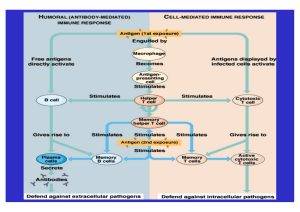
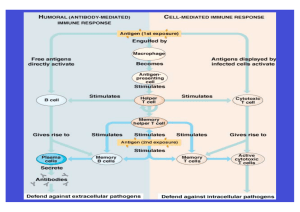
Zytokine und die immunmodulatorische Funktion des Vagusnervs
Werbung

Britische Zeitschrift für Anästhesie 102 (4): 453-62 (2009) Vorabdruck 3. März 2009 Zytokine und die immunmodulatorische Funktion des Vagusnervs G. R. Johnston und N. R. Webster* Institut für Medizinwissenschaften, Universität Aberdeen, Foresterhill, Aberdeen AB25 2ZD, UK E-Mails sind zu richten an den Autor: [email protected] Die Zytokinsynthese und -freisetzung sind die zentralen Aufgaben des angeborenen Immunystems. Eine gestörte und überschießende Produktion führt zu einer systemischen, den ganzen Körper betreffenden, Entzündungsreaktion, wodurch Fernorgane geschädigt werden. Aus der jüngsten Forschung geht hervor, dass dem Vagusnerv eine immunmodulatorische Funktion zukommt. Wird sein efferenter Schenkel aktiviert, kommt es zur Regulierung der Zytokinproduktion. Diese neuroimmune Kommunikation, als 'cholinerger antiinflammatorischer Weg' bezeichnet, hilft dem Wirt rasch, von sich aus und örtlich begrenzt, die Immunantwort zu regulieren und eine schwere Entzündung zu vermeiden. Die Stimulation des Vagusnervs verringert die Zytokinproduktion und verbessert die Überlebenschancen im Experiment bei Sepsis, beim hämorrhagischen Schock, bei einer Schädigung durch Ischämie/Reperfusion und bei anderen durch Zytokinüberschuss verursachten Erkrankungen. In der Forschung werden zur Zeit brandneue Therapeutika zur Stimulation des Vagusnervs entwickelt. Diese stimulieren ihn entweder direkt oder beziehen sich auf bestimmte Teile seiner Leitungsbahn. Stichwörter: Komplikationen, Infektion; Immunreaktion; parasympatisches Nervensystem, Vagus; Polypeptide, Zytokine Die Überlebenschancen sind abhängig davon, ob das Immunsystem imstande ist, den Körper gegen eindringende Pathogene und die Verletzung zu schützen. Das Ausmaß dieser Reaktion ist dabei von entscheidender Bedeutung. Eine Unterreaktion kann Sekundärinfektionen aufgrund einer Immunsuppression verursachen und eine Überreaktion kann gefährlicher sein als die ursprüngliche Verletzung. Die immunologische Reaktion kann in zwei verschiedene Reaktionstypen unterteilt werden (Tabelle 1). Die angeborene (unspezifische) Reaktion findet schnell statt. Sie unterscheidet nicht zwischen fremden Substanzen und steigert sich nicht, wenn ein Kontakt mit dem selben Antigen erneut stattfindet. Sie funktioniert auf der Grundlage natürlicher Killerzellen, welche von inflammatorischen Mediatoren und phagozytischen Zellen, etwa Neutrophilen, Monozyten und Makrophagen, freigesetzt werden. Im Gegensatz dazu wird die erworbene (spezifische) Immunreaktion durch Kontakt hervorgerufen. Sie ist für jedes in Berührung gekommene Antigen spezifisch. Sie verbessert sich, sobald ein neuerlicher Kontakt mit dem spezifischen Antigen stattgefunden hat. Mithilfe der antigenspezifischen B- und T-Lymphozyten wird das Antigen erkannt und eine Reaktion darauf ausgelöst. Zytokine Ein wesentliches Merkmal des angeborenen Immunsystems ist die Produktion und Ausschüttung von Zytokinen. Zytokine sind niedrigmolekulare Proteine, die nach der Bindung an bestimmte Rezeptoren die Immunzellteilung, Zellproliferation und -aktivität beeinflussen. Sie werden nicht gespeichert, sondern während der Aktivierung der Entzündungskaskade neu erzeugt und freigesetzt. Sie wirken multifunktional, aber im wesentlichen dirigieren sie die Entzündungsreaktion auf Infektions- und Verletzungsherde und verbessern ihre Wundheilung. Grob gesagt können Zytokine in jene mit entzündungsfördernder und in jene mit entzündungshemmender Wirkung unterteilt werden. Proinflammatorische Zytokine besitzen den Tumornekrosefaktor-α (TNF-α), das Interleukin-1 (IL-1), IL-6, und lL-8. Antiinflammatorische Zytokine besitzen den IL-1 Rezeptorantagonisten, IL-10, IL-13 sowie TNF-bindende Proteine 1 und 2. Der Tumornekrosefaktor-α Während der Infektion oder nach einer Verletzung ist TNF-α ein wichtiger und wirksamer Entzündungsmediator. Er wird hauptsächlich mittels Monozyten/Makrophagen und T-Zellen synthetisiert und hat innerhalb der Zirkulation eine Halbwertszeit von <20 min. TNF-α hat im Rahmen der Zytokinkaskade eine entscheidende Funktion, weil er die Entzündungsreaktion koordiniert und die distalen Mediatoren aktiviert. Er veranlasst in hohem Maße auch die Ausschüttung anderer proinflammatorischer Zytokine. TNF-α löst beträchtliche metabolische und hämodynamische Veränderungen aus und ist imstande eine Endorgandysfunktion hervorzurufen. Außerdem ist er verantwortlich für die Koagulationsaktivierung, den Muskelkatabolismus und die Kachexie. Interleukin-1 Es existieren zwei Formen von lL-1 (IL-α and IL-β), welche die selben Zelloberflächenrezeptoren identifizieren und aus diesem Grund verschiedene biologische Aktivitäten gemeinsam haben. IL-1 wird durch Monozyten, Leukozyten und andere Zelltypen synthetisiert und hat eine Halbwertszeit von ~6 min. Beide, IL-α and IL-β, rufen metabolische und hämodynamische Veränderungen, die jenen des TNF-α ähnlich sind, hervor, aktivieren die Bildung anderer Zytokine und mildern das Schmerzempfinden dadurch, dass sie die Freisetzung von β-Endorphinen fördern. Tabelle 1 Bestandteile des Immunsystems Angeboren Primärer Abwehrmechanismus Sofortige Reaktionszeit Unspezifische Reaktion – wird infolge unterschiedlicher fremder Substanzen aktiviert Kein immunologisches Gedächtnis Bedient sich natürlicher Killer-, Mastzellen, eosenophiler, basophiler und phagozytischer Zellen Erworben Zweite Verteidigungslinie Maximale Reaktionsverzögerung Spezifische Reaktion – antigen- und pathogen-spezifische Antwort verursacht durch vorangegangenen Kontakt Das immunologische Gedächtnis reagiert mit einer verbesserten Antwort auf wiederholten Kontakt mit einem spezifischen Antigen Bedient sich antigenspezifischer B- und T-Lymphozyten Interleukin-6 TNF-α und IL-1 veranlassen die IL-6-Synthese aus vielen Zelltypen, wie etwa den Lymphozyten, Fibroblasten und Monozyten. Diese Erscheinung kann 10 Tage nach der Verletzung beobachtet werden. Dadurch wird eine Reihe von biologischen Wirkungen hervorgerufen, wie etwa die Aktivierung von Neutrophilen, die Induktion einer hepatischen Akut-Phase-Reaktion und die Aktivierung der Koagulation. Interleukin-8 IL-8 ist ein Chemokin, das von einer Vielzahl von Zellen, wie etwa der Monozyten, Leukozyten und der endothelialen Zellen, als Reaktion auf unzählige Reize, den TNF-α, das IL-1 und das Endotoxin mit eingeschlossen, gebildet wird. IL-8 verursacht keine hämodynamische Dekompensation wie sie beim TNF-α und IL- 1 beobachtet wird, aber es aktiviert Neutrophile und zieht sie zu den Entzündungsherden. Interleukin-10 Mit überwiegend antiinflammatorischer Wirkung unterdrückt IL-10 hochgradig die Aktivierung von Makrophagen und blockiert ihre Eigenschaft, proinflammatorische Zytokine zu bilden und zu sezernieren. In vitro kann das lL-10 die Bildung des TNF-α, des IL-1, IL-6 und IL-8 hemmen. Deshalb ist es als der Zytokinsynthese-Inhibitionsfaktor beschrieben worden. lnterleukin-13 Ähnlich dem IL-10, gilt auch IL-13 als ein wirksames anti-inflammatorisches Zytokin, das die Aktivierung von Makrophagen mit der daraus resultierenden proinflammatorischen Zytokinbildung und -äußerung inhibiert. Zusätzlich kann IL-13 die Stickoxidproduktion (NO) inhibieren und Neutrophile zur Bildung des IL-1-Rezeptorantagonisten anregen. Die Wirkung der proinflammatorischen Zytokine In Phasen guter Gesundheit, aber auch Krankheit wird eine sensible Balance zwischen den pro- und anti-inflammatorischen Zytokinen gewahrt. Dies wird als Zytokinbalance bezeichnet. Proinflammatorische Zytokine wirken nahe an der Stelle, an der sie freigesetzt wurden. Sobald sich jedoch die Entzündungreaktion der örtlichen Kontrolle entzieht, wird eine systemische, den ganzen Körper betreffende, Reaktion ausgelöst. Um dies auf ein Minimum zu beschränken, haben TNF-α und IL-1 eine kurze Halbwertszeit. Ein System von Membranreservoiren halten die Zytokine nahe an der Stelle ihrer Freisetzung und die antiinflammatorischen Zytokine ändern die Entzündungsreaktion des Wirts. Das Herz betreffend TNF-α kann einen sofortigen wie auch verzögerten negativen inotropischen Effekt auf das myokardiale Gewebe auslösen. Dies hat eine Dysfunktion des linken Ventrikels zur Folge. TNF-α, IL-1β und IL-6 sind an der Myokarddepression beteiligt, indem sie die Myozyten direkt beeinflussen. (Tabelle 2) Die Gefäße betreffend Proinflammatorische Zytokine haben eine signifikante Wirkung auf den vaskulären Ton, hauptsächlich durch die Leitungsbahn des Stickoxids vermittelt, woraus eine Vasodilatation resultiert. Es ist beobachtet worden, dass TNF-α und IL-1β beide die Stickoxidproduktion ankurbeln. Es wurde berichtet, dass sie auf Vasokonstriktoren nicht reagieren können, nachdem sie über eine länger andauernde Phase diesen Zytokinen ausgesetzt waren. Die Atemwege betreffend Eine inflammatorische Lungenverletzung tritt auf, wenn aktivierte Neutrophile und Makrophagen vom Lungengefäßsystem in Alveolar- und Interstitialräume wandern. Makrophagen sekretieren TNF-α, IL-1 und IL-8, die ihrerseits mithilfe von Lungenepithel- und Mesenchymzellen eine weitere Zytokinbildung anregen. Es hat sich herauskristallisiert, dass IL-8 beim Auftreten eines Akuten respiratorischen Distress-Syndroms (ARDS) eine pathogene Rolle spielt, indem es eine durch Neutrophile vermittelte Lungenverletzung verursacht. Tabelle 2 Die Wirkung proinflammatorischer Zytokine auf Organsysteme Das Herz betreffend: Negative inotropische Wirkung auf den Herzmuskel Linke ventrikuläre Dysfunktion Die Gefäße betreffend: Vasodilatation Die Atemwege betreffend: Akute Lungenverletzung Pathogene Rolle bei ARDS Die Nieren betreffend: Glomeruläre Verletzung Tubulärer Zellschaden Die Leber betreffend: Vermehrte Synthese von Akut-Phase-Proteinen Koagulation: Regulierung