Hören/ auditives System
Werbung

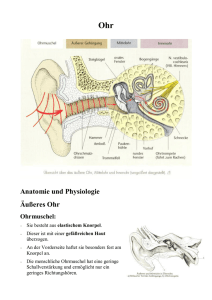
H ren.qxd 24.11.2003 15:10 Seite 1 H.P. Zenner Hören/auditives System Hören/ auditives System H.P. Zenner Hören und Sprechen sind die wichtigsten Kommunikationsmittel des Menschen. Das Gehör des Menschen erlaubt es, hochkomplexe, detaillierte Informationen aus der Umwelt zu extrahieren. In erheblich größerem Ausmaß als jeder andere Sinn ist das Gehör dabei für die menschliche Sprache und ihre Entwicklung verantwortlich. Aufgabe des Ohres Das Ohr ist das empfindlichste Sinnesorgan des Menschen. Der adäquate Reiz ist Schall. Er gelangt durch den äußeren Gehörgang an das Trommelfell, welches als Membran den Gehörgang abschließt und die Grenze zum luftgefüllten Mittelohr bildet. Durch die Gehörknöchelchen des Mittelohrs wird der Schall auf das Innenohr übertragen. Im flüssigkeitsgefüllten Innenohr läuft die Schallenergie als Welle – „Wanderwelle“ – weiter. Aufgabe der Sinneszellen des Innenohrs ist es, dieses mechanische Schallsignal in ein körpereigenes, bioelektrisches bzw. biochemisches Signal zu überführen. Nach diesem Transduktionsprozess gibt die Sinneszelle das Signal mittels eines Transmitters an den Hörnerv weiter. Hörnerv, Hirnstamm und Hörbahn leiten die Information als Folge von Aktionspotentialen, jedoch mehrfach durch Synapsen unterbrochen, über die Hörbahn bis zur Großhirnrinde. Die Schallleitung zum Innenohr Das Ohr des Menschen besteht aus dem äußeren Ohr, dem Mittel- und dem Innenohr [Abb. 1]. Der Schall gelangt durch die Luft des äußeren Gehörgangs bis zum Trommelfell [Luftleitung] und anschließend wird seine Energie durch Schwingungen von Trommelfell und Gehörknöchelchen bis zum ovalen Fenster des Innenohrs fortgeleitet. Gleichzeitig wird der niedrige Schallwellenwiderstand [Schallimpedanz] der Luft an die hohe Impedanz des flüssigkeitsgefüllten Innenohrs angepasst. Das Innenohr kann aber auch Schwingungen der Schädelknochen verarbeiten [Knochenleitung]. Das Mittelohr ist eine Schallbrücke, um den hohen Schallwellenwiderstand des Innenohrs zu überwinden. Im Mittelohr ist in das Trommelfell der Hammer [Malleus*] eingelassen und über den Amboss [In- Äußeres Ohr Mittelohr Nervus vestibulocochlearis Innenohr Trommelfell vestibuläres Labyrinth Gehörgang © Cochlea (Hörschnecke) Abb. 1. Schematische Darstellung des Ohrs. Längsschnitt durch den äußeren Gehörgang, räumliches Schema von Mittelohr und Cochlea. H ren.qxd 24.11.2003 15:10 Seite 2 Hören H.P. Zenner Steigbügel Cochlea Helikotrema Amboß Hammer Struktur der „entrollten“ Cochlea Trommelfell Basilarmembran und Corti-Organ ReissnerMembran Scala tympani (Perilymphe) (sog. Cochleäre Trennwand) ovales Fenster Scala media (Endolymphe) rundes Fenster Scala vestibuli (Perilymphe) Abb. 2. Schema von Mittelohr und Cochlea. Die Cochlea ist entrollt, um die Skalen besser zu sehen. cus*] mit dem Steigbügel [Stapes*] verbunden [Abb. 2]. Die Fußplatte des Steigbügels sitzt beweglich im ovalen Fenster* zum Innenohr. Beim Gesunden wird die Schallenergie im Mittelohr nicht durch Luftdichteschwankungen, sondern durch Schwingungen [Vibrationen] des Trommelfells und der Gehörknöchelchen fortgeleitet. Die Gehörknöchelchen sind anatomisch so gebaut, dass Schallenergie auf das Innenohr übertragen werden kann. Der Trommelfell-Gehörknöchelchen-Apparat passt die Impedanz der Luft an die Impedanz der Flüssigkeit des Innenohrs an. Diese