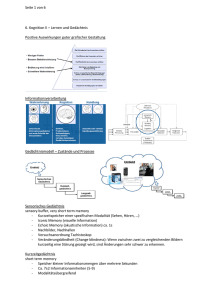
Der Einfluss von psychosozialem Stress und Cortisol auf das
Werbung

Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis Inaugural-Dissertation zur Erlangung des Grades eines Doktors der Naturwissenschaften in der Fakultät für Psychologie der Ruhr-Universität Bochum Vorgelegt von Dipl.- Psych. Diana Preuß Gedruckt mit Genehmigung der Fakultät für Psychologie der RUHR-UNIVERSITÄT BOCHUM Referent: Prof. Oliver T.Wolf Korreferent: PD Dr. Boris Suchan Tag der mündlichen Prüfung: 20.11.2009 DANKSAGUNG Mein Dank gilt allen, die mich während meiner Promotion begleitet und mich direkt oder indirekt unterstützt haben. Er gilt den Hilfskräften, die bei der Durchführung der Studien die entscheidende Rolle eingenommen haben und gestresste Versuchspersonen sichergestellt haben. All denjenigen, die bereitwillig an den Studien teilgenommen haben danke ich ebenfalls für die wertvollen Daten, die ich dadurch erhalten habe. Mein Dank geht an meinen Betreuer und Mentor Prof. Dr. Oliver T. Wolf, der mir stets mit Rat und Tat zur Seite stand und bereits meine Diplomarbeit betreut hat. Auch während meiner Promotion hat er mich unterstützt und gefördert und so die Arbeit erst möglich gemacht. Herzlichen Dank für die langjährige Begleitung! Frau Prof. Dr. Irene Daum danke ich herzlich, dass Sie sich trotz weiterer Verpflichtungen bereit erklärt hat meine Arbeit als Koreferentin zu begutachten. Weiterhin danke ich meinen Kollegen Serkan Het und Daniela Schoofs für die abwechslungsreiche und lehrreiche Zeit. Sie hatten stets offene Ohren und Augen sowie aufmunternde Worte, für die ich mich herzlich bedanke. Vielen Dank, dass ihr seid wie ihr seid! Mein inniger Dank gilt an dieser Stelle meiner Familie. Sie hat mich stets unterstützt, aufgebaut, angetrieben, hinterfragt, gefördert und mir den Rücken freigehalten. Ich danke meinen Eltern für die bedingungslose Unterstützung und Hilfe bei all meinen Vorhaben, die ich alleine nicht hätte bewältigen können. So habt ihr mir Kraft für meine Promotion gegeben. Ich danke meinem Bruder für seine klugen Worte und Fragen, die mir mehr als einmal neue Motivation und Antrieb gegeben haben. Und ich danke Ulrich Enke, meinem Partner und Gefährten auf allen Wegen des Lebens. Ich danke ihm für die Pausen, die ich dank ihm machen konnte und für das Ziel, das ich stets vor Augen hatte und habe. Ich DANKE euch! I N H AL T S Ü B E R S I C H T 1 Einleitung ...........................................................................................................1 2 Theoretische Einführung ..................................................................................3 2.1 Das Gedächtnis ........................................................................................3 2.1.1 2.2 2.1.1.1 Die zeitliche Aufteilung des Gedächtnisses ..............................................................3 2.1.1.2 Die inhaltliche Aufteilung des Gedächtnisses ...........................................................4 2.1.2 Gedächtnisphasen ............................................................................................ 5 2.1.3 Gedächtnisstrukturen ........................................................................................ 7 2.1.4 Das emotionale Gedächtnis .............................................................................. 9 Stress .....................................................................................................14 2.2.1 2.4 3 Stresskonzepte ............................................................................................... 14 2.2.1.1 Psychologisch orientierte Stresskonzepte ..............................................................14 2.2.1.2 Biologisch orientierte Stresskonzepte .....................................................................16 2.2.2 2.3 Gedächtnistheorien ........................................................................................... 3 Die physiologische Stressreaktion .................................................................. 19 2.2.2.1 Das sympathische Nervensystem...........................................................................20 2.2.2.2 Die Hypothalamus-Hypophysen-Nebennierenrinden-Achse...................................20 2.2.3 Stressoren ....................................................................................................... 23 2.2.4 Cortisoleffekte auf das Gehirn ........................................................................ 26 Der Einfluss von Stress auf das Gedächtnis...........................................30 2.3.1 Stresseffekte auf die Enkodierung .................................................................. 31 2.3.2 Stresseffekte auf die Konsolidierung .............................................................. 