extrinsischer Leitungsbahnen und jener des C-reaktiven Proteins Rolle bei der Entwicklung einer disseminierten intravasalen Koagulopathie (DIC) und Thrombose Die Nieren betreffend Aus Tierversuchen geht hervor, dass TNF-α ohne vorangegangene Nierenerkrankung glomeruläre Verletzungen der Nieren verursacht. Als Reaktion auf TNF-α und IL-6 produzieren glomeruläre Zellen freie Sauerstoffradikale, ein Komplement, Arachidonsäurederivative und NO, das die lokale Entzündung mit der daraus resultierenden glomerulären und tubulären Schädigung noch weiter verstärkt. Die Leber betreffend Vorwiegend IL-6, aber auch TNF-α und IL-1β stimulieren die Leber, um ihre Synthesefunktion zu verändern und die Bildung von Akut-Phase-Proteinen, wie etwa das Serum Amyloid A, α2-Makroglobuline und das C-reactive Protein (CRP), zu steigern. Die Substrate werden für diese verstärkte Produktion mithilfe der Zytokin vermittelten Skelettmuskelstörung ermöglicht. Daher wird TNF-α alternativ auch als Kachektin bezeichnet. Koagulation Zytokine sind nicht direkt in die Koagulationsbahnen involviert, aber es ist festgestellt worden, dass TNF-α und IL-1 die äußeren Leitungsbahnen der Koagulation und die Leitungsbahn des C-reaktiven Proteins reguliert. Es gibt Anzeichen dafür, dass diese Zytokine bei der disseminierten intravasalen Koagulopathie (DIC) und der Thrombose eine wichtige Rolle spielen. Es hat sich auch herausgestellt, dass IL-1 eine Fibrinolyse inhibiert. Ältere Menschen Die Folgen, die das Altern auf das Immunsystem hat, sind offensichtlich zu einem Großteil für die Morbidität und Mortalität bei älteren Menschen verantwortlich. Die TNF-α- und die IL-6-Konzentrationen sind bei älteren Menschen erhöht. Studien haben gezeigt, dass ältere Menschen als Reaktion auf ein Operationstrauma eine verstärkte und verspätete inflammatorische Zytokinreaktion haben. Krankheiten aufgrund eines Zytokinüberschusses Die Zytokinsynthese ist gestört, wenn eine exzessive oder ständige Aktivierung von Makrophagen und Neutrophilen stattfindet. Entzieht sich dies der örtlichen Kontrolle, gelangen Zytokine in die systemische Zirkulation. Dies führt zu einer umfassenden Aktivierung von inflammatorischen Kaskaden und dem Systemischen inflammatorischen Response-Syndrome (SIRS) (Abbildung 1). Dadurch wird eine zusätzliche Freisetzung von inflammatorischen Zytokinen ausgelöst. Es wird eine Abwärtsspirale von Organdysfunktionen in Gang gesetzt und schließlich kommt es zum Multiplen Organdysfunktionssyndrom (MODS). Die durch MODS verursachte Mortalität ist hoch. Bei Patienten mit ARDS allein beträgt die Mortalität etwa 50%. Mit jedem zusätzlichen Organversagen multipliziert sich dieses Risiko. Systemisches inflammatorisches Response-Syndrome Infektiöse Erkrankungen Sepsis - gram - ve - gram + ve Nicht-infektiöse Erkrankungen Operation Trauma Hämorrhagischer Shock Ischämie-Reperfusion Pankreatitis Verbrennungen Abb. 1 Krankheiten bei Zytokinüberschuss. SIRS können entweder infektiöse oder nicht-infektiöse Erkrankungen hervorrufen. TNF-α, IL-1β IL-6 und IL-8 sind stark unter dem Verdacht, septische Mediatoren zu sein. In Studien zur Sepsis sind erhöhte Konzentrationen zirkulierender Zytokine festgestellt worden. Weiters korrelieren anscheinend erhöhte proinflammatorische Zytokinwerte prinzipiell mit dem Schweregrad der Krankheit und dem Ergebnis, wobei IL-6 am stärksten mit der Mortalität verbunden wird. Erhöhte Plasmakonzentrationen proinflammatorischer Zytokine finden sich nach größeren Operationen und das Ausmaß der zytokin-vermittelten Entzündungsreaktion hängt offensichtlich damit zusammen, wie stark der operative Insult war. Konkret beeinflussen vermutlich hohe IL-6-Plasmakonzentrationen, die nach größeren Operationen auftreten, die postoperative Mortalität. Nach einem schweren Trauma sind die Serumwerte des TNF-α, IL-1β IL-6 und IL-8 bedeutend erhöht. Es gibt offenbar eine enge Beziehung zwischen der freigesetzten Menge der proinflammatorischen Zytokine, dem Schweregrad der Verletzung und der Krankenhausmortalität. Hämorrhagie führt zu einer ausgesprochen hohen Ausschüttung von proinflammatorischen Zytokinen in der Lunge und beeinflusst den Ausbruch von ARDS, wobei TNF-α eine wichtige Rolle bei der Pathogenese einer akuten Lungenverletzung nach einer Hämorrhagie sogar nach einer entsprechenden Reanimation innehat. Antiinflammatorische Mechanismen Es gibt eine Reihe von antiinflammatorischen Mechanismen infolge welcher Entzündungsmediatoren nicht in die Zirkulation gelangen können und eine unangemessene, überschießende Entzündungsreaktion vermieden werden kann. Antiinflammatorische Zytokine, wie etwa IL-10, das TNF-bindende Protein, der IL-Rezeptorantagonist (IL- 1RA) und der Transforming Growth Factor-β (TGF-β), die infolge einer normalen Immunantwort gebildet werden, können die Freisetzung von TNF-α und anderer proinflammatorischer Zytokine hemmen. Das TNF-bindende Protein stört die Bindung von TNF an seinen Rezeptor und hemmt damit seine Wirkung. 