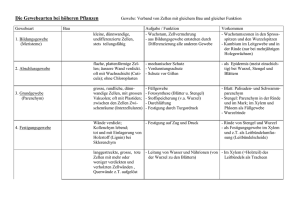
HISTOLOGIE
Werbung

55 HISTOLOGIE Auf die Besprechung der Pflanzenzelle, also die Cytologie, folgt die Lehre vom Bau der Gewebe, die man als Histologie bezeichnet. Die Histologie der Samenpflanzen, die uns hier allein interessiert, ist komplizierter als die der Tange oder Moose, aber immer noch weit einfacher als diejenige der höheren Tiere. Eine Samenpflanze beginnt ihr Leben als befruchtete Eizelle, also im einzelligen Stadium. Die Entwicklung zum Embryo und weiter zur reifen Pflanze erfordert Wachstumsvorgänge: Zellteilungen vermehren die Zahl der Zellen, Zellstreckungen vergrößern sie und Differenzierungen passen die Zellen an spezialisierte Aufgaben an. Die Zellen sind nicht unabhängig von einander, sondern in Stoffwechsel, Wachstum und Leistung aufeinander abgestimmt, um ihre Aufgaben im Pflanzenkörper erfüllen zu können. Man fasst Gruppen von Zellen zu Geweben zusammen, wenn sie gemeinsame Züge aufweisen. Die Histologie beschreibt und erforscht die pflanzlichen Gewebe. Es gibt freilich viele Möglichkeiten, die Gewebe einzuteilen. Man kann anatomische, physiologische, entwicklungsgeschichtliche oder funktionelle Aspekte bevorzugen; die einzelnen Betrachtungsweisen sind dabei grundsätzlich gleichberechtigt. Unsere Einteilung der Gewebe deckt sich daher auch nicht mit derjenigen in allen Lehrbüchern der Allgemeinen Botanik. Allgemein anerkannt ist die Gegenüberstellung von Bildungsgeweben oder Meristemen einerseits und Dauergeweben andererseits. Wir wollen uns zuerst den Meristemen zuwenden. Zu Beginn der Samenkeimung teilen sich Zellen im ganzen Körper des Embryos. Später wird jedoch die Neubildung von Zellen auf bestimmte Körperstellen beschränkt, während andere Teile neue Aufgaben übernehmen. Die erwachsene Pflanze enthält neben differenzierten weiterhin auch embryonale Gewebe. Diese, die Meristeme, erzeugen fortlaufend neue Zellen und daraus neue Gewebe und Organe, ohne sich dabei zu verbrauchen. Dies ist ein Unterschied zur Organisation der Tiere, die nach dem Jugendwachstums die Zellteilungen größtenteils einstellen und nur eine begrenzte Zahl von Organen haben. Meristeme sind nicht ständig aktiv. Außenbedingungen, etwa Winterkälte oder Sommertrockenheit, unterbrechen dieTeilungstätigkeit. Auch physiologische Einflüsse (Hormone; s.u.) können Meristeme für lange Zeit 55 56 inaktiv halten. So wird oft das Wachstum der Seitenknospen unterdrückt, solange der Haupttrieb wächst. Die embryonalen Zellen der Meristeme unterscheiden sich von voll ausgewachsenen Zellen in einigen Punkten. Die Zellen sind klein, im Verhältnis zum Volumen ist der Kern groß. Sie enthalten Proplastiden, aber keine Chloroplasten oder Chromoplasten. Die dünnen Wände haben keine Sekundärschichten. Vakuolen sind, wenn überhaupt vorhanden, meist klein. Es gibt verschiedene Typen von Meristemen, und auch auf ihre Einteilung sind verschiedene Prinzipien angewendet worden. Am einfachsten ist die Gliederung nach der Lage im Pflanzenkörper. Wir erhalten so: a) Apikalmeristeme oder Spitzenmeristeme an den Vegetationspunkten von Wurzel und Sproß; sie bauen neue Organe auf. b) Lateral- oder Flankenmeristeme an den Seiten der Organe; sie bauen neue Gewebe in bestehenden Organen auf. Wir werden Apikalmeristeme und Lateralmeristeme im Zusammenhang mit dem Bau der Organe näher besprechen, in denen sie auftreten. Daneben gibt es in Dauergewebe eingebettete Komplexe embryonaler Zellen, die zwar in einem Ruhezustand länger meristematisch bleiben als ihre Umgebung, sich jedoch dann nach wenigen Teilungen erschöpfen. Das sind die Restmeristeme oder interkalaren Meristeme. Dazu gehören die an der Basis der Blätter sitzenden Teilungsgewebe und das Gewebe im Grasknoten, das niedergedrückte Grashalme zur Aufrichtung befähigt. Nach wenigen Teilungen differenzieren sich die Zellen aus. Restmeristeme spalten sich fortlaufend von Apikalmeristemen ab, sind also deren Produkte. Sie enthalten oft auch differenzierte Elemente; so werden etwa die Grashalmknoten von wasserleitenden, also toten Gefäßen durchsetzt. All dies zeigt, daß die Interkalarmeristeme den Apikal- und Lateralmeristemen nicht gleichrangig sind. Die Dauergewebe der Samenpflanze entstehen aus den meristematischen Zellen durch Zellteilung, Streckung und Differenzierung. Alle Dauergewebe, die vor Einsetzen des sekundären Dickenwachstums gebildet werden, leiten sich von den Apikalmeristemen ab. Diese, die sogenannten Vegetationspunkte an den Spitzen des Sprosses und der Wurzel, geben laufend Zellen ab; die abgegliederten Zellen teilen sich noch eine Zeit lang weiter, sodaß die Enden von Sproßachse und Wurzel von einer in ständiger 56 57 Teilung begriffenen Meristemzone eingenommen wird. Diese Teilungszone ist allerdings nur Bruchteile eines Millimeters lang. Anschließend kann man an der unterschiedlichen Größe und Teilungsrichtung der Zellen schon erkennen, welche Gewebe der erwachsenen Pflanze aus ihnen entstehen werden. Auf diese Determinationszone folgen Streckungszone und Differenzierungszone. Die Zellen strecken sich unter Wasseraufnahme und Vakuolenbildung, sie verlieren ihre Teilungsfähigkeit und erreichen schließlich das endgültige Volumen und die Gestalt, die für Dauergewebezellen der einen oder der anderen Art charakteristisch sind. Nach Erreichen der endgültigen Zellgröße endet die Differenzierung erst, wenn die Zellen ihrer künftigen Aufgabe völlig entsprechen. Von der Gesamtheit der genetischen Möglichkeiten einer Zelle wird bei den Differenzierungsvorgängen jeweils nur ein begrenzter, je nach Zelltyp verschiedener Anteil verwirklicht. Mit anderen Worten heißt das, dass von allen Enzymproteinen, deren Struktur in der DNA des Kernes festgelegt ist, immer nur ein Teil gebildet wird. Dabei kommt es jedoch nicht nur darauf an, daß die Enzyme erzeugt werden, sondern die korrekte Differenzierung ist nur dann gewährleistet, wenn die Enzyme rechtzeitig und in der richtigen Reihenfolge zur Verfügung stehen. Andererseits muß ihre Produktion auch rechtzeitig wieder "abgedreht" werden. Die Hauptfrage der Differenzierung kann daher nur die Molekularbiologie beantworten. Sie lautet: Wie wird darüber entschieden, welche der zahlreichen möglichen Boten-RNA - Moleküle zu welchem Zeitpunkt gebildet werden? - Als überregionale Mittel zur Steuerung produzieren bestimmte Teile der Pflanze (besonders Blätter und Wurzeln) Botenstoffe, die Phytohormone. Man unterscheidet 5 Gruppen: Auxine, Giberelline, Cytokinine, Abscisinsäure, Ethylen. Dazu kommen noch einige Wachstumsregulatoren (Brassinolide, Jasmonsäure u.a.), die eher lokal begrenzt wirken. Ergebnis der Differenzierung sind voll ausgebildete, funktionsfähige Dauergewebe. Wenn sich ihre Zellen direkt von den Apikalmeristemen herleiten, nennt man sie primäre Gewebe. Die Teilung der Lateralmeristeme produziert sekundäre Gewebe. Wir können Dauergewebe nach ihrer Lage, Entwicklung und Funktion für unsere Zwecke in drei große Gruppen einreihen, die man Hautgewebe, Grundgewebe und Stranggewebe nennt. Wir wollen 57 58 Bau und Funktion dieser Gruppen der Reihe nach besprechen. Die Einteilung ist, wie gesagt, nicht verbindlich! Das primäre Hautgewebe Das primäre Hautgewebe ist die äußerste Zellschicht aller Organe der Spermatophyten und der Farne (zusammen: der Kormophyten). Es kommt in zwei verschiedenen Ausbildungen vor, die sich voneinander grundlegend unterscheiden. Die Funktionen dieser beiden Erscheinungsformen des Hautgewebes sind geradezu gegensätzlich, und das hat auch Auswirkungen auf den Bau. Daher bezeichnet man sie mit verschiedenen Namen: die Epidermis bedeckt die Organe des Sprosses, also Stämme und Blätter; die Rhizodermis oder Wurzelepidermis findet sich an den Wurzeln. Die Epidermis muß isolieren, die Rhizodermis stellt Kontakt mit dem Boden her. Die Epidermis wird vom Apikalmeristem des Sprosses, dem Sproßvegetationspunkt, angelegt. Die Zellen der äußersten Zellschicht in der Determinationszone, des Protoderms, bilden die neuen Wände bei der Teilung nur senkrecht zur Oberfläche, nicht jedoch parallel dazu. Das Hautgewebe bleibt daher einschichtig. Die Zellen schließen mit einer Ausnahme eng aneinander; diese Ausnahme bildet der Bereich einer Spaltöffnung. Die normalen Aufgaben der Epidermis an oberirdischen Pflanzenteilen sind folgende: a) Einschränkung der Transpiration b) Gasaustausch durch die Spaltöffnungen c) mechanischer Schutz d) Speicherung von Wasser und Stoffwechselprodukten Die Lebensdauer der Epidermis ist sehr verschieden. Organe, deren sekundäres Dickenwachstum unbedeutend oder nicht vorhanden ist, behalten ihre Epidermis, solange sie selbst leben. So kann die Epidermis mancher Kakteen viele Jahrzehnte alt werden, die von Koniferennadeln oder immergrünen Laubblättern der mediterranen Pflanzen mehrere Jahre. Stämme und Seitentriebe von Holzpflanzen hingegen beginnen schon im Jahr ihrer Ausbildung mit dem sekundären Dickenwachstum, also der Vermehrung ihres Leitgewebes mit Hilfe eines Lateralmeristems. Dadurch wird der Umfang des Stammes erweitert, und dem kann die Epidermis nicht lange folgen. Ihre Zellen 58 59 sind nämlich nicht mehr teilungsfähig und zerreißen bald unter dem Druck von innen, den die Vermehrung und die Streckung von Zellen in der Achse ausüben. Das müßte eigentlich zum Vertrocknen dieser Triebe führen, da die Epidermis sie nicht mehr schützt. Doch entsteht gleich nach Beginn des sekundären Dickenwachstums unter der Epidermis ein sekundäres Hautgewebe, das dem Wachstum durch Teilung seines Lateralmeristems folgt. Welche Zelltypen finden sich in der Epidermis? Die Hauptmasse des primären Hautgewebes besteht aus den eigentlichen Epidermiszellen, welche nur wenig spezialisiert sind. Dazwischen eingesprengt sind die Schließzellen der Spaltöffnungen, oft begleitet von besonders geformten Nebenzellen, sowie verschiedene Auswüchse, die als Trichome bezeichnet werden. a) Die eigentlichen Epidermiszellen: Im Querschnitt erscheinen diese Zellen meist flach. In der Aufsicht sind alle Übergänge vom isodiametrischen zum stark gestreckten Typ möglich. Langgestreckte Zellen sind meist an Pflanzenteilen mit ausgeprägtem Längenwachstum zu finden, etwa an Stengeln; die Epidermiszellen folgen nämlich dem Streckungswachstum der Organe. Häufig zeigen die Wände, die auf die Oberfläche senkrecht stehen, in der Aufsicht wellige Struktur. Die Fläche, an der benachbarte Zellen aneinander haften, wird auf diese Weise vergrößert, was die mechanische Festigkeit des Gewebes zweifellos erhöht. Wellige Wände finden sich besonders im Hautgewebe der Blätter, und tatsächlich sind Blätter durch scherende Kräfte, die der Wind ausübt, besonders beansprucht. Die Wände sind im allgemeinen ungleich dick. Die äußere Wand, der eigentliche Abschluß des Pflanzenkörpers, ist meist sehr stark ausgebildet. Die Undurchlässigkeit für Wasser und Wasserdampf wird durch das Cutin bewirkt, das in Form eines dünnen Häutchens, der Cuticula, außen der Zellwand aufgelagert ist, oft aber auch Teile der Zellwand durchtränkt, die dann Cuticularschichten enthält. Die Cuticula überzieht alle Teile des Sprosses (Achsen und Blätter) und verhindert durch ihren Cutingehalt die Wasserabgabe aus diesen Zellen bis auf einen geringen Restwert, die cuticuläre Transpiration. Die Entwicklung der Cuticula war einer der wesentlichen Schritte zur Eroberung des Landes durch die Pflanzen. Wir werden das noch besser begreifen, wenn wir Wassertransport und Wasserabgabe näher betrachten werden. - Die Oberfläche der Cuticula ist häufig noch mit kristallinen Wachsablagerungen bedeckt. Regentropfen 59 60 perlen von ihnen ab und nehmen dabei Staubteilchen und Mikroorganismen mit. Hingegen wäre ein Wasserfilm ein Keimmedium für Pilze und Bakterien. Auch könnte stehendes Wasser langsam einsickern und Schaden anrichten. So platzen die Früchte von Zuchtsorten gelegentlich durch übermäßige Wasseraufnahme. Dicke Wachsablagerungen finden sich tatsächlich besonders an Früchten; denken Sie etwa an blaubereifte Zwetschken oder Trauben. Im Rasterelektronenmikroskop zeigen die Wachse lockere, oft bizarre Strukturen. Sie bilden keinen geschlossenen Film, der die Transpiration verringern könnte. Cutin und Wachse wandern in flüssiger Form durch die zellulosische Zellwand und erstarren erst an der Oberfläche durch Reaktion mit dem Luftsauerstoff. Dicke der Wand, Cutinschichten und Wachse zeigen ökologische Anpassungen, besonders an die Beanspruchung des Wasserhaushaltes. Epidermiswände enthalten gelegentlich Lignin, so etwa bei den Koniferen; häufiger wird Kieselsäure (SiO2-Polymere) in Form amorpher Salze abgelagert; das ist charakteristisch für Schachtelhalme ("Zinnkraut"), echte Gräser und Riedgräser. Die Kieselsäure ist hier als Rekret mit Funktion anzusehen: Sie verleiht den Zellen mechanische Festigkeit. Eigentliche Epidermiszellen enthalten meist keine Chloroplasten. Die Vakuolen enthalten oft Anthocyane oder zwar farblose, aber stark UV-absorbierende Flavonverbindungen. Man hat experimentell bewiesen, daß die Epidermis als Schutzfilter für die UV-empfindlichen Chloroplastenpigmente in den darunterliegenden Assimilationszellen des Grundgewebes dient. Das paßt gut dazu, daß sich bei extremen Schattenpflanzen und bei submersen, also untergetaucht lebenden Wasserpflanzen sehr wohl funktionsfähige Chloroplasten in der Epidermis finden. Hier ist ja für die Chloroplastenpigmente keine Gefahren durch zu viel Licht oder durch Licht von schädlicher Wellenlänge zu befürchten. b) Der Spaltöffnungsapparat (Schließzellen und Nebenzellen): Die lückenlos aufgebaute Epidermis wird durch winzige Öffnungen durchbrochen, die von jeweils zwei hochspezialisierten Zellen eingefaßt sind. Man bezeichnet die Öffnungen als Spaltöffnungen, das Zellenpaar als Schließzellen. Sie enthalten als einzige Epidermiszellen Chloroplasten. Durch Formwechsel der Schließzellen wird die Größe der Öffnung verändert. Die funktionelle Einheit aus Schließzellen und Spaltöffnung wird als Stoma (griech. 60 61 "Mund", Mz. Stomata) bezeichnet. Die Spaltöffnung führt zunächst in einen Hohlraum, die Atemhöhle, von dem aus zahlreiche Interzellularen, luftgefüllte Kanäle, das Grundgewebe durchziehen. Die Stomata sind besonders typisch für die grünen Sproßteile, vor allem die Laubblätter, doch finden sie sich auch auf Blütenblättern und farblosen, unterirdischen Sproßteilen (Rhizomen), nicht jedoch auf Wurzeln. Ihre Verteilung über die Blattseiten ist verschieden. Meist sind sie auf der Blattunterseite gehäuft, manchmal sogar ausschließlich anzutreffen. Auch die Zahl der Spaltöffnungen ist artabhängig. Extremwerte dürften 50 und 900 Stück je mm 2 Blattfläche sein. Für ein Blatt der Sonnenblume ergeben sich ungefähr 13 Millionen Öffnungen. Insgesamt macht die Porenfläche jedoch nur 1 - 3% des gesamten Blattareals aus. Die Wuchsbedingungen können Zahl und Größe der Spaltöffnungen einer Art modifizieren, das Porenareal ist also veränderlich. Ein besonders kleines Porenareal besitzen Typen, die gut an Trockenstandorte angepaßt sind. Die Spaltöffnungen reagieren in einer ökologisch sinnvollen Weise auf eine ganze Reihe von Umwelteinflüssen. Es scheinen hier zwei Regelkreise zu wirken. Der CO2-Regelkreis spricht auf die Konzentration von CO2 im Inneren des Blattes an. Man kann ihn völlig rein studieren, wenn man abgezogene Epidermen im Dunkeln mit Luft von verschiedener CO2-Konzentration bespült. Die Luft der freien Atmosphäre hat eine CO2-Konzentration von ungefähr 360 ppm, das heißt 360 µl*l-1, sechsunddreißig Hunderttausendstel des Volumens, 0.036 Volumsprozent. (Ich nenne alle diese verschiedenen Formulierungen, da hier in der Literatur ziemliche Willkür herrscht). Wenn man nun die Konzentration der Luft im Experiment absenkt, dann bleiben die Spalten im Dunkeln bis etwa 220 - 250 ppm geschlossen, darunter öffnen sie sich. Im Licht sinkt bei den Pflanzen der CO2-Gehalt im Inneren des Blattes, da ja das CO2 bei der Photosynthese verbraucht wird. Daher öffnen sich die Spalten am Morgen und schließen sich nach Einbruch der Nacht, wenn die Atmung der Gewebezellen die CO2-Konzentration im Blattinneren erhöht und dieses Gas nicht mehr durch Photosynthese verbraucht wird. Eine Ausnahme bildet der CAM-Typ der Photosynthese (s.u.). 61 62 Anders funktioniert der H2O-Regelkreis. Wenn die Wasserversorgung der Zellen erschwert wird, sinkt ihr Wassergehalt. Es wird Abscisinsäure (ABA) freigesetzt, und diese induziert Turgorverlust der Spaltöffnungen, also eine Volumsverkleinerung. Das aber bewirkt, daß sich die Spalten verengen. Das versteht man erst, wenn man den Mechanismus der Spaltbewegungen näher angesehen hat. Hier gibt es eine Reihe von kleinen Abwandlungen eines Grundprinzips. Wir wollen nur Bau und Funktion des häufigsten Typs von Spaltöffnungen, des sogenannten Helleborus-Typs, besprechen. Die Schließzellen dieses Typs sind nierenförmig in der Aufsicht. Der Durchmesser der Spaltöffnung, die zwischen ihnen freibleibt, schwankt, wenn sich das Volumen der Schließzellen ändert. Wie kann sich das Volumen ändern? Natürlich durch Aufnahme oder Abgabe von Wasser; die Zelle füllt sich prall mit Wasser oder sie erschlafft: Ihr Turgor steigt oder sinkt. Man würde nun zunächst annehmen, daß durch Volumsvergrößerung der begrenzenden Zellen der Durchmesser der zwischen ihnen liegenden Öffnung verkleinert wird. Da aber die Rückwand der Zellen weit dünner ist als die anderen Wände, kann sich die Zelle nur dort verformen und ausdehnen. Sie zieht dabei die dünne Wandpartie im Bereich der engsten Stelle, des sogenannten Zentralspaltes, mit sich. Die Anordnung der Zellulosefibrillen in den Wänden unterstützt noch diese einseitige Ausdehnung. Was bewirkt die Volumszunahme? Es handelt sich um osmotische Vorgänge. Wenn die Konzentration von gelösten Stoffen im Zellsaft steigt, dann strömt in die Vakuole Wasser ein; wenn die Zellsaftkonzentration sinkt, dann wird Wasser an die Nebenzellen abgegeben. Öffnen und Schließen sind mit lebhaften Verlagerungen von Kaliumionen verbunden. Beim Öffnungsvorgang wird K+ durch aktiven Transport in die Schließzellen aufgenommen. Der Konzentrationsanstieg zieht passiv Wasser nach, das ja leicht durch die Plasmagrenzschichten in die Vakuole dringen kann, wo das Kalium gespeichert + wird. Das positiv geladene K kann freilich nicht allein aufgenommen werden, sondern zum Ladungsausgleich ist noch ein negativ geladenes Anion nötig. Diskutiert wird, welcher Säurerest sich hier beteiligt. Es scheint dies verschieden zu sein. Bei Gräsern wird meist Cl- mitgepumpt, bei anderen Arten werden aus der Stärke der Schließzellchloroplasten, die ja als einzige Plastiden 62 63 der Epidermis photosynthetisch aktiv sind, organische Anionen synthetisiert, anscheinend meist das Ion der Apfelsäure, das Malat. - Über die Rolle der Spaltöffnungen im Wasserhaushalt und über ihr Verhalten im Freiland wird noch gesprochen werden. Die Schließzellen der Spaltöffnungen grenzen oft nicht an eigentliche Epidermiszellen, sondern an kleinere, besonders geformte Zellen, die nach ihrer Lage neben den Spaltöffnungsschließzellen Nebenzellen heißen. Ihre Form scheint im Laufe der Ausdifferenzierung des Protoderms sozusagen erzwungen zu werden; sie haben trotz ihrer charakteristischen Gestalt keine eigenständige Funktion, sondern sie ähneln in allen wichtigen physiologischen Eigenschaften den typischen Epidermiszellen. Einzelne Spaltöffnungen können bei manchen Pflanzenarten einen Funktionswechsel erfahren und flüssiges Wasser ausscheiden. Man kann diesen Vorgang, die Guttation, besonders in feuchter Atmosphäre, also etwa am frühen Morgen auf der Wiese, beobachten. Die wasserausscheidenden Öffnungen bezeichnet man als Hydathoden oder Wasserspalten. c) Die Trichome (Pflanzenhaare und Emergenzen): Haare sind Anhangsgebilde des Pflanzenkörpers, die definitionsgemäß nur aus Hautgewebezellen bestehen. Dadurch unterscheiden sie sich von den Emergenzen, an deren Bildung auch das darunterliegende Grundgewebe beteiligt ist. In der Funktion ist zwischen beiden wenig Unterschied, wenn auch Emergenzen meist massiver sind. Wir wollen hier diese Auswüchse, die man zusammen Trichome nennt, gemeinsam besprechen,. Haare entstehen aus Zellen des Hautgewebes, und zwar sowohl der Epidermis wie der Rhizodermis. Epidermishaare zeigen sehr verschiedene Formen. So strecken sich manchmal die Epidermiszellen bloß zu langen Fadenzellen, oder sie werden durch Querteilungen gegliedert. Manche Haare verzweigen sich sogar und bilden dann Gestalten wie bizarre Bäumchen. Grundsätzlich muß man zwischen lebenden und toten Haaren unterscheiden, die sehr verschiedene Funktionen haben. Lebende Haare vergrößern die Oberfläche und steigern die Verdunstung. Solche Haare finden sich oft an frisch entfalteten Blättern, etwa bei der Buche oder der Platane. Sobald das Blatt erwachsen ist und die Spaltöffnungen funktionieren, verkahlen solche Blätter oft, oder die Behaarung 63 64 bleibt nur an wenigen Stellen erhalten. Tote Haare werden weiß, sobald Luft eindringt. Sie sind zunächst für den Strahlungshaushalt des Blattes wichtig, da sie das Licht reflektieren und damit bei voller Sonneneinstrahlung die Überhitzung verhindern oder wenigstens reduzieren. Weiße Haare sitzen manchmal auf beiden Blattseiten, öfter aber an der Blattunterseite, wo sie ihre Aufgabe der Strahlungsreflexion besonders bei jungen und bei welken Blättern erfüllen, weil dann das Licht auf sie fällt. An der Blattunterseite haben weiße Haare aber noch eine weitere Funktion: Sie verlangsamen die Luftbewegung an der Oberfläche, wodurch die Feuchtigkeit, die aus den Spaltöffnungen entweicht, nicht so rasch abgeführt wird. Dadurch ergibt sich ein Rückstau der Feuchtluft, der seinerseits die Transpiration des Blattes reduziert. Wir finden daher tote, weißfilzige Haarüberzüge vor allem an Pflanzen von Trockenstandorten. Diesen beiden Hauptfunktionen der Haare, der Steigerung der Transpiration im einen, ihrer Verringerung im anderen Fall, stehen eine große Anzahl von Spezialfunktionen gegenüber. An Samen sitzen oft Flughaare. Sehr häufig ist die Ausbildung von Drüsenhaaren, die in mehreren Familien auftreten. Bei den Labiaten sezernieren sie ätherische Öle, in den Knospenschuppen vieler Laubbäume, etwa der Roßkastanie, klebrige Substanzen, die die Knospe gegen Wasserverluste abdichten. Bei insektenfressenden Pflanzen werden Verdauungsenzyme von Haaren gebildet und ausgeschieden. Eine Sonderform der Drüsenhaare sind die Brennhaare der Brennessel. Ihr wichtigster Teil, eine riesige, flaschenförmig aufgetriebene Haarzelle, sitzt auf einem Sockel aus Grundgewebe. Das ganze Gebilde ist daher eine Emergenz. Die Haarzelle verjüngt sich nach oben stark und endet mit einem kleinen, schräg aufgesetzten Köpfchen. Die unmittelbar darunter liegende Stelle ist dünn und sehr spröde, da sie mit Kieselsäure inkrustiert ist. Wird das Köpfchen leicht berührt, so bricht es an dieser "Sollbruchstelle" ab. Das neue offene Haarende wirkt wie eine Injektionsspritze und dringt in die Haut ein, in die sich der Haarzell - Inhalt ergießt. Da dieser Histamin und Acetylcholin in hoher Konzentration enthält, schmerzt die Wunde und entzündet sich. Es ist schwer vorstellbar, daß ein solches Gebilde zufällig entstehen könnte; eher scheint es, daß diese Sekretzelle im Dienste der Verteidigung das Ergebnis einer längeren Optimierung durch Selektion darstellt. 