärbte Tetrazoliumsalz XTT (Natrium3`-[1-(phenylaminocarbonyl)-3,4-tetrazolium]-bis(4-methoxy-6-nitro)benzolsulfonsäure-hydrat) vorwiegend durch mitochondriale Dehydrogenasen intakter Zellen zu einem wasserlöslichen, stark gefärbten Formazan umgewandelt. Diese Farbstoffbildung wird gemessen, wobei die entstehende Farbintensität mit der Zahl lebender Zellen korreliert (MOSMANN, 1983). 2.7. MLR (Mixed Leukocyte Reaction) Mit Hilfe der heterologen MLR (Mixed Leukocyte Reaction) kann die Fähigkeit der unterschiedlich behandelten DC, T-Lymphozyten zur Proliferation anzuregen, untersucht werden. Dafür werden T-Lymphozyten einer NMRI-Maus mit DC einer BALB/c-Maus zusammen gebracht. Bei aktivierten DC werden die MHC-Moleküle der DC von den T-Lymphozyten als fremd erkannt. Dadurch kommt es zu einer Proliferation, also Zellteilung, der T-Lymphozyten. Die Proliferationsrate kann mit Hilfe des Farbstoffs CFSE (Carboxyfluorescein diacetate succinimidylester), mit dem die T-Lymphozyten markiert werden, sichtbar gemacht werden. Nach Diffusion in die Zelle wird die Acetatgruppe des CFSE von intrazellulären Esterasen gespalten, so dass ein fluoreszierender Farbstoff gebildet wird, der die Zytoplasmamembran nicht mehr passieren kann. Bei einer Zellteilung der T-Lymphozyten wird der Farbstoff zur Hälfte an die Tochterzelle weitergegeben und damit die Fluoreszensintensität auf die Hälfte reduziert. Mit Hilfe der FACS-Analyse kann die Fluoreszensintensität und somit die Proliferationsrate der T-Lymphozyten ausgewertet werden (Molecular Probes, Invitrogen Product Information). - 21 - Material und Methoden 3. Material und Methoden 3.1. Material 3.1.1. Geräte HERAcell®150, Heraeus, Hanau, D Brutschrank Binder, Tuttlingen, D Kühlzentrifuge Zentrifuge 5804 R, Eppendorf, Hamburg, D Zentrifuge 5403, Eppendorf, Hamburg, D Mikroskop Axiovert 25, Zeiss, Oberkochen, D Neubauer-Zählkammer VWR, Hannover, D Mikroplattenleser MRX Microplate Reader Dynatech, Denkendorf, D Ultrospec®2000, Pharmacia Biotech, Photometer Cambridge, UK Heraeus Lamin Air® TL 2448, Heraeus, Sterilbank Hanau, D Pipettierhilfe Accu-jet®pro, Brand, Wertheim, D Feinwaage ALS 120-4, Kern, Balingen, D Vortexer Heidolph Reax top, Heidolph, Schwabach, D Janke & Kunkel, IKA® Labortechnik, Staufen, D Wasserbad Büchi, Flawil, CH GFL®, Burgwedel, D Durchflusszytometer Coulter Epics XL, Beckman Coulter, FLA, USA - 22 - Material und Methoden 3.1.2. Verbrauchsmaterial Polypropylen-Röhrchen (15 ml/50 ml) Greiner, Frickenhausen, D Reaktionsgefäße (1,5/ 2 ml) Eppendorf, Hamburg, D Greiner, Frickenhausen, D Sarstedt, Nümbrecht, D Petrischalen (100 mm x 20 mm) tissue culture dish cell+, Sarstedt, D 6-,12-,24,-96-well-plates Greiner, Frickenhausen, D Pipetten Eppendorf, Hamburg, D (2,5/ 20/ 100/ 200/ 1000/ 2500 µl) LABMATETM, Abimed, Langenfeld, D Pipettenspitzen Eppendorf Tips, Eppendorf, Hamburg, (0,1-10/ 2-200/ 500-1000/ 500-2500 µl) D Pipettenspitzen (2-200/ 500-1000 µl) Greiner, Frickenhausen, D Plastikpipetten (10/ 25 ml) Greiner, Frickenhausen, D Zellschaber (24 cm) TPP, Schweiz Sterilfilter Minisart, Sartorius, Göttingen, D 96-well-plates, round-bottom Greiner, Frickenhausen, D 6-well-plates mit Einsätzen Greiner, Frickenhausen, D (8.0 µm Porengröße) Einmalspritzen, 2/10 ml Omnifix, Braun, Melsungen, D Einmal-Injektions-Kanüle (Gr.18) Braun, Melsungen, D Einmal-Injektions-Kanüle (19 G) Sterican®, Braun, Melsungen, D Einmal-Injektions-Kanüle, (Gr. 20) Omnifix, Braun, Melsungen, D Skalpellklinge (22) Bayha, Tuttlingen, D 3.1.3. Reagenzien RPMI-1640-Medium Biochrom, Berlin, D A. bidest Sigma-Aldrich Chemie, Steinheim, D Streptomycin Invitrogen GmbH, Karlsruhe, D Penicillin Invitrogen GmbH, Karlsruhe, D Fetales Kälberserum (FKS) PAA, Pasching, A - 23 - Material und Methoden Rinderserum Albumin (BSA) Sigma-Aldrich Chemie, Steinheim, D β-Mercaptoethanol Sigma-Aldrich Chemie, Steinheim, D Ethanol Roth, Karlsruhe, D GM-CSF R&D-Systems, Wiesbaden, D Polyvidon-Iod-Lösung Braunol, Braun, Melsungen, D Lipopolysaccharid (LPS, E. coli 0172:B8) Sigma-Aldrich Chemie, Steinheim, D Trypsin Sigma-Aldrich Chemie, Steinheim, D Trypan-Blau Sigma-Aldrich Chemie, Steinheim, D Tween20 Sigma-Aldrich Chemie, Steinheim, D Schwefelsäure 2N Merck, Darmstadt, D 3,3`,5,5-Tetramethylbenzidine (TMB) Liquid Sigma-Aldrich Chemie, Steinheim, D Dimethylsulfoxid (DMSO) Sigma-Aldrich Chemie, Steinheim, D Etoposide (VP-16) Merck, Darmstadt, D Blei(II)chlorid (MW = 278,10) Merck, Darmstadt, D Magnesium(II)chlorid Merck, Darmstadt, D Calcium(II)chlorid Merck, Darmstadt, D 3.1.4. Magnesiumbasislegierungen Mg(pur) (reines Magnesium) Institut für Werkstoffkunde, Garbsen, D MgCa0.6 (0,6 wt% Calcium) Institut für Werkstoffkunde, Garbsen, D MgCa0.8 (0,8 wt% Calcium) Institut für Werkstoffkunde, Garbsen, D MgCa1.0 (1,0 wt% Calcium) Institut für Werkstoffkunde, Garbsen, D MgCa1.2 (1,2 wt% Calcium) Institut für Werkstoffkunde, Garbsen, D - 24 - Material und Methoden 3.1.5. Puffer und Lösungen Zellkultur: DC-Medium: FKS 10% Penicillin/Streptomycin 1% Β-Mercaptoethanol 50 µM Das Medium wurde in RPMI-1640-Medium hergestellt. PBS-Puffer NaCl 137 nM KCl 2,7 mM Na2PO4 7,5 mM KH2PO4 1,5 mM Der Puffer wurde mit A. bidest. hergestellt; pH 7,4. Hämolyse-Puffer NH4Cl 0,15 M Na2-EDTA 0,13 mM Na HCO3 10 mM Der Puffer wurde mit A. bidest. hergestellt; pH 7,3. Erythrozyten-Lysepuffer NH4Cl 0,15 M EDTA 0,34 mM KHCO3 10 mM Der Puffer wurde mit A. bidest. hergestellt. - 25 - Material und Methoden FACS-Analyse: PBS/BSA-Waschpuffer BSA 1% Die Lösung wurde in PBS hergestellt; pH 7,4. Binding-Puffer (Annexin V-FITC) Herstellung des Puffers aus dem Annexin V-FITC Detection Kit I (BD Biosciences Pharmingen). Die Stammlösung wurde um den Faktor 10 mit PBS verdünnt. Elisa: Waschpuffer Tween 20 0,05% Die Lösung wurde in PBS hergestellt; pH 7,4. 10 x TBE Tris-HCl (pH 8,0) 0,9 M Borsäure 0,9 M EDTA (pH 8,0) 20 mM Die Lösung wurde bei Bedarf um den Faktor 10 mit H2O verdünnt. Reagent Diluent (ELISA) BSA 1% Die Lösung wurde in PBS hergestellt; pH 7,4. 3.1.6. Kits DuoSet® Mouse TNF-α ELISA R&D Systems, Minneapolis, MN, USA Annexin V-FITC Apoptosis Detection Kit I BD Biosciences Pharmingen™, Hamburg, D XTT Cell proliferation-kit Promega, USA Testkit Nanocolor Härte 20 Machery-Nagel, Düren, D - 26 - Material und Methoden 3.1.7. Antikörper FITC anti-mouse CD86 BD Biosciences Pharmingen™, Hamburg, D PE anti-mouse CD 11c BD Biosciences Pharmingen™, Hamburg, D Streptavidin-Peridin Chlorophyll-a Protein BD Biosciences Pharmingen™, (SAv-Per CP) Conjugate Hamburg, D CellTrace™ CFSE Invitrogen™, Oregon, USA 3.1.8. Mäuse Für die DC-Gewinnung wurden weibliche Mäuse des Stammes BALB/c, sowie für die MLR (Mixed Leukocyte Reaktion) weibliche Mäuse des Stammes NMRI verwendet. Die Tiere waren acht Wochen alt und wurden von Charles River (Sulzfeld, D) bezogen. 3.1.9. Software Expo 32 ADC XL 4 Color Beckman Coulter, FLA, USA Expo 32 ADC Analysis Beckman Coulter, FLA, USA Microsoft® Office 2003-Edition Microsoft®, Washingten, WA, USA GraphPad Prism®5 GraphPad Software, CA, USA - 27 - Material und Methoden 3.2. Methoden 3.2.1. Generierung von DC Für die Generierung von DC wurden nach LUTZ et al. (1999) aus dem Knochenmark von BALB/c-Mäusen DC-Vorläuferzellen gewonnen und im Brutschrank für 10 Tage bei 37 °C und 5 % CO2 kultiviert. Dazu wurde die BALB/cMaus durch zervikale Dislokation getötet und der Femur herauspräpariert. Mit Hilfe einer Skalpellklinge wurde der Femur von umliegender Muskulatur befreit und anschließend in 70 %-igem Ethanol 2-5 Minuten desinfiziert und in sterilem PBS gewaschen. Für die Entnahme der Stammzellen aus der Markhöhle wurden die Femurenden mit der Skalpellklinge abgetrennt. Mit Hilfe von Kanüle und Spritze konnte anschließend das Knochenmark mit 5 ml sterilem PBS aus dem Femur herausgespült werden. Dabei wurde die Zellsuspension in einem Zentrifugenröhrchen aufgefangen und die Zellen durch Pipettieren vereinzelt. Anschließend wurde die Zellsuspension zentrifugiert (290 g, 4 °C, 10 Minuten). Danach wurde der Überstand verworfen und das Zellpellett mit 5 ml RPMI-1640Medium resuspendiert. Für die Bestimmung der gewonnenen Zellzahl wurden zu einem Aliquot der Zellsuspension Erythrozyten-Lysepuffer gegeben (Verdünnung 1:2). Nach ca. 5 Minuten Inkubation wurden die Zellen in Trypanblau (Verdünnung 1:3) in der Neubauer-Zählkammer ausgezählt. Für einen Ansatz wurden 2x106 Zellen benötigt. Diese wurden in 10 Wells einer 12well-plate aufgeteilt, sodass sich pro Well 2x105 DC-Vorläuferzellen in 1 ml DCMedium befanden. Anschließend wurden die Stammzellen mit 20 ng/ml granulozytemacrophage colony-stimulating factor (GM-CSF) versetzt (INABA et al., 1992), um myeloide DC zu generieren. Nach LUTZ et al. (1999) entstehen dabei adhärente Makrophagen (MØ), nicht-adhärente reife DC (MHCIIhigh) und unreife DC (MHCIIlow). Es folgte eine zweitägige Inkubation im Brutschrank bei 37 °C und 5 % CO2. Am 3. Tag der Kultivierung erfolgte eine Zugabe von 1 ml DC-Medium pro Well und 20 ng/ml GM-CSF. An den Tagen 6, 8 und 10 wurde jeweils 1 ml Medium pro Well entnommen, das Zellpellett durch Zentrifugation gewonnen und nach Resuspension mit frischem Medium und GM-CSF zur Kultur zurückgegeben. - 28 - Material und Methoden 3.2.2. Gewinnung muriner T-Zellen Die Proliferationsrate der T-Zellen als Reaktion auf die behandelten DC wird mit Hilfe der Mixed Leukocyte Reaction (MLR) bestimmt. Hierbei handelt es sich um eine heterologe Reaktion, bei der T-Zellen einer NMRI-Maus mit DC einer BALB/c-Maus zusammengebracht werden. Die benötigten T-Zellen wurden nach GUNZER et al. (2001) aus der Milz von NMRIMäusen gewonnen. Dazu wurde die NMRI-Maus durch zervikale Dislokation getötet und die Milz herauspräpariert. Anschließend wurden die beiden Enden der Milzkapsel eröffnet und mit Hilfe einer Kanüle und einer Spritze die Milzpulpa mit 10 ml sterilem PBS ausgespült. Die Zellsuspension wurde in einem Zentrifugenröhrchen aufgefangen und zentrifugiert (1000 g, 4 °C, 10 Minuten). Der Überstand wurde verworfen und das Zellpellet mit 5 ml Hämolysepuffer versetzt, um die vorhandenen Erythrozyten zu lysieren. Nach 5 Minuten wurde durch Zugabe von 45 ml sterilem PBS der Hämolysepuffer neutralisiert und somit der Vorgang gestoppt. Nach erneuter Zentrifugation (1000 g, 4 °C, 10 Minuten) wurde das Pellet in 10 ml RPMI1640-Medium (+ 10 % FKS + 1 % Penicillin/Streptomycin) resuspendiert. Anschließend wurde die Zellsuspension in einer cell+-Petrischale im Brutschrank (37 °C, 5 % CO2) für 2 Stunden inkubiert. Nach der Inkubation wurden die nicht adhärenten Zellen vorsichtig mit einer Pipette gewonnen. Somit wurde eine TZellpopulation mit einer Reinheit von 60-70 % gewonnen. Anschließend wurden die T-Zellen nach erneuter Zentrifugation in 5 ml sterilem PBS resuspendiert und in Trypanblau, mit Hilfe der Neubauer-Zählkammer, gezählt. 3.2.3. Herstellung der Degradationsmedien Für die Herstellung der Degradationsmedien wurden zuerst die Legierungen im Autoklaven sterilisiert. Anschließend wurden für jede Legierung 10 ml DC-Medium in ein 50 ml Polypropylen-Röhrchen vorgelegt und ein Legierungsplättchen (Ø 8 mm, Höhe 2mm, Oberfläche 150,8 mm2) dazu gegeben. Für eine gleichmäßige Degradation wurden die DC-Medien für 72 h im Wasserbad (37 °C) unter ständiger - 29 - Material und Methoden Bewegung mit den Legierungen inkubiert. Nach der 72-stündigen Inkubation wurden die einzelnen Degradationsmedien mit Hilfe von Spritze und Sterilfilter von ungelösten Bestandteilen befreit. 3.2.3.1. Bestimmung der Magnesium- und Calciumkonzentration in den Degradationsmedien Die Magnesium- und Calciumkonzentration in den Degradationsmedien wurde mit Hilfe des Testkits Nanocolor Härte 20 (Machery-Nagel, Düren, D) mittels eines photometrischen Verfahrens bestimmt. Der Test beruht auf der Messung der Gesamthärte des Mediums, welche vor allem durch Magnesium- und Calcium-, aber auch geringfügig durch Barium- und Strontiumionen bestimmt wird. Dazu wird die Farbintensität von Phtaleinpurpur mittels Photometer gemessen, welche von der Gesamthärte des umgebenden Mediums abhängt. Anschließend werden die Calciumionen durch Zugabe eines speziellen Chelators maskiert, sodass bei wiederholter Messung die Härte nur noch vom Magnesiumgehalt abhängt. Der Calciumgehalt kann abschließend aus den Messwerten der Gesamthärte und des Magnesiumgehaltes errechnet werden. Für die Bestimmung der Magnesium- und Calciumkonzentration wurden zunächst jeweils fünf Kalibrationsstandards mit den Konzentrationen 0,2; 0,4; 0,6; 0,8 und 1,0 mmol/l angesetzt. Außerdem wurden zur Überprüfung von verschiedenen Magnesium/Calcium-Kombinationen 9 Kalibrationsstandards von Lösungen mit 0,5 bis 0,1 mmol/l Magnesium/Calcium angesetzt. Die Tabelle 4 soll einen Überblick über die Kalibrationsstandards geben. - 30 - Material und Methoden Tabelle 4: Überblick Kalibrationsstandards im DC-Medium MgCl2-Konz. [mmol/l] CaCl2-Konz. [mmol/l] MgCl2-/CaCl2-Konz. [mmol/l] 0.2 0.2 0.5/0.5 0.4 0.4 0.4/0.4 0.6 0.6 0.3/0.3 0.8 0.8 0.2/0.2 1.0 1.0 0.1/0.1 0.5/0.4 0.4/0.5 0.3/0.2 0.2/0.3 Danach wurden die Proben der Degradationsmedien vorbereitet. Dafür wurden zum einen unverdünnte Aliquots der Degradationsmedien aus Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 entnommen und zum anderen Aliquots mit einer Verdünnung 1:10 hergestellt. Anschließend wurde aus dem Testkit Nanocolor Härte 20 die Phtaleinpurpurlösung angesetzt und pro Probe 180 µl der Phtaleinpurpurlösung in die Wells einer 96-Well Platte vorgelegt. Danach wurden von jeder Probe 6 µl Lösung hinzu gegeben und gründlich durch auf und ab pipettieren gemischt. Anschließend wurde die Gesamthärte photometrisch (570 nm) gemessen. Nach der Bestimmung der Gesamthärte wurde die 96-Well-Platte aus dem Photometer entnommen und in jedes Probenwell 6 µl des Calciumchelators aus dem Testkit zugegeben. Danach wurden die Proben erneut bei 570 nm gemessen. Die daraus resultierten optischen Dichten werden aufgrund der vorherigen Maskierung der Calciumionen hauptsächlich durch Magnesium bestimmt. Für die Errechnung der Magnesiumkonzentration wurde aus den optischen Dichten der Standards eine Kalibrationsfunktion errechnet. Diese wurde anschließend zur Rückrechnung der an den Proben gemessenen Konzentrationen aus den gemessenen optischen Dichten benutzt. - 31 - Material und Methoden 2 1,8 Optische Dichte 1,6 1,4 y = 0,4259x + 1,3086 R2 = 0,9753 1,2 y = 0,461x + 1,296 R2 = 0,9666 1 0,8 0,6 0,4 0,2 0 0 0,2 Abb. 3.2.2.2.-a: 0,4 0,6 0,8 1 1,2 Salzkonzentration Errechnete Kalibrationsfunktion aus den an den Standards gemessenen optischen Dichten der Gesamthärtemessung und der Magnesiummessung. Y= optische Dichte, 2 X= Salzkonzentration, R = Bestimmtheitsmaß (>0,95). Für die Errechnung der Calciumkonzentration wurden für die Standards und jede Probe die bei der Magnesiummessung erfassten, optischen Dichten von den optischen Dichten der Gesamthärtemessung subtrahiert. Aus diesem Differenzwert der Standards konnte mittels linearer Regression eine Kalibrationsfunktion errechnet und mit dieser die Calciumkonzentration in den Proben errechnet werden. Die Magnesium- und Calciumkonzentrationsmessungen wurden jeweils achtmal durchgeführt. 