35 Herleitung der Fragestellung ..................................................................41 Studie 1: Associations between endogenous cortisol levels and memory in young women: influence of encoding instructions ..................................45 4 Studie 2: Post-learning psychosocial stress enhances consolidation of neutral stimuli ..................................................................................................55 5 Studie 3: The stressed student: Influence of written exams and oral presentations on salivary cortisol concentrations in university students .65 6 Diskussion und Ausblick ................................................................................87 6.1 Zusammenfassung der Studien..............................................................87 6.2 Diskussion der Ergebnisse .....................................................................89 6.2.1 6.2.1.1 Die hormonelle Stressreaktion ........................................................................ 89 Geschlechtsunterschiede........................................................................................91 6.2.2 Die Zusammenhänge zwischen Cortisol, Stress und freier Abrufaufgabe ..... 93 6.2.3 Die Cortisoleffekte auf hippocampusunabhängige Gedächtnisleistungen ..... 97 6.2.4 Der Einfluss des Geschlechts auf die Gedächtnisleistung ............................. 98 6.3 Kritische Betrachtung der Studien ..........................................................99 6.4 Ausblick ................................................................................................101 Literaturverzeichnis...............................................................................................104 Abkürzungsverzeichnis .............................................................................................I Abbildungsverzeichnis .............................................................................................II Tabellenverzeichnis..................................................................................................III Curriculum Vitae ...................................................................................................... IV Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 1 1 E I N LE I T U N G Der menschliche Organismus zeichnet sich durch die Fähigkeit aus adaptiv auf Veränderungen und Bedrohungen zu reagieren, um so ein bestmögliches Funktionsniveau sicherzustellen. In seiner Umwelt trifft jeder Mensch auf verschiedenste Einflüsse, die dieses optimale Funktionsniveau gefährden. Stress ist einer dieser Einflussfaktoren, der sowohl im Alltag, als auch im wissenschaftlichen Bereich viel Aufmerksamkeit erhalten hat. Neben Effekten auf den Organismus, nimmt Stress auch Einfluss auf verschiedene kognitive Prozesse und Funktionen. So beeinflusst er auch das Gedächtnis. Da sich das Gedächtnis durch verschiedene Systeme, Strukturen, Phasen und Funktionen definiert, stellt der Einfluss von Stress auf die Gedächtnisleistung keinen einfachen und eindeutigen Prozess dar. Vielmehr wird der Zusammenhang zwischen Stress und den damit einhergehenden Veränderungen im hormonellen Haushalt durch mehrere Faktoren moduliert. Da sich der Zusammenhang somit als sehr komplex darstellt, steht die experimentelle Forschung vor der Herausforderung die Bedingungen und Mechanismen der Stresseffekt auf das Gedächtnis zu identifizieren und zu beschreiben. Zu diesem Zweck wurde die vorliegende Arbeit durchgeführt. Die für die vorliegende Promotionsarbeit durchgeführten Studien beschäftigen sich zum einen mit dem Einfluss von Stress und Stresshormonen auf das emotionale Gedächtnis. Dazu wurde eine erste Studie durchgeführt, in der die Zusammenhänge zwischen basalen Cortisolspiegeln auf die Gedächtnisleistung für emotional erregende und neutrale Bilder betrachtet wurde. Zudem wurde der Einfluss von Enkodierungsstrategien auf diese Zusammenhänge untersucht. In dieser Studie zeigte sich ein positiver Zusammenhang zwischen basalen Cortisolspiegeln und der Gedächtnisleistung für emotional erregende Bilder in einem 24 Stunden verzögerten freien Gedächtnisabruf. Dieser Zusammenhang zeigte sich allerdings nur für eine intentionale Enkodierung. In der zweiten Studie wurde darauf aufbauen untersucht, wie sich ein psychosozialer Stressor, der nach der Enkodierung der Bilder eingesetzt wurde, auf die Konsolidierung der Bilder auswirkt. Zusätzlich wurde der Einfluss des Geschlechts der Versuchspersonen betrachtet. Hier zeigte sich ein positiver Effekt des Stressors auf die Konsolidierung der neutralen Bilder, auf die emotional erregenden positiven und negativen Bilder zeigte sich kein Effekt. Bei einer für die Geschlechter getrennten Ana- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 2 lyse zeite sich, dass der Effekt nur bei den Männern zu beobachten war. Für beide Geschlechter zeigte sich jedoch ein positiver korrelativer Zusammenhang zwischen