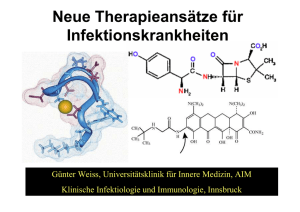
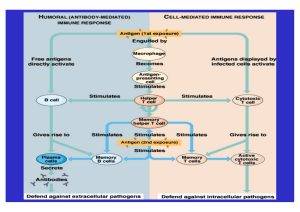
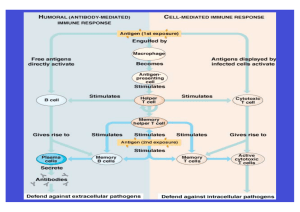
Band 41
Werbung

ZivilschutzForschung Schriftenreihe der Schutzkommision beim Bundesminister des Innern Herausgegeben vom Bundesverwaltungsamt – Zentralstelle für Zivilschutz – im Auftrag des Bundesministerium des Innern Neue Folge Band 41 W. König, A. Drynda, B. König, R. Arnold, P. Wachtler, M. Köller Einfluss von Zytokinen und Lipidmediatoren auf die Kontrolle und Regulation spezifischer Infektabwehr bei Brandverletzung ISSN 0343-5164 ZIVILSCHUTZFORSCHUNG Neue Folge Band 41 ZivilschutzForschung Schriftenreihe der Schutzkommision beim Bundesminister des Innern Herausgegeben vom Bundesverwaltungsamt – Zentralstelle für Zivilschutz – im Auftrag des Bundesministerium des Innern Neue Folge Band 41 W. König, A. Drynda, B. König, R. Arnold, P. Wachtler, M. Köller Einfluss von Zytokinen und Lipidmediatoren auf die Kontrolle und Regulation spezifischer Infektabwehr bei Brandverletzung ISSN 0343-5164 Herausgeber: Bundesverwaltungsamt – Zentralstelle für Zivilschutz – Deutschherrenstr. 93–95, 53177 Bonn Telefon: (0 18 88) 3 58-0 Telefax: (0 18 88) 3 58-58 03 Internet: www.bundesverwaltungsamt.de Die Arbeit gibt die Meinung der Autoren wieder. Sie stellt keine Äußerung des Herausgebers dar und ist auch nicht als solche auszulegen. © 2001 Bundesverwaltungsamt – Zentralstelle für Zivilschutz – Bonn Satz und Druck: Druckhaus Dresden GmbH 4 Inhaltsverzeichnis Einleitung. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 7 Kurze Darstellung zur Aufgabenstellung: . . . . . . . . . . . . . . . . . . . . . . 7 Voraussetzungen unter denen das Vorhaben durchgeführt wurde . . . . . . . 9 Planung und Ablauf des Vorhabens. . . . . . . . . . . . . . . . . . . . . . . . . . . 9 Wissenschaftlicher und technischer Stand. . . . . . . . . . . . . . . . . . . . . 10 Zusammenarbeit mit anderen Stellen.. . . . . . . . . . . . . . . . . . . . . . . . 10 Ergebnisse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 Einleitung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11 Mikrobielle Pathogenität und Pathogenitätsfaktoren . . . . . . . . . . . . 13 Untersuchungen zu mikrobiellen Pathogenitätsfaktoren. . . . . . . . . . 15 Untersuchungen zum Staphylokokken Leukozidin. . . . . . . . . . . . . . 15 Pathogenitätsfaktoren von Pseudomonas aeruginosa . . . . . . . . . . . . 16 Untersuchungen zur Bedeutung mikrobieller Superantigene . . . . . . 18 Modulation der Immuneffektorreaktionen durch RSV (Respiratory Syncytial Virus).. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20 Zusammenfassung: . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20 Bedeutung von Pharmaka (Zytokine, lösliche Rezeptoren, Antagonisten u.a.) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21 Bedeutung von Wachstumsfaktoren:. . . . . . . . . . . . . . . . . . . . . . . . . 21 Untersuchungen zum PAF-Rezeptorantagonisten. . . . . . . . . . . . . . . 22 Zusammenfassung: . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22 Therapeutische Interventionen bei der traumabedingten Immunparalyse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23 Untersuchungen zum Einfluss von γ-Interferon beim Brandverletzten . . 23 Immunmodulation – Untersuchungen zur Immunmodulation im Rahmen der Ernährungstherapie.. . . . . . . . . . . . . . . . . . . . . . . . . 24 Erfolgskontrollbericht . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27 Voraussichtlicher Nutzen, insbesondere Verwertbarkeit des Ergebnisses und der Erfahrungen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 29 Kurzfassung . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31 Literatur . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33 Die Autoren . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39 Anhang . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 41 5 Einleitung Kurze Darstellung zur Aufgabenstellung: Unter dem Thema „Einfluss von Zytokinen und Lipidmediatoren auf die Kontrolle und Regulation spezifischer Infektabwehr bei Brandverletzung“ wurde ein theoretisch klinisches und klinisch-experimentelles Projekt gefördert. Aus langjährigen Arbeiten wissen wir, dass die traumabedingte Immunparalyse, wie sie z.B. bei Polytrauma oder bei der Brandverletzung auftritt, durch eine Dysbalance und u.U. durch ein Versagen der Mediatorkaskaden zustande kommt. Am Beispiel der Brandverletzung wird dabei offensichtlich, dass nach einer Phase der Kolonisierung entweder im Rahmen der Stabilisierung der Infektabwehr keine mikrobielle Invasion stattfindet oder unter Zunahme des Pathogenitätserwerbs von Mikroorganismen und Veränderung der Infektabwehr es zur mikrobiellen Invasion kommt. Die Aufgabenstellung sollte deshalb mikrobielle Pathogenitätsfaktoren berücksichtigen, die die Immunparalyse einleiten und andererseits zellbiologische und immunologische Parameter erarbeiten, die die Immunparalyse anzeigen. Daraus resultierend sollen dann diagnostische Kriterien erarbeitet werden, die als frühe Prädiktivmarker der Immunparalyse eingesetzt werden können. Auf dem Boden der Erkenntnisse zur traumainduzierten Immunparalyse sollen für die Zukunft neue therapeutische Ansätze erarbeitet werden. Die vorliegenden Ergebnisse umfassen deshalb Arbeiten zur Validität von mikrobiellen Pathogenitätsfaktoren, wobei als Indikatorzellen die Immunzellen normaler Donoren eingesetzt wurden. Die Wechselwirkung dieser mikrobiellen Pathogenitätsfaktoren mit den Zielzellen erlaubt es, anhand von zellbiologischen Charakteristika die Aktivität der mikrobiellen Pathogenitätsfaktoren zu beurteilen. In einem weiteren Ansatz wurden bekannte Stimulatoren eingesetzt und Zielzellen von Patienten mit Brandverletzung, Polytrauma oder großflächigen operativen Eingriffen analysiert. Aus den erzielten Ergebnissen kommen wir zu neuen Erkenntnissen hinsichtlich der Pathogenität von mikrobiellen Exoprodukten, der Bedeutung von inflammatorischen Mediatoren in der Regulation wie aber auch in der Paralyse zellbiologischer Funktionen sowie zu neuen Denkkonzepten hinsichtlich therapeutischer Ansätze zur Vermeidung der Immunparalyse. Selbstverständlich besteht auch für die Zukunft großer Forschungsbedarf. 7 Voraussetzungen unter denen das Vorhaben durchgeführt wurde Die Arbeitsgruppe für Infektabwehr (Leiter Prof. Dr. med. W. König) hat über Jahre die zellbiologischen und immunologischen Voraussetzungen etabliert, um die mikrobielle Pathogenität in der Wechselwirkung mit humanen Effektorzellen zu quantifizieren und zu analysieren. So wurden die Mediatorkaskaden (Lipidmediatoranalyse, Zytokinanalyse, Adhäsinrezeptoren u.a.) in der Abteilung etabliert und in vielen Patientenstudien eingesetzt. Eine sehr enge Zusammenarbeit hinsichtlich des Arbeitsprojektes war durch die Interaktion mit der Abteilung für Plastische Chirurgie (Prof.Dr.med. Steinau), den Städtischen Krankenanstalten Dortmund, Universitätsklinik Marienhospital Herne sowie der Universitätsklinik Josefs-Hospital u.a. gegeben. Durch die Interaktion mit den klinischen Kollegen der dortigen Abteilung wurde ein entsprechendes Krankengut zusammengestellt und wesentliche Fragen hinsichtlich der Wechselwirkung mikrobieller Pathogenitätsfaktoren an immungestressten Effektorzellen analysiert. Fernerhin hatten wir durch diese enge Verzahnung die Möglichkeit, neue Therapieansätze (Dipeptid, Eicosapentaensäure u.a.) in klinischen Studien zu analysieren. Im Rahmen einer „Multicenterstudie“, wurde der Einfluss von (-Interferon auf die Reaktivierbarkeit von humanen Effektorzellen bei Brandverletzung analysiert. Als wesentlicher Marker der zellulären Reaktivität wurde die Expression von MHCII analysiert. Planung und Ablauf des Vorhabens Im Wesentlichen wurde das Vorhaben über zwei thematisch verzahnte Bereiche durchgeführt; es hatte sich in der Vergangenheit erwiesen, dass eine moderne Betrachtung zur klinischen Pathophysiologie der Immunparalyse z.B. der Brandverletzten nur dann getroffen werden kann, wenn gleichzeitig die dazugehörige Grundlagenforschung aktiviert und gefördert wird. Aus diesem Grunde wurde das Projekt zweigleisig angelegt: a) zum einen wurde der Einfluss mikrobieller Pathogenitätsfaktoren an humanen Immuneffektorzellen analysiert, b) zum anderen wurde die Reaktivität humaner Immuneffektorzellen unter dem Trauma analysiert. Dies Vorgehen erwies sich als ausserordentlich vernünftig, da man die Reaktivität normaler Immuneffektorzellen hinsichtlich unterschiedlicher Pathogenitätsfaktoren kennen muss um dann zu analysieren, inwieweit an verschiedenen Signaltransduktions- oder Schaltelementen hinsichtlich der Mediatorgenerierung, Produktion oder des Verhältnisses der Mediatoren untereinander bei immungestressten Patienten ein Defizit, ein Überschießen oder ein Missverhältnis auftritt. Es war also unser Ziel die Ergebnisse aus beiden Bereichen so abzurunden, dass wir diese basistheoretischen Ergebnisse für die Klinik verfügbar machen, um gleichermaßen eine Vertiefung des pathophysiologischen Wissens zu erreichen. 9 Infolge des dualen Ansatzes (klinisch-theoretisch, klinisch) wurden die Arbeiten parallel zueinander vorangetrieben. Es wurden die klinisch-theoretischen Ansätze an normalen Spenderzellen durchgeführt. Zu diesem Zweck wurden normale Spender einbestellt, es wurde Blut (200 ml) entnommen. Nach bewährten Methoden der Zellauftrennung wurden die Zellen isoliert und mit den mikrobiellen Pathogenitätsfaktoren, die in der Abteilung oder mit anderen Institutionen (national, international) erarbeitet wurden, inkubiert. Die Auswertungen zu den erzielten Resultaten (Lipidmediatoren, Zytokine, Lymphozyten, Funktionsanalysen u.a.) wurden in der Arbeitsgruppe für Infektabwehr durchgeführt. Parallel zu diesen Experimenten, die also stringent anhand der erhaltenen Ergebnisse weiterentwickelt wurden, waren wir auf die Spenderzellen aus den unterschiedlichen Kliniken angewiesen. Wenn diese Spender nach Rückkoppelung mit den Klinikern in den Abteilungen vorlagen, wurde Blut im Rahmen der Routineuntersuchung bei den Patienten entnommen und dann zur Arbeitsgruppe für Infektabwehr gebracht. Bei einem Großteil der Patienten haben wir den Transfer der Proben selber durchgeführt. Dies beinhaltete teilweise auch Fahrten von Bochum bis Essen (Universitätsklinik) oder von Bochum nach Dortmund (Städt. Krankenanstalten). Wissenschaftlicher und technischer Stand Die Arbeitsgruppe für Infektabwehr besteht seit 1979. Sie untersteht einem Universitätsprofessor (Prof. Dr. med. W. König). Der wissenschaftliche und technische Stand ist voll gewährleistet und viele internationale Publikationen resultierten aus der vorliegenden Förderung. Hinsichtlich der verwendeten Fachliteratur, Informations-, Dokumentationsdienste benutzen wir die moderne Kommunikation auf allen unseren Fachgebieten sowie insbesondere Kontakte zu internationalen Wissenschaftlern durch Vorstellen der Resultate bei internationalen Konferenzen. Aufgrund dieser langjährigen Interaktionen auf den vorliegenden Gebieten haben wir Kooperationen mit Arbeitsgruppen in Europa sowie in den USA aufgebaut. Zusammenarbeit mit anderen Stellen Hier sind als erstes zu nennen die Universitätskliniken der Ruhr-Universität Bochum, die Universitätsklinik in Essen, die Städtischen Krankenanstalten Dortmund; hinsichtlich der theoretisch-klinischen Arbeiten sind zu nennen die Arbeitsgruppen aus den USA (Colorado, NIH), aus Europa (Institut Pasteur, Paris, Institut für Mikrobiologie, Straßburg). 