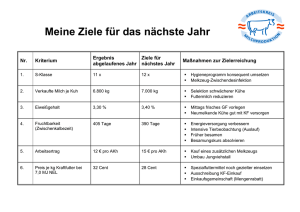
Einfluss einer wiederholten Applikation von Gonadotropin
Werbung

Tierärztliche Hochschule Hannover Einfluss einer wiederholten Applikation von Gonadotropin - Releasing Hormon auf die Fruchtbarkeit azyklischer Milchkühe INAUGURAL - DISSERTATION zur Erlangung des Grades einer Doktorin der Veterinärmedizin - Doctor medicinae veterinariae ( Dr. med. vet. ) vorgelegt von Hanna Bläsing Kassel Hannover 2011 Wissenschaftliche Betreuung: 1. Gutachterin: Univ.- Prof. Dr. Martina Hoedemaker, Ph. D. Klinik für Rinder Univ.- Prof. Dr. Martina Hoedemaker, Ph. D. 2. Gutachterin: Apl. Prof. Dr. Dagmar Waberski Tag der mündlichen Prüfung: 17.11.2011 Diese Arbeit wurde durch Mittel der Fa. Intervet Deutschland GmbH unterstützt. Meinen Eltern Inhaltsverzeichnis 1 Einleitung 13 2 Schrifttum 15 2.1 Sexualzyklus beim Rind 15 2.1.1 Endokrine Regulation des Zyklus 16 2.2 Hormonelle Aktivität post partum 18 2.2.1 FSH/LH und Östrogene 18 2.2.2 Progesteron 20 2.2.3 Ovartätigkeit post partum 21 2.2.4 Follikelwachstum und erste Ovulation/erste Brunst post partum 22 2.3 Postpartaler Anöstrus - unterschiedliche Definitionen 25 2.3.1 Anöstrus 26 2.3.2 Suböstrus/unbeobachtete Brunst/stille Brunst 26 2.3.3 Azyklie/eigentlicher Anöstrus/Ovardystrophie/anovulatorische Kühe 27 2.4 Einflüsse auf das Wiedereinsetzen der Ovarfunktionen post partum bzw. das Entstehen von Azyklie und Anöstrie 27 2.4.1 Einfluss von Fütterung und Körperkondition 27 2.4.2 Einfluss von Laktation/Milchleistung/säugendem Kalb 29 2.4.3 Einfluss von Krankheiten 30 2.4.4 Einfluss der Jahreszeit 31 2.4.5 Einfluss der Anzahl der Abkalbungen 31 2.4.6 Einfluss der Trockenstehzeit 32 2.4.7 Einfluss sonstiger Faktoren 32 2.5 GnRH als körpereigenes Hormon und Freisetzung von FSH/LH 33 2.5.1 GnRH 33 2.5.1.1 Verschiedene Formen von GnRH 33 2.5.1.2 GnRH-Rezeptor 34 2.5.1.3 Beeinflussung der GnRH-Sekretion auf Ebene des Hypothalamus 34 2.5.2 Synthese und Sekretion von FSH und LH 34 2.5.3 Unterschiede in der Regulation von FSH und LH 2.5.4 Einfluss von Östradiol, Progesteron und anderen Faktoren auf die Freisetzung von FSH/LH 2.6 35 37 Therapeutischer Einsatz von GnRH-Analoga/Beeinflussung der LH-Freisetzung 38 2.6.1 GnRH als Medikament 38 2.6.2 Therapie der Azyklie 41 2.6.3 Therapie von Anöstrus/Suböstrus 42 2.6.4 Therapie von Anöstrie und Azyklie ohne den Einsatz von GnRH 43 3 Material und Methode 45 3.1 Zielsetzung 45 3.2 Auswahl der Betriebe, Auswahl und Untersuchung der Versuchstiere 45 3.3 Weitere Datenerfassung 49 3.4 Statistische Auswertung 50 4 Ergebnisse 51 4.1 Gesamtpopulation und Stichprobenumfang 51 4.2 Ergebnisse in den Versuchsgruppen 53 4.2.1 Anzahl der Kühe mit Gelbkörper am Tag 65 oder 75 p.p. in den Versuchsgruppen 53 4.2.2 Rastzeit, Güstzeit und Verzögerungszeit in den Versuchsgruppen 54 4.2.3 Mittlere Anzahl der Besamungen in den Versuchsgruppen 55 4.2.4 Erstbesamungserfolg (EBE) in den Versuchsgruppen 57 4.2.5 Gesamtträchtigkeitsraten und Trächtigkeitsindizes in den Versuchsgruppen 57 4.2.5.1 Gesamtträchtigkeitsraten in den Versuchsgruppen 58 4.2.5.2 Trächtigkeitsindizes in den Versuchsgruppen 59 4.2.6 Anzahl besamter Tiere, Untersuchungen und Behandlungen am 4.3 Tag 100 post partum 60 Ergebnisse in Milchleistungsgruppen 62 4.3.1 Aufteilung der Kühe in Leistungsgruppen 4.3.2 Anzahl der Tiere mit Gelbkörper am Tag 65 oder 75 p.p. in den 62 Leistungsgruppen 62 4.3.3 Rastzeit, Güstzeit und Verzögerungszeit in den Leistungsgruppen 64 4.3.4 Mittlere Anzahl der Besamungen in den Leistungsgruppen 65 4.3.5 Erstbesamungserfolg in den Leistungsgruppen 66 4.3.6 Gesamtträchtigkeitsraten in den Leistungsgruppen 67 4.3.7 Trächtigkeitsindizes in den Leistungsgruppen 68 4.4 Ergebnisse der Färsen und Kühe 69 4.4.1 Aufteilung der Tiere nach Laktationsnummer und Versuchsgruppe 69 4.4.2 Anzahl der Färsen und Kühe mit Gelbkörper am Tag 65 oder 75 post partum 70 4.4.2.1 Vergleich der Färsen und Kühe aller Versuchsgruppen zusammen 70 4.4.2.2 Vergleich der Erstlaktierenden und der Mehrkalbskühe innerhalb der Versuchsgruppen 70 4.4.2.3 Vergleich der Erstlaktierenden der Versuchsgruppen untereinander 71 4.4.2.4 Vergleich der Mehrkalbskühe der Versuchsgruppen untereinander 72 4.4.3 Vergleich der Rastzeit, Güstzeit und Verzögerungszeit von Färsen und Kühen 72 4.4.3.1 Vergleich der Färsen und Kühe aller Versuchsgruppen zusammen 72 4.4.3.2 Vergleich der Rastzeit, Güstzeit und Verzögerungszeit der Färsen und Kühe innerhalb der Versuchsgruppen 4.4.3.3 Vergleich der Rastzeit, Güstzeit und Verzögerungszeit der Färsen der Versuchsgruppen untereinander 4.4.3.4 73 75 Vergleich der Rastzeit, Güstzeit und Verzögerungszeit der Mehrkalbskühe der Versuchsgruppen untereinander 75 4.4.4 Anzahl der Besamungen bei Färsen und Kühen 75 4.4.4.1 Vergleich der Färsen und Kühe aller Versuchsgruppen zusammen 75 4.4.4.2 Vergleich der Anzahl an Besamungen bei Färsen und Kühen innerhalb der Versuchsgruppen 76 4.4.4.3 Vergleich der Anzahl der Besamungen bei den Färsen der Versuchsgruppen untereinander 4.4.4.4 78 Vergleich der Anzahl der Besamungen bei den Mehrkalbskühen der Versuchsgruppen untereinander 78 4.4.5 Erstbesamungserfolg der Färsen und Kühe 78 4.4.5.1 Erstbesamungserfolg der Färsen und Kühe in den Versuchsgruppen 79 4.4.5.2 Erstbesamungserfolg der Färsen und Kühe zwischen den Versuchsgruppen 80 4.4.6 Gesamtträchtigkeitsraten der Färsen und Kühe 80 4.4.6.1 Vergleich der Gesamtträchtigkeitsraten der Färsen und Kühe innerhalb der Versuchsgruppen 4.4.6.2 81 Vergleich der Gesamtträchtigkeitsraten der Färsen und Kühe zwischen den Versuchsgruppen 81 4.4.7 Trächtigkeitsindizes der Färsen und Kühe 82 4.4.7.1 Trächtigkeitsindizes der Färsen und Kühe innerhalb der Versuchsgruppen 4.4.7.2 83 Trächtigkeitsindizes der Färsen und Kühe zwischen den Versuchsgruppen 83 4.5 Ergebnisse in Körperkonditionsgruppen 83 4.5.1 Aufteilung der Tiere nach Körperkondition 83 4.5.2 Anzahl der Tiere mit Gelbkörper am Tag 65 oder 75 post partum in den BCS-Gruppen 84 4.5.3 Rastzeit, Güstzeit und Verzögerungszeit in den BCS-Gruppen 85 4.5.4 Mittlere Anzahl der Besamungen in den BCS-Gruppen 87 4.5.5 Erstbesamungserfolg in den BCS-Gruppen 88 4.5.6 Gesamtträchtigkeitsraten in den BCS-Gruppen 88 4.5.7 Trächtigkeitsindizes in den BCS-Gruppen 89 5 Diskussion 91 5.1 Gesamtpopulation und Stichprobenumfang 91 5.2 Ergebnisse in den Versuchsgruppen 93 5.3 Aufteilung der Kühe nach Milchleistung 97 5.4 Aufteilung der Tiere nach Laktationsnummer 98 5.5 Aufteilung nach Körperkondition 99 5.6 Schlussfolgerung 101 6 Zusammenfassung 102 7 Summary 105 8 Literaturverzeichnis 108 Verzeichnis der verwendeten Abkürzungen BCS Body Condition Score bzw. beziehungsweise ca. circa cAMP cyclisches Adenosinmonophosphat CFI interval from calving to first insemination CIDR Controlled Internal Drug Release Cl Corpus luteum cm Zentimeter DO days open EBE Erstbesamungserfolg eCG equines Choriongonadotropin engl. Englisch et al. et alii Fa. Firma FCM fat corrected milk FIC interval from first insemination to conception FSC first service conception rate FSH Follikestimulierendes Hormon FSP FSH-Suppressing-Protein GABA Gamma-Aminobuttersäure GnRH Gonadotropin-Releasing Hormon GTR Gesamtträchtigkeitsrate GZ Güstzeit hCG humanes Choriongonadotropin HF Holstein Friesian IE Internationale Einheiten i.m. intramuskulär KB künstliche Besamung kg Kilogramm LH Luteinisierendes Hormon LH-RH Luteinisierendes Hormon-Releasing Hormon µg Mikrogramm mg Milligramm ml Milliliter mRNA messenger Ribonucleic acid ng Nanogramm OvSynch Ovulationssynchronisation PGF2α Prostaglandin F2α PI pregnancy index PMSG Preganant Mare's Serum Gonadotropin p.p. post partum PPI postpartales Intervall PR pregnancy rate PRID Progesterone releasing intravaginal device RZ Rastzeit SD Standard Deviation sog. sogenannte SPSS Statistical Package for the Social Science T. Tag Tab. Tabelle TI Trächtigkeitsindex TMR Total Mixed Ration TU Trächtigkeitsuntersuchung u. und UBB unbeobachtete Brunst vs. versus VZ Verzögerungszeit z. B. zum Beispiel 1 Einleitung In Milchviehbetrieben spielt die Fruchtbarkeit der Kühe eine große Rolle. Es ist wichtig, dass die Kühe nach der Abkalbung schnell wieder tragend werden, um eine ökonomisch sinnvolle Zwischenkalbezeit zu erreichen. Die Kühe müssen also nach kurzer Zeit wieder eine Brunst zeigen, damit sie besamt werden können. Nach der Abkalbung folgt aber zuerst eine unterschiedlich lange Phase der ovariellen Inaktivität bis die zyklische Eierstockstätigkeit wieder einsetzt und die Kuh wieder äußerlich erkennbare Brunstsymptome zeigt (OPSOMER u. DE KRUIF 1999). Bei vielen Kühen wird aber auch im weiteren Lauf der Laktation keine Brunst beobachtet. Es ist somit schwierig zu erkennen, wann die Kuh besamt werden kann. Bei Brunstlosigkeit muss unterschieden werden zwischen Anöstrie und der Azyklie im Sinne einer Ovardystrophie. Bei der Anöstrie läuft ein ovarieller Zyklus ab, die Kuh zeigt allerdings keine äußeren Brunstsymptome (sog. stille Brunst). Einige Kühe zeigen nur für kurze Zeit oder sehr schwach ausgeprägte Brunstsymptome und werden fälschlich als anöstrische Tiere eingeordnet. Bei der Azyklie im Sinne einer Ovardystrophie findet neben dem Fehlen der äußeren Brunstanzeichen auch keine zyklische Ovartätigkeit statt (GRUNERT 1999; MAYER et al. 1987). Seit vielen Jahren werden zur Behandlung von Fruchtbarkeitsstörungen bei Kühen das Hormon Gonadotropin-Releasing Hormon (GnRH) oder dessen synthetische Analoga verwendet. Das körpereigene Hormon GnRH wird vom Hypothalamus pulsatil sezerniert und bewirkt die Ausschüttung der für den Sexualzyklus wichtigen Hormone Follikelstimulierendes Hormon (FSH) und Luteinisierendes Hormon (LH) (PETERS 2005). In dieser Arbeit soll untersucht werden, ob die Azyklie bei Milchkühen mit dem GnRHAnalog Buserelin therapiert werden kann. Dabei werden die Kühe einer Versuchsgruppe einmal behandelt. Den Kühen einer zweiten Versuchsgruppe wird das Medikament in kleinen Dosen an fünf aufeinanderfolgenden Tagen verabreicht. Dabei soll untersucht werden, ob die häufige Applikation kleiner Mengen von Buserelin ähnlich wie die körpereigene pulsatile Sekretion von GnRH wirkt und so einen besseren Effekt auf die Fruchtbarkeit der azyklischen Milchkühe hat als die einmalige Injektion von Buserelin. 13 Es werden von den behandelten Tieren und von Kühen einer Kontrollgruppe verschiedene Fruchtbarkeitsparameter ausgewertet, um den Erfolg der Behandlungsschemata bewerten und vergleichen zu können. 14 2 Schrifttum 2.1 Sexualzyklus beim Rind Rinder europäischer Nutzungsrassen sind ganzjährig polyöstrische Tiere. Die Brunstzyklen treten regelmäßig über das Jahr verteilt auf, wobei ein Zyklus durchschnittlich 21 (18 bis 24) Tage dauert (CHRISTENSEN et al. 1974; GARVERICK u. SMITH 1993; EVANS et al. 1997; GRUNERT 1999). Im Laufe dieser Zeit kommt es zu Änderungen im Verhalten sowie an den äußeren und inneren Geschlechtsorganen. Abhängig von Art und Ort der Veränderungen unterscheidet man deshalb den äußeren Zyklus vom ovariellen und vom Schleimhautzyklus. Zeitlich wird der Zyklus in vier Phasen gegliedert: - Östrus (Brunst) - Post- oder Metöstrus (Nachbrunst) - Inter- oder Diöstrus - Präöstrus (Proöstrus, Vorbrunst) (BOUSQUET et al. 1976; GRUNERT 1999; MITKO et al. 2008). Der Präöstrus beginnt mit Verhaltensänderungen und dauert ca. zwei bis drei Tage bis zum Dulden des Aufsprungs. Die Rinder sind oft nervös, versuchen andere Tiere zu belecken und zu bespringen. Weiterhin kommt es oft zur Ödematisierung und Hyperämie der Vulvaschleimhaut und zum Abgang von Schleim (SWANSON u. HAFS 1971; GRUNERT 1999). Der Östrus dauert ca. 18 bis 23 Stunden (CHRISTENSEN et al. 1974; GARVERICK u. SMITH 1993) und kann abhängig von Rasse, Alter, Milchleistung oder Jahreszeit variieren. In dieser Zeit ist das Tier paarungsbereit und duldet die Begattung oder das Bespringen durch andere Tiere. Außerdem sind häufig Unruhe, Brüllen und Abgang von Brunstschleim sichtbar. Mit dem Abklingen der Paarungsbereitschaft beginnt der zwei bis drei Tage dauernde Postöstrus, während dessen die inneren und äußeren Brunstsymptome verschwinden. Ca. 30 bis 35 Stunden nach Beginn des Östrus, bzw. sechs bis 12 Stunden nach Ende der Brunst 15 kommt es zur Ovulation An den Postöstrus schließt sich der Interöstrus (Diöstrus) an, der mit ca. 16 Tagen die längste Zyklusphase ist und in der keine sexuelle Aktivität stattfindet. (SWANSON u. HAFS 1971; AURICH et al. 1996; GRUNERT 1999). Im Laufe des Zyklus treten hormonell bedingte Änderungen an der Genitalschleimhaut auf, welche die Voraussetzung für die Befruchtung, die Einnistung und Ernährung des Embryos schaffen und als Schleimhautzyklus bezeichnet werden. Östrogen bewirkt eine Proliferation, an die sich unter Einwirkung von Progesteron von ca. Tag drei bis Tag 14 des Zyklus die Sekretionsphase anschließt, der eine ungefähr fünf Tage dauernde Regenerationsphase folgt. Hauptsächlich von der Zervix wird Schleim gebildet, der unter Progesteroneinwirkung viskös ist und durch Einfluss von Östrogenen flüssiger wird, und oft während des Östrus als Brunstschleim abgeht. Im Postöstrus fließt bei einigen Tieren Schleim aus der Scheide, der mit Blut aus dem Uterus vermischt ist. Das Blut entstammt Kapillarblutungen im Uterus, die aufgrund von östrogenbedingter Hyperämie entstehen (GRUNERT 1999). 2.1.1 Endokrine Regulation des Zyklus Die Steuerung des Sexualzyklus unterliegt verschiedenen Hormonen, die von Hypothalamus, Hypophyse, Ovarien und Uterus gebildet werden (GARVERICK u. SMITH, 1993). Man unterteilt den hormonellen Zyklus in Follikereifungs-, Ovulations- und Gelbkörperphase (GRUNERT 1999). Das Dekapeptid Gonadotropin-Releasing Hormon (GnRH) wird vom Hypothalamus pulsatil sezerniert und gelangt über Portalgefäße zum Hypophysenvorderlappen (BÉDÉCARRATS u. KAISER 2003). Korrespondierend zur stoßweisen GnRH-Ausschüttung werden in der Hypophyse die Gonadotropine Follikelstimulierendes Hormon (FSH) und Luteinisierendes Hormon (LH) sezerniert (HANSEL u. SNOOK 1970; GARVERICK u. SMITH 1993). FSH bindet hauptsächlich an den Granulosazellen der Ovarien und stimuliert dort das Follikelwachstum und die Ausbildung von LH-Rezeptoren auf den Follikelzellen (THIBAULT et al. 1975; PETERS 1985). Ein Anstieg der FSH-Konzentration bewirkt das Wachstum mehrerer Follikel zur gleichen Zeit. Ein Follikel dieser Welle wird dabei zum 16 dominanten Follikel (SIROIS u. FORTUNE 1988) und hemmt, unter anderem durch Ausschüttung von Inhibin, das Wachstum der anderen Follikel (AURICH et al. 1996). Dabei treten beim Rind zwei oder drei, zum Teil auch vier Follikelwellen auf (LUCY et al. 1992; GARVERICK u. SMITH 1993). BLEACH et al. (2004) beschreiben auch das Vorkommen von sechs Follikelwellen. Im Interöstrus, beim Vorhandensein eines Gelbkörpers und damit einer hohen Progesteronkonzentration, findet keine Ovulation des dominanten Follikels statt, da Östradiol in dieser Zeit über einen negativen Rückkopplungsmechnismus die LH-Freisetzung aus der Hypophyse hemmt (SCHAMS et al. 1977; KESNER et al. 1982; WALTERS et al. 1984). Es kommt zur Atresie des dominanten Follikels. Daraufhin folgen eine oder mehrere neue Wellen, deren dominanter Follikel nach der Regression des Gelbkörpers ovuliert (SIROIS u. FORTUNE 1988). Das Glykoprotein LH bindet an Membranrezeptoren der Thekazellen und stimuliert die Reifung des dominanten zum Graafschen Follikel, dessen Ovulation und später die Entstehung und Erhaltung des Gelbkörpers. Der Graafsche Follikel bildet Östrogene, unter deren Einfluss es nach Brunstbeginn zu einer vermehrten Freisetzung von LH aus der Hypophyse kommt (PETERS 1985; GRUNERT 1999). Diese präovulatorische LH-Welle dauert ca. sechs bis zehn Stunden (WALTERS u. SCHALLENBERGER 1984). Nach dem höchsten LH-Anstieg, der als LH-Peak bezeichnet wird, kommt es ca. 24 Stunden später zur Ovulation (CHENAULT et al. 1974; CHRISTENSEN et al. 1974; SCHAMS et al. 1977). Somit findet die Ovulation in der Regel ca. 29,6 Stunden nach Beginn der Duldung des Aufsprunges (SWANSON u. HAFS 1971) beziehungsweise sechs bis 12 Stunden nach Ende der Brunst statt (AURICH et al. 1996). Der Einfluss proteolytischer Enzyme führt zur Ausdünnung der Follikelwand und später zu deren Einreißen und damit zum Austreten der befruchtungsfähigen Eizelle (THIBAULT et al. 1975; GARVERICK u. SMITH 1993). Aus Granulosa- und Thekazellen der rupturierten Follikelwand entsteht unter Einfluss der Gonadotropine der Gelbkörper, der das Hormon Progesteron sezerniert, das ungefähr vom vierten bis 17. Tag das Zyklusgeschenhen dominiert (HENDERSON u. MOON 1979; PETERS 1985). Progesteron bewirkt außerdem am Endometrium das Ablaufen der Vorgänge, die zur Implantation des Embryos notwendig sind (GEISERT et al. 1992; AURICH et al. 1996). 17 Die Progesteronsekretion wird durch luteotrophe und luteolytische Signale reguliert. Dabei ist beim Rind LH das vorherrschende luteotrophe Hormon (HANSEL u. CONVEY 1983). Weitere regulatorische Stimuli kommen von Uterus, Ovar und Embryo (GARVERICK u. SMITH 1993). Das gebildete Progesteron reduziert über die Hemmung der GnRH-Produktion im Hypothalamus die LH-Sekretion (AURICH et al. 1996). Der Gelbkörper erreicht seine maximale Größe von ca. 2,5 cm ungefähr an Tag sieben bis acht des Zyklus (GRUNERT 1999). In der späten Lutealphase um den 17. Zyklustag steigt die Produktion von Prostaglandin F2α (PGF2α) im Endometrium, das über die Uterin- und Ovargefäße zum Gelbkörper gelangt. PGF2α wirkt durch die Reduktion des Blutflusses zum Gelbkörper luteolytisch. Zusätzlich reguliert PGF2α über Bindung an Rezeptoren auf dem Gelbkörper die Progesteronsekretion (ALILA et al. 1988a, b; GARVERICK u. SMITH 1993). An den Ovarien bewirkt PGF2α die Ausschüttung von Oxytocin, das am Uterus ebenfalls zu einer Zunahme der Prostaglandinsekretion führt (WATHES u. LAMMING 1995; AURICH et al. 1996). 2.2 Hormonelle Aktivität post partum Nach der Abkalbung kommt es zu einer Umstellung der hormonellen Vorgänge von einer Erhaltung der Trächtigkeit zu einer Rückkehr der zyklischen Ovartätigkeit (GARVERICK u. SMITH 1993). 2.2.1 FSH/LH und Östrogene Während der Trächtigkeit und Geburt hemmen hohe Plasmakonzentrationen von Steroidhormonen die Hypothalamus–Hypophysen–Achse, was zu niedrigen LH- und FSHKonzentrationen führt, die auch direkt nach dem Abkalben bestehen bleiben. Wenige Tage post partum (p.p.) steigt die FSH- Konzentration an, stimuliert das Follikelwachstum und bewirkt so ein Ansteigen der Östrogenkonzentration. Die Östrogene sensibilisieren die Hypophyse für Reize des Hypothalamus und stimulieren so als positive Feedbackreaktion die LH-Sekretion (OPSOMER u. DE KRUIF 1999). 18 Frequenz und Amplitude der pulsatilen GnRH-Sekretion und die Ansprechbarkeit der Hypophyse auf GnRH nehmen in den ersten zwei Wochen post partum zu und führen über eine Steigerung der Gonadotropinsekretion zur Stimulation des Follikelwachstums (GOEHRING et al. 1999). Somit steht die ansteigende LH-Konzentration in direktem Zusammenhang mit dem Wiedereinsetzen der Ovartätigkeit nach der Kalbung (LAMMING et al. 1981). Der Anstieg der LH- und FSH- Pulsfrequenz erhöht sich bis auf mindestens einen Puls pro Stunde, was das Einsetzen des ersten Zyklus post partum ermöglicht (OPSOMER u. DE KRUIF 1999). Die häufigsten und höchsten LH-Peaks treten ca. zwei Wochen vor dem ersten Östrus auf (HUMPHREY et al. 1983). Das erste Ansteigen der Plasma FSH - Spiegel findet bei den meisten Kühen bereits innerhalb der ersten fünf bis zehn Tage nach der Abkalbung statt (LAMMING et al. 1981). Laut WEBB et al. (1980) setzt die episodische LH-Sekretion bereits kurz nach der Geburt wieder ein und die Amplitude steigt stufenweise bis ca. zehn Tage post partum (p.p.) an. Eine erste LH-Welle, die dem ersten kompletten ovariellen Zyklus vorausgeht, ist über den oben genannten positiven Feedbackmechanismus mit dem follikulären Östradiol verbunden, während der FSH-Gehalt im Plasma nicht mit dem Einsetzen des ersten Zyklus p.p. in Zusammenhang steht (WEBB et al. 1980). SAVIO et al. (1990a) beschreiben ebenfalls den Zusammenhang von pulsatiler LH-Sekretion und Östradiolkonzentration. Die Anzahl der LH-Pulse nimmt bei den meisten Kühen mit dem Ansteigen der Östrogenkonzentration zu. Dabei ist die Höhe der LH-Konzentration im Plasma in den ersten zwei Wochen post partum niedriger als später. Die Autoren gehen von einer begrenzten Menge an freisetzbarem LH p.p. aus. Da nicht alle Kühe auf höhere Östrogenkonzentrationen mit höherer LH-Freisetzung reagieren, halten die Autoren eine Phase der postpartalen Insensitivität variabler Länge auf den Östrogenstimulus für möglich (SAVIO et al. 1990a). Die während der Geburt hohe Östradiolkonzentration im Blut fällt post partum schnell ab und steigt erst kurz vor der ersten Brunst wieder an (HUMPHREY et al. 1983). 19 2.2.2 Progesteron Viele Autoren verwenden den Progesterongehalt in Blut (Plasma oder Serum) oder Milch als Mittel zur Messung und Beurteilung der ovariellen Aktivität. Die Progesteronkonzentration steht in direktem Zusammenhang mit der Gelbkörperfunktion und ist somit ein Indikator für die Ovartätigkeit (PETERS 1984). Aufgrund der Ungenauigkeit der rektalen Untersuchung alleine sehen MCLEOD und WILLIAMS (1991) die Untersuchung des Progesterongehaltes als geeignete Methode zur Beurteilung ovarieller Funktionen bzw. Fehlfunktionen. Dabei bezeichnen sie einen Progesterongehalt der Milch unter 4,0 ng/ml über einen Zeitraum von über 30 Tagen p.p. als Indikator für ein verlängertes anöstrisches Intervall. Auch OPSOMER et al. (2000) bewerten die Ovarfunktionen p.p. anhand des Progesterongehalts in der Milch. HUMPHREY et al. (1983) berichten über ein Absinken der während der Trächtigkeit hohen Serumprogesteronkonzentration ca. zwei Tage ante partum auf ca. 0,1 ng/ml. Auf dieser Höhe bleibt die Konzentration bis kurz vor der ersten Brunst p.p. Es kommt bei vielen Kühen zwischen 4,6 und 0,6 Tagen vor dem ersten Östrus nach der Abkalbung zu einem Progesteronanstieg. Der Autor vermutet, dass das Progesteron aus nicht palpierbarem Gelbkörpergewebe, möglicherweise aus luteinisierten kleineren Follikeln im Ovar, stammt. (HUMPHREY et. al. 1983). Diesem kurzfristigen Anstieg der Plasmaprogesteronkonzentration geht vier bis fünf Tage vorher eine LH-Welle voraus (WEBB et al. 1980). Auch CHOI et al. (1977) stellten fest, dass vor der ersten Brunst p.p. ein Anstieg der Plasmaprogesteronkonzentration auf 1-3 ng/ml erfolgt, der auf eine Ovartätigkeit hinweist. Während der ersten Brunst nach der Kalbung sinkt der Progesteronwert unter 0,25 ng/ml und steigt danach auf einen Höchstwert von ca. 5,4 ng/ml (CHOI et al. 1977). Die Plasmaprogesteronkonzentration bei 21 der von SAVIO et al. (1990a) untersuchten 22 Kühe waren während der Phase des postpartalen Anöstrus niedrig. Lediglich bei einer untersuchten Kuh trat ein Anstieg der Progesteronkonzentration vor der ersten Ovulation auf. 20 STUMPF et al. (1992) definieren den Beginn der zyklischen Ovartätigkeit über die Progesteronkonzentration im Serum. Dabei wird ein Progesterongehalt ≥ 1ng/ml in zwei aufeinanderfolgenden Blutproben gefolgt von zyklischem Anstieg und Abfall als Wiedereinsetzen der normalen zyklischen Tätigkeit gewertet. DARWASH et al. (1997) beschreiben den Beginn der Lutealaktivität über den Milchprogesterongehalt, wenn dieser in zwei aufeinanderfolgenden Proben über 3 ng/ml liegt. Dabei steigt der Progesteronwert vier bis fünf Tage nach der Brunst an. Die durchschnittliche Plasmaprogesteronkonzentration während des ersten Zyklus p.p. ist niedriger als während des zweiten Zyklus nach der Kalbung. Die Autoren folgern daraus, dass im ersten Zyklus noch nicht die maximale Gelbkörperfunktion erreicht wird (STAPLES u. THATCHER 1990). 2.2.3 Ovartätigkeit post partum Nach der Abkalbung folgt beim Rind eine Phase der Inaktivität der Ovarien und der sexuellen Ruhe, bevor der Zyklus wieder einsetzt (PETERS 1984). Post partum kommt es zur Involution des Uterus und zur Wiederaufnahme des ovariellen Zyklus. Dies sind Voraussetzungen, damit eine erneute Trächtigkeit möglich ist. Die Phase von der Abkalbung bis zum ersten Östrus mit Ovulation wird als postpartales Intervall (PPI) bezeichnet. Sie ist unterschiedlich lang und wird von verschiedenen Faktoren beeinflusst (PETERS 1984; RHODES et al. 2003). In der anovulatorischen postpartalen Phase variabler Länge kommt es zur Regression des Trächtigkeitsgelbkörpers, bevor die zyklischen Ovarfunktionen wieder einsetzen und die erste Ovulation nach der Abkalbung stattfindet (SAVIO et al. 1990a). Die Dauer des postpartalen Intervalls wird in der Literatur mit unterschiedlichen Längen angegeben. Dabei zeigt sich vor allem ein Unterschied in der Nutzungsrichtung der Tiere. OPSOMER et al. (2000) untersuchten hochleistende Kühe in belgischen Milchviehbetrieben. Bei diesen Tieren war der erste signifikante Anstieg der Progesteronkonzentration in der Milch (Progesteron ≥ 15 ng/ml) nach ca. 37 Tagen nachweisbar. Somit ist von einer Ovulation ungefähr 32 Tage nach der Abkalbung auszugehen. (OPSOMER et al. 2000). 