64 65 Typische Emergenzen sind die Stachel, wie man sie bei der Rose und der Brombeere findet. Sie können Wirbeltiere abwehren, sie können aber auch als Kletterhilfe dienen. Der Botaniker unterscheidet den Stachel, eine Bildung aus epidermalen und subepidermalen Geweben eines Organes, vom Dorn, bei dem ein ganzes Organ zu einem stechenden Gebilde umgewandelt ist. In diesem Sinne ist jede Rose ohne Dornen! Soviel zur typischen Epidermis der Sproßorgane. Die Wurzelepidermis oder Rhizodermis bedeckt den Teil der Wurzel, mit dem diese Wasser aus dem Boden aufnimmt. Während also die Epidermis den Wasseraustausch mit der Umgebung stark einschränkt, ist er für die Rhizodermis eine Hauptaufgabe. Ihr entspricht die Struktur: Die Wände sind dünn und nicht von einer Cuticula bedeckt. Spaltöffnungen fehlen, da ja der Gasaustausch nicht von der Cuticula behindert wird. Ein Filz einzelliger, lebender, schlauchförmiger Haare, der Wurzelhaare, vergrößert die Oberfläche um mindestens den Faktor 10 (etwa bei der Erbse auf das Zwölffache!). Die Rhizodermis schließt also den Pflanzenkörper nicht ab, sondern bringt ihn in engen Kontakt mit der Außenwelt. Wir werden über die Rhizodermis und ihr Zusammenspiel mit den übrigen Geweben der Wurzel später noch mehr erfahren. Das sekundäre Hautgewebe Wir wissen bereits, daß die Epidermis der Sproßachse nur die grünen Pflanzenteile bedeckt und oft nur eine beschränkte Lebensdauer hat. Sie kann nämlich dem Wachstum von Organen, die sich stark verdicken, nicht folgen. Ihre Zellen sind kaum teilungsfähig und zerreißen, sobald der Druck von innen zu groß wird. Sekundäre Ersatzgewebe, die Periderme, sichern dann weiter den Abschluß. Auch sie sind wasserundurchlässig. Diese Abdichtung bewirkt das Suberin in der Zellwand. Wir finden Periderme dort, wo die Achse sekundär wächst, also an Stämmen von Dikotylen und Gymnospermen. Auch dicke primäre Speicherknollen (Kartoffel) und Früchte (Alexanderbirne) bilden Periderme. Die Monokotylen, die zweite Unterklasse der Angiospermen, haben kein sekundäres Dickenwachstum und daher keine Periderme. Suberineinlagerungen in Einzelzellen oder Zellkomplexen kommen dagegen häufig auch bei Monokotylen und Farnen vor. Man muß also Verkorkung (=Suberineinlagerung) und Ausbildung eines echten Korkgewebes (eines 65 66 Periderms) auseinanderhalten, ebenso wie Verholzung (=Lignineinlagerung) nicht nur im Holz, sondern auch in anderen Teilen der Pflanze auftreten kann und auch bei Pflanzen, die kein Holz bilden. Verkorkung von Einzelzellen und Zellschichten tritt bei allen Kormophyten nach Verwundung auf, da eine zerstörte Epidermis "geflickt" werden muß, damit keine Trockenschäden an den darunterliegenden Zellen auftreten. Und nun zum echten Periderm. Das erste Periderm an einem jungen Stamm ersetzt die Epidermis; nach einiger Zeit des Dickenwachstums wird dieses erste Periderm meist durch Folgeperiderme abgelöst. Jedes Periderm besteht aus zwei bis drei Teilen: a) dem Phellogen, einem Lateralmeristem, das das Periderm erzeugt, b) dem Phellom oder Kork, dessen Zellen vom Phellogen nach außen abgeschieden werden, und manchmal auch c) dem Phelloderm, das vom Phellogen nach innen abgeschieden wird. Der Anfang der Peridermbildung ist die Anlage eines Phellogens. Ein Ring von Zellen im Stamm teilt sich durch Wände parallel zur Oberfläche des Organs. Je eine der beiden Tochterzellen erweist sich durch dünne Wände, Plasmareichtum und kleine Vakuolen als Phellogenzelle und schnürt nach außen Phellomzellen, nach innen Phellodermzellen ab. Das erste Phellogen entsteht in den meisten Fällen in der Subepidermis, der äußersten Schicht des Grundgewebes unter der Epidermis. Jedoch kann auch die Epidermis selbst oder aber eine tiefere Schicht des Grundgewebes Bildungsort dieses Phellogens sein. Die Phellogene der Folgeperiderme, die bei stärkerem Dickenwachstum angelegt werden, entstehen stets in tieferen Schichten des Rindengewebes oder im Bast, also im sekundären Stranggewebe. Das führt schließlich zur Bildung einer Borke (s.u.). Wie sind nun die verschiedenen Zellen des Periderms gebaut, und welche Funktion erfüllen sie? Phellogen: Das Phellogen, ein Lateralmeristem, besteht aus einer Schicht meristematischer Zellen, die im Querschnitt rechteckig sind. Die Zellen teilen sich überwiegend tangential, geben also neue Zellen nach außen und innen ab. Gelegentliche Teilungen durch senkrecht auf die Oberfläche stehende Wände vermehren die Phellogenzellen, sodaß sich das Meristem dem Um66 67 fangswachstum der Achse anpassen kann. Die Zahl der nach außen abgeschnürten Phellomzellen ist stets größer als die der nach innen gebildeten Phellodermzellen. Das Phelloderm kann auch überhaupt fehlen. Phellom: Wenn eine nach außen abgeschiedene Tochterzelle des Phellogens zur Korkzelle umgebildet wird, streckt sie sich in radialer Richtung, bleibt aber gleich breit. Dadurch liegen alle Korkzellen mit der Phellogenzelle, von der sie abstammen, streng in einer Reihe. Korkzellen stoßen lückenlos, ohne Interzellularen, aneinander. Die besondere Eignung als Zellen eines Abschlußgewebes erhalten die Korkzellen durch das Suberin in ihren Wänden. Diese Substanz wird in Form von Lamellen über der Primärwand abgelagert. Die Suberinlamellen selbst sind wieder von Wachsschichten durchzogen, die nur elektronenmikroskopisch sichtbar sind. Die Zelle verkorkt in der Weise, daß nach Abschluß des Streckungswachstums auf die zellulosische Primärwand Schichten von reinem Suberin und Wachs aufgelagert werden. Dabei bleiben zunächst Tüpfel ausgespart, durch die den Phellom-Protoplasten, die ja selbst keine Photosynthese betreiben, das Material für die Synthese von Suberin und Wachs geliefert wird. Zuletzt werden auch die Tüpfel mit Suberin ausgefüllt, worauf die Zellen absterben. Das Lumen füllt sich mit Luft. Suberin und Wachse können über 40% des Trockengewichtes der Wand ausmachen. Häufig werden Phlobaphene, Oxydationsprodukte der Gerbstoffe, eingelagert, die die Wände braun färben. Alle diese Substanzen verleihen den Wänden beträchtliche Resistenz gegen den Angriff von Mikroorganismen. Durch die Verkorkung zahlreicher hintereinander liegender Zellen wird die Transpiration noch wirksamer eingeschränkt als durch die Cutinisierung der Epidermis. Selbst dünne Korklagen setzen die Wasserabgabe wesentlich herab: Ein geschälter Erdapfel gibt in der Zeiteinheit 64 mal mehr Wasser ab als ein ungeschälter. Auch übermäßige Erwärmung durch Sonnenstrahlung verhindert der isolierende Kork wirksam. Alles in allem könnte man geschlossene Korkschichten gut mit dem Kunststoff Styropor vergleichen. Einige Worte zum technischen Kork. Dieser ist das Produkt der mediterranen Korkeiche, Quercus suber. Der junge Baum bildet "männlichen Kork", eine rissige, harte Borkenschicht wie bei den heimischen Eichenarten ohne wirtschaftlichen Wert. Erst nach dem Schälen dieser Schicht, im Alter von 67 68 10 bis 15 Jahren, entstehen aus einem neuen Phellogen mächtige und doch gleichmäßige Schichten weichen Korks. Dieser "weibliche Kork" wird in Abständen von etwa 9 Jahren wiederholt durch Schälen geerntet. Das Periderm schließt als Abschlußschicht den darunter liegenden Pflanzenkörper luft- und wasserdicht von der Außenwelt ab. Ein gewisser Gasaustausch ist aber weiterhin erforderlich: Die lebenden Zellen des Stammes müssen mit Sauerstoff versorgt werden, um ihren Energiebedarf aus der Atmung decken zu können. Oft enthalten sie auch Chloroplasten; dann benötigen sie auch CO2. In das Periderm sind daher Bereiche eingeschaltet, die durch lockere, interzellularenreiche Anordnung der Zellen Gasaustausch ermöglichen. Diese Bereiche werden als Korkporen oder Lentizellen (von lat. lenticella = kleine Linse) bezeichnet, da sie in der Aufsicht meist linsenförmig erscheinen. Lentizellen entstehen häufig unterhalb der Stomata des primären Hautgewebes. Ihr lockeres Füllgewebe wird vom Lentizellenmeristem produziert, das mit dem Phellogen allseitig zusammenhängt. Die Füllzellen sind verkorkt oder unverkorkt; für den Gasaustausch hat das keine Bedeutung, da die Luft an den Zellen vorbei durch die Interzellularen streicht. Dort, wo die Stämme ein langandauerndes Dickenwachstum zeigen, also bei Sträuchern und Bäumen, wird die erste Peridermschicht, die das primäre Hautgewebe ersetzt, meist mehr oder weniger bald zerrissen. Buche, Hainbuche und Hasel stellen als sogenannte „Peridermbäume" Ausnahmen dar. Bei ihnen bleibt das erste, die Epidermis ersetzende Korkgewebe dauernd tätig und produziert eine glatte Rinde, weil es der Umfangerweiterung folgen kann. Bei den übrigen heimischen Holzgewächsen kommt es zur Borkenbildung. Neue Periderme werden im Inneren der Stämme, zuerst im Rindengewebe, dann darunter im Bast, einem Teil des sekundären Stranggewebes, angelegt. Diese Periderme sind entweder uhrglasförmig und trennen relativ kleine Partien aus dem lebenden Bast heraus, oder sie sind ringförmig angeordnet und bilden sich innerhalb des geschlossenen Periderms aus, das sie in seiner Funktion ablösen sollen. Im ersten Fall finden wir eine Schuppenborke, etwa bei Kiefer, Lärche, Eiche und Esche. Im zweiten Fall liegt eine Ringelborke vor, die wir bei der Birke finden. Was umfaßt nun der Begriff Borke? Stets wird durch die Folgeperiderme lebendes Gewebe vom Zusammenhang mit den wasserleitenden Elementen des Stranggewebes 68 69 abgeschnürt. Die verkorkten Peridermzellen verhindern die Nachleitung von Wasser und Nährstoffen zu diesen Geweben, die daher bald absterben und vertrocknen. Die Gesamtheit aus Peridermen und dazwischen eingeschalteten, abgestorbenen Gewebekomplexen, die ältere Stämme bedeckt, nennt man Borke. Die ökologische Bedeutung des tertiären Hautgewebes, wie man die Borke auch nennt, geht über die mechanische Schutzwirkung und über die Vermeidung von Wasserverlusten weit hinaus. Einlagerungen von Gerbstoffen und Phlobaphenen verleihen den Borken Widerstandsfähigkeit gegen Schädlingsbefall; Borke isoliert gegen Hitze und plötzliche Kälte. Andererseits sind Borkenschäden für Bäume recht gefährlich, da dann Mikroorganismen in die lebenden Gewebe eindringen können. Darauf ist bei Bauarbeiten im Bereich der Bäume zu achten: Die Stämme sind durch Abdeckung zu schützen, entstandene Wunden müssen mit Baumwachs behandelt werden. Das Grundgewebe Als Grundgewebe werden alle Gewebe zwischen Hautgewebe und Stranggewebe zusammengefaßt. Sie entwickeln sich aus dem Grundmeristem der Determinationszone. Anatomisch und physiologisch handelt es sich um eine sehr heterogene Ansammlung von Geweben, die man am besten zunächst nach ihrem Bau weiter aufgliedert. Wir können unterscheiden: a) Parenchym b) Kollenchym c) Sklerenchym Wir wollen diese Systeme der Reihe nach besprechen. Das Parenchym Das Parenchym besteht aus lebenden, dünnwandigen Zellen von isodiametrischer oder mäßig gestreckter Gestalt. Solche Zellen werden als "parenchymatisch" bezeichnet, sie finden sich jedoch nicht nur im eigentlichen Parenchym, sondern auch im sekundären Hautgewebe als Phelloderm und im Stranggewebe als Leitparenchym. Das Parenchym des Grundgewebes ist der Hauptsitz wichtiger Leistungen der lebenden Protoplasten. Die Zellen sind verschieden stark spezialisiert, doch stets weniger als im Hautgewebe oder im Stranggewebe. Zellen mit so einzigartigen Funktionen wie Schließzellen oder Korkzellen gibt es im Parenchym nie. Daher kommt es gelegentlich zum 69 70 Aufgabenwechsel, etwa von der Assimilation zur Speicherung. Protoplasten unspezialisierter Zellen können sich in Sonderfällen teilen. Das erklärt etwa die Bildung von Phellogenen im Parenchym nach Verwundung, die dann Wundkork bilden. Bei Beginn des sekundären Dickenwachstums entstehen Lateralmeristeme (Phellogen, Teile des Kambiums) im Stamm und in der Wurzel durch Teilung parenchymatischer Zellen. Das Parenchym benötigt für seinen Stoffwechsel Energie, die aus Photosynthese und Atmung stammt. Ausreichende Versorgung mit Luft ist für diese Prozesse eine Hauptvoraussetzung. Die Gase treten durch Stomata oder Lentizellen in den Pflanzenkörper ein und werden in Interzellularräumen weitergeleitet. Luftführender Interzellularen können auf zwei Weisen entstehen, nämlich schizogen oder rhexigen. Die Zellen des Apikalmeristems stoßen lückenlos aneinander. Durch enzymatische Auflösung der Mittellamellen an einzelnen Stellen bei Beginn der Streckung entsteht eine kleine Lücke, die sich durch Streckung der angrenzenden Primärwände zur schizogenen Interzellulare vergrößert. Wenn sich die Zellen, die an eine solche Interzellulare anstoßen, mehrfach teilen und strecken, können sehr große Lufträume entstehen, wie sie für Wasser- und Sumpfpflanzen charakteristisch sind. Durch mechanisches Zerreißen von ganzen Zellen entstehen die rhexigenen Interzellularen. So folgen etwa die Markzellen im Zentrum von Stengeln dem Streckungswachstum der äußeren Gewebe nicht; sie zerreißen unter Beteiligung enzymatischer Abbauvorgänge in der Zellwand und es entsteht eine lufterfüllte Markhöhle. Diese ist für manche Familien charakteristisch, etwa für die Halme der Gräser oder die Stengel der Doldenblütler. Es gibt einige funktionelle Sondertypen des Parenchyms: a) Assimilationsparenchym: Dieses findet man in allen grünen Pflanzenteilen, besonders aber in Blättern. Charakteristisch sind seine großen Interzellularen. Wir haben schon bei der Besprechung der Diffusionsvorgänge gesagt, daß sie in Wasser etwa 10 000 mal langsamer als in Luft ablaufen. CO2 sollte also auf dem Gaswege möglichst nahe an die grünen Zellen herangeschafft werden; daraus erklärt sich der Bau dieses Gewebes. 70 71 b) Speicherparenchym: Reservestoffe werden oft in spezialisierten Organen, etwa unterirdischen Knollen oder Rüben, gespeichert. Samen speichern für die Erstversorgung der nächsten Generation. In allen diesen Fällen speichern große, zusammenhängende Komplexe aus typischen Parenchymzellen. Da die Atmung dieser Zellen nicht sehr intensiv ist, besitzen sie meist keine großen Interzellularsysteme. Wasserspeichernde Parenchyme finden sich in Blättern und Stämmen sukkulenter Gewächse, etwa der Kakteen. Holzgewächse speichern übrigens nicht im Grundgewebe, sondern in lebenden Zellen des sekundären Stranggewebes (Holzparenchymzellen und Markstrahlparenchymzellen). c) Ausscheidungsparenchym: Hier muß man zwischen intrazellulärer und extrazellulärer Ausscheidung unterscheiden. Intrazelluläre Ausscheidung ist die Ablagerung in Vakuole oder Zellwand der Zelle, die das Ausscheidungsprodukt gebildet hat. Beispiele sind Kalziumoxalat und Milchsaft in Vakuolen, Kieselsäure in den Zellwänden. Oft sind einzelne Zellen mit Speicherfunktion für Ausscheidungsprodukte in Parenchyme mit anderen Aufgaben eingebettet. Man nennt solche Zellen Idioblasten. Extrazelluläre Ausscheidung erfolgt aus Drüsenhaaren des Hautgewebes oder aus Hydathoden ins Freie, oft aber in Interzellularen. Häufig sind dies schizogene Interzellularen, die mit speziellen Grundgewebezellen ausgekleidet sind. Diese produzieren Exkrete oder Sekrete. Hierher gehören die Harzgänge im Grundgewebe der Nadeln vieler Koniferen; ähnliche Harzgänge besitzen diese im Stranggewebe des Stammes. Korbblütler, Doldenblütler und andere Familien haben schizogene, mit aromatisch riechenden Ausscheidungsstoffen gefüllte Interzellularen. Daneben gibt es eigene Interzellularen für die Ausscheidung, die lysigenen Interzellularen. Durch Auflösung von Zellwänden und Degeneration der Protoplasten entstehen große Hohlräume. Besonders oft tritt dieser Typ dort auf, wo ätherische Öle im Grundgewebe entstehen, etwa in der Schale von Citrus-Früchten. d) Durchlüftungsparenchym: Die Wurzeln von Sumpf- und Wasserpflanzen stecken in einem sauerstoffarmen Milieu, die Wurzel hat aber hohen Sauerstoffbedarf. Daher sind auf anaeroben Standorten im Stengel und in der Wurzel riesige Interzellularsysteme ausgebildet, die man Aerenchyme nennt. Sie stehen über die Spaltöffnungen der Blätter mit der Außenluft in Verbindung. 