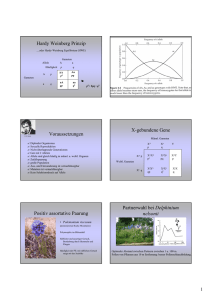
Chapter 4 - Biologie
Werbung

Kap. 6 aus meinem Buch “Do species exist? – Principles of taxonomic classification”, Wiley Blackwell 2012: die für das Modul 446 "Grundlagen der Biodiversität und Evolution" weniger wichtigen Teile sind blau markiert wichtige Begriffe, die auch Prüfungsstoff sind, sind rot markiert spezifische Hinweise zum Exkursionsinhalt sind grün markiert allgemein wichtige Aussagen sind fett markiert Kapitel 6. Merkmalsunterschiede, Polymorphismen und die polytypische Spezies Vorbemerkung Merkmalsunterschiede sind nicht notwendigerweise auch Artunterschiede Was sind Mutanten, Variationen, Aberrationen, Formen, Phasen und Phänons? Was sind Rassen, Unterarten und Subspezies? Was sind Morphen? Was sind Mutanten (im taxonomischen Sinn)? Geschlechtsdimorphismus Allele Vielfalt Welche evolutionäre Lebensdauer hat allele Vielfalt? Stabile Polymorphismen - Der Selektionsvorteil ist die Vielfalt Populationsunterschiede in der allelen Häufigkeitsverteilung im Vergleich zu Artunterschieden Das Beispiel der teilziehenden Vögel Innerartliche Morphen beim Widderchen Zygaena ephialtes Der Polymorphismus der Schnirkelschnecke Cepaea nemoralis Das Beispiel des Finkenschnabels Innerartlicher Polymorphismus beim Darwinfinken Geospiza fortis Die Morphen der Strumpfbandnatter Thamnophis ordinoides Die polymorphe genetische Grundlage der Verstädterung einiger Vogelarten Die Mimikry-Morphen bei weiblichen Schwalbenschwänzen der Gattung Papilio Die Morphen des brutparasitierenden Kuckucksweibchens (Cuculus canorus) Vorbemerkung Das vorliegende Kapitel handelt von den Widersprüchlichkeiten, die auftreten, wenn das Bewusstsein herrscht, dass das, was merkmalsverschieden ist, verschiedene Arten sein müssen. Die auch heute noch verbreitete Auffassung, deutlich merkmalsverschiedene Organismen seien verschiedene Arten, und weniger merkmalsverschiedene Organismen seien Unterarten, hält einer konsequenten Prüfung nicht stand. Auch auf DNA-Ebene können das Ausmaß der Unterschiede und die Gewissheit, dass diese den Artrang anzeigen, nicht gleichgesetzt werden. Nur ein winziger Bruchteil des Genoms hat etwas mit Artunterschieden zu tun. Der weitaus größte Teil des Genoms ist zwischen den Arten nicht verschieden, solange die Arten evolutionär nicht allzu alt sind. Bei verwandten Arten zeigen sich Artunterschiede oft nur in ganz wenigen Genen; der Großteil der Gene hat mit Artunterschieden nichts zu tun. Und umgekehrt gilt Folgendes: Fast alle Gene eines Organismus treten in verschiedenen Allelformen auf, und diese unterscheiden sich zwischen den Organismen. Das führt zu erheblichen Unterschieden zwischen den Organismen, auch wenn diese zu ein und derselben Art gehören. Vergleicht man nun zwei verwandte Arten miteinander, so sind erwartungsgemäß beide durch eine hohe innerartliche Diversität ausgezeichnet, und es fragt sich, welches nun intraspezifische und welches interspezifische Unterschiede sind. Merkmalsunterschiede allein definieren nicht die Einheiten, die man Spezies nennt. Da nun die meisten Gene nichts damit zu tun haben, dass es sich hier um zwei verschiedene Arten handelt, können zwei Individuen derselben Art zueinander genetisch signifikant verschiedener sein, als es zwei Individuen verschiedener Arten zueinander sind (Pääbo 2001 5884; Smith et al. 5775; Avise et al. 1998 5771). Auch äußerlich können sich zwei Organismen verschiedener Arten viel weniger voneinander unterscheiden als zwei Organismen derselben Art. Merkmalsunterschiede sind nicht notwendigerweise auch Artunterschiede In der ersten Auflage von „The Origin of Species“ sagt Darwin auf Seite 424 (zitiert in Ghiselin 1997 5325): „Jeder Biologe weiß, dass die beiden Geschlechter sich manchmal enorm in den meisten wichtigen Merkmalen unterscheiden. Kaum ein einziges Merkmal haben männliche adulte Tiere bei bestimmten Cirripediern (Rankenfußkrebse) mit den Hermaphroditen derselben Art gemeinsam, und trotzdem denkt niemand auch nur im Traum daran, sie zu trennen.“ Treffender kann nicht ausgedrückt werden, wie problemgeladen es ist, den Standpunkt zu vertreten: Dieser Organismus sieht anders aus; also müsste er eine eigene Art sein. Der Konkurrent, aber auch Freund Darwins, Alfred Russel Wallace, titulierte eine 1858 erschienene Publikation: „Über die Tendenz von Varietäten, unbegrenzt vom Originaltypus abzuweichen“ (Wallace 1858 5880). Damit kommt zum Ausdruck, dass Arten alles andere als homogen sind. Arten neigen dazu, in ihren Merkmalen „unbegrenzt“ zu variieren. Wie kann es dann angehen, das Phänomen der Merkmalsunterschiede zum Artkriterium zu machen? Zweifellos sind die Merkmale, die die Organismen innerhalb einer Art voneinander unterscheiden, nicht von anderer Natur als die Merkmale, die die verschiedenen Arten voneinander unterscheiden. In diesem Buch sind mit dem Begriff „Merkmal“ immer intrinsische Eigenschaften gemeint. Relationale Eigenschaften werden nicht mit dem Begriff „Merkmal“ bezeichnet (siehe Kap. 4). Das Spektrum der Merkmale reicht von komplexen Merkmalen (wie etwa die Schnabelfarbe eines Vogels), die in populären Artbestimmungsbüchern aufgelistet sind, bis hin zur modernen Methode des Barcoding, worunter man bestimmte DNA-Sequenzen versteht, an denen Arten fast automatisch identifiziert werden können, so wie die Ware am Waren-Code an der Kasse eines Supermarkts (siehe Kap. 5). Merkmale sind Beobachtungskonsequenzen aus primär anders definierten theoretischen Artkonzepten. Merkmalsunterschiede sind kein Kriterium einer Speziation, es sei denn, man wäre Anhänger des phänetischen Artkonzepts (Sneath and Sokal 5920 1973). Anhänger aller anderen Artkonzepte können nicht von Merkmalsunterschieden auf Artunterschiede schließen. Auch wenn es in der täglichen Praxis oft so ist, dass Artunterschiede an Merkmalsunterschieden erkannt werden können, so ist es doch prinzipiell nicht möglich, Merkmalsunterschiede zum Kriterium des Vorliegens unterschiedlicher Arten zu machen. Die Tatsache, dass Arten an Merkmalen unterschieden werden können, ist nicht gleichbedeutend mit der Annahme, dass Organismen mit anderen Merkmalen auch andere Arten sein müssen. Alle Artkonzepte außer dem phänetischen Artkonzept definieren Arten anders. Dass sich Arten oft an Merkmalen unterscheiden, ist eine sekundäre Folgeerscheinung, die keineswegs immer zutrifft. Es gibt sogenannte kryptische Arten, die sich an Merkmalen (fast) nicht unterscheiden lassen, und es gibt polytypische Arten, deren Organismen sich deutlich voneinander unterscheiden, obwohl sie zu ein und derselben Art gehören. Wie schon der klassische Evolutionsbiologe und Paläontologe G.G. Simpson gesagt hat: „Zwei Individuen sind nicht deswegen Zwillinge, weil sie sich so ähnlich sehen, sondern, ganz im Gegenteil, sie sehen sich so ähnlich, weil sie Zwillinge sind. Und ganz genauso gehören zwei Individuen nicht deswegen zu ein und demselben Taxon, weil sie sich so ähnlich sehen, sondern sie sehen sich so ähnlich, weil sie zu demselben Taxon gehören“ (Simpson 1961 5902). Die Annahme, die Angehörigen einer biologischen Spezies sähen einander grundsätzlich alle ähnlich, wird durch zahlreiche Ausnahmen widerlegt. Stabile Polymorphismen führen zur Ungleichheit vieler Merkmale, ohne dass dies auf verwandtschaftlicher Entfernung beruht oder darauf, dass Sexualbarrieren diese Unterschiede aufrecht erhalten. Intraspezifischer Polymorphismus scheint häufiger vorzukommen, als früher angenommen wurde (Smith et al. 2001 5775). Die Merkmale, die die unterschiedlichen Morphen (siehe unten) charakterisieren, sind für die menschliche Wahrnehmung und Wertung auffälliger oder weniger auffällig, aber das Phänomen des innerartlichen Polymorphismus selbst scheint bei vielen Arten realisiert zu sein. Merkmalsunterschiede innerhalb der Population zwischen den am gleichen Ort lebenden Morphen einer Art können durchaus stärker sein als zwischen den Individuen von geographisch entfernten Populationen (Rassen), und sie können auch stärker sein als zwischen verschiedenen Arten (Smith et al. 2001 5775). Polymorphismen (das sind Typenunterschiede am gleichen Ort) und ebenso auch Rassen (das sind Typenunterschiede der Organismen zwischen entfernten Orten) haben primär nichts mit Artbildung zu tun. Die Hervorbringung mehrerer Organismen in unterschiedlichen Formen und Verhaltensweisen kann durchaus ein Wesensmerkmal einer Spezies sein, mit einem Selektionsvorteil für die Spezies selbst, ohne dass dies das Anzeichen der Entstehung separater Arten wäre. Es kann für das Überleben mancher Arten wichtig sein, dass ihre Angehörigen in unterschiedlicher Gestalt existieren. Es ist nicht immer so, dass unterschiedliche Formen und Verhaltensweisen Kriterien für unterschiedliche Spezies wären, noch sind es immer die Vorstufen einer beginnenden Speziation. Angehörige ein und derselben Art können sich in durchaus mehr Merkmalen voneinander unterscheiden als Angehörige verschiedener Arten (Smith et al. 2001 5775; Avise et al. 1998 5771). Das liegt unter anderem daran, dass sich genetische Verschiedenheiten innerhalb ein und derselben Art über länger andauernde evolutionäre Zeitperioden ansammeln können, ohne dass es in diesen Zeiträumen zur Artaufspaltung kommen muss (Christoffersen 1995 5108). Substantielle Polymorphismen können im sympatrischen Verbreitungsbereich zu erheblichen innerartlichen Verschiedenheiten führen. Es können unterschiedliche Morphen entstehen, die die Art zu einer polytypischen Spezies machen. Zusätzlich können in geografischen Entfernungen auch Rassen entstehen, worunter man merkmalsverschiedene Populationen versteht, die als Anpassungen an bestimmte geografische Örtlichkeiten mehr oder weniger voneinander abweichen (siehe unten). Seien es nun Morphen oder Rassen, intraspezifische Verschiedenheiten sind durchaus normal für eine Art. Was sind Mutanten, Variationen, Aberrationen, Formen, Phasen und Phänons? Es werden unnötig viele Vokabeln verwendet, um innerartliche Varianten zu benennen. Man findet Begriffe wie Mutanten, Variationen, Aberrationen, Formen, Phasen, Phänon usw. Neben dem Geschlechtsdimorphismus als sehr drastischem Beispiel gibt es noch zahlreiche andere Beispiele für erhebliche äußere Unterschiede zwischen den Angehörigen ein und derselben Spezies. Jungtiere und Larven können sich deutlich von geschlechtsreifen Tieren unterscheiden. Das hat in klassischer Zeit bereits Linné dazu veranlasst, den immaturen Habicht (Accipiter gentilis) wegen seines längs gestreiften Bauchgefieders im Unterschied zum geschlechtsreifen Habicht mit quergestreiftem Bauchgefieder als eigene Art zu beschreiben (Mayr 2000 4643). Ein eindrucksvolles Beispiel für polymorphe Gefiederfärbung zeigen viele Arten der großen bis mittelgroßen Reiher. Hier gibt es sowohl Rassen als auch Morphen als auch deutliche Unterschiede zwischen noch nicht geschlechtsreifen immaturen Tieren und den fortpflanzungsfähigen Tieren. Viele Reiherarten können entweder weiß sein oder schwarz. Beim amerikanischen Blaureiher (Egretta caerulea) sind die immaturen Vögel im Jahr ihrer Geburt schneeweiß gefärbt, die geschlechtsreifen Tiere aber schwarz mit leicht zimtroten Ton (Farbbild). Noch eindrucksvoller aber sind bei anderen Reiherarten die verschiedenen Morphen, die gleichaltrig im selben Gebiet leben und jeweils ohne Paarungsbarriere miteinander zu ein und derselben Genflussgemeinschaft gehören. Bei fünf Reiherarten treten weiße und dunkle bis schwarze Morphen im adulten Federkleid nebeneinander auf. Beim Kanadareiher (Ardea herodias), einem nahe Verwandten des Altwelt-Graureihers (Ardea cinerea), tritt neben der normalen grauen Morphe eine komplett weiße Form auf. Der Seidenreiher (Egretta garzetta) ist normalerweise ganz weiß gefärbt. Es treten in seltenen Fällen aber auch total schwarze Morphen auf. Zentralamerikanische Rötelreiher (Egretta rufescens), afrikanische Küstenreiher (Egretta gularis) und australische Riffreiher (Egretta sacra) können entweder ein weißes Gefieder haben, oder sie können auch ganz schwarz sein (Farbbild). Beide Morphen treten bei den letzten drei Arten relativ häufig nebeneinander auf und verpaaren sich artspezifisch jeweils miteinander, offenbar ohne assortative Bevorzugung von weiß oder schwarz. Ebenso können Schneegänse (Anser caerulescens) entweder weiß oder dunkelbraun sein (Farbbild). Eleonorenfalken (Falco eleonorae) können auf der Unterseite schiefergrau oder orange sein. Die Weibchen des Tagfalters Kaisermantel (Argynnis paphia) können ockergelb oder grünschwarz sein. Letztere tragen den Namen „forma valesina“ und sind einer anderen Art zum Verwechseln ähnlich, nämlich dem Kardinal Argynnis pandora (valesina und pandora können wir evtl. in Bükk finden). Die unterschiedlichen innerartlichen Typen haben z.T. zusätzliche taxonomische Namen erhalten, die den wissenschaftlichen Art- und Unterartnamen angehängt werden: z.B. variatio, forma usw. Ernst Mayr verwendet den Begriff „Phänon“ und meint damit eine phänotypisch unterschiedliche Serie innerhalb der Art (Mayr und Ashlock 1975 4948). Nach Mayr kann eine Art also aus mehreren Phänons bestehen. Dieser Begriff hat sich aber nicht durchgesetzt. Die Vielfalt der in der Taxonomie verwendeten Begriffe - „variatio“, „aberratio“, „forma“ etc. - ist unnötig, zumal diese Begriffe von verschiedenen Autoren mit unterschiedlicher Bedeutung verwendet werden. Es ist durchaus uneindeutig, was mit „aberratio“ eigentlich gemeint ist. Man kommt zur Benennung innerartlicher Typen mit drei Begriffen aus und kann damit die polymorphe Vielfalt innerhalb einer Genflussgemeinschaft eindeutig bezeichnen. Die drei Begriffe sind Rasse, Morphe und Mutante. Alle weiteren Bezeichnungen sind für die Verständigung überflüssig. Was sind Rassen, Unterarten und Subspezies? Der Begriff der Rasse ist synonym zu Unterart oder Subspezies (siehe Kap. 7). Der Begriff bezeichnet eine geografische Teilpopulation innerhalb einer weit verbreiteten Genflussgemeinschaft. Die Entstehung von Rassen hat eine ganz bestimmte Ursache: Über geringe geografische Entfernungen schafft es der Genfluss, die Angehörigen der Population in ihrer Merkmalsausstattung homogen zu halten, weil die gegenseitige Vermischung der Organismen keine Sonderentwicklung von Subpopulationen aufkommen lässt. Jede abweichende Separatentwicklung wird durch Rückkreuzung mit Organismen der ursprünglichen genetischen Ausstattung wieder rückgängig gemacht. Sobald aber eine gewisse geografische Entfernung überschritten ist, wird der Genfluss zwischen den entfernten Organismen schwach. Er schafft es nicht mehr, separate Entwicklungen zu verhindern. Daher setzen sich als Anpassung an die unterschiedlichen lokalen Habitatgegebenheiten eigene Entwicklungswege in den geografisch entfernten Populationen durch. Solche