3.2.4. In-vitro-Untersuchungen mit Magnesium- und Calcium-Salzen Als Grundlage für die Versuche mit den Magnesium- und Calciumsalzen dienten Degradationsversuche mit Magnesiumbasislegierungen (Kapitel 3.2.3.), bei denen im Kulturmedium Magnesiumkonzentrationen zwischen 0,2 mmol/l und 10 mmol/l gemessen wurden. Durch die Zugabe von Magnesium- und Calciumsalzen in unterschiedlichen Konzentrationen zum Kulturmedium der DC sollte ein möglicher Einfluss auf die Zellfunktion von murinen DC untersucht werden. Zuerst wurde der Einfluss von Magnesium- und Calcium-Salzen auf die unterschiedlichen Phasen der DC-Maturation untersucht. Im Mittelpunkt standen zum einen die Vitalität der DC und die Expression diverser Oberflächenmoleküle untersucht in der Durchflusszytometrie, - 32 - Material und Methoden sowie das Auswanderverhalten der DC in einem Migrationsassay. Mit Hilfe der Mixed Leukocyte Reaction (MLR) konnte die Fähigkeit der DC, naive T-Zellen zur Proliferation anzuregen, untersucht werden. Versuche mit weiteren DC-Kulturen bestanden in der Gewinnung von Kulturüberständen zur Detektion des Zytokins TNF-α als Reaktion auf die Magnesium- und Calciumsalze. 3.2.4.1. Inkubation der DC mit MgCl2 und CaCl2 Für die Bestimmung der Vitalität, der TNF-α-Sekretion, der Migrationsbereitschaft, der Expression von CD11c/CD86 und der MLR wurden die DC pro Versuchsansatz ab dem 3. Tag der Kultivierung in jeweils fünf Gruppen für die Magnesiumchlorid- und Calciumchlorid-Behandlung unterteilt. Tabelle 5 gibt einen Überblick über die Kultivierung der DC-Vorläuferzellen und deren Behandlung. - 33 - Material und Methoden Tabelle 5: Übersicht zur Kultivierung der DC-Vorläuferzellen mit MgCl2 und CaCl2 einzelner Versuche Tag Inkubation mit CaCl2 der Inkubation mit MgCl2 Kultivierung Tag 0 Aussäen von 2x105 DC-Vorläuferzellen/ml in 10 Wells einer 12well-plate + GM-CSF Tag 3 Zugabe von 1 ml DC-Medium, Zugabe von 1 ml DC-Medium, GM-CSF und GM-CSF und 2+ Einstellung der Mg -Konz. im Einstellung der Ca2+-Konz. im Kulturmedium. Kulturmedium. Gruppe 1: 0,1 mmol/l MgCl2 Gruppe 6: 0,1 mmol/l CaCl2 Gruppe 2: 1,0 mmol/l MgCl2 Gruppe 7: 1,0 mmol/l CaCl2 Gruppe 3: 10,0 mmol/l MgCl2 Gruppe 8: 10,0 mmol/l CaCl2 Gruppe 4+5: unbehandelte Gruppe 9+10: unbehandelte Kontrolle Tag 6 Kontrolle Mediumwechsel, GMCSF und Mediumwechsel, GM-CSF und MgCl2. CaCl2. Tag 8 Behandlung wie Tag 6 Behandlung wie Tag 6 Tag 9 Je nach Versuchsdurchführung Behandlung der Gruppe 5 und 10 als Positiv-Kontrolle. 1. LPS-Stimulation (ELISA, CD11c/CD86) 2. Etoposide (Vitalitätstest) 3. Bleichlorid (Migrationsassay) Tag 10 Versuchsdurchführung: Zellzahlbestimmung der DC und der MØ Vitalitätstest, Migrationsassay, Ansetzen der MLR, Aufbewahrung der Kulturüberstände für Zytokinbestimmung Die Anzahl der durchgeführten Versuche sind im Anhang (Kapitel 9.) dargestellt. - 34 - Material und Methoden Für die Versuche mit MgCl2 wurden die Kulturmedien der Gruppen 1-3 auf unterschiedliche Mg2+-Konzentrationen eingestellt. In der Gruppe 1 enthielt das Kulturmedium 0,1 mmol/l MgCl2, das Kulturmedium der Gruppe 2 enthielt 1 mmol/l MgCl2 und das Kulturmedium der Gruppe 3 enthielt 10 mmol/l MgCl2. Für die Versuche mit CaCl2 wurden die Ca2+-Konzentrationen der Kulturmedien der Gruppen 6-8 eingestellt. In der Gruppe 6 enthielt das Kulturmedium 0,1 mmol/l CaCl2, das Kulturmedium der Gruppe 7 enthielt 1 mmol/l CaCl2 und das Kulturmedium der Gruppe 8 enthielt 10 mmol/l CaCl2. Die Gruppen 4 und 9 bildeten die unbehandelten Kontrollen, d.h. DC, die in unbehandeltem DC-Medium kultiviert wurden. Die DC der Gruppe 5 und 10 wurden als Positiv-Kontrollen verwendet. Dazu wurden die DC der Gruppe 5 und 10 ab Tag 9 der Kultivierung je nach Versuchsdurchführung unterschiedlich behandelt. In Anlehnung an HÜNERMANN (2006) wurde als PositivKontrolle für die TNF-α-Sekretion und die Expression von CD11c/CD86 dendritische Zellen mit 1 µg/ml LPS stimuliert, da LPS einen positiven Effekt auf die TNF-αSekretion hat und eine Steigerung der CD11c/CD86 positiven DC bewirkt. Als Positiv-Kontrolle für die Migrationsbereitschaft wurden die DC, ebenfalls in Anlehnung an HÜNERMANN (2006), mit 100 µmol/l Bleichlorid inkubiert, da Bleichlorid eine gesteigerte Migration bei DC bewirkt. Als Positiv-Kontrolle für die Bestimmung der Vitalität wurden die DC mit Etoposide [20 µg/ml] (Apoptose-inducer) inkubiert. Am Tag 10 der Kultivierung erfolgte die Durchführung der Versuche, wobei zunächst die Kulturüberstände der einzelnen Gruppen gewonnen und die DCZellzahl mit Hilfe der Neubauer-Zählkammer bestimmt wurde. Zusätzlich wurde die Anzahl der Makrophagen (MØ) ermittelt. Dazu wurden die adhärenten Zellen, die sich nach 10 Tagen der DC-Kultivierung am Boden der Wells der 12-well-plate befanden, mit Trypsin und Zellschaber vom Boden gelöst. Anschließend wurde die Zellsuspension zentrifugiert (400 g, 4 °C, 10 Minuten) und in 500 µl RPMI-1640-Medium resuspendiert. Danach wurden die MØ ebenfalls in Trypanblau mit Hilfe der Neubauer-Zählkammer gezählt. - 35 - Material und Methoden 3.2.5. In-vitro-Untersuchungen mit Magnesiumbasislegierungen Für die Versuche wurden fünf verschiedene Magnesiumbasislegierungen eingesetzt. Zum einen reines Magnesium (Mg(pur)) und zum anderen MagnesiumCalcium-Legierungen (Gewichtsprozent mit (wt%): vier 0,6; 0,8; unterschiedlichen 1,0; 1,2). Für Calciumkonzentrationen die Versuche wurden Legierungsplättchen mit einem Durchmesser von 8 mm und einer Höhe von 2 mm verwendet. Hergestellt wurden die Legierungen im Institut für Werkstoffkunde der Leibniz Universität Hannover. Die Zylinder wurden aus stranggepresstem Rohmaterial der entsprechenden Calciumgehalte durch spanende Bearbeitung hergestellt. Anschließend wurden die Werkstücke im Institut für Fertigungstechnik und Werkzeugmaschinen der Leibniz Universität Hannover auf einen Durchmesser von 8 mm gedreht und bei einer Höhe von 2 mm abgestochen. Für die Kultivierung der DC wurden zunächst Degradationsmedien aus den einzelnen Legierungen hergestellt (Kapitel 3.2.3.). Im Anschluss erfolgte die Messung der Magnesium- und Calciumkonzentration in den Degradationsmedien (Kapitel 3.2.3.1.). Um den Einfluss der Degradationsmedien aus den einzelnen Legierungen auf die Funktion der DC zu untersuchen, wurden die DC zunächst in den Degradationsmedien kultiviert. Anschließend wurden, wie bei den Magnesium- und Calcium-Salzen, die Zellzahl der DC und der MØ nach der Kultivierung bestimmt. Weiterhin wurden die Vitalität (Kapitel 3.2.8.1.) der DC und die Expression der Oberflächenmoleküle CD11c/CD86 (Kapitel 3.2.8.2.) in der FACS-Analyse untersucht. Außerdem wurden ein Migrationsassay (Kapitel 3.2.7.) und eine MLR (Kapitel 3.2.8.3.) durchgeführt. Die Kulturüberstände wurden mittels ELISA auf das Zytokin TNF-α (Kapitel 3.2.6.) untersucht. - 36 - Material und Methoden 3.2.5.1. Kultivierung der DC in den Degradationsmedien der Magnesiumbasislegierungen Nach der DC-Generierung (Kapitel 3.2.1.) wurden die DC ab Tag 3 der Kultivierung in sieben Gruppen unterteilt. Anschließend wurden die einzelnen Gruppen mit den unterschiedlichen Degradationsmedien versetzt. Die Tabelle 6 gibt einen Überblick über die Kultivierung der DC-Vorläuferzellen und deren Behandlung mit den Degradationsmedien. Die Anzahl der durchgeführten Versuche ist im Anhang (Kapitel 9) dargestellt. - 37 - Material und Methoden Tabelle 6: Übersicht über die Kultivierung der DC-Vorläuferzellen in den Degradationsmedien Tag der Behandlung der DC mit den Degradationsmedien Kultivierung Tag 0 Aussäen von 2x105 DC-Vorläuferzellen/ml in 7 Wells einer 12-wellplate + GM-CSF. Herstellung der Degradationsmedien (Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2) Tag 3 Gewinnung der Degradationsmedien und Sterilfiltration. Zugabe der Degradationsmedien zu den DC der einzelnen Gruppen + GM-CSF. Gruppe A: Degradationsmedien Mg(pur) Gruppe B: Degradationsmedien MgCa0.6 Gruppe C: Degradationsmedien MgCa0.8 Gruppe D: Degradationsmedien MgCa1.0 Gruppe E: Degradationsmedien MgCa1.2 Gruppe F, G: Zugabe von 1ml DC-Medium + GM-CSF Tag 6 Mediumwechsel, GM-CSF Tag 8 Behandlung wie Tag 6 Tag 9 Je nach Versuchsdurchführung Behandlung der Gruppe G als positive Kontrolle. 1. LPS-Stimulation (ELISA, CD11c/CD86) 2. Etoposide (Vitalitätstest) 3. Bleichlorid (Migrationsassay) Tag 10 Versuchsdurchführung: Zellzahlbestimmung der DC und der MØ Vitalitätstest, Migrationsassay, Bestimmung CD11c/CD86, Ansetzen der MLR, Zytokinbestimmung aus Kulturüberstände - 38 - Material und Methoden Die DC der Gruppen A bis E wurden in den Degradationsmedien der Legierungen Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 kultiviert. In der Gruppe F wurden die DC in unbehandeltem DC-Medium kultiviert und die Gruppe G diente als Positiv-Kontrolle. Dazu wurden die DC der Gruppe G, je nach Versuchsdurchführung, ab Tag 9 der Kultivierung unterschiedlich behandelt. In Anlehnung an HÜNERMANN (2006) wurde als Positiv-Kontrolle für die TNF-αSekretion und die Expression von CD11c/CD86 dendritische Zellen mit 1 µg/ml LPS stimuliert, da LPS einen positiven Effekt auf die TNF-α-Sekretion hat und eine Steigerung der CD11c/CD86 positiven DC bewirkt. Als Positiv-Kontrolle für die Migrationsbereitschaft wurden die DC, ebenfalls in Anlehnung an HÜNERMANN (2006), mit 100 µmol/l Bleichlorid inkubiert, da Bleichlorid eine gesteigerte Migration bei DC bewirkt. Als Positiv-Kontrolle für die Bestimmung der Vitalität wurden die DC mit Etoposide [20 µg/ml] (Apoptose-inducer) inkubiert. Am Tag 10 der Kultivierung erfolgte die Durchführung der Versuche, wobei zunächst die Kulturüberstände der einzelnen Gruppen gewonnen und die DC-Zellzahl mit Hilfe der NeubauerZählkammer bestimmt wurde. Zusätzlich wurde die Anzahl der Makrophagen (MØ) ermittelt. Dazu wurden die adhärenten Zellen, die sich nach 10 Tagen der DC-Kultivierung am Boden der Wells der 12-well-plate befanden, mit Trypsin und Zellschaber vom Boden gelöst. Anschließend wurde die Zellsuspension zentrifugiert (400 g, 4 °C, 10 Minuten) und in 500 µl RPMI-1640-Medium resuspendiert. Danach wurden die MØ ebenfalls in Trypanblau mit Hilfe der Neubauer-Zählkammer gezählt. 3.2.6. Zytokinbestimmung Für den Nachweis des Zytokins TNF-α in den Kulturüberständen wurde ein Enzymelinked Immunosorbent Assay (ELISA) durchgeführt. Dazu wurde der DuoSet® Mouse TNF-α ELISA (R&D Systems, Minneapolis, USA) verwendet. Als Positiv-Kontrolle für die TNF-α-Sekretion wurden die DC der Gruppe 5 und 10 (Tabelle 5) bzw. Gruppe G (Tabelle 6) am Tag 9 der DC-Kultivierung mit 1 µg/ml LPS stimuliert. Die Zellen wurden durch Zentrifugieren (400 g, 4°C, 10 min) aus den Kulturüberständen entfernt. - 39 - Material und Methoden Zusätzlich wurde eine Mikrotiterplatte mit 100 µl/Well Capture-Antikörper (TNF-α: 0,8 µg/ml in PBS) beschichtet und über Nacht bei RT inkubiert. Anschließend wurde die Platte drei Mal mit Waschpuffer gewaschen, mit 300 µl/Well Reagent Diluent 1 h bei RT blockiert und erneut gewaschen. Pro Well wurden 100 µl Probe bzw. Standard aufgetragen. Nach 2 h Inkubation bei RT wurde die Platte gewaschen und 100 µl/Well Detection-Antikörper (TNF-α: 150 ng/ml in Reagent Diluent) aufgetragen. Es folgte eine Inkubationszeit von 2 h bei RT. Anschließend wurde die Platte gewaschen und 100 µl/Well Streptavidin-HRP (Meerrettich-Peroxidase) aufgetragen. Nach 20 min wurde die Platte gewaschen und 100 µl/Well TMB-Substrat (Tetramethylbenzidin + H2O2) aufgetragen. Die enzymatische Reaktion wurde nach 10 bis 20 min durch Zugabe von 50 µl/Well 2N H2SO4 gestoppt und die optische Dichte bei 450 nm gemessen. Die Zytokinkonzentration ließ sich aus der Standardkurve berechnen. 3.2.7. Migrationsassay Um die Auswanderungsbereitschaft der unterschiedlich behandelten DC beurteilen zu können, wurde am Tag 10 der DC-Kultivierung ein Migrationsassay (GUTZMER et al., 2004) durchgeführt. Am Tag 8 der Kultivierung wurde vorher das Medium der Gruppe 5 und 10 (Tabelle 5) bzw. Gruppe G (Tabelle 6) als Positiv-Kontrolle mit 100 µmol/l Bleichlorid versetzt (HÜNERMANN, 2006). Am 10. Tag wurden die DC durch Zentrifugation (290 g, 4 °C, 10 Minuten) vom Medium getrennt und in 500 µl neuem DC-Medium resuspendiert. Anschließend wurden die Zellen in Trypanblau (Verdünnung 1:3) mit Hilfe der Neubauer-Zählkammer gezählt. Danach wurden in die Wells einer 6er Transwell-Platte jeweils 1 ml DC-Medium vorgelegt. Anschließend wurden pro Gruppe 200.000 DC in 300 µl Medium auf die Membran einer 6er Transwell-Platte (Porengröße 8 µm) gebracht und für 90 Minuten im Brutschrank (37 °C, 5 % CO2) inkubiert. Nach 90 Minuten Inkubation wurde die Membran vorsichtig entfernt und das Medium aus dem Well gewonnen und zentrifugiert (1000 g, 4 °C, 10 Minuten). Das Zellpellett wurde anschließend in neuem DC-Medium (100 µl) aufgenommen und in Trypanblau mit Hilfe der Neubauer-Zählkammer gezählt. - 40 - Material und Methoden 3.2.8. FACS-Analyse Die FACS-Analyse wurde für die Bestimmung der Vitalität (Annexin V-FITC und PI) (Kapitel 3.2.8.1), die Untersuchung der Expression von CD11c und CD86 (Kapitel 3.2.8.2.) und die Bestimmung der T-Zellproliferationsrate (Kapitel 3.2.9.) verwendet. Die verwendeten Antikörper, die bei den FACS-Analysen zum Einsatz gekommen sind, sind an fluoreszierende Farbstoffe (FITC, PE) gekoppelt und gegen die Oberflächenmoleküle CD11c (PE anti-mouse CD 11c), CD86 (FITC anti-mouse CD 86) und dem membranphospholipid Phosphatidylserin (PS) (Annexin V-FITC) der DC gerichtet. Die Markierung von Zellen für die FACS-Analysen in der hier vorliegenden Arbeit wird unter 3.2.8.1. bis 3.2.8.3. beschrieben. 3.2.8.1. Vitalitätstest (Annexin V-FITC und PI, XTT) Annexin V-FITC und PI: Für die Untersuchung wurde das Kulturmedium der DC der Gruppe 5 und 10 (Tabelle 5) bzw. Gruppe G (Tabelle 6) mit 20 µg/ml Etoposide (VP-16) 16 Stunden vor Versuchsdurchführung inkubiert. Da VP-16 ein Apoptose-Inducer ist, dienten die DC dieser Gruppen als Positiv-Kontrolle. Am Tag 10 der DC-Kultivierung wurden die Kulturmedien der einzelnen Gruppen mit den darin enthaltenen DC abgenommen und zentrifugiert (400 g, 4 °C, 10 Minuten). Anschließend wurden die DC in 500 µl PBS resuspendiert. Danach wurden Aliquots der einzelnen Gruppen in Trypanblau (Verdünnung 1:3) mit Hilfe der NeubauerZählkammer gezählt. Für die Färbung wurde das Annexin V-FITC Apoptosis Detection Kit I (BD Pharmingen™) verwendet. Dazu wurden pro Gruppe 100.000 DC in 200 µl PBS resuspendiert. Nach zweimaligem Waschen in PBS wurden die Zellen im Anschluss in 100 µl Binding-Puffer aufgenommen. Anschließend wurden die DC mit Annexin V-FITC und PI (beide in einer Verdünnung 1:20) versetzt und für 15 Minuten bei RT inkubiert. Danach wurde die Färbung durch Zugabe des vierfachen Volumens an Binding-Puffer gestoppt. Die Auswertung erfolgt mit Hilfe der FACSAnalyse. XTT: Für den Versuch wurden von jeder Gruppe 100.000 DC in 200 µl DCMedium auf eine 96-Well-Mikrotiterplatte gegeben. Anschließend wurde die - 41 - Material und Methoden gebrauchsfertige Bestimmungslösung des XTT Cell Proliferation-Kit (Promega, USA) aufgetragen. Nach einstündiger Inkubation im Brutschrank (37 °C, 5 % CO2) erfolgte die Auswertung in einem Mikrotiterplattenlesegerät (Wellenlänge= 450 nm). 3.2.8.2. Untersuchung der Expression von CD11c und CD86 Zur Untersuchung der DC-Aktivierung wurden die Oberflächenmoleküle CD11c (als DC-Marker) und CD86 (als DC-Aktivierungsmarker) mittels FACSAnalyse bestimmt. Die Fluoreszenzfärbung der Zelloberflächenantigene erfolgte direkt über Fluorescein-Isothiocyanat (FITC) und Phycoerthrin (PE) markierten Antikörpern. Die anschließend gemessene Intensität des Fluoreszenzsignals ist proportional zur Menge an gebundenen Antikörpern und gibt somit Auskunft über die Expression der Oberflächenmoleküle CD11c und CD86. Als Positiv-Kontrollen wurden die DC der Gruppen 5 und 10 (Tabelle 5) bzw. Gruppe G (Tabelle 6) mit LPS (1µg/ml) stimuliert. Anschließend wurden die DC der einzelnen Gruppen in PBS aufgenommen und auf eine 96-Well-Mikrotiterplatte gegeben. Danach wurde die Platte zentrifugiert (400 g, 4 °C, 6 Minuten) und der Überstand verworfen. Im Anschluss wurden die Zellen in PBS/BSA-Waschpuffer (1 %ig) gewaschen. Danach wurden die Antikörper PE (anti-mouse CD11c) und FITC (anti-mouse CD86) in PBS/BSA (Verdünnung 1:200) auf die DC gegeben. Nach 30 Minuten Inkubation wurden die Zellen zwei Mal in PBS/BSA gewaschen und in Reaktionsgefäße überführt. Anschließend erfolgte die Auswertung mittels FACSAnalyse. 