der Cortisolantwort und der Gedächtnisleistung für neutrale Bilder. Für die negativen Bilder zeigte sich hier ein Trend für die Gesamtstichprobe. Die dritte Studie wurde durchgeführt, um die hormonelle Stressreaktion bei naturlistischen Stressoren außerhalb des Laborkontextes zu untersuchen. Hier wurden zwei universitäte Stresssituationen betrachtet, einmal eine schriftliche Klausur und zum anderen eine Referatssituation. Zusätzlich wurde für die schriftliche Klausur beobachtet, ob eine antizipatorische Stressreaktion zu beobachten ist. Es zeigte sich für die schriftliche Klausur, dass einen Tag vor der Prüfung bereits erhöhte Cortisolwerte zu finden sind, jedoch nur bei Probanden, die die Prüfung bereits durchlaufen haben. Am Prüfungstag selber zeigten sich höhere Cortisolwerte vor der Prüfung, als an einem Kontrolltag. Nach der Prüfung fand sich kein Unterschied zur Kontrollsituation. Die Referatssituation führte zu einer signifikanten Cortisolausschüttung, die während des Referats weiter zunahm. Die vorliegende Arbeit zeigt so zum einen Effekte von Stress und Cortisol auf die Gedächtnisleistung und zum anderen die unterschiedliche Stressreaktion auf verschiedene naturalistische Stressoren. Für ein besseres Verständnis der Fragestellung wird im ersten Teil der Arbeit der theoretische Hintergrund dargestellt. Daran schließen sich die drei durchgeführten Untersuchungen an. Abschließend werden die Studien diskutuert und ein Ausblick für folgende Studien gegeben. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 2 3 THEORETISCHE EINFÜHRUNG 2.1 Das Gedächtnis Das menschliche Gedächtnis zeichnet sich durch die Fähigkeit aus, Informationen über einen langen Zeitraum zu speichern, zu integrieren und im richtigen Moment zum Abruf zur Verfügung zu stellen. Zahlreiche Forschungsbemühungen haben das Gedächtnis in den Fokus des Interesses genommen, um Aufbau, Strukturen und Prozesse, die mit dem Gedächtnis in Verbindung stehen, offenzulegen. Daraus haben sich Erkenntnisse zu Funktion und Struktur ergeben, die im folgenden Abschnitt der Arbeit dargestellt werden sollen. Zunächst werden zwei Ansätze zur theoretischen Einteilung des Gedächtnisses, daran anschließend die Phasen des Gedächtnisses erläutert und abschließend die mit dem Gedächtnis assoziierten Strukturen vorgestellt. 2.1.1 Gedächtnistheorien Die hier vorgestellten Gedächtnistheorien teilen das Gedächtnis in verschiedene Speicher ein und unterscheiden sich dabei in Bezug auf die Art der Einteilung. Neben der zeitlichen Einteilung des Gedächtnisses kann das Gedächtnis auch nach inhaltlichen Aspekten eingeteilt werden. 2.1.1.1 Die zeitliche Aufteilung des Gedächtnisses Ein Vertreter der zeitlichen Einteilung des Gedächtnisses ist die Theorie von Atkinson&Shiffrin (1968). Nach dieser Theorie lässt sich das Gedächtnis in drei Speicher einteilen, den sensorischen Speicher, das Kurzzeitgedächtnis und das Langzeitgedächtnis. Der sensorische Speicher stellt denjenigen Speicher dar, in dem die Informationen flüchtig gehalten werden. Informationen aus den Sinnesorganen werden für wenige Sekunden, abhängig von der Sinnesmodalität, im sensorischen Speicher behalten und zerfallen nach dieser kurzen Zeit. Nur Informationen, die genügend Aufmerksamkeit erhalten, gelangen weiter in das Kurzzeitgedächtnis (KZG). Im KZG werden die neu eingetroffenen Informationen mit Informationen aus dem Langzeitgedächtnis (LZG) verknüpft. Informationen im KZG werden 20-30 Sekunden gespeichert, sofern sie nicht aktiv aufrechterhalten werden. Eine aktive Wiederholung der Informationen führt zu einer längeren Speicherdauer. Neben der zeitlichen Begrenzung ist auch die Kapazität des KZG auf 7±2 Items begrenzt (Miller, 1994). Ein Item beschreibt dabei eine Informati- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 4 onseinheit, die aus mehreren zusammengefassten Einzelinformationen bestehen kann. So können z.B. einzelne Ziffern zu größeren Zahlen zusammengefasst werden, um die Kapazität des KZG optimaler auszunutzen. Im Langzeitgedächtnis (LZG) schließlich werden diejenigen Informationen gespeichert, die durch Elaboration in die bereits vorhandenen Gedächtnisstrukturen integriert wurden. Das LZG unterliegt im Gegensatz zum sensorischen Speicher und KZG keinen zeitlichen und kapazitätsbedingten Begrenzungen. Vor hier werden relevante Informationen zur Nutzung ins KZG übertragen. Diese zeitliche Einteilung des Gedächtnisses beschreibt verschiedene, aufeinander aufbauende und interagierende Gedächtnissysteme. Eine zweite Alternative zur Einteilung des Gedächtnisses besteht in der Aufteilung des LZG nach inhaltlichen Aspekten. 