Kommunikation der Zellen
Werbung

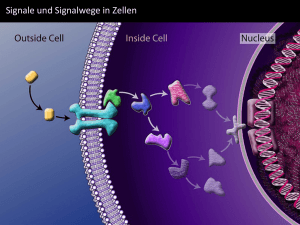
Kommunikation der Zellen INHALT: 1. 2. 3. 4. Die Grundlagen Komponenten der Signalwegen Nervenzellenkommunikation Komplexe Signalwege 1. Vorlesung 1.Vorlesung 2. Vorlesung 3. Vorlesung Grundlagen der Zellkommunikation 1. Einleitung 2. Typen der Zellkommunikationen 3. Signalweitergabe 4. Regulierung der Signalwegen Einleitung zur Grundlagen der Zellkommunikation Der Organismus der mehrzelligen Lebewesen wird von Zellen gebildet, welche durch ihre interaktive Kommunikation im Laufe der Ontogenese die Entwicklung und den Funktionsmechanismus des Organismus in kohärenter Einheit sichern. Die Zellen spalten sich, differenzieren sich, überleben, sterben, bewegen sich, erhalten die biochemischen und genetischen Mechanismen aufrecht, usw. Der Sinn der mehrzelligen Lebensform ist, dass die Zellen die unterschiedlichen Funktionen teilen, unterschiedliche Zellenarten bilden. Die spezialisierte Aufgabe können die Kontraktion, die Antwortgabe auf elektrische Impulse, die Entgiftung, der Gasaustausch, die Sekretion von unterschiedlichen Stoffen, der Schutz den fremden Eindringlingen gegenüber, die Sicherung der Körperstruktur, usw. Die erwähnten Lebensabläufe können die Zellen versorgen, indem sie Signale annehmen und absenden, wodurch sie sich verändern, ihren Funktionsmechanismus mit den Nachbarzellen synchronisieren, oder sich von diesen gerade ganz abgrenzen. Das Thema enthält sehr viele Details, deshalb versuchen wir in diesem Material statt den vollständigen Prozessen die Hauptsache und Logik der Funktionsmechanismen vorzustellen. Folie 1-11 Die Arten der Kommunikation der Zellen Innerhalb von einem Organismus können die Zellen - durch Senden von Signalen - den eigenen Funktionsmechanismus und den von anderen Zellen regulieren. Während der Regulation des eigenen Funktionsmechanismus durch Signale (autokrine Kommunikation) geht es oft darum, dass das eigentlich an die Nachbarzellen gesendete Signal auch, durch ein negatives Feed-back, auf die Rezeptoren der sendenden Zelle wirkt. Das heißt, die sendende Zelle nimmt einfach wahr, ob sie genügend Signalmoleküle in die Umgebung gestoßen hat. Wenn eine Zelle die Zellen aus ihrer nahen Umgebung reguliert, also das Signalmolekül nicht weit wandert, dann reden wir über eine parakrine Wirkung. Ein spezieller Fall der nahen Wirkung ist, wenn die miteinander kommunizierenden Zellen physischen Kontakt miteinander haben: entweder kann von juxtakriner Wirkung - nämlich die Interaktion zwischen Ligand und Rezeptoren -, oder von der Gap- Junction - wenn die Kommunikation über die Membrane überbrückenden speziellen Ionenkanäle der Nachbarzellen stattfindet - gesprochen werden. 1 Die ferne Verbindung wird durch die endokrine Kommunikation gesichert, während welcher die endokrinen Drüsen Hormone in das Blut ausstoßen, welche sich den Zellen, die über sie erkennenden Rezeptoren verfügen, anknüpfen. Es ist schwierig die synaptische (=neurokrine) Verbindung in eine der Kategorien der nahen oder fernen Verbindungen einzuordnen, da es sich eigentlich um eine Verbindung zwischen den sendenden (presynaptischen) und den annehmenden (post-synaptischen) Neuronen handelt, aber das annehmende Neuron das Signal weiterleitet, und das Signal so auch fernliegende Ziele erreicht. Aus bestimmter Hinsicht liegt der