10 Ergebnisse Einleitung Die innovativen Erkenntnisse auf dem Gebiet der mikrobiellen Infektabwehr und hier insbesondere zur Pathophysiologie der traumabedingten Immunsuppression sind gekennzeichnet durch die vertieften Vorstellungen zu den unterschiedlichen Mediatorkaskaden. Die inflammatorischen Effektorzellen, wie z.B. Neutrophile bilden erste Abwehrbarrieren für Mikroorganismen. Mikroorganismen können durch bakterielle Pathogenitätsfaktoren ihrerseits wie auch insbesondere durch Exoprodukte (Toxine, Superantigene u.a.) die Reaktivität der Effektorzellen verändern. Unter den Immuneffektorzellen spielen nicht nur neutrophile Granulozyten, Monozyten sondern auch Entzündungszellen wie Basophile, Mastzellen und Eosinophile eine bedeutsame Rolle. Die letzteren Zellen, die zunächst vornehmlich als relevant für allergische Reaktionen bekannt waren, spielen auch bei traumainduzierten Immunreaktionen eine besondere Rolle. Nach Aktivierung von Entzündungszellen über pseudoallergische Aktivatoren kommt es zur Freisetzung vielfältiger Mediatoren. Die Freisetzung der Entzündungsmediatoren beeinflusst wiederum den Reaktivitätserwerb im Rahmen der spezifischen Infektabwehr. Die mikrobielle Infektabwehr ist gekennzeichnet durch die unspezifische sowie durch die spezifische Infektabwehr. Träger der unspezifischen Infektabwehr sind einerseits inflammatorische Immuneffektorzellen, Serumkomponenten, Zytokine u.a.; Träger der spezifischen Infektabwehr sind Lymphozyten, die einerseits die humorale oder die zellvermittelte Immunantwort einleiten; verknüpfende Komponenten für die unspezifische und spezifische Infektabwehr sind die vielfältigen Entzündungsfaktoren. (Abb.1) In der Vergangenheit war den Lipidmediatoren besondere Aufmerksamkeit geschenkt worden. Vielfältige aktivierende Stimuli können rezeptorvermittelt oder nichtrezeptorvermittelt Zellen so stimulieren, dass es über eine Phospholipaseaktivierung zu einer Translokation der 5-Lipoxygenase mit der Freisetzung von Leukotrienen kommt. Diese Mediatoren haben chemotaktische und spasmogene Einwirkungen. Nach neueren Arbeiten können Lipidmediatoren wiederum Zytokine in ihrer Aktivität regulieren. Somit entsteht ein Konzept, dass verschiedene Mediatorkaskaden interagieren, teilweise synergistisch, additiv oder sogar antagonistisch wirksam sind. (Abb.2) Mediatoren ihrerseits können wiederum andere Mediatoren freisetzen oder die Zelle „primen“, so dass ein exogener Stimulus nun zu einer hyperreaktiven Freisetzung von Mediatoren führt. Viele Mediatoren führen ihre zellbiologische Funktion durch rezeptorvermittelte Wechselwirkung aus. Diese Rezeptoren können nach der Aktivierung von Zellen freigesetzt werden, wie es z.B. für den Tumornekrosefaktor (TNF) gezeigt wird. Der Mediator für sich hat ein duales Gesicht. Für die Homeoestase der Infektabwehr ist die balancierte Freisetzung unterschiedlicher Mediatorkaskaden notwendig. Eine zu starke Freisetzung von Mediatoren könnte zu einer überschießenden Entzündung führen. Ein Versagen der Mediatorkaskaden leitet die Immunparalyse ein. Unter den neugenerierten Mediatoren – hier handelt es sich meistens um Peptide – gibt es pro- und antiinflammatorische Mediatoren. (Tabellen 1,2,3) 11 Das Konzept, das Interleukine und Zytokine aus unterschiedlichen Vorläuferzellen stammen, hat zu einer didaktischen Einteilung hinsichtlich von Schutz und Schaden geführt. Aufgrund unterschiedlicher Infektionsmodelle weiß man, dass aus T-Helferzellen des Typs 1 die Zytokine IL-2 und Interferon-γ vornehmlich gebildet werden. Diese Zytokine werden für die protektive Infektabwehr in der Regel benötigt; aus T-Helferzellen des Typs 2 werden Zytokine wie IL-4, IL-5, IL-13 freigesetzt. (Abb.3) Diese Zytokine haben im Rahmen der allergisch-entzündlichen Reaktion eine herausragende Bedeutung. Die Interleukine IL-4 und IL-13 wirken als „Switch“-Faktoren bei der IgE-Synthese. Interleukin-5 hat darüber hinaus pleiotrope Funktionen und findet sich bei der mikrobiell induzierten Immunsuppression von der Konzentration her erhöht. Viele dieser Mediatoren haben also überlappende Funktionen und sind für unterschiedliche Krankheitsbilder bedeutsam. Interleukin-5 führt zur Eosinophilen Akkumulation, wie es bei allergisch-entzündlichen Reaktionen aber auch beim ARDS und Polytrauma gesehen wird. TH2-Zellen können nach ihrer Aktivierung auch Interleukin-10 wiederum freisetzen. Das Interleukin-10 supprimiert TH-1 vermittelte Zytokine und favorisiert damit die TH-2-Zellentwicklung. Aus Makrophagen und NK-Zellen wird Interleukin-12 nach der Aktivierung gebildet. Interleukin-12 wiederum induziert die Freisetzung von γ-Interferon. Somit findet sich für fast jede Mediatorkaskade ein aktivierender und gegenregulatorischer Bogen. Aufgrund dieser komplexen Ergebnisse ist es also verständlich dass nach Eckpfeilern gesucht werden muss, um den Status der Infektabwehr genügend genau zu analysieren. Für die hier vorliegenden Arbeiten hat es sich als günstig erwiesen, einerseits Zytokine wie Interleukin-1, Interleukin-6 zu studieren, das Chemotaxin Interleukin-8 als einen Vertreter der chemotaktischen Peptide, den Tumornekrosefaktor, der für Schock-Sepsis verantwortlich gemacht wird, allerdings auch benefizielle Funktionen aufweist sowie Interleukin-4 und Interleukin-13, fernerhin die Zytokine Interferon-γ und Interleukin-12. (Tabelle 4) In den vergangenen Jahren hatten wir gezeigt, dass ein frühes Merkmal der verbrennungsinduzierten Immunsuppression wie auch beim ARDS darin besteht, dass Immuneffektorzellen (Neutrophile) eine verminderte Freisetzung von chemotaktisch aktivem Leukotrien B4 zeigten. Der Neutrophile Granulozyt war in seiner Reaktivität hinsichtlich der LTB4-Freisetzung weitestgehend inaktiviert. Eine Reduktion von LTB4-Rezeptoren auf der Oberfläche der Neutrophilen wurde von uns beschrieben. Darüberhinaus konnten wir zeigen, dass im Vergleich zur Überlebensgruppe bei Brandverletzten ein irreversibler Anstieg von Interleukin-6 vorlag. Hinsichtlich des Differenzierungsmarkers von B-Zellen (CD23) zeigte sich im Verlauf der Brandverletzung (hier insbesondere vor der mikrobiellen Invasion) unter den Bedingungen von Schock und Sepsis eine Unfähigkeit der B-Lymphozyten nach Interleukin-4-Zugabe eine CD23-Expression aufzuzeigen. Auf dem Boden der hier vorliegenden Erkenntnisse ist es also verständlich, dass offenbar frühe zellbiologische Prädiktoren auftreten, die vor der mikrobiellen Invasion das Bild der drohenden Sepsis anzeigten. Die Entwicklung auf dem Gebiet zur Pathophysiologie der traumabedingten Immunsuppression war durch die Vielzahl der o.g. Zytokine erweitert worden. Neben der Wechselwirkung von Zytokinen mit Immuneffektorzellen sowie Zellen der spezifischen Abwehr kamen neue Ergebnisse zur Überlebenszeit von Zellen hinzu. Der Terminus der 12 „Apoptose (Zelltod)“ wurde geprägt. Ergebnisse aus der Grundlagenforschung hatten gezeigt, dass eine DNA-Fragmentierung auftritt, die z.B. durch Toxine unter nicht-toxischem Einfluss zustande kommt. Im Rahmen dieser Fragmentierung kommt es zu einer Abnahme der zellulären Leistung mit den entsprechenden zellulären Funktionen. Weiterführende Arbeiten konnten darüber hinaus darlegen, dass die „Apoptose“ durch Zytokine wie GM-CSF und G-CSF weitestgehend aufgehoben wird. Daraus ergab sich, dass Wachstumsfaktoren u.U. die Reaktivität von Immuneffektorzellen erhalten können. Mikrobielle Pathogenität und Pathogenitätsfaktoren In den vergangenen zwei Dekaden wurden enorme Fortschritte auf dem Gebiet der Zell- und Molekularbiologie hinsichtlich der Entzündungsmediatoren und ihrer pathophysiologischen Auswirkungen bei Infektionskrankheiten und immunpathologischen Stresssituationen erhoben. In diesem Zusammenhang kommt den Lipidmediatoren sowie in den vergangenen zehn Jahren den Zytokinen eine besondere Bedeutung zu. Die pathophysiologische Rolle der Zytokine wurde bei vielen Infektionen anhand von Infektionsmodellen analysiert. Es konnte festgestellt werden, dass beim gramnegativen septischen Schock die Lipopolysaccharide offenbar eine bedeutende Rolle spielen. Die pathologischen Auswirkungen des Endotoxinschocks wurden durch die Freisetzung von Interleukin-1, Interleukin-6 und des Tumornekrosefaktors erklärt. Diese Zytokine können wiederum andere Mediatoren induzieren, wie z.B. Prostanoide sowie auch endotheliale Adhesionsmoleküle. Die Molekularbiologie zur Kenntnis der Adhäsionsmoleküle hat dargelegt, dass Entzündungsmediatoren diese Strukturen an Endothelzellen und Epithelzellen regulieren, so dass es zu einer Stase der Leukozyten kommen kann. Als Folgereaktion kommt es zur Bildung und Freisetzung inflammatorischer Moleküle. Es ergibt sich eine Abnahme der Herzfunktion, des Blutdrucks und es kommt zu markanten Einflüssen an Immuneffektorzellen. Diese Veränderungen führen dann zum multiplen Organversagen und zum Zelltod des Patienten. Der septische Schock ist dabei eine akut auftretende Reaktion, wobei diese proinflammatorischen Zytokine ihre große Bedeutung entfalten; die Zytokine haben jedoch weitgespannte Auswirkungen und finden sich auch bei chronischen Entzündungsreaktionen und Infektionserkrankungen wie z.B. Lepra, Tuberkulose u.a. Zytokine führen ebenfalls zur Induktion der „Akute-Phase“-Antwort; dies ist ein integrierter Bestandteil der natürlichen Abwehr gegenüber Mikroorganismen. Durch molekularbiologische Untersuchungen hinsichtlich der mikrobiellen Erreger wie auch des Wirtes ist es heute möglich, die Bedeutung einzelner Pathogenitätsfaktoren und einzelner Zytokine tierexperimentell zu analysieren. Die Anwendung genetisch klonierter Mikroorganismen in der Wechselwirkung mit humanen Effektorzellen hat es darüber hinaus ermöglicht, eine Übertragung bisheriger und neuer Erkenntnisse zum Problem „Sepsis“ zu erhalten. Während in den vergangenen 30 – 40 Jahren den Endotoxinen eine ausschließliche Bedeutung zugesprochen wurde, ist heute allgemein akzeptiert, dass abseits vom Lipopolysaccharid andere mikrobielle Faktoren modulierende Funktionen ausüben und u.U. sogar wichtiger sind als das Lipopolysaccharid. Diese unterschiedlichen 13 mikrobiellen Faktoren können heute in vier Klassen eingeteilt werden; Adhäsine, Inversine, Aggressine und Impedine. (Abb.4 a,b) Adhäsine sind mikrobielle Pathogenitätsfaktoren, wie z.B. Fimbrien an E.coli oder Pseudomonas ssp., die sich an spezifische zelluläre Rezeptoren binden. Es handelt sich in der Regel um Karbohydrate und Karbohydrat-Wechselwirkungen, die wiederum zu zellbiologischen Veränderungen am Erfolgsorgan führen. Wir konnten nachweisen, dass definierte Adhäsine von E.coli Bakterien in sehr spezifischer Weise aus neutrophilen Granulozyten (PMN) sowie Lymphozyten, Monozyten, Basophilen (LMB), die Mediatoren wie Leukotriene und Histamin freisetzen. Damit war ersichtlich, dass die Adhäsin-Immuneffektor-Wechselwirkung zur Aktivierung der Zielzellen führt. Dies bedeutet, dass die Kolonisierung nur eine Voraussetzung für die nachfolgende Infektion ist und die entsprechende AdhäsinZellwechselwirkung die pathophysiologischen Reaktionen induziert. Invasine: Es sind Strukturen von Mikroorganismen, die für die Aufnahme des Mikroorganismus von wesentlicher Bedeutung sind. Es handelt sich ebenfalls um mikrobielle Oberflächenstrukturen, die in Wechselwirkung mit Epitopen der Zelle treten. Bekannt sind hier das Invasin von Listeria monocytogenes (Inv, ail) oder Yersinia ssp. (Yops), Salmonella ssp.. Invasine erlauben durch Wechselwirkung des Mikroorganismus mit den zellulären Strukturen eine Aktivierung innerhalb der Zelle, so dass der Mikroorganismus zur Aufnahme kommt. Innerhalb der Zelle kommt es zur Wechselwirkung des Mikroorganismus mit intrazellulären Strukturen und damit zu einer Modulation der zellulären Funktion. Aggressine: Es handelt sich in der Regel um mikrobielle Exotoxine oder auch Proteasen. Die Vielfältigkeit der Toxine hinsichtlich der Wechselwirkung an Immuneffektorzellen wurde in den vergangenen Jahren interdisziplinär erarbeitet. Es zeigte sich, dass viele Hämolysine durch Rezeptorwechselwirkung Zellen aktivieren können, dazu gehören z.B. das Pneumolysin oder Streptolysin O. Das letztere Toxin bindet sich an Cholesterol und führt zur Freisetzung von Leukotrienen. Andererseits können mikrobielle Exotoxine auch modulierend einwirken und die Mediatorkaskaden selektiv aktivieren oder inhibieren. Ein klassisches Beispiel ist das E.coli α-Hämolysin. Dieses Toxin supprimiert z.B. die Zytokinfreisetzung. Man kann sich also vorstellen, dass E.coli Bakterien mit spezifischen Adhäsinen und der α-Hämolysinausprägung zunächst Immuneffektorzellen aktivieren; die gleichzeitige Freisetzung von α-Hämolysin würde die Zytokinfreisetzung supprimieren und damit die spezifische Infektabwehr inhibieren. Impedine: Dies sind Faktoren, die die zelluläre Funktion supprimieren. Auf die duale Funktion vieler mikrobieller Exotoxine wurde eingegangen. Mikrobielle Superantigene oder z.B. das Alginat von Pseudomonas ssp. also mukoide Exoprodukte können phagozytäre Funktionen inhibieren, so dass damit trotz zirkulierender Phagozyten ein Defizit ihrer Funktion eintritt. Diese Substanzen würden dann die Aufnahme der Mikroorganismen inhibieren und häufig auch durch Wechselwirkung mit intrazellulären Strukturen (niedrigmolekulare G-Proteine, Aktin, respiratorischer Burst u.a.) die essentielle Funktion der Phagozyten blockieren. Die Kenntnis dieser multiplen Faktoren ist notwendig, um sich in der Zukunft Gedanken zu machen, wie die Pathophysiologie nach Kolonisierung eines Brandverletzten das Schicksal dieses Brandverletzten entscheiden kann. Es handelt sich 14 nicht um einen einzelnen mikrobiellen Faktor sondern um mehrere Faktoren; es ist nicht ein einzelnes Abwehrsystem sondern die Komplexität unterschiedlicher Mediatorkaskaden betroffen. Die Abwehrleistung des lokalen Wundfeldes wie auch der zirkulierenden Zellen sowie die Stabilität der natürlichen Immunität mit der Möglichkeit zum