21 WEBB et al. (1980) konnten bei Untersuchungen an Milchkühen in der zweiten Laktation bereits ca. 17 Tage nach der Kalbung eine zyklische Ovartätigkeit über den Plasmaprogesteronanstieg nachweisen. PETERS (1984) beschreibt bei Milchkühen das Wiedereinsetzen der Ovartätigkeit p.p. über den Milchprogesterongehalt. Die Dauer des postpartalen Intervalls war ca. 24 Tage. Bei Mutterkühen mit säugendem Kalb hingegen betrug die Dauer ca. 57 Tage. HUMPHREY et al. (1983) untersuchten den Blutprogesterongehalt von säugenden Mutterkühen unterschiedlicher Rassekreuzungen, die das zweite Mal gekalbt hatten. Die Dauer des postpartalen Intervalls betrug durchschnittlich 48 Tage. 2.2.4 Follikelwachstum und erste Ovulation/erste Brunst post partum SAVIO et al. (1990a) untersuchten das postpartale Follikelwachstum bei Milchkühen in der zweiten bis vierten Laktation. Dabei kam es bereits wenige Tage nach der Abkalbung zur Ausbildung kleiner Follikel, bevor sich ca. 12 Tage post partum ein erster dominanter Follikel bildete. Dieser ovulierte bei 14 von 19 untersuchten Kühen, und wurde bei vier von 19 Tieren zystisch. Bei den von WEBB et al. (1980) untersuchten Milchkühen in der zweiten Laktation waren im Durchschnitt 15,7 Tage nach der Kalbung Follikel durch rektale Palpation feststellbar. MURPHY et al. (1990) fanden bereits zwei bis acht Tage post partum bei säugenden Fleischkühen (zwei bis vier Abkalbungen) Follikel mittlerer Größe von ca. 5 bis 9 mm Durchmesser. Der erste dominante Follikel wurde ca. 10 Tage (sieben bis 18 Tage) nach der Kalbung festgestellt, ovulierte aber nur bei 12 % der untersuchten Kühe. SAVIO et al. (1990b) untersuchten bei Milchkühen den Zusammenhang von Zykluslänge und der Anzahl an Follikelwellen mit dominantem Follikel, die während eines ovariellen Zyklus auftreten. In kurzen Zyklen (neun bis 13 Tage) war meistens ein dominanter Follikel, in normal langen Zyklen (18 bis 24 Tage) waren meistens zwei, und in langen Zyklen (≥ 25 Tage) in der Regel drei dominante Follikel zu beobachten. Die erste Ovulation post partum fand bei 17 von 18 in dieser Studie untersuchten Kühen ohne sichtbare 22 Brunstsymptome statt. Dabei dauerte der erste Zyklus ca. 21 Tage. Die Zykluslänge war abhängig vom Zeitpunkt des Auftretens des ovulatorischen dominanten Follikels (SAVIO et al. 1990b). Zum Auftreten der ersten Ovulation post partum gibt es unterschiedliche Ergebnisse. DARWASH et al. (1997) berichten bei Milchkühen in Großbritannien (eine bis zwölf Laktationen) von einer Spanne von durchschnittlich 21,8 Tagen bis zur ersten Ovulation post partum. Die erste Ovulation nach der Kalbung trat bei Untersuchungen von SAVIO et al. (1990a) an Milchkühen in der zweiten bis vierten Laktation ungefähr an Tag 27 auf. OPSOMER et al. (2000) bestimmten den Zeitpunkt der ersten Ovulation über den Progesteronanstieg in der Milch. Die erste Ovulation fand bei den untersuchten hochleistenden Milchkühen ca. 32 Tage post partum statt. STEVENSON und BRITT (1980) geben bei Milchkühen eine Spanne von neun bis 34 Tagen nach der Kalbung an, in der die untersuchten Kühe zum ersten Mal ovulierten. Dabei wird die Dauer bis dahin vor allem vom Östrogenspiegel und der Follikelanzahl in den ersten beiden Wochen p.p. beeinflusst. Eine frühe Ovulation steht im Zusammenhang mit einem LHAnstieg in der ersten bis zweiten Woche post partum. In Untersuchungen von CHOI et al. (1977) an laktierenden Fleckviehkühen im Alter von zwei bis zweieinhalb Jahren trat die erste Brunst ca. 27 Tage nach dem Abkalben auf. Laut GARCIA und LARSSON (1982) fanden bei Milchkühen verschiedener schwedischer Rassen die erste, zweite und dritte Ovulation durchschnittlich an den Tagen 21, 39 und 58 nach der Abkalbung statt. Dabei dauerte der erste Brunstzyklus ca.17,5 Tage. KING et al. (1976) berichten vom Auftreten der ersten Ovulation post partum nach 19,5 (10-68) Tagen bei Milchkühen (erste bis fünfte Laktation). Dabei wurden bei kontinuierlich überwachten Tieren die ersten Brunstsymptome nach ca. 35 Tagen, bei nur gelegentlich beobachteten Kühen nach ca. 57 Tagen gesehen. Dies deckt sich mit Untersuchungen von MACMILLAN et al. (1996), nach denen hochleistende Milchkühe in der Regel innerhalb von 4 Wochen nach der Kalbung ovulieren, aber dabei keine Brunstsymptome beobachtet werden. Dies führt der Autor auf eine negative Energiebilanz post partum zurück. 23 STAPLES und THATCHER (1990) untersuchten die Ovarfunktionen mehrkalbiger Milchkühe in der Frühlaktation. Die Autoren stellten fest, dass Kühe mit einer besseren Energiebilanz schneller ovarielle Aktivität zeigen, zur Ovulation aber keine positive Energiebilanz notwendig ist. Bei säugenden Mutterkühen (zwei bis vier Abkalbungen) fand die erste Ovulation ungefähr an Tag 36 p.p. statt. 89 % der Tiere zeigten dabei keine Brunstsymptome. Die Dauer des ersten ovariellen Zyklus betrug nur ca. 12 Tage. Die erste sichtbare Brunst zeigten diese Kühe erst ca. 54 Tage nach der Kalbung (MURPHY et al. 1990). Die Dauer des postpartalen Intervalls, nach dem die ovarielle Aktivität nach der Kalbung wieder einsetzt, ist variabel (PETERS 1984). Um eine Zwischenkalbezeit von 365 Tagen zu erreichen, sollte aber die erste erfolgreiche Besamung nicht später als 80 bis 85 Tage nach der Abkalbung stattfinden (WEBB et al. 1980; PETERS 1984; MC LEOD u. WILLIAMS 1991; OPSOMER et al. 2000). Damit dieser Zeitrahmen eingehalten werden kann, ist es mittlerweile auf vielen Betrieben Routine, Kühe, die noch keine Brunst gezeigt haben, aber bald besamt werden können, zu untersuchen und gegebenenfalls zu behandeln (MC LEOD u. WILLIAMS 1991). Mittlerweile wird in der Regel statt einer Zwischenkalbezeit von 365 Tagen ein Wert von bis zu 385 Tagen akzeptiert, wobei die mittlere Rastzeit auch dann unter 85 Tagen liegen sollte (DE KRUIF et al. 2007). Eine unterschiedlich lange Phase des postpartalen Anöstrus wird als physiologisch angesehen. Gesunde Tiere sollten allerdings spätestens 40 bis 60 Tage nach der Kalbung wieder Brunstsymptome zeigen, um eine Zwischenkalbezeit von 365 Tagen zu erreichen (OPSOMER u. DE KRUIF 1999). MC LEOD und WILLIAMS (1991) bezeichneten die Phase der postpartalen Anöstrie („anoestrous“) als verlängert, wenn der Milchprogesterongehalt länger als 30 Tage post partum unter 4,0 ng/ml lag. 19 % der von ihnen untersuchten Tiere fielen in diese Kategorie. 42 Tage nach der Kalbung zeigten noch 7,6 % der Kühe keine zyklische Ovartätigkeit. Dabei wurden mehr Kühe als nichtzyklisch vorgestellt, als aufgrund von rektalen Untersuchungen der Ovarien und durch Bestimmung der Milchprogesteronwerte als azyklisch diagnostiziert wurden. 24 21,5 % der von OPSOMER et al. (2000) untersuchten Milchkühe zeigten in den ersten 50 Tagen post partum keinen Anstieg der Milchprogesteronkonzentration, sondern in diesem Zeitraum konstant niedrige Werte und wurden deshalb als Tiere mit verzögerter zyklischer Ovartätigkeit eingestuft. DE KRUIF (1977) beschreibt, dass bei 16 % der Kühe 50 bis 60 Tage nach der Geburt keine Brunst auftrat. Dabei bestanden große Unterschiede zwischen den Tieren einzelner Herden (8 bis 24 %). In amerikanischen Milchviehherden zeigten 42 % der Kühe bis 70 Tage nach der Abkalbung keine äußerlich erkennbare Brunst (BARTLETT et al. 1987). STAPLES und THATCHER (1990) bestimmten die Ovarfunktion über den Plasmaprogesterongehalt. Dabei hatten 28 % der Kühe bis 60 Tage post partum keine zyklische Ovartätigkeit. BALL und LAMMING (1983) geben einen Wert von 12,3 % der untersuchten Kühe an, die 35 Tage post partum noch keine ovarielle Aktivität zeigten. Über den Milchprogesterongehalt fanden BULMAN und LAMMING (1978) heraus, dass 5,2 % der Kühe 50 Tage nach dem Abkalben noch keine zyklische Eierstocksfunktion hatten, während weitere 5,2 % nach einer Phase der zyklischen Aktivität wieder azyklisch wurden. In den von MAYER et al. (1987) untersuchten israelischen Milchviehherden betrug der Anteil von Kühen mit unbeobachteter Brunst 18 %. Rund ein Viertel (26 %) dieser Tiere hatte bei der rektalen Untersuchung der Eierstöcke keine tastbaren Funktionskörper. Dies wurde von den Autoren als inaktive Ovarien bezeichnet, die Kühe wurden als azyklisch („echt anöstrisch“) eingestuft. Der weitaus größere Teil der Tiere hatte bei der Untersuchung Gelbkörper (58,6 %) oder Follikel (15,4 %). Somit hatte der Großteil der Kühe mit unbeobachteter Brunst trotzdem einen Zyklus und aktive Ovarien (MAYER et al. 1987). 2.3 Postpartaler Anöstrus - unterschiedliche Definitionen Beim Vergleich verschiedener Literaturstellen zum Thema Brunstlosigkeit fällt auf, dass die Autoren unterschiedliche Begriffe und Definitionen verwenden. Somit ist ein Vergleich einzelner Untersuchungen und Ergebnisse oft schwierig. 25 Ursache für differierende Prozentangaben von anöstrischen Tieren sind häufig verschiedene Untersuchungsmethoden, Definitionen und Berechnungen der Ergebnisse. Außerdem ergibt sich eine große Differenz durch die unterschiedliche Qualität der Brunsterkennung in den einzelnen Betrieben (OPSOMER u. DE KRUIF 1999). Im Folgenden werden in der Literatur verwendete Bezeichnungen und die Definitionen der Autoren beschrieben. 2.3.1 Anöstrus OPSOMER und DE KRUIF (1999) definieren Anöstrus (engl. „anestrus“) als das Fehlen des typischen Brunstverhaltens in einem Zeitabschnitt, in dem ein Östrusgeschehen eigentlich erwartet werden kann. Als Anöstrus vor der Besamung (engl. „preservice anestrus“) bezeichnen BARTLETT et al. (1987) Kühe, bei denen bis 70 Tage nach der Kalbung noch keine Brunst gesehen wurde. 2.3.2 Suböstrus/unbeobachtete Brunst/stille Brunst Der Suböstrus ist gekennzeichnet durch normale zyklische Ovartätigkeit, ohne dass die Tiere äußere Brunsterscheinungen zeigen (stille Brunst). Dies ist zu unterscheiden von der nicht wahrgenommenen Brunst, deren Rate sich durch Optimierung der Brunstbeobachtung verringern lässt (OPSOMER u. DE KRUIF 1999). MAYER et al. (1987) stellten die Diagnose „unbeobachtete Brunst“ (UBB) bei Kühen, bei denen 60 Tage post partum keine Brunsterscheinungen sichtbar waren, und die nach normaler Abkalbung keine krankhaften Erscheinungen wie Endometritis oder Nachgeburtsverhaltung hatten. Die ersten Ovulationen nach dem Wiedereinsetzen der Ovartätigkeit nach der Abkalbung verlaufen häufig unerkannt. SCHOPPER et al. (1993) ermittelten über den Progesteronanstieg in der Milch, dass 41,2 % der Ovulationen bei den untersuchten Kühen ohne erkennbare Brunstsymptome verliefen. 94,8 % der Tiere der Studie waren mindestens einmal stillbrünstig. 26 2.3.3 Azyklie/eigentlicher Anöstrus/Ovardystrophie/anovulatorische Kühe Azyklische Kühe haben sehr niedrige Progesteronwerte und kleine Ovarien mit niedriger follikulärer Aktivität. Dabei reicht eine einmalige Untersuchung zur sicheren Diagnose der Azyklie nicht aus (OPSOMER u. DE KRUIF 1999). BALL und LAMMING (1983) bezeichnen Kühe als azyklisch (engl. „acyclic“), wenn bis zum 35. Tag post partum kein Anstieg des Milchprogesterongehalts (zwei oder mehr aufeinanderfolgende Proben mit einem Gehalt von über 2 ng/ml Milch) erkennbar ist. Kühe mit inaktiven Ovarien werden als echte Anöstrus-Fälle (Azyklie) definiert (MAYER et al. 1987). MC DOUGALL et al. (2004) bezeichnen Kühe als anovulatorisch anöstrisch (engl. „anovulatory anoestrus“), bei denen per Ultraschall kein Corpus luteum erkennbar ist. GÜMEN et al. (2003) bezeichnen Kühe, die bei zweimaliger Untersuchung keinen palpierbaren Gelbkörper und einen niedrigen Plasmaprogesterongehalt haben, als anovulatorisch (engl. „anovular“). In der von GÜMEN et al. (2003) durchgeführten Studie waren 20 % der untersuchten Milchkühe ca. 60 Tage nach der Abkalbung anovulatorisch und hatten keine zyklische Ovartätigkeit. GRUNERT (1999) definiert eine Afunktion ehemals leistungsfähiger Eierstöcke als Folge belastender exogener und endogener Faktoren als Ovardystrophie. 2.4 Einflüsse auf das Wiedereinsetzen der Ovarfunktionen post partum bzw. das Entstehen von Azyklie und Anöstrie 2.4.1 Einfluss von Fütterung und Körperkondition Die von STAPLES und THATCHER (1990) untersuchten Milchkühe, die bis zum Tag 60 post partum eine normale zyklische Ovartätigkeit zeigten, hatten nach der Geburt ihre Futteraufnahme schneller gesteigert als Kühe, die zu diesem Zeitpunkt noch keine normalen Eierstocksfunktionen hatten. Diese Kühe nahmen in Relation zu ihrer Körpermasse weniger Futter auf. Die Autoren sehen deshalb einen Zusammenhang zwischen der Trockenmasseaufnahme nach der Kalbung und der Rückkehr zum normalen Zyklus, sowie 27 eine Koinzidenz von Körpermasseverlust und der damit einhergehenden negativen Energiebilanz während der ersten zwei Wochen post partum und der mangelnden Ovartätigkeit. Die Körperkondition von Milchkühen zum Zeitpunkt der Abkalbung wird von OPSOMER et al. (2000) nicht als Risikofaktor für das verspätete Einsetzen zyklischer Ovarfunktionen gesehen. Entscheidend ist vielmehr der Gewichtsverlust im Laufe der ersten zwei Wochen post partum. SHRESTHA et al. (2005) und SAMARÜTEL et al. (2008) beobachteten ein verzögertes Einsetzen der zyklischen Ovartätigkeit von Milchkühen, die in den ersten Wochen p.p. eine geringere Körperkondition (Body Condition Score, BCS) hatten oder nach der Abkalbung viel Gewicht verloren. Die prozentuale Veränderung des Körpergewichts von Milchkühen nach der Kalbung ist negativ korreliert zum Auftreten der ersten Ovulation (STEVENSON u. BRITT 1980). ZUREK et al. (1995) sehen nicht das Ausmaß des Gewichtsverlustes, sondern die dynamischen Veränderungen in der Energiebilanz als entscheidenden Faktor für den Zeitpunkt der ersten Ovulation post partum. Dies deckt sich mit den Untersuchungen von WHITAKER et al. (1993) und PATTON et al. (2007), die eine positive Korrelation von Energiebilanz und dem Beginn der Lutealaktivität nach der Abkalbung beschrieben. Bei schwerer Unterernährung oder als Folge einer negativen Energiebilanz kommt es durch eine Hemmung der GnRH-Sekretion zur verminderten Freisetzung von FSH und LH und damit zum verzögerten Wiedereinsetzen der Eierstocksfunktionen (JOLLY et al. 1995; GOEHRING et al. 1999). STUMPF et al. (1992) untersuchten die Auswirkungen der Körperkondition von Fleischkühen auf die Länge des postpartalen Intervalls. Kühe in besserer Kondition hatten eine kürzere Dauer des postpartalen Anöstrus im Vergleich zu Kühen mit niedrigerem Body Condition Score. Bei Fleischkühen in guter Kondition verlängert ein moderater Gewichtsverlust nach der Kalbung das postpartale Intervall nicht (HUMPHREY et al. 1983). RICHARDS et al. (1989) induzierten einen Anöstrus bei Herefordkühen durch restriktive Fütterung. Dabei kam es zu reduzierten LH-Konzentrationen und selteneren LH-Puls-Frequenzen. Bei Steigerung der Futtermenge setzten die Brunstzyklen wieder ein (RICHARDS et al. 1989). 28 Mit steigenden Futtermengen bei säugenden Fleischkühen sank das Intervall von Kalbung bis zum ersten Östrus post partum. Die unterschiedliche Futteraufnahme hatte allerdings keinen Effekt auf die LH-Freisetzung aus der Hypophyse. Wichtigeren Einfluss hatte die Erhaltung der Körperkondition p.p. unabhängig von der Nährstoffaufnahme. Fleischkühe, die ihr Gewicht nach der Abkalbung hielten, hatten eine kürzere Dauer des postpartalen Anöstrus als Kühe, die nach der Abkalbung Gewicht verloren (RUTTER u. RANDEL 1984). 2.4.2 Einfluss von Laktation/Milchleistung/säugendem Kalb Die Milchleistung in den ersten 100 Tagen p.p. war bei den von OPSOMER et al. (2000) untersuchten Kühen kein Risikofaktor für verzögerte postpartale Ovartätigkeit. Allerdings fanden die Autoren einen Zusammenhang zwischen dem Proteingehalt der Milch und verspäteten Eierstocksfunktionen und damit zwischen der Energiebilanz nach der Kalbung und der zyklischen Ovartätigkeit. In der Studie von STAPLES und THATCHER (1990) hatten Kühe, die bis 60 Tage p. p. keinen Zyklus hatten, eine niedrigere Milchleistung als Kühe, deren Ovarfunktionen schon früher wieder einsetzten. Deshalb folgerten die Autoren, dass der Energiestatus nach der Abkalbung wichtiger sei als die Milchleistung in Bezug auf das Einsetzen der Ovarfunktionen. Anöstrische Kühe hatten eine atypische Laktationskurve mit schnellem Anstieg, Peak in der zweiten Woche und schnellem Abfall. Diese Tiere bauten außerdem mehr Körpermasse ab, um die Milch zu produzieren. SMOLDERS et al. (1996) berichten hingegen von einer höheren Milchleistung (2,65 kg „fettkorrigierte“ Milch (fat corrected milk, FCM) mehr pro Tag als der Durchschnitt) anöstrischer Kühe im Vergleich zu Tieren, die bis zum 50. Tag post partum zyklische Ovartätigkeit wiedererlangt hatten. Bei den Untersuchungen von STEVENSON und BRITT (1980) zeigte sich nur ein geringer Einfluss von Energieaufnahme und Milchleistung während der ersten zwei Wochen post partum auf die Ovaraktivität. Die Autoren schließen aber die Gesamtlaktationsleistung als Einflussfaktor auf die Eierstocksfunktionen nach der Abkalbung nicht aus. Kühe, die gemolken werden, zeigen im allgemeinen schneller wieder zyklische Ovaraktivität als Tiere, die ein Kalb säugen. Dabei verzögert sich die Ovartätigkeit umso länger, je stärker die Saugaktivität ist (LAMMING et al. 1981). 29 Einige Autoren beschreiben einen inhibitorischen Effekt des Säugens auf die Aktivität des Hypothalamus, indem die pulsatile GnRH-Freisetzung und damit die LH-Freisetzung gehemmt wird. Dies verzögert das Einsetzen des Zyklus nach der Kalbung (LAMMING et al. 1981; WALTERS et al. 1982; HUMPHREY et al. 1983). So haben Milchkühe mit ca. 24 Tagen eine kürzere azyklische Phase post partum als säugende Kühe (ca. 57 Tage) (PETERS 1984). Die LH-Konzentration im Blut steigt bei Mutterkühen mit säugendem Kalb kurze Zeit nach dem Absetzen an. Auch die Frequenz der LH-Peaks im Blut war nach dem Absetzen der Kälber höher als bei säugenden Kühen, während die Kortisonspiegel in beiden Gruppen nahezu gleich blieben. Deshalb vermuten WHISNANT et al. (1985) keinen inhibitorischen Effekt von Serumkortison auf die LH-Sekretion bei säugenden Mutterkühen. 2.4.3 Einfluss von Krankheiten Das Auftreten von Krankheiten (Stoffwechselstörungen, klinische Mastitis, Lahmheit) in den ersten vier Wochen post partum ist ein Risikofaktor für das verzögerte Einsetzen der zyklischen Ovartätigkeit. Dabei gehen OPSOMER et al. (2000) davon aus, dass es während der Krankheit zu verminderter Futteraufnahme und damit zu einer negativen Energiebilanz kommt. Demgegenüber fanden STAPLES und THATCHER (1990) bei den von ihnen untersuchten hochleistenden Milchkühen keinen Zusammenhang zwischen dem Auftreten von Krankheiten und verzögerter Ovarfunktion. Kühe mit gestörter Geburts- und Puerperalphase zeigen den ersten deutlichen Progesteronanstieg ca. fünf Tage später als Kühe ohne Komplikationen. Bei Tieren ohne Endometritis puerperalis begann die Ovarfunktion post partum ungefähr neun Tage früher als bei Kühen mit Endometritis (BOSTEDT et al. 1985). DE KRUIF (1977) fand keinen Zusammenhang zwischen dem Verlauf von Geburt und Puerperium und dem Auftreten von Suböstrus und Azyklie. 30 2.4.4 Einfluss der Jahreszeit Das Intervall bis zur ersten Lutealaktivität post partum ist bei im Herbst gekalbten Milchkühen mit ungefähr 23 Tagen kürzer als zu jeder anderen Jahreszeit und am längsten im Frühjahr (ca. 28 Tage) (DARWASH et al. 1997). Dies wird von SAVIO et al. (1990) bestätigt, die bei im Herbst gekalbten Milchkühen signifikant früher einen dominanten Follikel fanden als bei Tieren, die im Frühjahr kalbten. (ca. sieben Tage p.p. im Herbst gegenüber ca. 20 Tagen p.p. im Frühling). Allerdings gab es in beiden Gruppen keinen signifikanten Unterschied im Zeitpunkt der ersten Ovulation post partum (SAVIO et al. 1990a). Auch CHOI et al. (1977) vermutet einen Einfluss der Jahreszeit auf die Länge des postpartalen Intervalls (PPI), da bei Untersuchungen an gemolkenen Fleckviehkühen im Sommer die erste Brunst post partum früher auftrat als bei Untersuchungen auf dem gleichen Betrieb im Winter. Milchkühe, die im Winter abkalben, zeigen häufiger ein verzögertes Einsetzen der zyklischen Ovartätigkeit als Kühe, die während der Weidesaison abkalben (OPSOMER et al. 2000). 2.4.5 Einfluss der Anzahl der Abkalbungen Zu diesem Thema gibt es in der Literatur sehr unterschiedliche Ergebnisse. Bei den von STAPLES und THATCHER (1990) untersuchten multiparen hochleistenden Milchkühen zeigte sich kein Zusammenhang zwischen dem Alter der Tiere und der Zeit bis zum Wiedereinsetzen der Ovartätigkeit post partum. Demgegenüber beschreiben DARWASH et al. (1997), dass das Intervall bis zum Beginn der Lutealaktivität bei den von ihnen untersuchten Milchkühen (erste bis zwölfte Laktation) mit der Anzahl der Abkalbungen länger wird. Im Gegensatz dazu zeigte sich in einer weiteren Untersuchung an Milchkühen bei den Färsen ein erster Anstieg der Progesteronkonzentration ca. 45 Tage nach der Kalbung, bei zweitkalbigen Tieren schon ca. 28 Tage post partum und ca. 39 Tage nach der Abkalbung bei Mehrkalbskühen (BOSTEDT et al. 1985). TANAKA et al. (2008) fanden bei ihren Studien eine negative Korrelation zwischen der Anzahl der Abkalbungen von Milchkühen und der Länge der Dauer bis zur ersten Ovulation post partum. 31 2.4.6 Einfluss der Trockenstehzeit Milchkühe, die nicht trockengestellt wurden und durchgehend mit hohen Mengen an Energie versorgt wurden, zeigten schneller Follikelwachstum und eine kürzere Zeit bis zur ersten Ovulation post partum (ca. 13 Tage) im Vergleich zu Kühen, die 28 oder 56 Tage trockengestellt wurden und während dieser Zeit weniger energiereiches Futter aufnahmen (GÜMEN et al. 2005). OPSOMER et al. (2000) fanden bei Untersuchungen an hochleistenden Milchkühen heraus, dass eine Trockenstehzeit von über 77 Tagen das Intervall bis zum Einsetzen der zyklischen Ovartätigkeit nach der Abkalbung verlängert. 2.4.7 Einfluss sonstiger Faktoren Weitere Faktoren, die die Länge des postpartalen Intervalls beeinflussen, werden von verschiedenen Autoren beschrieben. STUMPF et al. (1992) schlussfolgern, dass Fleischkühe in durchschnittlicher Körperkondition zum Zeitpunkt der Abkalbung auf die Anwesenheit eines Bullen in der Herde mit einem verkürzten postpartalen Anöstrus reagieren. Kühe mit niedrigerem Körpergewicht werden stärker von der Präsenz eines Bullen beeinflusst als Tiere mit einer besseren Körperkondition. Die Autoren vermuten eine Stimulation der GnRH-Sekretion und damit die Beeinflussung der Ovartätigkeit durch den Bullen. Ursache für die Differenzen in der Länge der postpartalen Intervalle bei einzelnen Tieren können laut DARWASH et al. (1997) vererbbare Unterschiede in der Sensitivität von Hypophyse und Ovar auf GnRH-Signale sein. Die Autoren vermuten, dass eine genetische Selektion auf kürzere Intervalle bis zur Lutealaktivität nach der Abkalbung möglich sei. OPSOMER und DE KRUIF (1999) beschreiben den Zusammenhang zwischen der Uterusinvolution nach der Abkalbung und dem Einsetzen der ovariellen Aktivität. Dabei verhindert vom Endometrium produziertes und sezerniertes PGF2α das Wiedereinsetzen der Ovartätigkeit. 32 2.5 GnRH als körpereigenes Hormon und Freisetzung von FSH/LH 2.5.1 GnRH Das Dekapeptid GnRH spielt als zentraler Initiator der reproduktiven Hormonkaskade eine wichtige Rolle bei der Regulation weiblicher und männlicher Fortpflanzungsfunktionen (CONN et al. 1984; MILLAR 2005). Es wird von neurosekretorischen Zellen des Hypothalamus gebildet und gelangt über die Portalgefäße zur Hypophyse, wo daraufhin FSH und LH freigesetzt werden, die an den Gonaden die Sekretion der Steroidhormone und von Inhibin stimulieren (CONN et al. 1984; GOEHRING et al. 1999; SOSNOWSKI et al. 2000). Diese Hormone wirken wiederum im Sinne eines geschlossenen Feedback-Systems auf Hypophyse und Hypothalamus. GnRH wird vom Hypothalamus pulsatil sezerniert und bewirkt damit auch eine episodische Ausschüttung der Gonadotropine (CONN et al. 1984). Jedem GnRH-Puls folgt ein LH-Puls. Die FSH-Pulse sind nicht so klar erkennbar (MILLAR 2005). 2.5.1.1 Verschiedene Formen von GnRH Bei den Wirbeltieren existieren 23 Formen von GnRH, die sich in ihren Aminosäuresequenzen unterscheiden (PAWSON u. MC NEILLY 2005). Bei den meisten Vertebratenspezies treten zwei dieser Formen auf. Analog dazu wird vermutet, dass zu den einzelnen Formen unterschiedliche Rezeptorsubtypen gehören. Die Formen unterscheiden sich neben den Aminosäuremustern auch in ihrer Lokalisation, Verteilung und dem embryologischen Ursprung. GnRH I wird als hypothalamische Form bezeichnet und ist speziesspezifisch. Die Funktion von GnRH II, das von Zellen des Mittelhirns gebildet und „Chicken-GnRH II“ genannt wird, ist bislang unklar. Als dritte Form ist bei einigen Vertebraten das „Salmon-GnRH III“ identifiziert worden (DUBOIS et al. 2002; MILLAR 2005; PAWSON u. MC NEILLY 2005; RAMAKRISHNAPPA et al. 2005). 