71 72 Die Wände der Parenchymzellen sind unverdickt. Bereits diese Zellen festigen den Pflanzenkörper: Sie werden durch den Turgordruck des Protoplasten straff gespannt, und ihre Zellwand ist in diesem Zustand sehr steif. Natürlich kann das nur bei lebenden Zellen funktionieren und auch nur dann, wenn die Zellen gut mit Wasser versorgt sind. Ist dies nicht der Fall, dann kommt es zu Welkeerscheinungen. Das Prinzip des Turgordruckes genügt also nicht einmal, um großen krautigen Pflanzen die nötige Festigkeit zu verleihen. Die Anforderungen sind ja hoch: Ein hohler Halm von Weizen oder Roggen hat bei einer Höhe über 1 m an seiner Basis weniger als 3 mm Durchmesser und muß dabei an seiner Spitze noch die Last der Ähre (rund 50% des Trockengewichtes!) tragen! Dazu ist hohe Festigkeit erforderlich. Daneben verfügt der Pflanzenkörper aber auch noch über eine beträchtliche Elastizität. Der lange Roggenhalm mit seiner schweren Ähre kann vom Wind vorübergehend gegen den Boden gedrückt werden, ohne zu knicken, und richtet sich danach mühelos wieder elastisch auf. Kollenchyme und Sklerenchyme Im Grundgewebe finden sich daher neben den Parenchymen auch Zellkomplexe, die mit Hilfe verdickter Zellwände festigend wirken. Die Zellen sind teils lebend, teils tot; die Verdickung kann gleichmäßig alle Wände erfassen oder sich auf einzelne Wände oder gar Wandteile beschränken. Und schließlich sind manche, aber nicht alle Wände lignifiziert. Wie soll man diese Vielfalt einteilen? Man hat sich für das einfachste Kriterium, nämlich das Erscheinungsbild im Mikroskop, entschieden und nennt Kollenchymzellen jene Zellen, deren Wände nur teilweise, nicht allseitig verdickt sind. Sie sind zu größeren Zellkomplexen, den Kollenchymen, zusammengefaßt, die an der Peripherie des Organes liegen, das sie stützen sollen. Meist sind sie lebend, meist langgestreckt, meist enthalten die Wände keine Lignin. Sklerenchymzellen haben allseitig gleichmäßig verdickte Wände, sind oft tot und infolge ihrer starken Verdickung ein sehr wirksames Element der Festigung. Sehr häufig werden die verdickten Zellwände durch Lignineinlagerung weiter versteift. Sklerenchymzellen bilden Sklerenchyme. Isodiametrische Sklerenchymzellen nennt man Steinzellen oder Sklereiden. Ihre Wände sind deutlich geschichtet, da das Wandmaterial rhythmisch abgelagert 72 73 wird. Sie finden sich in der Fruchtwand (dem Perikarp) von Nuß- und Steinfrüchten, sowie in Rinden und Borken. Sklerenchymfasern sind meist als Bündel in das Grundgewebe eingelassen. Sind diese Bündel zahlreich und wenig verholzt, so sind sie als Pflanzenfasern wirtschaftlich interessant (Blätter von Agave sisalana, Sisalagave, und Musa textilis, Manilahanf). Das Stranggewebe Das Stranggewebe dient hauptsächlich dem Stofftransport und der mechanischen Festigung der Pflanze, daneben auch der Speicherung. Es zieht sich als zusammenhängendes System von den dünnsten Wurzeln bis in die Blätter. Seine Entwicklung war einer der Hauptschritte bei der Evolution der höheren Landpflanzen, der Kormophyten, zu denen außer den Spermatophyten auch noch die Farnpflanzen oder Pteridophyten gehören. Stoffe werden in zwei getrennten Gewebeteilen geleitet, die auf den Ferntransport von Wasser und anorganischen Nährstoffen einerseits und auf den Ferntransport von Photosynthese-Produkten andererseits spezialisiert sind. Das Wasser mit den darin gelösten Nährstoffen wird von der Wurzel aufgenommen und im Xylem zu den oberirdischen Teilen der Pflanze befördert. Die bei der Photosynthese gebildeten Kohlenhydrate gelangen im Phloem von den Orten der Bereitstellung ("sources") zu den Orten der Entnahme ("sinks"). Besonders im Xylem, aber auch im Phloem finden sich langgestreckte Zellen mit verdickter Wand, die der Pflanze hohe mechanische Widerstandsfähigkeit gegenüber Zug, Biegung und Druck verleihen. Lebende Elemente des Stranggewebes können auch Reservestoffe speichern. Die Elemente des Stranggewebes sind im primären Zustand zu Gefäßbündeln vereinigt, die im Grundgewebe eingebettet sind. Jedes Gefäßbündel enthält Xylem und Phloem, die selbst wieder aus mehreren verschiedenen Zelltypen mit spezialisierten Aufgaben bestehen. a) Die Elemente des Xylems: Als hochspezialisierte Elemente der Wasserleitung dienen Tracheiden und Tracheen. Beide sind im funktionsfähigen Zustand tot und bestehen nur mehr aus den verdickten, stark verholzten Wänden. Tracheiden sind einzelne Zellen, die mit steilen, getüpfelten Schrägwänden aneinander grenzen; 73 74 Tracheen (Gefäße) sind dagegen Längsreihen tonnenförmiger Einzelzellen, der Gefäßglieder, die durch teilweise oder völlige Auflösung ihrer Querwände zu geschlossenen Röhren von oft beträchtlicher Länge vereinigt sind. Sie kommen nur bei Angiospermen vor. Funktionsfähige Tracheiden und Tracheen sind mit Wasser gefüllt, das auch Bodennährsalze enthält. Im Frühjahr werden auch organische Stoffe (Zucker), die aus der Mobilisierung von Reservestoffen stammen, vom Xylemwasser der Bäume zu den austreibenden Knospen und den jungen Trieben transportiert. Die Wände der Wasserleitungselemente besitzen verschieden ausgebildete, verholzte Verdickungen, die dem in Tracheen und Tracheiden bei der Transpiration der Pflanze entstehenden Unterdruck oder Zug von vielen bar standhalten. Was passiert, wenn man einen dünnwandigen Schlauch oder einen Plastiktrinkhalm zuhält und die Luft ansaugt? Halm oder Schlauch kollabieren, da der außen herrschende Überdruck die Wandung eindellt. Nun ist die Druckdifferenz zwischen Außen- und Innenseite bei einem solchen Experiment maximal 1 bar (Atmosphärendruck gegen Vakuum). Die Druckdifferenz zwischen der Xylemflüssigkeit und der Atmosphäre kann hingegen auch bei heimischen Pflanzen 10, 20 oder 30 bar betragen. Entsprechend druckresistent muß die Wandung der wasserleitenden Elemente gebaut sein. Das wird durch Verdickung und Verholzung erreicht. Bei krautigen Hygrophyten sind häufig nur kleine Bereiche der Wände verdickt; man spricht dann von Ring-, Schrauben- und Netzgefäßen oder tracheiden. Auf trockenen Standorten und generell bei Holzgewächsen findet sich überwiegend eine massive Form der Verdickung, die nur den Bereich der Tüpfel freiläßt. Es entstehen die Tüpfelgefäße und Tüpfeltracheiden. Das Xylemparenchym besteht aus lebenden und verholzten parenchymatischen Zellen. Es dient der Speicherung von Reservestoffen, wie Stärke und Fett, vermittelt aber auch die Querleitung des Wassers und der Nährsalze aus dem Bündel ins Grundgewebe. Als mechanische Festigungselemente dienen im Xylem die Libriformfasern. Es sind dies prosenchymatische, verholzte und dickwandige Zellen, die den Sklerenchymfasern ähneln. Sie finden sich nur bei Angiospermen. 74 75 b) Die Elemente des Phloems: Assimilate werden in wäßriger Lösung von den in Längsreihen angeordneten Siebröhrengliedern, den Elementen der Siebröhren, transportiert. Ihre Zellen enthalten Protoplasten mit Mitochondrien und stärkehaltigen Plastiden. Zellkern und Tonoplast werden in den hochentwickelten Siebröhren der Angiospermen frühzeitig aufgelöst; das Plasma erfüllt das gesamte Lumen der Zelle als aufgelockertes Maschenwerk in innigem Kontakt mit dem Zellsaft. Die Siebröhren sind stets unverholzt. Ihren Namen verdanken sie lokalen, siebartigen Durchbrechungen ihrer Querwände und ihrer Längswände. Die stark perforierten Querwände der Siebröhrenglieder bezeichnet man als Siebplatten, ähnliche, aber kleinere Abschnitte, die hier und da in den Längswänden auftreten, als Siebfelder. Dicke Plasmastränge durchziehen diese Wanddurchbrechungen und verbinden die Protoplasten. Man kann sich vorstellen, daß die innige Verbindung der Protoplasten im System der Siebröhren beim Stofftransport nützlich ist, man hat aber noch keine völlig gesicherte Vorstellung darüber, wie das Wasser und die darin gelösten Stoffe (meist Saccharose, in einigen Fällen auch Oligosaccharide, besonders Raffinose, und dazu eine Anzahl weiterer Substanzen, wie etwa Pflanzenhormone, Kaliumionen, Aminosäuren und sogar ATP), transportiert werden. Die meisten Autoren gehen heute von osmotischen Prozessen aus: Wenn das Röhrensystem an einer Stelle (der Source) mit gelösten Stoffen beladen wird, dann erhöht sich dort durch Ansaugung von Wasser der innere Druck. Die Druckwelle pflanzt sich fort und mit ihr werden Stoffe zu den Orten geringerer Konzentration (den Sinks) transportiert (Münch´sche Druckstromhypothese). Die Lebensdauer der Siebröhren ist in der Regel auf eine einzige Vegetationsperiode beschränkt. Dann degenerieren sie und müssen durch neue ersetzt werden. Bei den Angiospermen gehen die Siebröhren durch inäquale Teilung aus Siebröhrenmutterzellen hervor. Aus der größeren Zelle entsteht das Siebröhrenglied, aus der kleineren, die sich vorher auch noch quer teilen kann, entsteht schließlich die Geleitzelle. Geleitzellen sind viel englumiger als die Siebröhren, sehr plasmareich, haben große Zellkerne und stehen mit den Siebröhren durch zahlreiche Plasmodesmen in enger plasmatischer 75 76 Verbindung. Ihre polyploiden Kerne steuern den Stoffwechsel der kernlosen Siebröhrenglieder, wie man annimmt. Sie liefern also RNA oder Proteine, die in das Protoplasma des Siebröhrengliedes übertreten. - Die kernlosen Siebröhrenglieder und ihre Schwesterzellen, die Geleitzellen, finden sich nur bei den Angiospermen. Die Gymnospermen haben kernhaltige Siebzellen, die auch im Hinblick auf die Eiweißsynthese autark sein dürften. Auch im Phloem treten parenchymatische Zellen mit Speicher- und Leitfunktion auf, die größer als die Geleitzellen sind und als Phloemparenchym bezeichnet werden. Sklerenchymatische Elemente des Phloems, die ebenso wie die Libriformfasern des Xylems der mechanischen Festigung dienen, sind die Phloemfasern. Das sind prosenchymatische, an den Enden zugespitzte Zelen, die meist nur ein enges Lumen besitzen. Ihre Zellwände sind unterschiedlich verholzt: stärker bei Jute, schwächer bei Hanf, gar nicht bei Lein, um drei kommerziell genutzte Phloemfasern zu nennen. In der Regel sind sie tot, gelegentlich, so etwa beim Wein, kommen auch lebende Faserzellen vor. Phloemfasern sind bei geringem Durchmesser ungewöhnlich lang, im Durchschnitt 1 -2 mm, bei Lein 0.4 - 6,5 cm, bei der Brennessel bis 7,5 cm. Wie bereits erwähnt, halten den Längenrekord unter allen bekannten Pflanzenzellen die Phloemfasern der Gattung Boehmeria (Ramie), die bis 55 cm lang werden können. Ihre Länge erreichen die Fasern durch ein lange andauerndes Wachstum, bei dem ihre Spitzen sich auch zwischen andere Zellen schieben können, deren Mittellamellen aufgelöst werden ("gleitendes Wachstum"). Die Fasern liegen häufig als kompakter Strang an der Grenze vom Phloem zum Grundgewebe und können daher oft auch technisch genutzt werden. Zusammenfassend eine Gedächtnisstütze: Im voll entwickelten Zustand sind alle Elemente des Xylems mit Ausnahme der Xylemparenchymzellen tot, hingegen alle Elemente des Phloems mit Ausnahme der meisten Phloemfasern lebend. Die Gefäßbündel Wir kennen also jetzt die Elemente des Stranggewebes. Xylem und Phloem treten stets in unmittelbarer Nachbarschaft auf, bleiben aber scharf voneinander getrennt. Vor Einsetzen des sekundären Dickenwachstums lassen 76 77 sich im Sproß und der Wurzel Stränge von Leitgewebe, die Leitbündel oder Gefäßbündel, unterscheiden. Sie haben kreisförmigen oder elliptischen Querschnitt und verlaufen in Richtung der Längsachse der Organe, sind aber durch Querverbindungen zu einem kommunizierenden System verbunden. Abzweigungen reichen in Seitenäste und Blätter. Hier vollzieht sich der gesamte Ferntransport von Wasser und Assimilaten. Man teilt die Gefäßbündel nach den Lagebeziehungen zwischen Xylem und Phloem ein. a) Das kollaterale Gefäßbündel ist für die Stämme und Blätter der Gymnospermen und Angiospermen charakteristisch. Das Phloem ist nach aussen, das Xylem nach der Mitte des Stengels gerichtet. In den geschlossenen kollateralen Bündeln der Monokotylen grenzen Xylem und Phloem direkt aneinander, in den offenen kollateralen Bündeln der Gymnospermen und Dicotylen sind sie dagegen durch einen meristematischen Gewebestreifen, das faszikuläre Kambium, getrennt. b) Im konzentrischen Gefäßbündel ist ein Xylem- oder Phloemstrang von einem mantelförmigen Phloem- oder Xylemstrang umgeben. Liegt das Xylem im Bündel innen, so ist dieses hadrozentrisch, und nur dieser Typ ist wichtig, da er nämlich bei den meisten Farnen vorkommt. c) Das radiäre Gefäßbündel ist typisch für Wurzeln. Zum Unterschied von den oft sehr zahlreichen Bündeln im Stamm gibt es stets nur ein radiäres Bündel pro Wurzel. Dafür hat es aber mehrere getrennte Xylem- und Phloemstränge. Sie sind im Querschnitt wie die Speichen eines Rades und miteinander abwechselnd angeordnet. Die Xylemteile stoßen im Zentrum meist aneinander, das Xylem bildet daher eine sternförmige Figur. In den Buchten zwischen den Xylemsträngen liegt das Phloem, das vom Xylem durch Leitparenchymschichten getrennt ist. Die Entwicklung dieses Bündeltyps erfolgt zentripetal, von außen nach innen; am weitesten außen liegen daher die kleinen ersten Elemente, die Primanen, weiter innen die später ausdifferenzierten Folgeelemente von größerem Durchmesser. Zum Bündel gehört hier noch ein Meristemring aus teilungsfähigen Zellen, der Perizykel. Den Bau des sekundären Stranggewebes, das keine neuen Zelltypen enthält, wollen wir zusammen mit seiner Bildung etwas später besprechen. ORGANOGRAPHIE 77 78 Wir kommen zur Organographie des Pflanzenkörpers, der aus drei typischen Organen besteht: Stamm und Blatt sind enger verwandt, da sie auf ein gemeinsames Apikalmeristem zurückgehen. Sie werden als Spross zusammengefaßt. Dagegen ist die Wurzel als Produkt eines zweiten, anders strukturierten Apikalmeristems deutlich eigenständig. Die Grundform der drei Organe des Pflanzenkörpers und ihre übliche Funktion werden oft stark abgewandelt. Wir sehen also viele Funktionen und noch viel mehr Formen; mit der Gliederung dieser Vielfalt beschäftigt sich die Morphologie im engeren Sinn. Wir wollen zunächst die typischen Baueigentümlichkeiten der drei Organe untersuchen und so den „idealen Bau“ des ganzen Pflanzenkörpers erarbeiten. Sonderbildungen umfassen meist den ganzen Körper, also mehrere Organe, und sie sind oft Anpassungen an bestimmte Umweltansprüche. Der Samen Zunächst werden kurz Bau und Keimung des Samens besprochen, da ein junger Spermatophyt sich stets aus einem Samen entwickelt. Der Samen entsteht aus der Samenanlage; diese enthält eine Eizelle, die nach ihrer Befruchtung durch die männlichen Geschlechtszellen den vielzelligen Embryo aufbaut. Der Embryo braucht Schutz, vor allem gegen Fraßfeinde. Meist ist also die Diaspore (die Verbreitungseinheit) von einer widerstandsfähigen Hülle umgeben. Manchmal ist diese Hülle, die oft Sklereiden enthält, ein Teil der Frucht (etwa bei der Walnuß oder der Maroni), manchmal die Samenschale, die Testa. Im Inneren ruht der chlorophyllfreie, bleiche Embryo, der bei der Keimung auf Reservestoffe angewiesen ist. Diese werden von der Mutterpflanze mitgegeben. Entweder ist dafür ein spezielles Nährgewebe (Endosperm) ausgebildet, in das der Embryo eingebettet liegt, oder er speichert selbst, und zwar in den Keimblättern oder Cotyledonen. Nährgewebe enthalten zum Beispiel die Samen der Gräser, also auch der Getreidearten. Nährgewebelose Samen mit Speicherkotyledonen haben unter anderem die Stein- und Kernobstarten, die Hülsenfrüchtler wie Erbse und Bohne, die Buchen und Eichen, der Kürbis oder die Haselnuß. Die Keimblätter im Samen sind dann sehr verdickt und kaum mehr als Blattorgane zu erkennen. 78 79 Die Achse des Embryos besteht aus der Keimwurzel oder Radicula und dem Hypokotyl, die am Wurzelhals ineinander übergehen. Das Hypokotyl trägt die Keimblätter oder Cotyledonen, deren Zahl für die beiden Klassen der Angiospermen (Ein- und Zweikeimblättrige) charakteristisch ist. Nadelhölzer haben verschieden viele Keimblätter je nach Art. Alle Zellen des Embryos im Samen sind noch teilungsfähig. Nach der Fertigstellung geht diese junge Pflanze in einen Ruhezustand über, wobei der Samen meist stark austrocknet. Es folgt die Trennung der Diaspore von der Mutterpflanze und die Verbreitung durch Wind, Wasser oder Tiere. Nach der Verbreitung und der Einbettung in den Boden keimen viele Samen auch unter günstigen Bedingungen nicht sofort. Das gilt besonders für Samen, die spät im Jahr reifen. Diese Keimruhe kann verschiedene Ursachen haben, von noch nicht abgeschlossenen Wachstumsprozessen bis zu chemischen Hemmstoffen oder einer wasserundurchlässigen Testa. Solche Samen müssen eine Nachreife durchmachen, um keimfähig zu werden. In gemäßigten Breiten sind oft niedrige Temperaturen zur Auslösung der Nachreife erforderlich. Es können also viele Samen Anfang November auch bei Temperaturen um 10°C nicht keimen; dadurch wird die empfindliche Jungpflanze vor den niedrigen Wintertemperaturen geschützt. Der Samen keimt im Frühjahr, wenn nicht ein "Überliegen" erfolgt und er ein oder mehrere weitere Jahre in Ruhe bleibt. Die Keimung hat Voraussetzungen: 1) Wasser muß in genügender Menge und in günstigem Zustand vorhanden sein. 2) Die Sauerstoffversorgung muß gewährleistet sein. 3) Die Temperatur muß geeignet sein. Der Bereich ist an sich sehr groß (0 45°C), doch haben viele Samen ein Optimum bei 25 - 30°C. Die Keimung eines Samens wird durch die Aufnahme von Wasser ein- geleitet. Die trockenen Zellen im Samen vergrößern sich durch Quellung und sprengen die Testa. Das Zellmaterial wird durch Teilung vermehrt und durch Streckung vergrößert. Dabei ist zunächst das Wurzelwachstum gefördert. Die Primärwurzel tritt aus, verankert den Keimling im Boden und versorgt den wachsenden Embryo mit Wasser und Mineralstoffen. Substanzen und Energie für das Wachstum werden durch Umbau und Abbau der Reservestoffe gewonnen. Zuletzt streckt sich der Stamm. Die Blätter entfalten sich und treten aus der 79 80 Samenschale aus, die als leere Hülle abgeworfen wird. In manchen Fällen streckt sich das Hypokotyl nicht. Dann können auch die Cotyledonen in der Samenschale zurückbleiben, da diese Reservestoffbehälter nach ihrer Entleerung nutzlos sind. Man bezeichnet das Vorkommen eines gestauchten Hypokotyls als hypogäische Keimung ("Keimung unter der Erde", da ja der Same in vielen Fällen von einer dünnen Erdschicht bedeckt ist). In anderen Fällen, vor allem bei Samen mit Endosperm, werden die Keimblätter über die Erde erhoben. Sie werden grün und dienen als erste photosynthetisch tätige Organe der Pflanze. Dieses Verhalten bezeichnet man als epigäische Keimung ("Keimung über der Erde"). Wir sehen uns zunächst die weitere Entwicklung des Sprosses an. Ganz zu Anfang der Keimung sind noch alle Zellen des Keimlings teilungsfähig, doch bald werden allein von den Apikalmeristemen neue Zellen gebildet. Den Sprossvegetationspunkt nennt man seiner Gestalt wegen auch Vegetationskegel. Er legt an den Flanken unmittelbar unter seiner Spitze die Blätter an. Die sich streckenden jungen Blätter überragen und umhüllen das Apikalmeristem und schützen so das empfindliche Gewebe. Ein solches Gebilde nennt man Knospe. An jenen Stellen, die später in den Achseln der Blätter liegen, bleiben Zellen meristematisch und bilden ihrerseits einen Vegetationskegel und in weiterer Folge eine Knospe aus. Dann ruhen sie. Diese Knospen in den Achseln der Blätter bilden später die Seitensprosse, während der erste Vegetationskegel oft den Hauptspross aufbaut. Meist ist die so angelegte Hauptachse negativ gravitrop, sie wächst vom Erdmittelpunkt weg. Stellen der Blattabzweigung nennt man Knoten oder Nodien (Einzahl: Nodium). Sie unterscheiden sich anatomisch von den Zwischenknotenstücken oder Internodien. Die Länge der Internodien ist je nach dem Wuchstyp der Art verschieden. Bei vielen Kräutern sind die Blätter in einer grundständigen Rosette angeordnet, da die Hauptachse gestaucht bleibt und die Internodien sich kaum strecken. Oft ist die Rosette nur für das vegetative Wachstum charakteristisch. Sobald die Pflanze die Blühreife erlangt, verlängern sich die Internodien an den neugebildeten Sprossabschnitten, und es bildet sich ein Stengel, der mit einem endständigen Blütenstand abschließt. Dieser Stengel kann beblättert sein, wie beim Hirtentäschel, der Zuckerrübe oder dem Radieschen. Andere Rosettenpflanzen, wie etwa Primel, 80 81 Hungerblümchen, Erdbeere, Löwenzahn oder Wegerich, treiben Stengel ohne grüne Blätter, die von einem Blütenstand abgeschlossen werden. Blüten an gestreckten Stengeln sind wohl für Bestäuber leichter zu finden. Bei den meisten Pflanzen strecken sich aber alle Internodien beträchtlich. Daran ist auch die Neubildung von Zellen in Interkalarmeristemen beteiligt, die meist an der Basis der jungen Internodien knapp über dem Knoten sitzen. Pflanzen mit gestreckten Stengeln zeigen die Regeln der Blattstellung besser als Rosettenpflanzen. An jedem Knoten können ein, zwei oder mehrere Blätter stehen. Mehrere Blätter pro Knoten bezeichnet man als quirlständige Blätter. Zwei Blätter pro Knoten sind gegenständig; sie stehen einander immer diametral gegenüber. Dabei können die Blätter an aufeinanderfolgenden Knoten in rechtem Winkel zueinander stehen, was man als dekussierte Blattstellung bezeichnet, oder sie können übereinander stehen, wodurch sie zwei parallele Reihen bilden; das ist die distiche Anordnung. Wenn sich nur ein Blatt an jedem Knoten entwickelt (wechselständig), bilden sich entweder zwei gegenüberstehende Reihen, die Blätter sind also wieder distich, oder sie liegen schraubig auf einer Linie, die sich um die Achse herumzieht. Der Winkel zwischen zwei aufeinanderfolgenden Blattansatzstellen ist dann kleiner als 180 °C; er ist genetisch bedingt und sehr konstant. Allerdings kann einseitiger Lichteinfall die Stellung der Blattspreiten recht unterschiedlich machen. Der Blattstiel kann nämlich durch Wachstumsvorgänge die Spreite in eine optimale Stellung für die Photosynthese bringen. Die Blattstellung ist schon auf dem Vegetationskegel festgelegt. Um jede junge Blattanlage entsteht eine physiologische Sperrzone, die die Entstehung konkurrierender Anlagen unterdrückt. Die junge Anlage wirkt aber auch in die Tiefe des Stammgewebes und sorgt dafür, daß sie mit Leitgewebe versorgt wird, wofür eine besondere Ausbildung der Blattknoten nötig ist. Diese Entwicklungssteuerung beruht auf der Abgabe von Hormonen, die um die Anlage einen Konzentrationsgradienten bilden. Wir wollen jetzt den inneren Bau des Stammes betrachten, und zwar zunächst den Zustand vor dem Beginn der Tätigkeit von Lateralmeristemen. Dieser primäre Zustand bleibt bei allen Einkeimblättrigen und Farnen, aber auch bei zarten, einjährigen Kräutern unter den Dikotylen zeitlebens erhalten. 