Aufbau der spezifischen Abwehr sind zu beachten. All diese Komponenten sind letztendlich entscheidend, ob die Kolonisierung und mikrobielle Infektion kompensiert wird oder ob es letztendlich zu einer mikrobiellen Invasion mit dem Tod des Patienten durch die „Burn wound sepsis“ kommt. Es sind also allgemeingültige Fragen, die beim Brandverletzten wie auch beim Polytrauma zum Tragen kommt. Voraussetzung für die Beantwortung dieser Fragen ist die exakte Kenntnis der pathophysiologischen Vorgänge, die durch mikrobielle Pathogenitätsfaktoren ausgelöst werden sowie die Analyse der unterschiedlichen zellbiologischen Reaktionsformen auf mikrobielle Reize. (Abb. 5) Nur aus dieser komplexen Sicht heraus ist es möglich Prädiktivmarker der „Burn wound sepsis“ einzuführen und innovative therapeutische Ansätze zu erarbeiten. Die hier darzustellenden Ergebnisse betreffen also 1. weiterführende Arbeiten zu den mikrobiellen Pathogenitätsfaktoren in der Wechselwirkung mit humanen Immuneffektorzellen und 2. Studien zu den zellbiologischen Auswirkungen, die für den „Immunstress“ und die „Immunparalyse“ verantwortlich sein können sowie 3. den Einsatz neuer therapeutischer Ansätze zur Stabilisierung der Infektabwehr und zur Behandlung der traumabedingten Immunparalyse Untersuchungen zu mikrobiellen Pathogenitätsfaktoren Untersuchungen zum Staphylokokken Leukozidin Im Rahmen der mikrobiellen Kolonisierung bei Brandverletzten sind Pseudomonas ssp. und Staphylokokken prädominante Keime, die häufig im Verbund bei der mikrobiellen Invasion gefunden werden. In früheren Arbeiten hatten wir gezeigt, dass die Pathogenität der Mikroorganismen im Laufe der mikrobiellen Invasion offensichtlich zunimmt. Die Frage stellte sich, inwieweit definierte Pathogenitätsfaktoren die mikrobielle Invasion begünstigen, und welche Parameter der natürlichen Infektabwehr dabei moduliert werden. Im Folgenden wurde in einer Zusammenarbeit mit dem Institut Bactériologie der Medizinischen Fakultät der Universität Straßburg das Staphylokokken Leukozidin analysiert. Staphylokokkus aureus Stämme sind mit einer Vielzahl von Erkrankungen assoziiert, wie z.B. Lungeninfektion, Verbrennungen, Furunkel und Dermonekrosen. Die Pathogenitätsfaktoren von Staph.aureus, insbesondere die Toxine, spielen dabei eine besondere Rolle für die Auslösung des Krankheitsprozesses. In früheren Studien konnte gezeigt werden, daß Staphylokokkentoxine (z.B. α-Toxin, δ-Toxin, Toxic Shock Syndrome Toxin, die Immuneffektorzellen von Probanden hinsichtlich der Mediatorfreisetzung modulieren. Im Folgenden wurde das Staphylokokken Leukozidin, ein Zweikomponententoxin, analysiert. Es wurde das Panton-ValentineLeukozidin (Luk PV) dem Leukozidin R gegenübergestellt. Diese Zweikompo15 nententoxine (S und F) interagieren synergistisch mit Leukozyten. Beide Toxine wurden von Staph.aureus V8 isoliert. Nur die gleichzeitige Interaktion von Fund S-Komponenten des Leukozidins entfaltet biologische Aktivitäten. Histologisch ist die Dermonekrose durch einen zentralen nekrotischen Herd sowie durch eine massive Infiltration von neutrophilen Granulozyten und Makrophagen charakterisiert. Im Folgenden wurde analysiert, inwieweit humane Immuneffektorzellen sich hinsichtlich der Reaktivität gegenüber Staphylokokken Leukozidin verhalten. Wir können zeigen, dass das Panton-Valentine-Leukozidin pathophysiologisch aktiver als das Luk R ist. Offenbar bestimmt die Zusammensetzung der Bikomponenten Systeme die Aktivität des Toxins. In kürzlichen Arbeiten haben wir zeigen können, dass durch Austausch und Modifikation der einzelnen Toxinkomponenten die biologische Aktivität signifikant moduliert wird. Humane Immuneffektorzellen entlassen nach Wechselwirkung mit Leukozidin das chemotaktisch aktive Leukotrien B4; basophile Granulozyten entlassen Histamin (Abb. 6, 7) und unter den Zytokinen wird das chemotaktisch aktive Interleukin-8 freigesetzt. (Abb. 8) Leukozidin wirkt auch an permeabilisierten Leukozyten. Dies ist insbesondere wichtig, da im Rahmen der Wechselwirkung eines Toxins mit humanen Effektorzellen diese häufig permeabilisiert sind und damit das Toxin in die Zelle gelangt. Offenbar reicht aber der enzymatische Apparat aus, um auch bei permeabilisierten Leukozyten eine Freisetzung von Lipidmediatoren zu induzieren. Insgesamt gesehen hat Leukozidin also einen ausgeprägten proinflammatorischen Einfluss; erst in hohen Konzentrationen kommt es zur Lyse der Immuneffektorzellen. Zunächst werden also Mediatoren der Entzündung freigesetzt. Ausschlaggebend ist, dass im Rahmen der Wechselwirkung mit Leukozidin Zellen das „Heatschockprotein 70“ exprimieren. Leukozidin induziert ebenfalls die Apoptose von Zielzellen. Diese Apoptose wird durch Einwirkung von Wachstumsfaktoren inhibiert. (Abb. 9) Unter dem Einfluss von GM-CSF wird die Leukozidinwechselwirkung mit Effektorzellen und die Freisetzung von Mediatoren massiv verstärkt. Dies bedeutet, dass in einem bereits entzündlichen Wundfeld die Besiedlung mit Staph.aureus und die Sekretion von Leukozidin zu einer Aufschaukelung des Entzündungsprozesses führen kann. Man kann sich also vorstellen, dass am Wundfeld vielfältige Zytokine freigesetzt werden. Diese inhibieren die Apoptose. Durch Bindung des Leukozidins kommt es an den so geschützten Zellen zu einer massiven Mediatorfreisetzung, die eine weiterführende Entzündung begünstigen und damit die mikrobielle Invasion unterstützen. Pathogenitätsfaktoren von Pseudomonas aeruginosa In früheren Arbeiten hatten wir auf die Bedeutung von Pseudomonas ssp. für die Kolonisierung und mikrobielle Invasion hingewiesen. Wir konnten zeigen, dass Pseudomonas aeruginosa-Stämme von Brandverletzten zwei Hämolysine sezernieren, d.h. ein hitzelabiles Hämolysin (Phospholipase C) und ein hitzestabiles Hämolysin, ein Glykolipid. Beide Hämolysine verhalten sich unterschiedlich hinsichtlich der Mediatorfreisetzung. (Abb. 10) Wir untersuchten wiederum an humanen Immuneffektorzellen (neutrophile Granulozyten, humane periphere mononukleäre Zellen) den Einfluss unterschiedlicher Pseudomonas aeruginosa16 Stämme. Bekanntlicherweise unterscheiden wir mukoide und weniger mukoide Stämme. In Anlehnung an die Wachstumsbedingungen wird das Mukopolysaccharid von Pseudomonas aeruginosa gebildet. In klassischer Weise finden wir sehr viel Mukopolysaccharid (Alginat) bei Pseudomonas aeruginosa-Stämmen von Patienten mit zystischer Fibrose. Grundsätzlich ist aber für Pseudomonas aeruginosa das Mukopolysaccharid ein wichtiger Pathogenitätsfaktor. Im Folgenden wurden klinische Isolate analysiert, die einerseits mukoid und weniger stark mukoid ausgeprägt sind. Wir überprüften an humanen Immuneffektorzellen, ob durch Wechselwirkung mit Pseudomonas aeruginosa es zur Freisetzung des chemotaktisch aktiven Interleukin-8 kommt. Bekanntlicherweise gehört dieses Zytokin zu den niedrigmolekularen Chemotaxinen und ist außerordentlich aktiv für die Akkumulation von Entzündungszellen und Adhäsinausprägung an Granulozyten und Zielzellen. In unseren Untersuchungen können wir zeigen, dass eine definierte Bakterienzellkonzentration vorliegen muss, um eine optimale Freisetzung von Interleukin-8 zu induzieren. Unsere Daten belegen, dass Pseudomonas aeruginosa Bakterien die Freisetzung von Il-8 aus Granulozyten und Monozyten induzieren; sie sind damit in der Lage die Entzündungsreaktion zu amplifizieren und zu perpetuieren, indem zusätzliche Phagozyten an den Entzündungsherd geholt werden. Die Freisetzung von Interleukin-8 wird dabei nicht durch Alginat blockiert. Die Befunde sind für die Betrachtung der mikrobiologischen Situation bei Brandverletzungen von außerordentlicher Bedeutung. Das Wundfeld ist nach ca. einer Woche mit unterschiedlichen Mikroorganismen kolonisiert; die klinische Erfahrung zeigt, dass die Kolonisierung mit Pseudomonas aeruginosa ein sehr schlechtes Zeichen darstellt, weil es häufig zur mikrobiellen Invasion kommt. Man könnte nun davon ausgehen, dass die Bakterien selber zunächst durch Pathogenitätsfaktoren die Granulozyten an den Ort der Auseinandersetzung chemotaktisch akkumulieren; damit wäre eine erste Abwehrlinie gewährleistet. Gleichzeitig werden aber durch die Wechselwirkung des Mikroorganismus mit den Phagozyten Granulozyten zerstört, es kommt zur Freisetzung von Proteasen wie natürlich auch zur Generierung von weiteren chemotaktischen Faktoren, so dass die Entzündung sich aufschaukelt. Eine aufgeschaukelte Entzündung könnte zu einer verminderten Immunabwehr im Rahmen der neutrophilen Phagozytenabwehr führen. Die mikrobielle Invasion würde trotz des Abrufens von Phagozyten damit begünstigt. Nicht nur Granulozyten sondern auch Thrombozyten sind Träger von Mediatoren. Auf der Suche nach relevanten Pathogenitätsfaktoren von Pseudomonas untersuchten wir die Bedeutung des hitzelabilen Hämolysins und eines weiteren Pathogenitätsfaktors, der Lipase. Die Phospholipasen sind ubiquitäre Enzyme. Die Aktivierung von Phospholipasen führt wiederum zur Freisetzung von ungesättigten Fettsäuren. Die individuellen Zellpopulationen können je nach ihrer Enzymausstattung die ungesättigten Fettsäuren in unterschiedliche Metabolite umwandeln. Im Folgenden zeigen wir, dass neben dem bekannten Exotoxin von Pseudomonas aeruginosa (d.h. Exotoxin A, das mukoide Polysaccharid) das hitzelabile Hämolysin von außerordentlich großer pathophysiologischer Bedeutung ist. Pseudomonas aeruginosa Stämme können zwei lipolytische Enzyme produzieren, d.h. die Phospholipase C und die Lipase. Beide Toxine werden extrazellulär sezerniert. Die Wechselwirkung mit Thrombozyten führt zu einer massiven 17 Freisetzung von 12-Hydroxyeicosatetraensäure (12 HETE). (Abb. 11) Dieser Metabolit ist in der Aktivierung weiterer Entzündungszellen außerordentlich reaktiv. Die hier vorliegenden Untersuchungen zeigen also sehr deutlich, dass nicht nur Pseudomonas aeruginosa als Mikroorganismus sondern die relevanten Pathogenitätsfaktoren für die Freisetzung der Entzündungsmediatoren von außerordentlicher Bedeutung sind. In weiterführenden Arbeiten konnten wir diese Ergebnisse nicht nur an Thrombozyten sondern auch an Granulozyten und Monozyten bestätigen. Die Phospholipase C in ihrer Wechselwirkung mit Lipase führt zur Generierung von chemotaktisch aktivem Leukotrien B4, von Sauerstoffmetaboliten , der Histaminfreisetzung aus Basophilen. Die Analyse der Interleukin-8Freisetzung führte zu überraschenden Ergebnissen. Die Phospholipase selber zeigt für sich nur eine geringe Aktivierung der Interleukin-8-Freisetzung, obwohl in hohem Maße das chemotaktisch aktive Leukotrien B4 gebildet wird. In höheren Konzentrationen kam es sogar zu einer Suppression der Phospholipase C induzierten Interleukin-8-Freisetzung. (Abb.12,13) Diese duale Funktion eines Toxins hinsichtlich der Mediatorfreisetzung hatten wir auch für das E.coli α-Hämolysin beobachtet. Bekanntlicherweise sind Hämolysin-produzierende E.coli-Stämme in der Wechselwirkung mit ihrem Zielorgan besonders aggressiv. Pathophysiologische Reaktionen wie Meningitis, Sepsis werden nicht nur durch spezifische Adhäsine von E.coli ausgelöst sondern auch durch E.coli zusammen mit der α-Hämolysinproduktion. Die Untersuchungen belegen also, dass nicht nur die taxonomische Definition eines Mikroorganismus entscheidend ist für das, was sich beim Brandverletzten abspielt sondern dass die individuellen Pathogenitätsfaktoren mit den Komponenten der natürlichen und spezifischen Infektabwehr interagieren. Untersuchungen zur Bedeutung mikrobieller Superantigene Als Superantigene bezeichnet man eine heterogene Gruppe von Proteinen, die einen äußerst wirksamen Mechanismus der T-Lymphozytenstimulation benutzen. Sie binden über MHCII Moleküle an antigenpräsentierenden Zellen und vernetzen variable Anteile des T-Zellrezeptors auf CD4+ und CD8+-T-Zellen. Obwohl es unterschiedliche Typen von Superantigenen gibt, die untereinander kaum verwandt sind, benutzen sie die gleichen molekularen Mechanismen. (Abb. 14 a-d) Im Rahmen ihrer Bindung werden sie nicht prozessiert, wie dies für konventionelle Antigene üblich ist. Ihre pathophysiologischen Auswirkungen sind offenbar durch die ungewöhnlichen Mechanismen der T-Lymphozytenstimulierung vermittelt. Der Prototyp der Superantigene, die von Infektionserregern gebildet werden, sind Staphylokokken Enterotoxine sowie die pyrogenen Exotoxine, die aus Staphylokokken und Streptokokken gebildet werden. Andere Mitglieder dieser Familie sind das „Toxic-Shock-Syndrom“-Toxin von Staph.aureus und das Streptokokken Superantigen von S.pyogenes. Einige Toxine haben untereinander eine Sequenzhomologie z.B. SEA und SEB zu mehr als 50% im Unterschied zu TSST-1. In Folge der außerordentlichen Wirksamkeit der Toxine kann ein geringer bakterieller