Leseprobe zum Titel - content
Werbung

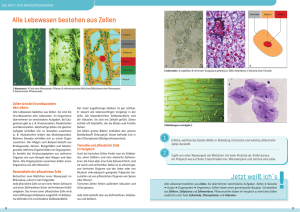
Seite 5 ZYTOLOGIE 1 Zelle Eine Zelle besteht aus einem Zellleib (Zytoplasma) und einem Zellkern. Das Zytoplasma beinhaltet das Zytosol, Zellorganellen und das Zytoskelett. Der Zellleib ist von der Zellmembran (Plasmamembran) umgeben, der Zellkern von der Kernmembran. 1.1 Zellmembranen Jede Zelle wird an ihrer Oberfläche durch eine Zellmembran (Plasmamembran, Plasmalemm) begrenzt. Diese besteht im Wesentlichen aus polaren Lipiden (hauptsächlich Phospholipiden) und Proteinen. Phospholipidmoleküle bilden eine flexible Doppelschicht, wobei die hydrophilen Anteile der Moleküle nach außen weisen, während ihr jeweiliger hydrophober Molekülanteil das Innere der Doppelschicht bildet. Diese Lipid-Doppelschicht bildet die Grundstruktur von biologischen Membranen. Sie muss flexibel sein, sich zum Beispiel Formveränderungen der Zelle anpassen können. Dazu müssen die Lipidmoleküle innerhalb ihrer Ebene beweglich sein (Fluidität der Biomembranen). Cholesterinmoleküle innerhalb der Membran vermindern diese Fluidität, sie dienen der Verfestigung, Versteifung der Membran. In die Lipid-Doppelschicht sind Proteine eingelagert. Ein Teil der Proteine reicht durch die LipidDoppelschicht hindurch (integrale Membranproteine), andere sind der Lipidschicht angelagert (periphere Membranproteine). Die Membranproteine sind für die meisten Funktionen der Membran verantwortlich. Sie fungieren als Strukturproteine, Enzyme, Kanäle, Transporter, Pumpen und als Rezeptoren (z.B. Endozytoserezeptoren, Hormonrezeptoren, Neurotransmitterrezeptoren, Immunrezeptoren). Die Glykokalix ist Bestandteil der Plasmamembran. Glykolipide in der äußeren Lipidschicht tragen an ihrem nach außen weisenden Ende Zuckerketten, Oligosaccharide. Ebenso besitzen manche der integralen Membranproteine Zuckerketten (Glykoproteine, Proteoglykane), mit denen sie über die äußere Lipidschicht hinausragen können. Die Gesamtheit aller dieser Zuckerketten an der äußeren Oberfläche der Plasmamembran wird als Glykokalix bezeichnet und ist für jede Zellart charakteristisch. Neben der Zellmembran an der Oberfläche sind auch im Inneren der Zelle derartige Membranen vorhanden, die den Zellkern und einige Zellorganellen (z.B. Mitochondrien, endoplasmatisches Retikulum, Golgi-Apparat, Lysosomen, Peroxisomen) begrenzen. Durch diese Membranen werden in der Zelle verschiedene Reaktionsräume oder Kompartimente geschaffen. Darstellung der Zellmembran Bei der Fixierung von Zellen mit Osmiumtetroxid sieht man die Lipiddoppelschicht in Ultradünnschnitten im Transmissionselektronenmikroskop (bei starken Vergrößerungen) als zwei elektronendichte, dunkle Linien, die durch eine dazwischen liegende helle Zone voneinander getrennt sind (Abb. 1). Die Glykokalix erscheint in solchen elektronenmikroskopischen Schnittbildern als filzartige Auflagerung an der Außenseite der Zellmembran. Im Gefrierätzpräparat sieht man die Membranproteine als kleine, meist punktförmige Erhebungen aus der Lipidschicht herausragen. © Hartmann et al., Institut für Zellbiologie, Histologie und Embryologie, Medizinische Universität Graz Seite 6 Die lichtmikroskopisch sichtbare Zellmembran ist ein Artefakt der Histotechnik. Dieser entsteht dadurch, dass sich Proteine des Zytoplasmas an die Membran anlagern und sie dadurch deutlicher erscheinen lassen. Abb. 1: Mikrovilli: Zellmembran Abb. 2: Oberflächendifferenzierung: Kinozilien 1.2 Oberflächendifferenzierungen Jede Körperzelle hat spezifische Aufgaben zu erfüllen, tritt mit ihrem Umfeld – mit anderen Zellen und mit dem Extrazellulärraum – in Wechselbeziehung, und besitzt dazu (neben ihrer Ausstattung mit bestimmten Rezeptoren und Kanalproteinen) speziell differenzierte Strukturen an ihrer Oberfläche. Viele Zellen sind polar differenziert, insbesondere trifft dies auf Epithelzellen zu. An diesen Zellen kann ein apikaler von einem basalen Pol unterschieden werden. An den jeweiligen Bereichen der Oberflächenmembran (apikal, lateral und basal) finden sich unterschiedliche Differenzierungen. 