3.2.8.3. MLR (Mixed Leukocyte Reaction) Für den Versuch wurden zunächst die T-Lymphozyten aus der Milz einer NMRI-Maus gewonnen und gezählt (Kapitel 3.2.2.). Anschließend wurden 4x106 TLymphozyten in 4 ml PBS aufgenommen und mit 0,5 µmol/ml CFSE versetzt. Damit der Farbstoff in die Zellen diffundieren konnte, erfolgte eine Inkubation von 10 Minuten im Brutschrank (37 °C, 5 % CO2). Anschließend wurde die Reaktion durch Zugabe des 5-fachen Volumens eiskalten RPMI-Medium und 5-minütiger Inkubation - 42 - Material und Methoden auf Eis gestoppt. Danach wurden die Zellen zentrifugiert (1000 g, 4 °C, 10 Minuten) und in 4 ml frischem RPMI-Medium resuspendiert. Dieser Schritt wurde zweimal wiederholt. Anschließend wurden die T-Lymphozyten in 4 ml DC-Medium aufgenommen. Für die Inkubation mit den unterschiedlich behandelten DC, wurden jeweils 105 T-Lymphozyten mit 104 DC pro Gruppe in Wells einer 96-Well-Platte (round-bottom) gegeben. Danach wurden die Zellen für 4 Tage im Brutschrank (37 °C, 5 % CO2) kultiviert. Für die Auswertung wurden die Zellen der einzelnen Gruppen am 4. Tag zentrifugiert und in frischem DC-Medium resuspendiert. Die Auswertung der Proliferationsrate der T-Lymphozyten erfolgte mit Hilfe der FACS-Analyse. 3.3. Statistische Auswertung Die Darstellungen der Ergebnisse (DC-Zellzahl und MØ-Zellzahl, TNF-αELISA, Migrationsassay und XTT-Test) erfolgten unter Angabe von Mittelwerte (MW) und Standardabweichungen (S.D.). Die statistische Auswertung der Ergebnisse erfolgte mit dem Programm GraphPadPrism. Die unterschiedlich behandelten Gruppen sind jeweils mit der Kontrollgruppe bzw. untereinander auf signifikante Differenzen untersucht worden. Eine Irrtumswahrscheinlichkeit von 5% galt als schwach signifikant (*); lag sie bei 1%, wurde sie als signifikant (**) bzw. bei 0,1% als hoch signifikant (***) eingestuft. Hierfür wurde zuerst eine parametrische Varianzanalyse (One-way-ANOVA) und im Anschluss der Dunnetts Test als posthoc-Test durchgeführt. Die Ausnahmen hierbei sind die Ergebnisse der MLR, CD11c/CD86, Annexin V-FITC- Vitalitätstest. Die Darstellungen der Ergebnisse dieser Tests sind repräsentative Ergebnisse aus drei identisch verlaufenden Versuchen. Alle Versuche wurden in mindestens drei unabhängigen Anläufen durchgeführt. Die Anzahl der Messwerte (n) im Ergebnisteil gibt den Stichprobenumfang identischer Versuche an. Die Einzelwerte aller durchgeführten Versuche sind in den Anhangstabellen (Kapitel 9) aufgeführt. Die Darstellung der Ergebnisse der Konzentrationsmessungen für Magnesium und Calcium sind Einzelwerte aus acht identisch durchgeführten - 43 - Konzentrationsmessungen. Ergebnisse 4. Ergebnisse 4.1. Magnesium- und Calciumkonzentration in den CaCl2 wurden Degradationsmedien Als Grundlage für die Versuche mit MgCl2 und Degradationsversuche mit reinem Magnesium (Mg(pur)) und vier verschiedenen Magnesiumbasislegierungen durchgeführt (MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2). Die Degradationsversuche zeigten keine signifikanten Unterschiede zwischen den einzelnen Legierungen in Bezug auf die Magnesiumkonzentration (Abb. 4.1.-a). In der Regel schwankten die Magnesiumkonzentrationen in den Degradationsmedien zwischen 0,6 und 1,5 mmol/l. Bei den Legierungen mit erhöhten wt% Calcium (MgCa1.0, MgCa1.2) zeigten sich jedoch größere Unterschiede bezüglich ihrer Degradationsgeschwindigkeit. So wurden bei dem Degradationsmedium der Legierung MgCa1.0 z.T. Magnesiumkonzentrationen bis zu 3,5 mmol/l gemessen. Durch die Kalibrationsstandards für CaCl2 zeigte sich, dass bis zu einer CaCl2Konzentration von 0,2 mmol/l die optische Dichte erfassbar und eindeutig zugeordnet werden konnte. Die Calciumkonzentration in den Degradationsmedien war nach 72stündiger Inkubation im Wasserbad unterhalb der Nachweisgrenze von 0,2 mmol/l. - 44 - Ergebnisse Mg (mmol/l) 4 3 2 1 0 a M gC a M gC a gC M gC M 1. 0 0. 8 0. 6 a M 1. 2 MgCa0.6 MgCa0.8 MgCa1.0 MgCa1.2 gP Mg(pur) Abb. 4.1.-a: Einzelwerte der Mg2+-Konzentration [mmol/l] in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2. Die Legierungsplättchen 2 (Höhe 2 mm, Ø 8 mm, Oberfläche 150,8 mm ) wurden über 72 h mit 10 ml DC-Medium im Wasserbad (37 °C) inkubiert. Dargestellt sind die Einzelwerte aus acht identisch durchgeführten Messungen. Die Einzelwerte siehe Tabelle 9.1. im Anhang. 4.2. Vergleichende Darstellung der In-vitro-Ergebnisse zwischen MgCl2, CaCl2 und den Magnesiumbasislegierungen Für einen besseren Vergleich der Versuche mit MgCl2, CaCl2 und den Magnesiumlegierungen, werden die Ergebnisse aus den einzelnen Versuchen zusammen dargestellt. 4.2.1. DC-Zellzahl und MØ-Zellzahl im Vergleich zwischen MgCl2, CaCl2 und Degradationsmedien Am Tag 10 der DC-Kultivierung wurden die unterschiedlich behandelten DC in Trypanblau mit Hilfe der Neubauer-Zählkammer gezählt. Zusätzlich wurden die MØ vom Boden der Wells mit Trypsin gelöst und ebenfalls mit Hilfe der NeubauerZählkammer gezählt. Bei den DC, die mit erhöhten MgCl2-Konzentrationen im Kulturmedium kultiviert wurden, zeigte sich ein signifikanter Rückgang der DC- - 45 - Ergebnisse Zellzahl (Abb. 4.2.1-a). Außerdem ist eine Dosisabhängigkeit erkennbar. Bei einer Mg2+-Konz. von 10 mmol/l im Kulturmedium ist der Rückgang der DC hoch signifikant. Bei der Auszählung der MØ-Zellzahl zeigte sich, dass bei einer Mg2+Konz. von 10 mmol/l im Kulturmedium der Anteil an MØ signifikant zunimmt (Abb. 4.2.1-b). Auch hier deutet sich eine Dosisabhängigkeit an. 1400000 1200000 * ** DC-Zellzahl 1000000 *** 800000 600000 400000 200000 0 Kontrolle MgCl 0.1 mmol/l MgCl 1 mmol/l MgCl 10 mmol/l Abb. 4.2.1.-a: DC-Zellzahlen nach 10-tägiger Kultivierung in mit MgCl2 angereichertem DCMedium. Die DC der Kontrollgruppe wurden unter Standardbedingungen kultiviert. Mit steigender MgCl2-Konzentration im Kulturmedium zeigt sich ein stärkerer Rückgang der DC-Zellzahl im Vergleich zur Kontrolle. Dargestellt sind die Mittelwerte aus drei identisch verlaufenden Versuchen. * P < 0,05; ** P < 0,01; *** P < 0,001 verglichen mit Kontrolle. Die Einzelwerte sind im Anhang Tabelle 9.2. aufgeführt. - 46 - Ergebnisse ** 120000 100000 Zellzahl 80000 60000 40000 20000 0 Kontrolle MgCl 0.1 mmol/l MgCl 1 mmol/l MgCl 10 mmol/l Abb. 4.2.1.-b: MØ-Zellzahl nach 10-tägiger Kultivierung der DC-Vorläuferzellen in mit MgCl2 angereichertem DC-Medium. Die MØ-Zellzahl der Kontrollgruppe sind MØ, die bei Standardkultivierung von DC-Vorläuferzellen entstanden sind. Bei einer MgCl2-Konzentration von 10 mmol/l im Kulturmedium zeigte sich, dass der Anteil an MØ aus den DC-Vorläuferzellen signifikant zunimmt. Dargestellt sind die Mittelwerte aus drei identisch verlaufenden Versuchen. ** P < 0,01 verglichen mit Kontrolle. Die Einzelwerte sind im Anhang Tabelle 9.3. aufgeführt. - 47 - Ergebnisse In der CaCl2-Gruppe zeigte sich ebenfalls, dass mit erhöhter Ca2+Konzentration im Kulturmedium der Anteil der DC nach 10-tägiger Kultivierung der DC-Vorläuferzellen abnimmt (Abb. 4.2.1.-c) Ab einer Konzentration von 1 mmol/l CaCl2 ist der Rückgang der DC signifikant. Eine Dosishabhängigkeit zeichnet sich ab. Bei der Auszählung der MØ zeigte sich, dass der Anteil an MØ aus den DCVorläuferzellen bei einer CaCl2-Konzentration von 10 mmol/l signifikant zunimmt (Abb. 4.2.1.-d). Auch in der CaCl2-Gruppe zeichnete sich eine Dosisabhängigkeit ab. 1400000 * 1200000 1000000 Zellzahl *** 800000 600000 400000 200000 0 Kontrolle CaCl 0.1 mmol/l CaCl 1 mmol/l CaCl 10 mmol/l Abb. 4.2.1.-c: DC-Zellzahlen nach 10-tägiger Kultivierung in mit CaCl2 angereichertem DCMedium. Die DC der Kontrollgruppe wurden unter Standardbedingungen kultiviert. Mit steigender CaCl2-Konzentration im Kulturmedium zeigte sich ein stärkerer Rückgang der DC-Zellzahl im Vergleich zur Kontrolle. Dargestellt sind die Mittelwerte aus drei identisch verlaufenden Versuchen. * P < 0,05; *** P < 0,001 verglichen mit Kontrolle. Die Einzelwerte sind im Anhang Tabelle 9.4. aufgeführt. - 48 - Ergebnisse 160000 *** 140000 120000 Zellzahl 100000 80000 60000 40000 20000 0 Kontrolle CaCl 0.1 mmol/l CaCl 1 mmol/l CaCl 10 mmol/l Abb. 4.2.1.-d: MØ-Zellzahlen nach 10-tägiger Kultivierung der DC-Vorläuferzellen in mit CaCl2 angereichertem DC-Medium. Die MØ-Zellzahl der Kontrollgruppe sind MØ, die bei Standardkultivierung von DC-Vorläuferzellen entstanden sind. Bei einer CaCl2-Konzentration von 10 mmol/l im Kulturmedium zeigte sich, dass der Anteil an MØ aus den DC-Vorläuferzellen signifikant zunimmt. Dargestellt sind die Mittelwerte aus drei identisch verlaufenden Versuchen. *** P < 0,001 verglichen mit Kontrolle. Die Einzelwerte sind im Anhang Tabelle 9.5. aufgeführt. - 49 - Ergebnisse Bei den Versuchen mit den Degradationsmedien wurden die DC- Vorläuferzellen ab Tag 3 der DC-Kultivierung in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2 angezüchtet. Am Tag 10 erfolgte die Auszählung der DC mittels Neubauer-Zählkammer. Es zeigte sich nur bei den Degradationsmedien der Legierungen MgCa1.0 und MgCa1.2 ein signifikanter Rückgang der DC-Zellzahl nach 10-tägiger Kultivierung der DC- DC Zellzahl am Tag 10 der Kultivierung Vorläuferzellen (Abb. 4.2.1.-e). 1200000 1000000 * ** MgCa 1.0 MgCa 1.2 800000 600000 400000 200000 0 Kontrolle Mg(pur) MgCa 0.6 MgCa 0.8 Abb. 4.2.1.-e: DC-Zellzahlen nach 10-tägiger Kultivierung der DC-Vorläuferzellen in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2. Für die Degradationsmedien wurden die einzelnen Legierungen über 72 h mit dem DC-Medium im Wasserbad (37 °C) inkubiert. DC der Kontrollgruppe wurden unter Standardbedingungen kultiviert. Dargestellt sind die Mittelwerte aus fünf identisch verlaufenden Versuchen. * P < 0,05; ** P < 0,01 verglichen mit Kontrolle. Die Einzelwerte sind im Anhang Tabelle 9.6. aufgeführt. - 50 - Ergebnisse Bei der Auswertung der MØ-Zellzahl zeigte sich, dass bei den DCVorläuferzellen, die im Degradationsmedium der Legierung MgCa1.0 kultiviert Adherente Zellen am Tag 10 der Kultivierung wurden, der Anteil an MØ tendenziell erhöht ist (Abb. 4.2.1.-f). 60000 * 50000 40000 30000 20000 10000 0 Kontrolle Mg(pur) MgCa 0.6 MgCa 0.8 MgCa 1.0 MgCa 1.2 Abb. 4.2.1.-f: MØ-Zellzahlen nach 10-tägiger Kultivierung der DC-Vorläuferzellen in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2. Die MØ-Zellzahl der Kontrollgruppe sind MØ, die bei Standardkultivierung von DC-Vorläuferzellen entstanden sind. Bei dem Degradationsmedium der Legierung MgCa1.0 zeigt sich, dass der Anteil an MØ aus den DC-Vorläuferzellen tendenziell zunimmt. Dargestellt sind die Mittelwerte aus fünf identisch verlaufenden Versuchen. * P < 0,05 verglichen mit Kontrolle. Die Einzelwerte sind im Anhang Tabelle 9.7. aufgeführt. - 51 - Ergebnisse 4.2.2. Vitalität (Annexin V-FITC und PI, XTT) Als Voraussetzung für die nachfolgenden Versuche wurde zunächst mittels FACS-Analyse (Annexin V-FITC) und XTT-Test die Vitalität der DC bestimmt. Annexin V-FITC: Als Positivkontrolle für die FACS-Analyse wurde ein Teil der DC am Tag 9 mit 20 µg/ml Etoposide versetzt. Die Markierung mit PI zeigte in der Analyse keinen Unterschied zwischen den einzelnen Versuchsansätzen und ist daher in den folgenden Darstellungen nicht dargestellt. MgCl2: Ab dem 3. Tag der DC-Kultivierung wurden die Kulturmedien der einzelnen Gruppen auf 0.1 mmol/l, 1.0 mmol/l und 10 mmol/l MgCl2 eingestellt. Am Tag 10 erfolgte die Untersuchung der Vitalität der DC. In Abbildung 4.2.2.-a sind die Histogramme von Annexin V-FITC markierten DC eines repräsentativen Versuches dargestellt. Die Tabelle 7 zeigt eine Zusammenstellung der prozentualen Annexin V-FITC positiven DC. Der Einfluss von MgCl2 auf die Vitalität der DC wurde in drei unabhängigen Versuchen dargestellt (Werte im Anhang Tabelle 9.8.). Bei den mit MgCl2 behandelten DC war im Vergleich zur Kontrolle und der Positivkontrolle kein Anstieg an apoptotischen Zellen zu erkennen. Bei der Positivkontrolle mit Etoposide wurde ein Anstieg der apoptotischen DC von über 50% gemessen. - 52 - Ergebnisse a) b) 25,6 % 50,5 % d) c) 20,6 % e) 17,1 % 17,2 % Abb. 4.2.2.-a: Histogramme der Analyse Annexin V-FITC von DC, die mit MgCl2 angereichertem Kulturmedium kultiviert wurden: a) unbehandelte Kontrolle (DC, die unter Standardbedingungen kultiviert wurden), b) Positivkontrolle (Zugabe von 20 µg/ml Etoposide 24 h vor Versuchsdurchführung), c) MgCl2 0,1 mmol/l, d) MgCl2 1,0 mmol/l, e) MgCl2 10 mmol/l. Dargestellt ist ein repräsentatives Ergebnis aus drei identisch verlaufenden Versuchen. Pro Ansatz wurden 5000 DC gemessen. Die Einzelwerte sind im Anhang Tab.9.8. aufgeführt. Tabelle 7: Zusammenstellung der prozentualen Annexin V-FITC positiven DC Probe Annexin V-FITC a) Unbehandelte Kontrolle 25,6 % b) Etoposide (Positiv-Kontrolle) 50,5 % c) MgCl2 0,1 mmol/l 20,6 % d) MgCl2 1,0 mmol/l 17,1 % e) MgCl2 10 mmol/l 17,2 % - 53 - Ergebnisse CaCl2 : Für die Untersuchungen mit CaCl2 wurden die DC ab Tag 3 der Kultivierung in mit 0,1 mmol/l, 1,0 mmol/l, 10 mmol/l CaCl2 versetzten Kulturmedien kultiviert. Am Tag 10 erfolgte die Durchführung des Vitalitätstest (Annexin V-FITC und PI). In der Abbildung 4.2.2.-b sind die Histogramme der Analyse Annexin V-FITC von DC eines repräsentativen Versuchs dargestellt. Die Tabelle 8 zeigt eine Zusammenstellung der prozentualen Annexin V-FITC positiven DC. Der Einfluss von CaCl2 auf die Vitalität der DC wurde in drei unabhängigen Versuchen dargestellt (Werte im Anhang Tabelle 9.9.). Im Vergleich zur Kontrolle zeigten die DC bis zu einer CaCl2-Konz. von 1,0 mmol/l keinen Rückgang der Vitalität. Bei 10 mmol/l CaCl2 zeigten die DC allerdings einen erheblichen Anstieg der apoptotischen DC auf über 70 %. - 54 - Ergebnisse b) a) 25,6 % c) 50,5 % d) e) 15,4 % 17,2 % 71,8 % Abb. 4.2.2.-b: Analyse Annexin V-FITC von DC, die mit CaCl2 angereicherten Kulturmedium ab Tag 3 der DC-Kultivierung kultiviert wurden: a) unbehandelte Kontrolle (DC die unter Standardbedingungen kultiviert wurden): b) Positivkontrolle (Zugabe von 20 µg/ml Etoposide 24 h vor Versuchsdurchführung), c) CaCl2 0,1 mmol/l, d) CaCl2 1,0 mmol/l, e) CaCl2 10 mmol/l. Dargestellt ist ein repräsentatives Ergebnis aus drei identisch verlaufenden Versuchen. Pro Ansatz wurden 5000 DC gemessen. Die Einzelwerte sind im Anhang Tabelle 9.9. aufgeführt. Tabelle 8: Zusammenstellung der prozentualen Annexin V-FITC positiven DC Probe Annexin V-FITC a) Unbehandelte Kontrolle 25,6 % b) Etoposide (Positiv-Kontrolle) 50,5 % c) CaCl2 0,1 mmol/l 15,4 % d) CaCl2 1,0 mmol/l 17,2 % e) CaCl2 10 mmol/l 71,8 % - 55 - Ergebnisse Degradationsmedien: Die DC wurden ab Tag 3 der Kultivierung in den Degradationsmedien der Legierungen Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 kultiviert. Für die Degradationsmedien wurden die Legierungen über drei Tage im DC-Medium inkubiert (Kap.:3.2.2.1.). Die Auswertung der Vitalität erfolgte am Tag 10 der DC-Kultivierung mittels FACS-Analyse. In der Abbildung 4.2.2.-c sind die Histogramme eines repräsentativen Versuchs der Analyse Annexin V-FITC dargestellt. Die Tabelle 9 zeigt eine Zusammenstellung der prozentualen Annexin VFITC positiven DC. Der Einfluss der Degradationsmedien auf die Vitalität der DC wurde in drei unabhängigen Versuchen dargestellt (Werte im Anhang Tab. 9.10.). Die Analyse zeigte, dass die Vitalität der DC, die in den Degradationsmedien kultiviert wurden, im Vergleich zu den Kontrollen, nicht wesentlich beeinträchtigt wurde. Allerdings stieg der Anteil an apoptotischen DC in der Gruppe Mg(pur) leicht auf 33,4 % an. Im Vergleich zur Positivkontrolle ist dieser Anstieg jedoch sehr gering. - 56 - Ergebnisse a) b) c) 25,6 % 50,5 % d) e) 33,4 % f) 28,0 % 26,6 % 24,8 % Abb. 4.2.2.