2.1.1.2 Die inhaltliche Aufteilung des Gedächtnisses Die inhaltliche Aufteilung des Gedächtnisses geht auf das Modell von Tulving (1995) zurück. Hiernach lässt sich das LZG in Systeme aufteilen, die sich inhaltlich unterscheiden. Hierbei werden das deklarative und das non-deklarative Gedächtnis unterschieden. Diese beiden Systeme lassen sich wiederum in weitere Subsysteme unterteilen. Das deklarative Gedächtnis, welches bewusste Informationen beinhaltet, umfasst das semantische und das deklarative Gedächtnis. Das semantische Gedächtnis wird auch als Wissenssystem beschrieben, da es Informationen umfasst, die sich auf Fakten und Wissen über die Welt beziehen. Dieses Wissen ist unabhängig vom Kontext des Erlernens. Im Gegensatz dazu beinhaltet das deklarative Gedächtnis kontextabhängiges Episodenwissen. Hier werden Informationen zur eigenen Person abgespeichert, die eng mit dem Ort und der Zeit des Lernens verbunden sind. Neben dem deklarativen Gedächtnis, in dem Informationen bewusst verarbeitet werden, postuliert Tulving (1995) auch ein non-deklaratives System, in dem Informationen ohne Bewusstsein verarbeitet werden. Dieses non-deklarative Gedächtnis umfasst Priming und das prozedurale Gedächtnis. Im prozeduralen Gedächtnis werden Informationen über motorische Abläufe gespeichert. Diese Abläufe sind automatisiert und laufen in der Regel unbewusst ab, sind aber auch bewusstseinsfähig. Priming schließlich beschreibt den Umstand, dass eine zuvor verarbeitete Information die Wahrscheinlich erhöht auf den Empfang derselben bzw. auf einer mit ihr verknüpften Information schneller zu reagieren. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 5 Die beiden hier vorgestellten Gedächtnistheorien unterscheiden sich bezüglich ihrer Betrachtungsweise des Gedächtnisses. Während die zeitliche Einteilung das Gedächtnis und die Gedächtnisbildung in ihren zeitlichen Verläufen beschreibt, betrachtet die inhaltliche Einteilung die Informationen, die im LZG gespeichert werden. Zeitliche und inhaltliche Aufteilung stehen daher nicht in Konkurrenz zu einander, sondern stellen sich ergänzende Betrachtungsweisen dar. Die zeitliche und inhaltliche Einteilung des Gedächtnisses sind in Abbildung 1 veranschaulicht. Abbildung 1: Schematische Einteilung des Gedächtnisses nach zeitlichen und inhaltlichen Aspekten. Nach zeitlichen Aspekten lässt sich das Gedächtnis in den sensorischen Speicher, das KZG und das LZG einteilen. Das LZG widerrum kann nach inhaltlichen Aspekten in das deklarative Gedächtnis mit den Subsystemen semantisches und deklaratives Gedächtnis sowie das non-deklarative Gedächtnis mit den Subsystemen prozedurales Gedächtnis und Priming unterteilt werden. 2.1.2 Gedächtnisphasen Aus der Darstellung der Gedächtnistheorien wurde bereits deutlich, dass es sich bei dem Gedächtnis nicht um ein allgemeines, unspezifisches Konstrukt handelt, sondern vielmehr um einen Überbegriff für verschiedene Subkonstrukte. Für das Langzeitgedächtnis lassen sich unabhängig von den verschiedenen Subsystemen drei Phasen des Gedächtnisses unterscheiden, die Enkodierung, die Konsolidierung und der Abruf (Lupien und McEwen, 1997). Der Gedächtnisprozess beginnt mit der Enkodierung der Informationen. Enkodierung bezeichnet hierbei die erste Aufnahme und Verarbeitung der neuen Informationen. In laborexperimentellen Untersuchungen stellt diese Phase die Lernphase dar, die entweder intentional oder implizit verläuft. Bei einer intentionalen Enkodierung sind sich die Versuchspersonen Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 6 darüber bewusst, dass die zu lernenden Informationen zu einem späteren Zeitpunkt abgefragt werden. Bei einer impliziten Enkodierung sind sich die Versuchspersonen über eine folgende Gedächtnistestung nicht bewusst und speichern die neuen Informationen daher nicht willentlich für einen späteren Abruf ab. Eine Schwierigkeit hierbei besteht darin sicherzustellen, dass den Probanden die Absicht der Gedächtnistestung verborgen bleibt und sie tatsächlich keine gedächtnisfördernden Strategien anwenden. Aus diesem Grund werden häufig Instruktionen verwendet, bei denen den Probanden andere Ziele der Studien vermittelt werden (Beckner, Tucker, Delville, Mohr, 2006; Cahill, Gorski, Le, 2003; Payne, Jackson, Hoscheidt, Ryan, Jacobs, Nadel, 2007). An die Enkodierung der neuen Informationen schließt sich die Konsolidierung an. In dieser Phase werden die Informationen gefestigt, mit bereits vorhandenen Strukturen verknüpft und in diese integriert. Je nachdem, wann in laborexperimentellen Gedächtnisstudien der Abruf der neu zu erlernenden Informationen stattfindet, ist die Konsolidierung unterschiedlich weit vorangeschritten. Die Messung der Gedächtnisleistung kann dabei zu unterschiedlichen Zeitpunkten durchgeführt werden, angefangen von Testungen unmittelbar im Anschluss an die Enkodierung (Wolf, Schommer, Hellhammer, McEwen, Kirschbaum, 2001) über Gedächtnistest Stunden bis wenige Tage (Buchanan, Karafin, Adolphs, 2003) nach der Enkodierung bis hin zu Wochen (Payne et al., 2007). Die dritte Phase des Gedächtnisses ist schließlich der Abruf der neu erlernten