Unterschied zwischen dem hormonellen und neuronalen Kontakt dort, dass im Gegensatz zu den endokrinen Drüsen die Nerven die chemischen Signale, welche oft Hormone sind, gezielt (und schneller) transportieren. Nicht nur die eigenen Zellen des Organismus können miteinander kommunizieren, sondern auch die Zellen des Embryos können mit denen des Uterus (Plazenta) der Mutter, und ebenso die pathogenen Bakterien (und die Viren, obwohl diese keine Zellen sind) mit den menschlichen Zellen. Bestimmte Zellen der einzelnen Exemplare kommunizieren miteinander auf chemischer Weise, über Pheromone, was hauptsächlich das Sexualverhalten reguliert. Auch von anderen Exemplaren ausgestoßene visuelle, und aus Kontakten stammende auditive Signale können Teil der Kommunikation bilden, diese treffen zwar nicht in Form von chemischen Signalen ein, aber während der Aufarbeitung wandeln sich diese in chemische Signale um. . Weitere allgemeine Merkmale der Signalprozesse FOLIE 13-20 Signaländerung Die in die Zelle eintreffende Information trifft in Form von einem chemischen Molekül ein. Nach der Ankoppelung an den Rezeptoren wandelt sich dieses Signal in ein anderes um. In weiterem Sinn ist die Konformationsveränderung des Rezeptors auch ein geändertes Signal, aber in engerem Sinn versteht man unter Signaländerung den Fall, wenn sich die sich an die Membran koppelnde Moleküle aktivieren. Manche sind der Ansicht, dass die Entstehung der sekundären Botenstoffe die veränderte Form des Signals bedeutet. Der Signaltransport (Relay) ist der Prozess, wenn die intrazellulären Signalmoleküle das eintreffende Signal aneinander, einander aktivierend weiterleiten. Die Signalamplifikation (Verstärkung) ist der Prozess, wo im Laufe dessen die Zahl der aktivierten Moleküle bei jedem Schritt wächst. Die Signalverstärkung ermöglicht, dass eine Zelle auch sehr schwache Inputs wahrnimmt. Die cAMP Route betrachtet produziert eine Adenylatcyclase Moleküle viele cAMP-s; eine Proteinkinase A Moleküle viele andere Kinasen, jede von diesen ist in der Lage mehrere Kinase-Enzyme zu aktivieren. Die Signaldivergenz bedeutet, dass ein Signal auf mehr als ein Effektormolekül wirkt, was dann weitere, voneinander unabhängige Wirkungen in der Zelle generiert. Während der Signalmodulation wirken bestimmte Proteinmoleküle auf einen gegebenen Signalprozess meistens hemmend. FOLIE 21-23 Interaktion zwischen den Komponenten der Signalwege Grundsätzliche Eigenheit der Signalwege ist die Interaktion: zeitlich gemessen wirkt ein vorher aktives (upstream) Element auf das darauffolgende (downstream). In einem Signalprozess gibt es aber mehrere solche Punkte, wo ein normalerweise später funktionierender Komponent auf die Funktion des vorher funktionierenden Komponents wirkt. Wenn die Rückwirkung verstärkend ist, dann redet man von einer positiven Rückwirkung (feedback), wenn diese hemmend ist, dann über eine negative. Die Signalwege kommunizieren miteinander (crosstalk= Kreuzender Effekt), können die Wirkung voneinander positiv oder negativ modulieren. Die unterschiedlichen Signalwege können sich so aneinanderknüpfen, dass sie über einem gemeinsamen Komponent verfügen. Man spricht über die Integration der Signale wenn ein 2 Effektor-Molekül die Inputs von zwei unterschiedlichen Signalwegen bekommt, und die Folge dieser bestimmt die Effektorwirkung. Die Integration kann auch