33 2.5.1.2 GnRH-Rezeptor GnRH stimuliert die Gonadotropinsekretion aus der Hypophyse in drei Schritten. Zuerst bindet GnRH an den Rezeptor auf der Plasmamembran, daraufhin wird Calcium als „Second Messenger“ mobilisiert. Im dritten Schritt werden FSH und LH freigesetzt (CONN et al. 1984; PAWSON u. MCNEILLY 2005). Die GnRH-Rezeptoren gehören zum Typ der G-Protein-gekoppelten Rezeptoren (COUNIS et al. 2005; RAMAKRISHNAPPA et al. 2005). Im Laufe des Zyklus variiert die Sensitivität der Hypophyse für GnRH nur über eine Änderung der Anzahl der GnRH-Rezeptoren, nicht über deren veränderte Bindungsaffinität. Die Anzahl der GnRH-Rezeptoren in der Hypophyse steigt während des Östrus deutlich an und bleibt im übrigen Zyklusverlauf gleich (CONN et al. 1984; LEUNG et al. 1984). Längerfristige kontinuierliche Verabreichung von GnRH oder Analoga führt zu einer ständigen Rezeptorbesetzung und damit zur Desensibilisierung, da die Rezeptoren schneller internalisiert werden als sie ersetzt werden können. Dies führt zu einer verminderten Gonadotopinsekretion (CONN et al. 1984; RISPOLI u. NETT 2005). 2.5.1.3 Beeinflussung der GnRH-Sekretion auf Ebene des Hypothalamus GnRH wird von hypothalamischen Neuronen pulsatil in die hypophysären Portalgefäße sezerniert. Dabei bilden die Neuronen ein diffuses Netzwerk, das als hypothalamischer Pulsgenerator bezeichnet wird und die episodische Freisetzung von GnRH reguliert. Die GnRH-Sekretion wird von verschiedenen Faktoren beeinflusst (MORI u. TANAKA 1995; KRSMANOVIC et al. 2009). Sie ist unter anderem calciumabhängig und wird durch cyclisches Adenosinmonophosphat (cAMP) stimuliert (KRSMANOVIC et al. 2010). Neben transsynaptischen Signalen wird die GnRH-Sekretion auch durch von Endothelzellen und Gliazellen freigesetzten Faktoren beeinflusst (BEAUVILLAIN u. PRÉVOT 2004). 2.5.2 Synthese und Sekretion von FSH und LH Die Glykoproteine FSH und LH bestehen jeweils aus einer α- und einer β-Untereinheit (PIERCE et al. 1976). Während die α-Untereinheit bei beiden Hormonen gleich ist, 34 unterscheiden sie sich in ihren β-Untereinheiten, die auch für die spezifische Rezeptorbindung verantwortlich sind (RYAN et al. 1988; COSOWSKY et al. 1995). Die Bindung von GnRH an die Rezeptoren bewirkt die gesteigerte Transkription der Gene der α-Untereinheit und der LH-β-Untereinheit (SOSNOWSKI et al. 2000). Neben einer basalen Sekretion von FSH und LH wird LH auch pulsatil ausgeschüttet und zeigt keinen Zusammenhang zur FSH-Sekretion. Dabei beträgt die Plasmahalbwertszeit von LH 20 Minuten und von FSH 150 Minuten. Die basale Sekretion findet unabhängig von GnRH statt, während die LH-Pulse abhängig von der pulsatilen GnRH-Freisetzung sind. Die FSH-Sekretion ist weitestgehend unabhängig von den GnRH-Pulsen (PAWSON u. MC NEILLY 2005) 2.5.3 Unterschiede in der Regulation von FSH und LH GnRH bewirkt die Sekretion beider Gonadotropine. Es ist außerdem nötig für die Bereitstellung der Untereinheiten und damit auch für die Synthese von FSH und LH (SANCHEZ et al. 1994). Trotzdem gibt es Unterschiede in der Regulation der Gonadotropine (WALTERS et al. 1984; CUPP et al. 1995). WALTERS et al. (1984) beobachteten das Vorkommen von FSH-Pulsen, die nicht von einem LH-Puls begleitet wurden. Sie vermuteten, dass diese zusätzlichen FSH-Sekretionen von LHPulsen unterhalb der Messgrenze begleitet wurden. Mittlerweile kennt man verschiedene Faktoren, die neben GnRH Einfluss auf die FSHSynthese nehmen. Das vom Follikel synthetisierte Protein Inhibin hemmt die FSH-Synthese in der Hypophyse, während das ebenfalls im Ovar gebildetete Protein Aktivin die FSHSynthese fördert (YING 1988; MUTTUKRISHNA u. KNIGHT 1991; EVANS et al. 1997). Ein weiteres Protein mit hemmender Wirkung ist Follistatin (FSH-Suppressing-Protein, FSP), das von der Hypophyse sezerniert wird. Follistatin bindet an Aktivin und hemmt die FSHSekretion, ohne die GnRH- oder LH-Ausschüttung zu beeinflussen (WANG et al. 1990; BARATTA et al 2001; PADMANABHAN et al. 2002). WANG et al. (1990) vermuten außerdem einen hemmenden Einfluss von FSP auf GnRH über Enzymhemmung und Reduzierung von GnRH-Rezeptoren. 35 PADMANABHAN et al. (2002) untersuchten die GnRH-unabhängige episodische FSHSekretion und vermuten, dass die zusätzlichen Pulse durch einen weiteren hypothalamischen Releasingfaktor hervorgerufen werden. Dieser Faktor könnte eine Variante von GnRH sein. Als weitere Möglichkeit vermuten die Autoren eine zusätzliche GnRH-Rezeptorform, an die GnRH II bindet, das bevorzugt die FSH-Sekretion stimuliert (PADMANABHAN et al. 2002). Der Einfluss der GnRH-Pulsfrequenz auf die Gonadotropinsynthese wurde von einigen Autoren untersucht. Bei In-vitro-Versuchen von KAISER et al. (1997) und BÉDÉCARRATS und KAISER (2003) steigerten höhere Pulsfrequenzen die Synthese der α- und der LH-βUntereinheiten, die LH-Sekretion und auch den Gehalt an GnRH-Rezeptor-mRNA in der Hypophyse. Bei niedrigeren Pulsfrequenzen stieg die Synthese der FSH-β-Untereinheiten und die FSH-Sekretion an. Die Autoren schlussfolgern, dass die Regulation der Gonadotropinsynthese über die Pulsfrequenz auf Ebene der Hypophyse wirkt (KAISER et al. 1997). Bei Schafen steigerte die erhöhte Pulsfrequenz eines GnRH- Agonisten den Gehalt an LH-β-Untereinheiten, hatte aber keinen Einfluss auf die Menge an FSH-β-mRNA (MOLTER-GÉRARD et. al. 1999). ASPDEN et al. (2003) untersuchten über einen Zeitraum von 31 Tagen die Langzeitanwendung eines GnRH-Agonisten bei Färsen. Die Bildung der LH-βUntereinheiten-mRNA wurde gehemmt, es waren aber keine signifikanten Auswirkungen auf die Synthese der FSH-β-mRNA festzustellen. Die Gonadotropingehalte wurden aber stärker gehemmt als die der Untereinheiten, so dass die Autoren von einer post-transkriptionalen Hemmung der Gonadotropinsynthese ausgehen (ASPDEN et al. 2003). Auch LANE et al. (2009) beschreiben den Zusammenhang zwischen der Regulation der GnRH-Pulsfrequenz und der LH-Pulsfrequenz. Der FSH-Serumgehalt reguliert sich selber über die Menge an FSH in der Hypophyse und vielleicht auch in Änderungen der Hypophysensensitivität gegenüber den zirkulierenden Steroiden über die Regulierung der Rezeptoranzahl (LANE et al. 2009). 36 2.5.4 Einfluss von Östradiol, Progesteron und anderen Faktoren auf die Freisetzung von FSH/LH Auch andere Hormone haben Einfluss auf die Ausschüttung von FSH und LH. So können Progesteron und Östradiol je nach Zyklusstand die Gonadotropinsynthese hemmen oder fördern (KESNER u. CONVEY 1982; CONN et al. 1984). Progesteron hat während der Gelbkörperphase bis ungefähr zum 17. Zyklustag einen negativen Rückkopplungsmechanismus auf die Gonadotropinsekretion der Hypophyse (HANSEL u. SNOOK 1970; KALTENBACH et al. 1974; WALTERS et al. 1984). Außerdem senkt Progesteron in der Lutealphase die Sensitivität der Hypophyse für GnRH (RISPOLI u. NETT 2005). Auch Östradiol hat bei hoher Progesteronkonzentration hemmenden Einfluss auf die hypophyseale Gonadotropinausschüttung (WALTERS et al. 1984; FINK 1988). Bei sinkendem Plasmaprogesterongehalt als Folge der Gelbkörperregression führt eine neue Follikelwelle zum Anstieg der Östradiolkonzentration. Östradiol bewirkt dann die Steigerung der Hypophysensensibilität für GnRH über die Erhöhung der GnRH-Rezeptorsynthese in der Hypophyse im Sinne eines positiven Rückkopplungsmechanismus (CHENAULT et al 1974; STUMPF et al. 1989; KOJIMA et al. 1995; RISPOLI u. NETT 2005). Aktivin und Inhibin verändern ebenfalls die Hypophysensensitivität für GnRH über Änderung der Rezeptoranzahl (RISPOLI u. NETT 2005). Neben den Feedback-Effekten der gonadalen Steroide haben noch andere Faktoren, zum Beispiel Stress, Ernährung oder Jahreszeit Einfluss auf die Funktionen der gonadotropinproduzierenden Zellen. Auch Serotonin, Dopamin, Gamma-Aminobuttersäure (GABA), Noradrenalin, Opioide oder Glutamat können eine modulierende Wirkung auf die Synthese und Sekretion von FSH und LH haben (CLARKE u. POMPOLO 2005). 37 2.6 Therapeutischer Einsatz von GnRH-Analoga/Beeinflussung der LH-Freisetzung 2.6.1 GnRH als Medikament Seit vielen Jahren ist bekannt, dass eine Injektion von GnRH innerhalb von 30 Minuten zu einer Freisetzung von FSH und LH führt. In der Veterinärmedizin werden am häufigsten GnRH oder das synthetische Nonapeptid Buserelin verwendet (PETERS 2005). Eine einzelne Injektion von GnRH und GnRH-Agonisten bewirkt eine Freisetzung von FSH und LH in den peripheren Blutkreislauf für einen Zeitraum von drei Stunden. Je nach Zyklusstand zum Zeitpunkt der GnRH-Applikation hat das Hormon variable Wirkungen. Die GnRH-induzierte LH-Sekretion kann die Gelbkörperfunktion beeinflussen oder zu Follikelreifung, Ovulation und Luteinisierung von Follikeln führen (THATCHER et al. 1993). GnRH kommt zu unterschiedlichen therapeutischen Zwecken zum Einsatz. Zum Zeitpunkt der Besamung oder kurz danach wird GnRH injiziert, um die Ovulation und die Luteinisierung zu induzieren und damit embryonale Verluste zu vermeiden. Im Rahmen des Ovulationssynchronisationsverfahrens (OvSynch) zur terminierten Besamung wird GnRH in Kombination mit Prostaglandinen eingesetzt. Außerdem wird GnRH verwendet, um die Ovarfunktionen post partum zu stimulieren. Dabei ist der Einsatz bei azyklischen Milchkühen effektiver als bei Fleischrassen. Weiterhin gilt GnRH als Mittel der Wahl bei zystischen Ovarerkrankungen, um die Ovulation und Luteinisierung eines neuen Follikels und/oder die Luteiniserung der Zyste zu bewirken. Somit bestehen also verschiedene Einsatzmöglichkeiten von GnRH und dessen Analoga zur Verbesserung der Fertilität von Kühen (PETERS 2005). THATCHER et al. (1993) betonen, dass die Verwendung dieser Medikamente zur Verbesserung der Fruchtbarkeitsleistung kein Ausgleich für Fehler im Management sein darf. Bereits 1977 beschrieben NAWITO et al. (1977) die Anwendung eines synthetischen GnRH Dekapeptids bei zyklischen Kühen und Färsen in der Lutealphase. Die Injektion bewirkte eine Sekretion von FSH. Schon bei niedrigeren Dosen wurde die Freisetzung von LH stimuliert, die im Vergleich zu FSH außerdem stärker war und länger andauerte. 38 Im gleichen Jahr untersuchten HUMKE und ZUBER (1977) den Einsatz des LH-RH-Analogs „Hoe 766“ bei azyklischen Kühen. Dabei kamen 67,3 % der einmal behandelten Kühe in einem Zeitraum von 24 Tagen nach der Injektion in Brunst, von denen 93,3 % tragend wurden. Bei einem weiteren Teil der Tiere wurde ein ovarieller Zyklus mit Ovulation induziert, sie zeigten aber erst drei Wochen später eine Brunst. RILEY et al. (1981) behandelten azyklische Fleischkühe mit pulsatilen GnRH-Injektionen im Abstand von zwei Stunden über einen Zeitraum von 48 Stunden. Dies führte zum Einsetzen von LH-Pulsen, aber nicht immer zu FSH-Pulsen. Vier von fünf Tieren zeigten einen regulären präovulatorischen LH- und FSH-Anstieg, auf den mindestens ein vollständiger Brunstzyklus folgte. D'OCCHIO et al. (1989) untersuchten die Wirkung von kontinuierlicher Langzeitgabe von GnRH oder Buserelin über 28 Tage bei azyklischen Fleischkühen. Dabei wurde der LHGehalt im Blut gesteigert, während der FSH-Gehalt sich nicht änderte. Der bei der Behandlung entstandene Gelbkörper hatte eine kürzere Lebensdauer als normale Gelbkörper. Eine Vorbehandlung mit Progesteron konnte die Lebensdauer des GnRH-induzierten Gelbkörpers aber verlängern. Während der Behandlung erwarteten die Autoren ungefähr am zehnten Tag eine zweite Ovulation, aus der ein Gelbkörper mit normal langer Lebensdauer hervorgehen sollte. Diese Ovulation trat jedoch nicht auf, möglicherweise als Folge einer ungenügenden FSH-Stimulation des frühen Follikelwachstums (D'OCCHIO et al. 1989). Die Behandlung azyklischer Fleischkühe bis 26 Tage nach der Abkalbung über einen Zeitraum von 48 Stunden entweder mit wiederholten Injektionen von GnRH oder über denselben Zeitraum mit kontinuierlicher GnRH-Infusion wurde von JAGGER et al. (1987) untersucht. Dabei kam es bei den meisten Tieren beider Gruppen zur einer LH-Freisetzung, die dem präovulatorischen LH-Peak ähnelte. Die LH-Anstiege bei den injizierten Kühen traten aber später auf als bei den infundierten Tieren. Beide Behandlungsmethoden konnten aber nicht bei allen azyklischen Tieren eine Ovulation induzieren. Bei den Versuchen von TWAGIRAMUNGU et al. (1995) bewirkte eine Buserelininjektion bei zyklischen und azyklischen Kühen ein größeres Volumen des induzierten Gelbkörpers im Vergleich zu dem unbehandelter Tiere. Die Autoren folgerten, dass die Anzahl großer Luteinzellen durch die Buserelin-induzierte LH-Freisetzung gesteigert wurde, und damit das Gelbkörpervolumen ansteigt. Die Autoren gehen weiterhin von einem kurzlebigen Buserelin- 39 induzierten Gelbkörper bei den azyklischen Tieren und einem normal lang existierenden Corpus luteum bei zyklischen Kühen aus (TWAGIRAMUNGU et al. 1995). Bei den Untersuchungen von GOEHRING et al. (1999) an Milchkühen im Interöstrus zeigte eine Dosis von 20 Mikrogramm Buserelin als Einzelinjektion die größte Wirkung auf die LHSekretion im Vergleich zu höheren und niedrigeren Dosen. Dabei dauerte der Buserelininduzierte LH-Peak ca. drei bis vier Stunden. Bei einer wiederholten Buserelinapplikation 72 Stunden nach der ersten Injektion kam es zu einem deutlichen Anstieg der LH-Freisetzung. Die Autoren schlussfolgern deshalb, dass es eine Refraktärphase nach der Stimulierung der LH-freisetzenden Zellen gibt und eine wiederholte Injektion erst 72 Stunden nach der ersten Behandlung sinnvoll ist, da die wiederholte Verabreichung von GnRH-Analoga der pulsatilen endogenen Sekretion anscheinend nur bedingt entspricht (GOEHRING et al 1999). MACMILLAN und THATCHER (1991) untersuchten die Effekte von Buserelin bei Färsen und zyklischen Kühen. Dabei hatte eine Buserelininjektion an Tag 12 des Zyklus keinen Einfluss auf die Zykluslänge, induzierte aber bei einigen Tieren eine Ovulation und hatte Einfluss auf die Größe der einzelnen Follikel. FERNANDES et al. (1978) behandelten Milchkühen zu unterschiedlichen Zeitpunkten nach der Abkalbung mit einer Injektion von 100 Mikrogramm GnRH. Bereits fünf Minuten post injectionem stiegen die Plasma LH-Spiegel an, 120 Minuten später kam es zu einem LHPeak. Die Werte kehrten etwa fünf bis sechs Stunden später zum Ausgangsniveau zurück. Dabei zeigten Kühe wenige Tage post partum (drei Tage) eine niedrigere LH-Ausschüttung nach GnRH-Applikation als Tiere, bei denen die Kalbung schon länger (zehn bis 40 Tage) zurücklag. Die Autoren folgerten, dass der LH-Gehalt der Hypophyse in der postpartalen Periode ansteigt und ca. zwei Wochen post partum seinen vollen Wert erreicht. Deshalb sollten Programme zum Wiedererlangen der ovariellen Funktionen nicht vor einem Abstand von zehn Tagen nach der Abkalbung begonnen werden (FERNANDES et al. 1978). Viele weitere Untersuchungen wurden zur Behandlung azyklischer und anöstrischer Kühe durchgeführt. Allerdings ist ein Vergleich der Studien aufgrund der unterschiedlichen Definitionen und Vorgehensweisen nicht immer einfach. 40 2.6.2 Therapie der Azyklie Die Behandlung azyklischer Kühe ist laut OPSOMER und DE KRUIF (1999) wenig erfolgreich. Dabei sollten unbedingt eventuell vorhandene Primärleiden behandelt werden. Mit einer Hormontherapie mit einer intravaginalen Progesteronspirale über zehn bis 14 Tage werden nach deren Entfernung die Hypothalamus-Hypophysen-Achse stimuliert und damit Östrus und Ovulation induziert. Auch eine Behandlung mit equinem Chorion-Gonadotropin (eCG) ist möglich, die Trächtigkeitsergebnisse sind danach aber nicht befriedigend. RILEY et al. (1981) behandelten azyklische Fleischkühe mit 5 Mikrogramm GnRH alle zwei Stunden über eine Dauer von zwei Tagen. Danach kam es zu einem Anstieg der pulsatilen LH-Sekretion und bei vier von fünf behandelten Kühen zur Ovulation und zu einem kompletten ovariellen Zyklus. Nach der Behandlung anovulatorischer Milchkühe mit dem OvulationssynchronisationsVerfahren (OvSynch, Injektion von GnRH, sieben Tage danach PGF2α und zwei Tage später erneut GnRH) ovulierten 94 % der Tiere nach der zweiten GnRH-Injektion (GÜMEN et al. 2003). MC DOUGALL et al. (2004) therapierten anovulatorisch anöstrische Milchkühe mit verschiedenen Dosen von Progesteron als intravaginale Verabreichung in Kombination mit Östradiolbenzoat. Nach der Behandlung wurden Follikelwellen mit dominantem Follikel und Ovulation beobachtet. XU et al. (2000) verglichen die Behandlung von nichtzyklischen („noncycling“) Milchkühen mit GnRH, einer intravaginalen progesteronfreisetzenden Spange (controlled internal drug releasing device, CIDR), Prostaglandin und Östradiolbenzoat in Kombinationen gegenüber CIDR und Östradiolbenzoat. Dabei war die Trächtigkeitsrate beim ersten Programm höher als beim zweiten (47 % gegenüber 29 %). BALL und LAMMING (1983) therapierten azyklische Milchkühe mit einer Injektion von GnRH oder einer progesteronfreisetzenden Spirale (Progesterone releasing intravaginal device, PRID) für zehn Tage. Dabei zeigten 10,3 % der PRID-Gruppe und 26,1 % der GnRHGruppe keine ovarielle Aktivität. 41 2.6.3 Therapie von Anöstrus/Suböstrus Der Suböstrus mit einem gut entwickelten Gelbkörper wird mit Prostaglandinen therapiert. Diese Behandlung führt bei ca. 80 % der Tiere innerhalb von zwei bis sechs Tagen zu einem fruchtbaren Östrus (OPSOMER u. DE KRUIF 1999). JUBB et al. (1989) behandelten anöstrische Milchkühe mit einer Progesteron freisetzenden Spange (CIDR) über 7 Tage, und beim Entfernen der Spirale mit einer Injektion von 400 IE Pregnant Mare’s Serum Gonadotropin (PMSG). Das Intervall bis zum Einsetzen des ersten Östrus post partum, der Erstbesamungserfolg und die Trächtigkeitsrate unterschieden sich nicht zwischen den behandelten und unbehandelten Tieren und Kühen, die ein Placebo erhalten hatten (JUBB et al. 1989). Bei der Behandlung anöstrischer Milchkühe mit entweder Norgestomet (CRESTAR) und 500 IE equinem Chorion-Gonadotropin (eCG) oder dem OvSynch-Verfahren wurde bei allen behandelten Tieren beider Gruppen eine Brunst induziert (OLIVEIRA et al. 2003). BISHOP und WETTEMANN (1993) behandelten Fleischkühe mit fütterungsbedingter Anöstrie mit kleinen Dosen GnRH (2 µg GnRH pro Stunde bzw. 2 µg GnRH alle vier Stunden) über 14 Tage. Die GnRH-Infusion steigerte die Amplitude der LH-Pulse, aber nicht die Pulsfrequenz. Die LH-Konzentration im Serum stieg bei behandelten Kühen im Vergleich zu Tieren, die Kochsalzlösung erhielten. WALTERS et al. (1982) behandelten anöstrische Fleischkühe über vier Tage mit 500 ng GnRH alle zwei Stunden. Diese Therapie verkürzte die Zeit bis zum ersten Östrus und der ersten Ovulation post partum im Vergleich zu unbehandelten Tieren. MIALOT et al. (1999) verglichen verschiedene Behandlungsmethoden an Kühen, die nach der Abkalbung noch keine Brunst gezeigt haben, in Betrieben mit unterschiedlich guter Brunstbeobachtung. Sie empfehlen die Therapie des Suböstrus mit Prostaglandinen und anschließender künstlicher Besamung in Betrieben mit guter Brunstbeobachtung und das OvSynch-Programm bei schlechter Brunstbeobachtung (MIALOT et al. 1999). Die Behandlung anöstrischer Kühe mit Progesteron (CIDR) über einen Zeitraum von zehn Tagen steigert die pulsatile LH-Freisetzung. Eine zusätzliche Injektion von Östradiolbenzoat reduziert das Auftreten von "persistierenden" dominanten Follikeln (RHODES et al. 2002). 42 Auch bei Kühen, die keine sichtbare Brunst zeigen, wird das OvSynch-Verfahren als erfolgsversprechende Behandlungsmöglichkeit beschrieben, da die Trächtigkeitsergebnisse sehr ähnlich denen des OvSynch bei zyklischen Tieren sind (SHEPHARD 2005). 2.6.4 Therapie von Anöstrie und Azyklie ohne den Einsatz von GnRH WITTKOWSKI et al. (1982) behandelten Kühe mit Ovardystrophie mit einer Progesteron und Östradiolbenzoat enthaltenden Spirale (PRID). Bei zwei Drittel der behandelten Tiere wurde am 13. Tag post medicationem ein funktionsfähiger Gelbkörper nachgewiesen. Die Kühe der Behandlungsgruppe zeigten gegenüber den Kontrolltieren eine verkürzte Güstzeit. Kühen mit inaktiven Ovarien wurden von GABOR et al. (2002) 1500 IE eCG injiziert. Daraufhin zeigten zehn von 12 Kühen innerhalb von zehn Tagen nach der Behandlung eine Brunst (GABOR et al. 2002). Nach der Behandlung anovulatorisch anöstrischer Milchkühe mit CIDR über acht Tage und einer Injektion von Östradiolbenzoat verbesserte sich die Fruchtbarkeit der behandelten Tiere bezüglich des Zeitpunkts der ersten Besamung post partum und der Trächtigkeitsrate. (SEGWAGWE et al 2006) Auch ALANKO und PYÖRÄLÄ (1980) behandelten Tiere in Suböstrus und Anöstrus mit PRID-Spiralen. Dabei waren die Trächtigkeitsraten der suböstrischen Kühe besser als die der anöstrischen. Bei der Behandlung anöstrischer Tiere mit entweder PRID oder 2500 IE hCG zeigten sich bei beiden Therapiemethoden nahezu gleiche Trächtigkeitsergebnisse. MC DOUGALL et al. (2005) fanden heraus, dass die Erhöhung der Progesterondosis keine Auswirkungen auf die Behandlung anöstrischer Kühe hat, die Dauer der Behandlung mit der PRID-Spirale jedoch eine Rolle spielt. So hatte eine achttägige Behandlung einen höheren Erstbesamungserfolg als eine sechstägige Behandlung (MC DOUGALL et al. 2005). Anovulatorische anöstrische Milchkühe mit niedrigem Body Condition Score wurden von NATION et al. (2000) für fünf Tage mit CIDR behandelt. Am ersten Tag der Behandlung kam es zu einem vorübergehenden Anstieg der LH-Pulsfrequenz und LH-Konzentration. Die Auswirkungen des freigesetzten Progesterons auf das Follikelwachstum waren abhängig von der Größe des dominanten Follikels während der Behandlung. Insgesamt ist die Progesterontherapie jedoch bei anovulatorischen Kühen mit schlechter Körperkondition nicht 43 effektiv in Bezug auf die Auslösung eines LH-Peaks, einer Brunst oder der Ovulation (NATION et al. 2000). 44 3 Material und Methode 3.1 Zielsetzung Ziel dieser Arbeit war, herauszufinden, ob eine mehrmalige Verabreichung von kleinen Dosen des synthetischen GnRH-Agonisten Buserelin (Receptal®, Firma Intervet, Unterschleißheim) einen positiven Effekt auf die Fruchtbarkeit azyklischer Milchkühe hat im Vergleich zu einer einmaligen Injektion mit höherer Dosierung oder keiner Behandlung. 3.2 Auswahl der Betriebe , Auswahl und Untersuchung der Versuchstiere Die Feldstudie wurde in einem Praxisgebiet im Landkreis Diepholz in Niedersachsen in den Jahren 2008 bis 2010 durchgeführt. Die untersuchten 871 Tiere befanden sich in neun Milchviehbetrieben mit Bestandsgrößen zwischen ca. 50 und 1100 Milchkühen. Alle Tiere wurden in Laufställen mit Liegeboxen oder Fressliegeboxen gehalten. Die Kühe erhielten ihr Futter entweder als Total Mixed Ration (TMR) oder als Mischfuttervorlage mit einer zusätzlichen transpondergesteuerten und leistungsbezogenen Kraftfuttergabe. Die mittlere Milchleistung von acht an der Milchkontrolle teilnehmenden Betrieben betrug im milchwirtschaftlichen Kontrolljahr 2008/2009 9605,50 kg (Standardabweichung (SD) 809,92 kg, Minimum 8630 kg, Maximum 10692 kg) und im Jahr 2009/2010 9556,88 kg (SD 815,21 kg, Minimum 8111 kg, Maximum 10698 kg). Ein Betrieb nahm nicht an der Milchkontrolle teil. Die abgelieferte Milchmenge betrug im Jahr 2008/2009 9517 kg und im Jahr 2009/2010 8885 kg. Die Brunsterkennung auf den Höfen fand entweder über Brunstbobachtung durch die Landwirte oder mittels Pedometer statt. Die Kühe gehörten der Rasse Schwarzbunte Holstein Friesian (HF) oder Rotbunte HF an und hatten mindestens einmal und maximal achtmal gekalbt. 45 Die Kühe wurden von den Landwirten zur Untersuchung vorgestellt, wenn sie bis 50 Tage nach der Abkalbung noch keine Brunstsymptome gezeigt hatten und noch nicht besamt waren. Am Tag 50 post partum (p.p.) wurde zuerst eine transrektale Palpation der inneren Geschlechtsorgane vorgenommen. Kühe mit einem fühlbaren Gelbkörper oder Zysten schieden bereits jetzt aus der Studie aus. Bei den Tieren ohne Funktionskörper oder mit einem und mehreren Follikeln auf den Ovarien wurde eine vaginale Untersuchung mit einem Röhrenspekulum nach Götze durchgeführt, um eine Endometritis auszuschließen. Als Zeichen für einen Genitalkatarrh und damit als Ausschlusskriterien galten Asymmetrie der Uterushörner mit Fluktuation bei der rektalen Untersuchung sowie purulenter oder mukopurulenter Vaginalausfluss mit oder ohne krankhafter Hyperämie der Schleimhaut von Vestibulum, Vagina und Zervix. Nur Kühe ohne Endometritis konnten weiter an der Studie teilnehmen. Am 60. Tag nach der Abkalbung wurden die Kühe erneut rektal untersucht, wenn die Landwirte in der Zwischenzeit keine Brunst festgestellt hatten und die Kühe nicht besamt worden waren. War auf den Eierstöcken wieder kein Corpus luteum fühlbar, galten die Kühe als azyklisch und wurden in die Studie aufgenommen. Waren an den Tagen 50 oder 60 post partum bei den Untersuchungen Zeichen für eine Brunst erkennbar, wurden die Tiere ebenfalls abgelehnt. Als Brunstanzeichen galten, neben dem typischen Brunstverhalten, erhöhte Kontraktilität des Uterus in Verbindung mit einem Follikel auf einem Ovar, Abgang von Brunstschleim, Ödematisierung der Vulva und Hyperämie der Schleimhaut. Bei der rektalen und vaginalen Untersuchung wurde das Beurteilungsschema von GRUNERT (1999) angewendet. Außerdem wurde an den Untersuchungstagen 50, 60, 65, 75 und 100 p.p. jeweils eine Beurteilung der Körperkondition (Body Condition Score, BCS) nach der Methode von EDMONDSON et al. (1989) durchgeführt. Die erhobenen Befunde wurden auf einem Formular festgehalten. 