und konsolidierten Informationen. In laborexperimentellen Studien stellt dies den Gedächtnistest dar. Dabei existieren verschiedene Aufgaben zur Messung der Gedächtnisleistung. Neben freien Abrufaufgaben, bei denen die Versuchspersonen ohne Hinweise die Stimuli abrufen sollen, werden Aufgaben eingesetzt, bei denen die Versuchspersonen aus verschiedenen Antwortmöglichkeiten die richtige Alternative auswählen sollen (mulitple-choice-Aufgaben), Abrufhinweise erhalten (cued-recall-Aufgaben) oder die während der Enkodierung präsentierten Stimuli wiedererkennen sollen (Rekognitionsaufgaben). Diese Aufgaben stellen unterschiedliche Anforderungen an die Probanden und sind mit unterschiedlichen Strukturen assoziiert (Aggleton und Brown, 2006). Während der freie Abruf eine hippocampusabhängige Gedächtnisaufgabe darstellt und vergleichsweise schwierig ist, da keinerlei Hinweise und Hilfen gegeben sind, stellen cued-recall-, Rekognitions- und multiple-choice-Aufgaben hippocampusunabhängige Aufgaben dar, die im Vergleich zum freien Abruf und auf Grund der Hinweisreize leichter zu lösen sind. Je nach Fragestellung sind also unterschiedliche Enkodierungsinstruktionen, Konsolidierungsintervalle und Abrufaufgaben notwendig, um valide Informationen sicherstellen zu können. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 7 Im Zusammenhang mit dem Abruf gespeicherter Informationen wird das Konzept der Rekonsolidierung diskutiert (Dudai, 2006; Nader, Schafe, LeDoux, 2000). Informationen scheinen bei ihrem Abruf und der damit einhergehenden Reaktivierung in einen erneuten labilen Zustand zu gelangen, in dem sie für Veränderungen anfällig werden. Informationen werden somit nicht nur einmal während der Enkodierung verarbeitet und unveränderlich abgespeichert, sondern bei jeder Reaktivierung erneut konsolidiert und den veränderten Umweltbedingungen angepasst. Der Prozess der Rekonsolidierug konnte dabei für verschiedene Gedächtnissysteme und bei unterschiedlichen Aufgaben gezeigt werden. Trotz dieser weitläufigen Befunde scheint Rekonsolidierung nicht, wie die Konsolidierung der Stimuli, zwingend stattzufinden und stellt damit nicht eine erneute Konsolidierung der Stimuli, sondern einen neuen und andersartigen Prozess dar, durch den die bereits gespeicherten und reaktivierten Informationen verändert werden können, aber nicht zwangsläufig verändert werden müssen. Das Konzept der Rekonsolidierung stellt einen stark diskutierten und umstrittenen Prozess dar, über den bis dato keine einheitliche Auffassung existiert. Nichtsdestotrotz muss die Möglichkeit einer durch den Abruf verursachten Veränderung der Informationen beachtet werden, da diese Überlegung für experimentelle Forschung eine große Bedeutung hat. . Eine häufig eingesetzte Methode im Rahmen von Gedächtnisstudien ist eine Kombination aus einer direkter Abrufaufgabe im Anschluss an die Enkodierung und einer verzögerter Abrufaufgabe in zeitlichem Abstand zu der Enkodierung (Elzinga, Bakker, Bremner, 2005; Kuhlmann, Kirschbaum, Wolf, 2005). Durch den Einsatz zweier Abrufaufgaben und der Reaktivierung durch den sofortigen Abruf entsteht so die Möglichkeit einer Veränderung der ursprünglichen Gedächtnisspuren. Die Möglichkeit der Rekonsolidierung muss also in der experimentellen Gedächtnisforschung weiter verfolgt werden, um Einflüsse auf die Ergebnisse zu verhindern und verschiedene Studien vergleichbar zu machen. In diesem Abschnitt der Arbeit wurden nach verschiedenen Gedächtnistheorien die drei Phasend es Gedächtnisses und deren Bedeutung für die experimentelle Gedächtnisforschung dargestellt. Im Folgenden Kapitel sollen nun die mit dem Gedächtnis assoziierten Strukturen und Prozesse dargestellt werden. Anschließend wird auf die Besonderheit des emotionalen Gedächtnisses eingegangen 2.1.3 Gedächtnisstrukturen An der Bildung des Gedächtnisses sind verschiedene interagierende Systeme beteiligt, die im Folgenden dargestellt werden sollen. Dabei wird zum einen auf die am Gedächtnis betei- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 8 ligten Strukturen, aber auch auf grundlegende Prozesse der Gedächtnisbildung eingegangen. Von den mit dem Gedächtnis assoziierten Gehirnstrukturen hat der Hippocampus am meisten Aufmerksamkeit erhalten. Er stellt diejenige Gedächtnisstruktur dar, die besonders eng mit dem deklarativen Gedächtnis assoziiert ist und eine besondere Bedeutung für diese Gedächtnisprozesse hat. Die Bedeutung dieser Gehirnstruktur wurde bereits 1957 (Scoville und Milner, 1957) erkannt, nachdem der Patient H.M. nach einer bilateralen Lobektomie des medialen Temporallappens eine schwerwiegende anterograde Amnesie erlitt. Sowohl human-, als auch tierexperimentelle Studien konnten die Bedeutung des medialen Temporallappens für das Gedächtnis weiter spezifizieren. Dabei zeigte sich, dass eine auf den Hippocampus beschränkte Läsion bereits zu