81 82 Vom meristematischen Vegetationspunkt der Sprossachse werden basalwärts Zellen abgegliedert, die in der darunterliegenden Determinationszone entsprechend ihren Aufgaben in einen peripheren Mantel aus künftigem Abschlußgewebe (Protoderm), eine Lage Grundmeristem der Rinde und einen zentralen Strang Grundmeristem des künftigen Markgewebes gesondert werden. Zwischen diesen bleibt bei den Dicotylen und Gymnospermen ein schmaler Zylindermantel erhalten, das Prokambium. Es spaltet sich später häufig in isolierte Einzelbündel auf, die aber mehr oder minder exakt auf einem Zylindermantel liegen bleiben. - Anders ist es bei den Monokotylen, wo kleinere meristematische Prokambiumstränge undifferenziert zwischen den späteren Grundgewebsbereichen erhalten bleiben. Nach der ersten Sonderung folgen die Zonen der Streckung und der Differenzierung, wo die Zellen des Grundmeristems zu parenchymatischen Dauerzellen werden. Bei Dikotylen und Gymnospermen nennt man die Bereiche außerhalb des Gefäßbündelringes das Rindenparenchym, die zentralen Bereiche das Mark. Aus der äußersten Schicht, dem Protoderm, entsteht die Epidermis. Gleichzeitig werden im Prokambium, dessen Zellen sich in die Länge strecken, Leitelemente angelegt. Zwischen den einzelnen Prokambiumsträngen entwickeln sich bei Dikotylen und Gymnospermen Streifen parenchymatischen Gewebes, die Markstrahlen. Mit zunehmender Entfernung vom Vegetationspunkt schreitet die Differenzierung der Prokambiumstränge fort, wobei auf der Innenseite Wasserleitungselemente, auf der Außenseite Siebröhren gebildet werden, deren Gesamtheit man Protoxylem und Protophloem nennt. Diese Elemente sind nur kurz in Funktion und werden beim weiteren Streckungswachstum zerrissen. Ihre Aufgabe wird dann von neugebildeten, längeren und breiteren Elementen in den Leitbündeln übernommen, deren Gesamtheit man als Metaxylem bzw. Metaphloem bezeichnet. In den Internodien der Gymnospermen und der Dikotylen sind die offenen kollateralen Bündel auf einem Zylindermantel angeordnet. Die Bündel können durch breite primäre Markstrahlen getrennt sein oder zu einem fast geschlossenen Zylinder zusammentreten. Der Bündelring umschließt das Mark, dessen Zellen oft Reservestoffe speichern. Gelegentlich bildet sich aber auch eine rhexigene Markhöhle. Außen ist der Bündelring oft von einem Ring skler82 83 enchymatischer Zellen umgeben. Alle diese Elemente zusammen bilden den Zentralzylinder. Um den Zentralzylinder liegt die Rinde, deren Parenchymzellen Photosynthese und Speicherung betreiben. Die äusserste Schicht des primären Stammes ist die Epidermis. Darunter finden sich im primären Stamm häufig Lagen von Kollenchymen und Sklerenchymen. Zusammen mit den verholzten Elementen des Leitgewebes (Tracheen, Tracheiden, Libriformfasern und Phloemfasern) verleihen sie dem Stamm Festigkeit. Anatomisch unterscheiden sich die Knoten der Dikotylen beträchtlich von den Internodien. Hier biegen die Bündelverzweigungen als sogenannte Blattspuren in die Blätter aus, ebenso Zweigspuren in die Seitenachsen, die ja in den Achseln von Blättern entstehen. Das Xylem, das im Stamm innen liegt, befindet sich im Blatt auf der ursprünglich dem Stamm zugekehrten Oberseite. Häufig sind die in den Internodien längs verlaufenden Bündel in den Knoten horizontal verbunden. Dadurch können Beschädigungen einzelner Abschnitte des Leitgewebes immer wieder umgangen werden. Der Stamm der Monokotylen enthält viele geschlossene kollaterale Bündel, die bei kompaktem Stengel über den gesamten Querschnitt verteilt sind. In Stengeln mit Markhöhle, wie etwa bei Gräsern, können Bündel natürlich nur am Rand liegen, bilden dort aber selten nur einen Kreis. Der Bündelverlauf durch den monokotylen Stamm ist höchst kompliziert. Während bei den Dikotylen von einem Netzwerk "stammeigener" Bündel zu den Blättern kurze Blattspurstränge abzweigen, verlaufen bei den Monokotylen die Bündel meist über lange Strecken im Stamm, werden aber immer wieder von Blattbündeln angezapft, die oft über mehrere Knoten parallel verlaufen und erst dann in die Blätter ausbiegen. Der Bau des Leitsystems ist also mindestens so kompliziert wie bei den Dikotylen, doch erfordert seine Analyse raffinierte Techniken. Der Eindruck, daß die Bündel regellos über den Stammquerschnitt verstreut seien, wie er sich bei der Betrachtung einzelner Querschnitte ergibt, ist jedenfalls falsch. Da die Gefäßbündel der Monokotylen kein Kambium enthalten, fehlt ihnen das Lateralmeristem für ein sekundäres Dickenwachstum. Monokotyle bleiben daher meist krautig. Dennoch können manche Arten dieser Unterklasse baumförmige Dimensionen erreichen. Das bekannteste Beispiel sind wohl die Palmen. Deren Stämme enthalten einzelne Leitbündel in einem verholzten 83 84 Parenchym. Sie bleiben nach der Keimung viele Jahre gestaucht; alle Assimilate werden zur Vergrößerung des Spitzenmeristems durch Teilung der Zellen verwendet. Wenn schließlich das Längenwachstum einsetzt, genügen Streckung und Erstarkung der Zellen, die von dem großen Vegetationskegel abgegeben werden, um einen mächtigen, säulenförmigen Stamm aufzubauen. Zum Unterschied vom Stamm dikotyler Bäume ist der Palmenstamm an der Basis nicht breiter als in der Krone; im Gegenteil, durch weiteres Wachstum des Vegetationskegels kann manchmal gegen die Spitze hin eine mäßige Verbreiterung eintreten. Dikotyle und Gymnospermen haben zeitlebens kleine Vegetationskegel, die nur wenig Zellmaterial abgeben. Im Verlauf des Differenzierungswachstums entstehen daraus die primären Gewebe. Wenn die Apikalmeristeme der Hauptachse und der Seitenachsen neue Zellen abgeben, werden jedoch die bereits fertigen Teile des Stammes immer stärker belastet: Leitelemente an der Stammbasis müssen mehr Wasser heranschaffen, mechanische Elemente müssen ein stets wachsendes Gewicht tragen. Wenn das primäre Wachstum begrenzt ist, dann reichen die direkt vom Apikalmeristem abstammenden Leitund Festigungsgewebe für diese Aufgabe aus. Hierher gehören aber nur relativ wenige einjährige dikotyle Kräuter. Bereits mehrjährige Stauden, noch mehr aber die hochwüchsigen Bäume, benötigen zusätzliche Leit- und Stützgewebe. Sie werden nicht mehr vom Apikalmeristem gebildet, sondern von einem Lateralmeristem, dem Kambium. Dieses beginnt in einiger Entfernung vom Apikalmeristem mit der Bildung sekundärer Dauergewebe, die den primären hinzugefügt werden. Das Kambium Das Kambium ist ein Lateralmeristem (von mehreren). Es liegt also parallel zur Außenseite des Stammes und gibt (wie das Phellogen) neue Zellen durch tangentiale Teilungen nach innen und nach außen ab. Das Kambium ist schon im primären Stamm der Dikotylen und Gymnospermen vorhanden, jedoch nur teilweise: Das Bündel- oder Faszikularkambium trennt in den offenen kollateralen Leitbündeln das Xylem vom Phloem. Es ist dies jener Rest des Prokambiumstranges, der bei der Bündeldifferenzierung nicht zu Xylem und Phloem umgewandelt wurde, sondern meristematisch blieb. 84 85 Die Bündel liegen im Stamm der Dikotylen und der Gymnospermen auf einem Zylindermantel. Zwischen den Bündeln laufen Parenchymstreifen, die primären Markstrahlen, die Rinde und Mark verbinden. Das sekundäre Dickenwachstum wird damit eingeleitet, daß Markstrahlzellen direkt neben dem Bündelkambium sich erneut teilen. Durch weitere Teilungen auch in der Mitte des Markstrahls entstehen Streifen von Interfaszikularkambium oder Zwischenbündelkambium, die zusammen mit den Bündelkambien einen geschlossenen Kambiumring bilden. Durch zeitlich abgestimmte Teilungen dieses Ringes vergrößert sich der Umfang der Achse gleichmäßig. Bei Arten mit einem im primären Zustand ringförmig angelegten Stranggewebe bildet das Faszikularkambium bereits einen geschlossenen Ring, der die schmalen primären Markstrahlen quert, so dass kein neues Interfaszikularkambium gebildet werden muss. Die Kambiumzellen sind typische Meristemzellen, treten aber in zwei unterschiedlichen Formen auf: Die Fusiforminitialen sind in der Längsrichtung des Stammes gestreckt, die kleineren Markstrahlinitialen isodiametrisch. Das Kambium hat meist keine Interzellularen, daher ist der Gasaustasch des Stamminneren erschwert und nur dort rasch, wo im Kambiummantel Lücken sind, etwa bei der Abzweigung von Seitensprossen. Ein aktives Kambium gibt nach innen und außen Zellen ab, die sich langsam zu Holz- oder Bastelementen ausdifferenzieren. Im Querschnitt sieht man daher meist nicht nur eine, sondern mehrere Lagen dünnwandiger Zellen in radialen Reihen hintereinander liegen. Die nach innen gerichteten Teilungen schieben das Kambium nach außen: Die Zellen des Holzes bleiben an ihrem Bildungsort, während die äußere der beiden Tochterzellen meristematisch bleibt, nach einiger Zeit wieder ihr ursprüngliches Volumen erreicht und sich erneut teilt. Es werden weit mehr Zellen nach innen als nach außen abgegeben, sodaß das Holz viel zellenreicher als der Bast ist. Die Steuerung der Produktion erfolgt durch Hormone: Verschiebt sich das Gleichgewicht zwischen Auxin und Gibberellin zum Auxin, dann bildet die Kambiumzelle ein Holzelement; nimmt die Gibberellin-Konzentration relativ zu, dann wird eine Bastzelle abgeschnürt. Da sich der Holzkörper verdickt, muss auch der Kambiumring an seinem Außenrand erweitert werden. Das geschieht durch tangentiales Wachstum der 85 86 Kambiumzellen und durch gelegentliche radiale Teilungen. Danach bleiben beide Tochterzellen im Kambiumring. Alle Gewebe außerhalb des Kambiums kommen durch die Umfangserweiterung unter Spannung. Wir haben bereits die Konsequenzen gesehen: Erst ersetzt ein Oberflächenperiderm die Epidermis, dann kommt auch dieses unter Spannung und wird durch Tiefenperiderme abgelöst. Sie schnüren das Rindengewebe und die älteren, außenliegenden Bastschichten ab und ergeben mit diesen zusammen eine tote, meist stark mit Gerbstoffen imprägnierte Schutzschicht, die Borke. Die Produkte des Kambiums Wir wollen jetzt sehen, was das Kambium eigentlich produziert. Hier sind uns zwei klare Definitionen hilfreich, die wir zunächst zur Kenntnis nehmen und erst später durchdenken wollen. Definition I: Alles vom Kambium beim sekundären Dickenwachstums nach innen abgeschiedene Gewebe heißt Holz, und zwar unabhängig vom Grade der Verholzung, also vom Ausmaß der Lignineinlagerung. Definition II: Alles vom Kambium nach außen abgeschiedene Gewebe heißt Bast. Man liest für Holz gelegentlich "sekundäres Xylem" und für Bast "sekundäres Phloem". Beides ist nicht ganz korrekt, wenngleich im Englischen üblich. In beiden Fällen kommen nämlich zu den primären Elementen des Xylems und des Phloems noch ganz neue Elemente, die Markstrahlen, hinzu. Wir finden also im Holz folgende Bestandteile: a) bei Gymnospermen: Tracheiden, Holzparenchym1, Markstrahlparenchym 1 b) bei Dikotylen: Tracheiden, Tracheen, Libriformfasern oder Holzfasern , 1 Holzparenchym , Markstrahlparenchym Auch im Bast kommen zu den Bestandteilen des Phloems noch die Markstrahlparenchymzellen hinzu. Wir finden also bei: 1 a) Gymnospermen: Siebzellen (mit Kern, ohne Geleitzellen) Bastfasern , 1 Bastparenchym , Markstrahlparenchym 86 87 1 1 b) Dikotylen: Siebröhren, Geleitzellen, Bastfasern , Bastparenchym , Markstrahlparenchym (1 Namenswechsel gegenüber den Elementen des primären Zustandes: Ersatz von "Xylem" durch “Holz", und von "Phloem" durch "Bast", am Bau ändert sich nichts!) Alle Elemente, die sich schon im primären Bündel finden (= alle außer den Markstrahlen), sind in achsialer Richtung gestreckt. Sie sind im sekundären Zustand Produkte der Fusiforminitialen. Nach der Abgliederung vom Kambium können sie entweder die Länge der meristematischen Fusiforminitialen beibehalten: Das tun die Tracheenglieder und die Siebröhrenglieder. Sie können sich verlängern und durch "gleitendes Wachstum" zwischen andere Zellen eindringen; so verhalten sich Fasern, teilweise auch Tracheiden. Und schließlich können sie sich ein- oder mehrmals teilen und kleinere Zellen liefern; dann entstehen Bastparenchym und Holzparenchym. Die Funktion aller dieser Zellen gleicht der ihrer Vorläufer in den primären Bündeln. Etwas Neues ist das Markstrahlparenchym, das Produkt der Markstrahlinitialen. Seine Zellen strecken sich quer zur Achse. Die Markstrahlen verbinden das Stamminnere mit den äußeren Teilen, also das Mark mit der Rinde und das Holz mit dem Bast. Diese Verbindung ist sehr wichtig für den Stoffaustausch. Durch sie erreichen Photosyntheseprodukte aus den Siebröhren die lebenden Holzzellen, wo sie als feste Reserven, nämlich als Stärke oder Fetttröpfchen, abgelagert werden. Wasser, Nährsalze und remobilisierte Reservestoffe gelangen hingegen vom Holz in den Bast. Im primären Stamm werden die Parenchymzonen zwischen den Bündeln als primäre Markstrahlen bezeichnet. Beim sekundären Dickenwachstum genügt es nicht, diese primären Markstrahlen nur zu verlängern. Ein alter Stamm hat einen großen Umfang, und die wenigen primären Markstrahlen könnten dort nur einen kleinen Teil des Bastes anzapfen und nur mit einem kleinen Teil des Holzparenchyms in innigen Kontakt treten. Um auch die Zwischenbereiche zu versorgen, teilen sich daher von Zeit zu Zeit Fusiforminitialen in Gruppen oder einzeln quer und werden zu kleineren Markstrahlinitialen. Das Kambium gibt dann an diesen Stellen nach innen und außen Markstrahlzellen ab. Diese sekundären Markstrahlen beginnen im Holz und enden im Bast. Es wurde gezeigt, daß immer dann, wenn Fusiforminitialen zu weit vom nächsten Markstrahl abrücken, durch die fehlende Versorgung mit 87 88 Stoffen aus diesen Markstrahlen eine Teilung dieser Initialen erzwungen wird. Das ergibt einen sogenannten "Mangelmustereffekt", der zu einer sehr regelmäßigen Verteilung der Markstrahlen im Holzkörper führt. Neben der Querleitung dienen die Markstrahlzellen auch der Reservestoffspeicherung während der Vegetationsruhe und der radialen Versteifung, da ihre Zellen klein und ihre Wände stark verdickt sind. Sie sind meist auch massiv verholzt. Starke Tüpfelung ermöglicht dennoch leichten Stoffaustausch. Rhythmen des Dickenwachstums Das sekundäre Dickenwachstum folgt Rhythmen. Viele Jungpflanzen verdicken schon im ersten Jahr die Basis ihrer Stengel durch die Tätigkeit des Kambiums, jedoch erst, wenn die Differenzierung der primären Gewebe beendet ist. Sie beginnen daher relativ spät im Jahr mit ihrem sekundären Dickenwachstum. Anders bei älteren Stämmen: Das Kambium der Holzpflanzen kann an einer Stelle des Stammes viele Jahre und Jahrzehnte hindurch neue Leitelemente bilden. Es beginnt sich im Frühjahr zu teilen, sobald die Bedingungen es zulassen; das ist je nach Breitengrad, Höhenlage und Witterung zeitlich verschieden. Ein grober Richtwert für Tieflagen in Mitteleuropa wäre ein Zeitpunkt im Monat April. Ebenso ist das Ende der Teilungen im Spätsommer nach Art und Standort verschieden. Meist tritt die Ruhepause Ende August bis Ende September ein. Man darf jedoch nicht annehmen, daß die Teilungen in der Zwischenzeit ständig im gleichen Rhythmus erfolgen; es gibt aktive und weniger aktive Phasen. So sind etwa für die Rotföhre (Pinus sylvestris) zwei aktive Phasen und zwei Pausen des Kambiums allein während der Frühjahrsmonate beschrieben worden. Die Pausen entsprechen dem Austrieb der Nadeln Ende April und der Entwicklung der weiblichen Zapfen Ende Mai. Das ist verständlich: Nadelschub und Zapfenwachstum benötigen organische Substanzen, die teils in die neugebildeten Zellen eingebaut, teils zum Energiegewinn veratmet werden. Nun richtet sich die Verteilung der Assimilate im Körper der Pflanze stets nach der Konkurrenzkraft der Verbraucherstellen, der "Sinks". Gegenüber Nadelschub und Zapfenwachstum zieht das Kambium also den Kürzeren und pausiert. 88 89 Bei der Föhre teilt sich die einzelne Zelle eines aktiven Kambiums etwa jede Woche, ihre Streckung hält etwa 2 - 3 Wochen an, und die Ausbildung der Tracheidenwände dauert nochmals etwa 3 - 4 Wochen. Alle diese Vorgänge werden durch Umweltfaktoren wie Temperatur und Wasserversorgung beeinflußt und modifiziert. Das Holz Wir haben erwähnt, daß die Kambiumzellen weit häufiger Holzzellen als Bastzellen bilden. Auch sind Bastzellen recht kurzlebig, und die äußere Borke verwittert langsam. Daher ist das Holzvolumen ein Mehrfaches des Bastvolumens. Holz ist ein besonders wertvolles und vielseitig nutzbares Naturprodukt. Wir haben auch gesehen, daß im Holz der evolutionär jüngeren Dikotylen neue Elemente entstanden. Dese Elemente verbesserten die Hauptfunktionen des Holzes, Leitung und Festigung. Bei den Gymnospermen müssen die Tracheiden eine Doppelfunktion erfüllen: Sie leiten Wasser und sie festigen den Pflanzenkörper. Beides tun sie bemerkenswert gut, wenn man bedenkt, daß hier eigentlich ein Widerspruch besteht. Leitrohre sollten große Innendurchmesser haben; Festigungselemente sollten dickwandig sein, und die aus ihnen aufgebauten Gewebepartien werden durch Hohlräume geschwächt. Die Tracheiden sind jedoch sehr variabel. Sie können dickwandig und englumig oder aber dünnwandig und weitlumig ausgebildet werden. Sie können also sowohl festigen wie leiten, doch sind sie für beides nicht optimal gebaut. Bei den Dikotylen leiten hauptsächlich die Tracheen. Sie haben Durchmesser zwischen 0.02 und 0.5 mm, während Tracheiden oft weniger als 0.1 mm erreichen. Auch Tracheen haben natürlich dicke Wände, da sie ja unter der inneren Spannung der Wassersäule nicht kollabieren dürfen. Die Masse der 3 Wände pro mm wäre aber gering, wenn das Gewebe nur aus den weitlumigen Tracheen bestünde. Daher festigen neben den weiter vorhandenen Tracheiden die phylogenetisch neuen Libriformfasern. Ihre Lumina sind überaus eng, sodaß sie für die Wasserleitung nicht mehr in Frage kommen; umso effizienter sind sie als Festigungselemente. Das Kambium arbeitet, wie wir gesehen haben, nur während des Sommerhalbjahres, etwa von April bis September. Der in diesem Zeitraum gebildete Holzzuwachs legt sich als Kegelmantel um die älteren Teile des Holzes; er 89 90 heißt Jahrring, da er am Querschnitt einen farblich abgesetzten Ring bildet. Im Jahrring lassen sich anatomische Unterschiede zwischen dem zu Beginn gebildeten Frühholz und dem Spätholz des Augusts und Septembers erkennen. Besonders deutlich unterscheiden sich Früh- und Spätholz bei den hauptsächlich aus Tracheiden aufgebauten Nadelhölzern. Das erklärt sich aus der großen Variabilität der Tracheiden. Frühholztracheiden sind weitlumig und dünnwandig; sie dienen in erster Linie der Wasserleitung. Die englumigen und dickwandigen Spätholztracheiden sind vor allem Festigungselemente. So werden auch die Jahrringgrenzen mit freiem Auge sichtbar: Sie werden dadurch gebildet, daß auf die engen Tracheiden des dünkleren Spätholzes sofort das weitlumige, helle Frühholz des nächsten Jahrringes folgt. Früh- und Spätholz unterscheiden sich auch bei den ringporigen Laubhölzern deutlich. Sie bilden im Frühjahr zunächst eine oder mehrere Lagen sehr weiter Gefäße, deren Lumina schon mit freiem Auge als Poren sichtbar sind. Diese Zone hebt sich vom dichten Spätholz mit engen Gefäßen auffällig ab und läßt auch die Jahrringgrenzen scharf hervortreten. Die engeren Tracheen der zerstreutporigen Laubhölzer sind dagegen gleichmäßiger über den Jahrring verteilt. Sie zeigen daher keine auffallenden Unterschiede zwischen Früh- und Spätholz und keine deutlich hervortretenden Jahrringgrenzen. Durch Jahrringzählung kann man das Alter von Bäumen bestimmen. Langlebig sind zum Beispiel folgende Arten: Ulmen bis 600 Jahre Eichen 1000 Jahre Linden 1000 Jahre Eiben 3000 Jahre Sequoiadendron (Mammutbäume, Nordamerika) 4000 Jahre Pinus longaeva (Borstenkiefer, Nordamerika) 4600 Jahre Hingegen ist die Lebensdauer von Pionierhölzern wie Grauerlen und manchen Weiden mit weniger als hundert Jahren begrenzt. Jahre, die für das Wachstum günstiger oder ungünstiger sind, führen zur Bildung unterschiedlich breiter Jahrringe. Es ergeben sich über längere Zeiträume charakteristische Muster, die für Bäume einer Art in einem bestimmten Wuchsgebiet übereinstimmen. Diese Muster bleiben auch in längst gefällten 90 91 Stämmen erhalten, deren Außenbereiche so alt sind wie die innersten Zonen jetzt noch