46 Insgesamt wurden 871 Kühe untersucht, von denen 555 Tiere bei der ersten Untersuchung und 153 Tiere bei der zweiten Untersuchung aufgrund der oben genannten Gründe ausschieden. Drei Tiere schieden aus anderen Gründen aus, und wurden bei den weiteren Berechnungen nicht berücksichtigt. 178 Tiere erfüllten die angesetzten Kriterien und wurden in die Feldstudie einbezogen. Die in die Studie aufgenommenen Kühe wurden nach dem Zufallsprinzip einer der drei Versuchsgruppen (Gruppe 50, 51 oder 55) zugeordnet. Tabelle 1 zeigt die untersuchten Tiere aufgeteilt nach Betrieben und die Einteilung in die Versuchsgruppen. Tab. 1: Anzahl ausgeschiedener und in die Studie aufgenommener Kühe aufgeteilt nach Betrieben und Versuchsgruppen Betrieb Kühe* (n = 4612) A (n = 104) B (n = 142) C (n = 204) D (n = 442) E (n = 226) F (n = 404) G (n = 448) H (n = 360) I (n = 2242) ausgeschiedene Kühe 1.Untersuchunga ausgeschiedene Kühe 2.Untersuchungb Gruppe 501 Gruppe 512 Gruppe 553 Gesamt (n = 555) 1 3 6 61 45 8 52 13 366 (n = 135) 1 1 6 13 8 0 12 0 94 (n = 66) 0 0 1 9 7 0 7 0 42 (n = 55) 0 2 2 5 7 0 6 0 33 (n = 57) 0 1 0 11 4 0 11 0 30 (n = 868) 2 7 15 99 71 8 88 13 565 *: Kühe ab 1. Laktation im Untersuchungszeitraum von zwei Jahren a,b: rektale Palpation der inneren Geschlechtsorgane an den Tagen 50 (1. Untersuchung) und 60 (2. Untersuchung) post partum 1: Kontrollgruppe 2: täglich 1 ml Receptal i.m. Tag 60 bis 64 post partum 3: 5 ml Receptal i.m. Tag 60 post partum Die Tiere der Gruppe 50 (n = 66) blieben als Kontrollgruppe unbehandelt. Den Kühen der Gruppe 51 (n = 55) wurde an fünf aufeinanderfolgenden Tagen (Tag 60 bis 64 p.p.) jeweils 1 ml Receptal® (Firma Intervet, Unterschleißheim) intramuskulär injiziert und die Tiere der Gruppe 55 (n = 57) bekamen an Tag 60 p.p. eine intramuskuläre Injektion mit 5 ml Receptal®. Der Wirkstoff von Receptal® ist das synthetische GnRH-Analog Buserelinacetat in einer Konzentation von 4 Mikrogramm (µg) Buserelin pro Milliliter Injektionslösung und bewirkt 47 nach Injektion die Freisetzung von Follikelstimulierendem Hormon (FSH) und Luteinisierendem Hormon (LH). An den Tagen 65 und 75 nach der Abkalbung wurden die Probanden erneut rektal untersucht und der BCS ermittelt. Eine weitere rektale Palpation der Geschlechtsorgane fand ungefähr am Tag 100 post partum statt. Kühe, die zu diesem Zeitpunkt nicht besamt waren, erhielten bei Vorhandensein eines Gelbkörpers eine intramuskuläre Injektion mit 2 ml Cyclix® (Firma Intervet, Unterschleißheim). Cyclix® enthält 250 µg Cloprostenol als Cloprostenol-Natrium und bewirkt als Prostaglandin F2α-Analogon eine Luteolyse. War zum Untersuchungszeitpunkt kein Corpus luteum fühlbar, wurden den Tieren 2 ml Receptal® injiziert. Die Untersuchungen und Behandlungen sind zusammengefasst in Tabelle 2 dargestellt. Nach der Besamung wurde eine Trächtigkeitsuntersuchung transrektal palpatorisch ab dem 40. Trächtigkeitstag oder ab dem 28. Tag mit einem Ultraschallgerät (Fa. Pie Medical) durchgeführt. Tab.2: Schematische Darstellung und zeitliche Einordnung der Untersuchungen und Behandlungen Tage post partum 50 60 61 62 63 64 65 75 100 rektale Palpation von Uterus und Ovarien X Vaginoskopie mittels Röhrenspekulum X Erfassung des Body Condition Score X X X X X X X X X Injektion von Receptal® 5 ml (Gr.55) X Injektion von Receptal® 5x1 ml (Gr.51) X nicht besamte Tiere mit Gelbkörper: Injektion von Cyclix® 2 ml nicht besamte Tiere ohne Gelbkörper: Injektion von Receptal® 2 ml X X X X X X 48 3.3 Weitere Datenerfassung Neben den Besamungsdaten wurden noch weitere Parameter von den Landwirten erfragt. Dazu gehörte die Milchleistung zum Zeitpunkt der zweiten Milchkontrolle nach der Abkalbung. Hier wurde anhand der Ergebnisse der Milchleistungsprüfung erfasst, ob die Kühe bei dieser Kontrolle mehr oder weniger als 40 kg Milch gaben. Außerdem wurde festgehalten, wie oft die Tiere bereits gekalbt hatten und eine Aufteilung in Tiere in der ersten Laktation und Kühe mit zwei und mehr Laktationen vorgenommen. Zusätzlich wurden die im Folgenden erläuterten Parameter ermittelt. Die Rastzeit (RZ) gibt den Zeitraum von der Kalbung bis zur ersten Besamung in Tagen an. Zielgröße ist eine maximale Dauer von 85 Tagen. Als Güstzeit (GZ) oder Zwischentragezeit bezeichnet man die Dauer von der Kalbung bis zum ersten Trächtigkeitstag. Zielgröße ist eine Zeit von unter 105 Tagen. Aus der Differenz von Rastzeit und Güstzeit errechnet sich die Verzögerungszeit (VZ), die bei unter 18 Tagen liegen sollte. Der Erstbesamungserfolg (EBE) ist der Prozentsatz nach Erstbesamung tragender Tiere im Verhältnis zur Anzahl tragender Tiere. Es wird ein Erstbesamungserfolg von mindestens 50 % angestrebt. Die Gesamtträchtigkeitsrate (GTR) ist die Anzahl der tragenden Tiere dividiert durch die Anzahl der besamten Tiere. Die GTR wird in Prozent angegeben und soll bei mindestens 90 % liegen. Es wurde die GTR aus allen Besamungen der beobachteten Laktation sowie die GTR aus Besamungen bis Tag 200 p.p. ermittelt. Der Trächtigkeitsindex (TI) gibt die Anzahl der Besamungen bei tragenden Tieren im Verhältnis zur Anzahl tragender Tiere an. Hierbei wird ein Wert von kleiner oder gleich 1,7 angestrebt. Der TI wurde für alle Besamungen der beobachteten Laktation und für Besamungen bis Tag 200 p.p. errechnet. Außerdem wurden die Anzahl der Besamungen pro Kuh in der gesamten Laktation und bis zum 200. Tag nach der Abkalbung erfasst (DE KRUIF et al. 2007). 49 3.4 Statistische Auswertung Die statistischen Berechnungen und Auswertungen wurden mit dem Programm Statistical Package for the Social Science SPSS, Version 15.0.1 1996, Fa. SPSS Inc., Somers, USA und dem Programm Excel® 2008 aus Office:mac 2008 Home and Student Edition, Fa. Microsoft Corporation, Redmond, USA durchgeführt. Zuerst wurden die Daten mit dem Kolmogorov-Smirnov-Test auf Normalverteilung überprüft. Zum Vergleich von Häufigkeiten wurde im Anschluss der Chi-Quadrat-Test durchgeführt. Die Mittelwerte der nicht normal verteilten Daten wurden mittels KruskalWallis-Test und Mann-Whitney-U-Test als post-hoc-Test verglichen. Als Signifikanzniveau wurde p kleiner oder gleich 0,05 festgelegt (FIELD 2009). 50 4 Ergebnisse 4.1 Gesamtpopulation und Stichprobenumfang In neun Betrieben mit insgesamt 4612 Tieren ab der ersten Laktation wurden in den Jahren 2008 bis 2010 868 Kühe mit dem Vorbericht „unbeobachtete Brunst in den ersten 50 Tagen nach der Abkalbung“ zur Untersuchung vorgestellt. Dies entspricht einem Anteil von 18,8 % an der zur Verfügung stehenden Gesamtpopulation. Bei der ersten Untersuchung am Tag 50 post partum (p.p.) schieden bereits 555 Tiere (12,0 % der Gesamtpopulation bzw. 63,9 % der vorgestellten Tiere) aus, die die angesetzten Kriterien nicht erfüllten. 313 Kühe wurden am Tag 60 p.p. erneut untersucht, von denen 135 Tiere (2,9 %, 15,6 %) ausgeschlossen wurden. Gründe für das Ausscheiden bei den Untersuchungen an den Tagen 50 oder 60 p.p. waren tastbare Gelbkörper, Endometritis, Anzeichen einer Brunst oder Ovarialzysten. Tabelle 3 zeigt die Anzahl der untersuchten, ausgeschlossenen und in die Studie aufgenommenen Tiere sowie die Gründe für das Ausscheiden. Von den 868 vorgestellten Tieren schieden insgesamt 690 Kühe im Laufe der ersten beiden Untersuchungen aus. Die Kriterien für Azyklie erfüllten 178 Kühe, die in die Studie einbezogen wurden. Dies entspricht einem Anteil von 3,9 % an der zur Verfügung stehenden Gesamtpopulation von 4612 Tieren beziehungsweise von 20,5 % der 868 zur Untersuchung vorgestellten Kühe. Die 178 Probanden wurden nach dem Zufallsprinzip in drei Gruppen aufgeteilt. In der Kontrollgruppe 50 waren 66 Kühe. Den 55 Tieren der Gruppe 51 wurden an fünf aufeinander folgenden Tage jeweils 1 ml Receptal® injiziert und die 57 Tiere der Gruppe 55 bekamen eine Injektion mit 5 ml Receptal®. 51 Tab. 3: Anzahl der untersuchten, ausgeschlossenen und in die Studie aufgenommenen Tiere sowie die Gründe für das Ausscheiden Kühe* (n) Untersuchung Tag 50 post partum (p.p.) insgesamt ausgeschieden am Tag 50 p.p. Aufteilung nach Ursachen Untersuchung Tag 60 p.p. insgesamt ausgeschieden am Tag 60 p.p. Aufteilung nach Ursachen Kriterien erfüllt und in Studie einbezogen (n = 4612) Anteil (%) an der Untersuchungspopulation (n = 868) 868 18,8 100,0 555 12,0 63,9 7,8 3,5 0,5 0,2 0,1 41,5 18,7 2,7 0,8 0,4 313 6,8 36,1 135 2,9 15,6 2,0 0,9 0,04 0,02 10,6 4,6 0,2 0,1 3,9 20,5 360 162 23 7 3 92 40 2 1 Gründe für den Ausschluss aus der Studie Gelbkörper Endometritis Brunst Ovarialzyste Sonstiges Gelbkörper Brunst Sonstiges Abgang 178 Anteil (%) an der Gesamtpopulation * Anzahl Kühe mit dem Vorbericht „unbeobachtete Brunst in den ersten 50 Tagen nach der Abkalbung“ von 4612 Tieren aus neun Betrieben. 52 4.2 Ergebnisse in den Versuchsgruppen 4.2.1 Anzahl der Kühe mit Gelbkörper am Tag 65 oder 75 p.p. in den Versuchsgruppen Die Probanden wurden an den Tagen 65 und 75 p.p. rektal untersucht, um festzustellen, ob auf den Ovarien ein fühlbarer Gelbkörper vorhanden war. In Tabelle 4 ist die Anzahl an Kühen, die am Tag 65 p.p. oder 75 p.p. ein tastbares Corpus luteum (Cl) hatten, aufgeteilt nach Versuchsgruppen und insgesamt dargestellt. Es besteht ein signifikanter Zusammenhang (p≤0,0001) zwischen den unterschiedlichen Versuchsgruppen und dem Vorhandensein eines Gelbkörpers an den Tagen 65 oder 75 nach der Abkalbung. Dabei ist bei den Tieren der Gruppe 51 signifikant seltener ein Gelbkörper zu diesem Zeitpunkt fühlbar als bei den Kühen der Gruppen 50 (p≤0,0001) und 55 (p≤0,0001). Bei den Kühen der unbehandelten Gruppe 50 ist signifikant seltener ein Cl am Tag 65 oder 75 p.p. tastbar als bei den Tieren der Gruppe 55 (p=0,033). Tab. 4: Anzahl der Tiere mit tastbarem Corpus luteum (Cl) am Tag 65 oder 75 post partum (p.p.) in den Versuchsgruppen Gruppe Kühe n Cl am Tag 65/75 p.p. % 50 51 55 Gesamt Kontrollgruppe 5 x 1 ml GnRH1) 1 x 5 ml GnRH2) 178 66 55 57 43,9 a 14,5 b 63,2 c 41,0 a-c: Werte mit unterschiedlichen Buchstabenindizes unterscheiden sich statistisch signifikant (p≤0,05). 1) Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. 53 4.2.2 Rastzeit, Güstzeit und Verzögerungszeit in den Versuchsgruppen Von n = 178 Probanden in drei Gruppen wurden 170 Tiere im Laufe der Laktation künstlich besamt. In der Kontrollgruppe 50 wurden 64 von 66 Kühen besamt, in der Gruppe 51 wurden 51 von 55 Tieren besamt, und in der Gruppe 55 wurden 55 von 57 Kühen besamt. Die mittlere Rastzeit (RZ) bei den Tieren aller Gruppen zusammen betrug 83,34 Tage, die mittlere Güstzeit (GZ) 150,08 Tage, und die mittlere Verzögerungszeit (VZ) lag bei 67,05 Tagen. In Tabelle 5 sind die mittleren Rastzeiten, Güstzeiten und Verzögerungszeiten mit Standardabweichung und Median für die einzelnen Gruppen und insgesamt dargestellt. Die mittleren Rastzeiten, Güstzeiten und Verzögerungszeiten unterscheiden sich zwischen den einzelnen Gruppen nicht signifikant (RZ: p=0,616, GZ: p=0,735, VZ: p=0,600). Tab. 5: Mittelwerte mit Standardabweichung (SD) und Median für Rastzeit, Güstzeit und Verzögerungszeit in den Versuchsgruppen Gruppe Kühe n Rastzeit in Tagen Mittelwert SD Median Güstzeit in Tagen Mittelwert SD Median Verzögerungszeit in Tagen Mittelwert SD Median 50 Kontrollgruppe 51 55 1) 5 x 1 ml GnRH 1 x 5 ml GnRH2) Gesamt 64 51 55 170 84,25 17,55 86,50 80,69 12,78 78,00 84,73 20,55 82,00 83,34 17,35 82,00 152,63 80,20 140,00 153,96 81,46 142,00 141,88 70,73 120,50 150,08 77,74 136,50 67,75 78,94 45,00 74,15 81,12 63,00 57,70 67,81 39,00 67,05 76,50 46,00 1) Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 54 4.2.3 Mittlere Anzahl der Besamungen in den Versuchsgruppen In jeder der drei Versuchsgruppen zeigten einige Tiere keine Brunstsymptome und wurden deshalb nicht besamt. Diese Kühe wurden bei der Berechnung der mittleren Anzahl an künstlichen Besamungen in den einzelnen Gruppen nicht mitgerechnet. Insgesamt wurden 170 von 178 Probanden künstlich besamt. Es wurde die Anzahl der Besamungen (KB) bis einschließlich Tag 200 p.p. sowie die Anzahl der Besamungen im Verlauf der gesamten Laktation ausgewertet. Tabelle 6 zeigt die Anzahl besamter Tiere in den Gruppen und die mittlere Anzahl an Besamungen pro Kuh bis Tag 200 p.p. und in der gesamten Laktation. Die mittlere Anzahl der Besamungen der Probanden aller drei Gruppen zusammen betrug im Mittel 3,43 KB pro Tier im Verlauf der gesamten Laktation. Die Mindestanzahl betrug eine Besamung und das Maximum waren 18 KB. Die mittlere Anzahl an Besamungen pro Kuh bis zum Tag 200 p.p. lag bei 2,63 KB pro Kuh mit einem Minimum von einer Besamung und einem Maximum von sieben Besamungen. Die mittlere Anzahl der Besamungen während der gesamten Laktation und bis zum Tag 200 p.p. unterscheidet sich zwischen den Gruppen 50, 51 und 55 nicht signifikant (KB insgesamt: p=0,738, KB Tag 200: p=0,450). 55 Tab. 6: Mittlere Anzahl Besamungen (KB) pro Kuh während der gesamten Laktation und bis Tag 200 post partum mit Standardabweichung (SD), Median, minimaler und maximaler Anzahl KB in den Versuchsgruppen Gruppe 50 51 55 Kontrollgruppe 5 x 1 ml GnRH1) 1 x 5 ml GnRH2) 66 55 57 178 64 51 55 170 Mittelwert SD Median Minimum Maximum Anzahl KB pro Kuh bis Tag 200 post partum 3,44 2,90 3,00 1 16 3,53 2,82 3,00 1 14 3,33 3,17 2,00 1 18 3,43 2,95 3,00 1 18 Mittelwert SD Median Minimum Maximum 2,64 1,43 2,00 1 7 2,84 1,70 3,00 1 6 2,42 1,42 2,00 1 6 2,63 1,52 2,00 1 7 Kühe n besamte Tiere n Anzahl KB pro Kuh während der gesamten Laktation 1) Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 56 Gesamt 4.2.4 Erstbesamungserfolg (EBE) in den Versuchsgruppen Von n = 178 Probanden wurden 170 Tiere künstlich besamt. Acht Kühe zeigten keine Brunstsymptome und wurden nicht besamt. Von den besamten Tieren waren 44 (25,9 %) tragend und 126 Kühe (74,1 %) nicht tragend von der ersten Besamung nach der Abkalbung. In Tabelle 7 sind die Erstbesamungserfolge in den Versuchsgruppen dargestellt. Die Erstbesamungserfolge in den drei Gruppen unterscheiden sich statistisch nicht signifikant (p=0,953). Tab. 7: Erstbesamungserfolg (EBE) in den Versuchsgruppen Gruppe Kühe n besamte Tiere n EBE (%) 50 51 55 Kontrollgruppe 5 x 1 ml GnRH1) 1 x 5 ml GnRH2) 66 55 57 178 64 51 55 170 25,0 27,5 25,5 25,9 1) Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. Gesamt Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 4.2.5 Gesamtträchtigkeitraten und Trächtigkeitsindizes in den Versuchsgruppen Von n = 178 Probanden wurden 170 besamt, von denen 117 Kühe tragend bis Tag 200 post partum waren. 144 Tiere waren tragend waren nach allen Besamungen im Verlauf der gesamten Laktation. 57 4.2.5.1 Gesamtträchtigkeitsraten in den Versuchsgruppen Die Gesamtträchtigkeitsrate (GTR) bis Tag 200 nach der Abkalbung lag bei den Probanden aller Gruppen bei 68,8 %, und die GTR aller KB der gesamten Laktation betrug 84,7 %. Die Gesamtträchtigkeitsraten der einzelnen Gruppen und aller Tiere zusammen sind in Tabelle 8 beschrieben. Die GTR bis Tag 200 p.p. unterscheiden sich zwischen den einzelnen Gruppen nicht signifikant (p=0,228). Die GTR aller KB der beobachteten Laktation unterscheidet sich signifikant zwischen den drei Gruppen (p=0,010). Die GTR aller Besamungen während der Laktation unterscheidet sich nicht signifikant zwischen der Kontrollgruppe 50 und der Gruppe 51 (p=0,575). Die GTR insgesamt der Gruppe 55 ist signifikant niedriger als in den Gruppen 50 und 51 (p=0,022, p=0,009). Tab. 8: Gesamtträchtigkeitsrate (GTR) bis Tag 200 post partum (p.p.) und GTR aller Besamungen (KB) der Laktation in den Versuchsgruppen Gruppe Kühe n besamte Tiere n GTR (%) bis Tag 200 p.p. GTR (%) aller KB der Laktation 50 51 55 Kontrollgruppe 5 x 1 ml GnRH1) 1 x 5 ml GnRH2) 66 55 57 178 64 51 55 170 73,4 72,5 60,0 68,8 89,1 a 92,2 a 72,7 b 84,7 1) Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. Gesamt a,b: Werte mit unterschiedlichen Buchstabenindizes unterscheiden sich statistisch signifikant (p≤0,05) 58 4.2.5.2 Trächtigkeitsindizes in den Versuchsgruppen Die mittlere Anzahl der Besamungen pro tragend gewordenem Tier (Trächtigkeitsindex, TI) lag für alle Gruppen zusammen bei 2,28 KB bis Tag 200 p.p. Das Minimum war eine Besamung, das Maximum bis Tag 200 waren sechs Besamungen. Der TI aus allen Besamungen der beobachteten Laktation lag bei 3,20 KB mit einer Mindestanzahl von einer Besamung und einem Maximum von 14 KB. Tabelle 9 zeigt die Trächtigkeitindices der einzelnen Gruppen. Die Trächtigkeitsindizes nach Besamungen bis Tag 200 p.p. und nach allen Besamungen der Laktation unterscheiden sich zwischen den Versuchsgruppen 50, 51 und 55 nicht signifikant (TI Tag 200 p.p.: p=0,645, TI alle KB: p=0,522). 59 Tab. 9: Mittlere Trächtigkeitsindizes bis Tag 200 post partum (p.p.) und nach allen Besamungen (KB) der gesamten Laktation mit Standardabweichung (SD), Minimum und Maximum in den Versuchsgruppen Gruppe 50 51 55 Kontrollgruppe 5 x 1 ml GnRH1) 1 x 5 ml GnRH2) 66 55 57 178 64 51 55 170 47 37 33 117 2,34 1,32 1 6 2,38 1,40 1 5 2,09 1,23 1 5 2,28 1,32 1 6 57 3,19 2,50 1 13 47 3,47 2,75 1 14 40 2,90 2,45 1 11 144 3,20 2,56 1 14 Kühe n besamte Tiere n tragende Tiere bis Tag 200 p.p. n Trächtigkeitsindex (TI) bis Tag 200 p.p. SD Minimum Maximum tragende Tiere nach allen KB der Laktation n TI aller KB SD Minimum Maximum 1) Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. Gesamt Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 4.2.6 Anzahl besamter Tiere, Untersuchung und Behandlung am Tag 100 post partum Von n = 178 untersuchten Kühen ging ein Tier aus Gruppe 55 wegen einer Ketose und ein Tier aus Gruppe 51 aufgrund schlechter Milchleistung vor der Untersuchung am Tag 100 p.p. ab. Sie wurden bei den Berechnungen der Ereignisse am Tag 100 nicht mitgerechnet. 60 Von den restlichen 176 Probanden waren bis zum Tag 100 nach der Abkalbung 145 Tiere besamt. In Gruppe 50 waren 53 (80,3 %) von 66 Tieren, in Gruppe 51 46 (85,2 %) von 54 Kühen und in Gruppe 55 46 (82,1 %) von 56 Tieren besamt bis Tag 100 p.p. Es besteht zwischen den Gruppen kein signifikanter Unterschied (p = 0,670) bei der Anzahl der bis Tag 100 p.p. besamten Tiere. Die zu diesem Zeitpunkt nicht besamten Tiere wurden untersucht und gegebenenfalls erneut behandelt. Tabelle 10 zeigt die Anzahl der besamten Tiere in den einzelnen Gruppen sowie die weiteren Befunde und Behandlungen der bis Tag 100 p.p. nicht besamten Tiere. Auf einen statistischen Vergleich der Anzahl der Kühe, denen Prostaglandin oder GnRH injiziert wurde oder die am Tag 100 p.p. in Brunst waren oder eine Ovarialzyste hatten, wurde verzichtet, da die Anzahl der Tiere in den einzelnen Gruppen bei diesen Parametern zu gering war. Tab. 10: Kühe (%), die bis Tag 100 post partum besamt wurden, sowie weitere Untersuchungsergebnisse und Behandlungen zu diesem Zeitpunkt Gruppe 50 51 55 Kontrollgruppe 5 x 1 ml GnRH1) 1 x 5 ml GnRH2) Gesamt Kühe n besamte Tiere bis Tag 100 post partum (%) Prostaglandin Injektion (%) GnRH - Injektion oder OvSynch (%) 66 54 56 176 80,3 85,2 82,1 82,4 15,2 1,9 3,6 7,4 3,0 13,0 3,6 6,3 Brunst (%) 1,5 0,0 8,9 3,4 Ovarialzyste (%) 0,0 0,0 1,8 0,6 1) Behandlung: täglich von Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. 61 4.3 Ergebnisse in Milchleistungsgruppen 4.3.1 Aufteilung der Kühe in Leistungsgruppen Anhand der Ergebnisse der Milchleistungsprüfung der zweiten Milchkontrolle nach der Abkalbung wurden die Kühe in zwei Leistungsgruppen eingeteilt. Die Tagesleistung der Gruppe 1 lag bei mehr als 40 kg Milch, die Kühe der Gruppe 2 gaben ≤ 40 kg Milch. Von den 178 Probanden wurden 46 Tiere (25,8 %) in Gruppe 1 und 132 Tiere (74,2 %) in Gruppe 2 eingeteilt. Aufgrund der geringen Tierzahl in Gruppe 1 wurde auf eine weitere Unterteilung der Leistungsgruppen in die einzelnen Versuchsgruppen verzichtet. 4.3.2 Anzahl der Tiere mit Gelbkörper am Tag 65 oder 75 p.p. in den Leistungsgruppen Von den Kühen der Leistungsgruppe 1 hatten 56,5 % am Tag 65 oder 75 nach der Abkalbung keinen tastbaren Gelbkörper. Bei den restlichen 43,5 % war zu diesen Untersuchungszeitpunkten ein Corpus luteum festzustellen. In der Gruppe 2 mit der niedrigeren Milchleistung war bei 59,9 % am Tag 65 oder 75 p.p. kein Gelbkörper fühlbar, 40,2 % hatten bei den Untersuchungen ein palpierbares Corpus luteum. In Tabelle 11 ist die Anzahl der Kühe mit und ohne Gelbkörper aufgeteilt nach Leistungsgruppen dargestellt. Statistisch lässt sich kein signifikanter Zusammenhang (p=0,693) zwischen den Leistungsgruppen und dem Vorhandensein eines tastbaren Gelbkörpers am Tag 65 oder 75 nach der Abkalbung feststellen. 62 Tab. 11: Kühe (%) mit tastbarem Corpus luteum (Cl) am Tag 65 oder Tag 75 post partum (p.p.) in den Leistungsgruppen Gruppe Kühe n Corpus luteum (Cl) am Tag 65 oder 75 p.p. (%) 1 Milchleistung > 40 kg bei der zweiten Leistungsprüfung p.p. 2 Milchleistung ≤ 40 kg bei der zweiten Leistungsprüfung p.p. Gesamt 46 132 178 43,5 40,2 41,0 Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 63 4.3.3 Rastzeit, Güstzeit und Verzögerungszeit in den Leistungsgruppen Von den 46 Tieren der Gruppe 1 mit einer Milchleistung über 40 kg wurden 45 Tiere besamt. In der Leistungsgruppe 2 (Milchleistung ≤ 40 kg) wurden von 132 Kühen 125 besamt. In Tabelle 12 sind die mittleren Rastzeiten, Güstzeiten und Verzögerungszeiten der Leistungsgruppen und insgesamt aufgeschlüsselt. Statistisch ließ sich kein Zusammenhang zwischen den Leistungsgruppen und der Rastzeit, Güstzeit oder Verzögerungszeit nachweisen (RZ: p=0,323, GZ: p=0,969, VZ: p=0,695). Tab. 12: Mittelwerte mit Standardabweichung (SD) und Median für Rastzeit, Güstzeit und Verzögerungszeit in den Leistungsgruppen Gruppe 1 Milchleistung > 40 kg bei der zweiten Leistungsprüfung post partum 2 Milchleistung ≤ 40 kg bei der zweiten Leistungsprüfung post partum Gesamt 46 132 178 45 125 170 Mittelwert SD Median Güstzeit in Tagen 85,62 18,32 80,00 82,51 16,99 82,00 83,34 17,35 82,00 Mittelwert SD Median Verzögerungszeit in Tagen 154,73 88,63 127,00 148,47 74,00 140,00 150,08 77,74 136,50 Mittelwert SD Median 68,59 89,98 37,00 66,51 71,72 50,00 67,05 76,50 46,00 Kühe n besamte Tiere n Rastzeit in Tagen Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 64 4.3.4 Mittlere Anzahl der Besamungen in den Leistungsgruppen In der Leistungsgruppe 1 mit der höheren Milchleistung wurden 45 von 46 Kühen besamt. Die Mindestanzahl lag bei einer Besamung, das Maximum bei 14 KB während der gesamten Laktation und bei sechs KB bis zum Tag 200 p.p. In der Leistungsgruppe 2 wurden 125 von 132 Tieren besamt. Hier lag das Minimum bei einer Besamung, das Maximum waren 18 KB insgesamt und sieben Besamungen bis zum Tag 200 p.p. In Tabelle 13 sind die mittleren Besamungshäufigkeiten nach Milchleistung aufgeteilt und insgesamt dargestellt. Die Mittelwerte der Besamungen insgesamt und bis Tag 200 p.p. unterscheiden sich zwischen den Leistungsgruppen nicht signifikant (KB insgesamt: p=0,837, KB bis T. 200: p=0,889). Tab. 13: Mittlere Anzahl Besamungen (KB) pro Kuh während der gesamten Laktation und bis Tag 200 post partum (p.p.) mit Standardabweichung (SD) und Median in den Leistungsgruppen Gruppe 1 Milchleistung > 40 kg bei der zweiten Leistungsprüfung p.p. 2 Milchleistung ≤ 40 kg bei der zweiten Leistungsprüfung p.p. Gesamt 46 132 178 45 125 170 Mittelwert SD Median Anzahl KB pro Kuh bis Tag 200 p.p. 3,67 3,21 2,00 3,34 2,86 3,00 3,43 2,95 3,00 Mittelwert SD Median 2,71 1,67 2,00 2,60 1,46 2,00 2,63 1,52 2,00 Kühe n besamte Tiere n Anzahl KB pro Kuh in der gesamten Laktation Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 65 4.3.5 Erstbesamungserfolg in den Leistungsgruppen In der Leistungsgruppe 1 mit der höheren Milchleistung wurden 45 Kühe künstlich besamt, von denen 26,7 % nach der ersten Besamung tragend waren. In der niedrigeren Leistungsgruppe 2 waren 25,6 % der Kühe tragend von der ersten Besamung. In Tabelle 14 ist der EBE in den Leistungsgruppen und insgesamt sowie die Anzahl besamter und tragender Kühe beschrieben. Es besteht kein signifikanter Zusammenhang zwischen der Milchleistung und dem Erstbesamungserfolg (p=0,889). Tab. 14: Erstbesamungserfolg (EBE) in den Leistungsgruppen Gruppe Kühe n besamte Tiere n EBE (%) 1 Milchleistung > 40 kg bei der zweiten Leistungsprüfung post partum (p.p.) 2 Milchleistung ≤ 40 kg bei der zweiten Leistungsprüfung p.p. Gesamt 46 132 178 45 125 170 26,7 25,6 25,9 Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 66 4.3.6 Gesamtträchtigkeitsraten in den Leistungsgruppen Von n = 170 besamten Tieren waren 45 Kühe in der höheren Leistungsgruppe 1 und 125 Kühe in Gruppe 2 mit einer Milchleistung von unter 40 kg bei der zweiten Kontrolle nach der Abkalbung. Die Gesamtträchtigkeitsrate (GTR) bis Tag 200 nach der Abkalbung betrug 66,7 % in der Gruppe 1 und 69,9 % in der Gruppe 2. Die GTR der Leistungsgruppe 1 lag bei 82,2 %, die der Gruppe 2 bei 85,6 % bezogen auf alle Besamungen in der beobachteten Laktation. In Tabelle 15 sind die Gesamtträchtigkeitsraten aufgeteilt nach Leistungsgruppen und insgesamt dargestellt. Die Gesamtträchtigkeitsraten bis Tag 200 p.p. und nach allen Besamungen im Verlauf der Laktation unterscheiden sich zwischen den Leistungsgruppen nicht signifikant (GTR Tag 200 p.p.: p=0,716, GTR insgesamt: p=0,589). Tab. 15: Gesamtträchtigkeitsrate (GTR) bis Tag 200 post partum (p.p.) und GTR nach allen Besamungen (KB) der Laktation in den Leistungsgruppen Gruppe Kühe n besamte Tiere n GTR (%) bis Tag 200 p.p GTR (%) aller KB der Laktation 1 Milchleistung > 40 kg bei der zweiten Leistungsprüfung post partum (p.p.) 2 Milchleistung ≤ 40 kg bei der zweiten Leistungsprüfung p.p. Gesamt 46 132 178 45 125 170 66,7 69,6 68,8 82,2 85,6 84,7 Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 67 4.3.7 Trächtigkeitsindizes in den Leistungsgruppen In Tabelle 16 sind die Trächtigkeitsindizes aus Besamungen bis Tag 200 p.p. und aus allen Besamungen der gesamten Laktation aufgeteilt nach Leistungsgruppen dargestellt. Die TI der Tiere mit hoher und niedrigerer Milchleistung unterscheiden sich nicht signifikant (TI Tag 200 p.p.: p=0,697, TI alle KB: p=0,966). Tab. 16: Mittlere Trächtigkeitsindizes mit Standardabweichung (SD), Minimum und Maximum bis Tag 200 post partum (p.p.) und nach allen Besamungen (KB) der Laktation in den Leistungsgruppen Gruppe Kühe n besamte Tiere n tragende Tiere bis Tag 200 p.p. n Trächtigkeitsindex (TI) bis Tag 200 p.p. SD Minimum Maximum tragende Tiere nach allen KB der Laktation n TI der gesamten Laktation SD Minimum Maximum 1 Milchleistung > 40 kg bei der zweiten Leistungsprüfung p.p. 2 Milchleistung ≤ 40 kg bei der zweiten Leistungsprüfung p.p. Gesamt 46 132 178 45 125 170 30 87 117 2,23 1,41 1 6 2,30 1,30 1 5 2,28 1,32 1 6 37 107 144 3,54 3,30 1 14 3,08 2,26 1 11 3,20 2,56 1 14 Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 68 4.4 Ergebnisse der Färsen und Kühe Durch Unterschiede in der Physiologie von erstlaktierenden Färsen und Kühen können die Fruchtbarkeitsparameter dieser Tiere in ihren Werten voneinander abweichen (MARKUSFELD 1987). Um zu überprüfen, ob die Laktationsnummer Einfluss auf die Behandlung der Azyklie hat, wurde in der vorliegenden Studie eine Unterteilung der Versuchstiere in Einkalbige und Mehrkalbskühe vorgenommen. 