Impedanzanpassung wird durch 2 Hauptmechanismen erzielt: ◗ Die Gehörknöchelchen wirken als Hebel. Dadurch übt die Steigbügelfußplatte auf das ovale Fenster eine größere Kraft aus, als die durch die Luft ursprünglich am Trommelfell erzeugte. ◗ Klinisch bedeutsamer ist es jedoch, dass die Fläche der Steigbügelplatte deutlich kleiner ist als die Fläche des Trommelfells. Da Druck = Kraft/Fläche ist, wird durch den Bau von Trommelfell und Gehörknöchelchen eine Druckerhöhung erreicht. Ohne Mittelohr würde 98 % des Schalls vom Ohr reflektiert und nicht aufgenommen werden. Ursache ist die viel höhere Impedanz des Innenohres im Vergleich zur Luft. Es ist also eine Impedanzanpassung erforderlich, für die Trommelfell und Gehörknöchelchen verantwortlich sind. Die Reflektion wird dadurch so drastisch verringert, dass 60 % der Schallenergie in das Innenohr eintreten kann. Die Schalltransduktion im Innenohr In der Cochlea* [Hörschnecke] des Innenohres bildet das Schallsignal eine Wanderwelle entlang des schlauchförmigen Corti-Organs aus. Das Amplitudenmaximum der Wanderwelle entsteht in Abhängigkeit von der jeweiligen Reizfrequenz an einem bestimmten Ort entlang des Corti-Organs. Die Schwingung des Corti-Organs löst eine Abbiegung der Sinneshärchen der Rezeptorzellen [Haarzellen*] im Corti-Organ aus. Dadurch wird das mechanische Schallsignal in elektrische und chemische Signale umwandelt [transduziert]. Als dessen Folge geben innere Haarzellen einen afferenten Transmitter an die afferenten Fasern des Hörnervs ab. Äußere Haarzellen sind für die aktive Verstärkung des Wanderwellenmaximums und die Stimulation der inneren Haarzellen verantwortlich. Die Innenohrschnecke ist ein aus mehreren Schläuchen aufgebautes Organ, das in Form eines Schneckenhauses in zweieinhalb Windungen aufgerollt ist. Im Querschnitt erkennt man, dass sie aus 4 übereinander liegenden „Schläuchen“ besteht [Abb. 3], aus 3 so genannten Skalen und dem Corti-Organ. Gegen das Mittelohr sind Scala vestibuli und Scala tympani durch die Steigbügelfußplatte am ovalen Fenster bzw. die Membran des runden Fensters abgegrenzt. Scala vestibuli und Scala tympani sind mit der aus dem Liquor stam- H ren.qxd 24.11.2003 15:10 Seite 3 H.P. Zenner Hören/auditives System menden Perilymphe gefüllt, einer Flüssigkeit, die Knochen sich ähnlich wie andere extrazelluläre Flüssigkeiten Reissner-Membran zusammensetzt, also viel Na+ enthält. Scala media Unterhalb der Scala vestibuli liegt die Scala meStria Scala vestibuli dia. Diese wird durch die Reissner-Membran und das vascularis Corti-Organ begrenzt. In der Scala media befindet sich die Endolymphe, eine auffällig K+-reiche FlüsTektorialmembran sigkeit, deren Zusammensetzung intrazellulären Flüssigkeiten ähnelt. Sie ist darüber hinaus gegenäußere über den übrigen Extrazellulärräumen des Körpers Haarzellen stark positiv geladen [etwa +85 mV]. Dieses ständig innere Haarzellen vorhandene Potential heißt endokochleäres Potential. Endolymphe und endolymphatisches Potential Basilarmembran Scala tympani werden durch die Stria vascularis, einem sehr gut Fasern des Hörnervs durchbluteten Bereich der Cochleawand, produziert. Zur Freisetzung des Kaliums besitzen die marginalen Abb. 3. Querschnitt durch die Cochlea. Striazellen Kalium-Ionenkanäle. Das Corti-Organ befindet sich zwischen Scala media und Scala tympani und enthält die Hörsinneszellen [Haarzellen]. Seine Grenzmembran