stehender Individuen. Man kann diese Überlappung in die Vergangenheit ausdehnen und Jahrringmuster für Perioden erarbeiten, die weit vor der Lebenszeit der ältesten Bäume liegen. Ausgehend von frisch gefällten Eichen bis hin zu subfossilen Stämmen aus dem Donauschotter ist es so gelungen, für Südwestdeutschland eine 7000jährige "Normkurve" zu erstellen, in die sich Eichenholz unbekannten Alters einordnen und damit datieren läßt. Baudaten von Dachstühlen mittelalterlicher Dome und von keltischen Hügelgräbern wurden so sehr präzise bestimmt. An der Boku wird derzeit Grubenholz aus Hallstatt datiert, nachdem in der Nähe durch Taucher uralte Stämme aus einem Bergsee geborgen worden waren. Bei den meisten Holzarten sind nicht alle im Laufe der Jahre gebildeten Jahrringe dauernd an der Wasserleitung beteiligt; oft ist im Stamm nur eine schmale, äußere Zone aktiv, die als Splint oder Splintholz bezeichnet wird, während die älteren, innen gelegenen Teile ihre Funktion verlieren. Das äußert sich so, daß die lebenden Elemente des Holzes absterben und sich die Leitelemente mit Luft füllen. Tracheen werden oft durch die Bildung von Thyllen verstopft. Dabei haben die Holzparenchymzellen eine entscheidende Rolle. Sie lösen enzymatisch eine Tüpfelmembran des Gefäßes auf, vergrössern ihr Volumen und wölben sich durch das Loch in das Lumen des Gefäßes vor. Durch Teilungen entsteht dort ein neues Gewebe, das die Tracheen ausfüllt. Man nimmt heute an, daß diese meist gerbstoffreichen Zellen vor allem das Wachstum von Pilzen in den Gefäßlumina blockieren sollen. Behält das nichtleitende Holz die gleiche Farbe wie der Splint, heißt es Reifholz (Fichte, Tanne, Weide), nimmt es aber durch Einlagerung von Gerbstoffen, Gummi, Harzen oder Farbstoffen eine dunklere Farbe an, nennt man es Kernholz (Eiche, Kiefer, Lärche, Tropenhölzer). Wir haben bisher den Querschnitt des Holzes betrachtet; der Aufbau des Holzes ist sehr kompliziert und erfordert zum vollen Verständnis eine räumliche Betrachtungsweise. Dabei muß man sowohl makroskopisch wie mikroskopisch von genau definierten Schnittführungen ausgehen. Die drei standardisierten Schnittebenen für holzanatomische Untersuchungen werden anatomische Hauptschnitte genannt. Es sind dies: 91 92 a) Der Querschnitt: Er wird senkrecht auf die Längsachse des Stammes geführt und zeigt vor allem den Verlauf und die Ausprägung der Jahrringe. Er schneidet Gefäße, Tracheiden und Fasern quer, kann also zur Messung der Durchmesser und Wandstärken herangezogen werden. Markstrahlen werden der Länge nach aufgeschnitten, ihre Breite und Länge läßt sich vermessen. b) Der Radial- oder Spiegelschnitt verläuft durch den Mittelpunkt des Sprosses als Längsschnitt. Er zeigt in abwechselnder Folge das längsgeschnittene Früh- und Spätholz in Form schmaler Streifen. Der Name "Spiegelschnitt" rührt von den Markstrahlen her, die bei geradem Verlauf lange Streifen bilden und sich bei makroskopischer Betrachtung des Holzes durch ihren Glanz von der Grundmasse des Holzes abheben. Radialschnitte zeigen nur die Höhe eines Markstrahles, nicht jedoch seine Breite. Die Länge der Leitelemente läßt sich bestimmen. c) Der Tangential- oder Fladerschnitt verläuft parallel zum Radialschnitt in der äußeren Zone der Achse. Er schneidet abwechselnd Früh- und Spätholz an, wodurch eine eigenartige Zeichnung des Holzes, die Fladerung, entsteht. Der Fladerschnitt trifft die Leitelemente in Längsrichtung und schneidet die Markstrahlen quer, sodaß deren Breite und Höhe bestimmt werden kann. Der Bast Der Bast zeigt wenig ausgeprägte Jahrringe, seine Struktur wird durch Tiefenperiderme gestört. Das Wort Bast wird vom Wortstamm "binden" abgeleitet und bezieht sich auf die Verwendbarkeit der Bastfasern mancher Arten, etwa der Linde. Der Aufbau von Sproßsystemen Wir haben die Entwicklung und den inneren Bau des Stammes besprochen. Dabei haben wir gleich zu Anfang die Gliederung des Stammes in Knotenbereiche (Nodien) und Zwischenknotenbereiche (Internodien) betrachtet. Sie ist für die Gestalt der Pflanze von großer Bedeutung: Die Internodien strecken sich verschieden stark, und die Achse kann sich im Bereich der Nodien verzweigen. Von dieser Verzweigung wollen wir jetzt reden. Der negativ gravitrope Stamm wird als Hauptachse bezeichnet. Seitensprosse gehen aus Knospen hervor, die in den Achseln ihrer Tragblätter 92 93 als meristematische Zonen angelegt sind. Die Knospen sind von Blattanlagen umhüllte Vegetationspunkte, die entweder bald austreiben oder länger inaktiv bleiben ("schlafen"). Knospen von Holzgewächsen treiben in der Regel frühestens im Jahr nach der Anlage aus. Die Größe der Knospen an ein und derselben Achse variiert stark. Sie ist bereits ein Hinweis darauf, wie lang der aus ihnen entstehende Seitentrieb wird. Bei der basitonen Förderung werden die basalen Knospen am Sproß stärker ausgebildet. Sie treiben früh aus und bilden kräftige, die Hauptachse überragende Sprosse. Eine Hauptstammbildung unterbleibt. Diese Verzweigung kann nur Sträucher als Wuchsform ergeben. Die meisten Holzarten zeigen jedoch eine akrotone Förderung, die Knospen werden von der Spitze des Triebes zur Basis kleiner. Die Seitenzweige akroton geförderter Arten werden umso länger, je näher sie der Triebspitze stehen. Die kleinsten Knospen an der Basis treiben nicht aus, sie sind hormonell gehemmt und „schlafen“. Bei Entfernung der hemmenden Endknospe kann es aber zur Bildung von Regenerationstrieben kommen, die man Stockausschläge nennt. Je nach Art bleiben die Knospen verschieden lang austriebsfähig. So kann sich im ostösterreichischen Eichen-Hainbuchen-Mischwald im Niederwaldbetrieb, das heißt bei einer Umtriebszeit von 30 Jahren, nur mehr die besonders austriebsfähige Hainbuche (Carpinus betulus) halten. Gut sichtbar wird die akrotone Förderung bei Ahorn- und Eschenarten. Die ökologische Bedeutung der Akrotonie dürfte zunächst in der raschen Durchdringung des Luftraums für den Lichtgewinn liegen. Die Ausgaben für Zweige und Blätter im lichtschwachen Kroneninneren werden reduziert, was vor allem im Eigenschatten der mächtigen Wipfel alter Bäume wichtig wird. Bei manchen Bäumen sind die Unterschiede zwischen geförderten Trieben der Spitzenregion und den weniger geförderten der Mittelregion eher graduell, wie wir das etwa bei Ahorn und Esche gerade gesehen haben. In anderen Fällen werden die Unterschiede aber extrem; man unterscheidet dann Langtriebe und Kurztriebe. Bei dieser zweiten Form strecken sich die Internodien des Kurztriebes kaum, sodaß eine rosettenartige Anordnung der Blätter resultiert. Bei der Buche und einigen anderen Waldbäumen wachsen die Endknospen eines Kurztriebes im nächsten Jahr erneut als gestauchter Kurztrieb weiter. Das ergibt eine dichte Aufeinanderfolge von feinen Blattnarben der Knospenschuppen am Zweig, unterbrochen von einigen größeren Narben 93 94 der Laubblätter. Bei vielen Nadelbäumen treiben nur die der Endknospe benachbarten 4 - 6 Seitenknospen zu Langtrieben aus, die tiefer inserierten Knospen jedes Jahreszuwachses am Haupttrieb treiben nicht aus oder werden zu Kurztrieben. Die Folge ist ein stockwerkartiger Aufbau. Bei den heimischen Kiefern (Pinus-Arten) sitzen grüne Nadeln zu je zweien (bei der Zirbe zu fünft) an Kurztrieben. Die Langtriebe tragen nur Schuppenblätter. Eine wirtschaftlich bedeutende Rolle spielen Kurztriebe bei Kernobst (Apfel, Birne) und Steinobst (Kirsche, Zwetschke) aus der Familie der Rosengewächse. Man nennt sie hier Fruchtholz, da nur sie Blüten und Früchte tragen können; Langtriebe bilden ausschließlich grüne Blätter. Der obstbauliche Schnitt und die dadurch erreichte Kronenausformung optimieren Zahl und Stellung der Kurztriebe für die Obstproduktion. Wir haben jetzt den einzelnen Trieb und seine Seitensprosse betrachtet. Wachstum unter Verzweigung führt allmählich zum Aufbau eines Sproßsystems. Es ermöglicht die Durchdringung des Luftraums und eine Vergrößerung der photosynthetisch aktiven Oberfläche. Ausschlaggebend für den Aufbau des Sproßsystems ist das Verhalten der Endknospe, mit der der Jahrestrieb im Spätsommer abschließt. Es gibt zwei Haupttypen: Beim ersten Typ entsteht der längste Trieb des neuen Jahres aus der Endknospe. Der Haupttrieb verlängert sich also jedes Jahr und die Seitentriebe bleiben kürzer. Es entsteht ein Monopodium. Monopodial verzweigte Bäume haben einen durchgehenden, gegenüber den Seitenästen stets dominierenden Schaft. Hierher gehören alle Nadelhölzer, ebenso Erle, Esche und Ahorn. Der Umriß ihrer Krone ist typischerweise pyramidal. Den Gegensatz bildet das Sympodium. Hier werden beim Neuaustrieb die obersten Seitenknospen gegenüber der Endknospe gefördert. Beim Monochasium übernimmt nur ein Seitensproß die Führung. Er schließt im Herbst mit einer geschwächten, verkümmernden Triebspitze ab. Der Fortsetzungstrieb entsteht wieder unter der Spitze und schwenkt in die Wachstumsrichtung der alten Hauptachse ein. Dieses Verzweigungssystem ist oft nur bei genauem Studium von einem Monopodium zu unterschieden. Hainbuche, Ulmen und Linden sind Beispiele. Beim Dichasium setzen die beiden obersten Seitenknospen in den Achseln gegenständiger Blätter das Wachstum fort. Das sehen wir beim Flieder, beim Kreuzdorn oder bei der 94 95 Mistel. Die Folge ist ein Verzweigungssystem ohne klar erkennbare Hauptachse, daher ergibt sich wieder strauchiger Wuchs. Besonders anschauliche Beispiele für die verschiedenen Verzweigungsmöglichkeiten liefern die Blütenstände oder Infloreszenzen. Man versteht darunter Sproßsysteme, die der Blütenbildung dienen und dementsprechend verändert sind. Ihre Verzweigung unterscheidet sich meist deutlich vom vegetativen Bereich. Es fällt hier etwas weg, was für die photosynthetisch aktiven Teile des Pflanzenkörpers wichtig ist: Die optimale Einstellung der Blätter zum Licht spielt keine Rolle mehr. Das erlaubt eine große Zahl an "spielerischen" Abwandlungen, die teilweise unglaublich kompliziert sind. Wir können hier nicht näher darauf eingehen. Einige häufig vorkommende Typen von Blütenständen werden Sie im Sommersemester kennenlernen. Was wir bis jetzt vom Bau des Stammes besprochen haben, war das Typische, das sich bei den meisten Arten mit leichten Abwandlungen findet. Doch kann der Stamm, ebenso wie die anderen Organe, radikal umgebaut werden, um ihn an besondere physiologische oder ökologische Anforderungen anzupassen. Solche Veränderungen des typischen Bauplans nennt man Metamorphosen. Meist bleiben solche Abwandlungen nicht auf ein Organ beschränkt; wir werden daher erst nach Besprechung des typischen Bauplanes der beiden anderen Organe, des Blattes und der Wurzel, die Metamorphosen des Kormus zusammenfassend besprechen. Das Blatt Das Blatt ist morphologisch und anatomisch das variabelste Pflanzenorgan. Man kann sehr verschieden gebauter Blätter an einer Pflanze finden, die im Laufe der individuellen Entwicklung in festgelegter Abfolge auftreten. Alle sind, ebenso wie der Stamm, an dem sie wachsen, Produkte des Sproßvegetationspunktes. Das Apikalmeristem wird also öfter umgestimmt (umprogrammiert), es erzeugt nacheinander verschiedene Arten von Blättern. Die Blattfolge am Stamm Die Keimblätter oder Kotyledonen sind die ersten Blätter der jungen Pflanze. Ihre Funktion wurde bereits besprochen. 95 96 Auf die Kotyledonen folgen die Laubblätter, zunächst oft einfach gebaute Primärblätter, deren Hauptfunktion die Photosynthese ist, ebenso wie bei den reicher gegliederten Folgeblättern. Die Laubblätter sind die größten Blätter. So wie die Hauptachse des Keimlings beginnen auch die Seitensprosse ihr Wachstum mit einfach gebauten, stark reduzierten Blättern, den Knospenschuppen der Winterknospen, bevor sie Laubblätter bilden. Die Internodien zwischen diesen Schuppen sind ungestreckt, sie sitzen also dicht aneinander. Solche Blätter werden auch an allen Vegetationspunkten des Haupttriebs und der Seitentriebe im Herbst angelegt, um das Meristem während der Vegetationsruhe durch Ausbildung einer Winterknospe vor Wasserverlust zu schützen. Zahl, Form und Farbe der Knospenschuppen sind für die Holzpflanzen sehr charakteristisch und daher ein wichtiges Hilfsmittel für die Bestimmung von Bäumen und Sträuchern im Winterzustand. Geht der Sproß zur Blütenbildung über, dann verändert sich erneut die Blattgestalt. Die Hochblätter unterscheiden sich von den Laubblättern durch einfachere Form, oft geringere Größe und manchmal auch durch die Farbe, wenn sie Schaufunktionen übernehmen. Oft entspringen die Blüten in den Achseln von Hochblättern. Blüten sind Sprosse, und zwar meist Seitensprosse. Gelegentlich beendet aber auch das Apikalmeristem des Haupttriebes seine Tätigkeit mit der Ausbildung von Blütenorganen. In der Blüte selbst findet man dann Kelchblätter und Kronblätter mit Schutz- und oft Schaufunktion, Staubblätter, die den Pollen produzieren, und Fruchtblätter, die die Samenanlagen tragen. Mit der Bildung der Fruchtblätter erschöpft sich der Vegetationskegel der Blüte und stellt sein Wachstum ein. Die Entwicklung und der Bau des Blattes Nach der Anlage des Blattes an der Sproßspitze wächst es durch Zellteilung, Streckung und Differenzierung. Bei den Spermatophyten differenziert sich die Blattspitze früh aus, das Wachstum wird mit Hilfe eines oder mehrerer interkalarer Meristeme fortgesetzt. Das starke Breitenwachstum vieler Blätter kommt daher, daß meristematische Zellen an den Blatträndern sich noch lange als "Randmeristem" teilen. Unterschiedliche Aktivität dieser interkalaren Randmeristeme bewirkt Zähnung, Lappung oder Fiederteiligkeit der Spreite. 96 97 Wir beschränken uns im folgenden auf die Besprechung des äußeren und des inneren Baus der typischen Laubblätter. Die übrigen Blattorgane zeigen meist ähnlichen, aber vereinfachten Bau. Wir müssen jedoch die Laubblätter der Angiospermen und der gymnospermen Koniferen getrennt besprechen, da zwischen ihnen ziemlich große Unterschiede bestehen. Angiospermen: Das typische Laubblatt der Monokotylen und Dikotylen ist ein flächig ausgebreitetes Assimilationsorgan. Bereits in einem sehr frühen Entwicklungsstadium sieht man an den Blattanlagen zwei deutlich getrennte Abschnitte, Oberblatt und Unterblatt, aus denen später Blattspreite und Blattstiel einerseits, Blattgrund und Nebenblätter andererseits hervorgehen. Aus dem Unterblatt entstehen also der verbreiterte Blattgrund, mit dem das Blatt dem Stengel aufsitzt und das diesen bei manchen Arten als Scheide umfaßt, sowie zwei kleine Anhänge, die Nebenblätter, die nur selten fast laubblattartig ausgebildet sind, in vielen Fällen aber auch fehlen können. Aus dem Oberblatt entsteht zunächst der Blattstiel. Er leitet Wasser und Photosyntheseprodukte. Daneben kann er durch Wachstum die Blätter im Raum orientieren und optimal zum Licht einstellen. Es entsteht so ein Blattmosaik zur Ausnützung des einfallenden Lichtes. Liebhaber von Topfpflanzen wissen, daß diese ein ständiges Drehen der Töpfe, also eine Änderung der Orientierung gegenüber dem einfallenden Licht, sehr übel nehmen. Sie können die richtige Lichtorientierung nicht erreichen und reagieren meist mit Blattabwurf auf die Überforderung der Blattstiel-Reaktion. Der Blattstiel ist jedoch keineswegs überall vorhanden. Den breit angesetzten Blättern vieler Monokotyler, etwa der Gräser, fehlt er völlig. In diesen Fällen geht der Blattgrund, eine verlängerte Blattscheide, direkt in die Spreite über. In einigen morphologisch interessanten Sonderfällen, etwa bei Akazien der afrikanischen Trockensavannen, wird der Blattstiel verbreitert und übernimmt an Stelle der verkümmerten Spreite die Photosynthese. Die Blattspreite ist in der Regel dorsiventral gebaut und zeigt an der Oberseite ein dunkleres Grün. Sie ist meist eine geschlossene Fläche, die vom Rand her mannigfaltig gezähnt, gelappt oder geschlitzt sein kann. Gelegentlich löst sie sich in mehrere Teilblättchen (Finger- oder Fiederblättchen, die der Mittelachse des Blattes, der Rhachis, ansitzen) auf. Die Monokotylen haben vorwiegend einfache Blätter (Ausnahme: viele Palmen), während 97 98 Fiederblättchen für manche dikotyle Familien, etwa die Hülsenfrüchtler oder Leguminosen, charakteristisch sind. Das folgende Schema zeigt den Bau der Laubblätter in Übersicht: Blattspreite (oder Blattachse und Fiedern) Oberblatt: Blattstiel (kann auch fehlen) Blattgrund Unterblatt: Nebenblätter (können fehlen) Die Histologie der Spreite Die Anordnung der drei Gewebesysteme zeigt große Mannigfaltigkeit, die sowohl auf systematische Unterschiede wie auf Anpassungen an ökologische Anforderungen zurückgeführt werden kann. Wir wollen zunächst das Laubblatt heimischer Angiospermen besprechen, deren Photosynthese dem C3-Weg folgt. Die subtropischen C4-Pflanzen (etwa Mais und Sorghum) haben einen abweichenden Blattbau, der mit physiologischen Besonderheiten zusammenhängt und später besprochen werden soll. Hautgewebe ist die Epidermis. Periderme treten nur nach Verwundung auf. Die wichtigsten Strukturen der Blattepidermis sind die Spaltöffnungen, die den Blättern die Photosynthese ermöglichen. Sie regeln die Abgabe von H2O und die Aufnahme von CO2. Die Spaltöffnungen sind über die Gesamtfläche des Blattes verschieden verteilt. Untergetauchte Wasserblätter haben oft überhaupt keine funktionsfähigen Spaltöffnungen; sie nehmen die gelösten Gase über die gesamte, nur sehr schwach kutinisierte Epidermis auf. Schwimmblätter, etwa die von See- und Teichrose, tragen Spaltöffnungen ausschließlich auf der Oberseite, sind also epistomatisch. Die Gasdiffusion ist in Flüssigkeiten viel langsamer als in Gasen. Es ist daher logisch, daß Schwimmblätter, deren Evolution ja sozusagen den Weg ihrer Versorgung mit Sauerstoff und Kohlendioxid frei wählen konnte, sich über die Gasphase versorgen. Andererseits hängt bei Landpflanzen die Verteilung der Spaltöffnungen von der Orientierung des Blattes im Raum ab. Steilstehende Blätter haben 98 99 meist Stomata auf beiden Seiten, sind also amphistomatisch. Steht das Blatt eher waagrecht, ist es oft hypostomatisch mit Öffnungen an der Unterseite. Der ökologische Vorteil von Stomata auf der lichtabgewandten Seite des Blattes liegt in geringerer Überhitzung, daher auch geringerer Wasserabgabe der Blattzellen, die den Spaltöffnungen benachbart sind. Das Grundgewebe des Blattes ist das Mesophyll. Seine Hauptmasse ist gewöhnlich ein Assimilationsparenchym, das durch Interzellularen mit Luft versorgt wird. Dieses Parenchym ist in Lagen geteilt: Das Palisadenparenchym besteht aus zylindrischen Zellen senkrecht auf die Epidermis. Die Zellen des Schwammparenchyms haben unregelmäßige Formen und sind von besonders großen Interzellularen umgeben. Der Name vergleicht das Gewebe mit dem Badeschwamm, der ebenfalls reich an Hohlräumen ist. Meist tritt das Palisadenparenchym in einer oder mehreren Reihen an der Oberseite, das Schwammparenchym an der Unterseite des Blattes auf. Solche Blätter heißen dorsiventral oder bifazial. Viele Blätter sind aber auch beiderseits gleich gebaut: man nennt sie äquifazial. Nur an den Bündeln, in denen das Xylem zur Oberseite gekehrt ist, kann man sich hier orientieren. Hierher gehören einmal viele Wasserpflanzen, bei denen das Palisadenparenchym ganz fehlt. Andererseits finden sich äquifaziale Blätter auch an Landpflanzen. Das sind meist Arten, deren Blätter aufrecht stehen, wie Schwertlilien oder Bogenhanf (Sansevieria). Bei ihnen grenzt an beide Epidermen eine Schicht von Palisadenparenchym. Die Anteile von Palisadenparenchym und Schwammparenchym am Blattquerschnitt können in Blättern derselben Pflanze verschieden sein. Hierfür sind einmal Außenfaktoren verantwortlich. Wassermangel und Starklicht begünstigen die Ausbildung von Palisadenparenchym; man spricht dann von Sonnenblättern, da angespannter Wasserhaushalt meist mit starker Belichtung einhergeht. Bei gedämpftem Licht und guter Wasserversorgung, etwa an der Basis der Baumkrone, werden Schattenblätter gebildet. Sie zeigen einen hohen Anteil von Schwammparenchym. Letzten Endes sind diese quantitativen Unterschiede im Bau hormonell vermittelt. Das sieht man unter anderem daran, daß ähnliche Unterschiede auch, unabhängig von der Umwelt, im Zuge der Entwicklung auftreten können. Bei vielen Pflanzen finden sich deutliche Unterschiede zwischen Jugendblättern (die eher den Schattenblättern 99 100 entsprechen) und Altersblättern von der Art der Sonnenblätter. Auch dafür sind hormonelle Umstimmungen verantwortlich, die besonders bei der Erlangung der Blühreife einsetzen. Der Efeu ist ein gutes Beispiel, bei ihm ändert sich auch die äußere Gestalt