4.4.1 Aufteilung der Tiere nach Laktationsnummer und Versuchsgruppe In den Versuchsgruppen und der Kontrollgruppe wurden die Kühe anhand der Anzahl der Abkalbungen noch einmal in zwei Laktationsgruppen aufgeteilt. Tiere, die einmal gekalbt hatten, bildeten die Untergruppe 1 (Färsen), Tiere mit zwei und mehr Abkalbungen waren in Untergruppe 2 (Kühe). Von n = 178 untersuchten Kühen hatten 88 (49,4 %) einmal gekalbt und 90 Kühe (50,6 %) hatten zweimal oder häufiger gekalbt. Tabelle 17 zeigt die Aufteilung nach Laktation innerhalb der Versuchsgruppen. Tab. 17: Tiere (%) in der ersten Laktation in den Versuchsgruppen und insgesamt Gruppe Tiere n 1. Laktation (%) 50 Kontrollgruppe 51 5 x 1 ml GnRH1) 55 1 x 5 ml GnRH2) Gesamt 66 55 57 178 50,0 50,9 47,4 49,4 1) Behandlung: täglich Tag 60 bis Tag 64 post partum i.m. 2) Behandlung: Tag 60 post partum i.m. 69 4.4.2 Anzahl der Färsen und Kühe mit Gelbkörper am Tag 65 oder 75 post partum 4.4.2.1 Vergleich der Färsen und Kühe aller Versuchsgruppen zusammen Bei den Untersuchungen an den Tagen 65 und 75 p.p. hatten 34,1 % der Färsen und 47,8 % der Kühe ab der zweiten Laktation einen tastbaren Gelbkörper. Die Anzahl der Tiere mit fühlbarem Corpus luteum an einem oder an beiden Untersuchungstagen unterscheidet sich zwischen den Kühen und den Färsen aller drei Gruppen zusammen nicht signifikant (p=0,063). In Tabelle 18 ist die Anzahl der Kühe und Färsen mit und ohne Gelbkörper an den Untersuchungstagen 65 und 75 p.p. dargestellt. Tab. 18: Tiere (%) mit tastbarem Corpus luteum (Cl) am Tag 65 oder Tag 75 post partum (p.p.) in den Laktationsgruppen Laktation Tiere n Cl am Tag 65 oder 75 p.p. (%) 1. Laktation ab 2. Laktation Gesamt 88 90 178 34,1 47,8 41,0 Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 4.4.2.2 Vergleich der Erstlaktierenden und der Mehrkalbskühe innerhalb der Versuchsgruppen In der Kontrollgruppe 50 hatten 33,3 % der Färsen und 54,6 % der Mehrkalbskühe einen fühlbaren Gelbkörper am Tag 65 oder 75 p.p. Tabelle 19 zeigt die Anzahl der Färsen und Kühe mit und ohne Gelbkörper am Tag 65 oder 75 p.p. in den einzelnen Gruppen. Es besteht in Gruppe 50 kein signifikanter Zusammenhang zwischen der Laktationsnummer und dem Auftreten eines Corpus luteum zu diesem Zeitpunkt (p=0,083). In der Versuchsgruppe 51 war bei 7,1 % der Färsen und bei 22,2 % der Kühe ein Gelbkörper palpierbar. In dieser Gruppe besteht kein signifikanter Zusammenhang zwischen der 70 Laktationsnummer und dem Vorhandensein eines Gelbkörpers nach der Behandlung (p=0,113). In der Versuchsgruppe 55 hatten 63,0 % der Erstlaktierenden und 63,3 % der Mehrkalbskühe einen Gelbkörper. Auch in dieser Gruppe besteht kein signifikanter Zusammenhang zwischen der Anzahl der Abkalbungen und der Ausbildung eines Corpus luteum nach der Behandlung (p=0,977). Tab. 19: Tiere (%) mit tastbarem Corpus luteum (Cl) am Tag 65 oder 75 post partum (p.p.) aufgeteilt nach Laktationsnummern und Gruppen Gruppe Tiere n Cl am Tag 65/76 p.p. 50 51 55 Gesamt Kontrollgruppe 1. ab 2. Laktation Laktation 5 x 1 ml GnRH1) 1. ab 2. Laktation Laktation 1 x 5 ml GnRH2) 1. ab 2. Laktation Laktation 33 33 28 27 27 30 33,3 a 54,5 1 7,1 b 22,2 2 63,0 c 63,3 1 1) Behandlung: täglich Tag 60 bis Tag 64 post partum i.m. 2) Behandlung: Tag 60 post partum i.m. 41,0 a-c: Werte mit unterschiedlichen Buchstabenindizes unterscheiden sich statistisch signifikant (p≤0,05). 1,2: Werte mit unterschiedlichen Zahlenenindices unterscheiden sich statistisch signifikant (p≤0,05). 4.4.2.3 Vergleich der Erstlaktierenden der Versuchsgruppen untereinander In der unbehandelten Gruppe 50 hatten 33,3 % der Färsen einen tastbaren Gelbkörper am Tag 65 oder 75 nach der Abkalbung, in der Versuchsgruppe 51 war bei 7,1 % der Färsen und in der Gruppe 55 war bei 63,0 % der Erstkalbinnen ein Corpus luteum zu fühlen. Es besteht ein signifikanter Zusammenhang zwischen der Versuchsgruppe und dem Vorhandensein eines Gelbkörpers am Tag 65 oder 75 p.p. (p≤0,0001). 71 Dabei war bei den Tieren in Gruppe 51 signifikant seltener ein Corpus fühlbar als bei den Tieren der Gruppen 55 (p≤0,0001) und 50 (p=0,013). Die Färsen der unbehandelten Gruppe 50 hatten signifikant seltener einen Gelbkörper als die Färsen der Gruppe 55 (p=0,022). 4.4.2.4 Vergleich der Mehrkalbskühe der Versuchsgruppen untereinander Von 33 Kühen ab der 2. Laktation hatten in der Gruppe 50 am Tag 65 oder 75 nach der Abkalbung 54,6 % einen tastbaren Gelbkörper, in der Gruppe 51 war bei 22,2 % ein Corpus luteum vorhanden. Bei 63,3 % der Mehrkalbskühe der Gruppe 55 war ein Gelbkörper zu fühlen. Es besteht bei den Kühen ab der 2. Laktation ein signifikanter Zusammenhang zwischen der Versuchsgruppe und dem Vorhandensein eines Gelbkörpers am Tag 65 oder 75 p.p. (p=0,005). Kühe der Gruppe 51 hatten zu diesem Zeitpunkt signifikant seltener einen fühlbaren Gelbkörper als Tiere in Gruppe 50 (p=0,011) oder in Gruppe 55 (p=0,002). Bei den Gruppen 50 und 55 besteht kein signifikanter Zusammenhang zwischen der Versuchsgruppe und dem Vorhandensein eines Gelbkörpers (p=0,479). 4.4.3 Vergleich der Rastzeit, Güstzeit und Verzögerungszeit von Färsen und Kühen 4.4.3.1 Vergleich der Färsen und Kühe aller Versuchsgruppen zusammen Von 88 Färsen wurden 84 Tiere besamt, in der Gruppe der Kühe wurden 86 von 90 Tieren besamt. Tabelle 20 zeigt die mittlere RZ, GZ und VZ der Färsen und der Kühe ab der zweiten Laktation. Die RZ, GZ und VZ der Färsen und Kühe unterscheiden sich nicht signifikant (RZ: p=0,927, GZ: p=0,441, VZ: p=0,654). 72 Tab. 20: Mittelwerte für Rastzeit, Güstzeit und Verzögerungszeit mit Standardabweichung (SD) und Median in den Laktationsgruppen Gruppe Tiere n besamte Tiere n Rastzeit in Tagen Mittelwert SD Median Güstzeit in Tagen Mittelwert SD Median Verzögerungszeit in Tagen Mittelwert SD Median 1. Laktation ab 2. Laktation Gesamt 88 90 178 84 86 170 82,23 13,23 85,00 84,42 20,63 79,00 83,34 17,35 82,00 144,88 73,94 141,00 156,04 82,05 135,00 150,08 77,74 136,50 63,23 72,09 46,00 71,43 81,61 45,00 67,05 76,50 46,00 Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 4.4.3.2 Vergleich der Rastzeit, Güstzeit und Verzögerungszeit der Färsen und Kühe innerhalb der Versuchsgruppen Von 33 Erstlaktierenden der Kontrollgruppe 50 wurden 32 Tiere künstlich besamt und von 33 Mehrkalbskühen dieser Gruppe wurden 32 Kühe besamt. In Tabelle 21 sind die mittleren Rastzeiten, Güstzeiten und Verzögerungszeiten der Färsen und Kühe der Kontrollgruppe und der Versuchsgruppen aufgeführt. Es besteht kein signifikanter Unterschied bei den Fruchtbarkeitskennzahlen Rastzeit (p=0,514), Güstzeit (p=0,250) und Verzögerungszeit (p=0,283) zwischen den Färsen und den Mehrkalbskühen der Kontrollgruppe.In der Versuchsgruppe 51 wurde von 26 von 28 Färsen und 25 von 27 Mehrkalbskühen besamt. Zwischen den mittleren Rastzeiten, Güstzeiten und Verzögerungszeiten der Färsen und der Kühe dieser Gruppe besteht kein signifikanter Unterschied (RZ: p=0,322, GZ: p=0,932, VZ: 73 p=0,803). In der Versuchsgruppe 55 wurden 26 von 27 Erstlaktierenden und 29 von 30 Kühen besamt. Es besteht auch in der Gruppe 55 kein signifikanter Unterschied zwischen den oben genannten Fruchtbarkeitskennzahlen der Färsen und denen der Mehrkalbskühe (RZ: p=0,258, GZ: p=1,000 VZ: p=0,638). Tab. 21: Mittelwerte für Rastzeit, Güstzeit und Verzögerungszeit mit Standardabweichung (SD) und Median aufgeteilt nach Versuchsgruppen und Laktationsnummern Gruppe Laktationsnummer Tiere n besamte Tiere n Rastzeit in Tagen Mittelwert SD Median Güstzeit in Tagen Mittelwert SD Median Verzögerungszeit in Tagen Mittelwert SD Median 50 51 55 Kontrollgruppe 1. ab 2. Laktation Laktation 5 x 1 ml GnRH1) 1. ab 2. Laktation Laktation 1 x 5 ml GnRH2) 1. ab 2. Laktation Laktation 33 33 28 27 27 30 32 32 26 25 26 29 83,34 11,28 88,00 85,16 22,29 80,50 81,81 11,35 83,50 79,52 14,27 77,00 81,27 17,08 79,00 87,83 23,08 85,00 147,97 91,55 133,00 157,81 66,74 144,00 144,92 56,45 148,00 165,14 105,07 127,00 140,43 67,87 138,00 143,47 75,60 118,00 64,67 89,11 41,50 71,19 67,40 47,00 63,12 53,71 60,00 87,81 105,71 64,00 61,33 67,98 38,00 53,68 69,24 40,00 1) Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). Die in 4.4.3.1 und Tabelle 21 angegebenen Werte gelten auch für die Berechnung von 4.4.3.2 und 4.4.3.3. 74 4.4.3.3 Vergleich der Rastzeit, Güstzeit und Verzögerungszeit der Färsen der Versuchsgruppen untereinander Zwischen den Färsen der Gruppen 50, 51 und 55 gibt es keine signifikanten Unterschiede bei der mittleren Dauer der Rastzeit (p=0,585), der Güstzeit (p=0,737) und der Verzögerungszeit (p=0,732). 4.4.3.4 Vergleich der Rastzeit, Güstzeit und Verzögerungszeit der Mehrkalbskühe der Versuchsgruppen untereinander Die Mittelwerte von Rastzeit, Güstzeit und Verzögerungszeit unterscheiden sich bei den Mehrkalbskühen in den einzelnen Versuchsgruppen nicht signifikant (RZ: p=0,294, GZ: p=0,670, VZ: p=0,486). 4.4.4 Anzahl der Besamungen bei Färsen und Kühen 4.4.4.1 Vergleich der Färsen und Kühe aller Versuchsgruppen zusammen In der Gruppe der Erstlaktierenden wurden 84 von 88 Tieren besamt. Die mittlere Anzahl an Besamungen lag bei 3,07 KB (Standardabweichung (SD) 2,18, Median 2,50) pro Tier mit einer Mindestanzahl von einer Besamung und einem Maximum von 11 KB im Verlauf der gesamten beobachteten Laktation. Bei der Anzahl der Besamungen bis Tag 200 p.p. betrug der Mittelwert 2,58 KB (SD 1,42, Median 2,00), das Minimum war eine Besamung und das Maximum waren sechs KB. In Gruppe der Kühe mit zwei und mehr Laktationen wurden 86 von 90 Tieren besamt. Die mittlere Anzahl waren 3,78 KB (SD 3,52, Median 3,00) pro Tier mit einer minimalen Anzahl von einer Besamung und einem Maximum von 18 KB während der gesamten Laktation. Bei der Anzahl der Besamungen bis Tag 200 p.p. war der Mittelwert bei 2,67 KB (SD 1,61, Median 2,00), mit einer Mindestanzahl von einer KB und einem Maximum von sieben Besamungen. Es besteht bei den Probanden aller drei Gruppen zusammengefasst kein 75 signifikanter Unterschied zwischen den Laktationsnummern bei der mittleren Anzahl der Besamungen bis Tag 200 p.p. (p=0,921) und bei der mittleren Anzahl der Besamungen insgesamt (p=0,611). 4.4.4.2 Vergleich der Anzahl der Besamungen bei Färsen und Kühen innerhalb der Versuchsgruppen Bei den 32 besamten Erstlaktierenden der Kontrollgruppe 50 lag die mittlere Anzahl an Besamungen bis zum Tag 200 p.p. bei 2,22 KB pro Tier und bei 2,81 Besamungen während der gesamten Laktation. Die 32 Mehrkalbskühe der Gruppe 50 wurden bis zum Tag 200 p.p. im Mittel 3,06 Mal besamt. Der Mittelwert aller Besamungen der gesamten Laktation lag bei diesen Tieren bei 4,06 KB pro Kuh. Die mittlere Anzahl an Besamungen bis zum Tag 200 nach der Abkalbung ist bei den Erstlaktierenden signifikant geringer als bei den Mehrkalbskühen (p=0,031). Bei der mittleren Anzahl der Besamungen in der gesamten Laktation gibt es keinen signifikanten Unterschied zwischen den Färsen und den Tieren ab der zweiten Laktation (p=0,086). Tabelle 22 zeigt die mittlere Anzahl der Besamungen pro Tier aufgeschlüsselt nach Laktationsnummern und Versuchsgruppen. Die mittlere Anzahl an Besamungen bis Tag 200 p.p. lag bei den 26 besamten Färsen der Gruppe 51 bei 2,88 KB pro Tier und während der gesamten Laktation bei 3,12 Besamungen pro Tier. Bei den Mehrkalbskühen wurden in dieser Gruppe 25 Tiere besamt. Der Mittelwert bis Tag 200 nach der Abkalbung betrug 2,80 KB pro Kuh und 3,96 Besamungen pro Tier insgesamt. Die Mittelwerte der Anzahl der Besamungen pro Tier bis zum Tag 200 p.p. und in der gesamten Laktation unterscheiden sich zwischen den Erstkalbinnen und den Mehrkalbskühen nicht signifikant (KB bis Tag 200 p.p.: p=0,728, KB insgesamt: p=0,863). Die 26 besamten Färsen der Versuchsgruppe 55 hatten bis zum Tag 200 p.p. eine mittlere Anzahl von 2,73 KB pro Tier. Innerhalb der gesamten Laktation lag der Mittelwert bei 3,35 KB pro Färse. Bei den 29 besamten Kühen dieser Gruppe lag der Mittelwert bei 2,14 Besamungen pro Tier bis Tag 200 p.p. und bei 3,31 KB pro Kuh in der gesamten Laktation. 76 Es gibt keinen signifikanten Unterschied zwischen den mittleren Besamungshäufigkeiten pro Tier bis Tag 200 p.p. und während der gesamten Laktation zwischen den Färsen und den Mehrkalbskühen der Gruppe 55 (KB bis Tag 200 p.p.: p=0,145, KB insgesamt: p=0,402). Tab. 22: Mittlere Anzahl der Besamungen (KB) pro Tier bis Tag 200 post partum (p.p.) und in der gesamten Laktation aufgeteilt nach Versuchsgruppen und Laktationsnummern mit Standardabweichung (SD), Median, Minimum und Maximum Gruppe Laktation Tiere n besamte Tiere n KB pro Tier bis Tag 200 p.p. 50 51 55 Kontrollgruppe 1. ab 2. Laktation Laktation 5 x 1 ml GnRH1) 1. ab 2. Laktation Laktation 1x 5 ml GnRH2) 1. ab 2. Laktation Laktation 33 33 28 27 27 30 32 32 26 25 26 29 Mittelwert SD Median Minimum Maximum KB pro Tier in der gesamten Laktation 2,221 1,10 2,00 1 5 3,062 1,61 3,00 1 7 2,881 1,58 3,00 1 6 2,801 1,85 2,00 1 6 2,731 1,54 2,00 1 6 2,141 1,27 2,00 1 5 Mittelwert SD Median Minimum Maximum 2,811 2,18 2,00 1 10 4,061 2,40 3,50 1 16 3,121 1,84 3,00 1 7 3,961 3,55 2,00 1 14 3,351 2,53 2,50 1 11 3,311 3,70 2,00 1 18 1) Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. 1,2: p≤0,05 (1. Laktation vs. Laktation > 1 innerhalb der Versuchsgruppen) Die Werte aus 4.4.4.2 und Tabelle 22 gelten auch für die Berechnung von 4.4.4.3 und 4.4.4.4. 77 4.4.4.3 Vergleich der Anzahl der Besamungen bei den Färsen der Versuchsgruppen untereinander Die Mittelwerte der Anzahl an Besamungen in der gesamten Laktation und bis zum Tag 200 nach der Abkalbung unterscheiden sich bei den Färsen der Gruppen 50, 51 und 55 nicht signifikant (KB insgesamt: p=0,581, KB bis Tag 200 p.p.: p=0,274). 4.4.4.4 Vergleich der Anzahl der Besamungen bei den Mehrkalbskühen der Versuchsgruppen untereinander Bei den Mehrkalbskühen der Gruppen 50, 51 und 55 gab es keine signifikanten Unterschiede bei der mittleren Anzahl der Besamungen insgesamt (p=0,287) und bis zum Tag 200 p.p. (p=0,072). 4.4.5 Erstbesamungserfolg der Färsen und Kühe Von n = 178 Probanden wurden 170 Tiere besamt. Der Erstbesamungserfolg der Tiere aller Gruppen zusammen lag bei 25,9 %. Es wurden 28,6 % der Färsen und 23,3 % der Mehrkalbskühe von der ersten Besamung nach der Abkalbung tragend. Der EBE unterscheidet sich nicht signifikant zwischen den Kühen und den Färsen der drei Gruppen zusammen (p=0,429). Tabelle 23 zeigt den EBE aufgeteilt nach Laktationsnummer und Versuchsgruppe. 78 Tab. 23: Erstbesamungserfolg (EBE) der Färsen und der Mehrkalbskühe in den Versuchsgruppen Gruppe Laktation besamte Tiere n EBE (%) 50 Kontrollgruppe 1. ab 2. Laktation Laktation 51 1) 5 x 1 ml GnRH 1. ab 2. Laktation Laktation 55 2) 1 x 5 ml GnRH 1. ab 2. Laktation Laktation Gesamt 1. Laktation ab 2. Laktation 32 32 26 25 26 29 84 86 31,3 18,8 26,9 28,0 26,9 24,1 28,6 23,3 1) Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). Die Werte aus Tabelle 23 gelten auch für die Berechnungen in den Abschnitten 4.4.5.1 und 4.4.5.2. 4.4.5.1 Erstbesamungserfolg der Färsen und Kühe in den Versuchsgruppen In der Kontrollgruppe 50 lag der EBE der Färsen bei 31,3 %, der EBE der Mehrkalbskühe war 18,8 %. Es gibt keinen signifikanten Zusammenhang zwischen der Laktationsnummer und dem EBE (p=0,248). In der Versuchsgruppe 51 waren 26,9 % der Färsen und 28,0 % der Kühe tragend aus der Erstbesamung. Es besteht kein signifikanter Zusammenhang zwischen der Laktationsnummer und dem EBE (p=0,931). In der behandelten Gruppe 55 betrug der EBE der Färsen 26,9 %, während 24,1 % der Kühe von der ersten Besamung tragend waren. Auch hier besteht kein signifikanter Zusammenhang zwischen Laktationsnummer und EBE (p=0,813). 79 4.4.5.2 Erstbesamungserfolg der Färsen und der Kühe zwischen den Versuchsgruppen Der Erstbesamungserfolg der Färsen unterscheidet sich zwischen den Versuchsgruppen nicht signifikant (p=0,913). Auch bei den Erstbesamungserfolgen der Kühe gibt es keinen signifikanten Unterschied zwischen den Gruppen 50, 51 oder 55 (p=0,708). 4.4.6 Gesamtträchtigkeitraten der Färsen und Kühe Von n = 178 Tieren wurden 84 Färsen und 86 Mehrkalbskühe besamt. Die Gesamtträchtigkeitsrate bis Tag 200 p.p. war in allen Gruppen zusammen bei den Färsen 78,6 % gegenüber 59,3 % bei den Kühen. Bei allen Besamungen im Verlauf der Laktation lag die GTR der Färsen bei 91,7 %, die GTR der Kühe betrug 77,9 %. Die GTR bis Tag 200 p.p. und nach allen Besamungen im Verlauf der Laktation sind bei den Färsen signifikant höher als bei den Kühen (GTR Tag 200 p.p.: p=0,007, GTR insgesamt: p=0,013). In Tabelle 24 sind die GTR der Färsen und Kühe insgesamt und aufgeteilt nach Versuchsgruppen beschrieben. Tab. 24: Gesamtträchtigkeitsrate (GTR) bis Tag 200 post partum und GTR nach allen Besamungen (KB) der Laktation aufgeteilt nach Färsen und Mehrkalbskühen und Versuchsgruppen Gruppe Laktation besamte Tiere n GTR (%) bis Tag 200 post partum GTR (%) aller KB der Laktation 50 Kontrollgruppe 1. ab 2. Laktation Laktation 51 1) 5 x 1 ml GnRH 1. ab 2. Laktation Laktation 55 2) 1 x 5 ml GnRH 1. ab 2. Laktation Laktation Gesamt 1. Laktation ab 2. Laktation 32 32 26 25 26 29 84 86 81,3 1 65,6 1 84,6 1 60,0 2 69,2 1 51,7 1 78,6 1 59,3 2 93,8 84,4 1 100 1,a 84,0 2 80,8 1,b 65,6 1 91,7 1 77,9 2 1,a,b 1) Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m. 2) Behandlung: Tag 60 p.p. i.m. 1,2: 1. Laktation vs. Laktation > 1 innerhalb der Versuchsgruppen (p≤0,05) a,b: Gruppenvergleich Färsen (p≤0,05) 80 Die Angaben der Tabelle 24 gelten auch für die Berechnungen der Werte in den Abschnitten 4.4.6.1. und 4.4.6.2. 4.4.6.1 Vergleich der Gesamtträchtigkeitsraten der Färsen und Kühe innerhalb der Versuchsgruppen Die GTR bis Tag 200 p.p. und nach allen KB der Laktation unterscheiden sich zwischen den Färsen und den Kühen innerhalb der Gruppen 50 und 55 nicht signifikant (KB bis Tag 200 p.p.: Gruppe 50: p=0,157, Gruppe 55: p=0,186, alle KB: Gruppe 50: p=0,230, Gruppe 55: p=0,205). In der Gruppe 51 sind beide GTR bei den Färsen der Gruppe signifikant höher als bei den Kühen (GTR Tag 200 p.p.: p=0,049, GTR alle KB: p=0,034). 4.4.6.2 Vergleich der Gesamtträchtigkeitsraten der Färsen und der Kühe zwischen den Versuchsgruppen Die GTR bis Tag 200 p.p. der Färsen der Gruppen 50, 51 und 55 unterscheiden sich nicht signifikant (p=0,359). Bei den GTR aller KB der Laktation gibt es signifikante Unterschiede zwischen den Färsen der einzelnen Gruppen (p=0,037). Dabei ist die GTR in der Versuchsgruppe 51 signifikant höher als bei den Färsen der Gruppe 55 (p=0,019). Die GTR aller KB der Färsen der Kontrollgruppe 50 unterscheidet sich nicht signifikant von denen der Gruppe 51 (p=0,195) und der Gruppe 55 (p=0,131). Die GTR der Kühe ab der zweiten Laktation bis zum Tag 200 p.p. und nach allen Besamungen der Laktation unterscheiden sich nicht signifikant zwischen den Versuchsgruppen 51 und 55 sowie der Kontrollgruppe 50 (GTR Tag 200 p.p.: p=0,542, GTR alle KB: p=0,142). 81 4.4.7 Trächtigkeitsindizes der Färsen und Kühe Von 84 besamten Färsen wurden 66 Tiere tragend bis Tag 200 p.p. und 77 Färsen trugen nach allen Besamungen der gesamten Laktation. Der Trächtigkeitsindex (TI) betrug 2,35 bis Tag 200 p.p. und 2,99 nach allen Besamungen. Es wurden insgesamt 86 Mehrkalbskühe besamt, davon trugen 51 Tiere bis Tag 200 p.p. und 67 Tiere wurden nach allen KB der Laktation tragend. Der TI lag bei 2,20 bis Tag 200 p.p. und bei 3,45 nach allen KB der Laktation. Die Trächtigkeitsindizes bis Tag 200 p.p. und nach allen KB der Laktation unterscheiden sich zwischen den Kühen und den Färsen aller Gruppen nicht signifikant (TI Tag 200 p.p.: p=0,565, TI alle KB: p=0,673). Tabelle 25 zeigt die Trächtigkeitsindizes aufgeteilt nach Laktationsnummer und Versuchsgruppe. Tab. 25: Mittlerer Trächtigkeitsindex (TI) bis Tag 200 post partum (p.p.) und nach allen Besamungen (KB) der Laktation aufgeteilt nach Färsen und Mehrkalbskühen und Versuchsgruppen mit Standardabweichung (SD), Minimum und Maximum Gruppe Laktation besamte Tiere n tragende Tiere bis Tag 200 p.p. n TI bis Tag 200 p. p. SD Minimum Maximum tragende Tiere nach allen KB der Laktation n TI aller KB der Laktation SD Minimum Maximum 1) 50 Kontrollgruppe 1. ab 2. Laktation Laktation 51 1) 5 x 1 ml GnRH 1. ab 2. Laktation Laktation 55 2) 1 x 5 ml GnRH 1. ab 2. Laktation Laktation Gesamt 1. Laktation ab 2. Laktation 32 32 26 25 26 29 84 86 26 21 22 15 18 15 66 51 2,15 1,19 1 5 2,57 1,47 1 6 2,64 1,50 1 5 2,00 1,20 1 4 2,28 1,40 1 5 1,87 0,99 1 4 2,35 1,35 1 5 2,20 1,28 1 6 30 27 26 21 21 19 77 67 2,83 3,59 3,12 3,90 3,05 2,74 2,99 3,45 2,25 1 10 2,75 1 13 1,84 1 7 3,58 1 14 2,52 1 11 2,42 1 10 2,17 1 11 2,95 1 14 Behandlung: täglich Tag 60 bis Tag 64 p.p. i.m., 2) Behandlung: Tag 60 p.p. i.m. Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 82 Die Werte in Tabelle 25 gelten auch für die Berechnungen der Abschnitte 4.4.7.1 und 4.4.7.2. 4.4.7.1 Trächtigkeitsindizes der Färsen und Kühe innerhalb der Versuchsgruppen In den Versuchsgruppen 51 und 55 sowie der unbehandelten Gruppe 50 gibt es keinen signifikanten Unterschied zwischen den Färsen und den Kühen ab der zweiten Laktation bei den TI bis Tag 200 p.p. und nach allen KB der gesamten Laktation (Gr. 50: TI Tag 200 p.p.: p=0,352, TI alle KB: p=0,227, Gr.51: TI Tag 200 p.p.: p=0,199, TI alle KB: p=0,931, Gr.55: TI Tag 200 p.p.: p=0,481, TI alle KB: p=0,635). 4.4.7.2 Trächtigkeitsindizes der Färsen und Kühe zwischen den Versuchsgruppen Die TI bis Tag 200 p.p. und nach allen KB der Laktation der Färsen in den Gruppen 50, 51 und 55 unterscheiden sich nicht signifikant (TI Tag 200 p.p.: p=0,547, TI alle KB: p=0,632). Auch zwischen den Trächtigkeitsindizes der Kühe in den Gruppen 50, 51 und 55 gibt es keinen signifikanten Unterschied (TI Tag 200 p.p.: p=0,281, TI alle KB: p=0,400). 4.5 Ergebnisse in Körperkonditionsgruppen 4.5.1 Aufteilung der Tiere nach Körperkondition Zur Untersuchung des Einflusses der Körperkondition auf die Fruchtbarkeit azyklischer Milchkühe wurden die Probanden anhand ihrer Körperkondition am Tag 60 nach der Abkalbung in drei Gruppen (BCS-Gruppen) eingeteilt. Kühe mit einem BCS von 2,5 oder weniger bildeten die Gruppe 1, Tiere mit einem BCS von 2,75 oder 3,0 waren in Gruppe 2 und Kühe deren BCS 3,25 oder größer war wurden in Gruppe 3 eingeordnet. In Gruppe 1 befanden sich 41 Tiere, in Gruppe 2 waren 106 Kühe und in Gruppe 3 waren 31 Tiere. Aufgrund der geringen Tierzahl in Gruppe 1 und 3 wurde auf eine weitere Aufteilung nach den Versuchsgruppen verzichtet. 