1.2.1 Mikroplicae sind kleine faltenförmige Aufwerfungen der Oberflächenmembran, die – wie Mikrovilli – Aktinfilamente enthalten und der Oberflächenvergrößerung dienen. 1.2.2 Mikrovilli sind etwa 0,1 µm dicke, fingerförmige, 1-2 µm lange Fortsätze an der apikalen Zelloberfläche, die ebenfalls der Vergrößerung der Zelloberfläche dienen. Sie besitzen ein inneres Stützgerüst aus Aktinfilamenten. (Abb. 1) In kleiner Anzahl finden sich Mikrovilli bei unterschiedlichen Zellarten, wie zum Beispiel bei Endothelzellen, Lymphozyten oder Leberzellen. Manche dieser Mikrovilli sind möglicherweise nur temporär vorhandene Strukturen. Bei Zellen, die vorrangig im Dienste der Resorption stehen – Darmepithelzellen, Epithelzellen des Tubulus proximalis der Niere –, bilden Mikrovilli einen dichten Rasen an der Zelloberfläche, in der Lichtmikroskopie als Bürstensaum bezeichnet. Diese Mikrovilli besitzen ein inneres Stützskelett aus Aktinfilamentbündeln, die von der Spitze der Mikrovilli bis in den apikalen Zytoplasmabereich reichen und hier in einem Filamentnetz (Terminal web) verankert sind. © Hartmann et al., Institut für Zellbiologie, Histologie und Embryologie, Medizinische Universität Graz Seite 7 In die vergrößerte Oberflächenmembran dieser Zellen sind für spezifische Resorptionsprozesse verschiedene Enzyme und Transportproteine eingebaut. Im Gefrierbruchpräparat treten sie als kleine Partikel in Erscheinung. An der Mikrovillimembran ist eine besonders deutlich ausgebildete Glykokalix vorhanden. 1.2.3 Stereozilien sind ebenfalls fingerförmige, 0,2 µm dicke und bis zu 10 µm lange Fortsätze und besitzen – wie Mikrovilli – ein Binnengerüst aus Aktinfilamenten. Samenweg-Stereozilien kommen an der apikalen Oberfläche der Epithelzellen des Ductus epididymidis und am Beginn des Ductus deferens vor. Sie bilden häufig Büschel, die aus einem gemeinsamen Sockel kommen können. Büschel solcher Stereozilien sind lichtmikroskopisch zu sehen. Innenohr-Stereozilien an Sinneszellen (Haarzellen) des Innenohres sind steife Stereozilien. Sie besitzen ein Skelett aus auffallend vielen, dicht gepackten Aktinfilamenten, die in einer sogenannten Kutikularplatte (entspricht dem Terminal web bei Epithelzellen) verankert sind. Sie sind sowohl am Hörvorgang als auch an der Funktion des Gleichgewichtsorgans beteiligt. 1.2.4 Kinozilien („Flimmerhärchen“) sind etwa 0,25 µm dicke, bis 5 µm lange, wimpernförmige Zellfortsätze, die mit einem axialen Mikrotubulus-Dynein-Bewegungsapparat schlagende Bewegungen ausführen können. Epithelzellen mit vielen Kinozilien an der apikalen Oberfläche (Abb. 2) werden als Flimmerzellen bezeichnet. Sie kommen im Epithel der Atemwege, des Eileiters und der Ductuli efferentes des Nebenhodens vor. Jeweils ein Kinozilium ist an den Sinneszellen des Gleichgewichtsorgans im Innenohr vorhanden. Die Bewegung eines Kinoziliums besteht aus einem raschen Schlag und einer langsamen Rückholbewegung. Alle Kinozilien einer Zelle bzw. eines Zellverbandes schlagen rhythmisch in einer genetisch festgelegten Richtung, die Schläge erfolgen koordiniert nacheinander, metachron. Der Bewegungsapparat einer Kinozilie wird aus Mikrotubuli und dem Motorprotein Dynein gebildet. Das Mikrotubulussystem im Inneren einer Kinozilie besteht aus einer zylinderförmigen Anordnung von 9 Mikrotubulus-Paaren (Doubletten) und 2 zentral gelegenen Einzeltubuli (9x2plus2-Struktur). Bei den Doubletten ist jeweils ein Mikrotubulus mit 13 Protofilamenten vollständig