-c: Histogramme der Analyse Annexin V-FITC von g) DC, die in den Degradationsmedien der Legierungen Mg(pur), 24,5 % MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2 ab Tag 3 der DCKultivierung kultiviert wurden: a) unbehandelte Kontrolle (DC die unter Standardbedingungen kultiviert wurden): b) Positivkontrolle (Zugabe von 20 µg/ml Etoposide 24 h vor Versuchsdurchführung), c) Mg(pur), d) MgCa0.6, e) MgCa0.8, f) MgCa1.0, g) MgCa1.2. Dargestellt ist ein repräsentatives Ergebnis aus drei identisch verlaufenden Versuchen. Pro Ansatz wurden 5000 DC gemessen. Die Einzelwerte sind im Anhang Tabelle 9.10. aufgeführt. Tabelle 9: Zusammenstellung der prozentualen Annexin V-FITC positiven DC Probe Annexin V-FITC a) Unbehandelte Kontrolle 25,6 % b) Etoposide (Positiv-Kontrolle) 50,5 % c) Mg(pur) 33,4 % d) MgCa0.6 28,0 % e) MgCa0.8 26,6 % f) MgCa1.0 24,8 % g) MgCa1.2 24,5 % - 57 - Ergebnisse XTT-Test: Zum Nachweis der Zellvitalität wurde zusätzlich ein XTT-Test durchgeführt (Kapitel 3.2.1.7.). Bei den Darstellungen handelt es sich um Zusammenfassungen der Mittelwerte aus vier Versuchen. Die Vitalität der Kontrolle wurde auf 100% gesetzt um die Versuche besser vergleichen zu können. MgCl2: Für die Versuche mit MgCl2 wurden die Kulturmedien der DC ab Tag 3 der Kultivierung auf unterschiedliche MgCl2-Konz. (0,1 mmol/l, 1,0 mmol/l, 10 mmol/l) eingestellt. Am Tag 10 erfolgte die Durchführung des XTT-Test. Die DC zeigten unabhängig von der MgCl2-Konz. keine Beeinträchtigung bezüglich ihrer Vitalität im Vergleich zur Kontrolle (Abb. 4.2.2.-d). 120 % vitale Zellen 100 80 60 40 20 0 Kontrolle MgCl 0.1 mmol/l MgCl 1 mmol/l MgCl 10 mmol/l Abb. 4.2.2.-d: XTT-Test nach Inkubation der DC mit MgCl2 ab Tag 3 der DC-Kultivierung. Die Kontrolle sind DC, die unter Standardbedingungen kultiviert wurden. Im Vergleich zur Kontrolle zeigten die DC, die mit MgCl2 behandelt wurden, keinen signifikanten Rückgang ihrer Vitalität. Dargestellt sind die Mittelwerte der prozentualen Anteile vitaler Zellen. n=4. Die Einzelwerte sind im Anhang Tabelle 9.11 aufgeführt. - 58 - Ergebnisse CaCl2: Für die Versuche mit CaCl2 wurden die Kulturmedien ab Tag 3 der DCKultivierung auf 0,1 mmol/l, 1,0 mmol/ und 10 mmol/l CaCl2 eingestellt. Am Tag 10 der Kultivierung erfolgte der XTT-Test. Der Vitalitätstest zeigte einen Rückgang der vitalen Zellen bei einer Konz. von 10 mmol/l CaCl2 im Vergleich zur Kontrolle (Abb. 4.2.2.-e). 120 ** % vitale Zellen 100 80 60 40 20 0 Kontrolle CaCl 0.1 mmol/l CaCl 1 mmol/l CaCl 10 mmol/l Abb. 4.2.2.-e: XTT-Test nach Inkubation der DC mit CaCl2 ab Tag 3 der DC-Kultivierung. Die Kontrolle sind DC, die unter Standardbedingungen kultiviert wurden. Im Vergleich zur Kontrolle war bei einer Konz. von 10 mmol/l CaCl2 ein signifikanter Rückgang der vitalen Zellen zu beobachten. Dargestellt sind die Mittelwerte der prozentualen Anteile vitaler Zellen. n=4. ** P < 0,01 verglichen mit Kontrolle. Die Einzelwerte sind im Anhang Tabelle 9.12. aufgeführt. - 59 - Ergebnisse Degradationsmedien: Für den XTT-Test wurden die DC ab dem 3. Tag der Kultivierung in den Degradationsmedien der Legierungen Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 kultiviert. Am 10. Tag erfolgte die Auswertung der Vitalität der DC mittels XTT-Test. Hierbei konnte kein signifikanter Rückgang der vitalen Zellen bei den unterschiedlich behandelten DC im Vergleich zur Kontrolle festgestellt werden (Abb. 4.2.2.-f). 120 % vitale Zellen 100 80 60 40 20 0 Kontrolle Mg(pur) MgCa0.6 MgCa0.8 MgCa1.0 MgCa1.2 Abb. 4.2.2.-f: XTT-Test der DC nach Inkubation in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2 ab Tag 3 der DC-Kultivierung wurden. Die Kontrolle sind DC, die unter Standardbedingungen kultiviert wurden. Die DC, die in den unterschiedlichen Degradationsmedien kultiviert wurden, zeigten keinen signifikanten Rückgang vitaler Zellen. Dargestellt sind die Mittelwerte der prozentualen Anteile vitaler Zellen. n=5. Die Einzelwerte sind im Anhang Tabelle 9.13. aufgeführt. - 60 - Ergebnisse 4.2.3. Freisetzung von TNF-α Für die Untersuchung der Freisetzung des Zytokins TNF-α wurden die Kulturüberstande der unterschiedlich behandelten DC gewonnen und mittels ELISA (Kapitel 3.2.1.4.) untersucht. Als Negativ-Kontrolle blieb ein Ansatz unbehandelt und als Positiv-Kontrolle wurde eine Gruppe der DC am Tag 9 der Kultivierung mit LPS (1 µg/ml) stimuliert. MgCl2: Die DC wurden ab Tag 3 der DC-Kultivierung in Kulturmedien mit 2+ Mg -Konz. von 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l kultiviert. Nach 10-tägiger Kultivierung wurden die Kulturüberstände gewonnen und mittels ELISA auf das proinflammatorische Zytokin TNF-α untersucht. Einen signifikanten Anstieg der TNFα-Ausschüttung konnte nur bei der Positiv-Kontrolle festgestellt werden. Die Ausschüttung von TNF-α bei den mit MgCl2 behandelten DC blieb auf dem Niveau der Negativ-Kontrolle (Abb. 4.2.3.-a). 1000 TNF alpha [pg/ml] 900 *** 800 700 600 500 400 300 200 100 0 Ko LPS Ko MgCl 0.1 mmol/l MgCl 1 mmol/l MgCl 10 mmol/l Abb. 4.2.3.-a: TNF-α [pg/ml] in Kulturüberständen von DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l MgCl2 ab Tag 3 der DC-Kultivierung. Ein signifikanter Anstieg der TNF-α zeigte sich nur bei den DC, die mit LPS stimuliert wurden. Dargestellt sind die Mittelwerte der Messungen aus fünf Kulturüberständen. *** P < 0,001 verglichen mit Kontrolle. Die Einzelwerte sind im Anhang Tabelle 9.14. aufgeführt. - 61 - Ergebnisse CaCl2: In der CaCl2-Gruppe wurden die DC ab Tag 3 der DC-Kultivierung in Kulturmedien mit Ca2+-Konz. von 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l kultiviert. Nach 10-tägiger Kultivierung wurden die Kulturüberstände gewonnen und auf das Zytokin TNF-α untersucht. Auch in den Kulturüberständen der mit CaCl2 behandelten DC wurde keine signifikante Erhöhung der TNF-α-Freisetzung im Vergleich zur Negativkontrolle gemessen (Abb. 4.2.3.-b). 1000 TNF alpha [pg/ml] 900 *** 800 700 600 500 400 300 200 100 0 Ko LPS Ko CaCl 0.1 mmol/l CaCl 1 mmol/l CaCl 10 mmol/l Abb. 4.2.3.-b: TNF-α [pg/ml] in Kulturüberständen von DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l CaCl2 ab Tag 3 der DC-Kultivierung. Ein signifikanter Anstieg der TNF-α zeigte sich nur bei den DC, die mit LPS stimuliert wurden. Dargestellt sind die Mittelwerte der Messungen aus fünf Kulturüberständen. *** P < 0,001 verglichen mit Kontrolle. Die Einzelwerte sind im Anhang Tabelle 9.15. aufgeführt. - 62 - Ergebnisse Degradationsmedien: Die DC wurden ab Tag 3 der Kultivierung in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 kultiviert. Nach 10-tägiger Kultivierung wurden die Kulturüberstände gewonnen und mittel ELISA auf die Freisetzung von TNF-α untersucht. Im Vergleich zur Negativ-Kontrolle konnte kein signifikanter Anstieg der Zytokinausschüttung festgestellt werden (Abb. 4.2.3.-c). 900 *** 800 TNF alpha [pg/ml] 700 600 500 400 300 200 100 0 Ko LPS Ko Mg(pur) MgCa0.6 MgCa0.8 MgCa1.0 MgCa1.2 Abb. 4.2.3.-c: TNF-α [pg/ml] in Kulturüberständen von DC nach Kultivierung in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2 . Ein signifikanter Anstieg der TNF-α zeigte sich nur bei den DC, die mit LPS stimuliert wurden. Dargestellt sind die Mittelwerte der Messungen aus fünf Kulturüberständen. *** P < 0,001 verglichen mit Kontrolle. Die Einzelwerte sind im Anhang Tabelle 9.16. aufgeführt. - 63 - Ergebnisse 4.2.4. Migrationsassay Um die Auswanderungsbereitschaft der DC nach MgCl2-, CaCl2- und Degradationsmedium-Inkubation beurteilen zu können, wurde am Tag 10 der DCKultivierung ein Migrationsassay durchgeführt. Als Negativ-Kontrolle wurde ein DCAnsatz unter Standardbedingungen kultiviert, als Positiv-Kontrolle wurde ein DCAnsatz am Tag 9 der Kultivierung mit 100 µmol/l Bleichlorid (Pb) versetzt. Bei den DC, die mit MgCl2 inkubiert wurden, zeigte sich bei einer Konzentration von 10 mmol/l MgCl2 ein signifikanter Anstieg der Auswanderungsbereitschaft der DC (Abb. 4.2.4.-a). Auch die DC, die mit CaCl2 inkubiert wurden, zeigten ab einer Konzentration von 10 mmol/l CaCl2 einen signifikanten Anstieg der Migrationsbereitschaft (Abb. 4.2.4.-b). Bei den DC, die in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 kultiviert wurden, zeigte sich bei MgCa1.2 ein schwach signifikanter Anstieg der Auswanderungsbereitschaft der DC (Abb. 4.2.4.-c). 45000 *** Ausgewanderte DC/ml 40000 ** 35000 30000 25000 20000 15000 10000 5000 0 Kontrolle Pb 100 µmol/l MgCl 0.1 mmol/l MgCl 1.0 mmol/l MgCl 10 mmol/l Abb. 4.2.4.-a: Migrationsassay der DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l MgCl2 ab Tag 3 der DC-Kultivierung. Die Kontrolle sind DC, die unter Standardbedingungen kultiviert wurden. Als Positiv-Kontrolle dienten DC, die am Tag 9 der Kultivierung mit 100 µmol/l Bleichlorid (Pb) inkubiert wurden. Im Vergleich zur Kontrolle zeigte sich ein signifikanter Anstieg der Auswanderungsbereitschaft der DC bei einer Konzentration von 10 mmol/l MgCl2. Dargestellt sind die Mittelwerte aus sechs identisch durchgeführten Versuchen. ** P < 0,01; *** P < 0,001 verglichen mit Kontrolle. Einzelwerte sind im Anhang Tabelle 9.17. aufgeführt. - 64 - Ergebnisse 45000 *** Ausgewanderte DC/ml 40000 *** 35000 30000 25000 20000 15000 10000 5000 0 Kontrolle Pb 100 µmol/l CaCl 0.1 mmol/l CaCl 1 mmol/l CaCl 10 mmol/l Abb. 4.2.4.-b: Migrationsassay von DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l CaCl2 ab Tag 3 der DC-Kultivierung. Die Kontrolle sind DC, die unter Standardbedingungen kultiviert wurden. Als Positiv-Kontrolle dienten DC, die am Tag 9 der Kultivierung mit 100 µmol/l Bleichlorid (Pb) inkubiert wurden. Im Vergleich zur Kontrolle zeigte sich ein signifikanter Anstieg der Auswanderungsbereitschaft der DC bei einer Konzentration von 10 mmol/l CaCl2. Dargestellt sind die Mittelwerte aus sechs identisch durchgeführten Versuchen. *** P < 0,001 verglichen mit Kontrolle. Einzelwerte sind im Anhang Tabelle 9.18. aufgeführt. - 65 - Ausgewanderte DC/ml Ergebnisse 50000 45000 40000 35000 30000 25000 20000 15000 10000 5000 0 *** * Kontrolle Pb 100 µmol/l Mg(pur) MgCa0.6 MgCa0.8 MgCa1.0 MgCa1.2 Degradationsmedien Abb. 4.2.4.-c: Migrationsassay der DC, die in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 ab Tag 3 der DC-Kultivierung kultiviert wurden. Die Kontrolle sind DC, die unter Standardbedingungen kultiviert wurden. Als Positiv-Kontrolle dienten DC, die am Tag 9 der Kultivierung mit 100 µmol/l Bleichlorid (Pb) inkubiert wurden. Im Vergleich zur Kontrolle zeigte sich ein schwach signifikanter Anstieg der Auswanderungsbereitschaft der DC, die im Degradationsmedium der Legierung MgCa1.2 kultiviert wurden. Dargestellt sind die Mittelwerte aus neun identisch durchgeführten Versuchen. . * P < 0,05; *** P < 0,001 verglichen mit Kontrolle. Einzelwerte sind im Anhang Tabelle 9.19. aufgeführt. 4.2.5. Expression der Oberflächenmoleküle CD11c und CD86 Mittels FACS-Analyse wurde die Expression der Oberflächenmoleküle CD11c als DC-Marker und CD86 als DC-Aktivierungsmarker nach MgCl2-, CaCl2- und Degradationsmedium-Inkubation gemessen. Als Positiv-Kontrolle wurde eine Gruppe der DC am Tag 9 der Kultivierung mit LPS stimuliert. Die Darstellungen sind repräsentative Ergebnisse aus drei identisch durchgeführten Versuchen und werden als Dotplots dargestellt, wobei der Quadrant G2 die CD11c- und CD86-positiven DC darstellt. In der Abbildung 4.2.5.-a sind die Dotplots von DC, die ab Tag 3 ihrer Kultivierung in Kulturmedien mit Konzentrationen von 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l MgCl2 kultiviert wurden. Die Tabelle 10 zeigt eine zusammenfassende Darstellung der CD11c/CD86-positiven DC nach MgCl2-Inkubation. In der MgCl2Gruppe konnte keine Erhöhung der CD11c/CD86-positiven Zellen im Vergleich zur - 66 - Ergebnisse Negativkontrolle festgestellt werden. In der Abbildung 4.2.5.-b sind die Dotplots von DC, die mit CaCl2-Konz von 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l inkubiert wurden. Die Tabelle 11 zeigt eine zusammenfassende Darstellung der CD11c/CD86-positiven DC nach CaCl2-Inkubation. Hier zeigte sich ebenfalls keine Erhöhung der CD11c/CD86-positiven Zellen innerhalb der CaCl2-Gruppe im Vergleich zur Negativkontrolle. In der Abbildung 4.2.5.-b sind die Dotplots von DC, die in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 kultiviert wurden. Die Tabelle 12 zeigt eine zusammenfassende Darstellung der CD11c/CD86-positiven DC nach Kultivierung in den entsprechenden Degradationsmedien. Bei den DC, die in den Degradationsmedien kultiviert wurden, zeigte sich bei allen eine leichte Erhöhung der CD11c/CD86-positiven Zellen im Vergleich zur Negativkontrolle. In Bezug auf die LPS-stimulierten DC ist die Steigerung allerdings gering. - 67 - Ergebnisse b) a) CD86 6,1 16,6 % % CD11c c) d) 6,5 e) % 4,6 % 6,1 % Abb. 4.2.5.-a: Expression von CD11c und CD86 von DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l MgCl2 ab Tag 3 der DC-Kultivierung: a) unbehandelte Kontrolle, b) PositivKontrolle (LPS-Stimulation) c) 0,1 mmol/l MgCl2, d) 1,0 mmol/l MgCl2, e) 10 mmol/l MgCl2. Als Positivkontrolle wurden DC am Tag 9 der Kultivierung mit 1 µg/ml LPS stimuliert. Der Quadrant G 2 zeigt den Prozentsatz CD11c/CD86-positiven DC. Es wurde keine Steigerung der CD11c/CD86positven Zellen festgestellt. Dargestellt ist ein repräsentatives Ergebnis aus 3 unabhängigen Versuchen. Pro Ansatz wurden 5000 Zellen gemessen. Einzelwerte sind im Anhang Tabelle 9.20. dargestellt. Tabelle 10: Zusammenfassende Darstellung der CD11c/CD86-positiven DC Probe Annexin V-FITC a) Unbehandelte Kontrolle 6,1 % b) LPS (Positiv-Kontrolle) 16,6 % c) MgCl2 0,1 mmol/l 6,5 % d) MgCl2 1,0 mmol/l 4,6 % e) MgCl2 10 mmol/l 6,1 % - 68 - Ergebnisse CD86 a) b) 6,1 % 16,6 % CD11c c) e) d) 5,9 % 5,9 % 7,6 % Abb. 4.2.5.-b: Expression von CD11c und CD86 von DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l CaCl2 ab Tag 3 der DC-Kultivierung: a) unbehandelte Kontrolle, b) Positivkontrolle (LPS-Stimulation) c) 0,1 mmol/l CaCl2, d) 1,0 mmol/l CaCl2, e) 10 mmol/l CaCl2. Als Positiv-Kontrolle wurden DC am Tag 9 der Kultivierung mit 1 µg/ml LPS stimuliert. Der Quadrant G 2 zeigt den Prozentsatz CD11c/CD86-positiven DC. Es wurde keine Steigerung der CD11c/CD86positven Zellen festgestellt. Dargestellt ist ein repräsentatives Ergebnis aus 3 unabhängigen Versuchen. Pro Ansatz wurden 5000 Zellen gemessen. Einzelwerte sind im Anhang Tabelle 9.21. dargestellt. Tabelle 11: Zusammenfassende Darstellung der CD11c/CD86-positiven DC Probe Annexin V-FITC a) Unbehandelte Kontrolle 6,1 % b) LPS (Positiv-Kontrolle) 16,6 % c) CaCl2 0,1 mmol/l 5,9 % d) CaCl2 1,0 mmol/l 5,9 % e) CaCl2 10 mmol/l 7,6 % - 69 - Ergebnisse CD 86 a) b) f) 8,4 % 7,4 % CD 11c d) c) 9,2 e) % 9,2 % 6,7 % Abb. 4.2.5.