der Blätter. – Oft ist ein Teil des Mesophylls ein Festigungsgewebe aus Kollenchym oder Sklerenchym. Sehr häufig liegt eine Bündelscheide um das Stranggewebe; weniger oft, und zwar besonders bei Pflanzen von Trockenstandorten, sind es die Zelllagen unmittelbar unter der Epidermis, die als Hypodermis verdickt sind und festigen. Das Stranggewebe des Blattes dient der Wasser- und Nährstoffversorgung, der Ableitung der Assimilate und der Festigung. Die Blattspreite wird von Bündeln in verschiedener Zahl und Größe durchzogen. Man bezeichnet die Blattbündel als Adern oder Nerven. Gewöhnlich springen die dickeren Adern, die man auch Rippen nennt, auf den Unterseiten leistenförmig vor; oberseits entsprechen ihnen oft Vertiefungen oder Furchen. Oft verläuft in der Mitte der Spreite ein besonders kräftiger Strang, die Mittelrippe. Im Frühling findet man auf dem Boden zahlreiche Blätter, deren Epidermis und Grundgewebe ausgefault sind, von denen sich aber das zarte Maschenwerk der Nerven als "Skelett" erhalten hat. Dieser Anblick zeigt uns, daß die Nerven auch die Aufgabe der Festigung und der Formgebung erfüllen; bei zarten Blättern ist hierfür zusätzlich der Turgordruck sehr wichtig. - Die Vielfalt der anatomischen Bezeichnungen fällt auf; das Stranggewebe des Blattes besteht also aus "Adern", "Nerven" oder "Rippen", die funktionell kaum Ähnlichkeiten mit den gleichnamigen Strukturen bei Mensch und Tier zeigen. Das ist historisch zu verstehen: Die Pflanzenanatomie begann weit später als die medizinische und tierische Anatomie und wurde zunächst von Ärzten betrieben, für die die Pflanze ein Lieferant von Heilmitteln war. Man war damals über die Funktionen selbst im Tierkörper nur schlecht informiert, aber über den Pflanzenkörper wußte man gar nichts. Daher suchte man nach oberflächlichen Ähnlichkeiten: Dem einen schien der Vergleich mit Nerven naheliegend, dem anderen der mit Adern. Ähnliche etwas ungeschickte Begriffe finden sich in der Pflanzenanatomie allenthalben: Denken Sie an die "Haare" oder an die "Stomata", was ja wörtlich "Münder" bedeutet. Die großen Pflanzengruppen unterscheiden sich durch Verlauf und Verzweigung der Nerven in der Spreite. Die Angiospermen haben 100 101 "vereintläufige" Nerven, die untereinander verbunden sind, die Farne und Gymnospermen "getrenntläufige". Streben die Hauptnerven, die schon makroskopisch gut sichtbar sind, unabhängig voneinander der Blattspitze zu, so spricht man von streifiger Nervatur. Diese ist für die meisten Monokotylen bezeichnend. Die Querverbindung durch feinere Nerven erfolgt hier nach Art dünner Leitersprossen. Hingegen sind bei den Dikotylen die Hauptnerven maschenartig verbunden, was man netzige Nervatur nennt. Die starken Nerven lassen zwischen sich Interkostalfelder frei, in die blind endende feine Nerven eindringen. Die C4-Blätter, die es nur bei Angiospermen gibt, unterscheiden sich von den oben besprochen C3-Blättern durch eine ganz andere Anordnung des photosynthetisch aktiven Grundgewebes, des Mesophylls. Es gibt keine Schichtung von Palisadenparenchym und Schwammparenchym, die chloroplastenführenden Zellen ordnen sich um die Nerven an, die im Querschnitt von einem Kranz aus Mesophyll umgeben sind ("Kranzanatomie"). Auf die physiologische Bedeutung dieses Bauplanes gehen wir im Rahmen der Ökophysiologie ein. Koniferen: Die Nadelblätter der heimischen Koniferen werden als Anpassung an die Trockenheit gedeutet. Es fehlen Nebenblätter und Stiel. Immergrüne Koniferen transpirieren auch im Winter, wenn Wasser aus dem gefrorenen Boden schwer oder nicht nachgeleitet wird. Der Nadelbau schränkt diese Transpiration stark ein. Die Oberfläche ist klein im Verhältnis zum Volumen, das Interzellularensystem mäßig entwickelt. Die Epidermis ist tot, sehr stark kutinisiert und so massiv verdickt, daß die Lumina fast verschwinden. Ähnlich verdickt ist die Subepidermis, die auch Hypodermis heißt. Die Stomata sind tief eingesenkt, wodurch vor ihnen eine windgeschützte Höhle entsteht. Oft ist diese durch poröse Wachse verstopft. Die Tanne und einige exotische Arten haben zwei Reihen von Spaltöffnungen mit weißlichen Wachsschichten an der Unterseite der Nadeln. Die Nerven der Farne und Gymnospermen sind "getrenntläufig": In flächigen Blättern (Farne, Ginkgo) verzweigen sich die Bündel nach dem Eintritt in das Blatt, die einzelnen Äste werden aber nicht mehr verbunden. Dieser 101 102 primitive Typ ist offensichtlich nicht sehr günstig, da bei Beschädigungen durch Insektenfraß, Pilzbefall oder Hagel, die einzelne Nerven durchtrennen, ganze Blattbezirke ohne Zu- und Ableitung bleiben müssen. Bei den heimischen Koniferen sind ein bis zwei unverzweigte Bündel in ein chlorophyllfreies Grundgewebe aus lebenden (assimilatleitenden) und toten (wasserleitenden) Elementen, das Transfusionsgewebe, eingebettet, das Stoffe zwischen Bündel und Mesophyll vermittelt. Zum Assimilationsparenchym hin ist das Transfusionsgewebe von einer Zellschicht abgegrenzt, die als Endodermis bezeichnet wird, ein Name, der uns auch für ein Gewebe der Wurzel begegnen wird, aber bloß "Innenhaut" bedeutet. Koniferennadeln sind der Länge nach von Harzgängen durchzogen; das sind schizogene Interzellularen zwischen lebenden, harzproduzierenden Sekretionszellen. Sie fehlen der Eibe. Blattaustrieb und Blattfall Die Blätter entstehen als Anlagen am Vegetationskegel. Viele Arten, darunter Ahorn, Roßkastanie, Buche und Eibe, legen alle Blätter für das nächste Jahr schon ab Juni oder Juli an, sobald die Blätter für dieses Jahr entfaltet sind. Andere Arten, wie Birke oder Liguster, legen nur einen Teil der Blätter vor der Winterruhe an, die übrigen Anlagen werden im Austriebsjahr gebildet. Die Blattanlagen treiben in Sonderfällen vorzeitig aus, und zwar dann, wenn die entfalteten Blätter einer Katastrophe, wie Spätfrost oder Insektenfraß, zum Opfer fallen. Baumarten unterscheiden sich stark in dieser Regenerationsfähigkeit. Linde, Flieder und Esche sind nur zu schwacher Erneuerung in der Lage; Eiche, Ulme und Ahorn können ihre Blätter einmal im Jahr erneuern, Birke, Erle und Robinie sogar öfter. Starke Regenerationsfähigkeit ist übrigens auch günstig für die Resistenz gegen Industrieabgase. Gegen Ende ihres Lebens altern die Blätter physiologisch. Beim Altern hören Nukleinsäure- und Proteinsynthese (außer von abbauenden Enzymen) fast völlig auf. Die Synthese wird durch Phytohormone gehemmt, dagegen gehen Abbauvorgänge weiter. Aminosäuren, Zucker und Ionen werden freigesetzt und im Phloem verlagert. Sommergrüne speichern diese wertvollen Stoffe im Herbst vor allem im Stamm und in der Wurzel. Bei den Immergrünen, 102 103 deren alte Blätter nach mehreren Jahren der Tätigkeit über die ganze Saison verteilt abgestoßen werden, bauen junge Blätter diese Nährstoffe ein. Schließlich wird an der Basis des Blattstieles eine schon vorhandene Trennschicht aktiviert. Sie besteht aus kleinen Parenchymzellen mit wenigen Interzellularen. Die eigentliche Abtrennung ist ein aktiver Prozeß, bei dem Mittellamellen oder ganze Zellwände aufgelöst werden. Diese Vorgänge sind hormonell gesteuert, wobei Auxine und Abscisinsäure zusammenwirken. Nötige Enzyme, also Pektinasen und manchmal auch Zellulasen, werden neu gebildet. Das benötigt sowohl Zeit als auch Energie, die durch Atmung beigestellt wird. Plötzliche Kälteeinbrüche im Herbst, etwa zu Anfang November, überraschen die Bäume oft noch voll beblättert. Während der Kälte können die Blätter mangels an Stoffwechselaktivität nicht abgeworfen werden; wenn dann noch Schnee fällt, kann es zu argen Fällen von Schneebruch kommen. Die Blattalterung und der Blattfall werden durch zwei Umweltfaktoren beeinflußt: neben der Temperatur spielt auch die Tageslänge eine Rolle. Dabei reagieren offenbar manche Arten eher auf tiefe Temperaturen (wie etwa manche Walnußsorten, die die Tageslänge nicht kümmert, solange nur die Tage und Nächte einigermaßen warm bleiben), andere stärker auf die Tageslänge. Dazu gehören manche Alleebäume, die gelegentlich durch Straßenlaternen übertölpelt werden: Äste, die sehr nahe an die Lampe heranwachsen, bleiben dann weit länger beblättert als die weiter entfernten. Blattnarben am Zweig zeigen die Stellen, wo früher Blätter ansetzten. Sie werden durch Verkorkung (Suberineinlagerung) der Wände an der Wundfläche verschlossen, oft entsteht auch ein echtes Periderm. Form und Größe der Blattnarben kann als Bestimmungsmerkmal dienen. Die Wurzel Dieses Organ ist das Produkt des Wurzelvegetationspunktes und unterscheidet sich stark von Stamm und Blatt, die vom gemeinsamen Sproßvegetationspunkt erzeugt werden. Die Wurzeln verankern die Pflanze im Boden, und sie nehmen dort Wasser und Nährsalze auf, die sie dem Sproß zuleiten. Wurzeln entwickeln sich meist unter der Bodenoberfläche, doch gibt es Beispiele für Luftwurzeln ebenso wie für unterirdische Stammteile. Aus dem Bodenleben und den Aufgaben erklären sich einige Baueigentümlichkeiten: Die 103 104 Festigungsgewebe liegen zentral und nicht peripher, da die Wurzel im Boden nicht durch Biegung, sondern durch Zug beansprucht wird. Sie bewahrt damit Baueigenheiten, die in frühen Epochen der Evolution (etwa im Karbon) auch Stämme primitiver Bäume zeigten. Das Hautgewebe bewirkt engen Kontakt mit der Umgebung und nicht Abschluß. Stomata und Blattanlagen fehlen. Auch Dickenwachstum und Verzweigung der Wurzel zeigen eigene Züge. Die verborgene Lebensweise hat die Erforschung der Wurzel sehr behindert: Zwar wird auch ihre Physiologie immer besser untersucht, doch sind noch viele Fragen offen. Wir betrachten zuerst die Gliederung der Wurzel in der Längsrichtung. Nahe an der Spitze liegt das Apikalmeristem. Es braucht für seine zarten Zellen besonderen Schutz gegen mechanische Verletzung durch scharfkantige Bodenteilchen. Diesem Schutz dient eine Kappe aus parenchymatischen Dauerzellen, die Wurzelhaube oder Kalyptra. Das Meristem liegt also im Inneren der Wurzelspitze und scheidet zur Spitze hin Kalyptrazellen, zur Basis hin die Vorläufer der Dauergewebezellen ab. Die ältesten Zellen der Wurzelhaube lösen sich außen ab, sie mazerieren und werden von innen her durch neue Zellen ersetzt. Dabei bildet sich ein hochmolekularer KohlenhydratSchleim, der den Reibungswiderstand beim Eindringen der Wurzel in den Boden vermindert. Das Apikalmeristem bildet die Teilungszone der Wurzel. Es folgt die Determinationszone, deren Zellen sich zwar noch teilen, aber bereits die künftige Abgrenzung der Gewebe zeigen, da sie je nach Bestimmung unterschiedlich groß sind und bestimmte Teilungsrichtungen bevorzugen. Daran schließt unmittelbar die Streckungszone an. Sie ist nur einige Millimeter lang Dadurch wird die Wurzel beim Eindringen in den Boden nicht so leicht in dieser wenig gefestigten Zone, in der ja die Zellwände für das Wachstum erweicht werden müssen, verformt. Verlängerung der Zellen in der Streckungszone treibt die Spitze der Wurzel vorwärts. Abschnitte hinter der Streckungszone verändern ihre Lage relativ zum Boden nicht mehr. An die Streckungszone schließt die Differenzierungszone an. Ihren Namen hat sie davon, daß hier die inneren Gewebe (Phloem, Xylem und Grundgewebe) fertiggestellt werden. Äußerlich ist die Differenzierungszone an einer Rhizodermis mit Wurzelhaaren zu erkennen. Sodann folgt die Zone der älteren 104 105 Primärwurzel, deren Abschlußgewebe nicht mehr die Rhizodermis, sondern die Exodermis ist. Hier beginnt später auch die Bildung von Seitenwurzeln. Bei Gymnospermen und Dikotylen kommt vor dem Übergang in den Stamm noch die Zone der Sekundärwurzel. Wir sehen uns jetzt den inneren Bau der Wurzel an. Natürlich kann man die Wurzel an verschiedenen Stellen ihrer Längserstreckung quer schneiden und dort ihr Inneres untersuchen. In der Kalyptra finden wir nur lebende Zellen, die den Parenchymzellen des Grundgewebes ähneln. In der Teilungszone zeigt der Querschnitt nur Meristemzellen, in der Determinationszone und der Streckungszone verschieden große Zellen mit Primärwänden und Vakuolen, aber ohne deutliche Unterschiede im Bau der Protoplasten oder der Wände. Sehr verschieden sind die Zellen hingegen in der Differenzierungszone. Wir wollen uns daher einen Querschnitt durch diese Region ansehen, am besten durch den Endbereich, wo schon alle Zellen fertiggestellt sind. In der Differenzierungszone besteht das Hautgewebe der Wurzel aus Rhizodermiszellen, die eine einschichtige Lage bilden. Bezeichnend sind die dünnen Außenwände, denen eine Cuticula fehlt. Auch fehlen Stomata. Hingegen ist die Rhizodermis reichlich mit einzelligen, lebenden Wurzelhaaren ausgestaltet, die die Oberfläche vergrößern. Gase und Wasser samt den darin gelösten Salzen des Bodens können durch die dünnen Wände leicht aufgenommen werden. Unter der Rhizodermis findet sich eine meist mächtige Lage von Grundgewebe, die Wurzelrinde. Ihre äußerste Region tritt bereits in der Differenzierungszone durch abweichenden Bau hervor; es fehlen diesen 1 3 Schichten meist Interzellularen, und die Zellen sind auffällig klein. Das sind frühe anatomische Anzeichen dafür, daß die Zellen auch physiologisch abweichen und in der älteren Wurzel zu einem typischen Abschlußgewebe umgestaltet werden. Man nennt diese Schicht Exodermis. Die darunter liegende Hauptmasse der Rinde kann, wie jedes Grundgewebe, verschiedene Funktionen übernehmen. Häufig ist sie ein Speichergewebe; bei Sumpf-und Wasserpflanzen sind die Interzellularen oft so stark entwickelt, daß man das Gewebe Aerenchym nennt. Daneben gibt es auch Exkretion. Photosynthese ist natürlich bei Erdwurzeln aus Lichtmangel unmöglich, sie beziehen Stoffe vom 105 106 Sproß. Hingegen können Luftwurzeln Chloroplasten bilden und Photosynthese betreiben. Die innerste Rindenschicht vor dem Stranggewebe ist bei allen Wurzeln als Endodermis ausgebildet. Ihre Funktion hängt eng mit der Aufnahme von Mineralstoffen zusammen. Dieser Vorgang muß daher kurz besprochen werden. Die Zellwände der Rhizodermis und der Wurzelrinde sind für die Bodenlösung frei zugänglich: Wasser und Ionen können in den Hohlräumen der Zellwand (aber nicht in den luftgefüllten Interzellularen zwischen den Zellwänden!) bis zur Endodermis diffundieren. Alle lebenden Zellen können Stoffe aus der Zellwand in ihre Protoplasten aufnehmen. Das Wasser tritt durch Permeation ein, die Ionen der anorganischen Nährsalze werden durch aktiven Transport aufgenommen, wobei nur die von der Pflanze wirklich benötigten ausgewählt werden, da nur für sie die spezifischen Trägersysteme im Plasmalemma vorhanden sind. Nach der Aufnahme werden die Ionen durch das Protoplasma der Zellen und durch die Plasmodesmen von Zelle zu Zelle befördert. Transportiert wird also im Symplasten, der Gesamtheit der miteinander verbundenen Protoplasten. In der Wand bleiben jene Stoffe zurück, für die es keine Verwendung gibt, etwa Kieselsäure und Aluminiumsalze in Silikatböden oder der Überschuß von Calcium auf Kalkstandorten. Diese Stoffe wären eine Belastung für den Sproß; daher müssen sie zurückgehalten werden, bevor sie durch Zellwanddiffusion das Xylem und damit den "Schnelltransportweg" zu den Blättern erreicht haben. Die notwendige Sperre ist in die Zellwände der Endodermis eingebaut. Die Endodermiszellen schließen ohne Interzellularen aneinander. Ihre Wände haben eine eigentümliche Einlagerung von wasserabstoßendem Material, den Casparyschen Streifen. Er zieht sich als Band durch alle Radialwände der Zelle, also durch jene vier der sechs Wände, die von innen nach außen führen. Im Querschnitt sieht man schwache Wandverdickungen. Das eingelagerte Material ist chemisch dem Suberin ähnlich; oft findet sich auch Lignin. Die Einlagerung hydrophober Stoffe verhindert den Transport von Wasser und Ionen durch diese Zellwände. Nicht imprägniert sind nur die beiden Tangentialwände, über die aber kein direkter Zugang zum Wurzelinneren besteht. Stoffe, die die Endodermis passieren sollen, müssen also unbedingt 106 107 irgendwo in das Protoplasma eintreten und sind daher der Auswahl durch die Carrier unterworfen. Nicht aufgenommene Ionen diffundieren aus dem Zellwandraum wieder zurück in den Boden. Freilich sind die Trägersysteme nicht perfekt, es werden daher hin und wieder nicht benötigte Ionen in das Protoplasma aufgenommen und zum Xylem gebracht, jedoch in viel geringerer Menge, als dies ohne den Casparyschen Streifen der Fall wäre. Innerhalb der Endodermis liegt das radiäre Gefäßbündel der Wurzel. Unmittelbar auf die Endodermis, die innerste Grundgewebeschicht, folgt eine Schicht dünnwandiger Zellen, der Perizykel, ein Lateralmeristem. Neue Gewebe bildet er erstens bei der Entstehung von Seitenwurzeln, zweitens bei der Anlage von Peridermen und drittens beim sekundären Dickenwachstum der Wurzel. Das Phloem liegt in einzelnen Strängen nahe am Perizykel. Das mächtigere Xylem ist ebenfalls in Stränge aufgelöst, die mit den Phloemsträngen abwechseln und von ihnen durch Parenchymzellen getrennt sind. Jeder Strang liegt sozusagen auf einem eigenen, vom Zentrum ausgehenden Radius, daher der Name des "radiären" Bündels. Die Xylemstränge verschmelzen im Zentrum miteinander, wobei dort große Gefäße oder Xylemparenchym liegen können. Die Zahl der Stränge ist variabel und hat Beziehungen zur Systematik: Für Dikotyle und Gymnospermen sind niedere Zahlen von zwei bis fünf charakteristisch, während die Monokotylen Bündel mit höheren Strangzahlen enthalten. Am Beginn der Differenzierungszone ist das radiäre Bündel noch nicht voll funktionsfähig. Die ersten fertigen Siebröhren und Tracheen liegen an der Außenseite. Hier können also bereits Stoffe transportiert werden, sobald sich die ersten Wurzelhaare entwickeln. Im Verlauf der Differenzierungszone kommen immer neue Leitelemente dazu, bis das Bündel voll funktioniert und das ganze Xylem Wasser transportiert. Ich möchte ausdrücklich festhalten, daß wir bisher kein Wort vom Kambium gesagt haben. Dieses fehlt nämlich in der primären Wurzel aller Pflanzen, also auch der Dikotylen und Gymnospermen. Wir legen jetzt einen weiteren Querschnitt durch die Wurzel, diesmal in der Zone der älteren Primärwurzel. Hier hat sich einiges verändert. Die Rhizodermis ist sehr kurzlebig: Die Wurzelhaare funktionieren oft nur wenige Tage, dann stirbt das gesamte Hautgewebe ab. Der Wurzelkörper verliert damit 107 108 seine zur Umgebung hin offene Hautschicht und bildet durch Umbau der Exodermis ein Abschlußgewebe. Jetzt lagern ihre Zellwände Suberin ein, die Zellen bleiben aber dennoch am Leben, da die Wände stark getüpfelt sind. Die hydrophoben Exodermiswände verhindern die Wasserabgabe an trockene Bodenschichten. Allgemein wachsen ja die feinen Saugwurzeln in die wasserreichsten Bodenschichten. Die nahe der Oberfläche liegenden Schichten trocknen stärker aus als die tieferen. Ältere Wurzeln müssen das Wasser bis zum Übergang ins Hypokotyl, dem Wurzelhals, leiten und dabei auch diese trockenen oberen Bodenschichten durchqueren. Wurzelhaare würden diesen Schlauch "durchlöchern", das Suberin der Exodermis dichtet ihn ab. Seitenwurzeln werden erst in der älteren primären Wurzel angelegt. Das ist ein Unterschied zu den Seitensprossen, die schon am Sproßvegetationspunkt angelegt werden und als oberflächennahe Meristeme in den Blattachseln ruhen, bis sie austreiben. Die Seitenwurzeln entstehen im Inneren der Mutterwurzel aus dem Perizykel. Dieses Lateralmeristem baut durch lokale Teilungen ein neues Apikalmeristem auf, den Vegetationspunkt der Seitenwurzel. Die Anlagen durchbrechen dann durch Streckung die Rinde ihrer Mutterwurzel, wobei auch Mazerationsvorgänge eine Rolle spielen. Bei den Gymnospermen, den dikotylen Holzgewächsen und einigen dikotylen Kräutern findet in der Wurzel sekundäres Dickenwachstum statt. Voraussetzung ist die Bildung eines Kambiumringes. Im Stamm gibt es schon im primären Zustand ein Fascicularkambium; in der Wurzel ist dies anders. Hier entwickelt sich erst bei Beginn des Dickenwachstum ein Kambium, und zwar aus den parenchymatischen Zellen zwischen Xylem und Phloem des radiären Gefäßbündels. Über den Xylemstrahlen stößt dieses Meristem auf den Perizykel, dessen Zellen in das Kambium einbezogen werden. So entsteht ein geschlossener Kambiumring, der im Querschnitt sternförmig ist. Wie im Stamm bildet das Kambium auch in der Wurzel nach innen Holz und nach außen Bast. Durch verstärkte Teilungen in den Buchten des "Kambiumsternes" entsteht ein runder Holzzylinder, der von einem Kambiumring umgeben ist. Auch hier gibt es Fusiforminitialen und Markstrahlinitialen, und das Holz der Wurzel bildet (sekundäre) Markstrahlen. Wurzelholz