ausgebildet: A-Tubulus. Der zweite, B-Tubulus, ist mit 11 Protofilamenten inkomplett und mit dem A-Tubulus verbunden. Das Mikrotubulusgerüst der Kinozilie ist aus dem knapp unterhalb der Basis der Kinozilie im apikalen Zytoplasma gelegenen Kinetosom ausgewachsen. Ein Kinetosom (Basalkörperchen) ist aufgebaut wie ein Zentriol, zylinderförmig, mit einer Wand bestehend aus 9 längsverlaufenden Mikrotubulus-Tripletten, wobei jeweils eine Triplette aus einem kompletten A-Tubulus, einem inkompletten B-Tubulus und einem inkompletten C-Tubulus besteht (9x3-Struktur). Die 9 Mikrotubulusdoubletten der Kinozilie sind jeweils Fortsetzungen der A- und B-Tubuli des zugehörigen Kinetosoms. Die Bewegung von Kinozilien erfolgt durch Gleitbewegungen zwischen benachbarten Tubulusdoubletten, die durch das Motorprotein Dynein verursacht werden. An jedem A-Tubulus sind „Dyneinarme“ befestigt, die – unter Spaltung von ATP – an den B-Tubuli der jeweils benachbarten Doublette Richtung Basis der Kinozilie entlangwandern, wodurch Doubletten gegeneinander „gleiten“. Da die Mikrotubulus-Doubletten der Kinozilie jedoch nicht frei beweglich, sondern mit den Mikrotubuli des Kinetosoms verbunden sind, wird aus der Gleitbewegung © Hartmann et al., Institut für Zellbiologie, Histologie und Embryologie, Medizinische Universität Graz Seite 8 eine Verbiegung. Diese findet abwechselnd in der einen bzw. anderen Hälfte der Zilie statt, was einen Vor- bzw. Rückschlag bewirkt. Die Kinozilien von Flimmerzellen sind lichtmikroskopisch erkennbar. An der Basis der Kinozilien ist ein stärker färbbarer Streifen zu sehen, der durch die Summe der Kinetosomen (Basalkörperchen) hervorgerufen wird. 1.2.5 Geißel (Flagellum) Eine Geißel ist eine einzelne, extrem lange, spezialisierte Kinozilie im etwa 55 µm langen Schwanzstück eines Spermiums, mit deren Hilfe sich das Spermium vorwärts bewegen kann. 1.2.6 Crusta ist ein lichtmikroskopischer Begriff für spezielle Oberflächendifferenzierungen an der apikalen Oberfläche von Deckzellen des Urothels (Übergangsepithels): der apikale Zytoplasmasaum dieser Zellen färbt sich stärker an als der übrige Zellleib. An der apikalen Zellmembran der Deckzellen sind dicht aneinander liegende, plattenförmige Areale (Plaques) ausgebildet, zwischen diesen sind jeweils schmale, flexible Membrananteile (Gelenke) gelegen. Im Bereich der Plaques sind in der Plasmamembran dicht gepackte, integrale Membranproteine (Uroplakine) vorhanden. Diese Plaques können eingestülpt und in Form von scheibenförmigen Vesikeln im Zytoplasma unterhalb der Zellmembran „gespeichert“ und bei Bedarf wieder in die Plasamamembran eingefügt werden. Die Vesikel, wie auch ein im apikalen Zytoplasma vorhandenes dichtes Netz von Intermediärund Aktinfilamenten sind ursächlich für die stärkere Anfärbbarkeit des oberflächlichen Zytoplasmas, die im Lichtmikroskop als Crusta gesehen wird. Die Crusta dient in den harnableitenden Organen (Nierenbecken, Ureter, Harnblase, proximaler Bereich der Urethra) als Schutz vor potentiell schädigenden Substanzen im Harn, mit dem dieses Epithel ständig in Kontakt ist. 1.2.7 Vergrößerungen der basolateralen Membranoberfläche Nachbarzellen können lateral mehr oder minder kompliziert durch Einfaltungen und Ausstülpungen miteinander verschränkt sein (z.B. im Nieren- und Darmepithel), was zu einer Vergrößerung der Oberfläche führt. In Nierenkanälchen (proximaler und distaler Tubulus) und im Streifenstück des Ausführungsgangsystems von Speicheldrüsen gibt es zur basalen Oberflächenvergrößerung basale Einfaltungen der Zellmembran, die Platz für Ionenpumpen schaffen. Zwischen den Membraneinstülpungen sind in Reihen viele Mitochondrien angeordnet. Beide Strukturen zusammen bilden die lichtmikroskopisch sichtbare basale Streifung. Weitere Beispiele für Strukturen zur Membranoberflächenvergrößerung sind schlauchförmige Einsenkungen der Zellmembran (T-Tubuli) in Skelettmuskulatur und Herzmuskulatur. 