Gedächtnisdefiziten führt, was die Bedeutung anderer mediotemporaler Strukturen für das Gedächtnis einschränkt (z.B.: Zola-Morgan und Squire, 1986). Der Hippocampus scheint allerdings nicht für alle Gedächtnissysteme gleich wichtig, sondern speziell für das deklarative Gedächtnis von Bedeutung zu sein. Bei einer Schädigung des Hippocampus zeigen sich Einschränkungen im expliziten Gedächtnis, wohingegen andere Gedächtnisformen wie Priming, Konditionierung oder das prozedurale Gedächtnis nicht beeinträchtigt sind (Squire, 1992). Zur Erfassung der Gedächtnisleistung existieren verschiedene Aufgaben (siehe Kapitel 1.1.2). Diese Aufgaben sind mit unterschiedlichen Gedächtnisleistungen assoziiert, da sie unterschiedliche Gedächtnissysteme und Strukturen beanspruchen. Während Rekollektionsaufgaben, also Abrufaufgaben, bei denen sich die Probanden explizit an die Stimuli erinnern müssen, wie z.B. freie Abrufaufgaben, mit dem Hippocampus assoziiert werden, können Rekognitionsaufgaben auf zwei Arten gelöst werden. Auch hier können explizite Erinnerungsleistungen vorliegen, wenn die Probanden die Stimuli wiedererkennen und sich bewusst an sie erinnern. Eine zweite Alternative ist das Wiedererkennen auf Grund von Vertrautheit. Die Probanden erinnern sich in diesem Fall nicht bewusst an das Item, sondern erkennen es auf Grund eines Gefühls der Bekanntheit (Wissen, dass sie es bereits gesehen haben, ohne sich explizit daran zu erinnern) wieder. Mit diesen Wiedererkennensleistungen ist nicht der Hippocampus, sondern der perirhinale Cortex assoziiert (Aggleton und Brown, 2006) .Somit wird deutlich, dass den verschiedenen Gedächtnisleistungen unterschiedliche Strukturen zu Grunde liegen, was diese für unterschiedliche Einflussfaktoren zugänglich macht. Ein Mechanismus, welcher der Gedächtnisbildung zu Grunde liegt ist die synaptische Plastizität. Werden Neurone aktiviert, so kann es zu Veränderung ihrer Effizienz kommen. Lang- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 9 zeitpotenzierung (LTP) ist eine Form der synaptischen Plastizität, die als grundlegend für die Gedächtnisbildung angesehen wird. LTP stellt dabei eine schnelle und lang anhaltende Form synaptischer Plastizität dar (Bliss und Gardner-Medwin, 1973; Gustafsson und Wigstrom, 1988) und beschreibt die Fähigkeit von Neuronen auf Reizungen mit einer verstärkten und zeitlich längeren Aktivierung zu reagieren und so ihre Effizienz zu vergrößern (Lynch, 2004). LTP tritt dann auf, wenn mehrere Synapsen gleichzeitig in kooperativer Weise aktiviert werden und bleibt auf diese aktivierten Zellen beschränkt. So verstärkt LTP die Bildung von Verbindungen zwischen Synapsen, wodurch neue Assoziationen erzeugt werden (Squire, 1992). Der Hippocampus ist eine Struktur, in der LTP besonders leicht auslösbar ist, was deren Bedeutung für die Gedächtnisbildung weiter unterstreicht (Lynch, 2004; Thompson, 2005). Der Hippocampus unterstützt somit die Gedächtnisbildung, idem er die Formung neuer Assoziationen zwischen Stimuli und so die Bildung neuer Episoden ermöglicht (Squire, 1992). Neben der LTP stellt die Neurogenese, die Entstehung neuer Neurone im erwachsenen Gehirn, eine weitere gedächtnisrelevante Funktion dar. Da der Gyrus dentatus des Hippocampus eine der Strukturen ist, in denen auch im erwachsenen Gehirn noch neue Zellen entstehen (Altman und Das, 1965; Dayer, Ford, Cleaver, Yassaee, Cameron, 2003) wird eine Assoziation mit Gedächtnisfunktionen angenommen (Barnea und Nottebohm, 1994). Zum einen zeigten sich Assoziationen zwischen der Anzahl neuer Neurone und der Leistung in hippocampusabhängigen Gedächtnisaufgaben (Gould, Tanapat, Hastings, Shors, 1999). So konnten hippocampusabhängige Lernaufgaben wie Spurenkonditionierung oder räumliche Lernaufgaben die Anzahl neuer Zellen steigern (Gould, Beylin, Tanapat, Reeves, Shors, 1999; Lemaire, Koehl, Le, Abrous, 2000). Zudem konnten verschiedene Studien zeigen, dass eine Entfernung neuer Neurone die Leistung in hippocampusabhängigen Lernaufgaben verschlechtert (Madsen, Kristjansen, Bolwig, Wortwein, 2003; Raber, Fan, Matsumori, Liu, Weinstein, Fike, Liu, 2004; Raber, Rola, LeFevour, Morhardt, Curley, Mizumatsu, VandenBerg, Fike, 2004; Shors, Miesegaes, Beylin, Zhao, Rydel, Gould, 2001; Shors, Townsend, Zhao, Kozorovitskiy, Gould, 2002; Snyder, Hong, McDonald, Wojtowicz, 2005). Der Hippocampus stellt den Ausführungen folgend eine wichtige Struktur für das Gedächtnis dar. Allerdings ist seine Bedeutung auf das deklarative Gedächtnis beschränkt und ist in diesem Zusammenhang vor allem für emotional ungefärbte neutrale Informationen relevant. Auf das emotionale Gedächtnis wird im nächsten Abschnitt eingegangen. 