83 4.5.2 Anzahl der Tiere mit Gelbkörper am Tag 65 oder 75 post partum in den BCS-Gruppen Von n = 178 Tieren hatten 59,0 % am Tag 65 oder 75 p.p. keinen palpierbaren Gelbkörper, bei 41,0 % der Kühe war ein Corpus luteum fühlbar. In Gruppe 1 mit den niedrigsten BCSWerten waren 41 Kühe, von denen 65,1 % einen palpierbaren Gelbkörper hatten. In Gruppe 2 mit mittlerer Körperkondition hatten 40,6 % von 106 Tieren ein Corpus luteum. Die 31 Probanden der Gruppe 3 hatten die höchsten BCS-Werte, bei 22,6 % war ein Gelbkörper tastbar. Tabelle 26 zeigt die Anzahl der Tiere mit und ohne Gelbkörper am Tag 65 oder 75 p.p. in den BCS-Gruppen. Bei der Aufteilung nach Körperkondition besteht ein signifikanter Zusammenhang zwischen der BCS-Gruppe und dem Vorhandensein eines Gelbkörpers an Untersuchungstag 65 oder 75 p.p. (p=0,016). Dabei haben die Tiere der Gruppe 1 signifikant häufiger einen Gelbkörper am Untersuchungstag als die Kühe der Gruppe 3 (p=0,004). Beim Vergleich der Gruppe 1 mit Gruppe 2 sowie der Gruppen 2 und 3 besteht kein signifikanter Zusammenhang zwischen dem Vorhandensein eines Gelbkörpers und der Zugehörigkeit zu einer der Gruppen (p=0,090, p=0,067). Tab. 26: Kühe (%) mit tastbarem Corpus luteum (Cl) am Tag 65 oder 75 post partum (p.p.) aufgeteilt nach Körperkondition Gruppe Kühe n Cl am Tag 65 oder 75 p.p. (%) 1 Body Condition Score (BCS) ≤ 2,5 2 3 Gesamt BCS 2,75 oder 3,0 BCS ≥ 3,25 41 107 31 178 56,1 %a 40,6 %a,b 22,6 %b 41,0 a,b: Werte mit unterschiedlichen Buchstabenindizes unterscheiden sich statistisch signifikant (p≤0,05). 84 4.5.3 Rastzeit, Güstzeit und Verzögerungszeit in den BCS-Gruppen Von 41 Kühen der BCS-Gruppe 1 mit der niedrigsten Körperkondition wurden 95,1 % besamt. In der Gruppe 2 wurden 94,3 % von 106 Kühen besamt. Alle 31 Probanden der BCS-Gruppe 3 mit der höchsten Körperkondition wurden besamt. In Tabelle 27 sind die Rastzeiten, Güstzeiten und Verzögerungszeiten der einzelnen BCSGruppen angegeben. Die RZ, GZ und VZ unterscheiden sich signifikant zwischen den drei BCS-Gruppen (RZ: p=0,028, GZ: p=0,002, VZ: p=0,020). Zwischen den BCS-Gruppen 1 und 2 gibt es keinen signifikanten Unterschied bei der Länge der Rastzeit (p=0,103), Güstzeit (p=0,289) oder Verzögerungszeit (p=0,556). Die Rastzeiten, Güstzeiten und Verzögerungszeiten der Gruppe 1 sind signifikant länger als die der Gruppe 3 (RZ: p=0,004, GZ: p=0,001, VZ: p=0,010). Bei der Dauer der Rastzeit gibt es keinen signifikanten Unterschied zwischen den BCS-Gruppen 2 und 3 (p=0,150), die Länge der Güstzeit und der Verzögerungszeit ist bei den Kühen der Gruppe 2 aber signifikant länger als bei denen der Gruppe 3 (GZ: p=0,005, VZ: p=0,013). 85 Tab. 27: Mittlere Rastzeit, Güstzeit und Verzögerungszeit mit Standardabweichung (SD) und Median in den BCS-Gruppen Gruppe 1 Body Condition Score (BCS) ≤ 2,5 2 3 BCS 2,75 oder 3,0 BCS ≥ 3,25 41 106 31 178 39 100 31 170 Mittelwert SD Median Güstzeit in Tagen 88,59 a 18,78 87,00 83,11 a, b 17,78 81,00 77,45 b 11,58 76,00 83,34 17,35 82,00 Mittelwert SD Median Verzögerungszeit in Tagen 168,91 a 78,12 146,00 155,64 a 81,46 143,00 109,74 b 48,05 86,00 150,08 77,74 136,50 Mittelwert SD Median 80,18 a 80,40 56,00 72,95 a 81,03 54,50 32,63 b 41,33 19,00 67,05 76,50 46,00 Kühe n besamte Tiere n Rastzeit in Tagen Gesamt a,b: Werte mit unterschiedlichen Buchstabenindizes unterscheiden sich statistisch signifikant (p≤0,05) 86 4.5.4 Mittlere Anzahl der Besamungen in den BCS-Gruppen Zwischen den Tieren in den drei BCS-Gruppen wurde die mittlere Anzahl der Besamungen bis zum Tag 200 nach der Abkalbung sowie die Anzahl der Besamungen während der gesamten beobachteten Laktation vergleichen. Die genauen Werte für die einzelnen Gruppen sind in Tabelle 28 dargestellt. Es besteht kein signifikanter Unterschied bei der mittleren Anzahl der Besamungen bis zum Tag 200 p.p. (p=0,433) und der Anzahl der Besamungen in der gesamten Laktation (p=0,147) zwischen den drei BCS-Gruppen. Tab. 28: Mittlere Anzahl der Besamungen (KB) pro Kuh während der gesamten Laktation und bis Tag 200 post partum mit Standardabweichung (SD), Median, Minimum und Maximum in den BCS-Gruppen Gruppe 1 Body Condition Score (BCS) ≤ 2,5 41 2 3 BCS 2,75 oder 3,0 BCS ≥ 3,25 106 31 178 39 100 31 170 Mittelwert SD Median Minimum Maximum Anzahl KB pro Kuh bis Tag 200 post partum 3,36 2,30 3,00 1 10 3,70 3,32 3,00 1 18 2,65 2,24 2,00 1 9 3,43 2,95 3,00 1 18 Mittelwert SD Median Minimum Maximum 2,69 1,38 2,00 1 6 2,68 1,54 2,00 1 7 2,39 1,61 2,00 1 6 2,63 1,52 2,00 1 7 Kühe n besamte Tiere n Anzahl KB pro Kuh während der gesamten Laktation Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 87 Gesamt 4.5.5 Erstbesamungserfolg in den BCS-Gruppen Von den 39 besamten Tieren der Gruppe 1 mit der niedrigsten Körperkondition waren 17,9 % tragend von der ersten Besamung nach der Abkalbung. In der Gruppe 2 waren 24,0 % von 100 besamten Kühen tragend von der Erstbesamung und in Gruppe 3 mit der höchsten Körperkondition und 31 besamten Tieren lag der Erstbesamungserfolg bei 41,9 %. In Tabelle 29 sind die EBE in den BCS-Gruppen und insgesamt aufgelistet. Es besteht kein signifikanter Zusammenhang zwischen der BCS-Gruppe und dem Erstbesamungserfolg (p=0,060). Tab. 29: Erstbesamungserfolg (%) in den BCS-Gruppen Gruppe Kühe n besamte Tiere n Erstbesamungserfolg (%) 1 Body Condition Score (BCS) ≤ 2,5 2 3 Gesamt BCS 2,75 oder 3,0 BCS ≥ 3,25 41 106 31 178 39 100 31 170 17,9 24,0 41,9 25,9 Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 4.5.6 Gesamtträchtigkeitsraten in den BCS-Gruppen In der Gruppe 1 mit der niedrigsten Körperkondition wurden 39 von 41 Tieren besamt, von 106 Probanden der Gruppe 2 wurden 100 Tiere besamt, und in Gruppe 3 mit den höchsten BCS-Werten wurden alle 31 Tiere besamt. In Tabelle 30 sind die Gesamtträchtigkeitsraten in den BCS-Gruppen aufgeführt. 88 Die GTR bis Tag 200 p.p. und nach allen Besamungen der Laktation unterscheiden sich zwischen den einzelnen BCS-Gruppen nicht signifikant (GTR Tag 200 p.p.: p=0,133, GTR insgesamt: p=0,916). Tab. 30: Gesamtträchtigkeitsrate (GTR) bis Tag 200 post partum (p.p.) und GTR aller Besamungen (KB) der Laktation in den BCS-Gruppen Gruppe Kühe n besamte Tiere n GTR (%) bis Tag 200 p.p. GTR (%) aller KB der Laktation 1 Body Condition Score (BCS) ≤ 2,5 2 3 Gesamt BCS 2,75 oder 3,0 BCS ≥ 3,25 41 106 31 178 39 100 31 170 66,7 65,0 83,9 68,6 84,6 84,0 87,1 84,7 Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 4.5.7 Trächtigkeitsindizes in den BCS-Gruppen Tabelle 31 zeigt die Trächtigkeitsindizes aufgeteilt nach BCS-Gruppen. Zwischen den BCS-Gruppen besteht kein signifikanter Unterschied bei den Trächtigkeitsindizes bis Tag 200 p.p. (p=0,342) und insgesamt (p=0,065). 89 Tab. 31: Mittlere Trächtigkeitsindizes mit Standardabweichung (SD), Minimum und Maximum bis Tag 200 post partum (p.p.) und in der gesamten Laktation in den BCS-Gruppen Gruppe Kühe n besamte Tiere n tragende Tiere bis Tag 200 p.p. n Trächtigkeitsindex (TI) bis Tag 200 p.p. SD Minimum Maximum tragende Tiere aller KB der Laktation n TI der gesamten Laktation SD Minimum Maximum 1 Body Condition Score (BCS) ≤ 2,5 2 3 BCS 2,75 oder 3,0 BCS ≥ 3,25 41 106 31 178 39 100 31 170 26 65 26 117 2,58 1,47 1 6 2,26 1,28 1 5 2,04 1,25 1 5 2,28 1,32 1 6 33 84 27 144 3,42 2,40 1 10 3,43 2,82 1 14 2,22 1,55 1 7 3,20 2,56 1 14 Die Unterschiede zwischen den Gruppen sind nicht signifikant (p>0,05). 90 Gesamt 5 Diskussion In dieser Arbeit wurde untersucht, ob die Behandlung azyklischer Milchkühe mit dem GnRH-Analog Buserelin die Fruchtbarkeit der Tiere im Vergleich zu einer Kontrollgruppe verbessert. Weiterhin wurde überprüft, ob die wiederholte Applikation kleiner Dosen von Buserelin einen besseren Effekt hat als eine einzelne Injektion. 5.1 Gesamtpopulation und Stichprobenumfang Auf neun Milchviehbetrieben in einem Praxisgebiet in Niedersachsen wurden während eines Zeitraumes von zwei Jahren Kühe untersucht und behandelt. Die zur Verfügung stehende Gesamtpopulation bestand aus ca. 4600 Kühen ab der ersten Laktation. Davon wurden 18,8 % zur Untersuchung vorgestellt, weil sie in einem Zeitraum von 50 Tagen nach der Abkalbung noch keine Brunstsymptome gezeigt hatten. Dieses Ergebnis ähnelt dem von MAYER et al. (1987), der von einem Anteil von 18,0 % der Tiere mit unbeobachteter Brunst während der ersten 60 Tage post partum berichtet. Bei den Untersuchungen von DE KRUIF (1977) war bei 16,0 % der untersuchten Tiere bis Tag 50 bis Tag 60 p.p. noch keine Brunst beobachtet worden. Der Autor beschreibt aber eine große Variation von 8,0 % bis 24,0 % anöstrischer Kühe zwischen den untersuchten Herden. Der Anteil von 18,8 % der Tiere mit unbeobachteter Brunst in dieser Studie ist deutlich niedriger als in der Untersuchung von BARTLETT et al. (1987) an 22 amerikanischen Milchviehherden beschrieben, in der bis 70 Tage nach der Abkalbung bei 42,0 % der Tiere keine Brunst beobachtet wurde. Grund für die verschiedenen Ergebnisse ist wahrscheinlich eine unterschiedliche Qualität der Brunstbeobachtung in den einzelnen Herden (MAYER et al. 1987; OPSOMER u. DE KRUIF 1999). Ziel der Dissertation war die Untersuchung der Buserelinwirkung auf die Fruchtbarkeit azyklischer Tiere. Die angesetzten Kriterien für Azyklie waren neben nicht vorhandenen äußeren Brunstsymptomen auch das Fehlen von tastbaren Gelbkörpern auf den Ovarien bei zwei Untersuchungen im Abstand von 10 Tagen. 91 Viele Kühe, die wegen unbeobachteter Brunst vorgestellt wurden, erfüllten die Voraussetzungen für Azyklie im Sinne einer Ovardystrophie nicht und schieden nach der Untersuchung an den Tagen 50 oder 60 p.p. aus. Die Hauptursache für den Ausschluss war das Vorhandensein tastbarer Gelbkörper. Insgesamt schieden 52,1 % der vorgestellten Kühe bei den ersten beiden Untersuchungen aus, weil ein Corpus luteum fühlbar war. Ein ähnliches Ergebnis erzielten MAYER et al. (1987) bei der Untersuchung von Kühen in israelischen Milchviehherden, in denen 58,6 % der Kühe mit unbeobachteter Brunst einen Gelbkörper hatten. Insgesamt erfüllten nur 178 Kühe die aufgestellten Kriterien für Azyklie. Das entspricht einem Anteil von 3,9 % an der Gesamtpopulation, beziehungsweise von 20,5 % der vorgestellten Tiere. Dies ist zum Teil erheblich weniger als die bisher von einigen Autoren beschriebenen Werte. Bei Untersuchungen von BULMAN und LAMMING (1978) hatten 10,4 % der Milchkühe keinen Zyklus am Tag 50 p.p. Die Autoren fanden über den Milchprogesterongehalt heraus, dass 5,2 % zu diesem Zeitpunkt noch keine zyklische Ovartätigkeit hatten und weitere 5,2 % zuvor zyklisch waren, die Eierstocksfunktionen aber wieder eingestellt hatten. STAPLES et al. (1990) fanden über die Bestimmung des Plasmaprogesterongehaltes einen Anteil von 28,0 % azyklischer Kühe am Tag 60 nach der Abkalbung. GÜMEN Plasmaprogesterongehaltes et al. eine (2003) führten ultrasonografische neben der Untersuchung Bestimmung des der auf Ovarien Lutealgewebe durch. Fünfzehn Prozent der mehrkalbigen Kühe und 28,0 % der Einkalbinnen waren in dieser Studie azyklisch. MAYER et al. (1987) fanden einen Anteil von 25,9 % azyklischer Tiere mit inaktiven Ovarien an der Gesamtheit der wegen unbeobachteter Brunst vorgestellten Kühe. Da nicht in allen durchgeführten Studien eine einheitliche Definition für Azyklie verwendet wurde, und die Untersuchungsmethoden und der Untersuchungszeitpunkt teilweise verschieden waren, ist ein Vergleich der Werte nicht immer aussagefähig. Insgesamt liegt der in dieser Dissertation ermittelte Anteil von 3,9 % azyklischer Milchkühe an der Gesamtpopulation unterhalb der in vergleichbaren Studien gefundenen Werte. Die Tiere wurden mit dem Vorbericht „unbeobachtete Brunst in den ersten 50 Tagen nach der Abkalbung“ vorgestellt. Bei den meisten kann von einer zyklischen Ovartätigkeit 92 ausgegangen werden, da bei der rektalen Untersuchung ein Gelbkörper tastbar war. Unklar ist, ob die Brunstsymptome nicht erkannt wurden, oder ob die Kühe zwar einen ovariellen Zyklus hatten, aber äußerlich keine oder wenige Brunstanzeichen zeigten. In einer Untersuchung von SCHOPPER et al. (1993) verliefen 41,2 % der durch Progesteronmessung retrospektiv ermittelten Ovulationen bei Kühen in der postpartalen Phase ohne erkennbare Brunstsymptome (SCHOPPER et al. 1993). Bei über der Hälfte der in der eigenen Studie untersuchten Tiere fand die Brunsterkennung mittels Pedometer statt, das bei den Kühen keine Brunstanzeichen registriert hat. Es ist also davon auszugehen, dass ein Großteil der Tiere mit ovariellem Zyklus keine oder nur schwach ausgeprägte äußerlich sichtbare Brunstsymptome gezeigt hat. 5.2 Ergebnisse in den Versuchsgruppen Die in die Studie aufgenommenen azyklischen Kühe wurden nach dem Zufallsprinzip einer von drei Gruppen zugeordnet. Die Tiere der Kontrollgruppe 50 blieben unbehandelt, die Tiere der Gruppe 55 bekamen eine Injektion mit 20 Mikrogramm Buserelin (5 ml). Den Kühen der Gruppe 51 wurden an fünf aufeinanderfolgenden Tagen jeweils 4 Mikrogramm Buserelin (1 ml) injiziert. Nach der Behandlung mit Buserelin wurden die Kühe an den Tagen 65 und 75 p.p. erneut rektal untersucht. Ein zu diesem Zeitpunkt tastbarer Gelbkörper wurde als Zeichen für eine zyklische Ovartätigkeit gewertet. Bei den Tieren der einmal behandelten Gruppe 55 wurde signifikant häufiger ein Zyklus induziert als bei den Kühen der anderen beiden Gruppen. Die Tiere der Kontrollgruppe hatten zudem häufiger (p≤0,05) einen tastbaren Gelbkörper als die Tiere der fünfmal injizierten Gruppe 51. Die in dieser Studie untersuchte Frage war, ob die Gabe mehrer Einzeldosen von Buserelin die körpereigene pulsatile GnRH-Sekretion nachahmen kann und somit die Ovartätigkeit besser stimuliert als eine einzelne Injektion des Hormons. 93 Im Vergleich zur Einzelinjektion und zur unbehandelten Gruppe hat die Injektion von fünf kleinen Dosen Buserelin bei signifikant weniger azyklischen Kühen zu einer Induktion der Ovartätigkeit geführt. In früheren Versuchen wurde gezeigt, dass schon niedrigere Dosen als die hier verwendeten 4 Mikrogramm Buserelin einen Effekt auf die Gonadotropinsekretion haben (CAVESTANY u. FOOTE 1985; D'OCCHIO et al. 1989). Die adäquate Dosierung für eine ovulationsauslösende LH-Sekretion wurde auf 8 Mikrogramm Buserelin getestet (CAVESTANY u. FOOTE 1985). Eventuell hätte eine Erhöhung der Einzeldosis auf 2 ml (8 Mikrogramm Buserelin) im Vergleich zur Einzeldosis von 1 ml die Gelbkörperbildung bei einer größeren Anzahl von Kühen bewirkt. Ein möglicher Grund für die niedrige Anzahl von Gelbkörpern in der Gruppe 51 könnte sein, dass die zyklische Ovartätigkeit erst mehr als sieben Tage nach der letzten Buserelininjektion an Tag 64 p.p. begonnen hat. Da in den ersten vier Tagen eines Zyklus der Gelbkörper noch nicht palpierbar ist (GRUNERT 1999), besteht die Möglichkeit dass an den Untersuchungstagen 65 und 75 p.p. noch kein Corpus luteum tastbar war, obwohl eine Ovartätigkeit induziert wurde. Dagegen sprechen aber die Ergebnisse von TWAGIRAMUNGU et al. (1994), die bereits drei bis sechs Tage nach einer Injektion von 8 Mikrogramm Buserelin bei azyklischen Kühen Gelbkörper nachweisen konnten. Zudem ist fraglich, ob in der vorliegenden Studie ein Einsetzen der Ovaraktivität über eine Woche nach der letzten Injektion noch auf die Buserelininjektion zurückzuführen ist, oder ob die Eierstockstätigkeit spontan wieder eingesetzt hat. Die mittlere Rastzeit (RZ), Güstzeit (GZ) und Verzögerungszeit (VZ) unterscheiden sich zwischen den Versuchsgruppen nicht signifikant. Die mittlere RZ aller drei Gruppen zusammen von 83,34 Tagen liegt unter dem oberen Richtwert von bis zu 85 Tagen (DE KRUIF et al. 2007). Die mittlere GZ liegt bei allen Tieren zusammen bei 150,08 Tagen und überschreitet den Zielwert von unter 105 Tagen (DE KRUIF et al. 2007) deutlich. Die mittlere GZ in dieser Studie ist auch deutlich länger als bei den azyklischen Kühen, die von BALL und LAMMING (1983) mit einer Einzeldosis GnRH behandelt wurden und die eine GZ von ca. 90 Tagen hatten. Bei der GZ gibt es einen numerischen Unterschied zwischen der Gruppe 55 (141,88 Tage) und den Gruppen 51 (153,96 Tage) sowie 50 (152,63 Tage). Die meisten Tiere haben somit, unabhängig von der Buserelinbehandlung, im Verlauf 94 der Laktation eine Brunst gezeigt, und sind besamt worden. Der klinische Erfolg bei der Aktivierung der Ovartätigkeit durch Injektion von 5 ml Buserelin, der sich in einer signifikant höheren Anzahl an Tieren mit Gelbkörpern nach der Behandlung zeigt, hat sich bei der Betrachtung der Rastzeit wieder relativiert. Diese unterscheidet sich nicht signifkant zwischen den Gruppen. Die Tiere der Gruppe 55 wurden allerdings etwas früher tragend, der Unterschied ist nur numerisch. Der mittlere Erstbesamungserfolg (EBE) aller Tiere zusammen liegt bei 25,9 %. Es gibt keinen signifikanten Unterschied zwischen den Gruppen. Der EBE liegt damit weit unter dem Richtwert von mindestens 50,0 % (DE KRUIF et al. 2007). Der EBE ist auch deutlich schlechter als bei den von BALL und LAMMING (1983) mit einer Einzeldosis GnRH behandelten azyklischen Kühen, die einen EBE von 53,6 % hatten. Insgesamt ist aber zu beachten, dass die Konzeptionsraten auch bei anderen Programmen, zum Beispiel dem Ovsynch-Programm oder Variationen davon, in der Regel nicht beständig über 40,0 % liegen (MACMILLAN 2010), selbst wenn diese nicht nur bei azyklischen Kühen angewendet werden. Der Trächtigkeitsindex (TI) bis Tag 200 p.p. liegt bei allen Tieren zusammen bei 2,28 KB, und damit über dem angestrebten Wert von ≤1,7 KB (DE KRUIF et al. 2007). Der TI nach allen KB der Laktation liegt bei 3,20 KB. Die TI unterscheiden sich zwischen den einzelnen Gruppen nicht signifikant. Die Gesamtträchtigkeitsrate (GTR) bis Tag 200 p.p. liegt bei allen Tieren zusammen bei 68,8 % und unterscheidet sich nicht signifikant zwischen den Gruppen. In der Gruppe 55 ist sie numerisch etwas schlechter als in den Gruppen 51 und 50 (60,0 % vs. 72,5 % und 73,4 %). Die GTR aller KB der Laktation liegt insgesamt bei 84,7 % und damit nur knapp unter dem angestrebten Wert von mindestens 90 % (DE KRUIF et al. 2007). Allerdings wurden einige Tiere über einen langen Zeitraum hinweg immer wieder besamt (Maximum 14 KB). Dieses wiederholte Besamen führt nicht nur zu langen Zwischenkalbezeiten, sondern kann durch lange Laktationsdauer bei abnehmender Milchleistung zu einer Überkonditionierung der Kühe und damit verbundenen postpartalen Störungen führen. Die GTR aller Besamungen der 95 Laktation ist bei den Tieren der Gruppe 55 signifikant niedriger als in den Gruppen 50 und 51 (72,7 % vs. 89,1 % und 92,2 %). Der Erfolg, der bei der Zyklusinduktion durch eine Einzeldosis von 5 ml Buserelin erzielt wurde, und sich in einem höheren Anteil von Tieren mit Gelbkörper am Tag 65 oder 75 p.p. zeigte, setzt sich nicht in einer besseren Fruchtbarkeit im Verlauf der weiteren Laktation fort. Die Tiere der einmal behandelten Gruppe 55 hatten sogar die niedrigsten GTR. Die Kühe der Gruppe 55, die tragend geworden sind, hatten allerdings eine ca. 11 Tage kürzere Güstzeit als die Tiere der anderen beiden Gruppen. Die Injektion mehrerer Einzeldosen bewirkt im Vergleich zur Kontrollgruppe keinen nennenswerten Unterschied bei den Parametern EBE, TI, Anzahl KB und GTR, führt also auch nicht zu besseren Resultaten als keine Behandlung der Azyklie. Insgesamt wiesen die Tiere aller drei Versuchsgruppen keine zufriedenstellenden Ergebnisse bei den untersuchten Fruchtbarkeitskennzahlen auf. Die untersuchten Tiere waren also nicht nur nach der Abkalbung azyklisch, sondern ließen auch im weiteren Verlauf der Laktation schlechte Fruchtbarkeitsergebnisse erkennen. Die Resultate der mit Buserelin behandelten Kühe sind nicht bedeutend besser als die der Kontrollgruppe. Es stellt sich also die Frage, welchen Nutzen eine Behandlung der Azyklie überhaupt hat. MC DOUGALL und COMPTON (2006) fanden heraus, dass eine Behandlung zur Zyklusinduktion bei Kühen mit unbeobachteter Brunst das Risiko verringert, dass die Tiere in der darauffolgenden Laktation erneut keine Brunstsymptome zeigen. Ergebnisse der Studie von LÓPEZ-GATIUS et al. (2008) zeigten, dass die Tiere, die nach der Behandlung von Anöstrie aufgrund von ovarieller Dysfunktion mit einer Brunst reagierten, ähnliche Trächtigkeitsergebnisse hatten, wie Kühe, die eine spontane Brunst zeigten. Die Resultate dieser Studien belegen, dass der Versuch bei azyklischen Kühen einen Zyklus zu induzieren, durchaus sinnvoll sein kann. Im Gegensatz dazu beurteilen OPSOMER und DE KRUIF (1999) die Behandlung der Azyklie bei Milchkühen als wenig erfolgreich. Um eine praxistaugliche Behandlungsstrategie für brunstlose Tiere zu entwickeln ist allerdings zu beachten, dass viele Kühe, die keine Brunstsymptome zeigen gar nicht azyklisch 96 sind, sondern einen ovariellen Zyklus haben und lediglich schwach ausgeprägte oder gar keine Östrusanzeichen aufweisen. Dies verdeutlicht auch die vorliegende Studie, bei der nur ca. 20 % der als brunstlos vorgestellten Kühe wirklich azyklisch waren. Um azyklische und anöstrische Tiere klar unterscheiden zu können, sind mehrere Untersuchungen der Kühe notwendig. Die Alternative zu mehrmaligen Untersuchungen ist ein Programm, mit dem man sowohl die azyklischen als auch die anöstrischen Tiere gleich gut behandeln kann. Bisher wurden als Alternative zu GnRH alleine verschiedene Behandlungsregimes bei Azyklie und Anöstrie untersucht. OZTÜRK et al. (2010), MC DOUGALL (2010) und BARTOLOME et al. (2005) untersuchten das Ovsynch-Programm mit PGF2α und GnRH in unterschiedlichen Variationen. Außerdem wurden Progesteron freisetzende Spiralen, zum Teil in Kombination mit Prostaglandin und Östrogen, getestet (WITTKOWSKI et al. 1982; SEGAWAGWE et al. 2006; WALSH et al. 2007). Eine weitere Behandlungsmöglichkeit ist eCG, auch in Kombination mit Progesteron und Östrogen (BRYAN et al. 2010; GABOR et al. 2002). Alle diese Programme waren mehr oder weniger dazu geeignet, bei brunstlosen Tieren einen Zyklus zu induzieren. Allerdings darf die medikamentöse Therapie der Azyklie kein Ausgleich für Fehler im Management sein, die eine Azyklie möglicherweise erst hervorrufen. Auch eventuelle Primäererkrankungen sollten ebenfalls behandelt werden (OPSOMER u. DE KRUIF 1999). Haltungsbedingte Faktoren, die das Einsetzen der zyklischen Ovartätigkeit beeinflussen können, müssen ebenfalls berücksichtigt werden. Vor allem die Futteraufnahme ist dabei von hoher Bedeutung, um die Körperkondition der Kühe im Verlauf der Laktation zu optimieren (DOMECQ et al. 1997; PRYCE et al. 2001; BUCKLEY et al. 2003; RHODES et al. 2003). 5.3 Aufteilung der Kühe nach Milchleistung Anhand der Ergebnisse der Milchleistungsprüfung der zweiten Milchkontrolle nach der Abkalbung wurden die Tiere in zwei Leistungsgruppen (Gruppe 1: Tagesgemelk der zweiten Kontrolle p.p. > 40 kg, Gruppe 2: Tagesgemelk ≤ 40 kg) eingeteilt. Zwischen den beiden Gruppen gab es bei keinem der untersuchten Parameter (Cl am Tag 65 oder 75 p.p., RZ, GZ, 97 VZ, Anzahl KB bis Tag 200 p.p. und insgesamt, GTR Tag 200 und insgesamt, TI Tag 200 und insgesamt und EBE) signifikante Unterschiede. Die Milchleistung scheint die Fruchtbarkeit azyklischer Milchkühe sowohl mit als auch ohne Behandlung nicht entscheidend zu beeinflussen. Die Tiere beider Leistungsgruppen haben annähernd gleich auf die Behandlung reagiert. Es befanden sich weniger Tiere in der hochleistenden Gruppe als in der Gruppe mit der niedrigeren Leistung (46 in Gruppe 1 vs. 132 in Gruppe 2). Die Ergebnisse lassen also nicht darauf schließen, dass besonders viele hochleistende Kühe azyklisch sind und somit eine hohe Milchleistung die Entstehung von Azyklie begünstigt. Dies deckt sich mit Untersuchungen von AEBERHARD et al. (2001), in denen sich die Fruchtbarkeit von hochleistenden Kühen und Kühen mit durchschnittlicher Milchleistung ähnelte. Auch LEBLANC (2010) stellte keinen klaren Zusammenhang zwischen Milchleistung und Fruchtbarkeit fest. In der Literatur gibt es aber auch andere Ergebnisse über den Einfluss der Milchleistung auf die Fertilität. LÓPEZ-GATIUS et al. (2006) fanden eine positive Korrelation von Leistung und Fruchtbarkeit. Kühe mit einer hohen Milchleistung wurden in dieser Studie früher tragend als Kühe mit niedrigerer Leistung (LÓPEZ-GATIUS et al. 2006). Demgegenüber fanden SCHOPPER et al. (1993) einen signifikant negativen Einfluss der Leistung auf verschiedene Fertilitätsparameter. Sie führen dies vor allem auf reduzierte oder fehlende Brunstsymptome und daraus resultierende schlechtere Fruchtbarkeitskennzahlen bei den hochleistenden Kühen zurück. 