ist von Stammholz selbst mikroskopisch oft schwer zu 108 109 unterscheiden. Wenn man einen ganzen Wurzelquerschnitt zur Verfügung hat, sieht man im Zentrum die Xylemstränge des radiären Bündels. Außen ist die sekundäre Wurzel von Periderm bedeckt. Dieses entsteht jedoch nicht, wie im Stamm, an der Peripherie. Das erste Periderm der Wurzel bildet vielmehr meist der Perizykel, der als Phellogen dient und nach außen Korkzellen abgibt. Die gesamte Wurzelrinde von der Exodermis bis zur Endodermis wird dabei abgestoßen. Tiefenperiderme schneiden dann in den Bast ein und bilden eine Borke. Wurzelsysteme Wir wissen bereits, wie sich die Wurzel der Pflanze verzweigt. Die Verzweigungen sind nicht auf die Hauptwurzel beschränkt, sondern auch die Seitenwurzeln verzweigen sich erneut, so daß schließlich ein Wurzelsystem von beträchtlichem Umfang entstehen kann. Addiert man Längen und Oberflächen sämtlicher Wurzeln, so kommt man zu überraschend hohen Werten; so ist zum Beispiel die Länge der Wurzeln einer einzeln stehenden Weizenpflanze auf 80 km berechnet worden. Das ist natürlich weit mehr als die Gesamtlänge der oberirdischen Teile, doch muß man bedenken, daß diese weit mehr Masse besitzen. Das zeigt sich auch im Oberflächenverhältnis: beim Roggen ist die Oberfläche des Wurzelsystems 130 mal so groß wie die der Sproßteile. Der zarte Bau der Wurzeln ist nur dadurch möglich, daß sie sich nicht selbst tragen müssen. Durch die zentralen Festigungszellen im radiären Gefäßbündel widerstehen sie der Zugbelastung und verankern den Pflanzenkörper wirksam im Boden. Bei Gymnospermen und Dikotylen geht das Wurzelsystem auf die Radicula des Embryos zurück, die die Primärwurzel oder Hauptwurzel bildet und sich reichlich verzweigt. Es entsteht ein allorhizes Wurzelsystem. Ein allorhizes Wurzelsystem, bei dem die Hauptwurzel länger und deutlich dicker als die Seitenwurzeln ist, nennt man ein Pfahlwurzel-System. Die Pflanzen nennt man auch Tiefwurzler, da die Hauptwurzel senkrecht nach unten wächst und sehr lang werden kann. Hierher gehören Kren, Löwenzahn und Zichorie unter den Kräutern, Eiche, Kiefer und Tanne unter den Bäumen. Bei den Flachwurzlern bleibt dagegen die Hauptwurzel bald in ihrem Wachstum zurück; mehrere flach streichende Seitenwurzeln übernehmen die 109 110 Führung. Hierher gehören der Erdapfel unter den Krautigen, die Fichte und die Pappelarten unter den Bäumen. Eine dritte Bewurzelungsform stellt das Herzwurzel-System dar. Hier gibt es keine führende Pfahlwurzel, aber ebenso fehlen die flach streichenden Seitenwurzeln. Vielmehr krümmen sich starke Seitenwurzeln bogig in die Tiefe. Dieser Typ ist von Erle (mit sehr tiefem System), Lärche, Birke, Linde und den Kernobstarten verwirklicht. Das Wurzelsystem der Monokotylen ist ganz anders gebaut: Es ist homorhiz. Die Keimwurzel stirbt früh ab oder bleibt sehr kurz und unverzweigt. Sie wird durch sprossbürtige Wurzeln ersetzt, die aus dem Hypokotyl und darüber liegenden Internodien oder Nodien (bei Mais 6 - 8 Knoten) hervorgehen. Das System besteht daher aus einer großen Zahl etwa gleichstarker Wurzeln, die bogenförmig in den Erdboden eindringen. Sie werden als Büschelwurzeln bezeichnet. Sprossbürtige Wurzeln finden sich nicht nur bei den Monokotylen. Viele Dikotyle bilden regelmäßig, zusätzlich zu ihrem allorhizen Wurzelsystem, sprossbürtige Wurzeln. Beispiele sind die Bodenspross- und Haftwurzeln des Efeus oder die Luftwurzeln vieler Tropenpflanzen. Auch an unterirdischen Sprossen (Rhizomen) und Ausläufern (etwa bei Erdbeere und Brombeere) entstehen regelmäßig Wurzeln. Die Ausbildung des Wurzelsystems hat größte Bedeutung zunächst für die Wasser- und Nährstoffversorgung der Pflanze: Arten, die in verschiedene Bodenschichten vordringen, werden die Vorräte insgesamt besser ausnützen und sich dabei weniger konkurrenzieren, als solche, die im gleichen Bestand denselben Bewurzelungstyp zeigen. Das ist einer der Vorteile von Mischbeständen, wie Wiesen oder naturnahen Wäldern, gegenüber Monokulturen, wie Getreidefeldern oder Fichtenforsten. So hat ein FichtenBuchen - Mischbestand eine höhere Gesamtwurzelzahl als Reinbestände der beiden Arten. Flachwurzler können kurze Niederschläge ausnützen, Tiefwurzler das absinkende Wasser nach langen Regenperioden verwerten. Tiefwurzler können so den Wasserspiegel absenken und Staunässe im Boden vermeiden helfen, wie sie auf ungeeigneten Standorten mit Fichtenmonokultur auftreten kann. Man hat sogar nachgewiesen, daß Tiefwurzler Wasser an die oberen Bodenschichten abgeben, von dem Flachwurzler in Trockenperioden profitieren können (Luzerne - Mais, "Intercropping"). Für Verbauungszwecke ist das 110 111 Verhalten der Wurzeln auch für den Kulturtechniker interessant; leider ist das Spezialwissen über diese Fragen noch nicht lückenlos; immerhin kann man öfters beobachten, dass Wurzeln Steine sprengen können. Die Forstwirte wissen viel über Waldbäume, da hier die praktische Bedeutung bald erkannt wurde. So leidet etwa die Fichte als Flachwurzler im Reinbestand häufig unter Windwurf, wobei der ganze Wurzelteller von stärkeren Stürmen ausgehoben wird. Die Tanne und die Föhre sind als Tiefwurzler viel weniger anfällig. Wir wissen, dass die Hauptachse von Bäumen aufrecht wächst, während sich die Seitenachsen unter einem Winkel einstellen. Ebenso gibt es Wachstumsunterschiede zwischen Haupt- und Nebenwurzeln: Die einen wachsen senkrecht nach unten, die anderen stellen sich unter einem Winkel zur Senkrechten ein. Es läßt sich nachweisen, daß hier tatsächlich die Schwerkraft das Wachstum der Pflanzenorgane steuert. Man bezeichnet diese richtende Wirkung als Gravitropismus (Geotropismus). Die Einstellung von Organen in Richtung auf den Erdradius bezeichnet man als Orthotropismus. Hauptwurzeln reagieren meist positiv, sie wachsen zum Erdmittelpunkt; Hauptsprosse sind negativ gravitrop. Bildet die Längsachse der Organe einen Winkel mit der Lotlinie (Seitentriebe, Seitenwurzeln 1. Ordnung), so spricht man vom Plagiotropismus. "Agravitrop", also von der Schwerkraft nicht beeinflußt, sind Seitenwurzeln 2. oder höherer Ordnungen und Saugwurzeln; sie reagieren “hygrotropisch“ auf den Wassergehalt. Man kann die Schwerkraft im Experiment ausschalten (im Weltraum) oder durch Zentrifugalkräfte überlagern; das beweist, daß hier eine Massenbeschleunigung wahrgenommen wird. Dafür sind natürlich Rezeptoren nötig. Die gängige Hypothese sagt, dass Amyloplasten mit Stärkekörnern im Zentrum der Wurzelhaube sich unter der Wirkung der Schwerkraft verlagern und Druck auf geeignete Rezeptoren am ER ausüben. Dies löst in der Streckungszone einseitige Streckungen aus, die das Organ in die vorprogrammierte Richtung krümmen. Unklar ist, wie die Information aus der Kalyptra die Streckungszone erreicht. Wuchsstoffe sind daran beteiligt, daß Wurzelbildung an ungewöhnlichen Orten ausgelöst werden kann. So entstehen Wurzeln auch an Blättern und Stammstücken nach Verletzung, und zwar entweder sofort oder erst nach Hormonbehandlung. Man spricht dann von Adventivwurzeln und kann diese zur vegetativen Vermehrung durch Stecklinge nützen. 111 112 Stämme und Blätter bilden also Wurzeln. Andererseits kann aber auch die Wurzel neue Sprosse bilden. Sie entstehen wie die Seitenwurzeln aus dem Perizykel. Ein neu entstehendes Sprossmeristem baut eine Knospe und dann einen beblätterten Sproß auf, der die Erdoberfläche durchbricht und zu einer jungen Pflanze heranwächst. Diese Wurzelbrut findet sich auch bei Bäumen. Pappeln und Weiden, besonders Silber- und Zitterpappel, zeichnen sich durch starke Wurzelbrut aus. Da alle Wurzelschößlinge genetisch mit dem Mutterbaum völlig identisch sind, sieht man im Auwald gruppenweise Klone, die der Forstmann je nach ihren Eigenschaften begünstigen oder zurückdrängen kann. Andere Arten bilden Wurzelbrut reichlich bei Verletzung der Wurzeln aus. Sind dies “Unkräuter“, wie die Ackerwinde oder die Kratzdistel, dann werden sie durch die Wurzelbrut sehr schwer bekämpfbar. Die Metamorphosen Die drei Pflanzenorgane sind sehr flexibel konstruiert, sie können sich an besondere Lebensweisen oder Standortsfaktoren anpassen. Man nennt extreme Abweichungen im Bau Metamorphosen (Gestaltumwandlungen). Dabei können Spezialbildungen, etwa Speicherorgane oder Dornen, verblüffend ähnlich werden, auch wenn sie auf verschiedene Organe zurückgehen. Sie sind dann zwar analog (funktionsgleich), aber nicht homolog (ursprungsgleich). Standortfaktoren, an die sich die Kormophyten habituell und physiologisch anpassen mussten, sind die Wasserversorgung, die Temperatur, die Einstrahlung und die Versorgung mit Mineralstoffen. Wir wollen einiges davon besprechen, so weit es in Mitteleuropa zu finden ist. A. Anpassungen an die Wasserversorgung: 1. Wasserpflanzen (Hydrophyten): Hier gibt es eine gleitende Skala der Formen. Submerse leben ganz untergetaucht, Arten mit Schwimmblättern haben Kontakt zur Atmosphäre, und amphibische Formen können an Land steigen; sie leiten zu den Sumpfpflanzen über, die nur mit ihren unteren Teilen im wassergetränkten Boden stehen. Alle echten Wasserpflanzen bilden ihre submersen Sproßteile besonders aus. Das Wasserleitungsgewebe wird reduziert, Mineralstoffe und Gase (CO2, O2) werden durch die kaum cutinisierte Epidermis aufgenommen. Die großen Interzellularen in Stamm und Blättern geben Auftrieb und erlauben aufrechten 112 113 Wuchs. Das Aerenchym ist bei Arten mit Schwimmblättern auch für die Sauerstoffversorgung wichtig. Bekanntlich ist O2 im Wasser schlecht löslich, luftgesättigtes Wasser hat nur etwa 3% der Atmosphärenkonzentration. Sauerstoff wird zu den Wurzeln tief unter der Wasseroberfläche besser in der Gasphase geleitet; das erforderte aber große Interzellularen. Manche Arten haben jedoch eine radikal andere Lösung gewählt: Sie wurden sekundär aus bewurzelten Pflanzen zu Schwimmpflanzen. Die Gattung Lemna bildet noch eine verkümmerte Wurzel aus, während sie bei der kleinsten Blütenpflanze, Wolffia arrhiza, völlig fehlt. Die Mineralstoffaufnahme ist ganz auf die Sproßepidermis übergegangen, aber nicht sehr effizient: Massenentwicklung dieser Wasserlinsen gibt es nur in überdüngten Teichen. 2. Feuchtpflanzen (Hygrophyten): Sie müssen die Transpiration fördern, um Mineralstoffe zu den Blättern zu bringen. Wir finden also an luft- und bodenfeuchten Standorten große, dünne und saftreiche Blattspreiten mit lebenden Haaren. Die Spaltöffnungen sind oft über die Umgebung erhoben. Typische Hygrophyten finden sich im Unterwuchs des tropischen Regenwaldes. 3. Mesophyten: Das sind Pflanzen "mittlerer Standorte" mit ausgewogenem Wasserhaushalt und daher ohne besondere morphologische Anpassungen. 4. Trockenpflanzen (Xerophyten): Wir finden eine Vielzahl von Strategien der Pflanzen, um mit dauernder oder vorübergehender Trockenheit fertigzuwerden. Arten, die wenigstens zeitweise große Trockenheit des Bodens ertragen, heißen Xerophyten. Eine Reihe von Baueigenheiten gelten als Anpassung an die Trockenheit, als Xeromorphosen. Besonders xeromorph sind viele Wüsten- und Steppenarten, aber auch heimische Bewohner von offenen Felsen und Sandfluren und sogar heimische Immergrüne. Diese, etwa die Nadelhölzer, müssen ja Frostperioden im Winter ertragen, die die Wasseraufnahme aus dem Boden erschweren oder unmöglich machen. Anatomisch sind viele Xerophyten (wenn auch keineswegs alle!) durch eine Kombination von teilweise bereits bekannten Zügen charakterisiert. Hierzu gehören: derbe oder gerollte Blätter, dicke, stark cutinisierte Außenwände der Epidermis, subepidermale Sklerenchyme, Wachsüberzüge, eingesenkte Spaltöffnungen, Haarfilze aus toten Haaren. Ein wirksamer Transpirationsschutz ist die Reduktion der transpirierenden Oberflächen. Abgeschwächt und vorübergehend kann man 113 114 das oft beobachten; bei Bäumen spricht man geradezu vom "frühsommerlichen Laubfall", wenn sie in den ersten trockeneren Junitagen zahlreiche Blätter abwerfen. Das sind dann meist solche, die im Schatten des Kroneninneren wenig produktiv sind. Subtropische Savannenarten können zu Beginn der Trockenzeit alle Blätter verlieren, und auch den Blattfall der heimischen Laubbäume im Herbst kann man als günstige Reaktion auf die Frosttrocknis betrachten: Sie müssen nicht so viel Material zur Vermeidung der Austrocknung investieren wie immergrüne Blätter. In Extremfällen wird die Oberflächenreduktion zur Daueranpassung. Es entstehen blattlose Rutensträucher, etwa der mediterranen Besenginster (Spartium junceum), bei dem die grünen Zweige Photosynthese betreiben. Noch günstiger wird das Verhältnis von Oberfläche zu Volumen, wenn ein Organ stark verdickt wird und Wasser speichert. Dies sind die sukkulenten Pflanzen; sie sind durch besondere Wassergewebe charakterisiert (etwa die Agave). Blattsukkulenten gibt es auch unter unseren Fels- und Trockenrasenarten, besonders in den Gattungen Sedum (Fetthenne) und Sempervivum (Hauswurz) aus der Familie der Steinbrechgewächse. Bei starker Trockenheit sieht man, daß die älteren Blätter dieser Arten schrumpfen und schließlich absterben, während junge Blätter oft sogar noch weiter wachsen können. Das Wasser wird also in die produktivsten Pflanzenteile umgelagert. Bei den Stammsukkulenten wird der Achsenkörper als Wasserspeicher ausgebildet und ist im Extremfall fast kugelig; die Kugel hat ja die kleinste Oberfläche für ein gegebenes Volumen. Meist sind die Blätter der Stammsukkulenten stark reduziert, oder sie werden sofort abgeworfen, wenn die Trockensaison beginnt. Die bekanntesten Stammsukkulenten sind die amerikanischen Kakteen, von denen sich einige (so z.B. die Gattung Opuntia) auch ans europäische Mittelmeer und nach Australien ausgebreitet haben. In Afrika treten an ihre Stellen säulen- und kandelaberartige Euphorbiaceen und Asclepiadaceen, die manchen Kakteen zum Verwechseln ähnlich sehen. Die in verschiedenen Verwandtschaftskreisen und Kontinenten als Anpassung an extreme Trockenheit entwickelte Kaktusform ist eines der besten botanischen Beispiele für Konvergenz. Darunter versteht man gleiche morphologische Ausgestaltung miteinander nicht näher verwandter Arten. Auch physiologische Anpassungen sind vielen Sukkulenten gemeinsam; so zeigen sie oft einen 114 115 speziellen Photosyntheseweg, den man als CAM bezeichnet. Er wird später noch genauer besprochen werden. Es ist merkwürdig, daß Xerophyten auch sehr häufig Dornen ausbilden. Es mag sein, daß der spärlichere Pflanzenwuchs der Trockengebiete Schutzmittel gegen den Tierfraß erfordert. Andererseits begünstigen offenkundig Trockenheit und Starklicht die Ausbildung von Dornen durch hormonelle Umstellungen. Im Prinzip kann jedes der drei Organe zum Dorn umgewandelt werden. Wurzeldornen sind selten und finden sich nur an Luftwurzeln von Exoten. Sprossdornen gehen bei heimischen Holzgewächsen meist aus Kurztrieben hervor, so bei der Schlehe (Prunus spinosa), bei den beiden Weißdornarten (Crataegus laevigata und monogyna), beim Wildapfel (Malus silvestris) und der Wildbirne (Pyrus pyraster). Langtriebdornen hat der Kreuzdorn (Rhamnus cathartica). Außer ganzen Sprossen werden aber auch Blätter zu Dornen umgewandelt. Bei der Robinie sitzen die beiden Nebenblätter als massive Dornen neben dem zarten Blattstiel. Bei der Berberitze (Berberis vulgaris) stehen die Dornen an Langtrieben. Sie sind reduzierte Blätter, in deren Achseln noch im gleichen Jahr weichlaubige Kurztriebe austreiben. – Woran kann man Dornen von Stacheln unterscheiden? Als umgewandelte Organe enthalten sie stets Stranggewebe, während die Stachel als Emergenzen allein aus Hautgewebe und Grundgewebe bestehen. B. Anpassungen an den Lichtgewinn Licht ist für die Photosynthese der Pflanze unbedingt nötig. Im Freiland fällt während der Vegetationsperiode auf alle Standorte genug Strahlung ein, doch beschatten sich die Pflanzen gegenseitig. Nur physiologisch besonders angepaßte Schattenpflanzen ertragen Dauerbeschattung. Es ist daher oft vorteilhaft, der Beschattung durch Konkurrenten räumlich oder zeitlich auszuweichen. Das einfachste (und „ehrlichste“) Mittel ist es, rasch zu wachsen. Dazu muß aber in kurzer Zeit ein massiver und stabiler Körper aufgebaut werden. Es gibt eine freilich kleine Gruppe von Pflanzen, die diese Notwendigkeit umgehen. Einige von ihnen sitzen hoch oben auf den Ästen der Bäume und leben als Epiphyten. Das erschwert die Wasser- und Nährstoffversorgung; man findet daher in der heimischen Flora zwar Algen und Flechten auf Bäumen, Angiospermen aber fast ausschließlich in den Tropen mit ihren täglichen 115 116 Regenfällen. Epiphyten müssen mit jenen Mineralstoffen auskommen, die sie aus angewehtem Staub und abgefallenen Pflanzenresten erhalten. In unserem Klima sind unter den höheren Pflanzen ausschließlich Kletterpflanzen zu finden. Sie wurzeln im Boden und klimmen mit dünnen Stämmen an anderen Pflanzen, aber auch an Felsen und Mauern empor. Sie sparen so Material gegenüber Arten, die ihr gesamtes Gewicht selbst tragen. Das Klettern erfolgt mit verschiedenen Mechanismen. Bei den Spreizklimmern stehen Körperteile steif ab. Bei Solanum dulcamara, dem Bittersüßen Nachtschatten, sind dies Sprosse, bei der großen Sternmiere (Stellaria holostea) Blätter. Kletterrosen und Brombeeren verhaken sich mit Stacheln, also Emergenzen, das Klettlabkraut (Galium aparine) mit starren, nach unten gerichteten Borstenhaaren. Wurzelkletterer bilden sproßbürtige Haftwurzeln. Wir haben das schon beim Efeu, dem einzigen heimischen Vertreter, kennengelernt. Windepflanzen haben lange Internodien; die Triebspitzen wachsen kreisend und der gesamte Sproß umschlingt die Stütze. Die Blätter entfalten sich bei Windepflanzen verzögert; die langen Triebe müssen also nicht auch noch das Gewicht ausgewachsener Blätter tragen (Hopfen, Bohne). Rankenpflanzen schließlich haben spezielle Befestigungsorgane. Eine Ranke ist ein fadenförmiges Gebilde, das eine Stütze umwindet und damit den Sproß verankert. Die Ranken der Weingewächse sind verzweigte Sproßachsen. Und zwar wird in Abständen das Sproßende zur Ranke umgebildet; da das Sproßsystem ein Monochasium ist, also eine Seitenknospe den Sproß verlängert, wird die Ranke in eine seitliche Stellung gedrängt. Dabei umschlingen die Ranken von Vitis vinifera, dem Wein, das stützende Gerüst oder (in der Natur) die tragende Pflanze (ebenso beim Wilden Wein oder der Mauerrebe, Parthenocissus quinquefolia), während die Ranken von Parthenocissus tricuspidata, der Mauerkatze, an den Enden Haftscheiben tragen, die mit Hilfe eines Klebesekrets an Mauern oder rauhen Borken haften. Blattranken bilden zahlreiche Hülsenfrüchtler, etwa die Erbse, bei der sich die endständigen Blattfiedern zu Ranken umwandeln, oder viele Wickenarten. Manche Arten von Lathyrus (Platterbse) bilden das ganze Oberblatt zu Ranken um und betreiben Photosynthese nur mit den großen Nebenblättern. Die Waldrebe (Clematis), eine holzige Ranunculacee mit Fiederblättern, windet die Blattstiele und die Blättchenstiele der Fiedern um die 116 117 Stützen. Auch die Ranken der Kürbisgewächse haben Blattnatur. Wild wächst bei uns die Zaunrübe (Bryonia dioica), bei der man die Reizbarkeit der Ranken gut studieren kann: Junge Ranken sind ausgestreckt. Streicht man mehrmals mit einem Stück Holz darüber, dann beginnen sie sich alsbald einseitig einzurollen. Diese Reaktion auf mechanische Reize nennt man Thigmotropismus. Wurzelranken gibt es in der heimischen Flora nicht, sie kommen aber in den Tropen vor. Sie entstehen natürlich aus sprossbürtigen Wurzeln. C. Anpassungen an ungewöhnliche Ernährungsbedingungen. Höhere Pflanzen sind zum Unterschied von Bakterien, Pilzen und Tieren im typischen Fall autotroph. Sie müssen also zum Energiegewinn und zum Aufbau ihrer eigenen Körpersubstanzen kein organisches Material aus dem Stoffwechsel anderer Organismen aufnehmen. Der Gegensatz dazu ist die Heterotrophie, die in mehreren verschiedenen Ausbildungen und Formen vorkommt. Die Mannigfaltigkeit des heterotrophen Verhaltens ist besonders bei den niederen Organismen sehr groß. Manche benötigen nur Kohlenstoffverbindungen, andere sind auch auf organisch gebundenen Stickstoff angewiesen. Die extremsten Ansprüche stellen jene Organismen, die bestimmte lebenswichtige organische Substanzen nicht selbst erzeugen können. Solche Verbindungen werden Vitamine genannt; sie sind für Bakterien ebenso bekannt wie für den Menschen, wenn es sich auch um verschiedene Verbindungen handelt. Allgemein teilt man die heterotroph lebenden Pflanzen, Pilze und Bakterien in drei Gruppen ein: Saprophyten, Parasiten und Symbionten. 