lokal angepassten Gruppen nennt man Rassen, Unterarten oder Subspezies. Rassen können nicht am selben Ort nebeneinander vorkommen, weil sie sich dann vermischen würden. Rassen sind also immer geografische Sonderentwicklungen. Rassen überschneiden sich in ihrem Vorkommen, aber immer nur in den Grenzgebieten. Falls Rassen überhaupt nicht geografisch getrennt sind, dann ist das ein Zeichen sekundärer Verlagerung der ursprünglichen Wohngebiete, und dieser Zustand ist wegen der Vermischung nicht von langer evolutionärer Dauer. Die Rassen vermischen sich und verschwinden in absehbarer Zeit. Falls es nicht zur Vermischung kommt, dann liegt das daran, dass Reproduktionsbarrieren bestehen. Dann jedoch spricht man konsequenterweise von Arten und nicht von Rassen. Ein verbreitetes Missverständnis von Art und Rasse ist es, stark unterschiedene Populationen als Arten und weniger unterschiedene Populationen als Rassen zu bezeichnen. Diese Sichtweise führt schon nach wenigen Anwendungen zu Inkonsequenzen und Missverständnissen. Der Begriff der Art beinhaltet immer Mechanismen, die die Angehörigen verschiedener Populationen daran hindern, ihre Gene auszutauschen. Solche Populationen sind dann deswegen verschiedene Arten, weil intrinsische oder extrinsische Barrieren den Genfluss zwischen ihnen verhindern. Etwas ganz anderes ist die Rasse. Der Begriff der Rasse beinhaltet immer eine Einschränkung des Genaustausches wegen der geografischen Entfernung, so dass sich aus diesem Grund evolutionäre Eigenentwicklungen durchsetzen. Zwischen den Rassen aber gibt es fließende Übergänge, dort, wo die Rassen geografisch aufeinanderstoßen. Diese Übergangszonen stellen eine klinale Verbindung zwischen den Rassen dar. In den Übergangszonen kommt es zu unbegrenzter Vermischung der Angehörigen verschiedener Rassen miteinander, weil keine assortative Paarung herrscht (siehe Kap. 7). Zwar kommt es auch zwischen Arten zu Vermischungen, solange die Arten noch nahe genug miteinander verwandt sind; jedoch sind Art-Vermischungen (Bildung von Hybriden) immer Ausnahmen, weil wegen der Paarungsbarrieren assortative Paarung herrscht. Ein eindrucksvolles Beispiel dafür, dass sich entfernte Populationen an die lokalen Gegebenheiten anpassen und dann in ihren Merkmalen voneinander differieren, liefert der Haussperling (Passer domesticus). Dieses Beispiel dokumentiert auch gleichzeitig, wie schnell es zu Anpassungen und damit zur Rassenbildung kommt. Die ursprüngliche Vorstellung, Rassenbildung sei ein Prozess der Jahrzehntausende, muss gründlich revidiert werden (Johnston und Selander 1964 5866). Der Haussperling war ursprünglich eine paläarktische Vogelart. In der Mitte des 19. Jahrhunderts wurden die ersten Exemplare in Nordamerika eingeführt und haben sich dann zu Beginn des 20. Jahrhunderts über den ganzen nördlichen Kontinent ausgebreitet. Im Laufe von nur etwa hundert Generationen entwickelten sich dann beträchtliche lokale Anpassungen hinsichtlich Färbung, Schnabel- und Flügelgröße, die einzelne Rassen voneinander unterscheiden. Jedoch sind alle Rassen über klinale Übergänge miteinander verbunden. Unter einem klinalen Übergang versteht man den allmählichen Übergang von einer geografischen Rasse in die andere, ohne dass eine scharfe Grenze besteht. In der Zone des klinalen Übergangs leben Mischtypen zwischen den beiden benachbarten Rassen. Leider ist in diesem Falle, wie in vielen ähnlich gelagerten Fällen, nichts über die genetische Kompatibilität der einzelnen Rassen miteinander bekannt. Es wäre von besonderem Interesse, ob die entfernten Rassen überhaupt noch miteinander erfolgreich kreuzbar sind: Die (vor allen Dingen postzygotische; siehe Kap. 7) genetische Kompatibilität ist jedoch weder mit klassischen Kreuzungsversuchen noch mit den moderneren Techniken der Messung des Genflusses zwischen den Rassen je gemessen worden. Es wäre von besonderer Wichtigkeit zu wissen, ob die F1-Nachkommen einer Kreuzung der entfernt lebenden Organismen eine Schnabel- und Flügelgröße aufweisen, die sie gegenüber reinrassigen Artgenossen konkurrenzfähig macht. Schnabel- und Flügelgröße sind offenbar sehr sensible Anpassungen an die spezifischen örtlichen Gegebenheiten. Außerdem ist zu erwarten ist, dass die Rassenhybride hinsichtlich dieser Merkmale phänotypisch intermediär ausgestattet sind. Die Konsequenz von beidem wäre, dass die Angehörigen entfernter Rassen trotz ihrer klinalen Verbindung nicht miteinander kreuzbar sind. In der Rückschau ist es bemerkenswert, dass die Entdeckung der distinkten Rassenunterschiede beim Haussperling in Nordamerika bereits fast ein halbes Jahrhundert zurückliegt (Johnston und Selander 1964 5866) und damit in die Zeit der „großen Synthese“ hineinfällt (siehe Kap. 7), als das Artkonzept der Reproduktionsgemeinschaft seine Blütezeit hatte. Die Spezies wurde damals als ein fest verbundener Genpool betrachtet (Mayr 1963 5377), aus dem die einzelnen Organismen nicht ausscheren können, weil sie durch genetische Rekombination immer wieder in den Genpool zurückgeholt werden und damit alle Organismen einer Art immer wieder einander angeglichen werden (siehe Kap. 7). Obwohl Daten über die Nicht-Kreuzbarkeit fehlen, hätte die Annahme einer zumindest eingeschränkten Kreuzbarkeit zwischen den entfernten Populationen einer Art nahe liegen müssen. Und diese Erkenntnis hätte stärkere Implikationen auf das Verständnis der biologischen Art als Reproduktionsgemeinschaft haben müssen. Rasse und Art unterscheiden sich dadurch, dass die einzelnen Rassen, so verschieden sie auch sein mögen, durch klinale Übergänge miteinander verbunden sind. Demgegenüber sind Arten Populationsgruppen, zwischen denen der Genfluss abgebrochen ist. Dieser Definitionsunterschied zwischen einer Rasse und einer Art bedeutet aber nicht, dass alle konkret existierenden Beispiele eindeutig als Rasse oder Art voneinander unterschieden werden können. Wie immer in der Biologie, ist die Existenz einer klaren Definition nicht gleichbedeutend damit, dass auch die real in der Natur existierenden Dinge ebenso klar voneinander unterschieden werden können. Es gibt Grenzfälle, die sowohl die Definitionskriterien einer Rasse als auch die einer Art erfüllen, aber eben keiner von beiden Definitionen hundertprozentig genügen, so dass es der subjektiven Entscheidung des Einzelnen überlassen ist, diese Grenzfälle als Rasse oder als Art einzustufen. Dieser unbefriedigende Zustand ist jedoch nicht gleichbedeutend mit der Folgerung, dass auch die Definition unscharf sei. Die Definitionen für Art und Rasse sind scharf, so wie jede Definition scharf ist (siehe Kap. 3). Ein Beispiel dafür ist das Artenpaar Rabenkrähe-Nebelkrähe (Corvus corone und Corvus cornix). [Im Bükk befinden wir uns im Nebelkrähen-Gebiet] Die beiden Arten sind geografisch getrennt, berühren sich aber in einer schmalen Kontaktzone, wo sie miteinander ziemlich erfolgreich hybridisieren (siehe Kap. 7). Die geografische Trennung und der klinale Übergang sind Kriterien dafür, die beiden Krähen als Rassen einzustufen. Jedoch ist die Berührungszone an der Grenze des gesamten Verbreitungsgebietes von Raben- und Nebelkrähe nur etwa 50 km breit. Zudem sind die Mischpaarungen nicht völlig zufällig, sondern signifikant assortativ (Saino und Villa 1992 4466). Das heißt, dass sich in erster Wahl Rabenkrähe mit Rabenkrähe und Nebelkrähe mit Nebelkrähe verpaaren. Erst in zweiter Wahl, wenn der conspezifische Partner nicht gefunden wird, z.B. weil etwas später in der Brutperiode die meisten Partner bereits vergeben sind, kommt es zu den Mischpaarungen. Daher überwiegen die Kriterien, die beiden Krähen als Arten statt als Rassen einzustufen. An dieser Stelle sei hinzugefügt, dass es keinen Erkenntnisgewinn bringt, das RabenkräheNebelkrähe-Problem mit der Bemerkung abzutun, hier läge halt ein noch laufender Speziationsprozess vor, der noch nicht vollendet sei. Diese Auffassung entspringt dem rein menschlichen Verlangen nach Stabilität, die in der Natur aber nicht gegeben ist. Alles fließt, mal langsamer und mal schneller, und jede Art befindet sich in einem noch laufenden Speziationsprozess, der nicht zu Ende ist. Alte historische Daten sprechen dafür, den gegenwärtigen Zustand des Nebeneinanders von Raben- und Nebelkrähe als eher stabil zu bezeichnen. Es ist eher wahrscheinlich, dass ein gleichartiges Nebeneinanders von Raben- und Nebelkrähe schon vor Jahrhunderten existiert hat und auch in weiteren Jahrhunderten noch so bleiben wird (Saino und Villa 1992 4466). Es ist in der Biologie eher angebracht, nach den Gesetzmäßigkeiten zu suchen, die ein Artenpaar trotz gegenseitiger Hybridisierung langfristig bestehen lassen, als dem Problem auszuweichen und von einem vorübergehenden Zustand zu sprechen. Das klarste Beispiel dafür, dass die Rassenvielfalt in einer Spezies der Spezies selber zugute kommen kann und eben nicht so zu verstehen ist, dass dies auf eine kommende Artbildung hinausläuft, liefern die Guppies (Poecilia reticulata) auf Trinidad (Magurran 1999 4334). Artbildung ist nicht die unausweichliche Folge einer hohen Populationsdifferenzierung. Die Guppies auf Trinidad zerfallen in eine Vielzahl kleiner Rassen, die in ihrer Merkmalsverschiedenheit die Diversität gut etablierter Arten übersteigen. Die molekularen Analysen haben ergeben, dass die einzelnen Rassen seit einer halben Million Jahre bestehen. Sie sind seit mehr als einer Million Generationen getrennt. Trotzdem sind die Rassen reproduktiv in breiten Übergängen miteinander verbunden. Es kommt weder zu einer homogenisierenden Versmischung der Rassenunterschiede, noch kommt es zum Aufbau von Reproduktionschranken wie bei einer anderen Fischfamilie, den afrikanischen Cichliden, die einer raschen Speziation unterliegen (siehe Kap.5 und7). Es ist ein naturwissenschaftlich völlig ungeklärtes Geheimnis, warum bestimmte Tiergruppen in evolutionär kurzen Zeiten viele neue Arten bilden, während andere, manchmal nahe verwandte Tiergruppen über lange Zeiten keine Arten bilden (Seehausen et al. 2008 5875; Verheyen et al. 2003 5360). Warum kommt es bei den Guppies nicht zur Speziation? Möglicherweise spielen die rigorosen Verfolgungsjagden eine Rolle, mit der die Guppymännchen die Weibchen für die Fortpflanzung „erobern“. Dieses Verhalten setzt den üblichen biologischen Mechanismus des „female choice“ weitgehend außer Kraft (Magurran 1999 4334). Normalerweise ist die selektive Auswahl der Männchen durch die Weibchen die Hauptursache dafür, dass es zur Speziation kommt. Abweichende Männchen werden bei den meisten höheren Tierarten von den Weibchen nicht zur Fortpflanzung zugelassen, wodurch sich Artschranken bilden können, die dann erhalten bleiben. Dieses Wahl- und Entscheidungsverhalten geht bei den meisten Arten von den Weibchen aus. Es wird „female choice“ genannt (siehe Kap. 7). Es ist eine der der entscheidenden Kräfte, die zur Artbildung führen. Die Männchen wirken gegen diese Kraft. Bei den Guppies aber durchbrechen die Männchen diese Schranken und erhalten damit den Genfluss auch zwischen sehr verschieden differenzierten Rassen aufrecht. Was sind Morphen? Der zweite wichtige Begriff für eine ganz andere Form innerartlicher Polytypie ist der Begriff der Morphe. Während jedoch Rassen immer geografisch voneinander entfernt leben, kommen Morphen nebeneinander (syntop) am gleichen Ort vor. Ein wichtiger Unterschied zwischen Rassen und Morphen ist, dass Rassen sich nicht voneinander abgrenzen müssen, weil sie ohnedies durch Distanz weitgehend isoliert sind, auch wenn es noch fließende Übergänge gibt, allerdings nur an den geografischen Berührungszonen. Morphen dagegen müssen durch Mechanismen geschützt sein, die die Integrität von zwei oder wenigen Typen aufrecht erhalten, ohne dass es zu intermediären Mischlingen kommt. Das ist ein elementarer Unterschied zwischen einer Rasse und einer Morphe. Es ist aber auch ein Unterschied zur Art. Denn Morphen haben keine Reproduktionsbarriere zwischen den verschiedenen Typen. Hätten sie das, dann wären es Arten. Morphen verdanken ihre Di- oder Polytypie also nicht dem über die Entfernung schwach gewordenem Genfluss, sondern besonderen Mechanismen, die die Polytypie trotz starkem Genfluss erhalten. Existieren nur zwei Morphen, dann ist es ein „Dimorphismus“, gibt es mehr als zwei, dann ist es ein „Polymorphismus“. Das bestbekannte Beispiel für Morphen ist der Geschlechtsdimorphismus, die gleichzeitige Existenz von Weibchen und Männchen. In den meisten Fällen liegt dem Di- bzw. Polymorphismus auch ein genetischer Polymorphismus zugrunde. Es werden dann zwei oder mehrere sehr distinkte Typen hervorgebracht, ohne dass Intermediäre entstehen können, wie das z.B. bei der Entstehung von Männchen und Weibchen im Falle der genetischen Geschlechtsbestimmung realisiert ist. Sollten trotzdem intermediäre Zwischentypen zwischen den beiden Geschlechtern entstehen, so geht das auf einen (meist genetischen) Unfall zurück. Aber es muss nicht immer eine genetische Grundlage für die Aufrechterhaltung distinkter Typen ohne Mischtypen gegeben sein. Es gibt auch andere Fälle für die Aufrechterhaltung distinkter Morphen, denen keine genetische Steuerung zugrunde liegt. Hier übernimmt die Selektion die Steuerung. Das bedeutet, dass zwar intermediäre Typen zwischen den distinkten Morphen entstehen, dass diese aber keine oder nur eine geringe Überlebenschance haben. Die intermediären Zwischentypen stellen keine optimale Anpassung an irgendwelche vorhandene Umweltgegebenheiten dar. Sie werden daher in jeder Generationenfolge nach ihrer Entstehung durch die Selektion ausgemerzt und treten kaum in Erscheinung. Eine besondere Form von Polytypie stellen die unterschiedlichen Phänotypen während der ontogenetischen Entwicklung (Larven und Imagines) und die zwei oder mehr unterschiedlichen Generationen im Lebenszyklus derjenigen Pflanzen und Tiere dar, die über einen Generationswechsel verfügen, wie z.B. bei Trematoden (Leberegeln), vielen Coelenteraten (Hohltieren) und Pflanzen. In diesen Fällen liegen keine genotypischen Unterschiede vor; es treten nur verschiedene Phänotypen in Erscheinung. Ein Beispiel für das Auftreten verschiedener Phänotypen während der ontogenetischen Entwicklung sind die Entwicklungsstadien holometaboler Insekten. Raupen und Puppen gleichen den fertigen Schmetterlingen überhaupt nicht. Immature Entwicklungsstadien vieler Tierarten sehen häufig ganz anders aus als die adulten geschlechtsreifen Tiere. Warum auch muss ein unreifer Organismus so aussehen wie ein geschlechtsreifer Organismus. Unreife Tiere leben manchmal in einer ganz anderen Umgebung als die erwachsenen Tiere, haben eine andere Ernährungsweise usw. Daher können es sich immature Tiere leisten, äußerlich anders auszusehen als die adulten Tiere. Noch wichtiger ist, dass es für