5.4 Aufteilung der Tiere nach Laktationsnummer Die Tiere wurden innerhalb der Versuchsgruppen anhand der Anzahl der Abkalbungen in zwei Gruppen aufgeteilt (Gruppe 1: erste Laktation, Gruppe 2: Kühe ab der zweiten Laktation). Es wurden bei allen Parametern die Färsen mit den Kühen insgesamt und innerhalb der Versuchsgruppen verglichen, außerdem jeweils die Färsen und Kühe der einzelnen Gruppen untereinander. Bei den Werten für RZ, GZ und VZ, bei der Anzahl der Besamungen insgesamt, Erstbesamungserfolg und Trächtigkeitsindex gibt es keine signifikanten Unterschiede. Es gibt 98 beim EBE allerdings einen numerischen Unterschied zwischen Färsen und Kühen der unbehandelten Gruppe 50 (31,3 % vs. 18,8 %). Außerdem liegen der EBE der Färsen (28,6 %) und der Kühe (23,3 %) jeweils deutlich unter den angestrebten Werten von ≥ 50 % (DE KRUIF et al. 2007). Unabhängig von der Versuchsgruppe hatten die Färsen signifikant bessere GTR als die Mehrkalbskühe. Beim Vergleich der Färsen und der Kühe innerhalb der einzelnen Gruppen waren die GTR der Färsen nur in der Gruppe 51 (fünf mal 1 ml Buserelin) signifikant höher als die GTR der Kühe. In der unbehandelten Gruppe 50 wurden die Kühe bis zum Tag 200 p.p. signifikant häufiger besamt als die Färsen. Weiterhin wurden jeweils die Färsen und die Kühe der einzelnen Gruppen untereinander verglichen. Dabei hatten sowohl die Färsen als auch die Kühe der Gruppe 51 signifikant seltener einen Gelbkörper am Tag 65 oder 75 p.p. als die Färsen der Gruppen 55 und 50. Die Färsen der Gruppe 50 hatten zudem signifikant seltener ein Cl als die Färsen der Gruppe 55. Die GTR aller Besamungen der Laktation ist bei den Färsen der Gruppe 51 höher als bei den Färsen der Gruppe 55 (p≤0,05). Insgesamt sind die Ergebnisse der Färsen etwas besser als die der Kühe. Das stimmt mit unterschiedlichen Studien überein, in denen die Fruchtbarkeit von Einkalbigen besser als die von mehrkalbigen Kühen ist (DOMECQ et al. 1997; GARCÍA-ISPIERTO et al. 2007; GALON et al. 2010; INCHAISRI et al. 2010). Möglicherweise haben also die Färsen keine besseren Ergebnisse als die Kühe, weil sie besser auf die Behandlung angesprochen haben, sondern weil sie insgesamt eine bessere Fruchtbarkeit haben als die mehrkalbigen Tiere. 5.5 Aufteilung nach Körperkondition Die untersuchten Tiere wurden anhand ihrer Körperkondition am Tag 60 p.p. in drei BCS Gruppen eingeteilt (Gruppe 1: BCS ≤ 2,5, Gruppe 2: BCS 2,75 oder 3,0, Gruppe 3: BCS ≥ 3,25). Die Kühe der Gruppe 1 mit der schlechtesten Körperkondition hatten signifikant häufiger einen tastbaren Gelbkörper am Tag 65 oder 75 p.p. als die Tiere der Gruppe 3. Die RZ, GZ und VZ der Tiere in Gruppe 1 ist allerdings signifikant höher als die der Kühe in Gruppe 3. Dieses Ergebnis stimmt nicht mit den Untersuchungen von SAKAGUCHI (2009) 99 und WALTNER et al. (1993) überein, die keinen Zusammenhang zwischen BCS und RZ oder GZ fanden. Die mageren Tiere haben häufiger einen Gelbkörper, werden also früher zyklisch als die dickeren Kühe, im weiteren Verlauf der Laktation werden sie aber später tragend als die besser konditionierten Tiere. Ähnliche Ergebnisse hatten PRYCE et al. (2001), GOSSEN et al. (2006) und VAN STRATEN et al. (2009), die bei Tieren mit niedriger Kondition eine längere RZ, GZ und VZ und schlechtere Trächtigkeitsergebnisse im Vergleich zu besser konditionierten Kühen beobachteten. Die Tiere mit der höchsten Körperkondition haben in der vorliegenden Studie zwar seltener einen Gelbkörper, werden also später zyklisch als die mageren Tiere, haben aber eine RZ (ca. 77 Tage), GZ (ca. 110 Tage) und VZ (ca. 33 Tage), die den angestrebten Richtwerten sehr nahe kommt. Auch die Tiere der mittleren BCS Gruppe haben eine signifikant längere GZ und VZ als die Tiere der Gruppe 3. Die mittlere Anzahl der Besamungen, EBE, GTR und TI unterscheiden sich nicht signifikant zwischen den drei Gruppen. Die dickeren Kühe werden insgesamt früher tragend; die GTR bis Tag 200 p.p. ist bei den Tieren der Gruppe 3 allerdings nur numerisch besser als in den Gruppen 1 und 3 (83,9 % vs. 65,0 % und 66,7 %). Auch beim EBE gibt es numerische Unterschiede zwischen den drei Gruppen (Gr.1: 17,9 %, Gr.2: 24,0 %, Gr.3: 41,9%). PRYCE et al. (2001) fanden bei Kühen mit unterdurchschnittlicher Körperkondition einen schlechteren EBE. Im Gegensatz dazu beschreiben BUCKLEY et al. (2003) einen besseren EBE bei Tieren mit niedrigem Gewicht, und HEUER et al. (1999) einen schlechteren EBE bei Kühen mit hohem BCS. Insgesamt zeigen in dieser Studie die Tiere der Gruppe mit der höchsten Körperkondition im Vergleich mit den dünneren Tieren etwas bessere Fruchtbarkeitsparameter. In dieser Gruppe waren Tiere mit einem BCS von mindestens 3,25 am Tag 60 p.p. Ein BCS von 3,25 liegt bei Kühen in der frühen Laktation noch im Normbereich (DE KRUIF et al. 2007), es sind also nicht ausschließlich fette Tiere in dieser Gruppe gewesen. Bei einer Einteilung nur mit Tieren mit deutlich überhöhtem BCS (z. B. BCS an Tag 60 p.p. ≥ 4,0) in einer Gruppe, wäre der in oben genannten Studien beschriebene Einfluss einer starken Überkonditionierung auf die Fruchtbarkeit möglicherweise deutlicher gewesen. 100 5.6 Schlussfolgerung Die wiederholte Applikation kleiner Dosen von Buserelin hat im Vergleich zur einmaligen Injektion keinen signifikanten Einfluss auf die untersuchten Parameter. Insgesamt war die Fruchtbarkeit der mit Buserelin behandelten Tiere nicht signifikant besser als die der Kontrollgruppe. Um zu überprüfen, ob die azyklischen Tiere allgemein schlecht auf eine medikamentöse Zyklusinduktion ansprechen, oder ob die Verwendung von Buserelin als einzigem Wirkstoff nicht effektiv ist, wären weitere Untersuchungen zum Vergleich verschiedener Therapiemöglichkeiten (z. B. Ovsynch, Progesteron etc.) sinnvoll. Die Fertilität der azyklischen Tiere ist insgesamt herabgesetzt, die Fruchtbarkeitskennzahlen sind bei den Färsen etwas besser als bei den Kühen, erreichen aber nicht die angestrebten Werte für eine zufriedenstellende Fruchtbarkeit. Die an der Studie beteiligten Betriebe sind insgesamt gut geführt, die Fruchtbarkeit der Herden liegt im regionalen Durchschnitt. Die unbefriedigenden Fruchtbarkeitskennzahlen der untersuchten und behandelten Tiere entsprechen somit nicht dem Herdenniveau der Betriebe, sondern scheinen die Folge einer individuell niedrigen Fertilität der azyklischen Kühe zu sein. 101 6 Zusammenfassung Hanna Bläsing: Einfluss einer wiederholten Applikation von Gonadotropin-Releasing Hormon (GnRH) auf die Fruchtbarkeit azyklischer Milchkühe. In dieser Arbeit wurde untersucht, ob die Behandlung azyklischer Milchkühe mit dem GnRH-Analog Buserelin die Fruchtbarkeit im Vergleich zu einer Kontrollgruppe verbessert. Weiterhin wurde überprüft, ob die wiederholte Applikation kleiner Dosen von Buserelin wirkungsvoller ist als eine einzelne Injektion. Für diese Feldstudie wurden auf neun Milchviehbetrieben in Niedersachsen 868 Kühe untersucht, bei denen in einem Zeitraum von 50 Tagen nach der Abkalbung noch keine Brunstsymptome festgestellt wurden. Wenn bei zwei rektalen Palpationen im Abstand von zehn Tagen keine Gelbkörper fühlbar waren, wurden die Tiere einer von drei Versuchsgruppen zugeordnet. Den Tieren der Gruppe 51 wurde an fünf aufeinanderfolgenden Tagen je 4 Mikrogramm (1 ml) Buserelin injiziert, die Tiere der Gruppe 55 bekamen eine Injektion von 20 Mikrogramm (5 ml) Buserelin und die Kühe der Kontrollgruppe 50 blieben unbehandelt. Insgesamt erfüllten 178 Tiere die Kriterien für Azyklie und wurden in die Studie einbezogen. Dies entspricht einem Anteil von 3,9 % an der Gesamtpopulation (n=4612) und von 20,5 % der als anöstrisch vorgestellten Kühe. Nach der Behandlung wurden die Kühe erneut untersucht (Tag 65 und 75 post partum); ein fühlbarer Gelbkörper wurde dabei als Zeichen für ein Wiedereinsetzen der Ovartätigkeit gewertet. Außerdem wurde eine Trächtigkeitsuntersuchung nach der Besamung durchgeführt und die Fruchtbarkeitskennzahlen Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ), Erstbesamungserfolg (EBE), Anzahl der Besamungen, Gesamtträchtigkeitsrate (GTR) und Trächtigkeitsindex (TI) erfasst. 102 Nach der Behandlung mit fünf mal 1 ml Buserelin hatten die Tiere der Gruppe 51 signifikant seltener einen Gelbkörper als die Tiere der anderen beiden Gruppen (14,5 % vs. 43,9 % Gruppe 50 und 63,2 % Gruppe 55). Die RZ, GZ und VZ, die mittlere Anzahl an Besamungen, der EBE und der TI unterschieden sich dagegen nicht signifikant. Die GTR aller Besamungen der Laktation war in der einmal behandelten Gruppe 55 signifikant niedriger als in den anderen beiden Gruppen (72,7 % vs. 89,1 % Gruppe 50 und 92,2 % Gruppe 51). Die Tiere, die mit fünf kleinen Dosen Buserelin behandelt wurden, haben zwar klinisch weniger auf die Behandlung reagiert, da sie signifikant seltener einen Gelbkörper hatten, später zeigten sich aber kaum Unterschiede bei den Fruchtbarkeitsparametern zwischen den einzelnen Gruppen. Die Tiere wurden außerdem nach Laktationsnummer in Gruppen aufgeteilt. Dabei wurden Färsen und Kühe ab der zweiten Laktation jeweils innerhalb der Versuchsgruppen und zwischen den Gruppen verglichen. Es gab keine signifikanten Unterschiede bei den Parametern RZ, GZ, VZ, Anzahl der Besamungen in der gesamten Laktation, EBE und TI. Sowohl die Färsen als auch die Kühe der fünfmal mit Buserelin behandelten Gruppe hatten jeweils signifkant seltener einen Gelbkörper als die Tiere der anderen beiden Gruppen. Die Kühe der Kontrollgruppe wurden bis zum Tag 200 p.p. signifikant häufiger besamt als die Färsen. Die GTR der Färsen aller Gruppen zusammen war signifikant höher als die GTR der Kühe. Die GTR aller Besamungen der Laktation war bei den Färsen der Gruppe 55 signifikant niedriger als bei den Färsen der Gruppe 51. Insgesamt war die Fruchtbarkeit der Färsen etwas besser als die der Kühe, die Unterschiede waren bei einigen Parametern aber nur numerisch. Bei der Aufteilung der Tiere nach der Milchleistung der zweiten Kontrolle nach der Abkalbung (Gruppe 1: > 40 kg Tagesgemelk, Gruppe 2: ≤ 40 kg Tagesgemelk) gab es bei keinem untersuchten Parameter signifikante Unterschiede zwischen den Kühen mit hoher und niedriger Leistung. Anhand der Körperkondition an Tag 60 nach der Abkalbung wurden die Kühe in drei Gruppen unterteilt (Gruppe 1: BCS ≤ 2,5, Gruppe 2: BCS 2,75 oder 3,0, Gruppe 3: BCS ≥ 103 3,25). Die Tiere mit den niedrigsten BCS-Werten hatten signifikant häufiger einen tastbaren Gelbkörper nach der Behandlung als die Tiere der Gruppe mit der höchsten Körperkondition. Allerdings war die RZ, GZ und VZ der Kühe mit den höchsten BCS-Werten signifikant geringer als in den anderen beiden Gruppen. Die dickeren Kühe waren zwar später zyklisch als die dünneren Tiere, wurden aber im Verlauf der Laktation früher tragend. Die Fertilität aller untersuchten azyklischen Kühe war insgesamt herabgesetzt, die meisten Fruchtbarkeitskennzahlen waren bei den Färsen numerisch oder signfikant besser als bei den Kühen, erreichten aber nicht die angestrebten Werte für eine zufriedenstellende Fruchtbarkeit. Die wiederholte Applikation kleiner Dosen von Buserelin hatte im Vergleich zur einmaligen Injektion keinen höheren Einfluss auf die untersuchten Parameter. Außerdem war die Fruchtbarkeit der mit Buserelin behandelten Tiere nicht signifikant besser als die der Kontrollgruppe. 104 7 Summary Hanna Bläsing: Effect of a repeated application of gonadotropin-releasing hormone (GnRH) on reproductive performance in acyclic dairy cows. It was the objective of this study to find out, whether or not the treatment with the GnRHanalogue Buserelin could improve the fertility of acyclic dairy cows in comparison with an untreated control group. Furthermore, it was tested, if the repeated application of small doses of Buserelin had a better effect than a single injection. For this field trial, 868 cows from nine dairy herds in Lower Saxony were examined. They had not been detected in estrus until 50 days postpartum. When there was no palpable corpus luteum at two manual transrectal examinations of the ovaries in a ten-day interval, cows were assigned to one of three groups. The cows in group 51 were injected with 4 µg (1 ml) Buserelin once daily for five days. The animals in group 55 received a single injection of 20 µg (5 ml) Buserelin and the cows in group 50 (control group) remained untreated. 178 cows were defined as acyclic and therefore, were included in the study. This is a percentage of 3.9 % of the population (n=4612) or 20.5 % of the 868 cows presented as anestrous. Cows were examined by rectal palpation five and ten days after the treatment (Days 65 and 75 postpartum). A palpable corpus luteum on either day was considered as a sign for the resumption of ovarian activity. Cows were subjected to the farm’s usual breeding program. Pregnancy diagnosis was performed after service and the measures of fertility were determined (interval from calving to first insemination (CFI), days open (DO), interval from first insemination to conception (FIC), number of services, first service conception rate (FSC), pregnancy rate (PR) and pregnancy index (PI)). The frequency of cows with a palpable corpus luteum was statistically significantly lower in group 51 (five injections of 1 ml Buserelin) than in the other groups (14.5 % vs. 43.9 % group 105 50 and 63.2 % group 55). There was no significant difference between the groups in the fertility measures CFI, DO, FIC, average number of services, FSC and PI. Pregnancy rate from all services during lactation was statistically significantly lower in cows receiving a single injection of 5 ml Buserelin (group 55) than in the other groups (72.7 % vs. 89.1 % group 50 and 92.2 % group 51). Even though the ovarian reactions to the treatment with five small doses of Buserelin seemed to be worse than in the other groups because of a lower percentage of a detectable corpus luteum, later on, there were hardly any differences in the measures of fertility between the groups. The animals were further assigned to groups according to their number of lactation (group 1: heifers, group 2: number of lactation ≥ 2). Cows and heifers were compared within and between the different treatment groups. The measures of fertility CFI, DO, FIC, number of services during the lactation, FSC and PI did not differ statistically significantly between the groups. Both, heifers and cows that received five injections of Buserelin (group 51) had significantly less often a palpable corpus luteum after treatment than animals in the other groups. Within the untreated control group, cows had significantly more services than heifers until day 200 postpartum. The pregnancy rate of heifers was significantly higher than that of the cows. The pregnancy rate of all services of heifers in group 55 (single injection of Buserelin) was significantly lower than that of the heifers which received five doses of Buserelin (group 51). Overall, fertility measures of heifers were slightly better than those of cows. The animals were also grouped according to their milk yield of the second test-day postpartum (group 1: > 40 kg milk per day, group 2: ≤ 40 kg milk per day). There was no significant difference in any of the analyzed parameters. In addition, the cows were also split into groups according to their body condition on Day 60 postpartum (group 1: body condition score BCS ≤ 2.5, group 2: BCS 2.75 or 3.0, group 3: BCS ≥ 3.25). The animals with the lowest body condition scores had significantly more often a palpable corpus luteum following treatment than the cows with the highest scores. However, the CFI, DO and FIC of the animals in group 3 (highest BCS) were significantly shorter than 106 in the other groups. The ovarian cycles of the well conditioned cows resumed later, but these cows became pregnant earlier than the cows in thinner condition. In conclusion, the fertility measures of all acyclic cows examined in this trial were far below the desired reference values with heifers yielding slightly better results than cows. Repeated injections of small doses of Buserelin did not have a significant effect on reproductive performance compared with a single injection. Fertility of acyclic cows that had been treated with Buserelin was not statistically significantly higher than that of acyclic cows without treatment. 107 8 Literaturverzeichnis AEBERHARD, K., R. M. BRUCKMAIER, U. KUEPFER u. J. W. BLUM (2001): Milk yield and composition, nutrition, body conformation traits, body condition scores, fertility and dieseases in high-yielding dairy cows - part 1. J. Vet. Med. A. Physiol. Pathol. Clin. Med. 48, 97 - 110 ALANKO, M. u. S. PYÖRÄLÄ (1980): The treatment of anoestrus and suboestrus in dairy cattle using a progesterone releasing intravaginal device (PRID) or gonadotrophins. Nord. Veterinaermed. 32, 444 – 452 ALILA, H. W., R. A. CORRADINO u. W. HANSEL (1988 a): A comparison of the effects of cyclooxygenase prostanoids on progesterone production by small and large bovine luteal cells. Prostaglandins 36, 259 – 270 ALILA, H. W., J. P. DOWD, R. A. CORRADINO, W. V. HARRIS u. W. HANSEL (1988 b): Control of progesterone production in small and large luteal cells separated by flow cytometry. J. Reprod. Fertil. 82, 645 – 655 ASPDEN, W. J., A. JACKSON, T. E. TRIGG u. M. J. D'OCCHIO (2003): Pituitary expression of LHbeta- and FSHbeta-subunit mRNA, cellular distribution of LHbetasubunit mRNA and LH and FSH synthesis during and after treatment with a gonadotropinreleasing hormone agonist in heifers. Reprod. Fertil. Dev. 15, 149 - 156 108 AURICH, J. E., D. AHLERS u. P. ANDRESEN (1996): Fruchtbarkeitsstörungen. In: GRUNERT, E. (1996): Buiatrik. 5. Aufl. Bd. 1. Euterkrankheiten, Geburtshilfe und Gynäkologie, Andrologie und Besamung. Verlag M. & H. Schaper Alfeld - Hannover, 217 - 241 BALL, P. J. H. u. G. E. LAMMING (1983): Diagnosis of ovarian acyclicity in lactating dairy cows and evaluation of treatment with gonadotrophin-releasing hormone or a progesterone-releasing intravaginal device. Br. Vet. J. 139, 522 – 527 BARATTA, M., L. A. WEST, A. M. TURZILLO u. T. M. NETT (2001): Activin modulates differential effects of estradiol on synthesis and secretion of folliclestimulating hormone in ovine pituitary cells. Biol. Reprod. 64, 714 - 719 BARTLETT, P. C., J. KIRK, P. COE, J. MARTENIUK u. E. C. MATHER (1987): Descriptive epidemiology of anestrus in Michigan Holstein-Friesian cattle. Theriogenology 27, 459 – 476 BARTOLOME, J. A., A. SOZZI, J. MCHALE, P. MELENDEZ, A. C. ARTECHE, F. T. SILVESTRE, D. KELBERT, K. SWIFT, L. F. ARCHBALD u. W. W. THATCHER (2005): Resynchronisation of ovulation and timed insemination in lactating dairy cows, II: assigning protocols according to stages of the estrous cycle or presence of ovarian cysts or anestrus. Theriogenology 63, 1628 - 1642 109 BEAUVILLAIN, J. C. u. V. PRÉVOT (2004): Des acteurs clés de la régulation de la sécrétion de GnRH: les cellules gliales et endothéliales de l'hypothalamus. Hypothalamic glial cells and endothelial cells as key regulators of GnRH secretion. J. Soc. Biol. 198, 68 - 72 BÉDÉCARRATS, G. u. U. B. KAISER (2003): Differential regulation of gonadotropin subunit gene promotor activity by pulsatile gonadotropin-releasing hormone (GnRH) in perifused LβT2 cells: role of GnRH receptor concentration. Endocrinology 144, 1802 - 1811 BISHOP, D. K. u. R. P. WETTEMANN (1993): Pulsatile infusion of gonadotropin-releasing hormone initiates luteal activity in nutritionally anestrous beef cows. J. Anim. Sci. 71, 2714 – 2720 BLEACH, E. C. L., R. C. GLENNCROSS u. P. G. KNIGHT (2004): Association between ovarian follicle development and pregnancy rates in dairy cows undergoing spontaneous oestrous cycles. Reproduction 127, 621 – 629 BOSTEDT, H., L. E. KOZICKI, K. H. FINGER u. H. KARG (1985): Untersuchungen über den Einfluss verschiedener Haltungsbedingungen auf postpartale Regenerationsvorgänge am Genitaltrakt von Milchkühen unter besonderer Berücksichtigung der Progesteronprofile. Zuchthygiene 20, 17 – 33 BOUSQUET, D., P. LAMOTHE u. P. GUAY (1976): Profil enzymatique des sécrétions utérines de la vache au cours du cycle oestral. Can. J. Comp. Med. Vet. Sci. 40, 198 – 201 110 BRYAN, M. A., G. A. BÓ, C. HEUER u. F. R. EMSLIE (2010): Use of equine chorionic gonadotrophin in synchronised AI of seasonal-breeding, pasturebased, anoestrous dairy cattle. Reprod. Fertil. Dev. 22, 126 - 131 BUCKLEY, F., K. O'SULLIVAN, J. F. MEE, R. D. EVANS u. P. DILLON (2003): Relationships among milk yield, body condition, cow weight, and reproduction in springcalved Holstein-Friesians. J. Dairy Sci. 86, 2308 - 2319 BULMAN, D. C. u. G. E. LAMMING (1978): Milk progesterone levels in relation to conception, repeat breeding and factors influencing acyclicity in dairy cows. J. Reprod. Fertil. 54, 447 – 458 CAVESTANY, D. u. R. H. FOOTE (1985): Reproductive performance of Holstein cows administered GnRH analog Hoe 766 (Buserelin) 26 to 34 days postpartum. J. Anim. Sci. 61, 224 - 233 CHENAULT, J. R., W. W. THATCHER, P. S. KALRA, R. M. ABRAMS u. C. J. WILCOX (1974): Transitory changes in plasma progestins, estradiol, and luteinizing hormone approaching ovulation in the bovine. J. Dairy Sci. 58, 709 - 717 CHOI, H. S., W. STÖCKL, K. ARBEITER, F. RITTMANSPERGER u. E. BAMBERG (1977): Der Plasmaprogesterongehalt bei Kühen in der Puerperalphase. Wien. tierärztl. Monatsschr. 64, 10 – 11 111 CHRISTENSEN, D. S., M. L. HOPWOOD u. J. N. WILTBANK (1974): Levels of hormones in the serum of cycling beef cows. J. Anim. Sci. 38, 577 – 583 CLARKE, I. J. u. S. POMPOLO (2005): Synthesis and secretion of GnRH. Anim. Reprod. Sci. 88, 29 – 55 CONN, P. M., A. J. W. HSUEH u. W. F. CROWLEY JR. (1984): Gonadotropin-releasing hormone: molecular and cell biology, physiology, and clinical applications. Fed. Proc. 43, 2351 – 2361 COSOWSKY, L., S. N. RAO, G. J. MACDONALD, H. PAPKOFF u. W. R. MOYLE (1995): The groove between the alpha- and beta-subunits of hormones with lutropin (LH) activity appears to contact the LH receptor, and its confirmation is changed during hormone binding. J. Biol. Chem. 270, 20011 - 20019 COUNIS, R., J. N. LAVERRIÈRE, G. GARREL, C. BLEUX, J. COHEN-TANNOUDJI, Y. LERRANT, M. L. KOTTLER u. S. MAGRE (2005): Gonadotropin-releasing hormone and the control of gonadotrope function. Reprod. Nutr. Dev. 45, 243 – 254 CUPP, A. S., F. N. KOJIMA, M. S. ROBERTSON, M. W. WOLFE, L. A. WERTH, R. J. KITTOK, H. E. GROTJAN u. J. E. KINDER (1995): Increasing concentrations of 17β-estradiol has differential effects on secretion of luteinizing hormone and follicle-stimulating hormone and amounts of mRNA for gonadotropin subunits during the follicular phase of the bovine estrous cycle. Biol. Reprod. 52, 288 - 296 112 DARWASH, A. O., G. E. LAMMING u. J. A. WOOLIAMS (1997): Estimation of genetic variation in the interval from calving to postpartum ovulation in dairy cows. J. Dairy Sci. 80, 1227 – 1234 DE KRUIF, A. (1977): Een onderzoek van runderen in anoestrus. Studies on cattle in anoestrus. Tijdschr. Diergeneeskd. 15, 247 – 253 DE KRUIF, A., R. MANSFELD u. M. HOEDEMAKER (2007): Tierärztliche Bestandsbetreuung beim Milchrind. 2. Aufl. Enke Verlag, Stuttgart D'OCCHIO, M. J., D. R. GIFFORD, C. R. EARL, T. WEATHERLY u. W. v. RECHENBERG (1989): Pituitary and ovarian responses of post-partum acyclic beef cows to continuous long-term GnRH and GnRH agonist treatment. J. Reprod. Fertil. 85, 495 – 502 DOMECQ, J. J., A. L. SKIDMORE, J. W. LLOYD u. J. B. KANEENE (1997): Relationship between body condition scores and conception at first artificial insemination in a large dairy herd of high yielding Holstein cows. J. Dairy Sci. 80, 113 - 120 DUBOIS, E. A., M. A. ZANDBERGEN, J. PEUTE u. H. J. GOOS (2002): Evolutionary development of three gonadotropin-releasing hormone (GnRH) systems in vertebrates. Brain Res. Bull. 57, 413 – 418 113 EVANS, A. C. O., C. M. KOMAR, S. A. WANDJI u. J. E. FORTUNE (1997): Changes in androgen secretion and luteinizing hormone pulse amplitude are associated with the recruitment and growth of ovarian follicles during the luteal phase of the bovine estrous cycle. Biol. Reprod. 57, 394 - 401 EDMONSON, A. J., I. J. LEAN, L. D. WEAVER, T. FARVER u. G. WEBSTER (1989): A body condition scoring chart for Holstein dairy cows. J. Dairy Sci. 72, 68 - 78 FERNANDES, L. C., W. W. THATCHER, C. J. WILCOX u. E. P. CALL (1978): LH release in response to GnRH during the postpartum period of dairy cows. J. Anim. Sci. 46, 443 – 448 FIELD, A. (2009) Discovering statistics using SPSS. 3. Aufl. Sage Publications Inc., Los Angeles, London FINK, G. (1988): Oestrogen and progesterone interactions in the control of gonadotrophin and prolactin secretion. J. Steroid Biochem. 30, 169 - 178 GABOR, G., J. P. KASTELIC, S. PINTAS, F. SZASZ, E. SZIGETI u. N. SOLYMOSI (2002): Improving reproductive performance in lactating dairy cows by synchronizing ovulation or inductiong oestrus. Acta Vet. Hung. 50, 231 – 234 114 GALON, N., Y. ZERON u. E. EZRA (2010): Factors affecting fertility of dairy cows in Israel. J. Reprod. Dev. 56, 8 - 14 GARCIA, M. u. K. LARSSON (1982): Clinical findings in post partum dairy cows. Nord. Veterinaermed. 34, 255 – 263 GARCÍA - ISPIERTO, I., F. LÓPEZ - GATIUS, P. SANTOLARIA, J. L. YÁNIZ, C. NOGAREDA u. M. LÓPEZ - BÉJAR (2007): Factors affecting the fertility of high producing dairy herds in northeastern Spain. Theriogenology 67, 632 - 638 GARVERICK, H. A. u. M. F. SMITH (1993): Female reproductive physiology and endocrinology of cattle. Vet. Clin. North Am. Food Anim. Pract. 