'Stress'-Hormone, wie etwa das Glucocorticoid, das Epinephrin, Norepinephrin und das α-melanozytenstimulierende Hormon hemmen die Zytokinbildung. Es ist nachgewiesen worden, dass zusätzlich zur TNF-α-hemmenden Produktion die Stimulierung des β-Adrenergorezeptoren die Bildung des lL-10 hochreguliert, wodurch die antiinflammatorische Wirkung erhöht wird. Andere lokale Effektoren, wie etwa Prostaglandin E2, Akut-Phase-Proteine, Hitzeschockproteine, Spermin und Feutin haben alle Zusatzaufgaben, nämlich die Immunantwort zu drosseln. Die Beeinträchtigung oder der Verlust einer dieser endogenen antiinflammatorischen Leitungsbahnen ist imstande, eine normale selbst-limitierende Reaktion in eine überschießende und potentiell schädigende umzukehren. Neuronale Regulation der Immunantwort Blalock wies darauf hin, dass das Immunsystem auch als Sinnesorgan fungiert. Es ist das sechste Sinnesorgan, das Bakterien, Viren und andere potentielle schädliche Zellen entdeckt, die er üblicherweise nicht sieht, hört, schmeckt, tastet oder riecht. Diese Information würde dann an das Zentrale Nervensystem (ZNS) übertragen, um eine entsprechende physiologische Antwort auszulösen. Jüngste Forschungen haben überraschenderweise eine autonome neuronale Leitungsbahn entdeckt, welche die Entzündungsreaktion beobachtet und reguliert. Diese wird als der 'inflammatorische Reflex' bezeichnet und hat sowohl immunsensorische als auch immunsuppressive Funktion. Verglichen mit den humoralen antiinflammatorischen Mechanismen, die langsam, diffus und vom Konzentrationsgradienten abhängen, ist die Leitungsbahn des inflammatorischen Reflexes schnell, lokalisiert und integriert (Abb. 2). DMN (Dorsaler Motonukleus) Efferente Nervenbahn des Vagus NTS (Nucleus tractus solitarii) Afferente Nervenbahn des Vagus Hemmt die Zytokinausschüttung ACh α7nAChR Pathogene Ischämie Verletzung Zytokinausschüttung Makrophage Abb. 2 Der inflammatorische Reflex. Pathogene, Ischämie und andere Formen von Verletzungen verursachen eine Zytokinfreisetzung aus Immunzellen. Dies wird von afferenten Nervenfasers des Vagus wahrgenommen. Diese Information wird an den Nucleus tractus solitarii (NTS) und anschließend an den dorsalen Motonukleus weitergeleitet, wodurch der Vagusnerv aktiviert wird. Die wird als der cholinerge antiinflammatorische Weg bezeichnet. Die efferente Vagusnervaktivität hemmt die Zytokinproduktion über den nikotinischen Acetylcholinrezeptor α7 (α7nAChR), der sich in Makrophagen und anderen Immunzellen findet. Der Name Vagusnerv leitet sich von der lateinischen Bezeichnung für 'umherschweifen' ab. Er hat seinen Ursprung in der Medulla oblongata im Hirnstamm und innerviert die inneren Organe, jene des retikuloendothelialen Systems miteinbezogen, wie etwa die Leber und die Milz. Als solcher ist er perfekt angelegt, um Informationen zwischen dem Immunsystem und dem ZNS weiterzuleiten. Er besitzt sowohl sensorische (afferente) sowie motorische (efferente) Bestandteile. Sensorische Fasern leiten die Information zum Hirn, worauf Fieber sowie andere Krankheitssymptome ausgelöst werden. Der efferente Schenkel des Vagusnervs wird traditionellerweise mit physiologischen Reaktionen, wie etwa der Bradykardie, Bronchokonstriktion, erhöhter gastrischer Motilität und Myose assoziiert. Daraus geht hervor, dass der Vagusnerv eine wichtige immunomodulatorische Rolle auch bei der Erkennung und Hemmung der inflammatorischen Reaktion innehat. Die immunsensorische Funktion der afferenten Fasern des Vagusnervs Es ist nachgewiesen worden, dass proinflammatorische Zytokine Signale zum Gehirn senden. Daran können mehrere Mechanismen beteiligt sein. Eine Reihe von humoralen Mechanismen steuern die Kommunikation zwischen dem Immun- und Nervensystem. Aus Studien geht hervor, dass TNF-α, IL-1 -α, IL-1β und IL-6 die Blut-Gehirn-Schranke (BBB) mittels saturierender Transportsysteme passieren. Zytokine können auch über zirkumventrikuläre Organe ins Gehirn gelangen, wie etwa über die Area postrema, wo eine Blut-Gehirn-Schranke (BBB) fehlt. Man geht davon aus, dass sie hier ihre Produktion starten und das Prostaglandin E2 (PGE2) freisetzen. In weiterer Folge aktiviert dieses die Hypothalamus-Hypophysen-Nebennierenrinden-Achse. Proinflammatorische Zytokine, speziell die Interleukine, können an zerebrale vaskuläre Endothelzellen andocken und deren Metabolismus verändern. Dadurch werden neuroaktive Substanzen auf der Gehirnseite der Blut-Gehirn-Schranke (BBB) freigesetzt. Aber für all die vorhin erwähnten Mechanismen sind die proinflammatorischen Zytokine in der Zirkulation erforderlich, damit diese ins Gehirn gelangen können. Es ist jedoch festgestellt worden, dass Entzündugsreaktionen auch ohne das Vorhandensein nachweisbarer Zytokinwerte im Blut auftreten können. Sensorische afferente Vagusfasern können jedoch niedrige Zytokinwerte und andere Entzündungsmediatoren aufspüren und über die