immature Tiere keinen Selektionsdruck für die Entwicklung artspezifischer Partnererkennungsmerkmale gibt. Daher brauchen immature Tiere nicht über die Fähigkeit zu verfügen, sich selber von den Angehörigen einer anderen Art zu unterscheiden. Daher sehen sich immature Tiere manchmal sehr ähnlich, ohne dass man sie als verschiedene Arten voneinander unterscheiden kann. Noch eindrucksvoller sind die verschiedenen Phänotypen, die bei den Pflanzen und Tieren auftreten, die durch einen Generationswechsel charakterisiert sind. Hier alternieren verschiedene Phänotypen miteinander innerhalb eines einzigen Lebenszyklus. Beispiele für solche Arten sind die Trematoden, viele Coelenteraten und Pflanzen. Der Lebenszyklus dieser Organismen besteht aus alternierenden Stadien mit jeweils distinktem Phänotyp, die sich auf unterschiedliche Weise fortpflanzen. In einigen Fällen wechseln sich diploide Stadien mit haploiden Stadien ab. In anderen Fällen alternieren Stadien, die sich vegetativ fortpflanzen, mit solchen Stadien, die sich bisexuell reproduzieren. In allen Fällen jedoch besteht der Lebenszyklus bei den Organismen mit Generationswechsel aus distinkten Phänotypen, die in ihrem Äußeren sehr verschieden aussehen. Für den Taxonomen sind diese Beispiele von Polytypie ein zusätzliches Erschwernis und ein erneutes Beispiel dafür, dass vom unterschiedlichen Aussehen nicht der Schluss auf das Vorliegen unterschiedlicher Arten gezogen werden kann. Die Natur hat die Arten nicht für die Menschen gestaltet, damit er sie auch erkennen kann. Auch in den Fällen, in denen im Zyklus eines Jahres mehrere sich jeweils fortpflanzende Generationen aufeinanderfolgen, sehen die Vertreter verschiedener Generationen (z.B. Frühjahrs- und Sommergeneration bei Schmetterlingen) oft verschieden aus. Ein eindrucksvolles Beispiel für einen derartigen Saisonpolymorphismus tritt bei vielen Tagfaltern auf, die (verursacht durch unterschiedliche Temperatureinwirkung während des Puppenstadiums) eine Frühjahrs- und dann eine anders aussehende Sommergeneration entwickeln. Das in Europa bekannteste Beispiel ist das Landkärtchen (Araschnia levana). Die kühlen Wintertemperaturen bedingen bei den im April schlüpfenden Imagines ein hellbraunes bis oranges Farbmuster, während die warmen Junitemperaturen zur Folge haben, dass die im Juli schlüpfende zweite Faltergeneration überwiegend schwarz gefärbt ist (diesen Falter sehen wir im Bükk). Bei den meisten Tagfaltern, die im Jahr zwei oder mehr Generationen ausbilden, gibt es Merkmalsunterschiede zwischen den Generationen. Sie sind jedoch meist nicht so augenfällig, wie das beim Landkärtchen der Fall ist. Diese Beispiele für Entwicklungs- und Generationspolymorphismen stellen jedoch nur zyklische Entwicklungsstadien dar. Es sind keine echten Polymorphismen, weil es lediglich aufeinanderfolgende Stadien ontogenetisch gesteuerter Entwicklungsprozesse sind. Ganz anders liegt die Sache, wenn die phänotypisch zu Tage tretende Polytypie einen genetischen Polymorphismus zur Grundlage hat, wenn den einzelnen Morphen also unterschiedliche Genome zugrunde liegen, wie das z.B. bei vielen Formen des Geschlechtsdimorphismus der Fall ist. In diesen Fällen ist die populationsgenetische Grundlage für die Existenz von Morphen ein stabiler Allel-Polymorphismus. Solche Beispiele einer genotypisch basierten Polytypie gibt es in der Natur in verschiedener Form. Neben dem Sexualdimorphismus gibt es den ökologische Polymorphismus. Verschiedene Morphen derselben Art sind an verschiedene ökologische Nischen angepasst und ernähren sich von verschiedenen Futterquellen. Daneben gibt es den MimikryPolymorphismus. Verschiedene Morphen derselben Art ahmen ganz verschiedene Tierarten nach, um dadurch in unterschiedlicher Form gegen Feinde geschützt zu sein. Trotzdem gehören sie alle zu ein und derselben Art. Was sind Mutanten (im taxonomischen Sinn)? Neben Rassen und Morphen gibt es noch einen weiteren innerartlichen „Morphotyp“, der allerdings im Gegensatz zur Rasse und Morphe keine taxonomische Bedeutung hat. Er ist als genetischer Unfall zu betrachten, hat selektive Nachteile und wird daher meist bald wieder ausgemerzt. Gemeint sind gelegentlich auftretende Mutanten, wie z.B. die Albinos, ein teilweise oder gänzlich farbloser Phänotyp aufgrund einer angeborenen Störung in der Biosynthese der Melanine. Albinos als genetische Unfälle sind nicht zu verwechseln mit den bei einigen Tieren vorkommenden weißen Morphen, z.B. bei mehreren Reiherarten (s.o.). Diese haben andere genetische Ursachen und sind nicht als „Unfälle“ zu betrachten. Der Begriff der Mutante ist in der Genetik sehr allgemein definiert und bezeichnet üblicherweise jede Form von genetischer Veränderung im Genom, von der Punktmutation bis zur Chromosomen- oder Genommutation. Für die taxonomische Gruppenbildung sind nur die Mutanten zu berücksichtigen, die einen diagnostisch klar erkennbaren Phänotyp erzeugen. Und nur das ist hier in diesem Kontext mit „Mutante“ neben Morphen und Rassen gemeint. Solche abweichenden Typen haben Taxonomen immer wieder verwirrt. Sie können fremde Arten vortäuschen (Farbbild athalia). Das Spektrum der Mutanten reicht von abweichenden Farben und Farbmustern bis hin zu entstellten Gestalten, wie z.B. dem sog. „Mongoloiden“, einem Menschen als Träger des "Down Syndroms" aufgrund des dreifachen Vorliegens des Chromosoms Nummer 21. Häufig tritt auch der sog. Melanismus auf. Darunter versteht man abweichende Typen mit dunkler bis schwarzer Körperfärbung, die z.B. bei verschiedenen Säugetieren beobachtet werden. Melanismus beruht auf übermäßiger Ablagerung von Melaninen. Besonders bekannt sind die melanistischen Mutanten bei mehreren Arten der Feliden (Katzenartige), und hier wiederum besonders beim schwarzen Panther, einer Mutante des Leoparden (Panthera pardus). Die genetische Ursache für den Melanismus beim Leoparden ist ein einziges Gen, das rezessiv vererbt wird. Interessanterweise treten schwarze Panther in unterschiedlichen geografischen Regionen in stark unterschiedlicher Häufigkeit auf. Da es sich um eine rezessive Mutante handelt, lässt das einen positiven Selektionsvorteil vermuten. Falls der melanistische Panther als polymorpher Angehöriger der Leopardenpopulationen eine evolutionäre Bedeutung hat, müsste er konsequenterweise als Morphe bezeichnet werden. Geschlechtsdimorphismus Das deutlichste Beispiel für drastische Merkmalsunterschiede trotz gleicher Artzugehörigkeit sind die Unterschiede zwischen Männchen und Weibchen. Beim Geschlechtsdimorphismus zeigen sich schon rein morphologisch (wie der Name sagt) deutliche Unterschiede zwischen den Organismen innerhalb ein und derselben Art. Bei sehr vielen Tierarten gelingt es uns leichter, die Männchen von den Weibchen zu unterscheiden, als die verschiedenen Arten auseinander zu halten. Ein Beispiel dafür sind die Enten, deren Weibchen oft nur von Spezialisten der Art nach zugeordnet werden können, während die Unterscheidung von Männchen und Weibchen der gleichen Art über die meiste Zeit des Jahres überhaupt kein Problem ist. Das zeigt, dass ein rein phänetisches Artkonzept (siehe Kap.5), das die biologische Spezies als nichts anderes betrachtet als eine Gruppe merkmalsähnlicher Organismen (Sneath and Sokal 1973 5920) nicht konsequent durchgehalten werden kann. Spätestens beim Geschlechtsdimorphismus endet die Möglichkeit, das eine Art zu nennen, was anders aussieht. Zwei dimorphe Geschlechter müssen nach den reinen Kriterien der Phänetik zu zwei verschiedenen Arten erklärt werden. Um dies zu vermeiden, muss zusätzlich zum phänetischen Artkonzept ein zweites Artkonzept mit hinzugezogen werden, nämlich das der Genflussgemeinschaft. Sonst würde ein biologisch wenig sinnvolles System aufgebaut werden. Dass der Mensch spontan zuerst einmal an verschiedene Arten denkt, wenn er das unterschiedliche Aussehen von Männchen und Weibchen wahrnimmt, beweist niemand besser als Linné selbst, der die beiden Geschlechter der Stockente (Anas platyrhynchos) zunächst für verschiedene Arten gehalten hat und ihnen unterschiedliche Namen gegeben hat (Mayr 2000 4643). Beim Geschlechtsdimorphismus handelt es sich aber nicht nur um Äußerlichkeiten. Die geschlechtsspezifisch unterschiedlichen Merkmale betreffen auch die Stoffwechsel- und Neurophysiologie, das Verhalten und die Muster der gewebespezifisch transkribierten Gene (Transkriptom) sowie der kodierten Proteine (Proteom). Körperform und Körpergröße, Nahrungsverwertung und Hirnfunktion, Reaktionen auf bestimmte Umweltreize und die Aktivitätsmuster der Gene in bestimmten Geweben. Alle diese Merkmale können zwischen den Geschlechtern verschieden sein. Bei nahe verwandten Arten kann die Gesamtheit der Unterschiede zwischen den zwei Geschlechtern die Unterschiede zwischen zwei verschiedenen Arten deutlich übersteigen (Billeter et al. 2006 5603). Allele Vielfalt Innerartliche Polymorphismen zeigen sich in verschiedener Form. Es beginnt damit, dass die Allele eines jeden Gens mutieren können und damit in den Genomen einer Population in unterschiedlichen Varianten vorliegen. Die Organismen einer Population sind also sehr verschieden. Dieses Phänomen wird multiple Allelie genannt. Bei zwei zufällig ausgewählten haploiden Genomen tritt beim Menschen im Durchschnitt nach allen 1250 Basen ein Unterschied auf (Venter et al. 2001 4976). Das bedeutet, dass die meisten Gene heterozygot vorliegen und sich damit die einzelnen Organismen einer Population alle voneinander unterscheiden. Bei Drosophila melanogaster und vielen anderen Tieren ist der mittlere Wert heterozygoter Nukleotidverschiedenheit sogar noch deutlich höher als beim Menschen (Aquadro et al. 2001 5659). Bei Drosophila melanogaster differieren selbst kleine, zueinander homologe Gene von nur 1000 Basenpaaren im Schnitt an vier Positionen pro Gen, sofern die Gene unabhängig voneinander sind (Powell 1997 4843). Der Mensch ist, auch im Vergleich zu seinem nächsten Verwandten, dem Schimpansen, genetisch ziemlich homogen. Das liegt daran, dass Homo sapiens bei der Auswanderung aus Afrika durch einen engen populationsgenetischen „Flaschenhals“ (bottle neck) gegangen ist, durch genetische Drift also viele Allele verloren hat. Nun ist zwar der Schluss vom Genotyp auf den Phänotyp schwierig. Die meisten phänotypischen Unterschiede zwischen verwandten Arten haben nichts mit den Proteinkodierenden Strukturgenen zu tun (siehe Kap. 5). Die Entwicklung des Bauplans wird vielmehr von Enhancern und den jeweils im Gewebe verfügbaren Transkriptionsfaktoren gesteuert, nicht vom Proteincode der Strukturgene (Carroll et al. 2009 5836). Wird ein Transkriptionfaktor nicht rechtzeitig exprimiert oder ist ein gewebespezifischer Enhancer mutiert, so kommt es zu Unterschieden im Körperbau, in der Physiologie und im Verhalten. Das liegt meist nicht daran, dass einzelne Strukturgene verändert sind, die den Merkmalen zugrunde liegen, sondern es liegt daran, dass die einzelnen Strukturgene, obwohl sie selbst unverändert bleiben, anders reguliert werden. Aber trotzdem kann man davon ausgehen, dass es jeweils viele Hunderte, wenn nicht Tausende von Merkmalen sind, in denen sich jeder einzelne Organismus vom anderen in einer Population unterscheidet. Jeder Organismus ist in seiner Komplexität eine Einmaligkeit. Was also ist ein innerartlicher Unterschied im Vergleich zu einem zwischenartlichen Unterschied? Das Ausmaß der innerartlichen Variabilität hängt von der Populationsgröße ab; denn Inzucht verringert schnell die allele Vielfalt. Alle Arten, die einmal durch einen genetischen Engpass gegangen sind (Flaschenhals = bottleneck), weil ihre Individuenzahl stark reduziert war, haben eine geringe allele Vielfalt: Ihre Genome sind weitgehend homozygot. Beispiele dafür sind mehrere Katzenartige (Felidae), die stark verfolgt wurden und deren Reviere zerstückelt wurden. So zum Beispiel der Puma (Puma concolor) in Nordamerika, der in einigen isolierten Verbreitungsgebieten der USA jeweils nur noch in wenigen Individuen vorkommt, unter anderem in Florida (Roelke et al. 1993 2590). Der afrikanische Gepard (Acinonyx jubatus) war einstmals von Afrika aus weit über Vorderasien bis Indien verbreitet. Er ist offenbar schon vor Jahrhunderten durch einen Flaschenhals gegangen, so dass alle heute in Afrika lebenden Exemplare von einer sehr kleinen Stammgruppe abstammen (Harvey and Reed 1988 976). Welche evolutionäre Lebensdauer hat allele Vielfalt? Allele Mutanten werden schon im genetischen Grundunterricht als die Ausnahmen neben dem Wildtyp als Standard dargestellt. Das ist oft nicht unbegründet; denn die meisten Mutationen haben eine Einschränkung der Fitness zur Folge. Daher werden die meisten Mutanten durch Selektion wieder beseitigt. Der seltene Fall einer Steigerung der Fitness führt dazu, dass oft schon nach wenigen Generationen das mutante Allel das bisherige Wildtyp-Allel in der ganzen Population ersetzt. Das neue Allel wird in der Population „fixiert“. Dieses Phänomen hat zur Folge, dass die allele Vielfalt meistens kurzlebig ist, zumindest in kleinen Populationen, weil die Selektion für das Überleben nur einer Variante sorgt, die sich als die Fitteste erweist. Nun gibt es aber neben den selektionskontrollierten Allelvarianten auch solche Allele, die selektionsneutral sind. Es sind Mutanten, die weder Vor- noch Nachteile haben. Sie werden also nicht durch die Selektion beseitigt oder gefördert und sollten demnach in einer Population ständig zunehmen, weil ja immer wieder neue Mutationen auftreten. Das ist aber nicht der Fall. Auch neutrale Allelvarianten gehen nach und nach verloren. Dafür ist die genetische Drift verantwortlich, die über gewisse Zeiträume zum Verschwinden vieler Allelvarianten führt, besonders schnell in kleinen Populationen. Würde man einmal fiktiv annehmen, dass es über einen langen evolutionären Zeitraum überhaupt keine Mutationen gäbe, so würde die allele Vielfalt durch Drift allmählich völlig verloren gehen. Man kann sich dies durch ein einfaches analoges Beispiel klarmachen (Cavalli-Sforza und Cavalli-Sforza 1994 4117). In einer Population leben viele Menschen, die alle einen unterschiedlichen Namen tragen. Die Nachkommen einer Ehe erhalten den Namen des Vaters. Die Folge wäre, dass der Name eines jeden ausstirbt, der keine Kinder erzeugt oder der eine Frau ist. Also verringert sich die Vielfalt der Namen von Generation zu Generation, und das hat nichts damit zu tun, dass die Namen umständlich oder gar schädlich sind. Sie verschwinden durch reinen Zufall. Schließlich würde die gesamte Population nur noch einen einzigen Namen tragen. Das ist genau das, was in der Populationsgenetik als genetische Drift verstanden wird. Um die Namensvielfalt zu erhalten, gibt es nur zwei Mittel: Zuwanderung fremder Populationen oder die