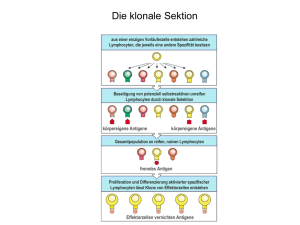
T-Zellen werden zur Kontrolle intrazellulärer Pathogene benötigt
Werbung

spezifische Antikörper • Komponenten und Aufbau des Immunsystems • Erkennungs und Effektormechanismen des Immunsystems bakterielle Toxine Bakterien im extrazellulären Raum Zelle mit Toxinrezeptoren • Lymphozytenentwicklung Neutralisierung Antikörper können auf drei Arten an der Immunabwehr beteiligt sein Bakterien im Plasma Makrophage Opsonisierung Aktivierung des Komlementssystems • Entstehung und Verlauf adaptiver Immunantworten Komplement T-Zellen werden zur Kontrolle intrazellulärer Pathogene benötigt und um B Zellen gegen die meisten Antigene zu aktivieren virusinfizierte Zelle Cytotoxische Zelle tötet infizierte Zelle Immunabwehr intrazelluärer Virusinfektionen cytotoxische T-Zelle Immunabwehr intrazellulärer Infektionen durch Mycobakterien infizierter Makrophage Lysosom aktivierter infizierter Makrophage Mycobakterium Virus infizierte Zelle abgetötete infizierte Zelle Antigen 1 Immunglobulinmolekühle bestehen aus zwei verschiedenen Arten von Polypeptidketten, den schweren Ketten und den leichten Ketten leichte Ketten Disulfidbrücken schwere Ketten Die Anitkörperarme sind durch ein flexibles Gelenk verbunden The Y-shaped immunoglobulin molecule can be dissected by partial digestion with proteases proteolytische Spaltung mit Papain Winkel zwischen den Armen 0° Winkel zwischen den Armen 90° proteolytische Spaltung mit Pepsin In den variablen Domänen gibt es definierte hypervariable Bereiche V-Region der schweren Ketten V-Region der leichten Ketten Variabilität Variabilität Winkel zwischen den Armen 60° Aminosäure Aminosäure 2 Antigene können in Taschen, Gruben oder an ausgedehnten Oberflächen innerhalb der Bindungsstellen von Antikörpern binden Der T-Zell-Rezeptor ähnelt einem membrangebundenem FarbFragment Die Struktur des T-Zell-Rezeptors Antigenbindungsstelle Antigenbindungsstelle Kohlenhydrat Antikörper α-Kette β-Kette variable Region (V) Antigenbindungsstelle konstante Region (V) Gelenk (H) T-ZellRezeptor Transmembranregion Cytoplasmatischer Schwanz T-Zelle Disulfidbrücke Unterschiede in der Antigenerkennung zwischen Immunglobulinen und T Zell Rezeptoren Zwei Hauptgruppen von T-Zellen erkennen Peptid, das an Proteinen von zwei unterschiedlichen Klassen von MHC Molekülen gebunden ist 3 MHC-Moleküle präsentieren die Peptidfragmente von Antigenen auf der Zelloberfläche MHC-I-Molekül präsentieren Antigene, die aus Proteinen im Zytosol stammen Im Zytosol werden virale Proteine synthetisiert Virus infiziert eine Zelle MHC-Klasse I MHC-Klasse II endoplasmatisches Reticulum Peptid MHC-I bindet Peptidfragmente viraler Proteine im ER MHC-I transportiert gebundene Peptide and die Zelloberfläche Zellmembran Zellkern Bakterium infiziert Makrophagen und dringt in Vesikel ein; Peptidfragmente entstehen MHC-II bindet bakterielle Fragmente in den Vesikel MHC-II transportiert gebundene Fragmente an die Zelloberfläche MHC class II molecules present antigen originating in intracellular vesicles Cytotoxische T-Zellen erkenn Antigene, die von MHC-I-Molekülen präsentiert werden, und töten die Zelle ab Cytotoxische T-Zelle erkennt Komplex aus viralem Fragment und MHC-I und tötet infizierte Zelle tötet Antigen wird aufgenommen und zu Peptidfragmente abgebaut an B-Zell-Rezeptor gebundenes Antigen Fragmente binden an MHC-II und werden an die Oberfläche transportiert MHCKlasse-I Antikörper B-Zelle TH1- und TH2-Zellen erkennen Antigene, die von MHC-II-Molekülen präsentiert werden Je nach Art des Antigens können Immunantworten nützlich oder schädlich sein Wirkung der Reaktion auf das Antigen TH1-Zelle erkennt Komplex aus bakteriellem Fragment und MHC-II und aktiviert Makrophagen aktiviert MHCKlasse-II TH2-Helferzelle erkennt Komplex aus antigenem Fragment und MHC-II und aktiviert die B-Zelle T-Helferzelle aktiviert MHCKlasse-II Antigen normale Reaktion ungenügende Reaktion Krankheitserreger Schützende Immunität wiederholte Infektionen harmlose Substanz Allergie keine Reaktion Transplantat Abstoßung Annahme körpereigenes Gewebe Autoimmunität Selbst-Toleranz Tumor Immunität gegen Tumoren Krebs 4 MHC-Moleküle präsentieren die Peptidfragmente von Antigenen auf der Zelloberfläche MHC-Klasse I Grundzüge der Strukturen der CD4- und CD8-Corezeptormoleküle MHC-Klasse II Peptid Zellmembran Die Expression von MHC-Molekülen unterscheidet sich in verschiedenen Geweben Gewebe MHC-Klasse I MHC-Klasse II Der T-Zell-Rezeptor-Komplex besteht aus Proteinen, die das Antigen erkennen, und unveränderlichen Proteinen, die Signale aussenden Lymphatisches Gewebe T-Zellen Erkennung B-Zellen Makrophagen andere antigenpräsentierende Zellen (z.B. Langerhans-Zellen) epitheliale Zellen des Thymus andere kernhaltige Zellen Neutrophile Zellen Hepatocyten Niere Gehirn kernlose Zellen rote Blutkörperchen Signalgebung Die Aggregation des T-Zell-Rezeptors und eines Corezeptors setzt die Verbreitung von Signalen in der Zelle in Gang in ruhenden T-Zellen sind die ITAMs nicht phosphoryliert Bindung des Liganden an den Rezeptor führt zur Phosphorylierung der ITAMs durch rezeptorassoziiete Kinasen ZAP-70 bindet an die phosphorzlierten ITAMs der ζKette; es wird durch Lck phosphoryliert und aktiviert, sobald der Corezeptor an den MHC-Liganden gebunden hat antigenpräsentierende Zelle T- Zelle 5