Kurzlehrbuch Histologie
Werbung

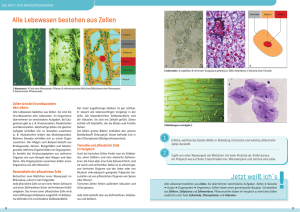
2 Zytologie 2.1 Einleitung 1 Lerncoach Bei der Erarbeitung der verschiedenen Kapitel zur Zytologie sollten Sie auf verschiedene Aspekte achten: Morphologie der Strukturen, molekularer Aufbau von Zellbestandteilen, Entstehung bestimmter Zellstrukturen, Struktur-Funktions-Beziehungen, molekulare Mechanismen von bestimmten Prozessen. Sie sollten sich bei diesen komplexen Themen zunächst einen Überblick verschaffen. Beachten Sie, dass Sie Details zu diesen Themen in verschiedenen Büchern unterschiedlich dargestellt finden. Manche Fakten und ihre Bedeutung werden Ihnen vielleicht erst bei der Bearbeitung der übrigen Histologiekapitel verständlich. Beachten Sie auch, dass Sie die Themen der Zytologie von verschiedenen Fachdisziplinen (Biologie, Biochemie, Physiologie) vorgestellt bekommen. Die Zellmembran ihrer Umgebung ab. Die Zelle untergliedert sich in Zellkern (Nukleus) und Zellleib (Zytoplasma). Im Zytoplasma finden sich die Zellorganellen (kleine „Organe“ der Zelle mit spezifischen Funktionen), ein Zytoskelett, Zelleinschlüsse (z. B. Stoffwechselprodukte) und ein flüssiges Grundplasma (Zytosol, Hyaloplasma). Als kleinste Funktionseinheit des Organismus besitzen Zellen die Fähigkeit zu Stoffwechselleistungen und zur Reizbeantwortung, sie können wachsen und sich vermehren. Im Organismus kommen verschiedene Zellarten vor, die sich durch ihre Form, Größe, Funktion und Lebensdauer voneinander unterscheiden. 2.2 Die Zellmembran 1 Lerncoach Der Aufbau und die Funktion von Biomembranen ist nicht nur eine wichtige Voraussetzung für das Verständnis der Vorgänge in der Zelle, sondern auch die Grundlage vieler biochemischer und physiologischer Prozesse. 2.2.1 Der Überblick 2.1.1 Die Zelle Die Zelle ist die kleinste selbstständig lebensfähige Baueinheit des Organismus (Abb. 2.1). Die Zellmembran (Plasmamembran, Plasmalemm) grenzt sie von Alle Biomembranen, d. h. neben der Zellmembran auch die Membranen der Zellorganellen, sind gleich aufgebaut (Einheitsmembran). Chemisch bestehen sie aus Lipid- und Proteinmolekülen. Die Grundlage Exozytose Mikrovilli Kinozilium Terminales Netzwerk Sekretvesikel Desmosom glattes endoplasmatisches Retikulum Golgi-Apparat Ribosomen Lysosom Kernpore Zelleinschlüsse Mitochondrien Zellkern raues endoplasmatisches Retikulum Endozytose Abb. 2.1 Zelle (Schema). (Nach: Ulfig N. Bewegungsapparat. Karger, Basel, 2002) Plasmamembran Nukleolus Zytoskelett 19 2 20 Die Zellmembran Zuckerketten 2 Zytologie Glykolipid Kanalprotein außen 2 Lipiddoppelschicht innen transmembranöses Protein Abb. 2.2 Phospholipid Cholesterin Aufbau einer Zellmembran (Schema). aller Zytomembranen bildet eine Lipid-Doppelschicht, in der die polaren Köpfe der Phospholipide nach außen, die apolaren Fettsäureketten nach innen, also aufeinander zu zeigen. Elektronenmikroskopisch lassen sich daher drei Schichten erkennen (trilamelläre Einheitsmembran). In die Membran sind Proteinkomponenten eingelagert. Man unterscheidet integrale Proteine, die die gesamte Doppelschicht durchsetzen von peripheren Proteinen, die in die äußere oder innere Fettschicht eingelagert sind. Ein Teil der äußeren peripheren Proteine sind Glykoproteine, deren Kohlenhydratseitenketten an der Bildung der Glykokalix auf der äußeren Oberfläche der Zellmembran beteiligt sind (Abb. 2.2). Die Zellmembran (Dicke: 8 nm) unterliegt einem ständigen Umbau, der mit dem Fluid-Mosaic-Modell beschrieben wird. Membranabschnitte können aus der Zellmembran herausgetrennt oder eingefügt