zur Scala tympani heißt Basilarmembran. Wird das Ohr beschallt, so schwingt der Stapes mit der ovalen Fenstermembran, sodass die Schallenergie durch das ovale Fenster in die Perilymphe der Scala vestibuli eintritt. Die Flüssigkeit ist nicht kompressibel und weicht daher aus; dabei werden Reissner-Membran, Scala media und Corti-Organ nach unten gedrückt [Abb. 2: weißer Pfeil; Abb. 3: roter und weißer Pfeil]. Dadurch wird auch die Flüssigkeit in der Scala tympani verdrängt. Diese ist ebenfalls inkompressibel, kann aber ausweichen, weil die Membran des runden Fensters gegen das Mittelohr gewölbt werden kann [Abb. 2]. Im weiteren Verlauf einer Schallschwingung schließt sich die umgekehrte Bewegung an: Steigbügel und ovales Fenster werden wieder nach außen, die Reissner-Membran und Corti-Organ nach oben, das runde Fenster nach innen bewegt. Da bei einem Schallereignis Schallschwingung auf Schallschwingung das ovale Fenster ein- und auslenken, führt dieser Vorgang zu einer ständigen Auf- und Abwärtsbewegung [Auslenkung] der Membranen und des Corti-Organs des Innenohrs. Im Querschnitt [Abb. 3, Abb. 4] sieht man die Rezeptorzellen [Haarzellen], die in Stützzellen eingebettet sind. Rezeptorzellen und Stützzellen bilden das Corti-Organ. Der Mensch besitzt 3 Reihen äußerer Haarzellen sowie eine Reihe innere Haarzellen, die an ihrem oberen Ende jeweils bis zu 100 haarähnliche, submikroskopische Fortsätze, die Stereozilien [Sinneshärchen], besitzen [Abb. 4]. Über ihnen (in der Scala media) befindet sich die Tektorialmembran [Abb. 3, Abb. 4], welche die Spitzen der längsten Stereozilien der äußeren Haarzellen soeben berührt. Dadurch befindet sich zwischen Tektorialmembran und Haarzellen ein schmaler, mit Endolymphe gefüllter Spalt. Die oben geschilderte schallinduzierte Auf- und Abwärtsbewegung [Auslenkung] von Scala media und Corti-Organ führt zu einer Relativbewegung [Scherbewegung] zwischen Tektorialmembran und CortiOrgan. Diese sind nämlich an unterschiedlichen übereinander liegenden Orten parallel aufgehängt [Abb. 5]. Wenn beide gleichzeitig ausgelenkt werden, entsteht eine Parallelverschiebung zwischen beiden Strukturen. Weil die Tektorialmembran die Spitzen der längsten Stereozilien der äußeren Haarzellen berührt, kann sie bei dieser Relativbewegung die Stereozilien umbiegen [abscheren, auslenken, deflektieren] und dadurch die Sinneszellen adäquat reizen [Abb. 5]. Endolymphe Tektorialmembran Stereozilien äußere Haarzellen innere Haarzelle efferente Synapsen Stützzellen Basilarmembran afferente Synapse Hörnerv Abb. 4. Querschnitt durch das Corti-Organ. Das Schema zeigt die Anordnung von Sinneszellen und afferenten Nervenfasern. Die äußeren Haarzellen haben Kontakt mit der Tektorialmembran, die inneren Haarzellen haben keinen Kontakt. Dadurch werden die äußeren Haarzellen durch die Tektorialmembran gesteuert. Die inneren Haarzellen werden durch die äußeren gesteuert (Pfeil). H ren.qxd 24.11.2003 15:10 Seite 4 Hören Die inneren Haarzellen hingegen haben keinen direkten Kontakt mit der Tektorialmembran. Man stellt sich vor, dass der schmale endolymphatische Flüssigkeitsfilm zwischen Tektorialmembran und Haarzellen auf Grund der Scherbewegung unter der Tektorialmembran hin- und hergleitet [subtektoriale Endolymphströmung, Pfeil in Abb. 5]. Dadurch sollen die Stereozilien der inneren Haarzellen mitgenommen und ausgelenkt werden. Man spricht von hydrodynamischer