-c: Expression von CD11c und CD86 der DC, die in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 ab Tag 3 der DCKultivierung kultiviert wurden: a) Mg(pur), b) MgCa0.6, c) MgCa0.8, d) MgCa1.0 e) MgCa1.2. Der Quadrant G 2 zeigt den Prozentsatz CD11c/CD86-positiven DC. Es wurde keine Steigerung der CD11c/CD86-positven Zellen festgestellt. Dargestellt ist ein repräsentatives Ergebnis aus 3 unabhängigen Versuchen. Pro Ansatz wurden 5000 Zellen gemessen. Einzelwerte sind im Anhang Tabelle 9.22. dargestellt. Tabelle 12: Zusammenfassende Darstellung der CD11c/CD86-positiven DC Probe Annexin V-FITC a) Mg(pur) 8,4 % b) MgCa0.6 7,4 % c) MgCa0.8 9,2 % d) MgCa1.0 9,2 % e) MgCa1.2 6,7 % - 70 - Ergebnisse 4.2.6. T-Zellproliferation in der MLR Mit Hilfe der heterologen MLR (Mixed Leukocyte Reaction) wurde die Fähigkeit der unterschiedlich behandelten DC, T-Lymphozyten zur Proliferation anzuregen, untersucht. Es wurden drei unterschiedliche Versuchsansätze untersucht. Zuerst wurden die DC mit 0,1 mmol/l, 1,0 mmol/ und 10 mmol/l MgCl2 ab Tag 3 der DC-Kultivierung inkubiert. Im zweiten Ansatz wurden die DC mit 0,1 mmol/l, 1,0 mmol/ und 10 mmol/l CaCl2 inkubiert und zuletzt wurden die DC in den Degradationsmedien von Mg(pur) und den Legierungen MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 kultiviert. Bei allen Versuchsansätzen wurde eine unbehandelte Kontrolle als Negativ-Kontrolle verwendet. Diese sind T-Lymphozyten, die mit unter Standardbedingungen kultivierten DC inkubiert wurden. Bei der Darstellung der Ergebnisse handelt es sich um Histogramme von repräsentativen Ergebnissen aus jeweils 3 Versuchen. In der Abbildung 4.2.6.-a sind Histogramme von T-Lymphozyten, die mit MgCl2-behandelten DC inkubiert wurden. Die Tabelle 13 zeigt eine zusammenfassende Darstellung der prozentualen T-Zellproliferation in der MgCl2-Gruppe. Bei allen drei MgCl2-Konzentrationen zeigte sich eine geringe Steigerung der T-Zellproliferation im Vergleich zur unbehandelten Kontrolle. Die Abbildung 4.2.6.-b zeigt die Histogramme der T-Lymphozyten nach Inkubation mit CaCl2-behandelten DC. Die Tabelle 14 zeigt eine zusammenfassende Darstellung der prozentualen T-Zellproliferation in der CaCl2-Gruppe Bei einer Konzentration von 10 mmol/l CaCl2 zeigte sich eine starke Erhöhung der TZellproliferation im Vergleich zur Kontrolle. In der Abbildung 4.2.6.-c sind die Histogramme der T-Lymphozyten nach Inkubation mit DC, die in den Degradationsmedien kultiviert wurden. Die Tabelle 15 zeigt eine zusammenfassende Darstellung der prozentualen T-Zellproliferation. Bei den Degradationsmedien, speziell bei Mg(pur) und MgCa1.2, zeigte sich eine leichte Erhöhung der Proliferationsrate im Vergleich zur unbehandelten Kontrolle. - 71 - Ergebnisse Zellzahl a) Intensität CFSE b) c) d) Abb. 4.2.6.-a: Histogramme von CFSE markierten T-Lymphozyten nach Ko-Inkubation mit MgCl2-behandelten DC: a) unbehandelte Kontrolle, b) MgCl2 0,1 mmol/l, c) MgCl2 1,0 mmol/l, d) MgCl2 10 mmol/l. Im Histogramm-Plot ist der Prozentsatz der proliferierten T-Lymphozyten angegeben. Im Vergleich zur Kontrolle zeigt sich eine minimale Steigerung der T-Zellproliferation. Pro Ansatz wurden 10.000 T-Lymphozyten gemessen. Repräsentatives Ergebnis aus 3 unabhängigen Versuchen. Einzelwerte sind im Anhang Tabelle 9.23. dargestellt. Tabelle 13: Zusammenfassende Darstellung der prozentualen T-Zellproliferation Probe Annexin V-FITC a) Unbehandelte Kontrolle 48,7 % b) MgCl2 0,1 mmol/l 61,6 % c) MgCl2 1,0 mmol/l 61,1 % d) MgCl2 10 mmol/l 53,8 % - 72 - Ergebnisse a) Zellzahl Intensität CFSE b) d) c) Abb. 4.2.6.-b: Histogramme von CFSE markierten T-Lymphozyten nach Ko-Inkubation mit CaCl2-behandelten DC: a) unbehandelte Kontrolle, b) CaCl2 0,1 mmol/l, c) CaCl2 1,0 mmol/l, d) CaCl2 10 mmol/l. Im Histogramm-Plot ist der Prozentsatz der proliferierten T-Lymphozyten angegeben. Starke Erhöhung der T-Zellproliferation bei einer Konz. von 10 mmol/l CaCl2. Pro Ansatz wurden 10.000 T-Lymphozyten gemessen. Repräsentatives Ergebnis aus 3 unabhängigen Versuchen. Einzelwerte sind im Anhang Tabelle 9.24. dargestellt. Tabelle 14: Zusammenfassende Darstellung der prozentualen T-Zellproliferation Probe Annexin V-FITC a) Unbehandelte Kontrolle 48,7 % b) CaCl2 0,1 mmol/l 55,5 % c) CaCl2 1,0 mmol/l 59,1 % d) CaCl2 10 mmol/l 77,5 % - 73 - Ergebnisse a) Zellzahl Intensität CFSE b) c) e) f) d) Abb. 4.2.6.-c: Histogramme von CFSE markierten T-Lymphozyten nach Ko-Inkubation mit in Degradationsmedien kultivierten DC: a) unbehandelte Kontrolle, b) Mg(pur), c) MgCa0.6 d) MgCa0.8, e) MgCa1.0, f) MgCa1.2. Im Histogramm-Plot ist der Prozentsatz der proliferierten T- Lymphozyten angegeben. Es zeigt sich eine minimale Erhöhung der T-Zellproliferation bei allen Degradationsmedien. Pro Ansatz wurden 10.000 T-Lymphozyten gemessen. Repräsentatives Ergebnis aus 3 unabhängigen Versuchen. Einzelwerte sind im Anhang Tabelle 9.25. dargestellt. - 74 - Ergebnisse Tabelle 15: Zusammenfassende Darstellung der prozentualen T-Zellproliferation Probe Annexin V-FITC a) unbehandelte Kontrolle 51,2 % b) Mg(pur) 58,4 % c) MgCa0.6 55,6 % d) MgCa0.8 53,5 % e) MgCa1.0 57,2 % f) MgCa1.2 58,7 % - 75 - Diskussion 5. Diskussion Ziel dieser Arbeit war es, Erkenntnisse über die Beeinflussung der Zellfunktionen von murinen DC und T-Zellen durch biokompatible, abbaubare Implantate aus Magnesium-Calcium-Legierungen zu gewinnen. Untersucht wurden eine reine Magnesiumlegierung (Mg(pur)) und vier Magnesium-Calcium-Legierungen mit steigenden Calciumkonzentrationen (MgCa0.6 mit 0,6 wt% Calcium, MgCa0.8 mit 0,8 wt% Calcium, MgCa1.0 mit 1,0 wt% Calcium und MgCa1.2 mit 1,2 wt% Calcium). Von den Legierungen wurden Degradationsmedien hergestellt, in denen die dendritischen Zellen kultiviert wurden. Parallel dazu wurden Versuche mit MgCl2 und CaCl2 in Konzentrationen von 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l durchgeführt. Als Grundlage für die Versuche mit MgCl2 und CaCl2 dienten Degradationsversuche mit Magnesiumbasislegierungen, bei denen Magnesiumkonzentrationen im Kulturmedium zwischen 0,2 mmol/l und 10 mmol/l gemessen wurden. 5.1. Degradationsversuche der Magnesium-Calcium-Legierungen Bei den Versuchen zeigte sich, dass das Korrosionsverhalten der Legierungen mit geringen wt% Calcium stabiler war als bei den Legierungen MgCa1.0 und MgCa1.2. Gerade die Legierung MgCa1.0 zeigte eine große Varianz bezüglich der gemessenen Magnesiumkonzentrationen im Degradationsmedium (Abb. 4.1.-a). Die Magnesiumkonzentrationen in den Degradationsmedien der Legierung MgCa1.0 variierten zwischen 0,9 mmol/l und 3,5 mmol/l. Gründe für die unterschiedlichen Degradationsgeschwindigkeiten der Legierung MgCa1.0 könnten unter anderen unterschiedliche Calciumkonzentrationen innerhalb einer Legierungscharge sein. So wurden bei Legierungen, die schneller degradierten, im Institut für Werkstoffkunde (Leibnitz Universität Hannover) tendenziell höhere Calciumkonzentrationen festgestellt als bei Legierungen, die stabiler degradierten (mündliche Mitteilung C. Krause, Institut für Werkstoffkunde der Leibniz Universität Hannover). Dieses Ergebnis zeigt, dass bei der Herstellung der Magnesium-Calcium-Legierungen mit höheren Calciumkonzentrationen eine - 76 - Variabilität bezüglich der Diskussion Calciumkonzentrationen innerhalb einer Charge besteht. Eine schnellere Degradation von Legierungen mit höheren Calciumkonzentrationen zeigte sich ebenfalls bei den In-vitro-Degradationsversuchen von DENKENA et al. (2005, 2006). Für den Degradationsversuch wurden von DENKENA Magnesium-Calcium- Legierungen mit steigendem wt% Calcium (MgCa0.4 mit 0,4 wt% Ca, MgCa0.8 mit 0,8 wt% Ca und MgCa2.0 mit 2,0 wt% Ca) für 72 Stunden einer 5%igen NaCl2Lösung ausgesetzt. Die Auswertung zeigte, dass bis zu einem Calciumanteil von 0,8 wt% die Legierungen gleichmäßig korrodierten. Bei den Legierungen mit höheren wt% Calcium verlief die Degradation stärker und inhomogener (DENKENA et al., 2005, 2006). VON DER HÖH (2008) untersuchte ebenfalls das Degradationsverhalten von Magnesium-Calcium-Legierungen, allerdings im In-vivoModell. Zum Einsatz kamen vier verschiedene Magnesium-Calcium-Legierungen (MgCa0.4 mit 0,4 wt% Ca, MgCa0.8 mit 0,8 wt% Ca, MgCa1.2 mit 1,2 wt% Ca und MgCa2.0 mit 2,0 wt% Ca). Dabei zeigte sich eine genaue gegensätzliche Degradation im Vergleich zu DENKENA et al. (2005, 2006). WITTE et al. (2006) beschrieb ebenfalls die gegensätzlichen Degradationseigenschaften von Magnesiumlegierungen in In-vitro- und In-vivo-Degradationsversuchen, sodass die tatsächlichen Degradationseigenschaften von Magnesiumlegierungen wahrscheinlich nur im Gewebe untersucht werden können. Aus diesem Grund würde sich das isoliert perfundierte Rindereuter für Degradationsversuche im Gewebe anbieten (BÄUMER und KIETZMANN, 2000). Dabei werden die Implantate in Hauttaschen des Rindereuters für eine Zeitspanne von 6-8 Stunden implantiert. Um die Implantate herum würden mit Hilfe eines Mikrodialyseschlauches freigesetzte Ionen herausgespült und quantifiziert werden. Durch die photometrische Messung der Kalibrationsstandards für CaCl2 zeigte sich, dass erst ab einer CaCl2-Konzentration von 0,2 mmol/l die optische Dichte erfassbar und eindeutig zugeordnet werden konnte. Die Calciumkonzentration in den Degradationsmedien war nach 72-stündiger Inkubation im Wasserbad unterhalb der Nachweisgrenze von 0,2 mmol/l. - 77 - Diskussion 5.2. DC-Zellzahl, MØ-Zellzahl und Vitalität der DC nach MgCl2- und CaCl2-Inkubation bzw. nach Kultivierung in den Degradationsmedien Die Kultivierung der DC-Vorläuferzellen zeigte, dass mit steigender Mg2+Konzentration die Zellzahl der DC nach sechstägiger Kultivierung signifikant abnimmt (Abb. 4.2.1.-a). Das gleiche Bild zeigten ebenfalls DC, die in steigenden Ca2+Konzentrationen kultiviert wurden. Auch hier nahm die Zellzahl der DC im Vergleich zur unbehandelten Kontrolle signifikant ab (Abb. 4.2.1.-c). Bei den DC, die in den Degradationsmedien der Legierungen MgCa1.0 und MgCa1.2 kultiviert wurden, zeigte sich ebenfalls ein schwach signifikanter Rückgang der DC-Zellzahl (Abb. 4.2.1.-e). Dieses Ergebnis lässt vermuten, dass Magnesium und Calcium in steigenden Konzentrationen und Magnesium-Calcium-Legierungen eventuell einen zytotoxischen oder apoptotischen Effekt auf DC haben könnten. Aus diesem Grund wurden die Vitalität der DC sowie die Zellfraktion der Makrophagen untersucht. Die Untersuchung der Vitalität der DC mittels FACS-Analyse und XTT-Test (Kapitel: 3.2.8.1.) zeigte, dass sowohl Magnesium als auch die Degradationsmedien keinen Einfluss auf die Vitalität der DC haben (Abb. 4.2.2.-a,-c,-d,-f). Bei den DC, die in dem Degradationsmedium von Mg(pur) kultiviert wurden, war der Anteil an apoptotischen Zellen leicht erhöht. Im Vergleich zur Positivkontrolle (Etoposide) war der Anstieg allerdings sehr gering (Abb. 4.2.2.-c). Der Anteil an nekrotischen Zellen war bei allen Versuchen im Vergleich zur Kontrolle nicht verändert. LI et al. (2008) untersuchten ebenfalls den Einfluss von Degradationsmedien von Magnesium-Calcium- Legierungen auf die Vitalität von murinen Fibroblasten (L929-Zellen). Dafür wurden die L929-Zellen in Dulbecco`s Modified Eagle`s Medium (DMEM) kultiviert. Ab Tag 1 der Kultivierung wurden die L929-Zellen mit unterschiedlichen Verdünnungsstufen (0 %, 50 %, 90 %) des Degradationsmediums der Legierung MgCa1 inkubiert. Am Tag 2, 4 und 7 wurde die Vitalität der Zellen mittels MTT bestimmt. Die Auswertung zeigte, dass MgCa1 keinen zytotoxischen Effekt auf L929-Zellen hat. Darüber hinaus schien die Vitalität der in unverdünnten Degradationsmedium kultivierten L929-Zellen besser zu sein als die Vitalität von L929-Zellen in der Kontrolle (LI et al., 2008). - 78 - Diskussion Dieses Phänomen konnte für DC nicht beobachtet werden. Allerdings zeigte sich bei der Untersuchung der Makrophagenpopulation (Kapitel: 4.2.1.), dass mit steigender Magnesiumkonzentration die Ausdifferenzierung der Makrophagen aus DC- Vorläuferzellen signifikant zunahm (Abb. 4.2.1.-b). Ebenso stieg der Anteil der Makrophagen bei der Kultivierung der DC-Vorläuferzellen im Degradationsmedium der Legierung MgCa1.0 signifikant an (Abb. 4.2.1.-d). Es scheint also, dass neben L929-Zellen auch Makrophagen die Anwesenheit von Magnesium favorisieren. CAICEDO et al. (2008) zeigten bei humanen Makrophagen, die mit metallischen Salzen und Legierungen inkubiert wurden, eine Aktivierung und eine Erhöhung der IL-1β-Sekretion. Zum Einsatz kamen Cobalt (Co), Chrom (Cr) und Molybdän (Mo)Salze sowie Co-Cr-Mo-Legierungen (CAICEDO et al., 2008). Ob Magnesium ebenfalls in höheren Konzentrationen zu einer Aktivierung von Makrophagen führte, wurde nicht untersucht. Im Gegensatz zu Magnesium zeigte Calcium bei einer Konzentration von 10 mmol/l einen signifikanten Rückgang der Vitalität sowohl in der FACS-Analyse als auch im XTT-Test. Dieses Ergebnis scheint allerdings in Bezug auf Magnesium-CalciumLegierungen keine (patho)physiologische Relevanz zu haben, da Calciumkonzentrationen von 10 mmol/l in den Degradationsmedien der Legierungen nicht erreicht wurden. 5.3. TNF-α-Sekretion der DC nach MgCl2- und CaCl2-Inkubation bzw. nach Kultivierung in den Degradationsmedien Die alleinige Inkubation der DC mit MgCl2 und CaCl2 zeigte, unabhängig der Konzentration, keine signifikante Steigerung der TNF-α-Freisetzung im Vergleich zur unbehandelten Kontrolle (Abb. 4.2.3.-a und -b). Im Mittel allerdings zeigten die DC, die mit MgCl2 und CaCl2 inkubiert wurden, eine höhere TNF-α-Sekretion als die unbehandelte Kontrolle. Die tendenzielle Steigerung ist nicht signifikant. Eine signifikant erhöhte Freisetzung von TNF-α konnte nur bei LPS-stimulierten DC beobachtet werden. Bei den DC, die in den Degradationsmedien der MagnesiumCalcium-Legierungen inkubiert wurden, zeigte sich ebenfalls im Mittel eine - 79 - Diskussion tendenzielle Steigerung der TNF-α-Freisetzung, die bei den Degradationsmedien der Legierungen MgCa1.0 und MgCa1.2 am größten war. Auch hier war die tendenzielle Steigerung nicht signifikant. Im Vergleich zur Positivkontrolle mit LPS war die TNF-αFreisetzung allerdings gering. Das proinflammatorische Zytokin TNF-α spielt eine wichtige Rolle bei der Aktivierung und Maturation, sowie der Expression diverser Oberflächenmoleküle von DC (AIBA 1998). Neben den DC sind auch aktivierte Makrophagen in der Lage TNF-α zu produzieren um im Entzündungsgeschehen weitere Zellen zu mobilisieren (SPIEKSTRA et al., 2005). Wie zuvor beschrieben, ist bei der Kultivierung der DCVorläuferzellen, gerade bei MgCl2, CaCl2 und den Degradationsmedien, der Anteil der Makrophagen signifikant gestiegen. Da Makrophagen ebenfalls in der Lage sind TNF-α zu produzieren, könnte die leichte Erhöhung von TNF-α auch von den Makrophagen stammen. 5.4. Auswanderungsverhalten von DC nach MgCl2- und CaCl2Inkubation bzw. nach Kultivierung in den Degradationsmedien In einem Migrationsassay wurde das Auswanderungsverhalten von DC nach Inkubation mit MgCl2, CaCl2 und den Degradationsmedien untersucht. Als PositivKontrolle wurde in Anlehnung an HÜNERMANN (2006) Bleichlorid (100 µmol/l) verwendet. Ein signifikanter Anstieg der Migrationsrate der DC konnte nach Inkubation mit MgCl2 bzw. CaCl2 in der jeweils höchsten Konzentration (10 mmol/l) im Vergleich zur Kontrolle gezeigt werden. Bei den DC, die im Degradationsmedium der Legierung MgCa1.2 kultiviert wurden, zeigte sich ebenfalls ein schwach signifikanter Anstieg der Migrationsrate. Normalerweise wird in einem Migrationsassay die Migrationsbereitschaft der DC auf ein Chemokin (z.B. MIP-3β) untersucht. Bei den durchgeführten Versuchen wurde auf diesen zusätzlichen Stimulus verzichtet, da bei früheren Untersuchungen im Institut der Pharmakologie, Toxikologie und Pharmazie (Tierärztliche Hochschule Hannover) gezeigt werden konnte, dass der Einsatz von Chemokinen im Vergleich zu keiner zusätzlichen Stimulation zu keinem qualitativen Unterschied bezüglich der Migrationsrate der DC - 80 - Diskussion führte. Voraussetzung für die DC-Migration ist die Auslösung ihrer Maturation. Durch die Aufnahme von Antigen kommt es auf Seiten der DC zur Veränderung des DCPhänotyps einhergehend mit einer verstärkten Expression costimulatorischer Moleküle (CD80, CD86, CD40 und MHCII) und Ausschüttung unterschiedlicher Zytokine (z.B. TNF-α, IL-6, IL-12) (AIBA, 1998). Aufgrund der eigenen Untersuchung wurde gezeigt, dass MgCl2 und CaCl2 in hohen Konzentrationen einen Stimulus auf die DC darstellten. Dadurch wurde die Maturation, in Form einer gesteigerten Migrationsbereitschaft, der DC beeinflusst. Die geringe Migration der DC, die mit niedrigen Konzentrationen von MgCl2 und CaCl2 inkubiert wurden, ist als Spontanmigration zu deuten und ist Ausdruck der physiologischen steady state migration. Spontanmigration zeigten ebenfalls DC, die in den Degradationsmedien inkubiert wurden. Nur die DC, die im Degradationsmedium der Legierung MgCa1.2 kultiviert wurden, zeigten eine schwach signifikante Erhöhung der Migrationsbereitschaft. Für eine (patho)physiologische Relevanz müsste die Legierung MgCa1.2 in weiteren Migrationsversuchen im Gewebe untersucht werden. 