werden. Diese Vorgänge spielen bei der Stoffaufnahme durch Endozytose und bei der Stoffabgabe durch Exozytose eine Rolle. An der Oberfläche bestimmter Zellen kommen Oberflächendifferenzierungen vor, die der Erfüllung spezifischer Aufgaben dienen. In Zellverbänden können die einzelnen Zellen untereinander über spezifische Zellkontakte mechanisch und funktionell gekoppelt sein. 2.2.2 Die Lipid-Doppelschicht und das FluidMosaic-Modell Drei Haupttypen von Lipiden bilden die Lipid-Doppelschicht, die Phospholipide, das Cholesterin und die Glykolipide. Alle Membranlipide haben ein hydrophiles Kopfende und hydrophobes Schwanzende (aus langen Fettsäureketten). Der bimolekulare Film, d. h. die Lipid-Doppelschicht, wird dadurch gebildet, dass die hydrophoben Schwanzenden aufeinander zu weisen (innere hydrophobe Zone). Die hydrophilen Köpfe sind nach außen (zur Zellumgebung) und nach innen (zum Zellinneren) gerichtet. Die Phospholipide sind (mengenmäßig) der Hauptbestandteil der Lipid-Doppel- schicht. Cholesterin beeinflusst die Fluidität der Membran; es führt zu einer geringen Versteifung der Membran. Die Zellmembran muss einerseits stabil, andererseits dynamisch und fluid (flüssig) sein. Aus der Fluidität ergibt sich die Möglichkeit einer fließenden Verlagerung der Membranproteine im Sinne von Lateralverschiebungen (Fluid-Mosaic-Modell). Durch diese Verschiebungen kann es örtlich zu einer Anhäufung bestimmter Membranbestandteile kommen. Die Fluidität der Membran ist von der Lipidzusammensetzung (besonders von der Cholesterinmenge) und von der Temperatur abhängig. Die Glykolipide beteiligen sich mit ihren Kohlenhydratketten an der Bildung der Glykokalix. 2.2.3 Die Membranproteine Die integralen Proteine (auch Transmembranproteine genannt) erstrecken sich durch beide Lipidschichten und verleihen der Membran eine selektive Durchlässigkeit. Diese Proteine bilden Kanäle, Transporter und Pumpen, verschiedene Rezeptoren. Durch Kanäle können Ionen ungehindert entlang einem Gradienten durch die Zellmembran diffundieren. Kanäle können durch bestimmte Signale geöffnet oder geschlossen werden. Die Passage eines Ions oder eines kleinen Moleküls durch einen Transporter (Carrier) dauert länger, weil es während des Transportes zu einer Konfigurationsänderung von Transporterproteinen kommt. Cotransporter sind Carrier, bei denen gleichzeitig ein weiteres Molekül oder Ion in die Zelle „mittransportiert“ (Symport) oder ein Molekül oder Ion aus der Zelle (Antiport) befördert wird. Pumpen können ein Ion gegen ein Konzentrationsgefälle aktiv und unter Energieverbrauch durch die Zellmembran bringen. Die Na+/K+-ATPase z. B. pumpt 3 Na+ aus der Zelle und 2 K+ in die Zelle. Pumpen können also ein spezifisches inneres Milieu schaffen. Wirkstoffe wie Hormone oder Neurotransmitter binden an die extrazelluläre Domäne eines jeweils spezifischen Rezeptors und lösen über Zwischenschritte (Signaltransduktion) einen bestimmten Effekt aus. Die peripheren Membranproteine können (angelagert an Transmembranproteinen) an der inneren oder äußeren Membranoberfläche liegen. Die inneren Membranproteine sind z. B. Adaptoren, über die das Zytoskelett mit Transmembranproteinen verbunden wird, die äußeren peripheren Membranproteine sind z. B. Verbindungsproteine zu Bestandteilen des Extrazellulärraumes. 2 Zytologie Die Zellmembran 21 2.2.4 Die Glykokalix Der Aufbau der Glykokalix 2 Die Glykokalix ist ein kohlenhydrathaltiger Film an der äußeren Oberfläche der Zellmembran. Sie wird gebildet von den Kohlenhydratketten der Glykoproteine und Glykolipide. Die Gesamtheit dieser Zuckerketten bildet die Glykokalix. Durch verschiedene Kombinationen der Zuckermoleküle entstehen Unterschiede in der Glykokalix verschiedener Zellarten. Diese Spezifität der Glykokalix ist die Grundlage für ihre Funktionen. 