Fokus bereits zu einer generellen Aktivierung des Immunsystems führen. Eine solche polyklonale T-Zell-Aktivierung kann zur Immunsuppression führen; 18 die Superantigene können Anergie oder Tod durch Apoptose an T-Zellen induzieren. Die Injektion von Staphylokokken Enterotoxin B in Mäusen führt zu einer partiellen Depletion von V8-T-Zellen. Die V8+ Zellen, die immer noch in der Milz vorliegen, antworten weder auf SEB noch auf T-Zellrezeptorantikörper. Die schockähnlichen Symptome, die durch Superantigene induziert werden, kommen durch die massive Freisetzung von Lymphokinen und Monokinen zustande. Ein kritischer Mediator ist wohl TNFα. Das TNF wird wahrscheinlich von T-Lymphozyten produziert. Superantigene können immunpathologische Phänomene sowie die Aktivierung von autoreaktiven B-Zellen wie z.B. Streptokokkenerkrankungen induzieren. In jüngster Zeit gibt es Hinweise, dass mikrobielle Superantigene auch für die Induktion und Persistenz der atopischen Dermatitis mitverantwortlich sein sollen. Im Folgenden untersuchten wir die Wechselwirkung von Toxic-Shock-Syndrom-Toxin und einiger Mutanten hinsichtlich der Mediatorfreisetzung aus humanen Immuneffektorzellen. Die mutierten TSST-Moleküle erhielten wir in der Zusammenarbeit mit Dr. Bonventre, USA. Diese Untersuchungen sind in sofern relevant weil man sich vorstellt, dass durch Veränderung von Superantigenen neue immuntherapeutische Anwendungen zu erreichen sind. Unsere Untersuchungen zeigen sehr deutlich, dass TSST-1 hinsichtlich der Freisetzung von unterschiedlichen Zytokinen (IL-6, TNF u.a.) hochaktiv ist und an wesentlichen Eckpunkten der natürlichen wie auch spezifischen Immunität Einfluss hat. Es gibt TSST-Mutanten, die diesen Einfluss nicht mehr haben. Die Frage stellt sich, inwieweit solche Mutanten in Zukunft für die Blockierung von Rezeptorstrukturen eingesetzt werden können, die durch mikrobielle Superantigene normalerweise okkupiert werden. Man könnte sich vorstellen, dass diese Mutanten sich binden und damit das natürliche Superantigen keine Bindungsstelle mehr hat und damit keine Reaktivität auslöst. (Abb.15 a) Weiterführende Arbeiten wurden in Zusammenarbeit mit Dr. Ulrich, NIH, USA, durchgeführt. Mit Dr. Ulrich wurden Mutanten des Staphylokokken Enterotoxin B (SEB) analysiert. (Abb. 15 b,c) Wir können zeigen, dass die Zytokinfreisetzung (TNF, IL-10, IL-12) von der Sekretion wie auch von der molekularbiologischen Expression durch diese Mutanten moduliert werden. In diesem Zusammenhang soll betont werden, dass Zytokine wie Interleukin-12 Hoffnungsträger sind, da sie die protektive Immunität offenbar erhöhen. Interleukin-12 induziert offenbar auch Interferon-γ. Interleukin-10 andererseits wirkt antiinflammatorisch aber begünstigt durch eine Stabilisierung des TH1-Zellpools die Aktivität eines TH2-Zellpools. Inwieweit durch eine verstärkte oder modulierte Interleukin-10-Freisetzung Zytokine freigesetzt werden, die eine Allergisierung favorisieren, kann an diesem Punkt nicht gesagt werden. Andererseits wissen wir heute dass Zytokine, die zu einer Allergisierung führen, wiederum potente inflammatorische Aktivität ausüben und im Rahmen der Immunsuppression offenbar eine große Rolle spielen. So findet man z.B. das Interleukin-4 nicht nur bei allergischen Prozessen sondern auch bei der Immunsuppression (HIV). Die hier vorgestellten Untersuchungen zeigen also sehr deutlich, dass pathogene Mikroorganismen ihrerseits über unterschiedliche Faktoren mit den Immuneffektorzellen in Wechselwirkung treten. Bei gramnegativen Bakterien spielt sicher das zellgebundene und lösliche Polysaccharid eine Rolle. Wenig bearbeitet wurden Strukturkomponenten der Mikroorganismen oder auch sezernierte Exopro19 dukte (Phospholipase, Lipasen u.a.). Das Zusammenwirken dieser Exoprodukte und Superantigene führt zu einer Modulation der Immunantwort. Für die künftige pathophysiologische Betrachtung von Schock und Sepsis wird man also nicht allein von dem klassischen Bild einer gramnegativen Sepsis (LPS) induzierten pathophysiologischen Veränderung ausgehen müssen, vielmehr wird man das Konzept vertreten müssen, dass mehrere Pathogenitätsfaktoren am Wirkort relevant sind und Immuneffektorzellen modulieren. Modulation der Immuneffektorreaktionen durch RSV (Respiratory Syncytial Virus). Virale Infektionen sind eine zusätzliche Komplikation bei immunsupprimierten Patienten; da RSV pathophysiologisch im frühen Lebensalter aber auch rekurrierend und rezidivierend bei Erwachsenen häufig der bakteriellen und respiratorischen Infektion vorausgeht, haben wir die Wechselwirkung von RSV mit bronchialen Epithelzellen und Immuneffektorzellen analysiert. Es wurde überprüft, ob die Wechselwirkung mit RSV Mediatoren der Entzündung entlässt. Dies ist in der Tat der Fall. Das chemotaktisch aktive Interleukin-8 wie auch der Tumornekrosefaktor (TNF) und lösliche Zytokinrezeptoren werden zeit- und dosisabhängig entlassen. Man muss sich also vorstellen, dass die Wechselwirkung des Epithels mit dem RSV zu einer Freisetzung von Entzündungsmediatoren führt. Diese sorgen für ein inflammatorisches Entzündungspotential. Diese Entzündungszellen können sich durch Aufregulation unterschiedlicher Adhäsine unterhalb des Epithels anlagern und innerhalb ihrer Aktivierung das Epithel durch weitere Mediatoren beeinflussen. RSV führt aber auch zu einer gezielten Freisetzung von Interleukin-10. (Abb. 16) Ein wichtiger Pathogenitätsfaktor ist hierbei das G-Protein. Mittels des isolierten G-Proteins konnten wir nachweisen, dass in der Tat das antiinflammatorische Zytokin Interleukin-10 freigesetzt wird. Man kann sich also vorstellen, dass bei schweren polytraumatischen Situationen (ARDS u.a.) Zytokine wie Interleukin-10 letztendlich ein TH2-“Pattern“ favorisieren. Dieses TH2-Zytokin-“Pattern“ konnte in der Tat im Rahmen der viralen Infektion nachgewiesen werden. Interleukin-5 führt z.B. zu einer Akkumulation von Eosinophilen. Diese Daten belegen und unterstreichen die Befunde, dass beim ARDS in der Lungenlavage es massiv zu einer Eosinophilie kommt. Die so aktivierten Eosinophilen führen wiederum mittels ihrer kationischen Proteine zu ernsten pulmonalen pathophysiologischen Veränderungen. Zusammenfassung: Anhand unterschiedlicher mikrobieller Pathogenitätsfaktoren haben wir zeigen können, dass neben Endotoxin wichtige Pathogenitätsfaktoren die Reaktivität von Immuneffektorzellen modulieren. Die Aktivierung von Entzündungsmediatoren ist eine wichtige Voraussetzung für die Balancierung der Infektabwehr. Eine überschießende Reaktion führt zu deletären Reaktionen. Eine persistierende Wechselwirkung von Pathogenitätsfaktoren mit humanen Immuneffektorzellen 20 führt zur Anergie und damit zur Immunparalyse. Diese Immunparalyse gibt der mikrobiellen Invasion Vorschub und dem Wachstum von opportunistischen Keimen. Es muss deshalb das Ziel sein, neben einer taxonomischen Beobachtung von Mikroorganismen an der Wundfläche die Pathologie der Entzündung durch unterschiedliche Pathogenitätsfaktoren verstehen zu lernen. Die Grundlagen eines besseren Verständisses werden dann zu innovativen therapeutischen Ansätzen führen sowie diagnostische Prädiktivmarker im Verlauf der Brandverletzung auch klinisch ermöglichen. Bedeutung von Pharmaka (Zytokine, lösliche Rezeptoren, Antagonisten u.a.) Unsere Untersuchungen haben dargelegt, dass die Wechselwirkungen unterschiedlicher Entzündungsmediatoren (Zytokine, Lipidmediatoren, zelluläre Adhäsionsmoleküle, lösliche Rezeptormoleküle u.a.) für die Balance der Infektabwehr wie aber auch für das Versagen der natürlichen und spezifischen Infektabwehr verantwortlich ist. Das Interleukin-4 als Wachstumsstimulationsfaktor hat eine herausragende Bedeutung. Interleukin-4 findet sich nicht nur als „Switch“Faktor sondern spielt auch im Rahmen der Suppression eine große Rolle. Die Inkubation von Interleukin-4 verändert das nachfolgende zell- und molekularbiologische Zytokinmuster nach Stimulation mit mikrobiellen Superantigenen. Interleukin-4 führt im Verlaufe der Wechselwirkung zu einer massiven Suppression. Daraus ergeben sich u.U. Möglichkeiten, durch Wegfangen von Interleukin-4 mittels mutierter Moleküle oder löslicher Il-4-Rezeptoren therapeutisch in vivo einmal eingreifen zu können. Anhand eines Brandverletzten wurden die Zellen ex vivo in vitro mit dem IL-4-Mutantenmolekül inkubiert und die Freisetzung von Mediatoren analysiert. (Abb.17) Auch hier eröffnen sich neue Perspektiven, die einer weiteren Analyse und Untersuchung bedürfen. Bedeutung von Wachstumsfaktoren: Die Wechselwirkung mikrobieller Exoprodukte führt nicht nur zur Aktivierung von Immuneffektorzellen sondern auch zum beschleunigten Zelltod, d.h. zur Apoptose. Im Rahmen dieser kurzen Lebenszeit kommt es zu einem Abknicken der Mediatorfreisetzung. Ein Versagen der natürlichen Infektabwehr ist die Folge. Die Zugabe von Wachstumsfaktoren könnte die Phagozyten stabilisieren. In Untersuchungen ex vivo in vitro zeigten wir, dass Toxine die Effektorzellen deaktivieren. Die Wechselwirkung von Effektorzellen mit Wachstumsfaktoren führt zu einer gesteigerten Freisetzung von Mediatoren. Hier liegt also nahe, dass u.U. Wachstumsfaktoren für therapeutische Ansätze von außergewöhnlicher Bedeutung sein könnten. Neben Zytokinen stellt sich die Frage, ob eine antientzündliche Therapie die zellbiologische Leistung moduliert. (Abb. 18) 21 Untersuchungen zum PAF-Rezeptorantagonisten Wir überprüften einen neuentwickelten PAF-Rezeptorantagonisten, der stereospezifisch mit dem PAF-Rezeptor interagiert sowie seinen Stereoisomer. Der Thrombozyten-aggregierende Faktor (PAF) aktiviert Immuneffektorzellen. Es kommt nach Wechselwirkung mit dem PAF-Rezeptorantagonisten zu einer zeitabhängigen Inhibition von Leukotrien B4 sowie auch Interleukin-8 aus PMN und LMB.(Abb.19,20) Diese Untersuchungen zeigen sehr deutlich, dass Substanzen, die Mediator-spezifisch ausgerichtet sind, aufgrund der Wechselwirkung mit bisher nicht bekannten zellbiologischen Strukturen die Freisetzung anderer Mediatoren moduliert. Der hier beschriebene PAF-Rezeptorantagonist könnte deshalb von außergewöhnlich großer Bedeutung sein, da Mediatoren abseits von PAF durch diese Substanz moduliert werden. Es bleibt abzuwarten, welche Bedeutung der PAF-Rezeptorantagonist für die Behandlung akuter sowie auch chronisch entzündlicher Reaktionen hat. In Anlehnung an die hier vorgestellten Daten überprüften wir auch einen H1-Rezeptorantagonisten, d.h. das Cetirizin. Das Cetirizin ist ein H1-Antagonist. Cetirizin moduliert humane Immuneffektorzellen. Es inhibiert die Freisetzung des chemotaktisch aktivem Leukotrien B4 wie auch des chemotaktisch aktiven Interleukin-8. Unsere Untersuchungen zeigen also sehr deutlich, dass ein klassisches Antihistaminikum offenbar abseits seiner bisherigen Spezifität (H-1) hinsichtlich der Modulation von Immuneffektorzellen außerordentlich wirksam ist. Zusammenfassung: Innovative therapeutische Ansätze können in der Entwicklung von Zytokinen, löslichen Zytokinrezeptoren oder auch Antizytokinen liegen. Die letztgenannte Therapie ist sicherlich sehr kostenaufwendig. Exakte Vorstellungen zu der Strukturwechselbeziehung eines Zytokins mit dem Rezeptor sind dagegen außerordentlich wichtig. Klassische oder neu zu entwickelnde Pharmaka könnten mit Rezeptorstrukturen von Zellen interagieren und damit die entzündliche Ausprägung von Immuneffektorzellen modulieren. In diesem Zusammenhang kommt also nicht nur Zytokinen, die zellulär protektiv oder Mediator neutralisierend sind, eine große Bedeutung zu sondern auch klassischen oder neu zu entwickelnden Pharmaka, die in einem breiteren Sinne entzündungsinhibierend sind ohne dass damit eine u.U. bestehende Immunsuppression weiterhin begünstigt wird. Zu bedenken ist hier, dass Mediatoren der Entzündung benötigt werden, um ein Gleichgewicht der Infektabwehr zu erzielen. Mediatoren haben also auch benefizielle Funktionen. Es ist notwendig dies Gleichgewicht herzustellen. 22 Therapeutische Interventionen bei der traumabedingten Immunparalyse Wie eingangs dargestellt, wurde das Projekt über zwei wesentliche Schienen bearbeitet: a) es wurden Arbeiten zum Einfluss mikrobieller Pathogenitätsfaktoren hinsichtlich der Freisetzung von Mediatoren der Entzündung mit der Fragestellung durchgeführt, über welche Mechanismen (Mediatoren, Zell-Zell-Interaktionen, Signaltransduktion, intrazelluläre Schaltmoleküle u.a.) die Immunparalyse zustande kommt. Aufgrund dieser Arbeiten wird es möglich sein, therapeutische Ansatzmöglichkeiten zu finden an denen man intervenieren kann. b) Ein zweiter wesentlicher Arbeitsbereich beschäftigte sich mit der Fragestellung, inwieweit man zum derzeitigen Zeitpunkt aufgrund des bisherigen Wissens die Trauma-induzierte Immunsuppression therapeutisch angehen kann. Wie bereits erwähnt, stehen bereits Zytokine oder lösliche Zytokinrezeptoren oder auch Antizytokine zur Verfügung. Im Folgenden wird über Arbeiten berichtet, die wir mit verschiedenen Universitätskliniken oder auch städtischen Krankenhäusern im Umfeld der RUB durchgeführt haben. Untersuchungen zum Einfluss von γ-Interferon beim Brandverletzten. Es handelte sich um eine Multicenterstudie, doppelblind und randomisiert sowie placebokontrolliert. Insgesamt sind 230 Patienten in Europa in diese Studie eingegangen. 