6 1.2 Strukturen der Nozizeption und der Schmerzverarbeitung
Werbung

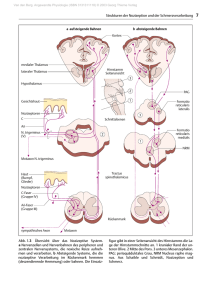
Van den Berg. Angewandte Physiologie (ISBN 3131311118) © 2003 Georg Thieme Verlag 6 1 Physiologie des Schmerzes und der Nozizeption 1.2 Strukturen der Nozizeption und der Schmerzverarbeitung Thomas Weiß und Hans-Georg Schaible 1.2.1 Übersicht über das nozizeptive System Wie oben dargestellt, bilden Nervenfasern und Nervenzellen, die ausschließlich oder vorwiegend der Aufnahme und der Verarbeitung noxischer Reize dienen, das nozizeptive System. Abb. 1.3 zeigt schematisch und stark vereinfacht eine Übersicht über das nozizeptive System. Auf der linken Seite ist das aszendierende System dargestellt. Hierzu gehören als erste Neurone die nozizeptiven Primärafferenzen, auch Nozizeptoren genannt. Sie dienen der Aufnahme noxischer Reize in den Organen, z. B. in der Haut. Nozizeptoren projizieren zum Rückenmark bzw., Nozizeptoren des Kopfbereichs projizieren zum Trigeminuskern im Hirnstamm. Im Rückenmark bzw. Trigeminuskern werden synaptisch nozizeptive Neurone erregt. Solche Neurone sind entweder Interneurone mit kurzer Projektion oder Neurone mit aszendierenden Axonen (sie bilden die diversen Schmerzbahnen) oder Neurone, die in motorische bzw. vegetative Reflexbögen eingebunden sind. Aszendierende Rückenmarkneurone aktivieren über die Schmerzbahn das nozizeptive thalamokortikale System. Letzteres ist erforderlich, um bewusste Schmerzempfindungen zu erzeugen. Neben dem aszendierenden System gibt es ein absteigendes (deszendierendes) System, das vom Hirnstamm seinen Ausgang nimmt (Abb. 1.3, rechte Seite). Deszendierende Bahnen sind entweder inhibitorisch oder exzitatorisch. Die nächsten Abschnitte werden die nozizeptiven Neurone in den einzelnen Ebenen des Nervensystems beschreiben. 1.2.2 Nozizeptive Primärafferenzen Nozizeptive Primärafferenzen, Nozizeptoren, sind Nervenzellen des peripheren Nervensystems, die auf die Detektion von potentiell oder aktuell gewebeschädigenden Reizen spezialisiert sind. Die meisten Organe bzw. Körperstrukturen sind mit Nozizeptoren versorgt. Die Dichte der Innervation ist allerdings für die verschiedenen Organe unterschiedlich. Reich innerviert mit Nozizeptoren ist die Haut, die Viszeralorgane besitzen dagegen weniger Nozizeptoren, und bei manchen Organen (Leber, Niere) scheint vor allem die Kapsel mit Nozizeptoren versorgt zu sein. Struktur der Nozizeptoren Nozizeptoren sind Primärafferenzen mit freien Nervenendigungen und langsam leitenden Axonen (Abb. 1.3). Eine freie Nervenendigung ist eine dünne unmyelinisierte Faserendigung, die anders als die Endigung niederschwelliger Mechanorezeptoren keine spezialisierte korpuskuläre Struktur aufweist. Teilweise sind diese Faserendigungen von Schwannzellen bedeckt, und teilweise grenzt die Membran der Nervenfaser direkt an das umgebende Gewebe. Es wird vermutet, dass an diesen Membranabschnitten die Transduktion der noxischen Reize erfolgt. Transduktion ist die Umwandlung mechanischer, thermischer und chemischer Reize in ein Rezeptorpotential oder Generatorpotential (siehe auch Bd. 2, Kap. 9). Dieses führt durch den Vorgang der Transformation zur Bildung von Aktionspotentialen. Neben der Aufnahme von Reizen haben freie Nervenendigungen eine weitere Funktion, nämlich die Freisetzung von Mediatoren aus der Nervenfaser (siehe unten). Diese Mediatoren beeinflussen Vorgänge im Gewebe. Während niederschwellige Mechanorezeptoren der Haut (z. B. Berührungsrezeptoren) und anderer Organe schnellleitende, myelinisierte Axone besitzen, haben Nozizeptoren entweder dünn myelinisierte oder unmyelinisierte Axone. Die dünn myelinisierten Nervenfasern sind Aδ-Fasern, die Leitungsgeschwindigkeiten von 2,5 – 30 m/sec aufweisen. In den Aδ-Fasern entsteht das Aktionspotential (Transformation) am ersten Schnürring. Die meisten Nozizeptoren besitzen unmyelinisierte Fasern, also C-Fasern. Diese haben Leitungsgeschwindigkeiten ⬍ 2,5 m/sec, die meisten im Bereich von etwa 1 m/sec. Es ist bis heute nicht bekannt, an welcher Stelle der C-Fasern die Transformation stattfindet, also die Umwandlung des Rezeptorpotentials in das Aktionspotential. Der Zellkörper eines Nozizeptors liegt im Spinalganglion (Abb. 1.3). Er steht über den axoplasmatischen Transport im Stoffaustausch mit den peripheren und spinalen Endigungen der Nervenfaser. Der axoplasmatische Transport wird durch ein intrazelluläres System bewerkstelligt, das Substanzen vom Zellkörper zu den Endigungen und umgekehrt transportiert. Auf diesem Weg werden Mediatoren und Moleküle für Ionenkanäle und Rezeptoren in die sensorischen Endigungen transportiert und dort in die Membran eingebaut. Interessanterweise werden viele Ionenkanäle und Rezeptoren auch in die Membran des Zellkörpers im Spinalganglion selbst integriert. Dies führt dazu, dass der Zellkörper ähnliche Rezeptoreigenschaften aufweisen kann wie die sensorische Endigung im Gewebe.








![Laminierung [Bearbeiten]](http://s1.studylibde.com/store/data/002675562_1-75537cc091b485163ea524f6877d4c25-300x300.png)