5.5. Expression von CD11c/CD86 auf DC nach MgCl2- und CaCl2Inkubation bzw. nach Kultivierung in den Degradationsmedien Um eine mögliche Aktivierung der unterschiedlich behandelten DC weiter zu untersuchen, wurden die DC auf die Expression von CD11c/CD86 untersucht. Die Expression von Charakteristisch CD86 für ist reife Ausdruck DC einer (mature stattgefundenen DC) ist die DC-Aktivierung. Expression des Zelloberflächenproteins CD11c, welches in der Lage ist, eine Adhäsion zwischen Zellen bei der Immunantwort zu ermöglichen (LAMBRECHT, 2001). LPS-stimulierte DC wurden bei diesem Versuch als Positiv-Kontrolle verwendet, da LPS eine Aktivierung und somit eine Reifung von immaturen DC in Richtung maturen DC bewirkt (AIBA, 1998). Eigene Untersuchungen zeigten, dass es bei DC, die mit unterschiedlichen Konzentrationen von MgCl2 oder CaCl2 inkubiert wurden, nicht zu einer Steigerung - 81 - Diskussion der Expression von CD11c/CD86 kommt (Abb. 4.2.5.-a). Vergleichbare Studien mit metallischen Salzen wurden von MANOME et al. (1999) durchgeführt. Hierbei kam es bei DC, die mit NiCl2, MnCl2 und MoCl2 inkubiert wurden, zu einer Hochregulation von CD86. Dies ging einher mit einer gesteigerten Produktion der Zytokine TNF-α, IL-1β und IL-12. In Anbetracht eigener TNF-α-Messungen, bei denen kein signifikanter Anstieg festzustellen war (Kapitel: 4.2.3.), liegt die Vermutung nahe, dass die CD86-Expression in Abhängigkeit von einem bestimmten Zytokinmilieu erfolgt. AIBA (1998) zeigte in diesem Zusammenhang die Notwendigkeit vom Vorhandensein der Substanzen TNF-α, GM-CSF und IL-1β für die CD86-Expression, indem die DC mit o.g. Zytokinen stimuliert und die Expression CD86 auf den Zellen untersucht wurden. In der Analyse der DC, die in den Degradationsmedien der Magnesium-Calcium-Legierungen kultiviert wurden, zeigte sich ebenfalls, dass der Anteil an CD11c/CD86 positiven DC auf dem Niveau der unbehandelten Kontrolle blieb (Abb. 4.2.5.-c). 5.6. Mixed Leukocyte Reaction mit DC, nach Präinkubation mit MgCl2 und CaCl2 bzw. nach Kultivierung in den Degradationsmedien Bei den Versuchen mit MgCl2 präinkubierten DC zeigte sich bei der Mixed Leukocyte Reaction (MLR) eine leichte Erhöhung der T-Zellproliferation im Vergleich zur unbehandelten Kontrolle (Abb. 4.2.6.-a). Ebenfalls zeigte sich bei den Versuchen mit DC, die in den verschiedenen Degradationsmedien kultiviert wurden, eine leichte Erhöhung der T-Zellproliferation (Abb. 4.2.6.-c). Vergleicht man allerdings diese TZellproliferationen mit den Ergebnissen aus der CaCl2-Gruppe, zeigte sich nur eine geringgradige Steigerung. Besonders für die T-Zellen, die mit 10 mmol/l CaCl2 präinkubierten DC gepaart wurden, zeigte sich eine starke Erhöhung der TZellproliferation (Abb. 4.2.6.-b). MANOME et al. (1999) beschrieben einen Zusammenhang zwischen einer erhöhten CD86-Expression von DC verbunden mit einer gesteigerten T-Zellproliferation in der MLR. In eigenen Untersuchungen konnte dieses jedoch nicht bestätigt werden, da die Expression von CD11c/CD86 bei allen Versuchsreihen auf dem Niveau der unbehandelten Kontrolle blieb (Kapitel: 4.2.5.). - 82 - Diskussion Neben maturen (CD11c/CD86 positiv) DC sind auch semimature DC in der Lage eine T-Zellantwort zu initiieren. Im Vergleich zu maturen DC fehlt ihnen die Fähigkeit proinflammatorische Zytokine (z.B. TNF-α) auszuschütten. Aufgrund dieser Tatsache sind sie in der Lage beispielsweise apoptotisches Zellmaterial zu den sekundären Lymphorganen zu transportieren und über dort ansässige T-Zellen eine Toleranz zu vermitteln (LUTZ und SCHULER 2002; STEINMAN et al., 2003a). Dies geschieht durch Reifung unreifer T-Zellen zu IL-10 produzierenden Supressor-Zellen (JONULEIT et al., 2000; DHODAPKAR et al., 2001). Sie spielen also eine wichtige Rolle in der Immuntoleranz. Verantwortlich für eine gesteigerte T-Zellproliferation könnten neben TNF-α auch andere Zytokine sein (z.B.: IL-2). LI et al. (2008) führten In-vivo-Versuche mit Magnesium-Calcium-Implantaten durch. Dabei untersuchten sie histologisch das Gewebe um die Implantationsstelle. Zwei Monate nach Implantation konnten sie einige Lymphozyten identifizieren, jedoch gab es keine Anzeichen auf eine erhöhte Immunreaktion. VON DER HÖH (2008) untersuchte ebenfalls die Biokompatibilität Kaninchenmodell. verschiedener Für den Magnesium-Calcium-Legierungen Versuch wurden aus vier im Magnesium-Calcium- Legierungen (MgCa0.4 mit 0,4 wt% Ca, MgCa0.8 mit 0,8 wt% Ca, MgCa1.2 mit 1,2 wt% Ca und MgCa2.0 mit 2,0 wt% Ca) Implantate hergestellt, mit drei verschiedenen Oberflächen (glatt, rau und gestrahlt). Zur Erstellung einer rauen Implantatoberfläche wurden die Implantate durch einen nachgelagerten Schleifprozess bearbeitet. Die gestrahlten Implantatoberflächen wurden durch Partikelbestrahlung von glatten Implantaten erzeugt (Partikeldurchmesser 300-400 µm, Strahlzeit 30 s). Anschließend wurden die Implantate für sechs Wochen in die medialen Femurkondylen beider Hintergliedmaßen eingesetzt. Unabhängig von den Calciumkonzentrationen in den Implantaten oder der Oberflächenstruktur zeigten die Implantate eine gute Verträglichkeit. Bei histologischen Untersuchungen dominierten bei allen Implantaten die Makrophagen. Dieser Befund deutet ebenfalls nicht auf eine erhöhte Abwehrreaktion hin. Nur bei gestrahlten Implantaten zeigten sich neben Makrophagen zusätzlich Fremdkörperzellen, Plasmazellen und Lymphozyten. Als Grund vermutete VON DER HÖH (2008) eine schnellere Degradation von gestrahlten Magnesium-Calcium-Legierungen. - 83 - Diskussion Zusammenfassend erwiesen sich bei der Herstellung der Degradationsmedien die Legierungen bis zu 0,8 wt% Calcium am stabilsten. Die Legierungen mit 1,0 und 1,2 wt% Calcium zeigten stärkere Degradation. Unabhängig von der Calciumkonzentration in den Legierungen konnte allerdings keine zytotoxische und apoptotische Wirkung auf DC festgestellt werden. Auch zeigte die Produktion des Zytokins TNF-α keinen signifikanten Unterschied zur unbehandelten Kontrolle. Bei der Untersuchung der Funktion von DC im Migrationsassay konnte eine leichte Erhöhung der Migrationsbereitschaft im Degradationsmedium der Legierung MgCa1.2 festgestellt werden. Auf die Expression von CD11c und CD86 hatten die Degradationsmedien der Legierungen ebenfalls keinen Einfluss. Nur in der MLR zeigte sich bei den T-Zellen ein geringer Anstieg der Proliferationsrate. Im Vergleich zu den T-Zellen, die mit 10 mmol/l CaCl2 behandelten DC inkubiert wurden, war der Anstieg der Proliferationsrate allerdings gering. Aufgrund der Ergebnisse scheinen Magnesium-Calcium-Legierungen bis zu 1,0 % Calcium aus immunotoxikologischer Sicht als unbedenklich, da sie keinen Einfluss auf die Funktion von murinen DC haben. Bei MgCa1.2 müssten im Bereich der MLR (Zytokinbestimmung in Überständen der MLR), und der Migration Untersuchungen durchgeführt werden. - 84 - (In-vivo-Migration) weitere Zusammenfassung 6. Zusammenfassung Klaas Feser Immunotoxikologische Untersuchung zur Biokompatibilität von abbaubaren Implantaten aus Magnesiumbasislegierungen Abbaubare Magnesiumbasislegierungen sind neue Materialien, die als Implantate in der Orthopädie und in der Weichteilchirurgie eingesetzt werden. In der vorliegenden Arbeit ist die Beeinflussung der Zellfunktionen muriner DC durch abbaubare Magnesiumbasislegierungen untersucht worden, da die DC im Organismus eine wichtige Rolle als antigenpräsentierende Zellen übernehmen. Für die Untersuchung wurden murine dendritische Zellen in Degradationsmedien aus reinem Magnesium (Mg(pur)) und aus Magnesium-Calcium-Legierungen mit steigendem wt% Calcium (0,6 %, 0,8 %, 1,0 % und 1,2%) kultiviert. In einem zweiten Versuchsansatz wurden murine DC mit MgCl2 und CaCl2 in steigenden Konzentrationen (0,1 mmol/l – 10 mmol/l) inkubiert. Die Kultivierung der DC über sechs Tage in den Degradationsmedien zeigte keine Beeinflussung der Vitalität sowohl im XTT-Test als auch im Annexin V-FITC Assay. Ebenso waren die Produktion von TNF-α und die Expression von CD86 nicht beeinflusst durch die Kultivierung der DC in den Degradationsmedien. Im Migrationsassay konnte gezeigt werden, dass die Migrationsrate der DC, die im Degradationsmedium der Legierung MgCa1.2 kultiviert wurden, leicht anstieg. Weiterhin zeigte sich in der Mixed Leukocyte Reaction bei allen Degradationsmedien nur eine maximal schwache Steigerung der Proliferationsrate. Bei den DC, die mit MgCl2 und CaCl2 in steigenden Konzentrationen inkubiert wurden, zeigte sich bei 10 mmol/l CaCl2 ein signifikanter Rückgang der Vitalität, sowie eine Steigerung der Migrationsrate und der TZellproliferation. Diese Ergebnisse haben jedoch wahrscheinlich keine (patho)physiologische Relevanz, da bei der Degradation der Legierungen solch hohe Konzentrationen nicht erreicht werden. Zusammenfassend zeigte sich im In-vitro- - 85 - Zusammenfassung Versuch mit murinen DC, dass Magnesium-Calcium-Legierungen bis zu 1,0 wt% Calcium aus immunotoxikologischer Sicht keinen Einfluss auf die Vitalität und Funktion von murinen DC haben. - 86 - Summary 7. Summary Klaas Feser Immunotoxicological studies on the biocompatibility of degradable implants from magnesium-based alloys Degrading magnesium alloys are new materials for implants used in orthopaedic and trauma surgery. The aim of this study was to investigate the influence of degradable magnesium alloys on the function of dendritic cells as these cells represent the major antigen presenting cells of the body. At first degradation media were made out of the alloys Mg(pur) (pure magnesium), MgCa0.6 (0,6 wt% Ca), MgCa0.8 (0,8 wt% Ca), MgCa1.0 (1,0 wt% Ca) und MgCa1.2 (1,2 wt% Ca). In parallel, murine bone marrow derived dendritic cells were incubated with increasing concentrations (0,1 mmol/l – 10 mmol/l) of magnesium chloride and calcium chloride, respectively. Incubation of DC with degradation media over 6 days had no influence on cell viability (XTT and Annexin V-FITC assay). Additionally, the production of TNFα and the expression of CD86 were not enhanced by incubation with degraded magnesium alloys. Only the DC migration was marginal influenced by degradation media of the alloy MgCa1.2. Furthermore, the T cell proliferation was marginally influenced by dendritic cells which were cultivated in degradation media. DC which were incubated with increasing concentrations of MgCl2 und CaCl2 showed only in the high CaCl2 concentration (10 mmol/l) an enhanced apoptosis and an increase of DC migration and T cell proliferation. This result is probably not of (patho)physiologically relevance, as these concentrations will not be achieved with degradation of magnesium alloys. Taken together magnesium-calcium-alloys up to 1,0 wt% calcium had no immunotoxicological effects on murine bone marrow derived dendritic cells. - 87 - Literaturverzeichnis 8. Literaturverzeichnis ABELN, I. (2006) Allergietestung von degradablen metallischen Magnesiumlegierungen als orthopädisches Implantatmaterial Hannover, Tierärztliche Hochschule, Diss. AIBA, S. (1998) Maturation of dendritic cells induced by cytokines and haptens Tohoku J. Exp. Med. 184, 159-172 AKBARI, O., N. PANJWANI, S. GARCIA, R. TASCON, D. LOWRIE und B. STOCKINGER (1999) DNA vaccination: transfection and activation of dendritic cells as key events for immunity J. Exp. Med. 189, 169-178 ALBERT, M. L., S. F. PEARCE, L. M. FRANCISCO, B. SAUTER, P. ROY, R. L. SILVERSTEIN und N. BHARDWAJ (1998) Immature dendritic cells phagocytose apoptotic cells via alphavbeta5 and CD36, and crosspresent antigens to cytotoxic T lymphocytes J. Exp. Med. 188, 1359-1368 AL-GHAMDI, S. M., E. C. CAMERON und R. A. SUTTON (1994) Magnesium deficiency: pathophysiologic and clinical overview. Am. J. Kidney Dis. 24, 737-752 - 88 - Literaturverzeichnis BACH, F. -W., T. HASSEL, A. GOLOVKO, C. HACKENBROICH und A. MEYERLINDENBERG (2005) Resorbierbare Implantate aus Magnesium durch Mikrolegieren mit Calcium, deren Verarbeitung und Eigenschaften Biomaterialien 6, 163 BÄUMER, W. und M. KIETZMANN (2000) Isolated perfused bovine udder as a moddel of dermal eicosanoid release Alternatives to Laboratory Animals 28, 643-649 BIESALSKI, H. K., P. FÜRST, H. KASPER, R. KLUTHE, W. PÖLERT, CH. PUCHSTEIN und H. STÄHELIN (1999) Ernährungsmedizin. 163-164, 170-172 Georg Thieme Verlag, Stuttgart BIESALSKI, H. K., J. KÖHRLE und K. SCHÜMANN (2002) Vitamine, Spurenelemente und Mineralstoffe. 132-137 Georg Thieme Verlag, Stuttgart BRANNAN, P. G., P. VERGNE-MARINI, C. Y. PAK, A. R. HULL und J. S. FORDTRAN (1976) Magnesium absorption in the human small intestine. Results in normal subjects, patients with chronic renal disease, and patients with absorptive hypercalciuria J. Clin. Invest. 57, 1412-1418. - 89 - Literaturverzeichnis BROCK, T. D., M. T. MADIGAN, J. M. MARTINKO und J. PARKER (2001) Klinische und diagnostische Mikrobiologie und Immunologie S. 978 f. Zellbiologie S. 82 f. Gentechnik und Biotechnologie S. 400 f. Prokaryotische Vielfalt: Bacteria S. 575 f. in: Mikrobiologie 9. Auflage Spektrum Akademischer Verlag, Heidelberg, Berlin CAICEDO, M. S., R. DESAI, K. MCALLISTER, A. REDDY, J. J. JACOBS und N. J. HALLAB (2008) Soluble and particulate Co-Cr-Mo-alloy implant metals activate the inflammasome danger signaling pathway in human macrophages: A novel mechanism for implant debris reactivity. J. Orthopaed. Res. 27, 847-854 CAUX, C., S. AIT-YAHIA, K. CHEMIN, O. DE BOUTEILLER, M. C. DIEU-NOSJEAN, B. HOMEY, C. MASSACRIER, B. VANBERVLIET, A. ZLOTNIK und A. VICARI (2000) Dendritic cell biology and regulation of dendritic cell trafficking by chemokines. Springer Semin. Immunopathol. 22, 345-369 CELLA, M., M. SALIO, Y. SAKAKIBARA, H. LANGNEN, I. JULKUNEN und A. LANZAVECCHIA (1999) Maturation, activation, and protection of dendritic cells induced by double-stranded RNA J. Exp. Med. 189, 821-829 CLAES, L. E. (1992) Mechanical characterization of biodegradable implants Clin. Mater. 10, 41-46 - 90 - Literaturverzeichnis CUMBERBATCH, M. und I. KIMBER (1995) Tumour necrosis factor-alpha is required for accumulation of dendritic cells in draining lymph nodes and for optimal contact sensitization Immunol. 84, 31-35 DENKENA, B., C. PODOLSKY, M. VAN DER MEER und A. LUCAS (2006) Permanent bioresorbable implants in association with the production process Annals of the German Academic Society for Production Engineering (WPG) Production Engeneering 8/1 Hannover. DENKENA, B., F. WITTE, C. PODOLSKY und A. LUCAS (2005) Degradable implants made of magnesium alloys Proc. of 5th euspen International Conference, Montpellier, France. DHODAPKAR, M. V., R. M. STEINMAN, J. KRASOVSKY, C. MUNZ und N. BHRADWAJ (2001) Antigen-specific inhibition of effector T-cell function in humans after injection of immature dendritic cells J. Exp. Med. 193, 233-238 DIEU, M. C., B. VANBERYLIET, A. VICARI, J. M. BRIDON, E. OLDHAM, S. AITYAHIA, F. BRIERE, A. ZLOTNIK, S. LEBECQUE und C. CAUX (1998) Selective recruitment of immature and mature dendritic cells by distinct chemokines expressed in different anatomic sites J. Exp. Med. 188, 373-386 DUBOIS, B., J. M. BRIDON, J. FAYETTE, C. BARTHELEMY, J. BANCHEREAU, C. CAUX und F. BRIERE (1999) Dendritic cells directly modulate B cell growth and differentiation J. Leukoc. Biol. 66, 224-230 - 91 - Literaturverzeichnis EGNER, W., J. L. MCKENZIE, S. M. SMITH, M. E. BEARD und D. N. HART (1993) Identification of potent mixed leukocyte reaction-stimulatory cells in human bone marrow: putative differentiation stage of human blood dendritic cells. J. Immunol. 