9, 223 – 247 GEISERT, R. D., G. L. MORGAN, E. C. SHORT u. M. T. ZAVY (1992): Endocrine events associated with endometrial function and conceptus development in cattle. Reprod. Fertil. Dev. 4, 301 – 305 GOEHRING, C., C. AURICH, J. LANGE, H.-O. HOPPEN u. J. E. AURICH (1999): Therapeutische und prophylaktische Beeinflussbarkeit der LH-Freisetzung beim Rind. Tierärztl. Prax. 27 (G), 25 – 29 GOSSEN, N., S. FIETZE, S. MÖSENFECHTEL u. M. HOEDEMAKER (2006): Beziehungen zwischen Körperkondition (Rückenfettdicke und Body Condition Scoring) und Fruchtbarkeit bei Milchkühen (Deutsche Holstein). Dtsch. Tierärztl. Wochenschr. 113, 171 - 172, 174 - 177 115 GRUNERT, E. (1999): Brunsterkennung. In: GRUNERT, E. u. A. DE KRUIF (Hrsg.): Fertilitätsstörungen beim weiblichen Rind. 3. Aufl. Parey Buchverlag im Blackwell Wissenschafts-Verlag, Berlin, Wien, 68 - 77 GRUNERT, E. (1999): Die gynäkologische Untersuchung. In: GRUNERT, E. u. A. DE KRUIF (Hrsg.): Fertilitätsstörungen beim weiblichen Rind. 3. Aufl. Parey Buchverlag im Blackwell Wissenschafts-Verlag, Berlin, Wien, 29 - 62 GRUNERT, E. (1999): Ovarielle Dysfunktionen. In: GRUNERT, E. u. A. DE KRUIF (Hrsg.): Fertilitätsstörungen beim weiblichen Rind. 3. Aufl. Parey Buchverlag im Blackwell Wissenschafts-Verlag, Berlin, Wien, 111 - 146 GRUNERT, E. (1999): Sexualzyklus. In: GRUNERT, E. u. A. DE KRUIF (Hrsg.): Fertilitätsstörungen beim weiblichen Rind. 3. Aufl. Parey Buchverlag im Blackwell Wissenschafts-Verlag, Berlin, Wien, 3 - 11 GÜMEN, A., J. N. GUENTHER u. M. C. WILTBANK (2003): Follicular size and response to Ovsynch versus detection of estrus in anovular and ovular lactationg dairy cows. J. Dairy Sci. 10, 3184 – 3194 116 GÜMEN, A., R. R. RASTANI, R. R. GRUMMER u. M. C. WILTBANK (2005): Reduced dry periods and varying prepartum diets alter postpartum ovulation and reproductive measures. J. Dairy Sci. 88, 2401 – 2411 HANSEL, W. u. R. B. SNOOK (1970): Pituitary ovarian relationships in the cow. J. Dairy Sci. 53, 945 - 961 HANSEL, W. u. E. M. CONVEY (1983): Physiology of the estrous cycle. J. Anim. Sci. 57, 404 – 424 HENDERSON, K. M. u. Y. S. MOON (1979): Luteinization of bovine granulosa cells and corpus luteum formation associated with loss of androgen-aromatizing ability. J. Reprod. Fert. 56, 89 – 97 HEUER, C., Y. H. SCHUKKEN u. P. DOBBELAAR (1999): Postpartum body condition score and results from the first test day milk as predictors of disease, fertility, yield, and culling in commercial dairy herds. J. Dairy Sci. 82, 295 - 304 HUMKE, R. u. H. ZUBER (1977): Die Behandlung von Anoestrie und Azyklie des Rindes durch ein LH-RH-Analog. Berl. Münch. tierärztl. Wochenschr. 90, 234 – 237 HUMPHREY, W. D., C. C. KALTENBACH, T. G. DUNN, D. R. KORITNIK u. G. D. NISWENDER (1983): Characterization of hormonal patterns in the beef cow during postpartum anestrus. J. Anim. Sci. 56, 445 – 453 117 INCHAISRI, C., H. HOGEVEEN, P. L. VOS, G. C. VAN DER WEIJDEN u. R. JORRITSMA (2010): Effect of milk yield characteristics, breed, and parity on success of the first insemination in Dutch dairy cows. J. Dairy Sci. 93, 5179 - 5187 JAGGER, J. P., A. R. PETERS u. G. E. LAMMING (1987): Hormone responses to low-dose GnRH treatment in post-partum beef-cows. J. Reprod. Fertil. 80, 263 – 269 JOLLY, P. D., S. MC DOUGALL, L. A. FITZPATRICK, K. L. MACMILLAN u. K. W. ENTWISTLE (1995): Physiological effects of undernutrition on postpartum anoestrus in cows. J. Reprod. Fertil. Suppl. 49, 477 – 492 JUBB, T. F., P. BRIGHTLING, J. MALMO, M. T. LARCOMBE, G. A. ANDERSON u. S. J. HIDES (1989): Evaluation of a regimen using a progesterone releasing intravaginal device (CIDR) and PMSG as a treatment for post partum anoestrus in dairy cattle. Aust. Vet. J. 66, 334 – 336 KAISER, U. B., A. JAKUBOWIAK, A. STEINBERGER u. W. W. CHIN (1997): Differential effects of gonadotropin-releasing hormone (GnRH) pulse frequency on gonadotropin subunit and GnRH receptor messenger ribonucleic acid levels in vitro. Endocrinology 138, 1224 - 1231 KALTENBACH, C. C., T. G. DUNN, T. E. KISER, L. R. CORAH, A. M. AKBAR u. G. D. NISWENDER (1974): Release of FSH and LH in beef heifers by synthetic gonadotrophin releasing hormone. J. Anim. Sci. 38, 357 - 362 118 KESNER, J. S. u. E. M. CONVEY (1982): Interaction of estradiol and luteinizing hormone releasing hormone on follicle stimulating hormone release in cattle. J. Anim. Sci. 54, 817 - 821 KESNER, J. S., V. PADMANABHAN u. E. M. CONVEY (1982): Estradiol induces and progesterone inhibits the preovulatory surges of luteinizing hormone and follicle-stimulating hormone in heifers. Biol. Reprod. 26, 571 - 571 KING, G. J., J. F. HURNIK u. H. A. ROBERTSON (1976): Ovarian function and estrus in dairy cows during early lactation. J. Anim. Sci. 42, 688 – 692 KOJIMA, F. N., A. S. CUPP, T. T. STUMPF, D. D. ZALESKY, M. S. ROBERSON, L. A. WERTH, M. W. WOLFE, R. J. KITTOK, H. E. GROTJAN u. J. E. KINDER (1995): Effects of 17β-estradiol on distribution of pituitary isoforms of luteinizing hormone and follicle-stimulating hormone during the follicular phase of the bovine estrous cycle. Biol. Reprod. 52, 297 - 304 KRSMANOVIC, L. Z., L. HU, P. K. LEUNG, H. FENG u. K. J. CATT (2009): The hypothalamic GnRH pulse generator: multiple regulatory mechanisms. Trends Endocrinol. Metab. 20, 402 - 408 KRSMANOVIC, L. Z., L. HU, P. K. LEUNG, H. FENG u. K. J. CATT (2010): Pulsatile GnRH secretion: roles of G protein-coupled receptors, second messengers and ion channels. Mol. Cell Endocrinol. 27, 158 - 163 119 LAMMING, G. E., D. C. WATHES u. A. R. PETERS (1981): Endocrine patterns of the post-partum cow. J. Reprod. Fertil. Suppl. 30, 155 – 170 LANE, E. A., T. SWEENEY, M. RYAN, J. F. ROCHE u. M. A. CROWE (2009): Relationship between serum gonadotropins and pituitary immunoreactive gonadotropins and steroid receptors during the first FSH increase of the estrous cycle and following steroid treatment in heifers. Anim. Reprod. Sci. 112, 66 - 82 LEBLANC, S. (2010): Assessing the association of the level of milk production with reproductive performance in dairy cattle. J. Reprod. Dev. 56 Suppl., 1 - 7 LEUNG, K., V. PADMANABHAN, E. M. CONVEY, R. E. SHORT u. R. B. STAIGMILLER (1984): Relationship between pituitary responsiveness to Gn-RH and number of Gn-RH-binding sites in pituitary glands of beef cows. J. Reprod. Fertil. 71, 267 - 277 LÓPEZ - GATIUS, F., I. GARCÍA - ISPIERTO, P. SANTOLARIA, J. YÁNIZ, C. NOGAREDA u. M. LÒPEZ - BÉJAR (2006): Screening for high fertility in high - producing dairy cows. Theriogenology 65, 1678 - 89 LÓPEZ - GATIUS, F., A. MIRZAEI, P. SANTOLARIA, G. BECH - SÀBAT, C. NOGAREDA, I. GARCIA - ISPIERTO, C. HANZEN u. J. L. YÁNIZ (2008): Factors affecting the response to the specific treatment of several forms of clinical anestrus in high producing dairy cows. Theriogenology 69, 1095 - 1103 120 LUCY, M. C., J. D. SAVIO, L. BADINGA, R. L. DE LA SOTA u. W. W. THATCHER (1992): Factors that affect ovarian follicular dynamics in cattle. J. Anim. Sci. 70, 3615 – 3626 MACMILLAN, K. L. (2010): Recent advances in the synchronization of estrus and ovulation in dairy cows. J. Reprod. Dev. 56, 42 - 47 MACMILLAN, K. L., I. J. LEAN u. C. T. WESTWOOD (1996): The effects of lactation on the fertility of dairy cows. Aust. Vet. J. 73, 141 – 147 MACMILLAN, K. L. u. W. W. THATCHER (1991): Effects of an agonist of gonadotropin-releasing hormone on ovarian follicles in cattle. Biol. Reprod. 45, 883 – 889 MARKUSFELD, O. (1987): Periparturient traits in seven high dairy herds. Incident rates, association with parity and interrelationships among traits. J. Dairy Sci. 70, 158 - 166 MAYER, E., G. FRANCOS u. A. NERIA (1987): Eierstocksbefunde und Fertilitäts-Parameter bei Kühen mit "unbeobachteter Brunst". Tierärztl. Umsch. 42, 506 – 509 MC DOUGALL, S. (2010): Effects of treatment of anestrous dairy cows with gonadotropin-releasing hormone, prostaglandin, and progesterone. J. Dairy Sci. 93, 1944 - 1959 121 MC DOUGALL, S. u. C. COMPTON (2006): Reproductive performance in the subsequent lactation of dairy cows previously treated for failure to be detected in oestrus. N. Z. Vet. J. 54, 132 - 140 MC DOUGALL, S., C. W. R. COMPTON u. F. M. ANNISS (2004): Effect of exogenous progesterone and oestradiol on plasma progesterone concentrations and follicle wave dynamics in anovulatory anoestrous post-partum dairy cattle. Anim. Reprod. Sci. 84, 303 – 314 MC DOUGALL, S., C. W. R. COMPTON, D. W. HANLON, P. J. DAVIDSON, D. J. SULLIVAN, A. H. GORE u. F. M. ANNISS (2005): Reproductive performance in anestrous dairy cows following treatment with two protocols and two doses of progesterone. Theriogenology 63, 1529 – 1548 MC LEOD, B. J. u. M. E. WILLIAMS (1991): Incidence of ovarian dysfunction in post partum dairy cows and the effectiveness of its clinical diagnosis and treatment. Vet. Rec. 128, 121 – 124 MIALOT, J. P., G. LAUMONNIER, C. PONSART, H. FAUXPOINT, E. BARASSIN, A. A. PONTER u. F. DELETANG (1999): Postpartum subestrus in dairy cows: comparison of treatment with prostaglandin F2 alpha or GnRH + prostaglandin F2 alpha + GnRH. Theriogenology 52, 901 – 911 MILLAR, R. P. (2005): GnRHs and GnRH receptors. Anim. Reprod. Sci. 88, 5 – 28 122 MITKO, K., S. E. ULBRICH, H. WENIGERKIND, F. SINOWATZ, H. BLUM, E. WOLF u. S. BAUERSACHS (2008) Dynamic changes in messenger RNA profiles of bovine endometrium during the oestrous cycle. Reproduction 135, 225 – 240 MOLTER-GÉRARD, C., J. FONTAINE, S. GUÉRIN u. C. TARAGNAT (1999): Differential regulation of the gonadotropin storage pattern by gonadotropin-releasing hormone pulse frequency in the ewe. Biol. Reprod. 60, 1224 - 1230 MORI, Y. u. T. TANAKA (1995): Electrophysiological approach to the hypothalamic GnRH pulse generator. J. Reprod. Fertil. Suppl. 49, 231 - 243 MURPHY, M. G., M. P. BOLAND u. J. F. ROCHE (1990): Pattern of follicular growth and resumption of ovarian activity in post-partum beef suckler cows. J. Reprod. Fert. 90, 523 – 533 MUTTUKRISHNA, S. u. P. G. KNIGHT (1991): Inverse effects of activin and inhibin on the synthesis and secretion of FSH and LH by ovine pituitary cells in vitro. J. Mol. Endocrinol. 6, 171 - 178 NATION, D. P., C. R. BURKE, G. PARTON, R. STEVENSON u. K. L. MACMILLAN (2000): Hormonal and ovarian responses to a 5-day progesterone treatment in anoestrous dairy cows in the third week post-partum. Anim. Reprod. Sci. 63, 13 – 25 123 NAWITO, M., E. SCHALLENBERGER u. D. SCHAMS (1977): Release of lutropin (LH) and follitropin (FSH) in cattle after administration of a new gonadoliberin (GnRH) analogue in comparison with the gonadoliberin decapaptide. Theriogenology 7, 277 - 291 OLIVEIRA, K. P. L., A. A. ARAUJO, P. A. MONTEZUMA JUNIOR, A. A. A. MOURA u. J. F. NUNES (2003): Comparacao de dois metodos de inducao e sincronizacao de estro e da ovulacao em vacas leiteiras mesticas no anestro pos-parto. Comparison of two methods of inducation and synchronisation of oestrus and the ovulation in cows in postpartum anoestrus. Revista Brasileira de Reproducao Animal 27, 453 – 454 OPSOMER, G. u. A. DE KRUIF (1999): Postpartaler Anöstrus bei Milchkühen – eine Übersicht. Tierärztl. Prax. 27 (G), 30 – 35 OPSOMER, G., Y. T. GRÖHN, J. HERTL, M. CORYN, H. DELUYKER u. A. DE KRUIF (2000): Risc factors for post partum ovarian dysfunction in high producing dairy cows in Belgium: a field study. Theriogenology 53, 841 – 857 OZTÜRK, O. A., U. CIRIT, A. BARAN u. K. AK (2010): Is Doublesynch protocol a new alternative for timed artificial insemination in anestrous dairy cows. Theriogenology, 73, 568 - 576 124 PADMANABHAN, V., D. BATTAGLIA, M. B. BROWN, F. J. KARSCH, J. S. LEE, W. PAN, D. J. PHILLIPS u. J. VAN CLEEFF (2002): Neuroendocrine control of follicle-stimulating hormone (FSH) secretion: II. Is follistatininduced suppression of FSH secretion mediated via changes in activin availability and does it involve changes in gonadotropin-releasing hormone secretion? Biol. Reprod. 66, 1395 - 1402 PATTON, J., D. A. KENNY, S. MCNAMARA, J. F. MEE, F. P. O'MARA, M. G. DISKIN, J.J. MURPHY (2007): Relationships among milk production, energy balance, plasma analytes, and reproduction in Holstein-Friesian cows. J. Dairy Sci. 90, 649 - 658 PAWSON, A. J. u. A. S. MC NEILLY (2005): The pituitary effects of GnRH. Anim. Reprod. Sci. 88, 75 - 94 PETERS, A. R. (1984): Reproductive activity of the cow in the post-partum period. I. Factors affecting the length of the post-partum acyclic period. Br. Vet. J. 140, 76 - 84 PETERS, A.R. (1985): Hormonal control of the bovine oestrous cycle. I. The natural cycle. Br. Vet. J. 141, 564 - 575 PETERS, A. R. (2005): Veterinary clinical application of GnRH – questions of efficacy. Anim. Reprod. Sci. 88, 155 – 167 125 PIERCE, J. G., M. R. FAITH, L. C. GIUDICE u. J. R. REEVE (1976): Structure and structure-function relationships in glycoprotein hormones. Ciba Found. Symp. 41, 225 - 250 PRYCE, J. E., M. P. COFFEY u. G. SIMM (2001): The relationship between body condition score and reproductive performance. J. Dairy Sci. 84, 1508 - 1515 RAMAKRISHNAPPA, N., R. RAJAMAHENDRAN, YUNG-MING LIN u. P. C. K. LEUNG (2005): GnRH in non-hypothalamic reproductive tissues. Anim. Reprod. Sci. 88, 95 - 113 RHODES, F. M., C. R. BURKE, B. A. CLARK, M. L. DAY u. K. L. MACMILLAN (2002): Effect of treatment with progesterone and oestradiol benzoate on ovarian follicular turnover in postpartum anoestrous cows and cows which have resumed oestrous cycles. Anim. Reprod. Sci. 15, 139 – 150 RHODES, F. M., S. MCDOUGALL, C. R. BURKE, G. A. VERKERK u. K. L. MACMILLAN (2003): Invited review: Treatment of cows with an extended postpartum anestrous interval. J. Dairy Sci. 86, 1876 - 1894 RICHARDS, M. W., R. P. WETTEMANN u. H. M. SCHOENEMANN (1989): Nutritional anestrus in beef cows: body weight change, body condition, luteinizing hormone in serum and ovarian activity. J. Anim. Sci. 67, 1520 – 1526 126 RILEY, G. M., A. R. PETERS u. G. E. LAMMING (1981): Induction of pulsatile LH release, FSH release and ovulation in post-partum acyclic beef cows by repeated small doses of GnRH. J. Reprod. Fertil. 63, 559 – 565 RISPOLI, L. A. u. T. M. NETT (2005) Pituitary gonadotropin-releasing hormone (GnRH) receptor: structure, distribution and regulation of expression. Anim. Reprod. Sci. 88, 57 - 74 RUTTER, L. M. u. R. D. RANDEL (1984): Postpartum nutrient intake and body condition: effect on pituitary function and onset of estrus in beef cattle. J. Anim. Sci. 58, 265 – 274 RYAN, R. J., M. C. CHARLESWORTH, D. J. MC CORMICK, R. P. MILIUS u. H. T. KEUTMANN (1988): The glycoprotein hormones: recent studies of structure-function relationships FASEB J. 2, 2661 - 2669 SAKAGUCHI, M. (2009): Differences between body condition scores and body weight changes in postpartum dairy cows in relation to parity and reproductive indices. Can. Vet. J. 50, 649 - 656 SAMARÜTEL, J., A. WALDMANN, K. LING, H. JAAKSON, T. KAART, A. LEESMÄE u. O. KÄRT (2008): Relationships between luteal activity, fertility, blood metabolites and body condition score in multiparous Estonian Holstein dairy cows under different management. J. Dairy Res. 75, 485 - 490 127 SANCHEZ, T., M. E. WEHRMAN, G. E. MOSS, F. N. KOJIMA, A. S. CUPP, E. G. BERGFELD, K. E. PETERS, V. MARISCAL, H. E. GROTJAN JR, J. E. KINDER u. D. L. HAMERNIK (1994): Differential regulation of gonadotropin synthesis and release in ovariectomized ewes after treatment with a luteinizing hormone-releasing hormone antagonist. Biol. Reprod. 51, 755 - 759 SAVIO, J. D., M. P. BOLAND, N. HYNES u. J. F. ROCHE (1990 a): Resumption of follicular activity in the early post-partum period of dairy cows. J. Reprod. Fertil. 88, 569 – 579 SAVIO, J. D., M. P. BOLAND u. J. F. ROCHE (1990 b): Development of dominant follicles and length of ovarian cycles in post-partum dairy cows. J. Reprod. Fertil. 88, 581 – 591 SCHAMS, D., E. SCHALLENBERGER u. B. HOFFMANN (1977): The oestrous cycle of the cow: hormonal parameters and time relationships concerning oestrus, ovulation, and electrical resistance of the vaginal mucus. Acta Endocrinol. 86, 180 – 192 SCHOPPER, D., R. SCHEMER, U. WEILER u. R. CLAW (1993): Einfluss der Milchleistung auf Fruchtbarkeitkriterien der Milchkuh post partum: Auswertung von Progersteronprofilen Zuchthygiene 28, 225 - 235 SEGWAGWE, B. V., J. MALMO, K. L. MACMILLAN u. P. D. MANSELL (2006): A comparative study of the effect of 2 hormonal treatment protocols on the reproductive performance of previously anoestrous dairy cows. J. S. Afr. Vet. Med. Assoc. 77, 33 – 39 128 SHEPHARD, R. W. (2005): A comparison of performance of the Ovsynch treatment program between cycling and noncycling cows within seasonally-calving dairy herds. Aust. Vet. J. 12, 751 – 757 SHRESTHA, H. K., T. NAKAO, T. SUZUKI, M. AKITA u. T. HIGAKI (2005): Relationships between body condition score, body weight, and some nutritional parameters in plasma and resumption of ovarian cyclicity postpartum during pre-service period in highproducing dairy cows in a subtropical region in Japan. Theriogenology 64, 855 -866 SIROIS, J. u. J. E. FORTUNE (1988): Ovarian follicular dynamics during the estrous cycle in heifers monitored by real-time ultrasonography. Biol. Reprod. 39, 308 – 317 SMOLDERS, E. A., M. S. THESINGH, P. L. VOS u. A. H. WILLEMSE (1996): Op gang komen van de ovariele cyclus na het kalben. The onset of the ovarian cycle following calving. Tijdschr. Diergeneeskd. 121, 610 – 614 SOSNOWSKI, R., P. L. MELLON u. M. A. LAWSON (2000): Activation of translation in pituitary gonadotrope cells by gonadotropin-releasing hormone. Mol. Endocrinol. 14, 1811 - 1819 STAPLES, C. R. u. W. W. THATCHER (1990): Relationship between ovarian activity and energy status during the early postpartum period of high producing dairy cows. J. Dairy Sci. 73, 938 – 947 129 STEVENSON, J. S. u. J. H. BRITT (1980): Models for prediction of days to first ovulation based on changes in endocrine and nonendocrine traits during the first two weeks postpartum in Holstein cows. J. Anim. Sci. 50, 103 - 112 STUMPF, T. T., M. L. DAY, M. W. WOLFE, A. C. CLUTTER, J. A. STOTTS, P. L. WOLFE, R. J. KITTOK u. J. E. KINDER (1989): Effect of estradiol on secretion of luteinizing hormone during the follicular phase of the bovine estrous cycle. Biol. Reprod. 40, 91 - 97 STUMPF, T. T., M. W. WOLFE, P. L. WOLFE, M. L. DAY, R. J. KITTOK u. J. E. KINDER (1992): Weight changes prepartum and presence of bulls postpartum interact to affect duration of postpartum anestrus in cows. J. Anim. Sci. 70, 3133 – 3137 SWANSON, L. V. u. H. D. HAFS (1971): LH and prolactin in blood serum from estrus to ovulation in Holstein heifers. J. Anim. Sci. 35, 1038 – 1041 TANAKA, T., M. ARAI, S. OHTANI, S. UEMURA, T. KUROIWA, S. KIM u. H. KAMOMAE (2008): Influence of parity on follicular dynamics and resumption of ovarian cycle in postpartum dairy cows. Anim. Reprod. Sci. 108, 134 – 143 THATCHER, W. W., M. DROST, J. D. SAVIO, K. L. MACMILLAN, K. W. ENTWISTLE, E. J. SCHMITT, R. L. DE LA SOTA u. G. R. MORRIS (1993): New clinical uses of GnRH and its analogues in cattle. Anim. Reprod. Sci. 33, 27 – 49 130 THIBAULT, G., M. GERARD u. Y. MENEZO (1975): Preovulatory and ovulatory mechanisms in oocyte maturation. J. Reprod. Fert. 45, 606 – 610 TWAGIRAMUNGU, H., L. A. GUILBAULT, J. G. PROULX u. J. J. DUFOUR (1995): Buserelin alters the development of the corpora lutea in cyclic and early postpartum cows. J. Anim. Sci. 73, 805 – 811 TWAGIRAMUNGU, H., L. A. GUILBAULT, J. PROULX, R. RAMKUMAR u. J. J. DUFOUR (1994): Histological populations and atresia of ovarian follicles in postpartum cattle treated with an agonist of Gonadotropin-Releasing hormone. J. Anim. Sci. 72, 192 - 200 VAN STRATEN, M., N. Y. SHPIGEL u. M. FRIGER (2009): Associations among patterns in daily body weight, body condition scoring, and reproductive performance in high-producing dairy cows. J. Dairy Sci. 92, 4375 - 4385 WALSH, R. B., S. J. LEBLANC, T. D. DUFFIELD, D. F. KELTON, J. S. WALTON u. K. E. LESLIE (2007): Synchronization of estrus and pregnancy risk in anestrous dairy cows after treatment with a progesterone-releasing intravaginal device. J. Dairy Sci. 90, 1139 - 1148 WALTERS, D. L., R. E. SHORT, E. M. CONVEY, R. B. STAIGMILLER, T. G. DUNN u. C. C. KALTENBACH (1982): Pituitary and ovarian function in postpartum beef cows. III. Induction of estrus, Ovulation and luteal function with intermittent small-dose injections of GnRH. Biol. Reprod. 26, 655 – 662 131 WALTERS, D. L. u. E. SCHALLENBERGER (1984): Pulsatile secretion of gonadotrophins, ovarian steroids and ovarian oxytocin during the periovulatory phase of the oestrous cycle in the cow. J. Reprod. Fert. 71, 503 – 512 WALTERS, D. L., D. SCHAMS u. E. SCHALLENBERGER (1984): Pulsatile secretion of gonadotrophins, ovarian steroids and ovarian oxytocin during the luteal phase of the oestrous cycle in the cow. J. Reprod. Fertil. 71, 479 - 491 WALTNER, S. S., J. P. MCNAMARA u. J. K. HILLERS (1993): Relationships of body condition score to production variables in high producing Holstein dairy cattle. J. Dairy Sci. 76, 3410 - 3419 WANG, Q. F., P. G. FARNWORTH, J. K. FINDLAY u. H. G. BURGER (1990): Chronic inhibitory effect of follicle-stimulating hormone (FSH)-suppressing protein (FSP) or follistatin on activin- and gonadotropin-releasing hormone-stimulated FSH synthesis and secretion in cultured rat anterior pituitary cells. Endocrinology 127, 1385 - 1393 WATHES, D. C. u. G. E. LAMMING (1995): The oxytocin receptor, luteolysis and the maintainance of pregnancy. J. Reprod. Fertil. Suppl. 49, 53 – 67 WEBB, R., G. E. LAMMING, N. B. HAYNES u. G. R. FOXCROFT (1980): Plasma progesterone and gonadotrophin concentrations and ovarian activity on post-partum dairy cows. J. Reprod. Fertil. 59, 133 – 143 132 WHISNANT, C. S., T. E. KISER u. F. N. THOMPSON (1985): Effect of calf removal on serum luteinizing hormone and cortisol concentrations in postpartum beef cows. Theriogenology 24, 119 – 129 WHITAKER, D. A., E. J. SMITH, G. O. DA ROSA u. J. M. KELLY (1993): Some effects of nutrition and management on the fertility of dairy cattle. Vet. Rec. 133, 61 – 64 WITTKOWSKI, G., E. GRUNERT u. A. BUKOWSKI (1982): Zur Zyklusinduktion bei Kühen mit Ovardystrophie durch Progesteron abgebende Vaginalspiralen (Abbovestrol©). Dtsch. tierärztl. Wochenschrift 89, 244 – 247 XU, Z. Z., G. A. VERKERK, J. F. MEE, S. R. MORGAN, B. A. CLARK, C. R. BURKE u. L. J. BURTON (2000): Progesterone and follicular changes in postpartum noncyclic dairy cows after treatment with progesterone and estradiol or with progesterone, GnRH, PGF2alpha, and estradiol. Theriogenology 54, 273 – 282 YING, S. Y. (1988): Inhibins, activins and follistatins: gonadal proteins modulating the secretion of folliclestimulating hormone. Endocr. Rev. 9, 267 - 293 ZUREK, E., G. R. FOXCROFT u. J. J. KENNELLY (1995): Metabolic status and interval to first ovulation in postpartum dairy cows. J. Dairy Sci. 78, 1909 – 1920 133 Erklärung Hiermit erkläre ich, dass ich die Dissertation "Einfluss einer wiederholten Applikation von Gonadotropin - Releasing Hormon auf die Fruchtbarkeit azyklischer Milchkühe" selbstständig verfasst habe. Bei der Anfertigung wurde keine Hilfe Dritter in Anspruch genommen. Ich habe keine entgeltliche Hilfe von Vermittlungs- bzw. Beratungsdiensten (Promotionsberater oder anderer Personen) in Anspruch genommen. Niemand hat von mir unmittelbar oder mittelbar entgeltliche Leistungen für Arbeiten erhalten, die im Zusammenhang mit dem Inhalt der vorgelegten Dissertation stehen. Ich habe die Dissertation an folgender Institution angefertigt: Klinik für Rinder der Tierärztlichen Hochschule Hannover. Die Dissertation wurde bisher nicht für eine Prüfung oder Promotion oder für einen ähnlichen Zweck zur Beurteilung eingereicht. Ich versichere, dass ich die vorstehenden Angaben nach bestem Wissen vollständig und der Wahrheit entsprechend gemacht habe. Hanna Bläsing 134 Danksagung Frau Prof. Dr. Martina Hoedemaker danke ich sehr herzlich für die Überlassung des interessanten Themas und die stets gewährte Hilfe und freundliche Betreuung bei der Verfassung dieser Arbeit. Mein großer Dank gilt Dr. Cord Müller und Dr. Ralph Müller und den Mitarbeitern der Gemeinschaftspraxis für die Möglichkeit, die Dissertation in der Praxis und während der Arbeitszeit anfertigen zu können, sowie für die ständige Unterstützung und Aufmunterung. Den beteiligten Landwirten und Mitarbeitern der Betriebe danke ich ganz herzlich für die Bereitstellung der Kühe und der Daten, die Hilfe bei den Untersuchungen und das Interesse an den Fortschritten dieser Arbeit. Bei der Firma Intervet Deutschland GmbH bedanke ich mich für die finanzielle Unterstützung der Dissertation. Vielen Dank an Dr. Annette Krech für das schnelle und hilfreiche Korrekturlesen und an Kathrin Johanning für die Korrektur der englischen Zusammenfassung. Ein besonderer Dank geht an meine Schwester Lena für die Beratung bei der statistischen Auswertung und für alles andere. Bei meiner Familie, vor allem bei meinen Eltern und meinem Opa möchte ich mich sehr herzlich bedanken. Ihr wart immer für mich da und habt mich unterstützt. Danke! 135