12 der Patienten wurden von uns hinsichtlich detaillierter immunologischer Parameter analysiert. γ-Interferon (100 µg täglich subkutan) wurde den Patienten bis Tag 20 appliziert. An den Tagen 1, 3, 7, 14, 30 sowie 50 wurden folgende Parameter analysiert; d.h. HLADR und HLADQ – Expression sowie Mediatorfreisetzung aus Granulozyten nach Stimulation mit dem Kalzium-Ionophor A23187. (Abb. 21) Im Verlauf des posttraumatischen Geschehens sieht man initial eine Suppression von HLADR und HLADQ, die sich über die Zeit normalisiert; diese Normalisierung ist unabhängig von der Interferon-γ-Applikation. Allerdings ist dieser Verlauf zeitlich schneller in der Verumgruppe. Unter der Annahme, dass MHCIIMoleküle für die antigene Präsentation und damit für die Auslösung der natürlichen wie auch der spezifischen Immunität verantwortlich sind, ist es natürlich wünschenswert, dass im Rahmen der Interferon-γ-Therapie es zu einer schnelleren Aufregulation von MHCII tragenden Zellen kommt. Die kritische Frage stellt sich an diesem Punkt, ob die hier vorgenommene Therapie (Zeitpunkt, Konzentration) in Bezug zum Patiententrauma sowie zu entsprechenden Einschluss- und Ausschlusskriterien überhaupt relevant sein kann. Die Diskussion anlässlich einer intereuropäischen Konferenz zeigte, dass bezogen auf den klinischen Verlauf keine positiven Effekte auf die infektionsbedingte Mortalität zu sehen ist. Die Einschlusskriterien enthielten die Bedingung, dass Patienten mit mehr als 90%iger Verbrennung aufgenommen sowie alle Altersstufen mitberück23 sichtigt wurden. Kritisch muss man anmerken, dass hier natürlich rigide Eckpfeiler gesetzt sind. Es wäre wünschenswert, wenn eine Folgestudie im Rahmen einer Subgruppierung von Patienten (Verbrennungsgrad, Patientenalter) durchgeführt wird. Es bleibt abzuwarten, ob und wann eine solche Studie durchgeführt werden kann, da diese mit außerordentlich hohen Kosten verbunden ist. Immunmodulation – Untersuchungen zur Immunmodulation im Rahmen der Ernährungstherapie. Man weiß aus vielfältigen Untersuchungen, dass die Ernährung einen gravierenden Einfluss auf Parameter der natürlichen wie auch spezifischen Infektabwehr ausübt. In diesem Zusammenhang ist die Applikationen ungesättigter Fettsäuren im Austausch der C20:4 Arachidonsäure vielfältig beschrieben worden. Andere Möglichkeiten bestehen z.B. in der Applikation definierter Peptide, die u.U. stimulatorisch an unterschiedlichen Zellen der Infektabwehr wirksam sind. Im Folgenden ist eine Zusammenstellung unserer Daten gegeben: a) Einfluss von Eicosapentaensäure (enteral, parenteral) Es wurden in der enteralen Studie n=80 Patienten mit Impact® (Sandoz, Bern) therapiert. Es wurden präoperativ, postoperativ (Tage 1, 5, 10) die Gesamtleukozyten von Patienten nach großen Oberbauchoperationen für die Stunden analysiert. Es wurden die Zellen mit dem Kalzium-Ionophor und dem bakteriellen Peptid fMLP stimuliert; Messparameter waren die Leukotriensynthese über die „Reverse Phase“ HPLC sowie die Produktion des chemotaktisch aktiven Zytokins Interleukin-8. Bei den Resultaten lässt sich nachweisen, dass es zu einer signifikanten Verschiebung der biologisch hochaktiven Serie-4 Leukotriene zugunsten der biologisch weniger aktiven Serie-5 Leukotrienen kommt. (Abb. 22) Es findet sich kein signifikanter Unterschied bei Interleukin-8. Hinsichtlich der klinischen Validität der Therapie lassen sich die Aussagen an diesem Punkt noch nicht konkretisieren. Unter der Annahme, dass die Leukotriene der 4 Serie in der Regel im Rahmen des Entzündungsgeschehens, falls überschießend gebildet, eher schadensbringende Funktionen ausüben, wäre die Akkumulation des Leukotriens der 5er Serie benefiziell zu werten. Man sollte betonen, dass es nicht um eine völlige Elimination von Mediatorkaskaden sondern um eine Dämpfung proinflammatorischer Mediatorkaskaden geht. Bei der parenteralen Therapie wurden über das gleiche Abnahmeschema 80 Patienten nach großen chirurgischen Oberbaucheingriffen untersucht. Die Resultate der parenteralen Therapie sind mit denen der enteralen Therapie vergleichbar. Zusätzlich wurden die systemischen Zytokine (TNF, IL-10, IL-6) in dieser Studie untersucht. Es fällt auf, dass in der Verumgruppe der Trauma-induzierte Anstieg von TNF, IL-6 und IL-10 gedämpft wird. Wir haben in früheren Studien gezeigt, dass Interleukin-6 bei Brandverletzten irreversibel ansteigt in der Nicht-Überlebensgruppe. Wir müssen deshalb davon ausgehen, dass dem Interleukin-6 eine wichtige Rolle bezüglich des Abschaltens wichtiger zellulärer Funktionen zukommt, die im Rahmen der Infektabwehr notwendig sind. 24 Man kann sich natürlich auch vorstellen, dass die Akkumulation von „AkutePhase-Proteinen“ die Zellen so übermäßíg stimuliert, dass nach längerer Expositionszeit diese deaktiviert werden. Insofern kommt einer Dämpfung der Interleukin-6-Freisetzung u.U. eine große Bedeutung zu. Hinsichtlich des TNF bestehen verschiedene Vorstellungen. TNF als Mediator von Kachexie, Sepsis und Schock hat natürlich auch benefizielle Funktionen. An diesem Punkt muss noch einmal darauf hingewiesen werden, dass die Therapie zu einer Dämpfung der TNF-Freisetzung führt und nicht zu einer kompletten Elimination von TNF. Eine Elimination von TNF z.B. mit anti-TNF oder löslichen TNF-Rezeptoren führt zum Anstieg opportunistischer Infektionen. Hinsichtlich des Interleukin-10 ist dieser Befund sehr interessant. Interleukin-10 hat antiinflammatorische Aktivitäten und supprimiert das Zytokinprofil der T1-Zellen. Dieser Zellpool generiert bekanntlicherweise Interleukin-2 und das Interferon-γ. Aufgrund der Dämpfung des „T1-Zytokinpools“ kommt es zu einer relativen Favorisierung des T2-Zytokinpools. Es ist bekannt, dass Zytokine aus dem T2-Zytokinpool, z.B. Interleukin-4, im Rahmen der Immunsuppression offenbar wichtige induzierende Funktionen haben. Insofern kommt der Dämpfung der Interleukin10-Funktion ebenfalls eine wichtige Interpretation zu. Hinsichtlich des klinischen Verlaufes zeigen sich in der Verumgruppe weniger posttraumatische Infektionen als in der Placebogruppe. Weiterführende Studien müssen durchgeführt werden, um die Signifikanz der beobachteten Resultate herauszuarbeiten. b) Einfluss des Dipeptids (Ala-GLN) In Zusammenarbeit mit der Abteilung für Intensivmedizin und Anästhesiologie der Universitätsklinik – Marienhospital, Herne, wurden 60 Patienten hinsichtlich der klinischen Verträglichkeit des Dipeptids überprüft. Von diesen 60 Patienten wurden 10 Patienten immunologisch analysiert. Die Untersuchungen wurden präoperativ und postoperativ an den Tagen durchgeführt. Als Messparameter der zellulären Aktivität diente die Analyse der Leukotriene sowie die Analyse des chemotaktisch aktiven Zytokins Interleukin-8. Folgende Resultate wurden erhoben: es kam posttraumatisch zu einer Verminderung der Zysteinylleukotriene; in der Verumgruppe wurde die verminderte Synthese aufgehoben. (Abb. 23) Dies spricht dafür, dass das Dipeptid offenbar einen modulierenden Einfluss hinsichtlich der Leukotriensynthese ausübt. Unter der Annahme, dass Phagozyten mit ihrer Stoffwechselleistung dem Druck der mikrobiellen Auseinandersetzung entgegenwirken, ist die Fähigkeit des Dipeptids zu einer stimulatorischen Aktivierung von phagozytären Leistungen natürlich interessant. Es müssen weiterführende Arbeiten durchgeführt werden, inwieweit diese Stimulation wie wir auch ex vivo in vitro an Normalspenderzellen zwischenzeitlich haben belegen können, einen protektiven Effekt hinsichtlich der postoperativen Phase ausübt. c) Zusammenfassung Aus den vorgegebenen Resultaten wird ersichtlich, dass es einige Therapieschemata gibt, die im Rahmen der natürlichen wie auch spezifischen Immunität einen dämpfenden wie auch einen aktivierenden Einfluss an verschiedenen Parametern haben. Wie eingangs erwähnt, sind die unterschiedlichen Mediatorkaskaden miteinander vernetzt. Die Methoden sind notwendig, um die Infektabwehrlage zu 25 stabilisieren. Es wird wichtig sein, ein Überschießen von Mediatorkaskaden entweder zu dämpfen oder eine supprimierte Mediatorkaskade im Verlauf einer bleibenden Infektion zu aktivieren. Selbstverständlich sind dies Anfangsschritte, die im Verbund mit den basis-theoretischen Untersuchungen ihren Sinn machen. Weiterführende Untersuchungen sind unbedingt notwendig, um den Einfluss dieser Therapie hinsichtlich unterschiedlicher Parameter (Immunstimulation, Dämpfung von Mediatorkaskaden u.a.) weitergehend zu analysieren. 26 Erfolgskontrollbericht Die hier vorliegenden Ergebnisse haben einen wichtigen Beitrag zur Frage der mikrobiellen Pathogenität sowie von Pathogenitätsfaktoren in der Auslösung und Persistenz des septischen Schocks erbracht. Die Analyse zur Freisetzung der Entzündungsmediatoren, die möglichen Ansätze der Therapie zur Stabilisierung der Infektabwehr sind allesamt innovativ. Es muss daher abgewartet werden, inwieweit im Verlaufe weiterer Arbeiten und Förderungen sich diese therapeutischen Ansätze nicht nur als innovativ sondern auch als klinisch wirksam erweisen. Die Arbeiten zu den klinisch-theoretischen Grundlagen der mikrobiellen Pathogenität, der Beantwortung der zellbiologischen Leistung sowie der Signaltransduktion sind Grundlage dafür, um für die Diagnostik die Prädiktivmarker des drohenden Schocks zu erarbeiten. Insofern stellen die Ergebnisse einen wesentlichen Beitrag zu den förderpolitischen Zielen des Förderprogramms dar. Die Traumainduzierte Immunparalyse bei Brandverletzten ist ein wichtiger Grund dafür, dass trotz moderner Kenntnisse der Intensivtherapie heute immer noch ein wesentlicher Prozentsatz der so Geschädigten trotz moderner derzeitiger Therapiemöglichkeiten verstirbt. Hinsichtlich des wissenschaftlichen und technischen Erfolges konnten wir wesentliche Ergebnisse und Erkenntnisse sammeln. Sie betreffen die Wertigkeit unterschiedlicher mikrobieller Pathogenitätsfaktoren von Bakterien wie Pseudomonas ssp., Staphylokokken sowie E.coli; fernerhin die Bedeutung von mikrobiellen Exotoxinen sowie Superantigenen. In der Vergangenheit war den Exotoxinen eine wesentliche Bedeutung für die Pathophysiologie von Schock/Trauma zugesprochen worden. Heute zeigt sich aber immer mehr, dass neben den Endotoxinen mikrobielle Exotoxine und Superantigene gemeinsam ihren Einfluss an Immuneffektorzellen nehmen. Der eine oder andere pathophysiologische Aktivator kann in diesem Zusammenhang relevanter sein. Insofern ist es wichtig die unterschiedlichen Segmente und Aktivatoren zu kennen, um aus dem unterschiedlichen Verhalten hinsichtlich der zellbiologischen Ansprechbarkeit diagnostische Erfahrungen zu sammeln und diese in der Zukunft in die Praxis umzusetzen. Hinsichtlich des Finanzierungs- und Zeitplans wurde der vorgegebene Zeitrahmen eingehalten. Die Finanzierung war leider für das 1. Jahr gekürzt. An dieser Stelle sollte bemerkt werden, dass naturgemäß die theoretisch-klinischen Untersuchungen vom Untersucher her geplant und anhand der vorliegenden Ergebnisse kontinuierlich weiterentwickelt werden können. Hinsichtlich der Patientenstudien sind wir darauf angewiesen, dass in der Zusammenarbeit mit der Klinik das Patientenmaterial anfällt und zum anderen natürlich bei groß angelegten Studien, dass diese Studien auch innerhalb eines Zeitrahmens zur Durchführung und zum Abschluss kommen. Dies ist in der vergangenen Förderperiode durch einen intensiven Einsatz der Mitarbeiter und durch Zusammenarbeit mit unterschiedlichen Kliniken sehr erfolgreich durchgeführt worden. So konnten wichtige klinische Studien initiiert aber auch abgeschlossen werden. Diese Zusammenarbeit 27 umfasste die Kliniken in Bochum selber, das Universitätsklinikum Essen, die Städt. Krankenanstalten in Dortmund und auf der europäischen Ebene hinsichtlich der γ-Interferon-Studie eine von der Firma Boehringer konzipierte Studie zum Wirkmechanismus von γ-Interferon mit unterschiedlichen Verbrennungs- und Forschungszentren in Europa. Die bisherigen Ergebnisse sind akademischer Natur und sicherlich auch schon kliniksnah. Einzelne Therapieschemata (z.B. Einsatz von ungesättigten Fettsäuren u.a.) sind im Einsatz und werden sicherlich in der Zukunft für verschiedene Fragestellungen in der Klinik ihren Fuß fassen. Im Rahmen der bisherigen Arbeiten wurden keine Erfindungen in Anspruch genommen. Es sind keine Schutzrechtsanmeldungen oder erteilten Schutzrechte wie deren Verwertung geplant. Insgesamt lässt sich anhand der Publikationen und Abstrakts festhalten, dass im Rahmen der vorgegebenen Förderung über die drei Jahre wir wesentliche Beiträge haben erarbeiten können, die theoretisch-klinisch relevant und auch für die Pathophysiologie der Immunparalyse von ausserordentlich hohem Interesse sind. Innovative Ansätze zu neuen Therapiemaßnahmen wurden iniziiert, durchgeführt und evaluiert. Selbstverständlich ist das Krankheitsbild der Immunparalyse nur dann zu beherrschen, wenn man die modernsten Erkenntnisse der Grundlagenforschung einbezieht. In Anbetracht der Tatsache, dass das gesamte Feld der Immunologie (Zellbiologie und Molekularbiologie) außerordentlich wichtige Erkenntnisse gebracht hat, sollte das Projekt in den nächsten Jahren fortgeführt werden. Nur so lassen sich die hier gewonnenen Erkenntnisse verifizieren und für kliniksrelevante Fragestellungen übertragen. 