aufsteigenden Fasern das Gehirn von der Entzündung an der Peripherie informieren. In der Tat reichen mitunter lokale Zytokinkonzentrationen im Gewebe aus, damit die sensorischen afferenten Vagusfasern aktiviert werden. Nicht immer ist ein systemischen Anstieg der Zytokinwerte erforderlich. Es ist nachgewiesen worden, dass subdiaphragmatische Vagotomie das Auftreten von Fieber bei Tieren, denen entweder niedere Intraabdominal-Dosen des IL-1 oder des Endotoxins verabreicht wurden, verhindert. Das Gehirn empfängt einfach nicht die Botschaft, dass hier eine Entzündung vorliegt. Es ist auch gezeigt worden, dass Vagus-Neuronen sensorischer Natur das IL-1 und den PGE2 Rezeptor mRNA wie auch IL-1 Bindungsstellen an Glomuszellen, die innerhalb der mit dem Vagusnerv verbundenen Paraganglien lokalisiert sind, exprimieren. Die Einbindung der afferenten und efferenten Bestandteile des inflammatorischen Reflexes findet hauptsächlich im medullären Nucleus tractus solitarii (NTS) statt, dem Hauptausgangspunkt der afferenten Fasern des Vagusnervs. Die NTS-Neuronen projizieren dann zum dorsalen Motonukleus des Vagus (DMN), wo die meisten efferenten präganglionischen Fasern des Vagusnervs ihren Ursprung haben. Die immunsuppressive Funktion der efferenten Fasern des Vagusnervs Der efferente Schenkel des antiinflammatorischen Reflexes ist als der 'cholinerge antiinflammatorische Weg' bezeichnet worden, weil Acetylcholin (ACh) der wichtigste Neurotransmitter des Vagusnervs ist. Makrophagen und andere Zytokin produzierende Zellen exprimieren ACh-Rezeptoren und werden inaktiviert, wenn sie dem ACh ausgesetzt sind. Studien zeigen, dass ACh posttranskriptionell die TNF-Synthese unterdrückt und die Ausschüttung von IL-1β, IL-6 und IL-8 hemmt, ohne die Freisetzung des antiinflammatorischen Zytokins IL-10 zu verhindern. ACh-Rezeptoren werden in zwei Klassen unterteilt: die Muskarin- und die Nikotinrezeptoren. Nikotinrezeptoren kann man des weiteren in α-Bungarotoxin empfindliche (das wiederum in das α1, α7 und α9 klassifiziert wird) und nichtempfindliche untergliedern. Sowohl die Muskarin- wie auch die Nikotinrezeptoren sind im ZNS und der Peripherie verteilt, jedoch auf verschiedenen synaptischen Standorten und mit vielfältigen Aufgaben bezüglich der cholinergen Transmission. Erste Studien zeigten, dass ACh die α-Bungarotoxin empfindlichen Nikotinrezeptoren menschlicher Makrophagen beeinflusst, aber diese spezifische Rezeptorenuntergruppe war bis unlängst unbekannt. Weiterführende Forschungen haben jedoch diese Rezeptorengruppe als den nikotinischen α7-ACh-Rezeptor (α7nAChR) identifiziert. Es hat sich gezeigt, dass eine elektrische Stimulation des Vagusnervs die TNF-Synthese in den Makrophagen von Wildmäusen hemmt, allerdings nicht bei Mäusen mit einem α7-Mangel. Daraus geht hervor, dass α7nAChR ein essenzieller peripherer Baustein des cholinergen antiinflammatorischen Wegs ist. Obwohl muskarinische Acetylcholinrezeptoren auch an Makrophagen und anderen Zytokin produzierenden Zellen exprimieren, bremst die Blockade der peripheren Muskarinrezeptoren nicht die antiinflammatorische Signalisierung des Vagusnervs. Anscheinend sind periphere Muskarinrezeptoren für die Zytokinmodulation des cholinergen antiinflammatorischen Weg nicht erforderlich. Unlängst ist dargelegt worden, dass zentrale Muskarinrezeptoren bei der Hemmung der systemischen Entzündung eine Rolle spielen. Es hat sich herausgestellt, dass die zentrale cholinerge Aktivierung durch muskarinische Rezeptorliganden bei endotoxämischen Ratten signifikant den systemischen TNF hemmt und die efferente Aktivität des Vagusnervs aktiviert. Bakterien und Endotoxin bleiben in erster Linie auf die Makrophagen in der Milz und Leber beschränkt. Aus Studien geht hervor, dass während der Endotoxämie hauptsächlich in der Milz die systemische TNF-Produktion stattfindet. Jüngste Forschungen zeigen, dass der cholinerge antiinflammatorische Weg funktionell fest mit der Milz über den gewöhnlichen Vagusast des Plexus coeliacus verdrahtet ist. Dieser vom α7nAChR abhängige Weg des Vagusnervs zur Milz kann die proinflammatorische Zytokinproduktion während der lethalen Endotoxämie und der polymikrobiellen Sepsis hemmen. Auch die Aktivität des Vagusnervs kann eine systemische humorale antiinflammatorische Antwort durch die Übertragung der Information an die medulläre retikuläre Formation, den Hypothalamus und den Locus caeruleus, auslösen. Daraus resultiert die Freisetzung des Adrenocorticotropins (ACTH) aus dem Hypophysenvorderlappen. Beweise für die Immunmodulation durch den Vagusnerv Den überzeugendsten Beweis dafür, wie wichtig der Vagusnerv bei der Modulation der Entzündungsreaktion ist, liefern Untersuchungen an Ratten, denen intravenös die tödliche Dosis Endotoxin verabreicht wurde. Die elektrische Stimulation der efferenten Fasern des Vagusnervs verringerte erheblich das TNF-α-Serum, reduzierte die Synthese des TNF-α in der Leber und