1. Die Saprophyten verwerten tote Teile von Organismen. Bakterielle und pilzliche Saprophyten sind als Fäulniserreger weit verbreitet. Meist scheiden sie Enzyme aus, die die Makromoleküle toter Organismen in die Bausteine, etwa Zucker oder Aminosäuren, spalten; diese werden dann durch aktiven Transport aufgenommen und genützt. Die Saprophyten haben größte Bedeutung für das Geschehen in der Biosphäre: Sie treiben den Kreislauf des Kohlenstoffs, des Stickstoffs, des Schwefels und des Phosphors. Ohne Abbau toter Organismen wäre alles CO2 längst aus der Atmosphäre verschwunden, Photosynthese wäre unmöglich und die Erde tot. Freilich erfolgt diese Remineralisierung nicht 117 118 restlos; das zeigen schon die fossilen Brennstoffe, die ja nicht abgebaute Reste von Lebewesen sind. Echte Saprophyten gibt es unter den höheren Pflanzen nicht, da diese ihre abbauenden Enzyme nicht durch die Wurzeln ausscheiden können. Hingegen gibt es bei einigen Orchideen und Pyrolaceen eine Symbiose (s.u.) zwischen der Wurzel und einem Pilz. Die Pflanze liefert dem Pilz Wirkstoffe, die er nicht synthetisieren kann, und der Pilz schließt den Humus des Waldbodens saprophytisch auf. Die von den Pilzhyphen gewonnenen Stoffe werden auch den symbiontischen Kormophyten zugeführt, die völlig chlorophyllfrei bleiben und auf diesem Umweg auch saprophytisch leben. Hierher gehören die beiden bleichen Orchideen Nestwurz (Neottia nidus-avis) und Korallenwurz (Corallorhiza trifida) und die Pyrolacee Fichtenspargel (Monotropa hypopitys). 2. Parasiten entziehen lebenden Wirten organische Verbindungen. Das tun die Krankheitserreger von Mensch, Tier und Pflanze unter den Bakterien und Pilzen. Der Wirt wird auf verschiedene Weise geschädigt. Für den Parasiten ist es optimal, wenn er dabei nicht abstirbt! Wenn der Parasit bloß begrenzte Stoffmengen entzieht (etwa durch lokale Welketoxine), ist er meist nicht besonders bedrohlich. Doch zerstört er dabei oft Gewebe des Wirtes, oder er verstopft die toten Elemente des Xylems, die für die Wasserleitung gebraucht werden. Dann kommt es, wie etwa beim Ulmensterben, zu katastrophalen Trockenschäden, die den Wirt töten. Es gibt auch parasitische Angiospermen. Man unterscheidet hier Halbschmarotzer (Hemiparasiten) und Vollschmarotzer (Holoparasiten). Halbschmarotzer passen nicht ganz in das Schema der Heterotrophie: Sie erzeugen organische Verbindungen durch Photosynthese, sind also kohlenstoffautotroph. Hingegen ersparen sie sich die Ausbildung und den Betrieb eines leistungsfähigen Wurzelsystems und entnehmen Wasser und anorganische Nährstoffe dem Xylem einer Wirtspflanze. Sie sparen also organische Substanz auf Kosten des Wirtes. Am besten sieht man dies bei Viscum album (der immergrünen Mistel) und Loranthus europaeus (der Eichenmistel oder Riemenblume). Hier sitzt der Parasit hoch auf den Ästen der Wirtspflanze und sendet flache Wurzeln in die Rinde. Von diesen treten kurze Senker in das Astholz und stellen den Anschluß an die wasserleitenden Elemente her. Die weißen oder gelben Beeren werden von Vögeln gefressen 118 119 und gelangen mit dem Kot auf andere Bäume. - Bodenbewohnende Hemiparasiten sind in der Familie der Rachenblütler oder Scrophulariaceen zu finden. Die Unterfamilie der Rhinanthoideen besteht nur aus Halbschmarotzern. Sie fallen freilich nicht auf, da sie unter der Erdoberfläche mit kurzen Wurzeln das Wurzelxylem ihrer Wirte anzapfen. Die Gattungen Rhinanthus (Klappertopf), Melampyrum (Wachtelweizen), Pedicularis (Läusekraut), Euphrasia (Augentrost) sind weit verbreitete grüne Halbparasiten; kurioserweise gibt es in dieser Unterfamilie auch einen bleichen Vollparasiten, der sich genauso verhält: die Schuppenwurz (Lathraea squamaria). Sie parasitiert auf Holzpflanzen und entwickelt sich im Frühjahr, wenn das Transpirationswasser auch große Mengen an remobilisierten Reservestoffen enthält. Vollparasiten aus anderen Familien, wie die Sommerwurz (Orobanche) und die Kleeseide (Cuscuta) bohren dagegen auch das Phloem der Wirtspflanzen an und entziehen diesem die Stoffe, die in den Siebröhren transportiert werden. 3. Symbionten leben in enger Gemeinschaft und ziehen daraus beiderseits Nutzen. Symbiosen beruhen nicht auf Verträgen unter Pflanzen, sondern auf wechselseitigem Parasitismus oder Angriff und Abwehr, wobei keiner der Gegner siegt. Sie entziehen einander Nähr- und Wirkstoffe und können auf diese Weise besser fortkommen als allein. Eine sehr innige Symbiose zwischen Organismen aus ganz verschiedenen systematischen Gruppen findet sich bei den Flechten. Hier entzieht eine Alge einem Pilzmyzel (einem Geflecht aus Pilzfäden) Wasser und anorganische Nährsalze. Der Pilz hingegen entsendet in einzelne Algenzellen kurze Fortsätze, mit denen er die Protoplasten verdaut. Die Gestalten der Flechtenkörper sind so charakteristisch, daß man sie als selbständige systematische Einheiten und nicht unter den Algen oder den Pilzen einreiht. Sogar bei den Fortpflanzung werden häufig Pilz und Alge gemeinsam verbreitet. Flechten wachsen vor allem auf offenen Extremstandorten, wie Felsen und Baumrinden (epiphytisch) und in alpinen Rasen. Pilze sind auch Partner in der Mykorrhiza, einer Symbiose mit der Wurzel einer höheren Pflanze; sie ist für sehr viele Pflanzen ernährungsphysiologisch wichtig. Ungefähr 80% der heimischen Kormophyten haben eine Mykorrhiza! Bei vielen Waldbäumen umspinnt ein Pilzmyzel die stumpfen, keulig angeschwollenen Wurzelenden. Die Pilze wachsen in den 119 120 Interzellularen der befallenen Wurzel bis zur Endodermis, dringen aber oft auch in die Zellen ein. Sie entziehen dem Wirt Photosyntheseprodukte, ersetzen ihm aber die Feinwurzeln und die Wurzelhaare, da ihre feinen Hyphen den Boden dicht durchlaufen und aufschließen. Welche Vorteile haben nun die Partner? Der Pilz profitiert von den organischen Substanzen der grünen Pflanze. Einerseits sind dies die Kohlenhydrate des Phloems, andererseits aber auch Wirkstoffe zur Fruchtkörperbildung, die der Pilz nicht synthetisieren kann. Der Gewinn der Bäume wird darin gesehen, daß der Pilz aus dem Humus anorganische Stoffe freisetzt, die er samt dem Wasser dem Baum zuleitet. Junge Zirben mit verpilzten Wurzeln enthielten bei einer Analyse in den Nadeln 86% mehr Stickstoff, 234% mehr Phosphor und 75% mehr Kalium als Pflanzen ohne Mykorrhiza. Verbesserte Ernährung und geringere Investitionen in die Wurzeln ermöglichen die Besiedlung ungünstiger Standorte; so hat man errechnet, daß die alpine Baumgrenze rund 2 - 300 m tiefer liegen würde, wenn die Bäume, die hier alle mykotroph sind, keine Pilzwurzeln hätten. Die Partner der Bäume sind meist Höhere Pilze (Basidiomyceten). Unter ihnen finden sich die Speisepilze, aber auch die bekanntesten giftigen Arten. Eine ökologisch und wirtschaftlich sehr wichtige Symbiose ist die zwischen Wurzeln und stickstoffbindenden Mikroorganismen. Höheren Pflanzen fehlen die Enzyme, die zum Einbau des Stickstoffs der Luft, also des Moleküls N2, in organische Verbindungen nötig sind. Sie sind daher auf Ammoniumsalze und Nitrate des Bodens angewiesen. Hingegen können gewisse Bakterienarten das N2 zu NH3 reduzieren, wozu sie allerdings sehr viel Atmungsenergie benötigen. Und hier setzt eine sehr profitable Symbiose ein: Die Bakterien befallen die Wurzel und entziehen ihr organische Verbindungen. Dafür erhält die Pflanze N-Verbindungen, die in den Sproß transportiert werden. Sie benötigt keinen Stickstoffdünger, und nach der Ernte erhöhen die verrottenden Wurzeln den Stickstoffgehalt des Bodens, sodaß der Anbau dieser Pflanzen den Ertrag später gebauter Kulturpflanzen verbessert. Die Leguminosen, die stets stickstoffbindende Symbionten besitzen, haben daher einen festen Platz im Fruchtwechsel. Sie werden oft auch zur Gründüngung angebaut und ohne Ernte in den Boden eingearbeitet. Als Erstsaat erleichtern sie die Begrünung von Rohböden (etwa auf frischen Wegböschungen). 120 121 Diese Symbiose entsteht zwischen Bakterien der Gattung Rhizobium und den Hülsenfrüchtlern oder Leguminosen. Die im Boden frei lebenden Bakterien dringen durch die Wurzelhaare in die Pflanze ein, indem sie die Wand des Haares enzymatisch auflösen. Ein Bakterienschlauch durchwächst die Rinde und gelangt bis zur Endodermis, die die Bakterien nicht durchdringen können. Der Befall regt die Rindenzellen zu Teilungen an; es entstehen Meristeme, die dicke Knöllchen produzieren. Die Bakterien teilen sich und befallen die Zellen des Knöllchens, wo sie von Membranen umschlossen werden und ständig Stickstoffverbindungen abgeben. Schließlich erlangt die Abwehr der Pflanze das Übergewicht und die Bakterien werden verdaut, was der Pflanze weiteren Stickstoff bringt. Und wo liegt der Vorteil der Bakterien? Nach dem Absterben der Wurzel gelangen weit mehr Bakterien in den Boden, als sich unter seinen kargen Bedingungen ohne Symbiose hätten bilden können. Das Beispiel zeigt, wie sich die Symbiose aus dem Parasitismus der Bakterien und der Abwehr der höheren Pflanze entwickelt hat. Von dem Kampfgleichgewicht profitieren beide Partner. Einige Bäume, vor allem die an Feuchtstandorten wichtigen Erlen, aber auch der Sanddorn und die Ölweide, bilden ähnliche Symbiosen mit der stickstoffbindenden Frankia alni, einer anderen Bakterienart. Auch hier entstehen an den Wurzeln Knöllchen, die bei der Erle die Größe von Tennisbällen erreichen können. Schließlich wollen wir noch die "fleischfressenden" Pflanzen (Insectivoren) erwähnen. Das sind Arten, die einen Teil ihrer Mineralstoffe aus Tieren (meist Insekten) decken, die sie mit besonderen Einrichtungen fangen und enzymatisch zerlegen. Alle diese Pflanzen kommen auf mineralstoffarmen Standorten vor, etwa bei uns in oligotrophen (nährstoffarmen) Gewässern und auf Hochmooren, oder aber im tropischen Regenwald, der nur fälschlich als üppiger Standort gilt. Auch diese Arten sind nicht heterotroph im engeren Sinn, da sie organische Substanzen durch Photosynthese aufbauen. Der Sonnentau (Drosera), eine Hochmoorpflanze, hat den Typus der Klebfallen verwirklicht. Die Pflänzchen tragen auf ihren Blättern Tentakel, Emergenzen, deren Drüsenköpfchen ein klebriges, honigduftendes Sekret absondern. Kleine Tiere werden angelockt, bleiben hängen und werden von immer mehr Tentakeln erfaßt, da diese auf die chemischen Stoffe, die das Tier 121 122 absondert, mit Krümmungen reagieren. Ähnlich ergeht es Tieren an den drüsig behaarten Blättern des Fettkrautes (Pinguicula). Die Tiere werden durch ausgeschiedene Enzyme rasch verdaut und die Substanzen der Weichteile werden absorbiert, die Chitinskelette bleiben über. Der Wasserschlauch (Utricularia), eine submerse Wasserpflanze, fängt Kleinkrebse (Daphnien) in Fangblasen. D. Anpassungen an die Temperatur I. Pflanzen heißer Zonen Man kann die Wirkung von Wassermangel und von hoher Temperatur kaum trennen, da beide über den Wasserverbrauch eng zusammenhängen. Je mehr sich das Blatt erhitzt, desto mehr Wasser verdunstet. Man weiß also oft nicht, ob morphologische Anpassungen (etwa ein weißer Haarfilz), die die Strahlungsbelastung reduzieren, eher die schädliche Überhitzung des Blattes verhindern sollen oder die große Wasserabgabe bei hoher Temperatur. //. Anpassung an die Winterkälte in gemäßigten Klimaten Der Winter ist bei uns sowohl durch die Kälte als auch durch den erschwerten Wasserhaushalt eine ungünstige Zeit. Ausdauernde Pflanzen passen sich morphologisch und physiologisch an, aber es ist oft schwer zu entscheiden, ob Abänderungen eher auf die Kälte oder auf die winterliche Trockenheit zurückzuführen sind. Im einzelnen sind die Anpassungen dieser Tropophyten recht verschieden. Mit der Physiologie der Kälteresistenz werden wir uns später beschäftigen. Morphologisch ist bedeutsam, welche Teile des Pflanzenkörpers der Kälte überhaupt ausgesetzt werden. Es macht einen Unterschied, ob die Blätter über den Winter behalten werden. Sie sind ja bei tiefen Temperaturen photosynthetisch inaktiv, transpirieren aber weiter und stellen damit eine Belastung dar. Der Neuaufbau aller Blätter im Frühling kostet freilich Energie und Material, besonders Mineralstoffe. Nadelhölzer und immergrüne Dikotyle statten ihre Blätter mit wassersparenden Einrichtungen aus und sorgen für physiologische Kälteresistenz. Auch das kostet Energie und Material. Dafür können sie diese kostspieligen Organe mehrere Jahre behalten und sind bei kurzen Vegetationszeiten, etwa im Gebirge, zu Beginn der günstigen Jahreszeit sofort zur Photosynthese bereit. Niederwüchsige Kräuter behalten oft wenigstens einige Blätter über den Winter. 122 123 Unabhängig davon, was die Blätter tun, unterscheiden sich Pflanzen darin, was aus den Stämmen und den an ihnen sitzenden Winterknospen wird. Dabei kommt es auf die Höhe über dem Boden an: Je weiter die Knospen in die Luft gestreckt werden, desto stärker sind sie wasserentziehenden, eisigen Winden ausgesetzt, desto weniger kann die gut isolierende Schneedecke sie schützen. Nach dem allgemeinen Habitus und der Lage der Erneuerungsknospen hat man die Kormophyten in fünf Gruppen eingeteilt. Der prozentuelle Anteil dieser Lebensformen ( die man besser Überlebensformen nennen sollte) an der Pflanzenwelt einzelner Standorte schwankt. a) Phanerophyten oder Luftpflanzen tragen ihre Erneuerungsknospen höher als 50 cm über der Oberfläche. In unserem Klima sind dies wintergrüne und winterkahle Bäume und Sträucher und die wenigen ausdauernden Kletterpflanzen wie Wein, Efeu und Waldrebe. b) Chamaephyten (Halb- und Zwergsträucher) tragen ihre Erneuerungsknospen näher am Boden, zwischen 10 und 50 cm Höhe. Das ist besonders in Regionen mit regelmäßiger, hoher Schneelage vorteilhaft: Die Knospen sind vor der extremen Kälte geschützt, besonders vor nächtlichen Strahlungsfrösten, da Schnee ein sehr schlechter Wärmeleiter ist. Ferner trocknen Pflanzenteile unter dem Schnee nicht aus. Ich möchte hier nur die Heidelbeere und die Heidekräuter (Calluna und Erica) nennen, doch gehören in den Alpen nach klassischen Untersuchungen Josias BRAUNS rund ein Viertel aller Pflanzen in diese Gruppe. Noch reichlicher vertreten sind in den Alpen, aber auch in tieferen Lagen der gemäßigten Zonen, die c) Hemikryptophyten oder Oberflächenpflanzen, deren Erneuerungsknospen dicht am Boden liegen. Sie werden schon von leichter Schneedecke oder Streu gegen den Bodenfrost geschützt. Hierher gehören die zweijährigen und ausdauernden Rosettenpflanzen, wie Löwenzahn oder Wegerich, die Stauden mit oberirdischen Ausläufern, etwa die Erdbeere, und die ohne Blätter überwinternden Schaftpflanzen, wie die Brennessel oder der Wermut; bei diesen liegen die Knospen am Grund des vertrockneten Stengels. In den Alpen gehören zu dieser Gruppe über 60% aller Pflanzenarten, allgemein in der gemäßigten Zone rund 50%. 123 124 d) Kryptophyten überdauern mit unterirdischen oder unter der Wasseroberfläche liegenden Organen und entwickeln an diesen die Erneuerungsknospen. Hierher gehören Hydrophyten (Wasserpflanzen) ebenso wie Geophyten: Rhizomgeophyten und Zwiebelgeophyten. Die unterirdischen Organe sind zum Zweck der Speicherung stark verändert. Kryptophyten sind eher selten, aber ihre wirtschaftliche Bedeutung für den Menschen ist groß. e) Die fünfte Gruppe, die Therophyten, überdauern die ungünstigste Jahreszeit als Embryonen in widerstandsfähigen Samen. Die einjährigen vegetativen Körper sterben völlig ab. Zu dieser Gruppe gehören viele unserer wichtigsten Feldfrüchte, zum Beispiel die Sommergetreide, aber auch Unkräuter, etwa der Klatschmohn, die Vogelmiere oder das gemeine Kreuzkraut. Meist sind dies Pflanzen (natürlich oder künstlich) offener Standorte. Die Therophyten sind also Einjährige (Annuelle). Dabei müssen wir Sommer- und Winterannuelle unterscheiden. An unseren Klimarhythmus angepaßte Therophyten sind nur die Sommerannuellen, die im Frühjahr keimen, sehr rasch blühen und fruchten und noch im Laufe des Sommers ihren Lebenszyklus beenden. Hierher gehören die Sommergetreide neben vielen unserer Wildpflanzen. Hingegen keimen die Winterannuellen bereits im Herbst, überdauern den Winter als Hemikryptophyten und blühen und fruchten im folgenden Jahr. Hierher gehören die Wintergetreide und verschiedene Ackerunkräuter von nahöstlicher oder mediterraner Herkunft. In diesen Gebieten ist nämlich nicht der Winter die ungünstigste Zeit, sondern der Sommer mit seiner langen Dürre. Annuelle sind offenbar in Anpassung nicht nur an die Kälte, sondern auch an lange Dürre entstanden. Für den Frühjahrsaustrieb ausdauernder Pflanzen werden Reservestoffe gebraucht; zu ihrer Speicherung sind lebende Zellen in großer Zahl nötig. Bei Phanerophyten und Chamaephyten werden dazu die lebenden Zellen im Stamm (Holz und Bast), in der Wurzel und eventuell auch in überwinternden Blättern herangezogen. Am anderen Ende der Lebensformen, bei den Therophyten, muß der Keimling mit dem auskommen, was im Samen gespeichert werden kann. Bei den Hemikryptophyten und den Kryptophyten, ausdauernden Kräutern, stehen zur Stoffspeicherung nur jene unterirdischen oder oberflächennahen oberirdischen Organe zur Verfügung, die als einzige überwintern. Um für einen raschen Wachstumsschub im nächsten Frühjahr 124 125 genug Material speichern zu können, entstehen verdickte Gebilde mit vielen Zellen. Einige dieser metamorphosierten Organe sind wichtige Nahrungsquellen des Menschen. Man unterscheidet fünf Gruppen: 1) unbegrenzt wachstumsfähige unterirdische Wurzelstöcke oder Rhizome 2) begrenzt wachsende ober- oder unterirdische Sproßknollen 3) Zwiebeln mit speichernden Blättern; 4) Wurzelknollen aus Nebenwurzeln; 5) Rüben, das sind verdickte Hauptwurzeln. 1. Wurzelstöcke oder Rhizome sind verdickte unterirdische Sprossachsen mit meist kurzen Internodien. Sie wachsen meist horizontal und können viele Jahre alt werden, da sie alljährlich apikale Knospen anlegen und im Frühjahr wieder austreiben. Alte Teile sterben im Laufe der Jahre ab; die Rhizome können sich jedoch verzweigen und so allmählich große Flächen mit ihren "Herden" von oberirdischen Sprossen bedecken. Rhizome bilden viele Kräuter, besonders Waldbodenpflanzen, wie Einbeere, Salomonsiegel, Maiglöckchen oder Bingelkraut. Rhizome tragen sprossbürtige Wurzeln und schuppenförmige Blätter. 2. Sprossknollen können oberirdisch oder unterirdisch angelegt werden. Sie bestehen entweder nur aus dem Hypokotyl oder aus einem oder mehreren Internodien des Hauptstammes oder eines Seitentriebes. a) Reine Hypokotylknollen finden sich bei der Zyklame, dem Radieschen und der roten Rübe. b) Eine oberirdische Sprossknolle aus mehreren grün beblätterten Sproßabschnitten bildet der Kohlrabi (Brassica oleracea var. gongylodes). c) Unterirdische Sprossknollen haben (wie Rhizome) Schuppenblätter. Sie sind dicker als Wurzelstöcke, meist nur einjährig und tragen keine sprossbürtigen Wurzeln. Bekanntester Vertreter ist die Kartoffel, bei der die Knollen an den Enden unterirdischer Seitenzweige, sogenannter Ausläufer, entstehen. Sie dienen der vegetativen Vermehrung: Nach Ausbildung der stärkereichen Knollen geht die Mutterpflanze zugrunde. Aus Knospen in den Vertiefungen der Knolle, den "Augen", entstehen nach einer Ruhezeit neue grüne Triebe mit sproßbürtigen Wurzeln. – Auch Topinambur bildet Sprossknollen. 125 126 3. Zwiebeln bestehen aus unterirdischen, sehr stark verkürzten Sprossachsen mit verdickten, fleischigen Speicherblättern. 4. Die Wurzelknollen ähneln äußerlich sehr den Sprossknollen. Sie sitzen an sprossbürtigen Wurzeln und unterscheiden sich von den Sprossknollen durch Wurzelhauben und das Fehlen von Schuppenblättern. Beispiele wären die Dahlie, die Feigwurz (Ranunculus ficaria) und die Batate (Ipomoea batatas), die in den Tropen die Kartoffel vertritt. Auch viele Orchideen bilden Wurzelknollen, die oft sogar verzweigt sind. 5. Rüben sind verdickte Hauptwurzeln. Es gibt sie daher nur bei den Dikotylen; die Hauptwurzel der Monokotylen stirbt ja ab. Meist ist die Rübe keine reine Hauptwurzel, sondern Stammteile beteiligen sich an ihrem Aufbau. Reine Wurzelrüben haben die Karotte und die Zuckerrübe. Bei der Futterrübe (Beta vulgaris var. crassa) und beim Rettich (Raphanus sativus) sind große Hypokotylanteile einbezogen. Bei der Sellerie (Apium graveolens) ist auch noch ein Teil des beblätterten Stammes rübenförmig verdickt. Es gibt also alle möglichen Übergänge zu den früher besprochenen reinen Hypokotylknollen (Radieschen) und Sproßknollen (Kohlrabi). Bei der Karotte (primärer Zustand) speichert das Rindenparenchym, beim Rettich (sekundärer Zustand) das Parenchym in dem ganz weich ausgebildeten und kaum verholzten Holz. 126