28 Voraussichtlicher Nutzen, insbesondere Verwertbarkeit des Ergebnisses und der Erfahrungen Diagnostische Prädiktivmarker, insbesondere bei Patienten mit Polytrauma, Immunsuppression als Folge der Brandverletzung sowie innovative therapeutische Ansätze können nur dann realisiert werden, wenn man die Pathophysiologie mikrobieller Erkrankungen verstehen lernt. Von den hier erhobenen Befunden lassen sich folgende Aussagen verwerten: 1. die taxonomische Zuordnung der Mikroorganismen alleine ist nicht ausreichend, um Prädiktivmarker für den möglichen Ausgang bei der Brandverletzung zu erheben, 2. es erscheint notwendig, mikrobielle Isolate hinsichtlich relevanter Pathogenitätsfaktoren zu analysieren, um das Ausmaß der pathophysiologischen Wechselwirkung quantifizieren zu können. 3. der Einstrom unterschiedlicher mikrobieller Pathogenitätsfaktoren (intakte Mikroorganismen, Exotoxine, Endotoxine Superantigene) ist für das Bild der Mediatoraktivierung wie auch für das Fortschreiten der Immunparalyse verantwortlich, 4. innovative therapeutische Ansätze umfassen die Gabe von Zytokinen, löslichen Zytokinrezeptoren, Antizytokinen, antiinflammatorischen Substanzen sowie Immunmodulatoren, 6. bei der Komplexität der Mediatorkaskaden ist zu überlegen, ob es Angriffsziele gibt, die auf einer breiteren Basis die Immunität stabilisieren können. Die hier erarbeiteten Ergebnisse lassen sich allesamt bereits verwerten. Sicherlich wird man eine realistische Durchführung zu den einzelnen Punkten erst dann haben, wenn der komplexe Ablauf der Sepsis verstanden ist. Die vorliegenden Ergebnisse stellen aber die Grundlage dar, um zu diagnostischen Prädiktivmarkern wie auch therapeutischen innovativen Ansätzen zu gelangen. Zusammenfassend haben diese Untersuchungen unser Verständnis zur Pathophysiologie der Trauma-induzierten Immunsuppression insbesondere bei der Brandverletzung vertieft. Die gegenwärtigen modernen zellbiologischen Werkzeuge wurden eingesetzt, um die Mediatorgenerierung sowie Reaktivität von Immuneffektorzellen zu quantifizieren. Therapeutische Ansätze wurden ex vivo in vitro an humanen Immuneffektorzellen von Normalpersonen wie auch Polytraumatisierten und Brandverletzten durchgeführt, mit der Zielsetzung eine Reaktivierung der Effektorfunktion zu erhalten. Die Ergebnisse belegen, dass diese Reaktivierung möglich ist, jedoch abhängig vom Zeitpunkt der Zugabe eines entsprechenden Zytokins ist. Zielsetzung müsste es sein, durch Zugabe von zu entwickelnden Pharmaka oder auch Infusionsschemata eine Balancierung der Mediatorkaskaden zu erreichen, ohne unbedingt den einzelnen Mediator für sich spezifisch zu inhibieren. Es sollte nicht das Ziel sein, die gesamten Mediatorkaskaden zu inhibieren, da die Mediatoren essentielle Bestandteile der Infektabwehr sind. Im Wesentlichen würde dies für eine Immunmodulation im breiteren Sinne sprechen, 29 wobei sicherlich Eckpfeiler zu definieren sind, an denen man die Modulation quantifizieren kann oder den Ausgangspunkt der Modulation auch in Relation zum Krankheitsbild wird beurteilen können. Es besteht also nach wie vor dringender Forschungsbedarf, um die vielfältig anwachsenden Daten zur Pathophysiologie der Immunparalyse zu verwerten und im Sinne der Bearbeitung diagnostischer Prädiktivmarker und innovativer Therapieansätze zu gestalten. 30 Kurzfassung Entzündungsmediatoren spielen eine essentielle Rolle im Ablauf der natürlichen und spezifischen Immunität. Ein ausgewogenes Verhältnis der unterschiedlichen Kaskaden ist notwendig, um der mikrobiellen Infektion entgegenzuwirken. Überschießende Reaktionen führen zu Schock und Sepsis; ein Nichtansprechen der Mediatorkaskaden oder der zellulären Antwort auf unterschiedliche Mediatoren führen zu opportunistischen Infektionen. Die Komplexität der pathophysiologischen Vorgänge bei Brandverletzten ist trotz der therapeutischen Erfolge nach wie vor unbestritten. Die mikrobielle Kolonisierung ist sehr häufig durch eine mikrobielle Invasion gefolgt. Parameter seitens des Mikroorganismus, wie mikrobielle Pathogenität oder ein Abknicken der Immunität sind für die Brandverletztensepsis verantwortlich. Die modernen Erkenntnisse der Molekular- und Zellbiologie und insbesondere die Einsicht in die komplexen Abläufe der Mediator-gesteuerten Signaltransduktionswege hat zu neuen diagnostischen Prädiktivmarkern sowie innovativen Ansätzen geführt. In der vorliegenden Arbeit wird berichtet, dass die mikrobiellen Pathogenitätsfaktoren, die für die Aktivierung von Entzündungszellen verantwortlich sind, unterschiedlicher Natur sind. Neben LPS spielen mikrobielle Adhäsine, Exotoxine und die intrazelluläre Infektion eine herausragende Rolle. Unterschiedliche Pathogenitätsfaktoren können dabei Entzündungsmediatorkaskaden abrufen oder auch inaktivieren. Aus dieser dualen Funktion mikrobieller Pathogenitätsfaktoren erklärt sich, warum es im Verlauf einer Kolonisierung u.U. zu einem Abknicken der Infektabwehr kommt. Neben den mikrobiellen Exoprodukten spielen Superantigene von Staphylokokken, Streptokokken eine herausragende Rolle. Sie können durch Wechselwirkung mit dem T-Zellrezeptor und MHCII-Strukturen eine Vielfalt von Zytokinen aus humanen Zellen aktivieren, die zu Schock und Sepsis führen. Neue therapeutische Ansätze könnten darin bestehen diese Superantigene durch mutierte Moleküle zu ersetzen, um damit im Rahmen der progredienten Aktivierung von humanen Immuneffektorzellen eine therapeutische Gegenoffensive zu starten. Die weitreichenden Kenntnisse zur Molekularbiologie und Immunologie unterschiedlicher Zytokine erlauben es im Krankheitsverlauf u.U. mutierte Zytokine einzusetzen oder Zytokine anzuwenden, die bei der Immunparalyse zu einer Reaktivierung von Immuneffektorzellen führen. Ein wichtiges Phänomen ist dabei der selektive Zelltod (Apoptose), der zu einer Inbalance der natürlichen und spezifischen Infektabwehr führt. Diese Mechanismen können durch eine Antizytokintherapie ausbalanciert werden. Die vorliegende Arbeit hat also unsere Kenntnisse zu den mikrobiellen Pathogenitätsfaktoren, die zur Immunparalyse führen, weitgehend vertieft, bereichert und erfordert innovatives Denken. Es ist nicht nur wichtig die taxonomische Zuordnung des Erregers zu evaluieren, sondern die diagnostischen Tore zur Evaluierung der mikrobiellen Pathogenität zu öffnen. Aus den Überlegungen zur mikrobiellen Pathogenität lässt sich dann folgern, inwieweit der Druck der Pathogenitätsfaktoren auf das Immunsystem stattfindet. Die Evaluierung unterschiedlicher Mediatorkaskaden erlaubt es, den Status des Immunsystems zu analysieren. 31 Erste therapeutische Ansätze wurden in klinischen Studien von uns erbracht. In einer „Multicenterstudie“ wurde der Einfluss von 8-Interferon bei Brandverletzten analysiert. Obwohl sich immunologische Veränderungen zeigen, ist die Wirksamkeit der 8-Interferontherapie nach wie vor umstritten. Vertiefte Untersuchungen sind jedoch notwendig; eine Subgruppierung der Patienten ist angezeigt. Darüberhinaus wurde die Immunmodulation durch eine nutritive Therapie analysiert. Auch hier zeigen sich immunologische Veränderungen hinsichtlich der Lipidmediatoraktivierung und der Zytokinfreisetzung. Zusammenfassend kann man sagen, dass die Arbeiten neue Perspektiven zur mikrobiellen Pathogenität und ihren Einfluss auf die Mediatorkaskaden eröffnet hat, fernerhin, dass es möglich ist anhand der Zytokine Eckpfeiler der natürlichen und spezifischen Immunität zu erheben und fernerhin, dass weitere Forschung durchgeführt werden muss, um die nutritive Immunregulation oder pharmakologische sowie biotechnologische Modulation der Infektabwehr zu analysieren und in die Praxis umzusetzen. Es besteht also nach wie vor dringender Forschungsbedarf, um die traumabedingte Immunparalyse besser zu verstehen und dieser entgegenzuwirken. Die Ergebnisse sind notwendig, um die Pathophysiologie der Sepsis, insbesondere bei Brandverletzten zu verstehen. Mit den neu gewonnenen Erkenntnissen lassen sich neue Therapieansätze entwickeln. 32 Literatur Eigene Arbeiten Prévost, G.Supersac, Colin, D.A., Couppié,P., Sire,S., Hensler,T., Petiau,P., Meunier,O., Cribier,B., König,W., Piemont,Y The new family of leucotoxins from Staphylococcus aureus: Structural and biological properties. In: Bacterial Protein Toxins, Freer et al (Eds) Zbl. Bakt. Suppl. 24, Gustav Fischer Verlag, Stuttgart, Jena, New York, p 284-293, 1994 Köller, M. und König, W. Die Grundlagen der Verbrennungsinduzierten Immunsuppression in S. Vogt, G. Metzner, A. Haverich (Hrsg.) Wolfgang Pabst Verlag, Lengerich, 130-140, 1994 Arnold,R., Humbert,B., Werchau,H., Gallati,H., König,W. Interleukin-8, IL-6, and sTNF-RI release from a human pulmonary epithelial cell line (A549) exposed to respiratory syncytial virus (RSV) Immunology, 82, 126-133, 1994 König,B., Jaeger,K.-E., König, W. Induction of inflammatory mediator release (12-Hydroxyeicosatetraenoic Acid) from human platelets by Pseudomonas aeruginosa Int.Arch. of All.Appl.Immunol., 104, 33-41, 1994 Arnold, R., Werner,F., Humbert,B., Krebs,H., Werchau,H., König,W. Effect of respiratory syncytial virus – antibody complexes on cytokine (IL-8, IL-6, TNF-α) release and respiratory burst in human granulocytes Immunology, 82, 184-191, 1994 König,B. und König W. Effect of growth factors (GM-CSF; G-CSF) on the E.coli (α-hemolysin induced mediator release from human inflammatory cells. Infect. Immun., 62, 2085-2093, 1994 Hensler, T., König, B., Prévost, G., Piémont, Y., König W. LTB4-generation and DNA-fragmentation induced by leukocidin from Staphylococcus aureus – the protective role of GM-CSF and G-CSF on human neutrophils Infect. Immun., 62, 2529-2535, 1994 33 König,B., Ludwig, A., Goebel,W.,and König, W. Pore formation by the Escherichia coli -hemolysin: role for mediator release from human inflammatory cells. Infect. Immun., 4612-4617, 1994 König,B., Köller,M., Prevost,G., Piemont,Y.,Alou,J.E.Schreiner, A. König,W. Activation of human effector cells by different bacterial toxins leukocidin, alveolysin, erythrogenic toxin A): generation of interleukin-8 Infect. Immun., 4831-4837, 1994 Hensler, T., Köller,M., Prévost,G., Piémont,Y., König, W. GTP-binding proteins are involved in the activation and deactivation of human neutrophils by leukocidin from Staphylococcus aureus Infect. Immun., 62, 5281-5289, 1994 König,B., Prévost,G., Piémont,Y.,König,W. Effects of Staphylococcus aureus leukocidins on inflammatory mediator release from human granulocytes The Journal of Infectious Diseases, 3, 607-613, 1995 Drynda,A., König,B., Bonventre, P.F., König,W. Role of a carboxy-terminal site of TSST-1 in elicting immune response of peripheral blood cells Infect. Immun., 63, 1095-1101, 1995 König,B., Ceska,M., König, W. Effect of Pseudomonas Aeruginosa on Interleukin-8 Release from Human Phagocytes Int.Arch.All.Appl.Immunol.,106, 357-365, 1995 Arnold,R., Werchau,H., König,W. Increase of intercellular adhesion molecule-1 on human pulmonnary epithelial cells (A549) after respiratory syncytial virus (RSV) infection. in „Effectors and Pathogenesis of Allergic Diseases“ Int.Arch.All.Appl.Immunol., 107,392-393, 1995 Hilger,R.A., Köller,M., Fränken,J., Rihoux, J.P., König,W. Interactions of cytokines and lipid mediators in acute and chronic inflammation. in „Effectors and Pathogenesis of Allergic Diseases“ Int.Arch.All.Appl.Immunol., 107,383-384 1995 Hilger, R.A., Köller, M., König,W. Immunosuppressive agents enhance the cytokine-induced priming of inflammatory cells.in „Allergens, Immunotherapy and Management“ Int.Arch.All.Appl.Immunol., 107,259-260, 1995 34 Arnold,R., König,B., Gallati,H., Werchau,H., König,W. Cytokine (Il-8, Il-6, TNF-α and sTNFR-I release from human peripheral blood mononuclear cells after respiratory syncytial virus (RSV) infection. Immunology 85,324-372, 1995 Brom, J., Köller, M., Schlüter, B., Steinau, H. U., König, W., Expression of the Adhesion Molecule CD11b and Polymerization of Actin by Polymorphonuclear Granulocytes of Burned and Septic Patients. Burns, 21, 427-431, 1995 Wachtler,P., Hilger, R.A., König W., Bauer, K.H., Kemen, M., Köller, M., Influence of a preoperative enteral supplement on functional activities of peripheral leukocytes from patients with major surgery Clinical Nutrition, 14, 275-282, 1995 Köller, M., König, W. Interaction of bacteria with host defense cells:mechanisms of burn wound sepsis In: Wound Healing and Skin Physiology, Altmeyer et al. (Eds.) Springer-Verlag Berlin Heidelberg, S. 413-421, 1995 Hilger,R., Köller,M., König,W. Inhibition of leuktriene formation and IL-8 release by the PAF-receptor antagonist SM-12502 Inflammation 20,57-70,1996 Köller, M., Hilger, R.A., König, W. Effect of the PAF-receptor antagonist SM-12502 on human platelets Inflammation, 20, 71-85,1996 Köller,M., Hilger,R.A., Rihoux,J.P., König,W. Cetirizine demonstrates anti-inflammatory properties on human neutrophils – Evidence for the involvement of G-proteins Int.Arch.All.Appl.Immunol., 110: 52-56,1996 Shinoda,I., Takase,M., Fukuwatari,Y.,Shimamura,S., Köller,M., König,W Effects of Lactoferrin and Lactoferricin“ on the release of Interleukin 8 from human polymorphonuclear leukocytes. Biosci.Biotech.Biochem., 60(3): 521-523,1996 König, B., Streckert,H.