milderte das Auftreten eines endotoxischen Schocks milderte. Im Gegensatz dazu führte eine Vagotomie zu wesentlich erhöhten TNF-α-Serumwerten in der Leber und beschleunigte die Ausbildung eines Schocks. Im Rahmen der Forschung wurde die Bedeutung des cholinergen antiinflammatorischen Wegs während einer Infektion und nicht während einer sterilen Entzündung ermittelt. Es wurde bei Mäusen mittels einer Intraperitonealinjektion von lebenden Escherichia coli eine abdominelle Sepsis hervorgerufen. In dieser Studie führte die zervikale Vagotomie zu einer verminderten Zytokinfreisetzung, einem Anstieg an Entzündungszellen, die an das Peritoneum rekrutiert waren und zu einem ausgedehnten Leberschaden. Die Aktivierung der peripheren Komponente des cholinergen antiinflammatory Wegs durch Nikotin verursachte eine erhöhte Zytokinproduktion und einen reduzierten Zustrom von Entzündungszellen. Ein Leberschaden blieb aus. Andere Untersuchungen zeigen, dass eine Stimulation des Vagusnervs (VNS) bei der Endotoxämie von Ratten die Aktivierung einer Koagulation und Fibrinolyse hemmt. Im Rahmen der Entzündung unterdrückt die cholinerge Stimulation die Rekrutierung von Leukozyten und die Aktivierung von endothelialen Zellen. Es wurde auch untersucht, wie der Vagusnerv die Entzündung beim hämorrhagischen Schock reguliert. Die VNS erhöhte beim tödlichen hämorrhagischen Schock von Ratten wesentlich die Überlebensrate, schützte gegen das Auftreten einer Hypotonie und verringerte TNF-α-Serumkonzentrationen. ACTH aktiviert den cholinergen antiinflammatorischen Weg. Es ist gezeigt worden, dass ACTH die kardiovaskulären und respiratorischen Funktionen verbessert, den Schockzustand beseitigt und die Überlebensrate beim hämorrhagischen Schock in weiteren Tierversuchen erhöht. Die VNS bei Tieren, bei denen durch einen vorübergehenden Aortenverschluss eine Ischämie/Reperfusion herbeigeführt wurde, milderte wesentlich die Synthese des TNF-α und das Auftreten eines Shocks. In einem Versuchsmodell verursachte die Stimulation des Vagusnervs bei einer ischämischen Herzerkrankung während einer Myokardischämie/Reperfusion eine Verringerung der Werte freier Radikale und reduzierte bedeutend die Fälle schwerer Arrhythmien und die Letalitätsrate. Der cholinerge antiinflammatorische Weg hat auch bei der Hemmung lokal auftretender Entzündung eine besondere Bedeutung. Bei lokaler Entzündung milderte in einem Versuchsmodell die VNS die Entzündungsreaktion und blockierte die Schwellung von Pfoten bei muriner Arthritis von Labormäusen. Mögliche therapeutische Auswirkungen der Immunmodulation des Vagusnervs Mit der Entdeckung des cholinergen antiinflammatorischen Wegs bietet sich die Chance zur Entwicklung von Therapien, die darauf abzielen, die Entzündungsreaktion zu regulieren. Die Einschränkung der proinflammatorischen Zytokinproduktion kann entweder durch die Veränderung der Aktivität des Vagusnervs oder durch das Targeting spezifischer Teile des Weges erreicht werden. Die Stimulation des Vaguservs Direkte elektrische Stimulation des Vagusnervs verringert die Produktion des TNF-α bei Versuchsmodellen im Falle der Endotoxämie, des hämorrhagischen Shocks und anderer Erkrankungen, die durch einen Zytokinüberschuss ausgelöst werden. Die Voltzahl und die Stimulationsfrequenz, die dazu dienen, den cholinergen antiinflammmatorischen Weg zu aktivieren, sind unter dem Schwellenwert, der benötigt wird, um die vagalen Herzfasern zu aktivieren. Aus diesem Grund sind keine wesentlichen Folgen für die Herzfrequenz beobachtet worden. Die VNS ist in den USA von der FDA, der amerikanischen Arzneimittelzulassungsbehörde, zur Behandlung von refraktorischer Epilepsie seit mehr als 10 Jahren zugelassen und wird derzeit in klinischen Experimenten bei resistenter Depression getestet. Die VNS erfordert die Implantation eines kleinen pacemaker-ähnlichen Geräts, das sicher, effizient und gut verträglich ist. Das VNS-Gerät wird durch einen Schnitt in den Nacken eingesetzt, wobei Drähte an den Vagusnerv gelegt werden. Ein elektrisches Element wird unterhalb des Schlüsselbeins implantiert. Komplikationen infolge des Implantationseingriffs sind atypisch. Häufig berichtete Nebenwirkungen sind Heiserkeit, eine veränderte Stimme, Hals- Kopf- oder Nackenschmerzen, Husten und Dyspnoe. Diese Nebenwirkungen treten hauptsächlich während der Stimulation auf, können jedoch durch eine Stimulationsveränderung verringert werden und lassen mit der Zeit nach. Überraschenderweise waren die Werte der zirkulierenden pro- und antiinflammatorischen Zytokine in einer Studie von VNS-Patienten mit einer resistenten Depression deutlich erhöht. Weiters war der Anstieg, wie ihn die CRP-Werte zeigten, kaum eine nicht-spezifische Entzündungsreaktion. Transkutane nicht-elektrische VNS kann die Serumwerte des TNF-α verringern und die Überlebenschancen von Labormäusen bei muriner Sepsis verbessern. Bei der erwähnten Studie wurden wache Mäuse mit transkutaner