Kopplung. Durch die Abscherung der Sinneshärchen wird der Transduktionsprozess in den Haarzellen eingeleitet. Befindet sich die Haarzelle in Ruhe, so beträgt das Ruhemembranpotential der Haarzellen [Abb. 6] zwischen rund -40 mV [innere Haarzellen] und rund -70 mV [äußere Haarzellen]. Eine Deflektion der Stereozilien infolge des Schallreizes führt zur Änderung des Membranpotentials [Rezeptorpotential, Abb. 7]. Für den zugrunde liegenden Transduktionsprozess wird angenommen, dass eine Abscherung der Zilien die Öffnung von Ionenkanälen an der Spitze der Zilien hervorruft. Interessanterweise ziehen kleine Fäden von den Spitzen der meisten Stereozilien zur Wandung der dahinter stehenden Zilie [sog. Tip links, Abb. 8]. Werden die Stereozilien in Erregungsrichtung deflektiert, so werden die Tip links gespannt. Man stellt sich vor, dass durch den Zug K+durchlässige Kanäle geöffnet werden und dass durch diese Kanäle K+-Ionen aus der K+-reichen Endolymphe in die Haarzelle einströmen und zu deren Depolarisation führen [Abb. 9]. Zur Repolarisation besitzt die Zelle kaliumspezifische Ionenkanäle [z. B. KCNQ4-Kanäle] an ihrer seitlichen Zellmembran. Eine Depolarisation der Haarzelle öffnet mehr dieser Kanäle. Dadurch können K+-Ionen die Haarzelle durch die seitliche Zellmembran wieder verlassen, und das Membranpotential wird wieder angehoben. Signaltransformation von der Sinneszelle zum Hörnerven Die durch die Abscherung der Stereozilien bewirkten Ionenströme und Potentialänderungen innerer Haarzellen – nicht jedoch äußerer Haarzellen – setzen an ihrem unteren Ende den Neurotransmitter* Glutamat frei [Abb. 9]. Glutamat diffundiert durch den dort befindlichen schmalen synaptischen Spalt und bindet an AMPA-Rezeptoren der Hör-Nervenzellmembran. Dadurch wird ein postsynaptisches Potential ausgelöst, das zu Nervenaktionspotentialen führt. Die Reizung der afferenten Nervenfasern und damit die Weitergabe der im Schallreiz enthaltenen Information erfolgt ausschließlich von den inneren Haarzellen. Interessanterweise haben die äußeren Haarzellen nämlich eine ganz andere Funktion, die später besprochen wird. Frequenzselektivität: Grundlage des Sprachverständnisses Das gesunde Ohr hat eine erstaunlich gute Fähigkeit, Tonhöhen zu unterscheiden, wenn die Töne sukzes- H.P. Zenner Tektorialmembran Endolymphe Perilymphe äußere Haarzelle Basilarmembran a Tektorialmembran Sog Endolymphe Auslenkung Perilymphe Deflexion der Stereozilien (Sinneshärchen) b Abb. 5. Erregungsmechanismus der Haarzellen. Schematischer Ausschnitt aus der Schneckentrennwand. Gezeigt ist die Anordnung der Haarzellen zwischen Tektorial- und Basilarmembran: A in Ruhe, äußere Haarzellen berühren die Tektorialmembran, innere berühren sie nicht; B bei Auslenkung der Schneckentrennwand. Die wanderwelleninduzierte Auslenkung der Schneckentrennwand einschließlich Haarzelle nach oben führt zu einer Deflektion der Stereozilien. Die Stereozilien der äußeren Haarzellen werden durch die Tektorialmembran deflektiert. Die Stereozilien der inneren Haarzellen schert der Sog der Endolymphströmung (Pfeil) ab. 0 mV [Na+] [K+] +85 mV Scala media [K +] [Na+] Scala vestibuli +155 mV –70 mV Haarzelle [Na+] [K+] 0 mV Scala tympani Abb. 6. Endokochleäres Potential. Die Scala media mit positivem endokochleären Potential und auffällig hoher Kaliumkonzentration in der Endolymphe. Das apikale Ende der Haarzellen ragt in die Scala media