2.1.4 Das emotionale Gedächtnis Emotionale Reize stellen eine besondere Form von Gedächtnisinhalten dar, da sie häufig besser erinnert werden, als neutrale Reize (LaBar und Cabeza, 2006). Dieser “emotional en- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 10 hancement effect“ konnte für eine Vielzahl von laborexperimentellen Stimuli wie Filmclips, Wörter oder Bilder gefunden werden (Hamann, 2001). Diese Befunde deuten darauf hin, dass emotionale Stimuli in besonderer Weise verarbeitet werden. Bei der Verarbeitung emotionaler Reize muss zwischen den Dimensionen Arousal und Valenz unterschieden werden. Während die Verarbeitung der Valenz der Stimuli (positiv vs. negativ vs. neutral) vorwiegend im prefrontalen Cortex stattfindet, wird die Dimension Arousal (erregend vs. nicht-erregend) in der Amygdala verarbeitet. Von dieser Struktur wird angenommen, dass sie maßgeblich für den „emotional enhancement effect“ verantwortlich ist (Kensinger und Corkin, 2004; LaBar und Cabeza, 2006). Evidenz für diese Annahme, dass die Amygdala eine bedeutende Rolle bei der Verarbeitung emotionaler Reize einnimmt, stammt von bildgebenden Verfahren, die eine verstärkte Aktivität der Amygdala bei der Enkodierung emotionaler Stimuli zeigen konnten. Die Stärke der Amygdalaaktivität korrelierte dabei mit der der Gedächtnisleistung für diese Bilder (Cahill, Haier, Fallon, Alkire, Tang, Keator, Wu, McGaugh, 1996; Canli, Zhao, Brewer, Gabrieli, Cahill, 2000; Hamann, Ely, Grafton, Kilts, 1999). Weitere Studien berichteten in diesem Zusammenhang von Geschlechtsunterschieden. Während Frauen eine Assoziation zwischen verbesserter Gedächtnisleistung und linkslateralisierter Aktivierung der Amygdala bei der Enkodierung emotionaler Stimuli zeigen, findet sich bei Männern dieser Zusammenhang mit einer rechtsseiten Aktivierung (Cahill, Haier, White, Fallon, Kilpatrick, Lawrence, Potkin, Alkire, 2001; Cahill, Uncapher, Kilpatrick, Alkire, Turner, 2004; Canli, Desmond, Zhao, Gabrieli, 2002). Zusätzlich konnte demonstriert werden, dass eine sympathische Aktivierung grundlegend für den „emotional enhancement effect“ ist. Adrenalin aktiviert bei der Verarbeitung emotional erregender Stimuli noradrenerge Mechanismen im basolateralen Kern der Amygdala und ermöglicht so eine verstärkte Verarbeitung emotional erregender Stimuli. Die Blockierung zentraler beta-adrenerger Rezeptoren vor der Enkodierung neutraler und emotionaler Stimuli durch die Verwendung eines Rezeptorantagonisten (Propanolol) hebt den „emotional enhancement effect“ auf, so dass keine Unterschiede zwischen neutralen und emotional erregenden Stimuli auftreten (Cahill, Prins, Weber, McGaugh, 1994; van Stegeren, Goekoop, Everaerd, Scheltens, Barkhof, Kuijer, Rombouts, 2005). Im Gegensatz dazu verbessert die Verabreichung eines beta-adrenergen Rezeptoragonisten das Gedächtnis für emotional erregende Reize (Cahill und Alkire, 2003; O'Carroll, Drysdale, Cahill, Shajahan, Ebmeier, 1999). Für diese beta-adrenerg vermittelten Effekte wird der Amygdala eine entscheidende Rolle zugesprochen, da sich bei einer Verabreichung von Propanolol gleichzeitig mit den Gedächtniseffekten eine reduzierte Aktivierung der Amygdala während der Enkodierung emotionaler Items zeigte (Strange und Dolan, 2004; van Stegeren et al., 2005). Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 11 Für die gedächtnismodulierenden Effekte von Emotionalität wird eine Interaktion zwischen Amygdala und Hippocampus angenommen. So konnten Strange und Dolan (2004) mit Hilfe von funktioneller Magnetresonanztomographie zeigen, dass die erfolgreiche Enkodierung emotional erregender Items mit einer Aktivierung der Amygdala einhergeht. Beim Abruf dieser Items zeigte sich auch eine im Vergleich zu neutralen Items verstärkte Aktivierung des Hippocampus. Der beta-adrenerge Rezepterantagonist Propanolol führte während der Enkodierung zu einem Ausbleiben der verstärkten Aktivierungen von Amygdala und Hippocampus (Strange und Dolan, 2004). Aufgrund der Befunde wird postuliert, dass die Amygdala die Bildung des LZG für emotional erregende Stimuli durch eine Modulation der hippocampalen Aktivität verbessert (Cahill und McGaugh, 1998; McGaugh, Cahill, Roozendaal, 1996; Packard und Teather, 1998; Phelps, 2004). Eine Interaktion zwischen Amygdala und Hippocampus scheint damit grundlegend für die Effekte auf das Gedächtnis für emotional erregende Reize zu sein. Betrachtet man den „emotional enhancement effect“ allerdings genauer, so stellt sich heraus, dass nicht alle Elemente eines emotional erregenden Stimuli von der Emotionalität profitieren. Vielmehr scheint es bei emotional erregenden Stimuli zu einer Aufmerksamkeitsverengung („attentional narrowing“) zu kommen, die die Aufmerksamkeit einer Person auf die zentralen Inhalte („gist“) des Reizes lenkt und die peripheren Details („details“) vernachlässigt. Nach der Easterbrook-Hypothese (Easterbrook, 1959) verursacht eine physiologische Erregung eine Reduzierung des Aufmerksamkeitbereichs und eine verringerte Sensitivität für Informationen, die sich in einer