Neuvergabe von Namen. Letzteres wäre in der Genetik die Mutation. Aus all dem folgt, dass allele Vielfalt meist kurzlebig ist und überhaupt nur deshalb existiert, weil sie durch Mutation ständig wieder neu erzeugt wird. Stabile Polymorphismen - Der Selektionsvorteil ist die Vielfalt Das Prinzip, dass allele Vielfalt evolutionär kurzlebig ist, wird allerdings durch eine wichtige Ausnahme unterbrochen. Diese Ausnahme ist dann gegeben, wenn die Selektion den Polymorphismus fördert, wenn die Selektion sozusagen daran „interessiert“ ist, dass mehrere Allelformen in der Population coexistieren. Diese Situation wird stabiler Polymorphismus genannt. Während die Selektion normalerweise nur das einzelne Allel sieht, das ein Organismus gerade besitzt, und dieses fördert oder benachteiligt, gibt es Fälle, bei denen nicht die Qualität des einzelnen Allels über die Fitness entscheidet, sondern stattdessen das Vorhandensein mehrerer Allelvarianten innerhalb der Population. Angriffsziel der Selektion ist in diesen bemerkenswerten Fällen nicht die Güte des Einzelallels, sondern der Überlebensvorteil, der durch das Vorhandensein mehrerer paralleler Varianten in der Population gegeben ist. Der selektive Vorteil ist, dass die Population Vielfalt erhält. Die Population hat sowohl Individuen mit dem einen Allel als auch Individuen mit dem anderen Allel aufzuweisen. Auf diese Weise kann die Population flexibler auf variable Umweltschwankungen reagieren, da sie immer einige Organismen bereit hält, die schon präadaptiv auf die möglicherweise kommenden Umweltveränderungen eingestellt sind. Solche Allele werden also sozusagen von der Population in Reserve gehalten, um im entscheidenden Augenblick bei Bedarf rekrutiert werden zu können. Dieser Sachverhalt wird Präadaptation genannt. Der Selektionsvorteil der allelen Vielfalt in der Population sowie die Präadaptation sind darwinistisch nicht einfach zu verstehen und folglich umstritten. Es geht darum, dass die einzelnen Organismen im Endeffekt von einem Populationsvorteil profitieren. Das läuft auf Gruppenselektion hinaus, und Gruppenselektion wird von vielen Evolutionsbiologen abgelehnt. Das Verhältnis zwischen der Fitness eines Einzelorganismus und der Fitness einer Gruppe ist eine umstrittene Angelegenheit. Die multiple Allelie und der Polymorphismus sind eine Sache der Gruppenfitness. Theoretiker sind darüber zerstritten, ob die Gruppenfitness als Summe (oder Durchschnitt) der Individualfitness definiert werden kann (Okasha 2009 5932). Agiert die Selektion primär an Einzelorganismen, an Gruppen, an Genen oder an ganzen Arten (Okasha 2006 5924)? Es wird argumentiert, dass nur solche Einheiten die Einheiten der Selektion sein können, die sich reproduzieren und dass es eine notwendige Bedingung der Reproduktion ist, dass die Einheit der Nachkommen die Elterneinheit überleben kann (Okasha 2003 5931). Reproduziert sich eine Population? Oder ist die Fortpflanzung nur eine Sache der Einzelorganismen? In der Gruppenselektion-Kontroverse geht es darum, ob die Selektion jemals auf der Ebene von Gruppen operieren kann oder nur auf der Ebene von Einzelorganismen. Die meisten gegenwärtigen Evolutionsbiologen stehen der Gruppenselektion höchst skeptisch gegenüber, weil sie biologisch nicht plausibel erscheint (Okasha 2001 5930). Wie kann ein Allel lange Zeit in der Population überleben, wenn es keine Vorteile, sondern in vielen Fällen sogar Nachteile mit sich bringt? Es müsste doch eigentlich in absehbarer Zeit durch die Selektion ausgemerzt werden und damit auf die Dauer verschwunden sein. Soweit Allelpolymorphismen bis heute verstanden sind, beinhalten viele von ihnen keinen zeitnahen direkten Vorteil für das Individuum, das ein bestimmtes Allel trägt. Der Überlebensvorteil liegt stattdessen darin, dass die Population im Falle einer Umweltveränderung nicht ausstirbt. Das Überleben einer Population kann bei einer Umweltveränderung davon abhängen, dass gerade die wenigen Individuen überleben, die bisher nur Nachteile hatten. Warum sind die bisher benachteiligten Organismen (und damit ihre Allele) nicht ausgestorben? Diese Problematik ist nicht einfach zu lösen. Welche genetischen Gesetzmäßigkeiten beinhalten Mechanismen, die das Überleben von zeitweise nachteiligen Allelen erlauben? Ein Mechanismus leuchtet unmittelbar ein. Es ist die Rezessivität. Viele Allele, auch nachteilige, können lange Zeit überleben, wenn sie im heterozygoten Zustand nicht exprimiert werden, weil das homologe dominante Allel den Phänotyp bestimmt und damit allein der Selektion ausgesetzt ist. Solche Allele überleben sozusagen unter der „rezessiven Tarnkappe“. Es gibt Autoren, die darin sogar den biologischen Sinn der Diploidie begründet sehen. Ein weiterer genetischer Mechanismus, der unvorteilhafte Allele langfristig erhalten kann, ist die feste Koppelung dieser Allele mit benachbarten anderen Genen, mit denen sie am Chromosom verbunden sind und nur selten oder gar nicht durch Rekombination von diesen abgeschnitten werden. Wenn diese Nachbargene dann einen eindeutigen Selektionsvorteil haben, ziehen sie die gekoppelten Gene von Generation zu Generation mit sich, auch wenn diese einen Selektionsnachteil haben, der allerdings nicht allzu groß sein darf. Vielleicht trägt das folgende einfache Beispiel zum Verständnis bei: Auch der Geschlechtsdimorphismus ist ein stabiler Polymorphismus. Es gibt genügend Situationen, in denen es evident ist, dass es selektive Nachteile hat, einem bestimmten (und nicht dem anderen) Geschlecht anzugehören. Trotzdem wird das eine Geschlecht nicht durch Selektion verdrängt, weil der selektive Vorteil eben der ist, dass es zwei Geschlechter gibt. Das durch angeborene Eigenschaften oder benachteiligende ethische gesellschaftliche Zwänge benachteiligte Geschlecht hat deshalb keinen Selektionsnachteil, weil es sein Überleben dem Vorteil verdankt, dass zwei Geschlechter nebeneinander existieren, so benachteiligt das eine auch sein mag. Stabile Polymorphismen beschränken sich nicht auf den Geschlechtsdimorphismus. Sie treten in unterschiedlichen Formen auf und bestimmen das Bild der intraspezifischen Merkmalsunterschiedlichkeit. Bekannte Beispiele für stabile Polymorphismen sind der Haupthistokompatibilitäts-Genkomplex (MHC) und die Blutgruppen. In beiden Fällen liegt eine Allelvielfalt vor, die bezüglich bestimmter Allele die Angehörigen verschiedener Arten einander ähnlicher macht als die Angehörigen innerhalb ein und derselben Art. Beim Haupthistokompatibilitäts-Genkomplex (MHC) ist die allele Vielfalt älter als die evolutionäre Aufspaltung von Mensch und Schimpanse. Hinsichtlich bestimmter MHC-Allele sind sich bestimmte Menschen mit bestimmten Schimpansen ähnlicher als bestimmte Menschen untereinander (Figueroa et al. 1988 5611). Während der Polymorphismus der MHC-Allele die Speziation höherer Primaten prädatiert, scheint sich allerdings der Blutgruppen-Allel-Polymorphismus bei den Primaten parallel entwickelt zu haben. A-, B- und 0-Allele treten beim Menschen und bei Rhesusaffen auf; Schimpansen haben nur A und 0. Im Gegensatz zu früheren Auffassungen prädatiert dieser Polymorphismus allerdings nicht den Zeitpunkt der Aufspaltung in die einzelnen Primatenarten. Es handelt sich wahrscheinlich nicht um die Weitergabe einer „uralten“ ancestralen multiplen Allelie, sondern um einen erst nach der Aufspaltung in die einzelnen Primatenarten mehrfach entstandenen, also konvergenten Polymorphismus (Kermarrec 5401). Umso deutlicher geht daraus aber hervor, dass der Polymorphismus des Blutgruppen-Gens in der Population einen starken Selektionsvorteil haben muss (und nicht das einzelne Allel); denn der Polymorphismus verschwindet nicht zugunsten eines der drei Allele. Stabile Polymorphismen erschweren die merkmalsorientierte Einteilung in taxonomische Gruppen. Sie sind ein Hindernis in der Taxonomie. Viele Arten sind „polytypisch“. Die Anhänger des phänetischen Artkonzepts stehen vor dem nicht unerheblichen Problem, mit der Existenz von diagnostizierbaren Gruppen innerhalb der Art fertig zu werden (siehe Kap. 5). Man muss daraus eine unausweichliche Konsequenz ziehen, die auch im Alltag von Bedeutung ist, nicht nur für Taxonomen. Die biologische Vielfalt kann nicht nur durch Artenreichtum ausgedrückt werden. Sie ist ebenso auch ein Ausdruck für den Reichtum innerhalb der Art (Lockwood and Pimm 1994 2103). Diese Erkenntnis hat Folgen für die Zielsetzung der Artenschutzprojekte. Wenn „Artenschutz“ wirklich so eng gesehen wird, wie es der Name ausdrückt, also der Schutz und die Erhaltung von „Arten“, dann wird diese Zielsetzung der Wirklichkeit der Biodiversität nicht gerecht. Populationsunterschiede in der allelen Häufigkeitsverteilung im Vergleich zu Artunterschieden Ein wichtiger Parameter ist nicht nur die allele Vielfalt, sondern auch die quantitative Verteilung der verschiedenen Allele innerhalb der Population. Nimmt man zum Beispiel an, dass ein Allel in einer Population in vier unterschiedlichen Varianten vorliegt, so müsste rein statistisch jedes der vier Allele im Verhältnis von 25% in der Population vertreten sein. Das ist aber nicht der Fall. Die Allelverteilung weicht durchaus von der erwarteten Gleichverteilung ab. Sie ist auch zwischen verschiedenen Populationen ein und derselben Art oft sehr verschieden. Auch ändert sie die Allelverteilung im Laufe der Zeit innerhalb der Population. Die Häufigkeitsverteilung der Allele variiert in Raum und Zeit. Sie ist alles andere als starr und konstant. Sie unterscheidet sich oft schon zwischen Populationen, die nicht weit voneinander entfernt leben. Zum Beispiel haben in Deutschland die Bayern bereits eine signifikant andere Blutgruppenverteilung als die Westfalen (Mourant et al. 1976 5424). Das Hardy-Weinberg-Gesetz sagt zwar aus, dass die allele Häufigkeitsverteilung innerhalb einer Population nicht durch Sexualprozesse geändert wird, weder durch die meiotische Rekombination noch durch die Genomverschmelzung bei der Befruchtung. Das setzt jedoch eine nach außen abgeschlossene und in jeder Hinsicht konstante Population voraus. Da diese in der Natur nie gegeben ist, ändert sich die allele Häufigkeitsverteilung in Raum und Zeit. Sie ändert sich: 1. durch Mutationen, die ein Allel in ein anderes umwandeln, 2. dadurch dass Individuen aus fremden Populationen eindringen und damit die Allelhäufigkeiten verändern, 3. durch assortative Paarung, also dadurch dass die Gameten in einer Population sich nicht zufällig zur Zygotenbildung zueinander finden, sondern dass bestimmte Geschlechtspartner bevorzugt ausgewählt werden, deren Gameten dann stärker vermehrt werden als die Gameten anderer Organismen, 4. indem die Selektion für unterschiedliche Überlebens- und Fortpflanzungserfolge der einzelnen Organismen sorgt, wodurch einige Allele auf Kosten anderer begünstigt vererbt werden, und 5. durch die genetische Drift, wodurch bestimmte Allele durch Zufall seltener oder häufiger werden können. Niemand, der Arten nach Merkmalsunterschieden voneinander abgrenzt, käme auf die Idee, zwei Populationen als verschiedene Arten zu bezeichnen, nur weil sie sich in der Häufigkeit bestimmter Allele unterscheiden, z.B. weil in der einen Population im AB0Blutgruppensystem die Blutgruppe 0 mit 50% vertreten ist, in der anderen Population aber nur mit 20%. Aber schon, wenn die Verteilungsverhältnissen extremer werden, sieht die Sache anders aus, obwohl ja von der Qualität her nichts anders geworden ist, ob nun zwei Populationen mit dem Unterschied 50% zu 20% verglichen werden, oder ob der Populationsunterschied 99% zu 1% ist. Wer aber sagt dem Käferforscher, der in einer Population in 99 Belegfunden Tiere mit dem Merkmal A gefunden hat und in einer anderen Population in 99 Belegfunden nur Tiere mit dem Merkmal B gefunden, dass dies lediglich ein Unterschied in der allelen Häufigkeitsverteilung sein kann und somit weder das Merkmal A noch das Merkmal B ein Bestimmungsmerkmal ist, das allen Individuen der Art gemeinsam sein muss? Wenn er weniger als 100 Exemplare gesammelt hat, dann würde er nicht einmal festgestellt haben, dass das Merkmal B auch in den Tieren der ersten Population vorkommt, wenn auch nur in 1% der Individuen. Alleler Häufigkeitsunterschied bedeutet, dass zwischen zwei verglichenen Gruppen zwar ein deutlicher Unterschied vorhanden ist, dass aber kein einziges Merkmal 100%ig für diesen Unterschied verantwortlich ist. Dies sei an folgendem Schema an den Gruppen a und b illustriert, die durch die Merkmale A und B charakterisiert sind: Sechs Individuen der Gruppe a haben die Eigenschaften: A A A A A B Sechs Individuen der Gruppe b haben die Eigenschaften: B B B B B A Kein einziges der beiden Merkmale A und B bedeutet einen qualitativen Unterschied zwischen den beiden Gruppen a und b. Die Merkmale A und B können zwischen den beiden unterschiedlichen Gruppen nicht als verlässliche artspezifische Bestimmungsmerkmale benutzt werden, obwohl sie zweifellos einen Merkmalsunterschied beinhalten. Fast 99% der nativen Indianer Lateinamerikas haben das 0-Blutgruppen-Allel; einige innerafrikanische Völker haben es nur zu 30% (Mourant et al. 1976 5424). Das kann für Anhänger von Artkonzepten, die Arten nach Merkmalsunterschieden voneinander abgrenzen, kein vernünftiger Unterschied zwischen zwei Arten sein. Wo aber fängt es an, dass Arten abgegrenzt werden können, wenn die Unterschiede nur in der allelen Häufigkeitsverteilung liegen? Bei 90%, bei 99%; oder sind es nur dann verschiedene Arten, wenn 100% aller Individuen das Merkmal A in der eine Art und 100% aller Individuen das Merkmal B in der anderen Art besitzen? In den meisten Fällen einer merkmalsorientierten taxonomischen Einteilung dürfte die Zahl der untersuchten Proben nicht ausreichen, um zwischen diesen Möglichkeiten zu unterscheiden. Vermutlich kann bei keiner Art ausgeschlossen werden, dass dem Artunterschied „lediglich“ ein alleler Häufigkeitsunterschied des betreffenden Merkmals zugrunde liegt. Einem statistisch willkürlich herausgegriffenen Individuum ist dann an einem einzelnen Merkmal nicht mit Sicherheit anzusehen, welcher Art es angehört. Das ist auch einer der Gründe dafür, dass es nicht möglich ist, Arten so zu definieren, dass irgendein Merkmal essentiell dafür verantwortlich ist, dass ein Organismus zu einer Art gehört. Daher können Arten keine „natural kinds“ sein (siehe Kap.4). Arten sind eben anders zu definieren als als Gruppen von Organismen mit essentiellen Merkmalen, wie das bei der Gruppenzugehörigkeit der chemischen Elemente möglich ist. Auch Morphen (also komplexere Systeme, in denen der Polymorphismus mehrerer Allele eine Rolle spielt) kommen in verschiedenen Populationen in variierender Häufigkeitsverteilung vor. Wenn eine Morphe in einer geografisch entfernten Population in einer