hinein. Beim Transduktionsvorgang öffnet die Haarzelle Ionenkanäle, sodass aufgrund der elektrochemischen Potentialdifferenz vermutlich Kaliumionen aus der Scala media in die Haarzelle einströmen. H ren.qxd 24.11.2003 15:10 Seite 5 H.P. Zenner Hören/auditives System Schallreiz 110 db SPL Rezeptorpotential Membranpotential [mV] -60 -70 -80 Abb. 7. Potentialmessung an Haarzellen mit Mikroelektroden. Schnelle positive und negative Potentialabweichungen vom -70 mV-Wert bei Beschallung. Diese Potentialänderungen heißen Rezeptorpotentiale. Abb. 8. Tip links. A Rasterelektronenmikroskopie der Tip links. Man sieht Fäden, die von der Spitze eines Stereoziliums zum dahinter stehenden Stereozilium ziehen (Abb. Dr. Koitchev, Tübingen). B Eine akustische Reizung führt zu einer Anspannung der Spitzenfäden (Tip links), die zur Öffnung von Ionenkanälen in den Spitzen von Stereozilien führen soll. C Eine Hemmung erlaubt eine Entspannung der Spitzenfäden mit Schluss von Ionenkanälen. Scala tympani Scala media medial lateral Tip link öffnet Kanal Stereozilium (Sinneshärchen) K+ Schallsignal Deflektion Endolymphe hohe [K+ ] Perilymphe niedrige [K+ ] Depolarisation öffnet Kanal Zellkern 0 mV K+ (Repolarisation) ? [Ca 2+ ] afferenter Transmitter [Gehirn] +85 mV Abb. 9. Transduktionschritte von Haarzellen. Das Schallsignal führt zu einer Deflektion des Haarbündels, wodurch sich apikale Ionenkanäle öffnen.Kaliumionen strömen in die Zelle.Die Folge ist eine Depolarisation der Zelle. Die Depolarisation führt (in inneren Haarzellen) zur Freisetzung des afferenten Transmitters (vermutlich Glutamat), wodurch die afferenten Nervenfasern stimuliert werden. Bei äußeren Haarzellen führt Zytoplasma der sie zur Kontraktion der Zellen. Gleichzeitig steigert die Depolarisation die Haarzelle Öffnungswahrscheinlichkeit von kaliumspezifischen Kanälen in der Hörnervenfaser laterobasalen Zellwand (in äußeren Haarzellen sind es z.B.Typ-C-Kanäle). Sie erlauben die Repolarisation der Zelle. Äußere Haarzellen elongieren, innere beenden die Transmitterfreisetzung. siv angeboten werden. Bei 1000 Hz können Änderungen um 0,3 %, also 3 Hz wahrgenommen werden. Ist diese Frequenzunterschiedsschwelle verschlechtert, kann der Kranke Sprache kaum noch verstehen. Für die Ausbildung dieser sog. Frequenzselektivität besitzt die Cochlea einen zweistufigen Mechanismus. Erklingt ein Ton, werden die schlauchförmige Scala media und Corti-Organ gleichzeitig in die bereits geschilderten ständigen Auf- und Abwärtsbewegungen, also in Vibrationen versetzt [um nicht alle H ren.qxd 24.11.2003 15:10 Seite 6 Hören Strukturen nennen zu müssen, sprechen manche auch verkürzend von Vibrationen der Basilarmembran, meinen jedoch alle genannten kochleären Strukturen]. Diese Vibrationen bleiben nun nicht auf den Bereich in unmittelbarer Nähe von Steigbügel und rundem Fenster beschränkt, sondern bilden eine Welle aus, die von der Schneckenbasis bis zur Schneckenspitze wandert [Wanderwelle]. Während ihrer Wanderung nimmt ihre Amplitude in einem ersten Schritt etwas zu, wird in einem zweiten Schritt bis zu tausendfach zu einer hohen Welle mit sehr scharfer Spitze verstärkt und nimmt im weiteren Verlauf plötzlich wieder ab [Abb. 10]. Diese Verstärkung ist bei niedrigen und mittleren Schalldrücken besonders auffällig. Die Vibration der scharfen Spitze der Wanderwelle soll dann den oben genannten subtektorialen Flüssigkeitsfilm