C4-Pflanzen
Werbung

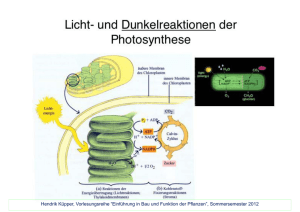
C4-Pflanzen aus Wikipedia, der freien Enzyklopädie Wechseln zu: Navigation, Suche Zu den C4-Pflanzen werden Pflanzen gezählt, bei denen sich im Vergleich zu den C3-Pflanzen ein weiterer Weg zur Kohlenstoffdioxid-Fixierung entwickelt hat. Da das erste fassbare Photosyntheseprodukt ein C4-Körper ist, das Oxalacetat, wurde die Bezeichnung zur Unterscheidung von den Pflanzen mit "normalem" Photosynthesestoffwechsel gewählt. Bekannte C4-Pflanzen sind Amarant, Hirse, Mais, Zuckerrohr, und Chinaschilf. C4-Pflanzen Mit dem Aufkommen der oxygenen Photosynthese vor ca. 1,5 Milliarden Jahren reicherte sich Sauerstoff in der Atmosphäre an. Das Kohlenstoffdioxid-fixierende Enzym RubisCO katalysiert in einer – allerdings sehr signifikanten – Nebenreaktion auch die Fixierung von Sauerstoff. Nun konnte die Assimilation nicht mehr so problemlos ablaufen, wie in der ursprünglichen sauerstoffarmen Atmosphäre, da durch die genannte Nebenreaktion im Stoffwechsel C2-Körper anfielen. Die Pflanze betreibt einen erheblichen Aufwand, um diese C2-Körper zurückzugewinnen. Da bei diesem Vorgang Sauerstoff verbraucht wird, nennt man ihn Photorespiration. Da die Affinität der RubisCO gegenüber Sauerstoff bei steigender Temperatur stärker zunimmt als für Kohlenstoffdioxid, ist dies besonders problematisch für Pflanzen in heißen und trockenen Gebieten, von denen einige einen alternativen Mechanismen zur Kohlenstoffdioxid-Fixierung entwickelten. Hier entstand eine "ATP-getriebene CO2-Pumpe", die einen Mikrokosmos herstellt, welcher jenem der Uratmosphäre entspricht. Zusätzlich zur Rubisco-Reaktion entwickelte sich in den Mesophyllzellen der PEP-Carboxylase-Weg zur Kohlenstoffdioxid-Fixierung. Dabei wird CO2 an Phosphoenolpyruvat – kurz PEP – fixiert, wobei Oxalacetat – kurz OA – entsteht, welches vier Kohlenstoffatome enthält, wovon sich analog zu den C3-Pflanzen der Name dieses Pflanzentyps ableitet. Das Oxalacetat gelangt über eine Malatschleuse in die Leitbündelscheiden, wo es durch das Malatenzym in Pyruvat – kurz Pyr – und CO2 gespalten wird. Dadurch entsteht dort eine erhöhte CO2-Konzentration, wodurch der reguläre Calvinzyklus effizienter abläuft. Der gesamte Weg wurde als zyklischer Prozess formuliert: Hatch-Slack-Zyklus Die Reaktionen dieses C4-Dicarboxylatweges, die sich über zwei benachbarte Zelltypen erstrecken, wurde in den 60er Jahren des 20. Jahrhunderts ergründet und nach seinen Entdeckern Marshall Davidson Hatch und Charles Roger Slack benannt. Nach seinem Beginn in den Mesophyllzellen, die nicht über RubisCO verfügen, setzt er sich, wie beschrieben, in den Leitbündelscheidenzellen fort, die aufgrund der Enzymausstattung von C3-Pflanzen den Calvin-Zyklus ausführen können, Abbildung: Verbindung der Hatch-Slack- und Calvin-Zyklen in C4-Pflanzen CAM-Pflanzen Eine weitere Optimierung des Prinzips der ATP-getriebenen CO2-Pumpe findet sich im CAM- Pflanzen (CAM = Crassulaceae Acid Metabolism). An trockenen Standorten bewerkstelligen Sukkulente statt der örtlichen eine zeitliche Trennung der CO2-Fixierungsschritte. So müssen die Spaltöffnungen (Stomata) zur CO2-Aufnahme nur nachts geöffnet werden, was eine Austrocknung verhindert. Der Tag (T)-Nacht (N)-Rhythmus gestaltet sich dabei wie folgt: Nachts: CO2-Aufnahme und Fixierung an PEP. Entstehendes Oxalacetat (OA) wird zu Malat reduziert und in Malatspeichern (Zellvakuolen) zwischengelagert. Der Prozess ist von einer Ansäurung begleitet und nutzt die tagsüber in der Lichtreaktion gebildeten Reduktionsäquivalente. Tags: die in der Zellvakuole deponierte