1 2 Die Funktionen der Glykokalix Die Glykokalix steuert Wechselwirkungen zwischen Zellen. Lektine (als Zucker bindende Proteine) binden selektiv an Bestandteile der Glykokalix; so werden Zell-Zell-Interaktionen ermöglicht (z. B. Adhäsion von Leukozyten am Endothel. Die Glykokalix enthält als Antigene wirksame Moleküle (z. B. Blutgruppensubstanzen). Rote Blutkörperchen fremder Blutgruppen (nach einer Bluttransfusion) werden erkannt und können abgebaut werden. Glykoproteine der Glykokalix können Rezeptorfunktion (z. B. für Hormone) haben. Des Weiteren ergibt sich die Asymmetrie der Zellmembran z. B. durch das Vorkommen der Glykokalix auf der äußeren Seite, die so eine Negativladung enthält. 2.2.5 Die Oberflächendifferenzierungen Bestimmte Zellarten zeigen eine Differenzierung ihrer Oberfläche, die mit ihrer spezifischen Funktion im Zusammenhang steht. Zu den Oberflächendifferenzierungen gehören Mikrovilli, Stereozilien, Kinozilien und basale Einfaltungen. Die Mikrovilli Die Mikrovilli sind fingerförmige Ausstülpungen der Zellmembran (Abb. 2.3). Sie dienen bei resorbierenden Epithelien der Vergrößerung der Zelloberfläche. Bei besonders stark resorptiv tätigen Zellen findet sich ein dichter Rasen gleichlanger Mikrovilli, der schon lichtmikroskopisch als Bürstensaum erkennbar ist. Ein Bürstensaum findet sich z. B. im Dünndarm und in Röhren (Tubuli) der Niere. Kurze einzeln stehende Mikrovilli finden sich bei einer Vielzahl von Zellarten. Im Inneren der Mikrovilli liegen Bündel längsorientierter Aktinfilamente. Diese sind im terminalen Netzwerk im apikalen Zytoplasma (Terminalgespinst, terminal web) verankert, das aus einem Filamentnetz (Aktinfilamente mit Spektrin) besteht. Untereinander sind die Aktinfilamente durch aktinbindende Proteine (z. B. Fimbrin) verbunden. 3 a b Abb. 2.3 Mikrovilli. a Mikrovilli im Längsschnitt und Zellkontakte im Darmepithel. 1, Zonula occludens; 2, Zonula adhärens; 3, Desmosom. Vergrößerung: 50 500-fach. b Mikrovilli im Querschnitt. Im Inneren Aktinfilamente, an der Querfläche Glykokalix. Vergrößerung 90 000-fach. Mikrovilli tragen eine gut ausgeprägte Glykokalix, sie sind bis zu 2 μm lang und etwa 100 nm dick. Die Stereozilien Die Stereozilien (bis 10 μm lang, unbeweglich) gleichen in ihrem Aufbau den Mikrovilli. Sie sind deutlich länger als Mikrovilli. Stereozilien kommen im Nebenhodengang vor. Sie beteiligen sich bei Resorptions- und Sekretionsvorgängen. Des Weiteren können sie als spezielle Oberflächenstrukturen von Sinneszellen der Aufnahme von Reizen im Innenohr dienen. Die Kinozilien Die Kinozilien sind feine, bewegliche Zellfortsätze (Abb. 2.4). Sie sind ca. 6 μm lang, also erheblich länger als Mikrovilli. Ihr Durchmesser beträgt etwa 0,3 μm. Elektronenmikroskopisch ist erkennbar, dass im Inneren der Kinozilien ein charakteristisches System von Mikrotubuli (mit assoziierten Proteinen) vorkommt, das als Axonema bezeichnet wird. Zwei zentrale Mikrotubuli (S. 30), das sog. Zentralpaar, werden von einem Ring aus 9 Paaren (Doubletten, Doppeltubuli) peripherer Mikrotubuli umgeben. Die- 22 Die Zellkontakte 2 Zytologie Jede Kinozilie ist an einem Basalkörperchen (Kinetosom) im Zytoplasma verankert. Die Basalkörperchen gleichen in ihrem Aufbau den Zentriolen (S. 30); sie bestehen aus 9 radiär angeordneten Tripletts (Dreiergruppen) von (kurzen) Mikrotubuli. Die Basalkörperchen können als Basalkörperchensaum lichtmikroskopisch lokalisiert werden. Kinozilien kommen in den Atemwegen, im Eileiter und im Nebenhoden vor. Geißeln ähneln in ihrem Feinaufbau den Kinozilien, sie dienen der Fortbewegung der Spermien (S. 189). Plasmalemm periphere Mikrotubuli 2 zentrale Mikrotubuli Kinetosom Klinischer Bezug Wurzelfaser B-Tubulus a A-Tubulus periphere Mikrotubuli zentrale Mikrotubuli Radiärspeiche Dyneinarme b Plasmalemm Zentralscheide Abb. 2.4 Nexine Kinozilien (Schema). a Längsschnitt; b Querschnitt. ser Aufbau wird als „9 × 2 + 2“-Struktur bezeichnet. Das Zentralpaar wird von zwei getrennten Mikrotubuli gebildet, während die Mikrotubuli der peripheren Doubletten teilweise miteinander verschmolzen sind. Die Mikrotubuli der Doubletten haben an ihrer Kontaktstelle eine gemeinsame Wandung. Der sogenannte A-Tubulus ist vollständig (aus 13 Untereinheiten aufgebaut), an ihm ist der unvollständige BTubulus (aus 11 Untereinheiten) angelagert. Benachbarte Doubletten sind über Nexine miteinander (ringförmig) verbunden. Außerdem verläuft von jedem A-Tubulus ein Proteinfaden als Radiärspeiche nach innen. Die Proteinfäden treten nahe an die Zentralscheide, die die beiden zentralen Tubuli umgibt. Vom A-Tubulus gehen Ärmchen aus, die aus dem Protein Dynein bestehen. Die Ärmchen können sich an den B-Tubulus der benachbarten Doublette anlagern, was zu einer Gleitbewegung zwischen den benachbarten Doubletten führt. Diese Gleitbewegung (Verschiebebewegung) ist die Grundlage des Zilienschlages, der aus einer schnellen Vorwärtsbewegung und einer langsamen Rückwärtsbewegung besteht. Die schnelle Vorwärtsbewegung dient dem Transport von Schleim oder Flüssigkeiten auf der Zelloberfläche. Kartagener-Syndrom: Beim Kartagener-Syndrom liegt ein angeborener Defekt der Dynein-Arme in Kinozilien (und Spermien) vor. Durch den Ausfall der zilienvermittelten Transportfunktion im respiratorischen Epithel kommt es u. a. zu häufigen Lungenentzündungen, Bronchitiden und Nasennebenhöhlenentzündungen. Basale Einfaltungen Mikrovilli, Stereozilien und Kinozilien liegen an der apikalen Oberfläche, d. h. an der Seite der Zelle, die zu einem Hohlraum (Organlichtung) gerichtet ist. Auf der gegenüber liegenden basolateralen Oberfläche (polare Differenzierung) können die basalen Einfaltungen als röhrchenförmige Einsenkungen der Zellmembran vorkommen. Sie führen zu einer Vergrößerung der Zelloberfläche, wo vermehrt Ionenund Wassertransporte stattfinden. Zwischen den Einfaltungen (basales Labyrinth) liegen schmale Zytoplasmaabschnitte, die aufgereihte Mitochondrien enthalten (liefern Energie für Transportvorgänge). Aus den tiefen Einfaltungen und den in Reihen angeordneten Mitochondrien ergibt sich lichtmikroskopisch das Bild der basalen Streifung. Solche basalen Einfaltungen (Invaginationen) kommen in Ausführungsgängen der Speicheldrüsen und in Nierentubuli vor. 2.3 Die Zellkontakte 1 Lerncoach In der Vorlesung, oder beim Nachschlagen in anderen Lehrbüchern können Ihnen sehr unterschiedliche Einteilungen und Bezeichnungen begegnen; im Folgenden ist eine aktuelle Einteilung dargestellt. Es lohnt sich wenig, diese verschiedenen Einteilungen im Detail abzugleichen. Konzentrieren Sie sich also auf die strukturellen und funktionellen Unterschiede zwischen den verschiedenen Zellkontakten. 