der Endolymphe deutlich verschieben [Endolymphströmung] und dadurch die inneren Haarzellen stimulieren. Letztere geben anschließend, nach dem oben genannten Transduktionsprozess, den afferenten Transmitter an die afferenten Hörnervenfasern weiter. Für das Verständnis der Frequenzselektivität ist von grundlegender Bedeutung, dass sich diese scharfe Spitze für jede Tonfrequenz an einem anderen Ort in Längsrichtung der Basilarmembran ausbildet. Hohe Frequenzen erzeugen das Maximum der Wanderwelle in der Nähe der Schneckenbasis, mittlere Frequenzen in der Schneckenmitte, tiefe Frequenzen an der Schneckenspitze. Man spricht daher vom Ortsprinzip [Ortstheorie, Tonotopie] der Wanderwelle. Das Maximum der Wanderwelle ist auf die äußeren Haarzellen zurückzuführen. Bei niedrigem Schalldruck erzeugen die äußeren Haarzellen nämlich zusätzliche mikromechanische Schwingungen in der Reizfrequenz. Äußere Haarzellen können sich bis zu 20.000-mal pro Sekunde [20 kHz] verkürzen und verlängern [Abb. 11]. Die zusätzliche Schwingungsenergie entsteht nur an dem jeweils frequenzcharakteristischen, eng umschriebenen Ort der Basilarmembran. Nur dort werden jeweils einige wenige, wahrscheinlich ca. 50, äußere Haarzellen durch die Tektorialmembran gereizt, die zusätzlich erzeugte Schwingungsenergie wird scharf lokalisiert an die inneren Haarzellen abgegeben: Die Wanderwelle wird in dem sehr eng umschriebenen Bereich verstärkt. Durch diesen kochleären Verstärkungsprozess wird die hohe Frequenzselektivität des gesunden Ohres, die Voraussetzung für das Sprachverständnis ist, erreicht. Der für diesen Verstärkungsprozess verantwortliche Motor ist das Protein Prestin [von ital. „presto“ = schnell] in der Zellmembran der äußeren Haarzellen. H.P. Zenner ovales Fenster rundes Fenster Scala vestibuli Wanderwelle Basilarmembran + Corti-Organ = Cochleare Trennwand Scala tympani Abb. 10. Die Wanderwelle in den kochleären Membranen. Die Wanderwelle startet nahe den Fenstermembranen und läuft die Basilarmembran entlang in Richtung Schneckenspitze. In Abhängigkeit von der jeweiligen Frequenz des Schallsignals bilden die kochleären Membranen ein Amplitudenmaximum an einem jeweils eng umschriebenen Ort aus. Abb. 11. Die Motilität äußerer Haarzellen als Grundlage des kochleären Verstärkers. Links Haarzelle in Ruhe; Mitte stimulierte äußere Haarzelle: die Haarzelle verkürzt sich;Rechts anschließend elongiert die Haarzelle. Die Längenänderungen „pumpen“ mechanische Energie in die Wanderwelle, wodurch diese tausendfach verstärkt und die Endolymphströmung unter der Tektorialmembran so stark wird, dass die inneren Haarzellen gereizt werden. (Abb: Dr. R. Zimmermann,Tübingen) Informationsübertragung und Verarbeitung im Zentralnervensystem Die von der inneren Haarzelle als Folge des Transduktionsprozesses ausgelöste Transmitterfreisetzung wird in Form einer neuronalen Erregung über Hörnerv, Hirnstamm und Hörbahn bis zum auditorischen Kortex H ren.qxd 24.11.2003 15:10 Seite 7 H.P. Zenner Hören/auditives System innere Haarzelle A B Aktionspotentiale bei geringem Schalldruck Rate der Aktionspotentiale nimmt zu Bestfrequenz hier! leise Töne afferente Nervenfaser Schalldruckverlauf laute Töne Abb. 12. Kodierung des Schalldrucks im Hörnerv. A Bei leisen Tönen werden nur die Fasern mit der dazugehörigen Bestfrequenz gereizt; B bei zunehmender Lautstärke nimmt die Zahl der Aktionspotentiale in den Fasern zu; C