signifikant anderen Häufigkeitsverteilung im Vergleich zu einer anderen Population vorkommt, dann entsteht die Frage, ob es sich hier um Rassen handelt. Können Rassenunterschiede lediglich Unterschiede in der Häufigkeitszusammensetzung der ihr angehörenden Morphen sein? Die Antwort heißt nein. Zwar können Morphen wie die Merkmale unterschiedlicher Rassen als Folge lokaler Adaptationen in ihrer Häufigkeitsverteilung geografisch variieren; aber es gibt einen klaren Unterschied zwischen Morphen und Rassen. Morphen vermischen sich nicht durch genetische Kreuzung zu intermediären Typen (siehe oben). Rassen dagegen erzeugen im Falle der Kreuzung intermediäre Typen. Das Beispiel der teilziehenden Vögel Bei den Eigenschaftsunterschieden zwischen Zugvögeln und Standvögeln scheint es sich ebenfalls um allele Häufigkeitsunterschiede zu handeln. Dahinter aber stecken noch viele ungelöste Probleme. Ornithologen stehen vor dem Phänomen, dass das Zugverhalten von Vögeln uneinheitlich ist. Viele Arten bestehen aus geografischen Populationen, die das Brutgebiet im Herbst verlassen, um im darauffolgenden Frühjahr wieder ins Brutgebiet zurückzukommen; es sind Zugvögel. In anderen geografischen Regionen jedoch bleiben die Tiere derselben Art im Winter im Brutgebiet; es sind Standvögel. Dieses Nebeneinander von Zug- und Standvögeln ist nicht der freien, individuellen Entscheidung der einzelnen Vögel überlassen, vielmehr ist es genetisch programmiert (Berthold und Querner 1981 2571). Die Zugbereitschaft der meisten Vögel, also ihr Beweggrund, das Brutgebiet zu verlassen, wird zumindest bei den Langstreckenziehern nicht als Reaktion auf die jahreszeitliche Klimaveränderungen ausgelöst. Nur bei den sogenannten „Kälteflüchtlingen“ sind Temperaturrückgang oder Futtermangel unmittelbarer Auslöser für den Wegzug. Dazu gehören in Europa bestimmte Vogelarten, die meist Kurzstreckenzieher sind, zum Beispiel viele Enten, Gänse und Schwäne (Anatidae), einige Reiher (Ardeidae) und die Große Rohrdommel (Botaurus stellaris). Vertreter dieser Arten bleiben oft so lange im Brutgebiet oder auch in Zwischenaufenthaltsgebieten westlich bis südlich des Brutgebiets, bis Fröste und Nahrungsknappheit sie im Herbst weiter nach Süden oder Westen ausweichen lassen. Bei den meisten Langstreckenziehern jedoch sind es eben gerade nicht die Nahrungsverknappung oder die einbrechende Kälte, die die Tiere dazu veranlassen, ihr Brutgebiet zu verlassen, um in südliche oder westliche Gefilde auszuweichen. Diese Zugvögel verlassen im Sommer oder Spätsommer ihr Brutgebiet bereits zu einem Zeitpunkt, wenn die Temperatur und das Nahrungsangebot noch optimal sind. Man denke nur an die Mauersegler (Apus apus), die aus Mitteleuropa bereits Mitte Juli fortziehen, wenn die Insekten in der Luft noch in ausreichenden Maßen zur Verfügung stehen. Diese Vögel ziehen also nicht weg, weil sich irgendwelche Lebensbedingungen verschlechtert haben oder weil sie hungern oder frieren. Sie werden stattdessen durch die abnehmende Tageslänge gesteuert. Dieses Zugverhalten hat eine genetisch fundierte Basis. Die Vögel werden unruhig wegen angeborener innerer Zeitgeber, und nicht wegen äußerer Zwänge. Die genetische Anlage für Zug- oder Standvogelverhalten erfordert mit hoher Wahrscheinlichkeit das Zusammenwirken mehrerer Gene. Über diese Gene ist fast nichts bekannt (Berthold 1992 2589). Allerdings dürften die Unterschiede zwischen ziehenden und sedentären Individuen einer Vogelart meist nicht in den Strukturgenen selbst liegen, sondern in deren Regulationselementen (Enhancern) sowie in den gewebespezischen Aktivitäten der jeweiligen Transkriptionsfaktoren. Zugvögel und Standvögel sind durch eine Fülle von Merkmalen unterschieden. Gibt es auch eine ebenso große Zahl genetischer Differenzen zwischen ziehenden und sedentären Individuen ein und derselben Vogelart? Ein Zugvogel braucht andere Instinkte als ein Standvogel. Er braucht solche, die die Zugunruhe, die Orientierung (Himmelsrichtung) und die Zugweite steuern. Ein Zugvogel braucht einen anderen Stoffwechsel als ein Standvogel. Er muss im Spätsommer ein Fettpolster als Kraftstoff- und Wasserdepot für den langen Zug anlegen. Ein Zugvogel braucht andere Körpermaße als ein Standvogel. So haben Mönchsgrasmücken (Sylvia atricapilla) auf den Kapverdischen Inseln, wo sie Standvögel sind, viel kürzere Flügel als die am europäischen Festland heimischen Vögel. Ihre kurzen Flügel würden ihnen einen langen Flug über das Meer gar nicht erlauben (Berthold 1992 2589). In Zugvögeln sind Gene aktiv, die zweimal im Jahr zum richtigen Zeitpunkt das instinktive Zugunruheverhalten steuern, das bei den entsprechenden Standvogelpopulationen nicht auftritt. Hinzu kommt die Aktivität von Genen, die die Zugrichtung und Zugweite steuern; denn zumindest viele Singvögel ziehen nachts ohne elterliche Anleitung. Sie müssen die Zugwege selber finden, ohne dass ihnen Führung oder Erfahrung helfen können. Das erfordert zwei in den Erbanlagen festgelegte Steuerungen: erstens die Zugrichtung und zweitens die Zugweite. Der Vogel muss instinktiv wissen, ob er in SW- oder in SO-Richtung ziehen muss. Und er muss zusätzlich wissen, wie viele Tage er ziehen muss. Andernfalls würde er zu früh oder zu spät mit der Wanderung aufhören und dann im falschen Winterquartier sein. Kleine Singvögel sind meist kurzlebig, so dass sie kaum von der Erfahrung des Vorjahres profitieren können. Standvögel brauchen solche Instinkte nicht. Die dafür zuständigen Gene sind inaktiv. Zugvögel müssen vor dem Herbstzug gewaltige Fettreserven aufbauen. Das erfordert eine Fülle von Enzymaktivitäten, die alle bei Standvögeln derselben Spezies reprimiert oder anders geschaltet sind. Es gibt also viele Genaktivitäten, die die für das Zugverhalten notwendigen Eigenschaften steuern. Wie ist die Sache unter dem Blickwinkel der taxonomischen Einteilung zu sehen? Was liegt taxonomisch vor, wenn eine Vogelart in bestimmten geografischen Regionen als Zugvogel in Erscheinung tritt, in anderen als Standvogel und wieder in anderen Regionen als Teilzieher? Unter Teilzieher versteht man eine Vogelpopulation in einem geografischen Überlappungsbereich, bei der ein Teil der Individuen das Zugvogelverhalten zeigt, ein anderer Teil der Vögel in derselben Population aber genetisch als Standvogel gesteuert ist. Vögel sind in ihrem Zugverhalten durch mehrere Gene bzw. deren Regulationselemente gesteuert, die zwischen ziehenden und sedentären Individuen in verschiedener Allelform vorliegen müssen. Nach der Auffassung von Berthold sollen die Allele für Zugverhalten und die für Standvogelverhalten in allen Vogelpopulationen vorhanden sein, auch in den Populationen, die entweder nur Zugvögel sind oder nur Standvögel sind (Berthold 1992 2589). Wenn also im Norden eine Vogelpopulation im Herbst das Brutgebiet verlässt und im Süden eine Population derselben Art das ganze Jahr im Brutgebiet bleibt, dann ist das nur eine Sache der allelen Häufigkeitsverteilung. Es handelt sich also um geografisch unterschiedlich verteilte Allelhäufigkeiten in den Populationen. Das bedeutet, dass auch im Norden mit unwirtlichen Wintern einige Vögel die Standvogel-Allele enthalten. Die Regulationselemente für die Gen-An- und -Abschaltung der Zugeigenschaften scheinen in allen Populationen vorhanden zu sein, jedoch erfolgt die Anschaltung dieser Regulationswege bei einer Zugvogelpopulation in viel mehr Individuen als in einer Standvogelpopulation (und umgekehrt). Es scheint keine Vogelpopulation zu geben, deren Genotypen zu 100% aus Zugvogelgenen oder zu 100% aus Standvogelgenen bestehen. Die genetische Anlage zum Standvogel kommt in einer Zugvogelpopulation möglicherweise nur deswegen nicht zur Wirkung, weil sie unter dem Deckmantel der Rezessivität verborgen bleibt. Treten in der Zugvogelpopulation jedoch Homozygotien dieser StandvogelSteuerelemente auf, so haben die davon phänotypisch betroffenen Organismen wenig Überlebenschancen und sterben vermutlich. Trotzdem aber überleben die StandvogelSteuerelemente im rezessiven Zustand in vielen Organismen, so dass die gesamte Zugvogelpopulation die Chance behält, im Falle einer Klimaänderung überwiegend zu Standvögeln zu werden. Und entsprechend umgekehrt ist es in den Standvogelpopulationen, in denen die Zugvogelallele nur in wenigen Organismen vorhanden sind. Wenn sich die Umweltbedingungen ändern, dann kann eine Standvogelpopulation durch Selektion in wenigen Generationen zur Zugvogelpopulation werden. Die Auffassung Bertholds wird auch dadurch gestützt, dass eine Zugvogelpopulation innerhalb von nur wenigen Generationen zur Standvogelpopulation werden kann; und das gleiche kann auch umgekehrt ablaufen, wenn die Klimaänderungen es so wollen (Berthold 1992 2589). Der allele Polymorphismus gibt den einzelnen Populationen eine bemerkenswerte Flexibilität über längere evolutionäre Zeiträume hinweg. Änderungen des lokalen Klimas können binnen weniger Generationen aus einem Zugvogel einen Standvogel machen, indem die Individuen mit der jeweils präadaptiven Merkmalsausstattung die anderen Individuen innerhalb der Population ersetzen, da sie plötzlich einen kolossalen Selektionsvorteil haben. Auch die in Mitteleuropa heutzutage als Überwinterer so standorttreuen Amseln (Turdus merula) waren zu Goethes Zeiten in Deutschland noch Zugvögel, als die Winter, vor allem in den damals noch nicht so aufgeheizten Städten, noch wesentlich kälter waren (Berthold 1992 2589). Die meisten Untersuchungen zum Zugpolymorphismus sind an Mönchsgrasmücken (Sylvia atricapilla) gemacht worden (Berthold und Querner 1981 2571). [sehen wir im Bükk] Mönchsgrasmücken haben ein weites Brutareal, das von den Kapverdischen Inseln auf der geografischen Breite Senegals über die Kanarischen Inseln, dann über fast ganz Europa bis in den Iran und ins Ob-Gebiet der Westsibirischen Tiefebene jenseits des Urals reicht (siehe Kap. 7). Auf den Kapverdischen Inseln sind die Mönchsgrasmücken Standvögel, in ganz Südeuropa sind es sowohl Zugvögel als auch Standvögel, und in Nordeuropa, Osteuropa und Asien sind es Zugvögel (Abb. 2 aus Sutherland 1988 5780). In Sibirien hätte eine daheim bleibende Mönchsgrasmücke im Winter wohl keine Überlebenschance, genauso wenig wie eine auf den Kapverden brütende Grasmücke kaum überleben würde, wenn sie auf den Ozean hinaus wandern würde. Aber die wenigen phänotypisch auftretenden „Fehlleistungen“ gefährden das Überleben der Allele nicht, wenn diese überwiegend rezessiv in der jeweiligen Population erhalten bleiben. Es entsteht die Frage, ob Zugvögel und Standvögel Rassen oder Morphen sind. Diese Frage wird im Folgenden am Beispiel der „echten“ Zugvögel untersucht, also der Zugvögel, die Langstreckenzieher sind und die in ihrem Zugverhalten weitestgehend genetisch gesteuert sind. Bei vielen Arten sind die im Norden brütenden Populationen Zugvögel, die im Süden brütenden Populationen dagegen Standvögel. Diese Zweiteilung in zwei unterschiedliche Populationen als lokale Anpassung an unterschiedliche geografische Umweltbedingungen erinnert zunächst an geografische Rassen. Aber im klinalen Übergangsbereich zwischen Zugund Standvögeln gibt es eine Mischpopulation. Das sind die sogenannten Teilzieher: Ein Teil der Individuen verlässt im Herbst das Brutgebiet und kehrt im Frühjahr des nächsten Jahres wieder zurück; es sind Zugvögel. Ein anderer Teil der Individuen derselben Population im selben Vorkommensgebiet verhält sich ganz anders; die Tiere ziehen nicht; es sind Standvögel. Eine Teilzieher-Population besteht zum Teil aus Zugvögeln mit den Erbanlagen des Zuges, und sie besteht zu einem anderen Teil aus Standvögeln, in denen diese Erbanlagen reprimiert sind. Es scheint also keine Mischtypen zu geben, zumindest nicht in großen Mengen. Zug- und Standvögel können also keine Rassen sein; denn Rassen sind geografisch getrennte Populationen, die sich im klinalen Übergangsbereich vermischen und dort intermediäre Genound Phänotypen hervorbringen. Wären Zug- und Standvögel Rassen, so müssten die Rassenunterschiede allmählich ineinander übergehen. Die Kreuzung zwischen Zug- und Standvögeln in der Mischpopulation bringt aber nach dem Entweder-Oder-Prinzip zumindest überwiegend nur Zugvögel oder Standvögel hervor. Daher müssen die Zug- und Standvögel als Morphen eingestuft werden, vergleichbar mit der Entstehung von Männchen und Weibchen beim Beispiel des Geschlechtsdimorphismus, wo es auch keine Zwischentypen gibt. Wie kann das funktionieren? Die zahlreichen Erbanlagen der Zug- und Standvögel müssten doch wechselseitig miteinander inkompatibel sei. Es sollte doch erwartet werden, dass Zugund Standvögel nicht erfolgreich miteinander kreuzbar wären, weil ihre Gene rekombinieren. Ein Teil der „Zugvogel-Allele“ rekombiniert mit einem Teil der „Standvogel-Allele“. Aber die Mischpopulationen der Teilzieher bilden entgegen der Erwartung einen lebensfähigen Reproduktionsverband, der eben nicht überwiegend letale Mischlinge erzeugt. Warum sind in der Teilzieher-Population die Zugvögeln und die Standvögel dauerhaft fruchtbar miteinander kreuzbar? Es sind wahrscheinlich viele Gene, die die Zugeigenschaften kontrollieren, und diese müssen alle zusammen gleichzeitig exprimiert sein, um einen Vogel zum funktionsfähigen Zugvogel zu machen. Und umgekehrt muss die gesamte Zahl dieser Gene reprimiert sein, um einen Standvogel zu erzeugen. Wenn ein Zugvogel mit einem Standvogel gekreuzt wird, sollte ein intermediärer Hybrid erwartet werden, der teilweise mit Zugvogeleigenschaften, aber auch teilweise mit Standvogeleigenschaften ausgestattet ist. Gemeint sind damit die Allele für Zugunruhe, Zugrichtung, Zugweite, für den Fettstoffwechsel, für die Flügellänge und andere Merkmale, die die Unterschiede zwischen Zug- und Standvögeln beinhalten. Wie ist es möglich, dass Zug- und Standvögel miteinander kreuzbar sind? Eine Denkmöglichkeit wäre, dass ein genetischer Mechanismus vorliegt, der ganze Komplexe aus mehreren Genen vor Rekombination schützt und damit als Kopplungsgruppe zusammenhält. Es kommt in der Teilzieherpopulation offenbar nicht zu Vermischungen der Kopplungsgruppen; denn es ist in der Population eine Entweder-OderEntscheidung möglich, wie das in vielen Beispielen von der genetischen Geschlechtsbestimmung her bekannt ist. Ein genetischer Mechanismus könnte dafür sorgen, dass Gruppen von Genen in einer bestimmten Allelstellung gekoppelt vererbt werden, ohne dass es mit anderen Allelen der gleichen Gengruppe zur Rekombination kommt. Eine andere Denkmöglichkeit wäre, dass alle „Zugvogel-Gene“ von einem übergeordneten Gen („master gene“) kontrolliert werden, das entweder an- oder abgeschaltet werden kann und damit einen gesamten Satz untergeordneter Gene reguliert. Andererseits jedoch haben künstliche Kreuzungsversuche an osteuropäischen