Säure (überwiegend Äpfelsäure, aber auch Asparaginsäure) wird decarboxyliert. Freigesetztes CO2 treibt den Calvinzyklus an. Bemerkenswert ist die Regenerierung von PEP aus Pyr, die zwei ATP-Äquivalente benötigt. In Pflanzen wird hier nicht die Zweischritt-Strategie der Gluconeogenese genutzt. Vielmehr entstand mit Pyruvat-Phosphat-Dikinase ein Enzym, das zwei energiereiche Phosphatbindungen eines Moleküls ATP und ein anorganisches Phosphat-Ion zu diesem Zweck nutzen kann. Abbildung: Kohlendioxid-Fixierung in C4 - und CAM-Pflanzen: Dieser Pflanzentyp nutzt zur Kohlendioxid-Fixierung Phosphoenolpyruvat (PEP; Kasten am linken Rand). Es entsteht die C4Verbindung Oxalacetat (OA). Bei deren Decarboxylierung erhöht sich der CO2-Partialdruck, so dass der Calvin-Zyklus (unten rechts eingezeichnet) effizient betrieben werden kann. Die Regenerierung von PEP erfolgt durch Pyruvat-Phosphat-Dikinase in einem Schritt. CAM-Pflanzen haben diese Reaktionen einem Tag-Nacht(T/N)-Rhythmus unterworfen. Tagsüber (T) wird die Lichtreaktion der Photosynthese betrieben, womit Energieäquivalente zur Synthese von Stärke bereitgestellt werden. Nachts (N)entsteht aus Stärke der Akzeptor PEP, daraus OA, das zu Mal umgesetzt und zwischengelagert wird. Ökonomische und ökologische Aspekte C4-Pflanzen sind den meisten C3-Pflanzen insofern überlegen, als sie CO2 und Wasser ökonomischer nutzen können: (WUE water-use-efficiency) Während C4-Pflanzen zur Bildung von 1 g Trockenmasse 230–250 ml Wasser benötigen, liegt der Bedarf für C3-Pflanzen zwei bis dreimal so hoch. Überlegene Wachstumsraten zeigt das Chinaschilf, das aus diesem Grunde Interesse für die Produktion von Biomasse für die Energiegewinnung gefunden hat: Unter günstigen Voraussetzungen lassen sich bis zu 45 Tonnen Trockenmasse je Hektar ernten. Die optimale Wachstumstemperatur liegt zwischen 30 °C und 45 °C, für C3-Pflanzen dagegen bei 15–25 °C. Wachsende Aufmerksamkeit gewinnen auch tropische C4-Futtergräser, die mit Stickstoff bindenden Bakterien vergesellschaftet sind und somit kaum einer Zusatzdüngung bedürfen. Obwohl zu den Gräsern gehörend, ist Reis keine C4-Pflanze. Um die Erträge zu steigern, gab man ihm die Genausstattung vom Mais und verzeichnete Ertragssteigerungen von bis zu 35 Prozent. In den letzten 30 Jahren ist eine Ausbreitung von C4-Pflanzen auch auf warmen, sonnigen Standorten in Mitteleuropa zu beobachten. Zumeist handelt es sich um hirseartige Gräser und Fuchsschwanzarten. Deren Ausbreitung wird zumindest bisher nicht als Gefahr für die heimische Flora gewertet. Isotopendiskriminierung C4-Pflanzen lassen sich durch das Verhältnis der beiden Kohlenstoff-Isotope 12C und 13C erkennen. Die beiden Isotope kommen in der Atmosphäre mit 98,89 % und 1,11 % vor (das radioaktive Isotop 14C spielt in diesem Zusammenhang keine Rolle). Das Enzym RubisCO reagiert mit 12C schneller als mit 13C, bei C3-Pflanzen ist daher das 13C abgereichert. Das Verhältnis 13C/12C wird als δ-13C-Wert ausgedrückt: Als Standard ist ein bestimmtes Kalkgestein definiert. Produkte der C3-Photosynthese besitzen δ13C-Werte von rund –28 ‰. In den C4-Pflanze wird jedoch fast das gesamte CO durch die PEP2 Carboxylase vorfixiert, die die Kohlenstoffisotope nicht diskriminiert. Durch die hohe interne CO2Konzentration in den Bündelscheidenzellen kommt auch die Diskriminierung der RubisCO nicht zum Tragen. Daraus ergibt sich für C4-Pflanzen ein δ-13C-Wert von durchschnittlich –14 ‰. Durch Bestimmung des δ-13C-Wertes mittels Massenspektrometrie kann man daher unterscheiden, ob Zucker aus der Zuckerrübe (C3) oder aus Zuckerrohr (C4) stammt.