-J., Krusat,T., König,W. Respiratory Syncytial-Virus G-protein modulates cytokine release from human peripheral blood mononuclear cells. J.Leukocyte Biol., 59, 403-408,1996 35 König,B., Jaeger, K.-E., Sage,A.E., Vasil,M.L., König,W. Role of P.aeruginosa lipase for inflammatory mediator release from human inflammatory effector cells (platelets, granulocytes, monocytes) Infect, Immun., 64, 3252-3258, 1996 König,B., Krusat,T., Streckert,H.J., König,W. Interleukin-8 release from human neutrophils by the Respiratory Syncytial Virus (RSV) is independent from viral replication.“ J.Leukocyte Biol., 60, 353-260, 1996 Arnold, R., König, W. (ICAM-1, LFA-3) expression and low molecular weight G protein activation of human bronchial epithelial cells (A549) infected with RSV J.Leukocyte Biol., 60, 766-771, 1996 König,B., Vasil, M.L., König,W. Role of hemolytic and nonhemolytic phospholipase C from Pseudomonas aeruginosa for IL-8 release from human monocytes. J.Med.Microbiol., 46, 471-478, 1997 König,B., Vasil,M.L., König,W. Role of hemolytic and nonhemolytic phospholipase C from Pseudomonas aeruginosa for inflammatory mediator release from human granulocytes Int. Arch.All.Appl.Immunol. 112, 115-124, 1997 Wachtler,P., König,W., Senkal,M., Kemen,M., Köller,M. A total parenteral nutrition enriched with ω-3 fatty acids modulates leukotriene – Synthesis of peripheral leukocytes and systemic Il-6, Il-10 and TNF-α levels in patients with major surgery. Journal of Trauma 42, 191-198, 1997 Arnold,R., König,W. Interleukin-8 release and low molecular weight G-Protein modulation in human neutrophils after phagocytosis of Listeria monocytogenes and Yersinia enterocolitica. J.Med.Microbiol., 47, 55-62, 1998 Köller,M., Wachtler,P., David,A., Muhr,G., König,W. Arachidonic acid induces DNA-fragmaentation in human polymorphonuclear neutrophil granolocytes Inflammation, 21, 463-474, 1997 König,B., Prévost,G., König,W. Composition of staphylococcal bi-component toxins determines pathophysiological reactions J.Med.Microbiol. 46, 479-485, 1997 36 Köller,M., König,W. Interaction of bacteria with host defense cells: mechanisms of burn wound sepsis. In: Wound Healing and Skin Physiology, Altmeyer et. al. (Eds.) Springer-Verlag Berlin-Heidelberg, S. 413-421, 1995 König,B., Jaeger,K.-E., Sage,A.E., Vasil,M.L., König,W. Role of P. aerugionosa lipase for inflammatory mediator release from human inflammatory effector cells (plate lets, granulocytes, monocytes) Infect. Immun., 64, 3252-3258, 1996 37 Die Autoren Wolfgang König Univ.-Professor Dr.med. Direktor des Instituts für Medizinische Mikrobiologie an der Otto-von-GuerickeUniversität Magdeburg, 39120 Magdeburg Manfred Köller Priv. Doz. Dr.rer.nat. ehemaliger Projektleiter der Abteilung Infektabwehrmechanismen für Medizinische Mikrobiologie der Ruhr-Universität Bochum, jetzt: Chirurgische Universitätsklinik, Bergmannsheil Bochum, 44789 Bochum Brigitte König Priv. Doz. Dr. rer. nat. Institut für Medizinische Mikrobiologie an der Otto-von-Guericke-Universität Magdeburg, 39120 Magdeburg Ralf Arnold Dr. rer. nat. Institut für Medizinische Mikrobiologie an der Otto-von-Guericke-Universität Magdeburg, 39120 Magdeburg Andreas Drynda Dr. rer. nat. Institut für Medizinische Mikrobiologie, Otto-von-Guericke-Universität Magdeburg, 39120 Magdeburg Paul Wachtler Dr. med. Marienhospital Herne, Medizinische Klinik I, Ruhr-Universität Bochum, 44625 Herne 39 Anhang Abbildung: 1 Die Abbildung zeigt die unterschiedlichen Wege der Infektabwehr auf; Neutrophile, Granulozyten und Makrophagen sind u.a. bestimmend für die Entzündungsreaktion. Die Wechselwirkung von dendritischen Zellen mit T-Zellen führt zur Einleitung der spezifischen Immunität; Makrophagen, „Natural Killer“Zellen sowie γδ-T-Zellen in der Mukosa sind zelluläre Elemente der natürlichen Immunität;TH2-Zellen in Wechselwirkung mit B-Zellen stimulieren die Antikörpersynthese; TH1-Zellen in Wechselwirkung mit Monozyten führen zur Freisetzung von Zytokinen und antibakteriellen Substanzen. 41 Abbildung: 2 Einfluss der Zytokine Zytokine bestimmen die Akkumulation von Zellen (Chemotaxis; die metabolische Aktivierung, die Proliferation, die Inhibition der Proliferation, die Differenzierung wie Zelltod (Apoptose) 42 Tabelle 1 Darstellung der Zytokinfamilien; dazu gehören die Interleukine, die zytotoxischen Zytokine, die koloniestimulierenden Faktoren, die Interferone, die Wachstumsfaktoren sowie die Chemokine 43 Tabelle 2 Zytokine mit proinflammatorischen Wirkungen 44 Tabelle 3 Zytokine mit antiinflammatorischen Wirkungen 45 Tabelle 4: Biologische Wirkung von Interleukin-1, Interleukin-6 und Tumornekrosefaktor 46 Abbildung: 3 Einfluss der T1- und T2-Zytokine auf Immunzellen. Neuroendokrine Faktoren (lokal, systemisch) wirken auf TH-0-Zellen und verändern das T1- sowie T2Zytokinmuster. Dies ist gemeinsam den Vorgängen bei Allergie wie auch Schock und Sepsis. 47 Abbildung: 4 a, b Einfluss mikrobieller Faktoren (Moduline) aus Immuneffektorzellen mit der Freisetzung von Zytokinen sowie der Modulation der zellulären Antwort. 48 Abbildung: 5 Einfluss von Bakterien und Toxinen sowie Virus auf Mastzellen, Neutrophile, Eosinophile. Es kommt zur Aktivierung der Zellen mit der Freisetzung multipler Zytokine und Mediatoren. Gleichzeitig werden diese Zellen geprimt. T-Lymphozyten werden ihrerseits wiederum programmiert. Es kommt zur Freisetzung vielfältiger Mediatoren durch die Wechselwirkung mikrobieller Exoprodukte mit Zellen der Infektabwehr. Diese entfalten chemotaktische Aktivitäten und sind verantwortlich für das mikrobielle „Killing“, führen zur Mediatorfreisetzung und der Freisetzung von Proteasen. Vorgänge der akuten und chronischen Entzündung werden dadurch reguliert. 49 Abbildung: 6 Histaminfreisetzung aus humanen Basophilen durch Luk-PV und Luk-R Eine humane Zellsuspension (LMB) (5x106) wurde mit Luk-PV oder Luk-R in den angegebenen Konzentrationen (0-, 5-, 50-, 500-, 1000-, 2000-, 3000 ng) für 1h bei 37°C inkubiert. Formylmethionylleucylphenylalanine (FMLP, 10-4M) diente als Positivkontrolle. Die Daten entsprechen den Mittelwerten von 3 unterschiedlichen Experimenten. * signifikanter Unterschied gegenüber Luk-R. 50 Abbildung: 7 Leukotrien B4 Freisetzung aus humanen Neutrophilen durch Luk-PV und Luk-R Humane PMN (107/500µl) wurden mit Luk-PV oder Luk-R in den angegebenen Konzentrationen (0-, 5-, 50-, 500-, 1000-, 2000-, 3000 ng) für 1h bei 37°C inkubiert. Formylmethionylleucylphenylalanine (FMLP, 10-4M) diente als Positivkontrolle. Die Daten entsprechen dem Gesamt-LTB4 (LTB4 + 20-COOH-LTB4 + OHLTB4) und sind Mittelwerte von 3 unterschiedlichen Experimenten. * signifikanter Unterschied gegenüber Luk-R. 51 Abbildung: 8 Interleukin-8 Freisetzung aus humanen Neutrophilen durch Luk-PV und Luk-R Humane PMN (107/500µl) wurden mit Luk-PV oder Luk-R in den angegebenen Konzentrationen (0-, 5-, 50-, 500-, 1000-, 2000ng) für 1h bei 37°C inkubiert. Die Daten sind Mittelwerte von 8 unterschiedlichen Experimenten. * signifikanter Unterschied gegenüber Luk-R. 52 Abbildung: 9 Zellbiologische Vorgänge zur Einleitung des Zelltodes (Apoptose) Dieser kann über pharmakologische, immunologische Aktivierung (T-Zellrezeptor) sowie Fas-ligand eingeleitet werden. Zytokine führen zur Aktivierung einer Sphingomyelinase mit der Freisetzung von intrazytoplasmatischem Ceramid; es kommt zum oxidativen Stress. Kortikoide führen ebenfalls zur gesteigerten Apoptose. Metabolite der Arachidonsäure und der Eicosapentaensäure regulieren und verzögern die Apoptose. 53 Abbildung: 10 Einwirkung von Pseudomonas aeruginosa Kulturüberstandsfraktionen auf die Interleukin-8-Freisetzung aus humanen Monozyten Humane Monozyten (4x105) wurden stimuliert mit konzentrierten (1:1) oder mit Verdünnungen (1:5, 1:100) der unterschiedlichen Kulturüberstandsfraktion; die Molekulargewichts „cut off“– Punkte belaufen sich auf >50.000; 30.000 – 50.000, 5.000-30.000; und unter 5.000 Da der entsprechenden Pseudomonas aeruginosa-Stämme (PAO1; PAO1∆SR, PAO1∆SRN). Die Kulturüberstände wurden isoliert nach 24 h und analysiert hinsichtlich von Interleukin-8 durch ELISA (Triplikatbestimmung). Die Ergebnisse zeigen Mittelwerte und Standardabweichungen aus drei unabhängigen Experimenten. *wesentlicher Unterschied im Vergleich zu unstimulierten Zellen; p<0,05 54 Pseudomonas aeruginosa Stämme Phospholipase-Typ PAO1 wild type hämolytisch, nicht-hämolytisch PAO∆SR nicht-hämolytisch PAO PLCN° hämolytisch PAO1∆SRN keine 55 Abbildung: 11 Einfluss von gereinigter Pseudomonas aeruginosa Lipase auf die Phospholipase C (Pseudomonas aeruginosa, C.perfringens) induzierte 12-HETE-Freisetzung Thrombozyten (108/500µl) wurden mit gereinigter Phospholipase inkubiert (P.aeruginosa, C. perfringens; 10 U) in der Anwesenheit von gereinigter Pseudomonas aeruginosa Lipase (0-, 0.43-, 4.3-, 108-, 216-, 433 U). Die Inkubation wurde über 60 Min. bei 37° C durchgeführt; die Werte für die unstimulierten Zellen wurden von den anderen Werten für die 12-HETE Bildung subtrahiert. *sind hier Unterschiede im Vergleich zu Phospholipase C stimulierten Thrombozyten in der Abwesenheit von Pseudomonas aeruginosa Lipase (p< 0,05). Ergebnisse sind Mittelwerte ± Standardabweichung (Balken) von drei unabhängigen Experimenten. 56 Abbildung: 12 Wirkung von gereinigter Pseudomonas aeruginosa Lipase auf die Phospholipase induzierte Leukotrien B4-Freisetzung aus humanen neutrophilen Granulozyten. Neutrophile (107/500µl) wurden mit gereinigter Phospholipase C (P.aeruginosa: 10 U; C.perfringens: 10 U) in der Anwesenheit von gereinigter Pseudomonas aeruginosa Lipase inkubiert. Die Inkubation lief über 30 Min. bei 37° C. Die Werte der unstimulierten Zellen wurden subtrahiert von den anderen Werten für die LTB4-Bildung. Die Ergebnisse sind Mittelwerte ± Standardabweichung (Balken) von drei unabhängigen Experimenten *signifikante Unterschiede im Vergleich zur Stimulation in der Abwesenheit von Lipase, (p< 0,05). 57 Abbildung: 13 Einwirkung von gereinigte hämolytischer Pseudomonas aeruginosa Phospholipase C auf die Interleukin-8-Freisetzung (ng/ml) aus humanen Monozyten. Humane Monozyten (4x105)/ml wurden mit gereinigter Phospholipase C von P.aeruginosa zu den angezeigten Konzentrationen über 24 h stimuliert. Die Ergebnisse zeigen die Standardwerte ± von drei unabhängigen Experimenten. *signifikante Unterschied im Vergleich zu unstimulierten humanen Monozyten (0 U PLC)(p<0,05) 58 Abbildung: 14 Zytokinfreisetzung (TNF-alpha, IL-6, IL-10, IL-12) aus humanen PBMC durch Superantigene Humane PBMC (1x106/ml) wurden mit den unterschiedlichen bakteriellen Superantigenen (SEB; TSST-1, ETA) in einer Konzentration von 10ng/ml für 24-, 48-, 72-, 96-, 120-, und 144h stimuliert. Die Überstände wurden mittels zytokinspezifischer ELISAs auf den Gehalt von TNF-alpha, IL-6, IL-10, IL-12 hin untersucht. Es ist ein repräsentatives Experiment gezeigt. 59 Abbildung: 15 a Exemplarische Darstellung zum Wirkmechanismus mikrobieller Superantigene und ihrer Mutanten. 