Stimulation durch die Veränderung des Direktdruckes senkrecht direkt neben dem Rand der Trachea behandelt. Schwerkranke Patienten sind meistens schlechte Operationskandidaten, sodass die Möglichkeit, den Vagusnerv über den transkutanen Pfad zu stimulieren, klinisch wertvoll sein kann. Eine Massage des Sinus caroticus kann durchgeführt werden, um Tachyarrhythmien durch die Aktivierung des efferenten Vagusnervreflexes zu beenden. Weitere Daten werden benötigt, damit die immunologische Effizienz dieser Methode beurteilt werden kann. Zusätzlich könnten Akupunkturtechniken angewendet werden, um die Immunomodulatorik des Vagusnervs zu stimulieren. Aus Studien geht hervor, dass Akupunktur gegebenenfalls einen messbaren Anstieg der Aktivität des Vagusnervs verursacht, sodass es wenigstens theoretisch möglich erscheint, dass auch Akupunktur den cholinergen antiinflammatorischen Weg aktiviert und die Immunantwort reguliert. Studien haben gezeigt, dass auch verschiedene Formen von Meditation den parasympathischen Bereich hinsichtlich der Herzfrequenzschwankungen verändern und vermutlich den autonomen Herzton positiv beinflussen können. Ähnlich können Entspannungstherapien und Biofeedback den parasympathischen Einfluss auf Herzfrequenzschwankungen signifikant erhöhen, was sich in einem Anstieg der Aktivität des Vagusnervs wiederspiegelt. Pharmakologische Agenzien Es sind einige potentielle Therapeutika entwickelt worden, die auf dem Targeting des cholinergen antiinflammatirischen Wegs beruhen, und zwar werden dabei zentrale Muskarinrezeptoren oder α7nACh-Rezeptoren in der Peripherie aktiviert. CNI-1493 Es ist gezeigt worden, dass CNl-1493, eine vierwertige Quanylhydrazonverbindung und ein wirksames antiinflammatorisches Agens und bedeutender Aktivator der cholinergen antiinflammatorischen Bahn aufgrund der Interaktion mit zentralen Muskarinrezeptoren ist. CNI-1493 hemmt die proinflammatorische Ausschüttung der Zytokine aus Monozyten und Makrophagen. Eine systemische Verabreichung ist bei der Behandlung von Tieren mit Endotoxämie und Sepsis im Versuchsmodell wirksam gewesen. Weiters ist beobachtet worden, dass eine intakte Nervenbahn des Vagus im ZNS notwendig ist, damit CNI-1493 nach der Verabreichung antiinflammatorisch wirken und eine verstärkte Aktivität des Vagusnervs stattfinden kann. Nikotin Nikotin ist ein relativ unspezifischer α7nAChR-Agonist, der nachgewiesenermaßen wesentlich TNF-α und andere proinflammatorische Zytokine aus mit Endotoxin stimulierten menschlichen Makrophagen hemmt. Darüber hinaus unterdrückt Nikotin auch die Ausschüttung des High-Mobility-Group-Proteins B1 (HMGB1) aus den menschlichen Makrophagen. Das HMGB1-Protein ist ein später Mediator der tödlich verlaufenden systemischen Entzündung bei Sepsis und der in Tiermodellen durch Versuche hervorgerufenen Sepsis. Durch die Behandlung mit Nikotin konnten die HMGB1-Serumwerte herabgesetzt werden, die klinische Manifestation der Sepsis gesenkt werden und die Überlebenschancen bei ausgeprägter Sepsis verbessert werden. Mit einer Nikotingabe konnte der Tod abgewendet werden. Außerdem wurde Nikotin erst 24 h nach der Induktion einer tödlich verlaufenden Peritonitis bei Mäusen verabreicht, woraus man herauslesen kann, dass der cholinerge antiinflammatorische Weg die Entzündungsantwort sogar bei schwerer Sepsis regulieren kann. Die Nikotinverabreichung schien einen Langzeitschutz zu bieten, denn keines der Tiere verendete im Nachhinein. Eine Entwicklung von Nikotin als therapeutische Intervention ist wegen seiner toxischen Nebenwirkungen und pharmakologischen Unspezifität eingeschränkt. Eine Studie zur Nikotinersatztherapie bei Rauchern, die stationär zu intensivmedizinischer Betreuung zugelassen wurden, ergab, dass eine Nikotinersatztherapie mit einer höheren Sterblichkeitsrate einher geht. Diese Forschungsergebnisse zeigen jedoch, dass α7nAChR-Agonisten möglicherweise als eine neue Klasse antiinflammatorischer Medikamente bedeutsam sind. GTS-21 Ist auch als DMXB oder DMXBA bekannt. GTS-21 ist ein gezielter α7nAChR-Agonist und wird im Gegensatz zu Nikotin von gesunden männlichen Probanden ohne wesentliche klinische Nebenwirkungen gut vertragen. GTS-21 hat sowohl die Serumwerte von TNF-α als auch HMGB1 verringert und bei der Sepsis von Mäusen deren Überlebenschancen verbessert. Ghrelin Ghrelin ist ein endogener Ligand des Wachstumshormon-Sekretagog-Rezeptors (GHSR). Die biologische Wirkung von Ghrelin wird wahrscheinlich durch GHSR erzielt. GHSR wird weitreichend zentral und peripher verteilt und findet sich in afferenten Neuronen des Ganglions nodosum. Nahe liegende Ghrelinsignale werden mittels vagaler afferenter Fasern an das Gehirn weitergeleitet. Aus Tiermodellen geht hervor, dass die Ghrelinkonzentration bei Sepsis in der Anfangs- und Schlussphase reduziert wird, obwohl der Ghrelinrezeptor in der Anfangsphase überaus aktiv ist. Es ist auch gezeigt worden, dass die vaskuläre Sensibilität