mit westeuropäischen Mönchsgrasmücken gezeigt, dass dabei Mischeigenschaften herauskommen (Berthold und Querner 1981 2571). Die Hydride zwischen osteuropäischen und westeuropäischen Mönchsgrasmücken zeigen teilweise die Zugeigenschaften der SüdostZieher und teilweise die der Südwest-Zieher. Das spricht gegen eine eine Entweder-OderEntscheidung. Wären solche Hybride in freier Natur geboren worden, so hätten sie keine Chance zum Überleben gehabt. Die genetische Kompatibilität zwischen Zug- und Standvögeln ist immer noch eine offene Frage, und daher bedarf die oben vorgenommene Klassifikation der Zug- und Standvögel als Morphen noch einer weiteren Überprüfung. Innerartliche Morphen beim Widderchen Zygaena ephialtes Viele Arten der Schmetterlings-Gattung Zygaena (Blutströpfchen oder Widderchen) sind morphologisch kaum voneinander zu unterscheiden. Daher werden sie als „kryptische Spezies“ bezeichnet. Ein Beispiel sind die drei Arten der Zygaena-transalpina-Gruppe: transalpina, hippocrepidis und angelicae (siehe unten und Abb. S. 286 in Ebert und Rennwald 1993 2592, Bd. 3). Nur Spezialisten sind in der Lage, diese drei Arten auseinander zu halten, und auch dann verbleibt noch eine gewisse Restunsicherheit (siehe Kap. 5). Andererseits gibt es gerade bei der Gattung Zygaena ein eindrucksvolles Beispiel eines intraspezifischen Polymorphismus. Der liegt bei der Art Zygaena ephialtes vor (diese Art werden wir wahrscheinlich im Bükk sehen). Hier existieren einige deutlich unterscheidbare Morphen. Es ist um ein Vielfaches leichter, diese innerartlichen Morphen voneinander zu unterscheiden, als bei den anderen Zygaenen die Artzugehörigkeit zu ermitteln. Wer sich vorgenommen hat, Zygaena-Arten zu bestimmen, der kann nur davon träumen, eine ganze Reihe von Arten so leicht voneinander unterscheiden zu können, wie er die einzelnen Morphen der Art Z. ephialtes unterscheiden kann. Zygaena ephialtes tritt in vier verschiedenen Morphen auf. Daher trägt Z. ephialtes auch den deutschen Trivialnamen „Veränderliches Widderchen“. Die verschiedenen Morphen sind durch zwei Merkmale gekennzeichnet, die ihrerseits in zwei alternativen Qualitäten auftreten. Das eine Merkmal ist die Farbigkeit oder Nicht-Farbigkeit der Hinterflügel, das andere ist die Qualität der Farbe. Die Hinterflügel sind entweder farbig oder nicht-farbig, und die Färbung der Vorder- und Hinterflügel ist entweder rot oder gelb (Abb. 14 aus Sbordoni und Forestiero 1984 5782). Das führt zu vier Morphen, die durch jeweils vier verschiedene Merkmalskombinationen gekennzeichnet sind. Die beiden Merkmale werden von zwei Genen bestimmt, jedes davon in zwei alternativen Allelformen (Ebert und Rennwald 1993 2592). Das erste Gen legt fest, ob der Hinterflügel farbig oder schwarz ist, wobei farbig dominant über schwarz ist. Das zweite Gen bestimmt die Farbe rot oder gelb, wobei rot dominant über gelb ist. Diese Gene sind im Erbgang frei kombinierbar, da sie auf verschiedenen Chromosomen liegen. Die genetische Kreuzung der reinerbigen Eltern entspricht genau dem Schulbeispiel der dritten Mendelregel, der Zweifaktorkreuzung, mit dem bekannten Resultat, dass in der F2Generation vier verschiedene Phänotypen im Zahlenverhältnis 9:3:3:1 auftreten. Das sind die vier Morphen: eine rote farbigflügelige Form (9mal; beide Gene bestimmen dominant den Phänotyp), eine gelbe farbigflügelige Form (3mal; dominant und homozygot rezessiv). eine rote schwarzflügelige Form (3mal; dominant und homozygot rezessiv) und eine gelbe schwarzflügelige Form (1mal; beide Gene sind homozygot rezessiv) (Ebert und Rennwald 1993 2592). Von den Erstbeschreibern wurden die einzelnen Morphen von Z. ephialtes durchweg als eigene Arten angesehen. Sie erhielten unterschiedliche wissenschaftliche Namen, wobei der Gattungsname damals noch nicht Zygaena, sondern Sphinx hieß: Sphinx peucedani, S. athamanthae, S. coronillae etc. Aus dem Vergleich der Merkmale allein konnte dieser Fehler nicht behoben werden. Hier zeigt sich erneut, wie wichtig es ist, Merkmalsunterschieden als alleinigem Kriterium einer Artzugehörigkeit zu misstrauen. Das Ansinnen, aus Merkmalsgleichheit Conspezifität bzw. aus Merkmalsungleichheit unterschiedliche Artzugehörigkeit herzuleiten, kann sehr schnell zu falschen Resultaten führen kann. Der Fall Zygaena ephialtes ist etwas außerordentlich Bemerkenswertes. Denn einerseits haben wir etwas ganz Normales vor uns: Das Auftreten von multiplen Allelen und das entsprechende Vererbungsschema mit unterschiedlichen Geno- und Phänotypen sind ein Normalfall bei allen diploiden Lebewesen. Andererseits aber gibt es nicht viele Organismen, die uns innerhalb der Art eine entsprechende Morphenvielfalt zeigen. Warum also sind nicht alle Tiere, zumindest alle Schmetterlinge, entsprechend polytypisch vertreten? Das würde die Übersichtlichkeit von Bestimmungsbüchern gewaltig erschweren. Offenbar sind die Allelpaare der beiden Gene von Z. ephialtes besonders stabil. Normalerweise sind multiple Allelien in der Population nicht sehr langlebig. Die Selektion und die genetische Drift sorgen in den meisten Fällen dafür, dass mutante Allele sich in der Population nicht fixieren können (siehe oben). Und wenn sie sich schon ausgebreitet haben, dann sind sie oft nicht langlebig. Ganz anders verhält es sich jedoch, wenn die Selektion den Polymorphismus fördert, wenn die Selektion sozusagen daran „interessiert“ ist, dass mehrere Allelformen in der Population coexistieren. Das muss bei Z. ephialtes der Fall sein. Bemerkenswerterweise sind die vier verschiedenen Morphen in verschiedenen geografischen Verkommensgebieten nicht so verteilt, wie es der 9:3:3:1-Häufigkeitsverteilung des Kreuzungsschemas entspricht. Das bringt zum Ausdruck, dass die Selektion in unterschiedlichen Regionen bestimmte Allele unterschiedlich fördert, so dass sich geografisch entfernte Populationen in den allelen Häufigkeiten voneinander unterscheiden. So besiedelt die farbigflügelige Morphe hauptsächlich Mittel- und Osteuropa, während die schwarzflügelige Morphe ihr Verbreitungsareal mehr im Mittelmeerraum und auf dem westlichen Balkan hat. Die vier verschiedenen Morphen von Z. ephialtes scheinen in verschiedene Mimikry-Systeme eingebunden zu sein, wobei bei der gelben Morphe mit schwarzen, nicht-farbigen Hinterflügeln die täuschende Ähnlichkeit zum Bärenspinner Syntomis phegea (diese Art werden wir mit Sicherheit im Bükk sehen) eine mögliche Rolle spielt. Syntomis phegea gehört zu einer ganz anderen Familie, nämlich den Arctiiden (Bärenspinner) statt den Zygaeniden (Widderchen). Hier wirkt sich offenbar eine Mimikry aus, die zu einer erstaunlichen Konvergenz in der Gestalt nicht-verwandter Arten geführt hat. Die Ähnlichkeit der gelben, schwarzflügeligen Morphe von Z. ephialtes mit dem Bärenspinner Syntomis phegea ist verblüffend. Das wäre aber nur die Erklärung für eine der vier Morphen. Und im übrigen gibt es auch Orte des gemeinsamen Vorkommens von Syntomis phegea und Zygaena ephialtes, ohne dass Z. ephialtes an diesen Orten in der „passenden“ Morphe vertreten wäre. Was die Gesamtheit der Zygaena-ephialtes-Morphen angeht und ihre spezifische Häufigkeitsverteilung an verschiedenen geografischen Orten, so ist bis heute kein schlüssiger Zusammenhang zwischen den jeweiligen Farbmustern und einem dazu passenden MimikrySystem gefunden worden. Welche möglichen Tierarten an verschiedenen Orten nachgeahmt werden und daher die unterschiedliche Häufigkeitsverteilung der Allele selektiv beeinflussen, ist weitgehend unbekannt. Der Polymorphismus der Schnirkelschnecke (Cepaea nemoralis) [Gruppenselektion] Warum gibt es einen so verbreiteten stark ausgeprägten genetischen Polymorphismus? Diese Frage stellt sich bei vielen Beispielen aus dem Tierreich. Historisch wurde der Polymorphismus oft als neutral ohne funktionale Relevanz betrachtet. Diese Auffassung muss heute als falsch bezeichnet werden (Jones 2009 5840). Stattdessen gilt innerartliche Vielfalt als eines der Schlüsselprinzipien des Überlebens mancher Tierarten. Ein Paradebeispiel sind die Gehäuse der Hain-Schnirkelschnecke (Cepaea nemoralis), deren extreme Farb- und Bänderungsvielfalt sehr auffällig ist [werden wir im Bükk sehen]. Die Schalen können an ein und demselben Vorkommensort gelb, rot oder braun sein. Sie können ohne Bänder sein oder durch ein, zwei, drei, vier oder fünf Bänder gemustert sein. Es gibt 20 bis 30 verschiedene Morphen in einer Population. Was sind die Gründe für diese Vielfalt? Wieso dominiert nicht ein einziger Typ, der am besten angepasst ist? Es kann doch eigentlich nicht sein, dass alle Typen mit ihren unterschiedlichen Färbungen und Zeichnungen alle die exakt gleichen Überlebens- und Fortpflanzungschancen haben. Warum verdrängt der eine Genotyp nicht den anderen? Die visuelle Selektion durch Fraßfeinde oder die klimatische Selektion müssten doch immer wieder den einen oder den anderen Phänotyp bevorzugen. Und wenn es sich schon um eine variable Anpassung an unterschiedliche ökologische Nischen handelt, warum führt die unterschiedliche Einnischung dann nicht zur assortativen Paarung und damit zur (sympatrischen) Artbildung (siehe Kap. 7)? Ob C. nemoralis Bänder trägt oder nicht, könnte mit der Singdrossel (Turdus philomelos) zusammenhängen. Singdrosseln sind die Hauptfeinde der Schnirkelschnecke (Anton und Bossdorf 2009 5824). Hier gibt es viele Untersuchungen, ob das Ausmaß der Bänderung oder die Färbung der Schneckenschale einen mehr oder minder guten Tarneffekt hat. In der Tat wurde festgestellt, dass die Singdrosseln die gebänderten Schnecken in der Vegetation schwerer entdecken können als die ungebänderten. Genau dieses Ergebnis aber erklärt nicht den Polymorphismus. Im Gegenteil, der Polymorphismus müsste über kurz oder lang verschwinden, zumindest an den Orten, wo die Singdrossel der Hauptfeind der Schnecke ist. Einen weiteren selektiven Einfluss darauf, ob C. nemoralis Bänder trägt oder nicht, könnten klimatische Faktoren nehmen. Dunkle Gehäuse absorbieren mehr Wärmestrahlung, gelbe Gehäuse reflektieren sie stärker. Die Farbe der Gehäuse beeinflusst also die Temperatur der darin lebenden Schnecke. Die Exposition zur Sonne ist entscheidend. Zu viel Sonne bedeutet den Wärmetod, zu wenig Sonne bedeutet, dass sie als wechselwarme Tiere zu starr sind, sich nicht genügend bewegen können und daher verhungern. Aber auch diese Erklärung ist unbefriedigend. Warum gibt es dann so viele unterschiedliche Morphen am selben Ort? Cepaea nemoralis hat ein großes Verbreitungsgebiet, das sich von Norwegen bis nach Spanien erstreckt und von der Meeresküste bis in 1200 Metern Höhe. Man findet C. nemoralis in so unterschiedlichen Lebensräumen wie Dünen, Wiesen oder Wäldern. Zwar sind die Morphen nicht gleichmäßig verteilt. Stattdessen dominieren in manchen Regionen bestimmte GehäuseVarianten. Aber es ist nicht so, dass jeder geografische Ort, jede Gebirgshöhe oder jedes Habitat ihre eigenen jeweils spezifischen Morphen haben. Zwar wurde auch beobachtet, dass sich die unterschiedlichen Genotypen ganz unterschiedlich lange in der Sonne aufhalten (Jones 2009 5840). Manche bevorzugen die kühlen Morgenstunden für ihre Aktivität, andere kriechen eher mittags herum, wiederum andere bewegen sich vorwiegend im Schatten. Aber ein schlüssig erkennbarer Zusammenhang zwischen der Temperatur und der Hell- oder Dunkelfärbung oder der starken bzw. schwachen Bänderung ist bisher nicht gefunden worden. Bis heute können die Wissenschaftler die spezifischen Farbmuster der Phänotypen in den meisten Fällen nicht den einzelnen physikalischen Faktoren des Klimas in den Lebensräumen zuordnen, in denen die jeweiligen Morphen sich aufhalten. Das Bild, das sich abzeichnet, ist zu komplex. Die treibende Kraft für die innerartlichen Vielfalt der Cepaea-Schnecken scheint zu sein, dass mit der Erhöhung der Typenvielfalt auch die Vielfalt der bewohnbaren Habitate erhöht wird. Dadurch erweitern die Schnecken ihre Ressourcen und verstärken damit ihre Populationsgröße. Das wäre zweifellos ein Selektionsvorteil. Aber dann bleibt die Frage offen, warum die unterschiedlichen Einnischungen nicht zur Artbildung geführt haben. Es gehört dies wohl zu den entscheidenden Fragen der Taxonomie. Das Beispiel des Finkenschnabels [Gruppenselektion] Die Schnabelhöhe bei Finkenvögeln ist ein von der Selektion stark kontrolliertes Merkmal. Schon Schwankungen um ein Zehntel Millimeter in der Schnabelhöhe können dazu führen, dass die betroffenen Vögel nicht mehr konkurrenzfähig gegenüber ihren Artgenossen sind, wenn es darum geht, bestimmte Samen zum Nahrungserwerb zu knacken. Die Höhe des Schnabels ist kein Ergebnis eines besonderen Trainings; vielmehr ist dieses Merkmal genetisch vorgegeben (Abzhanov et al. 2004 5736). Finkenschnäbel bieten ein schönes Beispiel für einen stabilen Polymorphismus innerhalb der Population, ebenso wie die oben erwähnten Blutgruppen. Die westafrikanischen Finken Pyrenestes ostrinus (Purpurastrild) sind geradezu das Schulbeispiel für adaptiven Polymorphismus, worunter das Nebeneinander von unterschiedlichen Habitatadaptationen durch Vertreter ein und derselben Art verstanden wird. In Kamerun treten zwei Morphen des Purpurastrilds auf, die durch unterschiedliche Schnabelgrößen charakterisiert sind. Beide Morphen kommen gleichzeitig nebeneinander in einer einzigen Population vor (Smith 1993 5773; Smith 2008 5774). Die kleinschnäbeligen Individuen ernähren sich hauptsächlich von weichen Samen, während die großschnäbeligen Vögel sich auf das Knacken harter Samen spezialisiert haben. Es handelt sich um einen Dimorphismus. Dieser Dimorphismus ist im Ergebnis vergleichbar mit dem Geschlechtsdimorphismus: Es treten nur die dünnschnäbeligen und dickschnäbeligen Morphen auf. Man beobachtet keine Tiere mit intermediärer Schnabelgröße, genauso wie auch beim Geschlechtsdimorphismus keine Zwischenformen auftreten. Allerdings gibt es einen deutlichen Unterschied. Das alleinige Auftreten zweier distinkter Typen ist beim Geschlechtsdimorphismus ein auf einen genetischen Mechanismus zurückzuführen, der über das Entweder-Oder entscheidet. Beim Finkenschnabel ist dagegen die Selektion dafür verantwortlich, dass keine Mischtypen auftreten. Die intermediären Individuen mit Schnäbeln mittlerer Größe entstehen zwar bei entsprechender Allelkombination, jedoch haben diese Tiere wegen der scharfen Kontrolle durch die Selektion keine Überlebenschance gegenüber ihren „reinen“ kleinschnäbeligen und großschnäbeligen Artgenossen, die jeweils auf ganz bestimmte Samen spezialisiert sind. Alle intermediären Phänotypen werden immer wieder von neuem ausgemerzt. Die Selektion „zerreißt“ sozusagen die Population in zwei Morphen. Es gibt bei den Finken weder einen genetischen Mechanismus für die Entstehung von nur zwei Schnabelgrößen, noch