so zustande kommen, dass z.B. einen Komponenten des Effektor-Moleküls der eine Weg, den anderen Komponenten der andere Weg produziert. Wenn einer nicht entsprechend funktioniert, entstehen keine funktionsfähigen Effektor-Moleküle. Auf der Folie sind die Interaktion und die Integration von wahrhaften Signalwegen zu sehen. Im Laufe der Vorlesung werden wir uns nicht mit jeder von diesen beschäftigen ( z.B. Wnt, Hedgehog, extrazelluläres Matrix), die anderen werden aus dem Ergänzungsmaterial dann verständlich. FOLIE 24-25 Die Desensibilisierung einer Zielzelle kann auf verschiedene Weisen vorkommen. Desensitisation geschieht auf verschiedene Arten: 1. Rezeptorsequestrierung: Ligandbindung kann zur Endozytose und zeitweisem Einschliessen in Endosomen führen 2. Rezeptor-Down-Regulierung: Lysosomen könnten weiter die Rezeptoren zerstören, was Rezeptor down-Regulation genannt wird 3. Die Rezeptoren werden sofort inaktiviert, z.B. durch Phosphorylierung 4. Veränderung und Inaktivierung des Signalproteins 5. Produktion eines Inhibitors, der den Transduktionsprozess stoppt. FOLIE 26 Ein Ligand wirkt auf mehrere Rezeptoren Die Zahl der Rezeptoren beträgt viel mehr als die der Liganden. Die mathematische Folge dieser Tatsache ist, dass ein Ligand auf mehrere Rezeptoren wirkt. Die Rezeptoren, die den gleichen Ligand erkennen sind meistens auf demselben Zellentypen platziert (nicht in jedem Fall, z.B. können an derselben Nervenzelle unterschiedliche Glutamatrezeptoren platziert sein). Wir stellen die Situation am Beispiel des Acetylcholin (Ach) Rezeptor dar. Wie man sieht ruft Ach in der glatten Muskulatur und im Herzmuskel eine Entspannung hervor, aber in der quergestreiften Muskulatur eine Kontraktion. Grund hierfür ist, dass, während in der Skelettmuskulatur das Ach sich an den Ionenkanal-gesteuerte Rezeptor koppelt, knüpft es in der glatten Muskulatur an den G-Protein gekoppelten Rezeptor an. Weiterhin löst Ach in der Speicheldrüse und der Bauchspeicheldrüse Sekretion aus (regt sowohl den Ausstoß von Enzymen, als auch den von Insulin an), und regt die Nervenzellen (meist Neuro-Transmitter) zum Abfeuern an. FOLIE 28-29 Das Schicksal einer Zelle hängt von den eintreffenden Signalkombinationen ab Zum am Leben bleiben der Zellen ist es notwendig, dass sie ständig unterschiedliche Signale ( Überlebensfaktoren ) bekommen. Zur Spaltung und Differenzierung sind außer den Überlebensfaktoren noch weitere Komponenten notwendig, dies sind die Differenzierungsfaktoren. Wenn die Zellen keine Überlebensfaktoren erhalten, oder Todessignale bekommen, dann ist die Folge ein programmierter Zellentod. Die Apoptose kann auch ohne äußere Faktoren, infolge der Wirkung von inneren Signalen ablaufen. FOLIE 30 Schnelle und langsame Signalwege Das Tempo eines Signalprozesses hängt davon ab, aus wie vielen Komponenten der Signalweg besteht. Die gegenseitige Aktivierung der Komponente ist aber ein sehr schneller Prozess, so ist der Unterschied zwischen einem kürzeren und einem längeren Signalweg nicht bedeutend. Das Tempo der Signalwege wird verlangsamt, wenn die Veränderung der Aktivität der Gene auch Zielpunkt des Signalweges ist. Ursprünglich nennt man die Signalwege schnell, bei welchen sich die DNS Aktivität nicht ändert, und die werden langsam genannt, bei welchen das Entstehen der neuen Proteine auch Teil des Signalprozess ist. 3