bei Tieren aufgrund einer Stimulation durch Ghrelin während der hyperdynamen Phase der Sepsis erhöht ist. In einer Studie mit Ratten wurde gezeigt, dass eine Behandlung mit Ghrelin bei Sepsis die Serum- und Peritonealflüssigkeitskonzentrationen von TNF-α und IL-6 bedeutet verringert, wobei zusätzliche Forschungsergebnisse zeigten, dass ein intakter Vagusnerv für die Hemmwirkung des Ghrelin auf die Ausschüttung proinflammatorischer Zytokine essenziell ist. Die Aktivierung und Manipulation der cholinergen antiinflammatorischen Bahn zur Entzündungshemmung kann sowohl in Fällen von Zytokinüberschuss von Vorteil sein, aber auch bei Patienten mit einem Schädel-Hirn-Trauma (SHT) eine Rolle spielen. Das Auftreten einer Infektion ist eine häufige und ernste Komplikation nach einem SHT und das Phänomen einer posttraumatischen Immunparalyse ist wahrscheinlich für die erhöhte Infektionsrate bei diesen Patienten verantwortlich. Aus Studien geht hervor, dass in der akuten Phase nach einem SHT die Immunantwort stark beeinträchtigt und die vagale Aktivität verstärkt ist. Dies ist vermutlich die Folge des erhöhten intrakranialen Drucks. Aufgrund dieser Beobachtungen ist die Annahme entstanden, dass das steigende Risiko einer Infektion nach einem SHT auf der vagal verursachten Suppression der Immunantwort beruht und das Eingreifen in den cholinergen antiinflammatorischen Weg eine Chance ist, betreffenden Patienten helfen zu können. Resümee Trotz der jüngsten Fortschritte in der medizinischen Intersivbehandlung kann das Multiorganversagen (MODS) wegen einer überschießenden und prolongierten proinflammatorischen Zytokinausschüttung in Fällen wie etwa der Sepsis, Ischämie/Reperfusion und dem hämorrhagischen Shock Ursache für eine hohe Sterblichkeitsrate sein. Proinflammatorische Zytokine, wie zum Beispiel TNF-α und IL-1β sind als 'frühzeitige' Mediatoren der Entzündungsreaktion identififiziert worden. Sofern sie durch den Einsatz spezifischer Antikörper neutralisiert werden, können sie einen septischen Schock, wie in Tiermodellen gezeigt wurde, verhindern. Leider sind diese Methoden nicht in effiziente Therapien für schwer kranke Patienten umgesetzt worden. Studien haben gezeigt, dass TNF-α-Antikörper unwirksam sind, wenn der Behandlungsbeginn nach der Reinigung des TNF-α-Serums erfolgt. Aufgrund der Identifizierung des HMGB1, das 'spät' als Entzündungsmediator auftritt, können therapeutische Agenzien entwickelt werden, die speziell auf das HMGB1 abzielen und das therapeutische Zeitfenster vergrößern. Nun, da die immunmodulatiorische Funktion des Vagusnervs entdeckt wurde, bietet sich die Chance zur Entwicklung neuer Therapeutika zur Behandlung von entzündlichen Erkrankungen. Die Kenntnisse über die immunsuppressive Funktion der efferenten Fasern des Vagusnervs, was als der 'cholinerge antiinflammatorische Weg' bezeichnet wird, eröffnen einen neuen Zugang im Zusammenhang mit der Regulierung der proinflammatorischen Zytokinproduktion. Es hat sich herausgestellt, dass die VNS die Zytokinproduktion verringert, das Aftreten des Schocks mildert und die Überlebenschancen bei Tieren mit einer Sepsis, einem hämorrhagischen Shock und einer Ischämia/Reperfusion erhöht. Der Vagusnerv kann direkt stimuliert werden, was bei refraktärer Epilepsie bereits Anwendung findet. Dabei handelt es sich jedoch um eine invasive Technik. Andere Methoden, wie zum Beispiel die transkutane Stimulation, verbessern die Überlebenschancen bei muriner Sepsis. Es muss noch untersucht werden, ob diese Methoden auch bei Menschen mit SIRS und MODS zur Behandlung eingesetzt werden können. Akupunktur, Meditation, Biofeedback und andere Formen alternativer Therapien verbessern dieVagusnervaktivität. Es wird interessant sein herauszufinden, ob diese Methoden den cholinergen antiinflammatorischen Weg aktivieren können. Der Vagusnerv kann chemisch stimuliert werden. Durch die Identifizierung bestimmter Teile des cholinergen antiinflammatorischen Wegs sind neue Möglichkeiten zur Entwicklung vielversprechender neuer Therapieansätze wie etwa Mittel, die auf wichtige Muskarinrezeptoren und bestimmte α7nAChR-Agonisten wirken, eröffnet worden. Es ist angeregt worden, eine neue Klasse antiinflammatorischer Präparate basierend auf dem GTS-21-Molekül zu synthetisieren. Auch gängige Arzneimittel, zum Beispiel Cholinergika, Anticholinergika, nicht-steroidale Antiphlogistika und Amiodarone, die alle auf die Aktivität des Vagusnervs Einfluss nehmen, können den cholinergen antiinflammatorischen Weg beeinflussen, was früher nicht erkannt wurde. Die Entdeckung der immunmodulatorischen Funktion des Vagusnervs stellt ein spannendes Unterfangen in der Entwicklung neuer Therapeutika zur Regulierung der Immunantwort dar. Es ist zwar unwahrscheinlich, dass die VNS die Standardbehandlung in der Internsivmedizin ersetzen wird, kann jedoch eine Bedeutung in der Begleittherapie haben und zukünftigen Patienten zum Vorteil gereichen.