bei weiterer Steigerung des Schalldrucks kann die Zahl der Aktionspotentiale nicht mehr gesteigert werden. Daher werden zusätzlich Nachbarfasern aktiviert (Rekrutierung). im Temporallappen weitergeleitet. Dabei sind wenigstens 5–6 hintereinander geschaltete, durch Synapsen* verbundene Neurone* beteiligt. Sie besitzen Kollaterale und Interneurone*, die zu einer ausgedehnten neuronalen Vernetzung des auditorischen Systems führen. Der Hörnerv überträgt die transduzierten Signale Radiatio aus der Cochlea ins Zentralnervensystem [ZNS]. Er auditiva besteht aus einer großen Zahl afferenter sowie teilCorpus genic. med. Colliculus inferior weise auch efferenter [d.h. aus dem Gehirn kommenLemniscus lateraalis Auditorischer der] Nervenfasern. 90 % der afferenten Nervenfasern Kortex im Gyrus Nuclei lemnisci later. haben nur eine Synapse mit einer einzigen, nämlich temporalis N. cochlearis dorsalis einer inneren Haarzelle. An das Gehirn werden also transversus N. cochlearis ventralis im Wesentlichen Informationen von den inneren Haarzellen weitergeleitet. Da jede Haarzelle nach Nucleus statoacusticus dem Ortsprinzip [siehe oben] einer ganz bestimmNucleus olivaris sup. Kochlea ten Tonfrequenz zugeordnet ist, wird die mit einer Corpus trapezoideum bestimmten Haarzelle synaptisch verbundene Hörnervenfaser bei Beschallung des Ohrs mit dieser Abb. 13. Schematische Darstellung der zentralen Hörbahn. ganz bestimmten Frequenz optimal erregt. Diese Frequenz heißt charakteristische Frequenz oder Bestfrequenz einer Einzelfaser. Die Zeitdauer eines Schallreizes wird durch die Zeitdauer der Aktivierung der Nervenfasern kodiert, die Höhe des Schalldruckpegels durch die Entladungsrate verschlüsselt [Abb. 12]. Allerdings kann eine einzelne Nervenfaser eine bestimmte Entladungsrate nicht überschreiten, sondern erreicht ab einem bestimmten Schalldruck einen Sättigungsbereich. Trotzdem kann die Information nach höherer Lautstärke weitergegeben werden, da dann eine zunehmende Zahl benachbarter Fasern aktiviert wird [Rekrutierung, Recruitment]. Die zweiten Neurone gehen vom ventralen Nucleus cochlearis aus. Ein Teil zieht zur oberen Olive der gleichen Seite, ein Teil kreuzt zur oberen Olive der anderen Seite [Abb. 13]. Ebenso kreuzen die afferenten Fasern vom dorsalen Kern zum Nucleus lemnisci lateralis der Gegenseite. Dadurch ist jedes Innenohr mit der rechten und der linken Hörrinde verbunden. Außerdem können in den Nervenzellen des Olivenkomplexes erstmals im Verlauf der Hörbahn binaurale [= von beiden Ohren aufgenommene] akustische Signale miteinander verglichen werden. Die höheren Neurone verlaufen von der oberen Olive zum Teil auf der gleichen Seite, zum Teil auf der Gegenseite nach jeweils neuer Umschaltung zum Colliculus inferior und anschließend zum Corpus geniculatum mediale. Schließlich ziehen die Afferenzen als Hörstrahlung [Radia- H ren.qxd 24.11.2003 15:10 Seite 8 Hören/auditives System tio acustica] zur primären Hörrinde [HeschlQuerwindung] des Temporallappens. H.P. Zenner Schallquelle Die einfache Kodierung des ersten und von Teilen des zweiten Neurons wandelt sich grundlegend ab dem dorsalen Nucleus cochlearis und weiter zunehmend mit jedem höheren Neuron. Zwar wird das Ortsprinzip bis zum auditorischen Kortex beibehalten, d.h., dass bestimmte Schallfrequenzen an bestimmten Orten der Hörrinde oder der auditorischen Kerne repräsentiert sind. Zusätzlich besitzen