60 Abbildung: 15 b TNF-alpha Freisetzung aus humanen PBMC durch SEB und SEB-Mutanten Humane PBMC (1x106/ml) wurden mit SEB oder SEB-Mutanten für 24-, 48-, und 72h bei 37°C stimuliert. Die Überstände wurden mittels ELISA auf ihren TNF-alpha Gehalt untersucht. 61 Abbildung: 15 c TNF-alpha Freisetzung aus humanen PBMC durch SEA und SEA-Mutanten Humane PBMC (1x106/ml) wurden mit SEA oder einer SEA-Mutante für 24-, 48-, und 72h bei 37°C stimuliert. Die Überstände wurden mittels ELISA auf ihren TNF-alpha Gehalt untersucht. 62 Abbildung: 16 Dosis- und zeitabhängiger Einfluss des gereinigten G-Proteins von RSV auf die Interleukin-10-Freisetzung aus humanen PBMC. Humane PBMC (1x106/ml) mit unterschiedlichen Konzentrationen von G-Protein (von 0 – 10 µg/106 PBMC) über 2-, 4-, 6-, 12-, 18-, 24 Stunden behandelt. Die ELISA-Werte sind Mittelwerte + Standardabweichung von drei unabhängigen Experimenten. IL-10-Spiegel sind signifikant verschieden (p< 0,05) von unbehandelten PBMC (p< 0,05) 63 Abbildung: 17 Einfluss einer teilaktiven IL-4-Mutante auf die Zytokinsynthese aus LMBs eines schwerbrandverletzten Patienten 1 x 106 LMBs/ml RMI 1640 wurden für 3 Tage in Abwesenheit oder Anwesenheit einer teilaktiven IL-4-Mutante (Y124 D) in Kultur genommen. Anschließend wurde die spontane IL-6 Freisetzung mittels ELISA quantifiziert. Patient: 39 J., männl., 35 % 2.° + 3.° Verbr., zusätzliches Polytrauma 64 Abbildung: 18 DNA-Fragmentierung in neutrophilen Granulozyten. 5 x 106 Zellen/1ml RPMI 1640 wurden für 18 h in Kultur genommen. A: Medium Kontrolle, B: in Anwesenheit von 40 µM Arachidonsäure, C: in Anwesenheit von 1µM MK 886 (Leukotien-Synthese-Inhibitor), D: in Anwesenheit von 1µM Indomethacin (Prostaglan-dinsynthese-Inhibitor). Anschließend wurden die Zellen mit Propidium-Jodid markiert und über FACS analysiert. 65 Abbildung: 19 a,b,c Einfluss von SM-12502 auf die Freisetzung von Leukotrienen aus neutrophilen Granulozyten. 1 x 107 Zellen/500µl PBS wurden mit verschiedenen Konzentrationen von SM12502 präinkubiert (20 min. 37° C in Anwesenheit von CA2+/Mg2+ (1mM/0,5 mM) und anschließend mit dem Ca-Ionophor A23187 (A, 5µM), fMLP (B) 1µM in Anwesenheit von 1µM Cytochalasin B) und mit NaF (C, 20mM in Anwesenheit von 10µM AlCl3) stimuliert. Die freigesetzten Leukotriene wurden über RPHPLC quantifiziert. 66 Abbildung: 20 a,b Einfluss von SM-12502 auf die Freisetzung von IL-8 aus peripheren Leukozyten Periphere Ganulozyten (A) oder Lymphozyten/Monozyten (B) wurden mit unterschiedlichen Konzentrationen von SM-12502 für 90 Min. in Anwesenheit von 1mM Ca++ und 0,5 mM Mg++ in PBS inkubiert. Die Freisetzung von IL-8 wurde anschließend im ELISA gemessen. 67 Abbildung: 21 Synthese von LTB4 aus Gesamtleukozyten schwerbrandverletzter Patienten (n=12) in Korrelation zur HLA-DR-Expression auf Monozyten. 1x 107 Gesamtleukozyten wurden mit dem Ca-Ionophor A23187 (5µM) in Anwesenheit von 1mM Mg stimuliert. Die gebildeten Leukotriene wurden über RPHPLC analysiert. Die Expression von HLA-DR auf Monozyten wurde mittels FACS-Analytik quantifiziert. Control: Placebogruppe Verum: IFN-γ 100µg sc. täglich 68 Abbildung: 22 Verhältnis der LTB4/LTB5-Mediatoren nach vollständiger parenteraler Ernährung (TPN) postoperativer Patienten1x107 Gesamtleukozyten/500µl PBS wurden mit dem Ca-Ionophor A23187 (5µM) in Anwesenheit von 1mM Ca und 0,8 mM Magnesium stimuliert. Die gebildeten Leukotriene wurden über RP-HPLC quantifiziert. Control: Kontrollgruppe 20 Patienten ohne Ω-3-Fettsäure-Supplementierung Verum: Verumgruppe 20 Patienten mit Ω-3-Fettsäuren-Supplementierung, p< 0.0001 69 Abbildung: 23 Einfluss eines Dipeptides AlaGln (Alanyl-Glutamin) auf die Synthesekapazität für LTE4 aus LMBs. 1 X 107 LMBs/500 µl PBS wurden mit unterschiedlichen Konzentrationen von AlaGln (Dipeptamin“, Fresenius) 15 Min präinkubiert; anschließend wurden die Zellen mit dem Ca-Iononphor A23187 (5µM) unter Zugabe von 1 µM Ca und 0,5 mM Magnesium stimuliert. Die freigesetzten Leukotriene wurden mittels RPHPLC quantifiziert. 70 Zivilschutz-Forschung, Neue Folge Schriftenreihe der Schutzkommission beim Bundesminister des Innern Herausgegeben vom Bundesverwaltungsamt – Zentralstelle für Zivilschutz – ISSN 0343-5164 im Auftrag des Bundesministeriums des Innern Band 50 – in Vorbereitung – R. Zech Entgiftung von Organophosphaten durch Phosphorylphosphatasen und Ethanolamin 2002, Broschur Band 49 – in Vorbereitung – G. Matz, A. Schillings, P. Rechenbach Task Force für die Schnellanalytik bei großen Chemieunfällen und Bränden 2002, Broschur Band 48 Zweiter Gefahrenbericht der Schutzkommission beim Bundesminister des Innern Bericht über mögliche Gefahren für die Bevölkerung bei Großkatastrophen und im Verteidigungsfall 2001, 92 Seiten, Broschur Band 47 J. Rasche, A. Schmidt, S. Schneider, S. Waldtmann Organisation der Ernährungsnotfallvorsorge 2002, 88 Seiten, Broschur Band 46 F. Gehbauer, S. Hirschberger, M. Markus Methoden der Bergung Verschütteter aus zerstörten Gebäuden 2002, Broschur Band 45 V. Held Technologische Möglichkeiten einer möglichst frühzeitigen Warnung der Bevölkerung – Kurzfassung – 2001, 144 Seiten, Broschur Band 44 E. Pfenninger, D. Hauber Medizinische Versorgung beim Massenanfall Verletzter bei Chemikalienfreisetzung 2002, Broschur Band 43 D. Ungerer, U. Morgenroth Empirisch-psychologische Analyse des menschlischen Fehlverhaltens in Gefahrensituationen und seine verursachende und modifizierenden Bedingungen sowie von Möglichkeiten zur Reduktion des Fehlverhaltens 2002, 300 Seiten, Broschur Band 42 45., 46. und 48. Jahrestagung der Schutzkommission beim Bundesminister des Innern – Vorträge – 2000, 344 Seiten, Broschur Band 41 W. König, M. Köller Einfluss von Zytokinen und Lipidmediatoren auf die Kontrolle und Regulation spezifischer Infektabwehr bei Brandverletzung 2002, 76 Seiten, Broschur Band 40 Institut der Feuerwehr Sachsen-Anhalt Entwicklung von Dekontaminationsmitteln und -verfahren bei Austritt von Industriechemikalien 2002, Broschur Band 39 TÜV Energie und Umwelt GmbH Optimierung des Schutzes vor luftgetragenen Schadstoffen in Wohngebäuden 2002, Broschur Band 38 W. Kaiser, M. Schindler Rechnergestütztes Beratungssystem für das Krisenmanagement bei chemischen Unfällen (DISMA®) 1999, 156 Seiten, Broschur Band 37 – in Vorbereitung – K.-J. Kohl, M. Kutz Entwicklung von Verfahren zur Abschätzung der gesundheitlichen Folgen von Großbränden 71 Band 36 M. Weiss, B. Fischer, U. Plappert und T. M. Fliedner Biologische Indikatoren für die Beurteilung multifaktorieller Beanspruchung Experimentelle, klinische und systemtechnische Untersuchung 1998, 104 Seiten, Broschur Band 35 K.Amman, A.-N. Kausch, A. Pasternack, J. Schlobohm, G. Bresser, P. Eulenburg Untersuchung der Praxisanforderungen an Atemund Körperschutzausstattung zur Bekämpfung von Chemieunfällen 2002, Broschur Band 34 W. Heudorfer Untersuchung der Wirksamkeit von Selbstschutzausstattung bei Chemieunfällen 1998, Broschur Band 33 J. Bernhardt, J. Haus, G. Hermann, G. Lasnitschka, G. Mahr, A. Scharmann Laserspektrometrischer Nachweis von Strontiumnukliden 1998, 128 Seiten, Broschur Band 32 G. Müller Kriterien für Evakuierungsempfehlungen bei Chemiekalienfreisetzungen 1998, 244 Seiten + Faltkarte, Broschur Band 31 G. Schallehn und H. Brandis Beiträge zur Isolierung und Indentifizierung von Clostridium sp. und Bacillus sp. sowie zum Nachweis deren Toxine 1998, 80 Seiten, Broschur Band 30 G. Matz Untersuchung der Praxisanforderung an die Analytik bei der Bekämpfung großer Chemieunfälle 1998, 192 Seiten, Broschur Band 29 D. Hesel, H. Kopp und U. Roller Erfahrungen aus Abwehrmaßnahmen bei chemischen Unfällen 1997, 152 Seiten, Broschur 72 Band 28 R. Zech Wirkungen von Organophosphaten 1997, 110 Seiten, Broschur Band 27 G. Ruhrmann, M. Kohring Staatliche Risikokommunikation bei Katastrophen Informationspolitik und Akzeptanz 1996, 207 Seiten, Broschur Band 26 43. und 44. Jahrestagung der Schutzkommission beim Bundesminister des Innern – Vorträge – 1997, 326 Seiten, Broschur Band 25 K. Buff, H. Greim Abschätzung der gesundheitlichen Folgen von Großbränden – Literaturstudie – Teilbereich Toxikologie 1997, 138 Seiten, Broschur Band 24 42. Jahrestagung der Schutzkommission beim Bundesminister des Innern – Vorträge – 1996, 205 Seiten, Broschur Band 23 K. Haberer, U. Böttcher Das Verhalten von Umweltchemikalien in Boden und Grundwasser 1996, 235 Seiten, Broschur Band 22 B. Gloebel, C. Graf Inkorporationsverminderung für radioaktive Stoffe im Katastrophenfall 1996, 206 Seiten, Broschur Band 21 Arbeiten aus dem Fachausschuß III: Strahlenwirkungen – Diagnostik und Therapie 1996, 135 Seiten, Broschur Band 20 Arbeiten aus dem Fachausschuß V I. – D. Henschler: Langzeitwirkungen phosphororganischer Verbindungen II. – H. Becht: Die zellvermittelte typübergreifende Immunantwort nach Infektion mit dem Influenzavirus III. – F. Hoffmann, F. Vetterlein, G. Schmidt; Die Bedeutung vasculärer Reaktionen beim akuten Nierenversagen nach großen Weichteilverletzungen (Crush-Niere) 1996, 127 Seiten, Broschur Band 19 Radioaktive Strahlungen I. – B. Kromer unter Mitarbeit von K. O. Münnich, W. Weiss und M. Zähringer: Nuklidspezifische Kontaminationserfassung II. – G. Hehn: Datenaufbereitung für den Notfallschutz 1996, 164 Seiten, Broschur Band 18 L. Clausen, W.R. Dombrowsky, R.L.F. Strangmeier Deutsche Regelsysteme Vernetzungen und Integrationsdefizite bei der Erstellung des öffentlichen Gutes, Zivil- und Katastrophenschutz in Europa 1996, 130 Seiten, Broschur Band 17 41. Jahrestagung der Schutzkommission beim Bundesminister des Innern – Vorträge – 1996, 197 Seiten, Broschur Band 16 F. E. Müller, W. König, M. Köller Einfluß von Lipidmediatoren auf die Pathophysiologie der Verbrennungskrankheit 1993, 42 Seiten, Broschur Band 15 Beiträge zur dezentralen Trinkwasserversorgung in Notfällen Teil II: K. Haberer und M. Drews 1. Einfache organische Analysenmethoden 2. Einfache Aufbereitungsverfahren 1993, 144 Seiten, Broschur Band 14 Beiträge zu Strahlenschäden und Strahlenkrankheiten I. – H. Schüßler: Strahleninduzierte Veränderungen an Säugetierzellen als Basis für die somatischen Strahlenschäden II. – K. H. von Wangenheim, H.-P. Peterson, L.E. Feinendegen: Hämopoeseschaden, Therapieeffekte und Erholung III. – T. M. Fliedner, W. Nothdurft: Präklinische Untersuchungen zur Beschleunigung der Erholungsvorgänge in der Blutzellbildung nach Strahleneinwirkung durch Beeinflussung von Regulationsmechanismen IV. – G. B. Gerber: Radionuklid Transfer 1993, 268 Seiten, Broschur Band 13 H. Mönig, W. Oehlert, M. Oehlert, G. Konermann Modifikation der Strahlenwirkung und ihre Folgen für die Leber 1993, 90 Seiten, Broschur Band 12 Biologische Dosimetrie I. – H. Mönig, Wolfgang Pohlit, Ernst Ludwig Sattler: Einleitung: Dosisabschätzung mit Hilfe der Biologischen Dosimetrie II. – Hans Joachim Egner et al.: Ermittlung der Strahlenexposition aus Messungen an Retikulozyten III. – Hans Mönig, Gerhard Konermann: Strahlenbedingte Änderung der Chemilumineszenz von Granulozyten als biologischer Dosisindikator IV. – Paul Bidon et al.: Zellmembranänderungen als biologische Dosisindikatoren. Strahleninduzierte Membranänderung im subletalen Bereich. Immunbindungsreaktionen an Lymphozyten 1993, 206 Seiten, Broschur Band 11 Beiträge zur Katastrophenmedizin vergriffen Band 10 W. R. Dombrowsky Bürgerkonzeptionierter Zivil- und Katastrophenschutz Das Konzept einer Planungszelle Zivil- und Katastrophenschutz 1992, 79 Seiten, Broschur Band 9 vergriffen 39. und 40. Jahrestagung der Schutzkommission beim Bundesminister des Innern Band 8 vergriffen Beiträge zur dezentralen Trinkwasserversorgung in Notfällen Teil I: K. Haberer und U. Stürzer Band 7 E. Pfenninger und F. W. Ahnefeld Das Schädel-Hirn-Trauma vergriffen 73 Band 6 O. Messerschmidt und A. Bitter Neutronenschäden vergriffen Band 5 vergriffen R. E. Grillmaier und F. Kettenbaum Strahlenexposition durch Ingestion von radioaktiv kontaminiertem Trinkwasser Band 4 vergriffen W. R. Dombrowsky Computereinsatz im Zivil- und Katastrophenschutz Band 3 B. Lommler, E. Pitt, A. Scharmann und R. Simmer Der Nachweis schneller Neutronen in der Katastrophendosimetrie mit Hilfe von Ausweisen aus Plastikmaterial 1990, 66 Seiten, Broschur Band 2 vergriffen Gammastrahlung aus radioaktivem Niederschlag Berechnung von Schutzfaktoren Band 1 vergriffen L. Clausen und W. R. Dombrowsky Zur Akzeptanz staatlicher Informationspolitik bei technischen Großunfällen und Katastrophen Katastrophenmedizin – Leitfaden für die ärztliche Versorgung im Katastrophenfall Neuauflage 2001, Broschur Broschüren und eine komplette Liste aller bisher erschienenen und bereits vergriffenen Bände können kostenlos bezogen werden bei: Bundesverwaltungsamt – Zentralstelle für Zivilschutz – Deutschherrenstraße 93–95 53177 Bonn 74 Notitzen 75 Notitzen 76