1.3 Zytoskelett Im Zytoplasma von Zellen ist elektronenmikroskopisch ein Netzwerk von Filamenten und Tubuli zu sehen, das einen internen Stütz- und Bewegungsapparat, das Zytoskelett oder Zellskelett, bildet. Das Zytoskelett ist verantwortlich für die mechanische Stabilisierung einer Zelle und ihrer äußeren Form, für Bewegungen der Zelle (Muskelkontraktion, Zellwanderung; Kinozilienbewegung) und für Transportvorgänge innerhalb der Zelle. © Hartmann et al., Institut für Zellbiologie, Histologie und Embryologie, Medizinische Universität Graz Seite 9 Zum Zytoskelett gehören: Aktinfilamente oder Mikrofilamente (7nm Durchmesser) Intermediärfilamente (10nm Durchmesser) Mikrotubuli (25nm Durchmesser) Alle Elemente des Zytoskeletts bestehen aus Einzelbausteinen (Proteinen), die sich zu Filamenten zusammenlagern (polymerisieren) und auch wieder auseinander fallen (depolymerisieren) können. Für jedes Filamentsystem gibt es spezifische Begleitproteine, die Polymerisation und Depolymerisation, wie auch Anordnung und Stabililtät des Filamentsystems regulieren. In Verbindung mit Motorproteinen dienen Aktinfilamente und Mikrotubuli der Bewegung von Zellen selbst und Bewegungsabläufen innerhalb der Zelle, z.B. dem intrazellulären Transport von Zellorganellen. In gebündelter Form können Filamente und Tubuli auch lichtmikroskopisch als „Fibrillen“ sichtbar sein (z.B. Neurofibrillen in Nervenzellen, Tonofibrillen in Epithelzellen). 1.3.1 Aktinfilamente oder Mikrofilamente Aktinfilamente sind in allen Zellen vorhanden. Sie bilden sich durch Aneinanderlagerung globulärer Aktinmoleküle (G-Aktin), von denen es verschiedene Isoformen gibt. Zwei α-helikal umeinander gewundene Stränge bilden ein Aktinfilament (F-Aktin) mit einem Durchmesser von 7nm. Aktinfilamente sind polar gebaut: am (+)-Ende (Plus-Ende) können sie sich rasch verlängern, aber auch ebenso rasch wieder zerfallen, am (–)-Ende (Minus-Ende) laufen die Vorgänge langsamer ab. Verschiedene Aktin-Begleitproteine regulieren Umbau und Zerfall der Aktinfilamente, verbinden die Filamente zu Bündeln oder Netzen und verankern sie an Transmembranproteinen der Plasmamembran. Die Motorproteine der Aktinfilamente sind die Myosine. Myosine, von denen es viele verschiedene Klassen und Unterklassen gibt, finden sich in fast jeder Zelle und sind in Verbindung mit Aktinfilamenten die Voraussetzung für die Kontraktilität von nicht-muskulären Zellen und Muskelzellen. In Muskelzellen bilden Myosine dicke Myosinfilamente. Aktinfilamente können einzeln oder in Bündeln durch das gesamte Zytoplasma ziehen und finden sich bei den meisten Zellen als dichtes Netz unter der Zellmembran (kortikales Netz). Es ist für die Aufrechterhaltung der Zellform und – durch rasche Umgestaltung der Aktinfilamente – für aktive Formveränderungen von Zellen verantwortlich, z.B. werden bei der Phagozytose und Zellwanderung kurzlebige Fortsätzen wie Pseudopodien bzw. Lamellipodien gebildet. Aktinfilamente dienen auch der Stabilisierung von Mikrovilli und Stereozilien und sind Baubestandteile von Zellkontakten (Zonulae adhaerentes, Fasciae adhaerentes, Fokalkontakten). 1.3.2 Mikrotubuli bestehen aus starren, unterschiedlich langen Röhrchen, die jeweils einen Durchmesser von 25 nm besitzen. Ihre Wand wird aus Molekülen der globulären Proteine α-Tubulin und ß-Tubulin gebildet, die zu Tubulin-Dimeren zusammengelagert sind. Die Tubulin-Dimere aggregieren zu langen Strängen, den Protofilamenten. 13 solcher Protofilamente bilden einen vollständigen Mikrotubulus. © Hartmann et al., Institut für Zellbiologie, Histologie und Embryologie, Medizinische Universität Graz