2 Zytologie Die Zellkontakte 2.3.1 Der Überblick In Zellverbänden können die einzelnen Zellen über spezifische Kontakte miteinander verbunden sein. Solche Verbindungen sind besonders dort ausgeprägt, wo Zellen dichte Verbände bilden (siehe Epithelgewebe). Bei den Zellkontakten handelt es sich um spezifische, d. h. strukturell und funktionell charakterisierte Zellverbindungen (Abb. 2.5, siehe auch Abb. 2.3). Dabei lassen sich aufgrund ihrer Funktion drei Gruppen von Zellkontakten unterscheiden: Kontakte zur mechanischen Verbindung benachbarter Zellen: die Desmosomen und die Adhärens-Kontakte, Kontakte zur metabolischen und elektrischen (ionalen) Kommunikation benachbarter Zellen: der Nexus, Verschlusskontakte (Barrierekontakte): die Zonula occludens, Zudem kommen Kontakte zwischen Zellen und Extrazellulärmatrix vor. 2.3.2 Die Kontakte zur mechanischen Verbindung Diese Kontakte heißen auch Verankerungs- oder Adhäsionskontakte (Haftkontakte), da sie die Zellen aneinander oder an einer Unterlage haften lassen (vgl. Membranproteine a Connexon Proteinkanäle b Intermediärfilamente Plaque c Interzellularspalt Mesophragma Verbindungsproteine Abb. 2.5 Zellkontakte (Schema). a Zonula occludens (Tight Junctions). b Nexus (Gap Junctions). c Desmosom (Macula adhaerens). auch klinischer Fall (S. 18). Dabei wird die Verbindung zwischen zwei Zellen durch transmembranöse Verbindungsproteine hergestellt. Die Teile der Verbindungsproteine benachbarter Zellen, die in den Interzellulärraum ragen, nehmen Kontakt miteinander auf. Der zytoplasmatische Teil der Verbindungsproteine ist (in beiden Zellen) in einer plattenartigen Zytoplasmaverdichtung (Plaque) verankert. Dieser (Anheftungs-)Plaque besteht aus Anheftungsproteinen, sie werden auch als Haftplatten oder submembranöse Verdichtungen bezeichnet. In den Plaque strahlen Bündel von Filamenten des Zytoskeletts ein. Man unterscheidet zwei Typen von Adhäsionskontakten: die Desmosomen und die Adhärenskontakte. Die Desmosomen (Macula adhaerens) Die Desmosomen (oder Fleckdesmosomen genannt) sind umschriebene Haftstellen von runder (Durchmesser: 0,1–0,5 μm) oder elliptischer Form (Abb. 2.5). Sie kommen vor allem in Epithelien und zudem zwischen Herzmuskelzellen vor. Der Interzellulärspalt des Desmosoms ist etwas weiter (30– 50 nm) als in Bereichen, in denen spezielle Zellkontakte fehlen (20 nm). Er ist mit feinfädigem Material (Desmogea) gefüllt, das in der Mitte des Interzellulärspaltes eine linienförmige Verdichtung zeigt (Mesophragma). Die Desmogea besteht aus Cadherinen (Verbindungsproteine: Desmocolline, Desmogleine). Sie ragen in den zytoplasmatischen Plaque, der von der Zytoplasmamembran durch eine schmale Aufhellungszone getrennt ist. Der Plaque besteht aus den Anheftungsproteinen (Adaptor-Proteine: Desmoplakin, Plakoglobin, Plakophilin, Plektin u. a. ). In den Plaque strahlen Intermediärfilamente (Zytokeratin in den Epithelzellen, Desmin in Herzmuskelzellen) ein. Verbinden sich die Zelladhäsionsmoleküle (CAMs) einer Zelle nicht mit den CAMs der Nachbarzelle, sondern mit der extrazellulären Substanz, der Basallamina (S. 40), so spricht man von Hemidesmosomen. Es handelt sich also um Zell-Matrix-Kontakte. Diese entsprechen auch strukturell etwa einer Desmosomenhälfte. Als Verbindungsproteine (an die Extrazellulärmatrix) finden sich hier Integrine. Die Adhärenskontakte Bei den Adhärenskontakten sind die integralen Verbindungsproteine ebenfalls aus der Gruppe der Cadherine (z. B. E-Cadherin im Epithel, N-Cadherin im Herzmuskel, VE-Cadherin im Gefäßendothel). Zu den Plaqueproteinen gehören Aktinin, Vinculin und Catenine. In den Plaque strahlen Aktinfilamente ein. Drei Formen von Adhärenskontakten können unterschieden werden: Zonula, Punctum und Fascia adhaerens. 23 2