150, 3043-3053 EL-RAHMAN, S. S. A. (2003) Neuropathology of aluminium toxicity in rats Pharmacol. Res. 47, 189-194 ENGERING, A. J., M. CELLA, D. FLUITSMA, M. BROCKHAUS, E. C. HOEFMIT, A. LANZAVECCHIA und J. PIETERS (1997) The mannose receptor functions as a high capacity and broad specificity antigen receptor in human dendritic cells Eur. J. Immunol. 27, 2417-2425 FANGER, N. A., K. WARDWELL, L. SHEN, T. F. TEDDER und P. M. GUYRE (1996) Type I (CD64) and type II (CD32) Fc gamma receptor-mediated phagocytosis by human blood dendritic cells J. Immunol. 157, 541-548 FATEMI, S., E. RYZEN, J. FLORES, D. B. ENDRES und R. K. RUDES (1991) Effect of experimental human magnesium depletion on parathyroid hormone secretion and 1,25-dihydroxyvitamin D metabolism J. Clin. Endocrinol. Metab. 73, 1067-1072 FAWCETT, W. J., E. J. HAXBY und D. A. MALE (1999) Magnesium: physiology and pharmacology Br. J. Anaesth. 83, 303-320 - 92 - Literaturverzeichnis FINE, K. D., C. A. SANTA ANA, J. L. PORTER und J. S. FORDTRAN (1991) Intestinal absorption of magnesium from food and supplements J. Clin. Invest. 88, 396-402. FLEET, J. C. und K. D. CASHMAN (2001) Magnesium. In: Present Knowledge in Nutrition Eight Edition. Bowman B.A., Russell R.M. (Eds.) ILSI Press, Washington , DC , 292-301 FLORIDO, M., A. S. GONCALVES und R. APPELBERG (2004) CD40 is required for the optimal induction of protective immunity to Mycobacterium avium Immunol. 111, 323-327 GAD, M., M. H. CLAESSON und A. E. PEDERSEN (2003) Dendritic cells in peripheral tolerance and immunity APMIS 111, 766-775 GEIJTENBEEK, T. B., D. S. KWON, R. TORENSMA, S. J. VAN VLIET, G. C. VAN DUIJNHOVEN, J. MIDDEL, I. L. CORNELISSEN und H. S. NOTTET (2000) DC-SIGN, a dendritic cellspecific HIV-1-binding protein that enhances trans-infection of T cells. Cell 100, 587-597 GUNZER, M., C. WEISHAUPT, L. PLANELLES und S. GRABBE (2001) Two-step negative enrichment of CD4+ and CD8+ T cells from murine spleen via nylon wool adherence and an optimized antibody cocktail. J. Immunol. Methods 258, 55-63 - 93 - Literaturverzeichnis GUTZMER, R., M. LISEWSKI, J. ZWIRNER, S. MOMMERT, C. DIESEL, M. WITTMANN, A. KAPP und T. WERFEL (2004) Human monocyte-derived dendritic cells are chemoattracted to C3a after upregulation of the C3a receptor with interferons. Immunol. 111, 435-443 HACKER, H., H. MISCHAK, T. MIETHKE, S. LIPTAY, R. SCHMID, T. SPARWASSER, K. HEEG, G. B. LIPFORD und H. WAGNER (1998) CpG-DNA-specific activation of antigen-presenting cells requires stress kinase activity and is preceded by non-specific endocytosis and endosomal maturation Embo. J. 17, 6230-6240. HEATON, F. W., (1993) Distribution and function of magnesium within the cell Birch, N.J. (Ed.), Magnesium and the Cell. Academic Press, London, S. 121-136 HÖH, N. VON DER (2008) Einfluss der Oberflächenbearbeitung von resorbierbaren Knochenimplantaten aus Magnesium-Calcium-Legierungen auf das Degradationsverhalten im Kaninchenmodell Hannover, Tierärztliche Hochschule, Diss. HÜNERMANN, K. (2006) Untersuchung zur Beeinflussung der Funktion dendritischer Zellen durch Schwermetalle Hannover, Tierärztliche Hochschule, Diss. - 94 - Literaturverzeichnis INABA, K., M. INABA, N. ROMANI, H. AYA, M. DEGUCHI, S. IKEHARA, S.MURAMATSU und R. M. STEINMAN (1992) Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. J. Exp. Med. 176, 1693-1702 INABA, K., M. INABA, M. NAITO und R. M. STEINMAN (1993) Dendritic cell progenitors phagocytose particulates, including bacillus CalmetteGuerin organisms, and sensitize mice to mycobacterial antigens in vivo. J. Exp. Med. 178, 479-88 JANEWAY, C. A. und P.TRAVERS (1997) Immunologie 2. Auflage Spektrum Akademischer Verlag, Heidelberg, Berlin JANEWAY, C. A., P. TRAVERS, M. WALPORT und M. SHLOMCHICK (2002) Grundbegriffe der Immunologie S. 3 ff. Die Werkzeuge des Immunologen S. 681 f. in: Immunologie 5. Auflage Spektrum Akademischer Verlag, Heidelberg, Berlin JANEWAY, C. A. und R. MEDZHITOV (2002) Innate immune recognition. Annu. Rev. Immunol. 20, 197-216 - 95 - Literaturverzeichnis JONULEIT, H., E. SCHMITT, G. SCHULER, J. KNOP und A. H. ENK (2000): Induction of interleukin 10-producing, nonproliferating CD4(+) T cells with regulatory properties by repetitive stimulation with allogeneic immature human dendritic cells. J. Exp. Med. 192, 1213-1222 KAESE, V. (2002) Korrosionsschützendes Legieren von Magnesiumwerkstoffen. Hannover, Leibnitz Universität, Diss. KRAUSE, A., C. HACKENBROICH, N. VON DER HÖH, S. WAGNER, D. BORMANN, T. HASSEL, H. WINDHAGEN und A. MEYER-LINDENBERG (2005) Seltene Erden-haltige Magnesiumlegierungen als degradable, intramedulläre Implantate in Kaninchentibiae Biomaterialien 6, 190 KRAUSE, A., C. KRAUSE, H. WINDHAGEN und A. MEYER-LINDENBERG (2006): In vivo- Degradationseigenschaften und Biokompatibilität von resorbierbaren Implantaten aus MgCa0,8 im Kaninchenmodell Gemeinsame Jahrestagung der Deutschen, Österreichischen und Schweizerischen Gesellschaften für Biomedizinische Technik. ETH Zürich, Schweiz KRUTZIK, S. R. und R. L. MODLIN (2004) The role of Toll-like receptors in combating mycobacteria. Semin. Immunol. 16, 35-41 LAMBOTTE, A. (1932) L’utilisation du magnesium comme materiel perdu dans l’osteosynthèse Bull. Mém. Soc. Nat. Chir. 28, 1325-1334 - 96 - Literaturverzeichnis LAMBRECHT, B. N. (2001) Allergen uptake and presentation by dendritc cells Curr. Opin. Allergy. Clin. Immunol. 1, 51-59 LANGERHANS, P. (1868) Über die Nerven der menschlichen Haut. Virchows Archiv für pathologische Anatomie und Physiologie, und für klinische Medicin 44, 325-337 LATCHUMANAN, V. K., B. SINGH, P. SHARMA und K. NATARAJAN (2002) Mycobacterium tuberculosis antigens induce the differentiation of dendritic cells from bone marrow. J. Immunol. 169, 6856-6864 LEBRE, M. C., T. BURWELL, P. L. VIEIRA, J. LORA, A. J. COYLE, M. L. KAPSENBERG, B. E. CLAUSEN und E. C. DE JONG (2005) Differential expression of inflammatory chemokines by Th1- and Th2-cell promoting dendritic cells: a role for different mature dendritic cell populations in attracting appropriate effector cells to peripheral sites of inflammation. Immunol. Cell Biol. 83, 525-535 LI, Z., X. GU, S. LOU und Y. ZHENG (2008) The development of binary Mg-Ca alloys for use as biodegradable materials within bone Biomaterials 29, 1329-1344 LIU, Y. J. (2001) Dendritic cell subsets and lineages, and their functions in innate and adaptive immunity. Cell 106, 259-262 - 97 - Literaturverzeichnis LUTTMANN, W., K. BRATKE, M. KÜPPER und D. MYRTEK (2006) Durchflusscytometrie S. 77 ff. Quantitative Immunoassays S. 113 ff. in: Der Experimentator: Immunologie 2. Auflage Spektrum Akademischer Verlag Heidelberg, Berlin LUTZ, M. B., N. KUKUTSCH, A. L. OGILVIE, S. ROSSNER, F. KOCH, N. ROMANI und G.SCHULER (1999) An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 223, 77-92 LUTZ, M. B. und G. SCHULER (2002) Immature, semi-mature and fully mature dendritic cells: which signals induce tolerance or immunity? Trends Immunol. 23, 445-449 MAHNKE, K., M. GUO, S. LEE, H. SEPULVEDA, S. L. SWAIN, M. NUSSENZWEIG und R. M. STEINMAN (2000) The dendritic cell receptor for endocytosis, DEC-205, can recycle and enhance antigen presentation via major histocompatibility complex class II-positive lysosomal compartments. J. Cell. Biol. 151, 673-684 MANOME, H., S. AIBA und H. TAGAMI (1999) Simple chemicals can induce maturation and apoptosis of dendritic cells. Immunol. 98, 481-490 - 98 - Literaturverzeichnis MCBRIDE, E. D. (1938) Absorbable Metal in Bone Surgery J. Am. Med. Assoc. 111, 2464-2467 MCCOLL, S. R. (2002) Chemokines and dendritic cells: a crucial alliance. Immunol. Cell. Biol. 80, 489-496 MCCARTHY, J. T. und R. KUMAR (1999) Divalent Cation Metabolism: Magnesium (Chapter 4) In: Atlas of Diseases of the Kidney: Vol.1 (Robert W. Schrier) University of Colorado School of Medicine Denver, Colorado MEYER-LINDENBERG, A., A. KRAUSE, C. KRAUSE, D. BORMANN und H. WINDHAGEN (2007) Entwicklung der mechanischen Eigenschaften von degradablen intramedullären Implantaten auf Magnesiumbasis nach unterschiedlicher Implantationsdauer Biomaterialien 8, 180 MEYER-LINDENBERG, A., H. WINDHAGEN, M. NIEMEYER, E. SWITZER, F. WITTE, V. KAESE, T. FABIAN und A. SCHIESZLER (2003) Untersuchungen zum Einsatz degradabler metallischer Osteosynthese-Materialien auf Magnesiumbasis FK-DVG, 49. Jahreskongress, Leipzig, 6-9, 174-176 MOSMANN, T. (1983) Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays J. Immunol. Methods 65, 55-63 - 99 - Literaturverzeichnis NATIONAL ACADEMY OF SCIENCES, INSTITUTE OF MEDICINE (1999) Dietary reference intakes for calcium, magnesium, vitamin D, and fluoride. The National Academy Press, Washington D.C. PAMER, E. und P. CRESSWELL (1998) Mechanisms of MHC class I--restricted antigen processing Annu. Rev. Immunol. 16, 323-358 PAVIL, P., L. MAXWELL, E. VAN DE POL und F. DOE (1996) Distribution of human colonic dendritic cells and macrophages. Clin. Exp. Immunol. 104, 124-132 PICCIOLI, D., S. SBRANA, E. MELANDRI und N. M. VALIANTE (2002) Contact-dependent stimulation and inhibition of dendritic cells by natural killer cells J. Exp. Med. 195, 335-341 PULENDRAN, B., E. MARASKOEAU und C. R. MALISZEWSKI (2001) Modulating the immune response with dendritic cells and their growth factors. Trends in Immunol. 22, 41-47 PRICKETT, T. C., J. L. MCKENZIE und D. N. HART (1988) Characterization of interstitial dendritic cells in human liver. Transplantation 46, 754-761 QUAMME, G. A. (1993) Magnesium homeostasis and renal magnesium handling. Miner. Electrolyte Metab. 19, 218-225 QUAMME, G. A. (1997) Renal magnesium handling: new insights in understanding old problems Kidney Int. 52, 1180-1195. - 100 - Literaturverzeichnis RESCIGNO, M., F. GRANUCCI, S. CITTERIO, M. FOTI und P. RICCIARDICASTAGNOLI (1999) Coordinated events during bacteria-induced DC maturation Immunol. Today 20, 200-203 RYAN, M. P. (1993) Interrelationships of magnesium and potassium homeostasis Miner Electrolyte Metab. 19, 290-295 SALLUSTO, F., M. CELLA, C. DANIELI und A. LANZAVECCHIA (1995) Dendritic cells use macropinocytosis and the mannose receptor to concentrate macromolecules in the major histocompatibility complex class II compartment: downregulation by cytokines and bacterial products. J. Exp. Med. 182, 389-400 SALLUSTO, F., P. SCHAERLI, P. LOETSCHER, C. SCHANIEL, D. LENIG, C. R. MACKAY, S. QIN und A. LANZAVECCHIA (1998) Rapid and coordinated switch in chemokine receptor expression during dendritic cell maturation Eur J. Immunol. 28, 2760-2769 SARIS, N. E., E. MERVAALA, H. KARPPANEN, J. A. KHAWAJA und A. LEWENSTAM (2000) Magnesium. An update on physiological, clinical and analytical aspects. Clin. Chem. Acta. 294, 1-26 SCHAAFSMA, G. (1997) Bioavailability of calcium and magnesium. Eur. J. Clin. Nutr. 51, 13-16 - 101 - Literaturverzeichnis SCHWEIGEL, M. und H. MARTENS (2000) Magnesium transport in the gastrointestinal tract. Front. Biosci. 5, 666-677. SERRE, C. M., M. PAPILLARD, P. CHAVASSIEUX, J. C. VOEGEL und G. BIOVIN (1998) Influence of magnesium substitution on a collageneapatite biomaterial on the production of a calcifying matrix by human osteoblasts J. Biomed. Mater. Res. 42, 626-633 SERTL, K., T. TAKEMURA, E. TSCHACHLER, V. J. FERRANS, M. A. KALINER und E. M. SHEVACH (1986) Dendritic cells with antigen-presenting capability reside in airway epithelium, lung parenchyma, and visceral pleura. J. Exp. Med. 163, 436-451 SHIKINAMI, Y. und M. OKUNO (1999) Bioresorbable devices made of forged composites of hydroxyapatite (HA) particles and poly-L-lactide (PLLA) Part I. Basic characteristics. Biomaterials 20, 859-877 SHILS, M. A. (1997) Magnesium. In: O`Dell, B.J., Sunde, R. (Eds.), Handbook of Nutritionally Essential Mineral Elements. Marcel Dekker, New York, Basel, Hong Kong, 117-152. SOJKA, J. E. und C. M. WEAVER ( 1995) Magnesium supplementation and osteoporosis Nutr. Rev. 53, 71-74 - 102 - Literaturverzeichnis SOZZANI, S., P. ALLAVENA, A. VECCHI und A. MANTOVANI (1999) The role of chemokines in the regulation of dendritic cell trafficking J. Leukoc. Biol. 66, 1-9 SPIEKSTRA, S. W., M. J. TOEBAK, S. SAMPAT-SARDJOEPERSAD, P. J. VAN BEEK, D. M. BOORSMA, T. J. STOOF, B. M. VON BLOMBERG, R. J. SCHEPER, D. P. BRUYNZEEL, T. RUSTEMEYER und S. GIBBS (2005) Induction of cytokine (interleukin-1-alpha and tumor necrosis factor-alpha) and chemokine (CCL20, CCL27 and CXCL) alarm signals after allergen and irritant exposure Exp. Dermatol. 14, 109-116 STAIGER, M. P., A. M. PIETAK, J. HUADMAI und G. DIAS (2006) Magnesium and its alloys as orthopedic biomaterials: a review Biomaterials 27, 1728-1734. STEINMAN, R. M. und Z. A. COHN (1973) Identification of a novel cell type in peripheral lymphoid organs of mice: Morphology, quantitation, tissue distribution. J. Exp. Med. 137, 1142-1162 STEINMAN, R. M. (1991): The dendritic cell system and its role in immunogenicity. Annu. Rev. Immunol. 9, 271-296 STEINMAN, R. M., D. HAWIGER und M. C. NUSSENZWEIG (2003a) Tolerogenic dendritic cells. Annu. Rev. Immunol. 21, 685-711 STEINMAN, R. M. (2003b) Some interfaces of dendritic cell biology. APMIS 111, 675-697 - 103 - Literaturverzeichnis STINGL, G. und P. R. BERGSTRESSER (1995): Dendritic cells: a major story unfolds. Immunol. Today 16, 330-333. STROGANOV, G. B., E. SAVITSKY, N. TIKHOVA, V. TEREKHOVA und V. E. A. FEDOROVNA (1972) Magnesium-base alloys for use in bone surgery Patentschrift 13.05.1971, (1953241) SWAMINATHAN, R. (2003) Magnesium Metabolism and its Disorders, Toxikologische Bedeutung Clin. Biochem. Rev. 24, 47-66 SWITZER, E. N. (2005): Resorbierbares metallisches Osteosynthesematerial Hannover, Tierärztliche Hochschule, Diss. TROITSKII, V. und D. TSITRIN (1944) The resorbing metallic alloy "Osteosintezit" as material for fastening broken bones Khirurgiya 8, 41-44 VAN VOORHIS, W. C., L. S. HAIR, R. M. STEINMAN und G. KAPLAN (1982) Human dendritic cells: enrichment and characterization from peripheral blood. J. Exp. Med. 155, 1172-1187 VERBRUGGE, J. (1934) Le Material Metalique resobable en Chirurgie osseuse La Presse Medicale 23, 460-465 - 104 - Literaturverzeichnis VERHEYEN, G. R., E. SCHOETERS, J. -M. NUIJTEN, R. L. V. D. HEUVEL, I. NELISSEN, H. WITTERS, V. V. TENDELOO, Z. N. BERNEMANN und G. E. R. SCHOETERS (2004) Cytokine transcript profiling in CD34+-progenitor derived dendritic cells exposed to contact allergens and irritants. Toxicology Letters 155, 187-194 VERMAELEN, K. und R. PAUWLS (2005) Pulmonary dendritic cells. J. Res. Crit. Care Med. 172, 530-551 WALLET, M. A., P. SEN und R. TISCH (2005) Immunoregulation of dendritic cells. Clin. Med. Res. 3, 166-175 WÄRING, J. (2008) Molekulare und evolutionäre Hintergründe der Kationenpermeation bei Mitgliedern der TRPM-Subfamilie Marburg, Philipps-Universität, Diss. WEISINGER J. R. und E. BELLORIN-FONT(1998) Magnesium and phosphorus Lancet. 