gibt es zwischen den beiden Morphen mit den zwei Schnabelgrößen irgendwelche Paarungsbarrieren. Beide Morphen gehören zu ein und derselben Art, weil sie Angehörige ein und derselben Genflussgemeinschaft sind. Das gleichzeitige Nebeneinander zweier Phänotypen innerhalb ein und derselben Population hat einen biologischen Sinn. Es ist nicht der Selektionsvorteil des dicken Schnabels, der sich hier äußert, noch ist es der Selektionsvorteil des dünnen Schnabels. Es ist stattdessen der Selektionsvorteil, dass es schlechthin zwei Varianten gibt, die die Gesamtpopulation bereichern. Die Existenz zweier Morphen mit unterschiedlichem Schnabeltyp erweitert das Nahrungsspektrum der Art. Würden alle Organismen den gleichen Schnabeltyp haben, so würden alle dasselbe fressen. Dadurch würden die Nahrungsressourcen im Habitat viel schneller erschöpft werden. Erneut tritt für die Taxonomie das ungelöste Problem auf, warum die Natur bei derartigen Adaptationen in bestimmte Nischen zwei Wege beschreitet, einmal den adaptiven Polymorphismus innerhalb der Art, andererseits aber auch die adaptive Speziation (Dieckmann et al. 2005 5454). Viele Beispiele belegen, dass genetische Anpassungen, die eine weitere Nahrungsquelle erschließen, nicht zum Polymorphismus, sondern zur Artbildung führen. Adaptive Einnischung ist ja eine der entscheidenden Voraussetzungen für die sympatrische Speziation (Tautz 2009 5838). Normalerweise besteht ein Selektionsdruck, die neu auftretenden Organismen mit Anpassungsmerkmalen an ein neues Habitat dadurch zu erhalten, dass sie vor Rückkreuzung mit ihren Artgenossen geschützt werden. Das geschieht, indem coevolutiv zu den Merkmalen für den neuen Nahrungserwerb auch Merkmale positiv selektiert werden, die eine assortativen Paarung bewirken, die neuen Organismen also davor schützen, sich weiterhin mit den „alten“ Organismen zu paaren (siehe Kap. 7). Das führt meist rasch zu getrennten Genflussgemeinschaften, also zur Aufspaltung in zwei neue Arten. Warum aber gibt es auch den zweiten Mechanismus, die Erhaltung der Adaptation an ein neues Habitat im Nebeneinander zu den Organismen, die das „alte“ Habitat bewohnen, durch den Mechanismus des stabilen Polymorphismus innerhalb ein und derselben Art? Was sind die Ursachen dafür, dass die Natur zwei verschiedene Wege beschreitet, um Neuadaptationen zu konservieren? Was sind die Mechanismen dafür, dass im Falle des adaptiven Polymorphismus die Artbildung umgangen werden kann? Das Beispiel des afrikanischen Purpurastrilds ist mit einem Phänomen zu vergleichen, das bei manchen Greifvögeln beobachtet wird, dass nämlich die Weibchen erheblich größer sind als die Männchen. Das ist z.B. bei Habichten, Sperbern und Wanderfalken der Fall. Offenbar bietet die erweiterte Vielfalt durch das Vorhandensein zweier unterschiedlicher Körpergrößen den betroffenen Vögeln die Möglichkeit, ihr Nahrungsspektrum zu erweitern. Die größeren Weibchen schlagen zum Teil eine andere Beute als die kleineren Männchen. Die Geschlechter konkurrieren weniger miteinander und nutzen insgesamt eine größere Vielzahl an Beuteobjekten unterschiedlicher Körpergrößen. Innerartlicher Polymorphismus beim „Darwinfinken“ Geospiza fortis [Gruppenselektion] Ein weiteres Beispiel erhöhter Flexibilität durch intraspezifischen Polymorphismus liegt beim „Darwinfinken“ Geospiza fortis (Mittlerer Grundfink) vor. Die Grundfinken auf der Galapagos-Insel Daphne Major bestehen genau wie beim afrikanischen Purpurastrild aus starkschnäbligen und dünnerschnäbligen Individuen. Im Unterschied zum Purpurastrild leben die starkschnäbligen und dünnerschnäbligen Grundfinken aber nicht gleichzeitig nebeneinander, sondern folgen einander in verschiedenen Jahren je nach Klimabedingungen. Das Forscherehepaar Grant hat in einer 30 Jahre andauernden Studie jährlich verfolgt, wie sich die Morphen von Jahr zu Jahr abwechseln (Grant und Grant 2002 5226). Auf der Insel Daphne Major verursachen periodische Schwankungen der Meeresströmung (El Nino) eine drei- bis fünfjährige Trockenperiode, die dann wieder von einer ebenso langen Feuchtperiode abgelöst wird. Das hat zur Folge, dass die Vegetation sich rhythmisch ändert. In den Jahren mit feuchter Witterung stehen den Finken kleine, weichere Samen zur Verfügung, in den Trockenperioden überwiegen Pflanzen mit dicken, harten Samen. Dieses Phänomen wird von einem bemerkenswerten Oszillieren der Schnabelgröße beim Mittelgrundfinken Geospiza fortis begleitet. Die Finken haben in den Feuchtjahren etwas dünnere Schnäbel, in den Trockenjahren dann dickere Schnäbel. Dies geht keineswegs auf Trainingseffekte zurück, sondern auf allele Unterschiede (Abzhanov et al. 2004 5736). Der Genpool der Population des Mittelgrundfinken enthält sowohl die Allele für Dickschnäbeligkeit als auch die Allele für Dünnschnäbeligkeit. Was sich in den einzelnen Perioden ändert, ist nur die Häufigkeitsverteilung der Allele in den zur entsprechenden Zeit lebenden Individuen. Sobald eine Trockenperiode anbricht, sterben die meisten Mittelgrundfinken, weil ihre Nahrungspflanzen mit weichen Samen nicht mehr verfügbar sind. Sie verhungern regelrecht. Hauptsächlich die großen starkschnäbligen Individuen überleben, weil sie in der Lage sind, extrem harte Samen zu knacken. Sie erzeugen ein paar Generationen lang Nachkommen mit größeren und breiteren Schnäbeln, bis nach ein paar Jahren die erneut auftretende feuchte Witterung wieder weichsamige Pflanzen wachsen lässt. Dann geschieht das Umgekehrte: Die inzwischen häufig gewordenen dickschnäbeligen Finken sterben größtenteils, und die wenigen überlebenden dünnschnäbligen Finken haben sofort einen Selektionsvorteil, produzieren höhere Nachkommenzahlen und beginnen, in der Gesamtpopulation zu dominieren, indem sie die dickschnäbeligen Individuen verdrängen und ersetzen. Das ist ein eindrucksvolles Beispiel für einen stabilen allelen Polymorphismus. Die Kraft eines starken Selektionsdrucks nimmt Einfluss auf die jeweilige Zusammensetzung der Allelverteilung in der Population, ohne aber eines der Allele völlig zu verdrängen. Zwei Morphen wechseln sich nacheinander in der Finkenpopulation innerhalb ganz weniger Generationen in ihrer Häufigkeit gegenseitig ab. Das Aussterben der Gesamtpopulation wird dadurch verhindert. Für den Fall, dass sich die Umwelt im Laufe der Jahre ändern sollte, ist die Population schon darauf vorbereitet. Die Population hält sozusagen immer einige Individuen parat, die einer möglichen Veränderung von vornherein (präadaptiv) angepasst sind. Diese Individuen können die nunmehr Benachteiligten in wenigen Generationen ersetzen. Das Interessante ist, dass keine der alternativen Allele, weder die für Dünnschnäbeligkeit noch die für Dickschnäbeligkeit, völlig aussterben, obwohl sie in den „falschen“ Jahren nur Nachteile und keine Vorteile mit sich bringen. Dieses Phänomen ist darwinistisch nur schwer zu verstehen; denn da in den Trockenperioden die Dünnschnäbeligkeit selektiv benachteiligt ist, müssten die Allele für die Dünnschnäbeligkeit in diesen Zeiten eigentlich aussterben, und umgekehrt müssten dies die Allele für Dickschnäbeligkeit in den Feuchtperioden. Keines der Allele stirbt aber aus, weder durch Selektion noch durch genetische Drift. Es handelt sich um einen stabilen Polymorphismus. Der Selektionsvorteil eines stabilen Polymorphismus ist nicht der Vorteil des einen oder des anderen Allels. Es ist der Vorteil der Vielseitigkeit, der Präadaptation an Bedingungen, die zurzeit nicht herrschen, aber kommen könnten. Wenn die Umweltbedingungen sich ändern, dann stehen der Population die jeweils vorteilhaften Allele bereits zur Verfügung. Das ist darwinistisch schwer erklärbar. Es ist das Problem der Gruppenselektion, auf dessen Kontroversität schon oben hingewiesen wurde (Okasha 2001 5930; Okasha 2006 5924). In den letzten Jahren hat es entscheidende Erkenntnisfortschritte gegeben, die die genetische Steuerung des Finkenschnabels verständlich machen (Abzhanov et al. 2004 5736). Die Morphogenese des Vogelschnabels geht in der Tat auf ein oder wenige Gene zurück. Das Schlüsselgen ist Bmp4. Dieses Strukturgen hat weitreichende Aufgaben in der Differenzierung der Zellen des cranialen Neuralkammes und damit in der cranialen Skelettmorphogenese. Das Gen steuert das Muster, das Wachstum und die Chondrogenese der mandibularen und maxillaren Vorsprünge des vorderen Kopfskeletts. Wie üblich bei der Morphogenese der Tiere, spielt dieses Gen verschiedene Rollen zu verschiedenen Entwicklungszeiten. Der aus Kasachstan stammende, in Harvard forschende Zoologe Arkhat Abzhanov hat sechs Arten der Darwinfinken hinsichtlich der artspezifischen Unterschiede in der Bmp4Genexpression untersucht. Unterschiede in Zeitpunkt und im quantitativen Ausmaß der Genexpression von Bmp4 korrelieren mit Unterschieden in der Schnabelmorphologie der verschiedenen Darwinfinkenarten. Für diese Unterschiede sind, wie üblich, regulierende DNA-Abschnitte (Enhancer) und gewebespezifische Transkriptionsfaktoren verantwortlich, die artspezifisch unterschiedlich im Epithel des sich entwickelnden Schnabels exprimiert werden. Die allelen Unterschiede zwischen den Schnabelmorphen der Finken sind also nicht im Strukturgen Bmp4 zu suchen, sondern in dessen Regulationselementen. Das Strukturgen wird lediglich variierend reguliert, ist ansonsten aber immer das gleiche. Experimentelle Eingriffe, die das Expressionsniveau von Bmp4 künstlich erhöht haben, lösten in der Tat Veränderungen in der Schnabelmorphologie hervor. Die Morphen der Strumpfbandnatter Thamnophis ordinoides [Gruppenselektion] Bei der nordamerikanischen Strumpfbandnatter (Thamnophis ordinoides) leben mehrere Morphen nebeneinander in derselben Population (Brodie 1989 4814). Sie unterscheiden sich deutlich in der Rückenzeichnung und -färbung. Außerdem zeigen die einzelnen Morphen ein unterschiedliches Verhalten. Sie haben ein großes oder kleineres Jagdrevier, bei Gefahr fliehen sie entweder oder bleiben gedrückt liegen, oder sie zeigen bei Gefahr ein aggressives Verhalten. Das Interessante ist, dass die einzelnen Schlangen mit Merkmalskombinationen ausgestattet sind, die zusammenpassen. Färbung und Verhalten sind aufeinander abgestimmt. Schlangen mit tarnfarbenem Rückenmuster zeigen ein geducktes Verhalten und bleiben bei Gefahr ruhig liegen, Schlangen mit auffälliger Färbung haben ein aggressives Verhalten, oder sie fliehen rechtzeitig. Die genetische Grundlage dafür ist die Koppelung der Gene für die entsprechenden Merkmale, wodurch diese vor Rekombination geschützt sind. Dies führt zu klar unterschiedenen Morphen innerhalb der Art, bei denen ein bestimmtes Farbmuster jeweils mit einem bestimmten Antiprädator-Verhalten kombiniert ist. Auch hier haben wir, wie bei den Finken, wieder ein Beispiel für Selektionsverhältnisse, die nicht einen bestimmten Phänotyp als den Fittesten bevorteilen und gegenüber konkurrierenden Phänotypen durchsetzen, sondern der Selektionsvorteil liegt stattdessen in der Diversität innerhalb der Population. Der Selektionsvorteil ist die Bereitstellung mehrerer Morphen innerhalb derselben Art, um für mehrere Fälle variabler Umweltgefahren „gerüstet“ zu sein. Auch hier kommt wieder die Frage auf, warum sich der adaptive Polymorphismus durchgesetzt hat und nicht die adaptive Speziation, die sympatrische Artbildung mit reproduktiv abgegrenzten Einheiten. Die polymorphe genetische Grundlage der Verstädterung einiger Vogelarten Präadaptive genetische Anlagen scheinen häufig für Verhaltensänderungen einer ganzen Population verantwortlich zu sein; es sind nicht immer Lernprozesse. So spricht einiges dafür, dass auch die heute vielerorts beobachtete Verstädterung der Vögel auf genetischer Präadaptation beruht. Die Scheu vieler Tiere vor ihren potentiellen Feinden hat spezifische genetische Grundlagen. Ein frisch aus dem Ei schlüpfendes Hühnchen kann einen Sperber von einer harmlosen Taube am Flugbild unterscheiden, ohne dies von den Eltern oder sonstigen Artgenossen gelernt zu haben. Ein aus Nordsibirien an der deutschen Wattenmeerküste im August eintreffender im selben Sommer erst geborener Strandläufer (Calidris spec.) kann einen Menschen oder Hund von einem am Deich grasenden Schaf unterscheiden, ohne dass er mit Menschen, Hunden oder Schafen je eine Erfahrung gemacht hat. Die Scheu vieler Tiere vor dem Menschen und bestimmten Raubtieren ist das Produkt aus genetischen Anlagen und diversen Erfahrungen. Entscheidend sind frühkindliche Prägungen, eigene Erlebnisse und die Erziehung durch Eltern und andere Artgenossen, aber ein Großteil der Scheu gegen mögliche Feinde ist genetisch verankert. Bei Vögeln zeigt sich, dass die Scheu vor dem Menschen in verschiedenen geografischen Regionen sehr unterschiedlich ist und auch historischen Veränderungen unterliegt. Obwohl diese Beobachtung beim ersten Ansehen eher den Standpunkt unterstützt, die Scheu sei ein Resultat unterschiedlicher Erfahrungen mit dem Menschen, sprechen auch viele Daten für eine vorgegebene genetische Veranlagung. Und diese genetische Grundlage scheint in den Organismen einer Population polymorph verankert zu sein. Als die Städte seit hundert und mehr Jahren zunehmend größere Räume einnahmen und in ihren Qualitäten ländliche Räume und Wälder überboten, wechselten mehrere Vogelarten ihr angestammtes Habitat. Sie zogen mehr und mehr in die Städte ein, die ein größeres Nahrungsangebot und einen besseren Schutz vor Feinden boten. Die Skala der Beispiele reicht vom Graureiher (Ardea cinerea) über mehrere Entenarten, das Blässhuhn (Fulica atra) und das Teichhuhn (Gallinula chloropus), die Ringeltaube (Columba palumbus) und die Misterdrossel (Turdus viscivorus) bis zur Rabenkrähe (Corvus corone). Die Ringeltaube heißt im Englischen bemerkenswerterweise heute noch „Woodpigeon“, obwohl sie in vielen Teilen Europas (nicht in allen Teilen) kaum noch in den Wäldern vorkommt, sondern ganz und gar die Städte und die stadtnahen Bereiche erobert hat. Die Verstädterung der Ringeltaube hat sich in Nordwestdeutschland in den 1950er und 1960er Jahren vollzogen. Vorher war diese Taube ein ganz und gar scheuer Waldbewohner. Unter Jägern sollte mit dem Ausspruch „das ist eine Ringeltaube“ eine besondere Auszeichnung zum Ausdruck gebracht werden, so wie ein erfolgreicher Abschuss einer Ringeltaube von Seltenheit und Können zeugt. Heute fressen einem die Ringeltauben fast aus der Hand. Warum aber hat es so lange gedauert, bis die Tauben die Städte erobert haben? Die geringere Gefahr eines Abschusses in den menschlichen Wohngebieten und ein deutlich höheres Nahrungsangebot hätten doch schon seit viel längerer Zeit als einem halben Jahrhundert zur Verfügung gestanden. Und auch heute noch gibt es Regionen in Europa, in denen die Ringeltaube nicht verstädtert ist. Z.B. gibt es im Süden und Südosten Mitteleuropas kaum Stadt-Ringeltauben (Bezzel and Kooiker 2003 5126). Allein dieser geografische Vergleich zeigt schon, dass die Verstädterung nicht allein auf Lernprozessen beruhen kann; denn diese waren überall in Europa annähernd gleich. Vertreter der Auffassung, dass die Menschscheu der Vögel allein auf negativen Erfahrungen beruht, finden kein Argument dafür, warum der Weißstorch (Ciconia ciconia) schon seit Menschengedenken zum Menschen zutraulich ist, während sein naher Verwandter, der Schwarzstorch (Ciconia nigra), geradezu den Extremfall eines menschenscheuen Vogels darstellt. In den meisten Brutgebieten und vor allen auf den Zugstrecken und in den Überwinterungsgebieten zeigt sich keineswegs, dass die eine Art geschützt wird, während die andere Art verfolgt wird. Menschscheue beruht also höchstwahrscheinlich zu einem Teil auf genetischer Veranlagung, und diese Veranlagung kann auf verschiedenen Individuen polymorph verteilt sein, so dass eine Zunahme wie auch eine Abnahme der Menschenscheue einem Selektionsvorteil unterliegt, so dass folglich in einer Population die eine allele Ausstattung die andere zurückdrängen kann. Der deutliche Selektionsvorteil, in den Genuss der nahrungsreichen und sicheren Städte zu kommen, hat bei mehreren Vogelarten in wenigen Generationsfolgen zu einer deutlichen Veränderung der allelen Häufigkeitsverteilung geführt. Die Mimikry-Morphen bei weiblichen Schwalbenschwänzen der Gattung Papilio [Gruppenselektion] Intraspezifische Morphen waren schon in der Vergangenheit von erheblicher Tragweite, als es darum ging, die Taxonomie von einer rein merkmalsorientierten Gruppierung abzubringen und ihr eine naturwissenschaftliche Grundlage zu verschaffen. Vor mehr als hundert Jahren erkannte Edward Bagnall Poulton die Inkonsequenz und Widersprüchlichkeit eines allein auf Merkmalen beruhenden Artkonzepts. Seine klassischen Publikationen „What is a species?