jedoch beispielsweise einige vom dorsalen Nucleus cochlearis ausgehende Neurone kollaterale Verschaltungen, die teils exzitatorisch, teils inhibitorisch zusätzliche Laufstrecke wirksam sind [On-off-Neurone]. Die Folge ist, dass Laufzeitdifferenz einzelne Neurone des dorsalen Cochleariskerns bei Schallreiz stets gehemmt werden. Eine grundsätzliche Eigenschaft der höheren Neurone der Hörbahn ist es, nicht auf reine Sinustöne, sondern auf bestimmte Eigenschaften eines Schallmusters [z. B. Sprachmuster] zu reagieren. So Abb. 14. Auditorische Raumorientierung. Die Laufzeitdifferenz eines gibt es Fasern, die bei einer bestimmten Schallfre- Tons zwischen beiden Ohren wird im zentralen auditorischen System quenz aktiviert, durch höhere oder tiefere Töne je- verarbeitet und dient der lateralen Schallquellenlokalisation. doch gehemmt werden. Auch gibt es Neurone, die auf eine Frequenzzunahme, und solche, die auf eine Frequenzabnahme [Frequenzmodulation] reagieren, wobei zusätzlich der Grad der Modulation von Bedeutung sein kann. Andere Zellen sprechen nur auf die Amplitudenänderung eines Tons an. Diese Spezialisierung von Neuronen auf bestimmte Eigenschaften eines Schallmusters ist im auditorischen Kortex noch ausgeprägter. Neurone können hochspezialisiert auf den Beginn oder das Ende, auf eine Mindestzeitdauer oder eine mehrfache Wiederholung, auf bestimmte Frequenz- oder Amplitudenmodulationen eines Schallreizes sein. Man nimmt daher an, dass diese bis zur Hörrinde zunehmende Spezialisierung der Neurone auf bestimmte Eigenschaften des Schallreizes es erlaubt, Muster innerhalb des Schallreizes herauszuarbeiten und für die kortikale Beurteilung vorzubereiten [Informationsverarbeitung]. Das gesprochene Wort oder Musik bestehen aus derartigen Mustern, die wir trotz eines Störschalls [z. B. Umgebungsgeräusche] erkennen können. Die Richtung einer Schallquelle kann geortet werden. Dieses räumliche Hören oder auditorische Raumorientierung geschieht durch das zentrale Hörsystem. Dort finden sich in bestimmten Bereichen, etwa der oberen Olive oder dem Colliculus inferior, auf Raumorientierung hochspezialisierte Neurone, welche die von den beiden Ohren ankommenden Folgen von Aktionspotentialen miteinander vergleichen. Dazu müssen zunächst einmal beide Ohren einigermaßen normal hören [binaurales Hören]. In der Regel liegen Schallquellen nicht genau in der durch den Kopf definierten Mittelebene [Mediansagittalebene], sondern irgendwie seitlich. Dann ist die Schallquelle von einem Ohr weiter entfernt als vom anderen. Der Schall trifft dadurch am entfernteren Ohr später und leiser ein [Abb. 14]. Das auditorische System ist dabei in der Lage, Intensitätsunterschiede von nur 1 dB und Laufzeitunterschiede bis hinab zu 3 x 10-5 s sicher zu erkennen. Eine derartig minimale Schallverstärkung tritt bei einer Abweichung der Schallquelle von 3 Grad von der Mittellinie auf. Laufzeit- und Intensitätsdifferenzen erlauben zwar die Bestimmung des Raumwinkels, nicht jedoch die Entscheidung, ob sich die Schallquelle oben, unten, vorne oder hinten befindet. Hierzu ist die Form der Ohrmuschel, die eine Richtcharakteristik besitzt, bedeutsam. Je nachdem, in welchem Winkel das Schallsignal auf die Ohrmuschel auftrifft, wird es minimal verformt. Offenbar können diese dadurch modulierten [„verzerrten“] Schallmuster zentral erkannt und ebenfalls zur Bildung eines Raumeindrucks verwandt werden.