352, 391-396 WILLIAMS, R. J. P. (1993) Magnesium: an introduction to its biochemistry, S. 15-30. In Nicholas J. Birch (ed.), Magnesium and the cell. Academic Press, Boston, Mass. - 105 - Literaturverzeichnis WINZLER, C., P. ROVERE, M. RESIGNO, F. GRANNUCCI, G. PENNA, L. ADORINI, V. S. ZIMMERMANN, J. DAVOUST und P. RICCIARDI-CASTAGNOLI (1997) Maturation stages of mouse dendritic cells in growth factor-dependent long-term cultures J. Exp. Med. 185, 317-328 WITTE, F., J. FISCHER, J. NELLESEN, H. A. CROSTACK, V. KAESE, A. PISCH, F. BECKMANN und H. WINDHAGEN (2006a) In vitro and in vivo corrosion measurements of magnesium alloys Biomaterials 27, 1013-1018 WITTE, F., H. ULRICH, M. RUDERT und E. WILLBOLD (2006b) Biodegradable magnesium scaffolds: Part I: Appropriate inflammatory response J. Biomed. Mater. Res. A. 81, 748-756 WITTE, F., V. KAESE, H. HAFERKAMP, E. SWITZER, A. MEYER-LINDENBERG, C. J. WIRTH und H. WINDHAGEN (2005) In vivo corrosion of four magnesium alloys and the associated bone response Biomaterials 26, 3557-3563 WOLF F. I. und A. CITTADINI (2003) Chemistry and biochemistry of Magnesium Mol. Aspects Med. 24, 3-9 WU, L. und A. DAKIC (2004) Development of dendritic cell system. Cell. Mol. Immunol. 1, 112-118 - 106 - Literaturverzeichnis YUMIKO, N., T. YUKARI, T. YASUHIDE, S. TADASHI und I. YOSHIO (1997) Differences in behavior among the chlorides of seven rare earth elements administered intravenously to rats Fundam. Appl. Toxicol. 37,106-116 ZITVOGEL, L. (2002) Dendritic and natural killer cells cooperate in the control/switch of innate immunity J. Exp. Med. 195, F9-F14 - 107 - Anhang 9. Anhang Tabelle 9.1.: Magnesiumkonzentration in den Degradationsmedien Die Legierungen Mg(pur); MgCa0.6; MgCa0.8; MgCa1.0 und MgCa1.2 wurden für 72h mit 10ml DCMedium im Wasserbad (37°C) inkubiert. Anschließend erfolgte die Bestimmung der Magnesium- und Calciumkonzentration mit Hilfe des Testkits Nanocolor Härte 20. n=8 Messung 1 2 3 4 5 6 7 8 MW S.D. Mg(pur) MgCa0.6 MgCa0.8 MgCa1.0 MgCa1.2 [mmol/l] [mmol/l] [mmol/l] [mmol/l] [mmol/l] 1,03 0,97 0,68 0,50 1,59 0,43 1,64 1,50 1,04 0,49 1,31 1,37 1,19 1,25 1,72 1,08 1,74 1,22 1,36 0,24 1,05 1,39 0,78 1,20 1,52 1,15 1,74 1,62 1,30 0,32 3,48 3,36 0,95 1,29 1,05 1,47 1,47 1,18 1,78 1,03 1,78 1,13 1,85 1,18 0,69 2,09 1,99 1,13 1,48 0,51 Tabelle 9.2.: DC-Zellzahlen nach 10-tägiger Kultivierung in, mit MgCl2 angereichertem, DC-Medium Die DC wurden ab Tag 3 der Kultivierung in MgCl2 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l kultiviert. Am Tag 10 wurde die Zellzahl der DC bestimmt. n=3 n1 DC-Zellzahl/Well n2 n3 Kontrolle 1269000 1255500 1327500 MgCl2 0,1 mmol/l 982599 1047500 904500 MgCl2 1,0 mmol/l 963000 949500 936000 MgCl2 10 mmol/l 630000 850500 751500 - 108 - Anhang Tabelle 9.3.: MØ-Zellzahl nach 10-tägiger Kultivierung der DC-Vorläuferzellen in, mit MgCl2 angereichertem, DC-Medium Die DC wurden ab Tag 3 der Kultivierung in MgCl2 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l kultiviert. Am Tag 10 wurde die Zellzahl der MØ bestimmt. n=3 Kontrolle n1 45000 MØ-Zellzahl/Well n2 30750 MgCl2 0,1 mmol/l 48000 34500 43500 MgCl2 1,0 mmol/l 55500 45750 45750 MgCl2 10 mmol/l 117000 74250 82500 n3 39750 Tabelle 9.4.: DC-Zellzahlen nach 10-tägiger Kultivierung in, mit CaCl2 angereichertem, DC-Medium Die DC wurden ab Tag 3 der Kultivierung in CaCl2 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l kultiviert. Am Tag 10 wurde die Zellzahl der DC bestimmt. n=3 Kontrolle n1 1269000 DC-Zellzahl/Well n2 1255500 n3 1327500 CaCl2 0,1 mmol/l 1113750 1147500 963000 CaCl2 1,0 mmol/l 1068750 982500 940500 CaCl2 10 mmol/l 472500 249375 657000 - 109 - Anhang Tabelle 9.5.: MØ-Zellzahl nach 10-tägiger Kultivierung der DC-Vorläuferzellen in, mit CaCl2 angereichertem, DC-Medium Die DC wurden ab Tag 3 der Kultivierung in MgCl2 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l kultiviert. Am Tag 10 wurde die Zellzahl der MØ bestimmt. n=3 Kontrolle n1 45000 MØ-Zellzahl/Well n2 30750 CaCl2 0,1 mmol/l 69000 33750 45750 CaCl2 1,0 mmol/l 81000 38250 53250 CaCl2 10 mmol/l 144000 87000 81750 n3 39750 Tabelle 9.6.: DC-Zellzahl nach 10-tägiger Kultivierung der DC-Vorläuferzellen in den Degradationsmedien der Legierungen Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2. Die DC wurden ab Tag 3 der Kultivierung in den Degradationsmedien der einzelnen Legierungen kultiviert. Am Tag 10 wurde die Zellzahl der DC bestimmt. n=5 Kontrolle n1 918000 DC-Zellzahl/Well n2 n3 999000 1089000 n4 841500 n5 1004000 Mg(pur) 778500 855000 1012500 598500 822000 MgCa0.6 738000 781500 963000 724500 696000 MgCa0.8 781500 913500 918000 607500 762000 MgCa1.0 657000 805500 855000 787500 657000 MgCa1.2 756000 774000 553500 657000 598500 - 110 - Anhang Tabelle 9.7.: MØ-Zellzahl nach 10-tägiger Kultivierung der DC-Vorläuferzellen in den Degradationsmedien der Legierungen Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2. Die DC wurden ab Tag 3 der Kultivierung in den Degradationsmedien der einzelnen Legierungen kultiviert. Am Tag 10 wurde die Zellzahl der MØ bestimmt. n=5 Kontrolle n1 38250 MØ-Zellzahl/Well n2 n3 n4 38250 33000 43500 Mg(pur) 36750 36750 35250 51000 39000 MgCa0.6 45750 45750 36000 46500 47250 MgCa0.8 43500 43500 34500 55500 49500 MgCa1.0 46500 39000 48000 48750 45750 MgCa1.2 45000 38250 54750 57000 48000 n5 35350 Tabelle 9.8.: Analyse Annexin V-FITC von DC, die mit MgCl2 angereicherten Kulturmedium kultiviert wurden Positiv-Kontrolle: Zugabe von 20 µg/ml Etoposide 24 h vor Versuchsdurchführung. Pro Ansatz wurden 5000 DC gemessen. n=3 Kontrolle Versuch 1 25,6 Annexin V-FITC [%] Versuch 2 23,6 Versuch 3 22,3 Etoposide 20µg/ml 50,5 56,6 54,6 MgCl2 0,1 mmol/l 20,6 17,1 23,8 MgCl2 1,0 mmol/l 17,1 17,0 21,1 MgCl2 10 mmol/l 17,2 20,1 22,8 - 111 - Anhang Tabelle 9.9.: Analyse Annexin V-FITC von DC, die mit CaCl2 angereicherten Kulturmedium kultiviert wurden Positiv-Kontrolle: Zugabe von 20 µg/ml Etoposide 24 h vor Versuchsdurchführung. Pro Ansatz wurden 5000 DC gemessen. n=3 Kontrolle Versuch 1 25,6 Annexin V-FITC [%] Versuch 2 23,6 Versuch 3 22,3 Etoposide 20µg/ml 50,5 56,6 54,6 CaCl2 0,1 mmol/l 15,4 17,1 21,7 CaCl2 1,0 mmol/l 17,2 20,1 25,5 CaCl2 10 mmol/l 71,8 64,3 68,5 Tabelle 9.10.: Analyse Annexin V-FITC von DC, die in den Degradationsmedien der Legierungen Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2 kultiviert wurden Positiv-Kontrolle: Zugabe von 20 µg/ml Etoposide 24 h vor Versuchsdurchführung. Pro Ansatz wurden 5000 DC gemessen. n=3 Kontrolle Versuch 1 25,6 Annexin V-FITC [%] Versuch 2 23,6 Versuch 3 22,3 Etoposide 20µg/ml 50,5 56,6 54,6 Mg(pur) 33,4 33,1 28,7 MgCa0.6 28,0 28,0 24,3 MgCa0.8 26,6 23,7 20,1 MgCa1.0 24,8 30,7 21,8 MgCa1.2 24,5 25,8 21,5 - 112 - Anhang Tabelle 9.11.: XTT-Test nach Inkubation der DC mit MgCl2 Die DC wurden ab Tag 3 der Kultivierung mit MgCl2 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l inkubiert. Am Tag 10 erfolgte die Auswertung der Vitalität mittels XTT-Test. n=4 Kontrolle Vitale DC [%] n2 n3 100 100 n1 100 n4 100 MgCl2 0,1 mmol/l 102,03 97,5 89,62 93,22 MgCl2 1,0 mmol/l 91,60 101,35 87,97 81,67 MgCl2 10 mmol/l 97,89 99,97 97,26 71,12 Tabelle 9.12.: XTT-Test nach Inkubation der DC mit CaCl2 Die DC wurden ab Tag 3 der Kultivierung mit CaCl2 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l inkubiert. Am Tag 10 erfolgte die Auswertung der Vitalität mittels XTT-Test. n=4 Kontrolle Vitale DC [%] n2 n3 100 100 n1 100 n4 100 CaCl2 0,1 mmol/l 89,15 101,27 92,90 72,81 CaCl2 1,0 mmol/l 92,06 90,66 83,64 77,88 CaCl2 10 mmol/l 84,52 80,31 77,95 64,36 Tabelle 9.13.: XTT-Test der DC nach Inkubation in den Degradationsmedien der Legierungen Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2 Die DC wurden ab Tag 3 der Kultivierung in den jeweiligen Degradationsmedien kultiviert Am Tag 10 erfolgte die Auswertung der Vitalität mittels XTT-Test. n=5 Kontrolle n1 100 n2 100 Vitale DC [%] n3 100 Mg(pur) 93,44 96,51 89,00 91,02 97,00 MgCa0.6 103,71 96,64 95,07 91,75 94,93 MgCa0.8 104,50 94,69 85,43 81,43 88,45 MgCa1.0 99,88 95,46 83,22 105,95 85,17 MgCa1.2 107,24 102,58 100,30 92,77 98,91 - 113 - n4 100 n5 100 Anhang Tabelle 9.14.: TNF-α [pg/ml] in Kulturüberständen von DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l MgCl2 Als Positivkontrolle wurden DC mit LPS (1 µg/ml) stimuliert. n=5 LPS Kontrolle MgCl2 0,1 mmol/l MgCl2 1,0 mmol/l MgCl2 10 mmol/l n1 486 n2 718 TNF-α[ pg/ml] n3 876 n4 n5 63 4 55 18 75 166 127 105 162 28 158 69 109 202 8 129 75 109 243 61 Tabelle 9.15.: TNF-α [pg/ml] in Kulturüberständen von DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l CaCl2 Als Positivkontrolle wurden DC mit LPS (1 µg/ml) stimuliert. n=5 LPS Kontrolle CaCl2 0,1 mmol/l CaCl2 1,0 mmol/l CaCl2 10 mmol/l n1 486 n2 718 TNF-α[ pg/ml] n3 876 63 4 55 18 75 178 119 24 97 75 160 125 136 81 59 99 111 117 117 71 - 114 - n4 n5 Anhang Tabelle 9.16.: TNF-α [pg/ml] in Kulturüberständen von DC nach Kultivierung in den Degradationsmedien der Legierungen Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0, MgCa1.2 Als Positivkontrolle wurden DC mit LPS (1 µg/ml) stimuliert. n=5 n1 486 n2 718 TNF-α[ pg/ml] n3 876 63 4 55 18 75 65 99 67 63 14 MgCa0.6 26 111 77 75 38 MgCa0.8 101 77 67 89 4 MgCa1.0 97 67 83 129 59 MgCa1.2 83 85 89 127 18 LPS Kontrolle Mg(pur) n4 n5 Tabelle 9.17.: Migrationsassay der DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l MgCl2 Die DC wurden ab Tag 3 der Kultivierung mit MgCl2 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l inkubiert. Am Tag 10 erfolgte die Auswertung der Migration. Als Positivkontrolle wurden DC mit 100 µmol/l Bleichlorid (Pb) inkubiert. n=6 PbCl2 Kontrolle MgCl2 0,1 mmol/l MgCl2 1,0 mmol/l MgCl2 10 mmol/l Ausgewanderte DC/ml n3 n4 39750 27000 n1 33000 n2 37500 n5 33750 n6 27750 18750 24000 15000 28500 15000 20250 28500 24000 27000 23250 18000 16500 27750 19500 23250 30000 27000 15750 36250 30750 33000 33750 23250 27750 - 115 - Anhang Tabelle 9.18.: Migrationsassay von DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l CaCl2 Die DC wurden ab Tag 3 der Kultivierung mit CaCl2 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l inkubiert. Am Tag 10 erfolgte die Auswertung der Migration. Als Positivkontrolle wurden DC mit 100 µmol/l Bleichlorid (Pb) inkubiert. n=6 Pb n1 33000 Ausgewanderte DC/ml n2 n3 n4 37500 39750 27000 n5 33750 n6 27750 Kontrolle 18750 24000 15000 28500 15000 20250 CaCl2 0,1 mmol/l 24000 27750 21000 33000 20250 18000 CaCl2 1,0 mmol/l 19500 30750 20250 29250 24000 19500 CaCl2 10 mmol/l 33750 29250 38250 38250 28500 33750 Tabelle 9.19.: Migrationsassay der DC, die in den Degradationsmedien der Legierungen Mg(pur), MgCa0.6, MgCa0.8, MgCa1.0 und MgCa1.2 kultiviert wurden Als Positivkontrolle wurden DC mit 100 µmol/l Bleichlorid (Pb) inkubiert. n=9 Kontrolle Ausgewanderte DC/ml n1 n2 n3 n4 n5 n6 n7 n8 n9 26250 16500 18000 27000 22500 32250 25500 17250 24000 Pb 39750 42000 41250 38250 43500 39750 30750 43500 33750 Mg(pur) 30750 29250 27000 29250 29250 30750 24750 23250 19500 MgCa0.6 21750 24750 24000 27750 24000 31500 25500 23250 24750 MgCa0.8 22500 27000 24000 30750 33750 29250 22500 24750 26250 MgCa1.0 38250 30750 31500 33750 29250 33750 19500 18000 22500 MgCa1.2 30750 31500 29250 23250 29250 27000 28500 27750 33750 - 116 - Anhang Tabelle 9.20.: Expression von CD11c und CD86 von DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l MgCl2 Positivkontrolle: Zugabe von 1 µg/ml LPS 24h vor Versuchsdurchführung. Pro Ansatz wurden 5000 DC gemessen. n=3 Kontrolle CD11c/CD86- positive DC [%] Versuch 1 Versuch 2 Versuch 3 6,1 8,6 8,0 LPS 16,6 34,6 20,7 MgCl2 0,1 mmol/l 5,6 10,0 7,2 MgCl2 1,0 mmol/l 4,6 6,8 5,6 MgCl2 10 mmol/l 6,1 6,7 4,5 Tabelle 9.21.: Expression von CD11c und CD86 von DC nach Inkubation mit 0,1 mmol/l, 1,0 mmol/l und 10 mmol/l CaCl2 Positivkontrolle: Zugabe von 1 µg/ml LPS 24h vor Versuchsdurchführung. Pro Ansatz wurden 5000 DC gemessen. n=3 Kontrolle CD11c/CD86- positive DC [%] Versuch 1 Versuch 2 Versuch 3 6,1 8,6 8,0 LPS 16,6 34,6 20,7 CaCl2 0,1 mmol/l 5,9 6,7 10,7 CaCl2 1,0 mmol/l 5,9 5,0 6,7 CaCl2 10 mmol/l 7,6 6,7 9,9 - 117 - Anhang Tabelle 9.22.: Expression von CD11c und CD86 von DC, die den Degradationsmedien der Legierungen Mg(pur), MgCa 0.6, MgCa 0.8, MgCa 1.0 und MgCa 1.2 kultiviert wurden. Positivkontrolle: Zugabe von 1 µg/ml LPS 24h vor Versuchsdurchführung. Pro Ansatz wurden 5000 DC gemessen. n=3 Kontrolle CD11c/CD86-positive DC [%] Versuch 1 Versuch 2 Versuch 3 5,8 8,8 8,0 LPS 16,6 34,6 20,7 Mg(pur) 8,4 9,5 7,6 MgCa0.6 7,4 8,0 5,8 MgCa0.8 9,2 6,4 7,4 MgCa1.0 9,2 8,1 8,6 MgCa1.2 6,7 6,0 5,8 Tabelle 9.23.: CFSE markierten T-Lymphozyten nach Ko-Inkubation mit MgCl2behandelten DC Pro Ansatz wurden 10.000 T-Lymphozyten gemessen. n=3 Kontrolle Versuch 1 48,7 T-Zellproliferation [%] Versuch 2 53,3 Versuch 3 37,6 MgCl2 0,1 mmol/l 61,6 52,3 41,3 MgCl2 1,0 mmol/l 61,1 52,1 42,6 MgCl2 10 mmol/l 53,8 49,6 53,6 - 118 - Anhang Tabelle 9.24.: CFSE markierten T-Lymphozyten nach Ko-Inkubation mit CaCl2behandelten DC Pro Ansatz wurden 10.000 T-Lymphozyten gemessen. n=3 Kontrolle Versuch 1 48,7 T-Zellproliferation [%] Versuch 2 53,3 Versuch 3 37,6 CaCl2 0,1 mmol/l 55,5 44,9 42,2 CaCl2 1,0 mmol/l 59,1 57,9 39,0 CaCl2 10 mmol/l 77,5 65,0 76,2 Tabelle 9.25.: CFSE markierten T-Lymphozyten nach Ko-Inkubation mit in Degradationsmedien kultivierten DC Pro Ansatz wurden 10.000 T-Lymphozyten gemessen. n=3 Kontrolle Versuch 1 51,2 T-Zellproliferation [%] Versuch 2 38,0 Versuch 3 44,9 Mg(pur) 58,4 42,3 46,2 MgCa0.6 55,6 42,4 47,3 MgCa0.8 53,5 42,2 46,7 MgCa1.0 57,2 43,4 51,4 MgCa1.2 58,7 45,8 53,3 - 119 - Hiermit erkläre Untersuchung ich, zur dass ich die Biokompatibilität Dissertation von „Immunotoxikologische abbaubaren Implantaten aus Magnesiumbasislegierungen“ selbständig verfasst habe. Ich habe keine entgeltliche Hilfe von Vermittlungs- bzw. Beratungsdiensten (Promotionsberater oder andere Personen) in Anspruch genommen. Niemand hat von mir unmittelbar oder mittelbar entgeltlich Leistungen für Arbeiten erhalten, die im Zusammenhang mit dem Inhalt der vorgelegten Dissertation stehen. Ich habe die Dissertation am folgenden Institut angefertigt: Institut für Pharmakologie, Toxikologie und Pharmazie der Stiftung Tierärztliche Hochschule Hannover Bünteweg 17, 30559 Hannover. Die Dissertation wurde bisher nicht für eine Prüfung oder eine Promotion oder für einen ähnlichen Zweck zur Beurteilung eingereicht. Ich versichere, dass ich die vorstehenden Angaben nach bestem Wissen vollständig und der Wahrheit entsprechend gemacht habe. Danksagung An dieser Stelle bedanke ich mich recht herzlich bei all denen, die zum Gelingen dieser Arbeit beigetragen haben, insbesondere: - Herrn Prof. Dr. Manfred Kietzmann für die Überlassung des interessanten Themas, der stets freundlichen und humorvollen Unterstützung und der abwechslungsreichen Konversationen. - Herrn Prof. Dr. Wolfgang Bäumer für die gewährte Unterstützung, der hervorragenden Betreuung sowie für die immerwährende Geduld. - Herrn Dr. Frank Niedorf für den Rat und Tat bei analytischen Fragen sowie den abwechslungsreichen Mittagspausen. - den stets hilfsbereiten und fröhlichen Mitarbeitern der Arbeitsgruppe – Viki, Caro, Isa und Hans-Herbert Bohr – für die Hilfe im Labor. - Meinen Mitdoktoranten für jede Menge Spaß und aufmunternde Gesprächen. - Meinen Chefs Ludger und Paul, die es möglich gemacht haben, Doktorarbeit und Praxis unter einen Hut zu bekommen und meinen Kollegen Armin, Mattias und Ruth, die, durch zahlreiche Überstunden, mir genügend Freizeit für die Doktorarbeit verschafften. - meinen lieben Eltern für die liebevolle Unterstützung und den Rückhalt, den sie mir all die Jahre boten. - Silvia für die Geduld, den liebevollen Rückhalt und das größte Geschenk: Unseren kleinen Sohn Pitt.