“ und „The conception of species as interbreeding communities” in den Jahren 1904 und 1938 in den „Proceedings of the Entomological Society of London” und den “Proceedings of the Linnean Society of London” (Mallet 2004 5330) können in ihrer Bedeutung kaum unterschätzt werden. Einerseits haben sie der Linnéschen Typologie die naturwissenschaftliche Grundlage entzogen. Sie haben die Einteilung nach Merkmalen entlarvt als das, was sie ist, eine subjektive Gruppierung nach unseren eigenen pragmatischen Zielen und Merkmalsbewertungen, oder (wie Darwin bereits 1859 gesagt hat): „made for our convenience“ (Darwin 1859 4355). Andererseits hat Poulton die wesentlichen Gedankengänge von Ernst Mayr bereits Jahrzehnte vorweggenommen, indem er das Grundkonzept der Art als Reproduktionsgemeinschaft vorgeschlagen hat. Poulton wurde von den Beobachtungen von Alfred Russel Wallace und Henry Walter Bates über die Mimikry-Morphen von Schmetterlingen beeinflusst (Mallet 2004 5330). Während Bates hauptsächlich südamerikanische Heliconiiden untersuchte, entdeckte Wallace die fascinierenden Mimikry-Morphen bei den südostasiatischen Schwalbenschwänzen Papilio memnon, P. polytes und P. aegeus. Bei diesen Arten tritt das Weibchen in mehreren sehr verschieden aussehenden Morphen auf. Die Männchen sind dagegen einheitlich gestaltet und treten nicht in unterschiedlichen Morphen auf. Die unterschiedlichen Weibchen-Morphen ahmen Schmetterlingsarten ganz anderer Familien nach, die für Vögel ungenießbar sind, weil ihre Raupen giftige Substanzen aufnehmen, die im Körper der fertigen Schmetterlinge (Imagines) noch enthalten sind. Aufgrund der Täuschung durch gleichartiges Aussehen sind die Nachahmer (genau wie die Nachgeahmten) vor Fressfeinden geschützt, obwohl sie selber gar keine Gifte enthalten. Dies hat einen erheblichen selektiven Vorteil. Es handelt sich hier um einen klassischen Fall (und historisch um einen der ersten publizierten Beispiele) für Bates’sche Mimikry, worunter die Nachahmung eines wehrhaften oder ungenießbaren Tieres durch ein harmloses Tier zur Täuschung von Feinden verstanden wird. Für Poulton lag die Bedeutung dieser Schmetterlinge vor allem darin, dass er erkannte, wie fehlerhaft es sein kann, vom Erscheinungsbild auf die Artzugehörigkeit zu schließen. Die Weibchen dieser Schwalbenschwänze unterscheiden sich innerartlich um ein Vielfaches stärker voneinander als sie sich von den Angehörigen anderer Schwalbenschwanzarten unterscheiden. Etwas später wurde dann dasselbe Phänomen beim afrikanischen Schwalbenschwanz Papilio dardanus entdeckt, der heute zu den bestuntersuchten Beispielen für Bates’sche Mimikry zählt (Abb., z.B. Tafel 81 in Sbordoni and Forestiero 1984 5782). Auch hier sind die Männchen einheitlich gestaltet, während die Weibchen polymorph sind. In Madagaskar und in Äthiopien gibt es nur ursprüngliche, monomorphe Populationen, ohne Mimikry, bei denen die Weibchen wie die Männchen Schwanzfortsätze am Hinterflügel und eine gelbschwarzes Färbung aufweisen. An anderen Orten in Ostafrika unterscheiden sich die Weibchen vom stets konstant gleichgestalteten Männchen drastisch und ahmen ungenießbare Schmetterlinge anderer Familien nach, z.B. die Danaiden Danaus chrysippus und Amauris niavius. Insgesamt wurden bei Papilio dardanus mehr als 30 deutlich unterscheidbare mimetische Morphen entdeckt (Salvato 1997 4228). In einigen Populationen von Papilio dardanus ahmen die Weibchen nur eine fremde Art nach, in anderen Populationen gleichzeitig mehrere. Diese Form des Mimikry-Polymorphismus ist nicht auf die Gattung Papilio beschränkt. Ein ganz analoges Beispiel tritt bei einem Schmetterling aus einer anderen Familie auf, nämlich aus der Familie der Nymphaliden. Bei diesem über mehrere Erdteile verbreiteten Schmetterling, Hypolimnas misippus, zeigen die Männchen ebenfalls keinen Polymorphismus, während die Weibchen mehrere Morphen ausbilden, darunter solche, die wiederum Danaus chrysippus nachahmen. Die genetische Grundlage für diesen geschlechtsgebundenen Polymorphismus beruht auf einem Komplex nicht rekombinierbarer Gene, die als genetische Kopplungsgruppe zusammenhängen. Jede Morphe exprimiert ihre eigene Gengruppe, so dass (ähnlich wie beim oberen Beispiel der Strumpfbandnatter) ein durch mehrere Gene bestimmtes Farbmuster, die Flügelform und auch bestimmten Verhaltensweisen aufeinander abgestimmt sind. Auf diese Weise kommt es zu komplexen Morphen, die ohne intermediäre Mischtypen auftreten. Es treten bei Papilio dardanus nebeneinander klar voneinander abgegrenzte Typen auf: neben der dardanus-Morphe existieren die Danaus-chrysippusMorphe, die Amauris-niavius-Morphe und viele weitere Morphen. Der ganze Genkomplex scheint Y-chromosomal lokalisiert zu sein, was die genetische Koppelung der Gene des Komplexes leicht erklären würde, da das Y-Chromosom weitgehend vor Rekombination geschützt ist. Bei Schmetterlingen (im Gegensatz zu den meisten anderen Tieren) sind die Weibchen das heterogametische Geschlecht. Sie haben also statt der XXKonstitution die XY-Konstitution, wie das bei den Säugetieren die Männchen haben. Meist tragen die Geschlechtschromosomen dann allerdings nicht die Bezeichnung XY, sondern ZW, wenn das heterogametische Geschlecht das Weibchen ist. Auf jeden Fall ist das Y oder WChromosom in hohem Maße vor genetischer Rekombination geschützt, so dass die Gene des Morphotypen-Komplexes nicht auseinandergerissen werden können. Das würde erklären, warum das Phänomen der Mimikry-Morphen bei den Schmetterlingen geschlechtsgebunden nur im Weibchen auftritt. Historisch wurde die Conspezifität der unterschiedlichen Papilio-Morphen daran entdeckt, dass die Kopulation der verschiedenen Morphen mit dem jeweils zugehörigen Männchen in der Natur beobachtet wurde. Das Vorkommen so unterschiedlicher Morphen, die alle zu ein und derselben Sexualgemeinschaft gehören, hat Poulton dazu veranlasst, neben der Gruppierung der Organismen in diagnostizierbare Einheiten neue Artkonzepte vorzuschlagen, die nicht reine Sortiereinheiten sind, sondern auf Naturgesetzlichkeiten beruhen. Er schlug vor, miteinander sich fortpflanzende Organismen zu Gruppen zusammenzufassen, die er „syngamisch“ nannte. Damit nahm er das Mayrsche Artkonzept der Reproduktionsgemeinschaft vorweg. Außerdem fasste er als ein weiteres Artkonzept die Nachkommen gemeinsamer Vorfahren zu einer Gruppe zusammen, die er „synepigonisch“ nannte. Das war, grob gesagt, die Vorwegnahme des cladistischen Artkonzepts. Die Morphen des brutparasitierenden Kuckucksweibchens (Cuculus canorus) Der Kuckuck (Cuculus canorus) ist eine zur Brutzeit über ganz Europa bis Ostasien verbreitete Vogelart. Sein auffälligstes Merkmal ist der Brutparasitismus. Der Kuckuck ist unfähig, seine Eier selbst zu erbrüten und die Jungen aufzuziehen. Stattdessen legt er seine Eier in die Nester fremder Vogelarten, wobei es sich fast ausschließlich um Angehörige der Unterordnung der Sperlingsvögel (Passeriformes) handelt. Diese Wirtsvögel erkennen die falsche Brut meistens nicht, brüten das Ei des Kuckucks aus und füttern den Jungvogel, bis dieser die Flugfähigkeit erreicht hat und auf die Betreuung durch die Zieheltern nicht mehr angewiesen ist. Der Kuckuck ist ein für die Speziesproblematik außerordentlich relevanter Fall, weil die Weibchen in zahlreichen Merkmalen an die jeweils spezifischen Wirtsvogelarten angepasst sind. Jedes Kuckucksweibchen hat nur eine Wirtsvogelart, in deren Nester sie die Eier legt. Das Kuckuckweibchen wechselt den Wirt nicht, selbst dann nicht, wenn es während der Legeperiode keine Nester für die Eiablage findet. Die herangewachsenen Jungkuckucke suchen im nächsten Frühjahr wieder die Nester der gleichen Wirtsvogelart auf. Allein in Europa gibt es ca. 90 verschiedene Wirtsvogelarten, an die die Kuckucksweibchen angepasst sind, und entsprechend gibt es 90 genetisch verschiedene Kuckucke. Allerdings werden nicht alle Wirtsvogelarten gleich häufig von Kuckucken parasitiert. Einige Arten werden besonders häufig von Kuckucksweibchen aufgesucht, andere Arten werden nur von wenigen Kuckucksweibchen aufgesucht. Und es gibt weitere Wirtsvogelarten, bei denen der Kuckucks-Nachwuchs nur eine geringe Chance hat, erfolgreich auszufliegen (Glutz von Blotzheim 2003 5180). In den asiatischen Ländern gibt es viele weitere Wirtsvogelarten, so dass die Gesamtzahl der Kuckuckswirte die 200 überschreitet. Die starke Anpassung an nur eine Wirtsvogelart ist in zahlreichen erblichen Merkmalen festgelegt. Das Kuckucksweibchen muss zur rechten Zeit fähig sein, seine Eier zu legen. Rotkehlchen (Erithacus rubecula), Hausrotschwänze (Phoenicurus ochruros) und Drosseln (Turdus spec.) als häufig aufgesuchte Wirte brüten in Mitteleuropa schon Ende April, Rohrsänger (Acrocephalus spec.) erst gegen Ende Mai. Darauf muss der Kuckuck hormonal eingestellt sein. Seine Eier dürfen nicht zu früh und nicht zu spät ablagereif sein. Der Kuckuck muss das Nest des richtigen Wirtsvogels finden und erkennen. Dazu verfügt er allenfalls über Erinnerungen an das Vorjahr, als er in einem solchen Nest groß geworden ist. Er hat aber selbstverständlich niemals eine elterliche Anleitung gehabt, wie er den geeigneten Brutbiotop findet und von anderen Habitaten unterscheidet und wo er den potentiellen Neststandort des Wirtsvogels zu finden hat, ob hoch im Baum oder im niedrigen Gebüsch oder am Erdboden. Kuckucksweibchen findet geeignete Nester durch Beobachtung, indem sie gegebenenfalls stundenlang an verborgener Stelle warten, um den Nestbau und den richtigen Zeitpunkt des Bebrütungsbeginns des Wirtvogels zu beobachten und sich darauf einzurichten. Auch ist wahrscheinlich, dass die Wirtsvögel akustisch am Gesang erkannt werden. Auch das muss größtenteils erblich festliegen, da keine elterliche Anleitung möglich ist, sondern allenfalls die Erinnerung an die Bedingungen der eigenen Nestlingszeit. Der Brutparasitismus kann weiterhin nur deshalb funktionieren, weil die Kuckuckseier in ihrer Färbung und Zeichnung den Eiern der Wirtvögel angepasst sind. Das heißt, dass es in Eurasien innerhalb der Art Cuculus canorus mehrere hundert verschiedene genetische Varianten gibt, die den Färbungsmodus der Eier festlegen. Das einzelne Weibchen einschließlich seiner weiblichen Nachkommen legt nur Eier eines ganz bestimmten Typs. Diese Mimikry der Kuckuckseier kann nur durch das Vorhandensein polymorpher Allele erklärt werden. Für das Verständnis des Artbegriffs liegt das Hauptproblem nicht allein in der perfekten Anpassung, sondern vielmehr darin, dass diese Anpassung nur durch die Kombination zahlreicher Gene erklärbar ist, die alle in einem Kuckucksweibchen zusammen wirken müssen. Der Zeitpunkt der Eiablage, die Erkennung des Wirtes und die Eifärbung sind durch genetische Anlagen unterstützt, die nicht durch Rekombination getrennt werden dürfen, weil ein bestimmtes Kuckucksweibchen nicht in der Hälfte seiner Merkmale an den einen Wirtsvogel und zur anderen Hälfte an einen anderen Wirtsvogel angepasst sein darf. Alle Anpassungsmerkmale, die auf einen bestimmten Wirtsvogel abgestimmten sind, dürfen nicht rekombinieren. Sie müssen genetisch als Ganzheiten untrennbar miteinander gekoppelt sein. Also gibt es genetisch unterscheidbare Rotkehlchen-Kuckucke, Hausrotschwanz-Kuckucke, Drossel-Kuckucke, Rohrsänger-Kuckucke usw. Warum also sind es nicht 200 bis 300 Kuckucksarten, die Eurasien besiedeln? Die Antwort darauf lautet, dass die Anpassung an bestimmter Wirtsvogelarten nur durch die weiblichen Kuckucke aufrechterhalten wird. Bei den Männchen gibt es keine Wirtsspezifität. Also handelt es sich wiederum, wie schon beim Mimikry-Polymorphismus von Schwalbenschwanz-Weibchen (siehe oben), um Morphen, und zwar wiederum um Morphen, die nur im weiblichen Geschlecht auftreten. Warum gerade im weiblichen Geschlecht? Es scheint kein Zufall zu sein, dass ausgerechnet die Schmetterlinge und die Vögel, bei denen die ans Weibchen gebundenen Morphen auftreten, jene Tiergruppen sind, die sich von den anderen Tiergruppen dadurch unterscheiden, dass bei ihnen die Weibchen das heterogametische Geschlecht sind, und nicht wie üblich die Männchen. Bei Schmetterlingen und Vögeln haben die Weibchen die XYKonstellation (manchmal ZW genannt), während die Männchen die homogametische XXKonstellation haben. Das Y-Chromosom (W-Chromosom) enthält einen großen Teil von Genen, die im Heterochromatin eingebettet sind und die daher vor der Rekombination geschützt sind und eine sog. Kopplungsgruppe bilden. Vermutlich sind hier alle Gene lokalisiert, die im Verbund miteinander die unterschiedlichen Morphen bestimmen. Das würde erklären, warum eine ganze Batterie von Genen beisammen bleibt und über die weibliche Linie vererbt wird. Wäre der Kuckuck wie ein Säugetier im weiblichen Geschlecht homogametisch, so könnte es vermutlich keinen Brutparasitismus geben.