Verengung der Aufmerksamkeit auf zentrale Informationen äußert. Dies führt bei emotional erregenden Items, die mit physiologischer Erregung einhergehen, zu einer schlechteren Gedächtnisleistung für periphere Details und einer besseren Gedächtnisleistung für zentrale Informationen dieser Stimuli (Burke, Heuer, Reisberg, 1992; Heuer und Reisberg, 1990). Für diesen Effekt ist, ebenso wie für den allgemeineren „emotional enhancement effect“, die Amygdala entscheidend. So untersuchten Adolphs et al. (2001) die Bedeutung der Amygdala für Gist und Detail Informationen bei Patienten mit und ohne Amygdalaschädigung. Die Autoren konnten zeigen, dass eine bilaterale Schädigung der Amygdala zu einem reduzierten Gedächtnis für Gist und gleichzeitig zu einer verbesserten Gedächtnisleistung für die Detail Informationen von emotional erregenden Stimuli führte. Die bei gesunden Personen beobachtete Aufmerksamkeitsverengung war bei Probanden mit Amygdalaschädigung nicht vorhanden. In einer weiteren Studie konnten Buchanan et al. (2003) die zu Grunde liegenden Prozesse auf Ebene der Amygdala weiter spezifizieren. Sie zeigten, dass eine Verabreichung von Benzodiazepinen, welche durch eine Erleichterung der Aktivität von GABAA-Rezeptoren die episodische Langzeitgedächtnisleistung verschlechtern, Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 12 auch die Gedächtnisleistung für Gist Informationen beeinflussen. Die Verabreichung der Benzodiazepine führte zu einer Verschlechterung der Gedächtnisleistung für Gist Informationen von emotional erregenden Stimuli, wohingegen Detail Informationen nur gering beeinflusst wurden. Aus diesen Ergebnissen lässt sich ableiten, dass auf Ebene der Amygdala die Aktivität der GABA-Rezeptoren einen entscheidenden Einfluss auf die Gedächtnisleistung für emotional erregende Stimuli nimmt. Auch in Bezug auf die Gedächtnisleistung für Gist und Detail Information zeigen sich Geschlechtsunterschiede. Hier konnten Studien demonstrieren, dass Männer bei emotional erregenden Stimuli im Vergleich zu neutralen Stimuli eine verbesserte Gedächtnisleistung für zentrale Gist Informationen zeigen, wohingegen dies bei Frauen nicht zu beobachten war. Dieser Geschlechtsunterschied lag jedoch in dieser Studie nicht im biologischen Geschlecht, sondern vielmehr in den Geschlechterrollen, die die Versuchspersonen einnahmen (Cahill, Gorski, Belcher, Huynh, 2004). Es wird angenommen, dass dieser Unterschied auf die Lateralisationsunterschiede zurückgeht. Während die Amygdala der rechten Hemisphäre Gist Informationen verarbeitet und bei Männern bei der Enkodierung emotional erregender Informationen stärker aktiviert ist, ist die Amygdala der linken Hemisphäre an der Verarbeitung der Detail Informationen beteiligt und bei Frauen stärker mit der Enkodierung emotional erregender Stimuli assoziiert (Cahill, 2003). Damit übereinstimmend konnten Cahill et al. (2003) zeigen, dass die Blockade der beta-adrenergen Rezeptoren der Amygdala bei Männern zu einer verschlechterten Gedächtnisleistung für zentrale Informationen, nicht aber für Detail Informationen emotional erregender Stimuli führte, wohingegen bei Frauen die Gedächtnisleistung für Detailinformationen durch den beta-adrenergen Rezeptorantagonisten verschlechtert wurde, nicht aber für zentrale Gist Informationen. In Bezug auf die Gedächtnisleistung zeigen sich demnach Geschlechtsunterschiede, die zu gegenteiligen Effektmustern führen. Das Geschlecht der Versuchsperson muss daher immer berücksichtigt werden, wenn die Effekte von Emotionalität auf die Gedächtnisleistung untersucht werden sollen. In diesem Abschnitt der Arbeit wurde das Gedächtnis näher betrachtet. Dabei wurden zunächst zwei theoretische Ansätze vorgestellt und im Anschluss auf die dem Gedächtnis zu Grunde liegenden Strukturen und Prozesse eingegangen, die in Abbildung 2 dargestellt sind. Es wurde dargestellt, dass an der Verarbeitung neutraler und emotional erregender Stimuli unterschiedliche Gehirnstrukturen beteiligt sind, die in interaktiver Weise das Gedächtnis beeinflussen. Während der Hippocampus vorwiegend in die Verarbeitung neutraler Informationen involviert ist, geschieht die Verarbeitung emotional erregender Informationen über eine Interaktion zwischen Hippocampus und Amygdala. Die Amygdala scheint dabei auch zur Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 13 Aufmerksamkeitseinschränkung dazu beizutragen, so dass die Aufmerksamkeit von irrelevanten Detailinformationen abgezogen und auf relevante zentrale Informationen gelenkt wird. In diesem Zusammenhang wurden auch Geschlechtsunterschiede beobachtet. Das folgende Kapitel der Arbeit beschäftigt sich nun mit Stress. Abbildung 2: Die mit dem Gedächtnis assoziierten Hirngebiet. (aus: Markowitsch, 2002) Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 2.2 14 Stress “Everybody knows what stress is and nobody knows what it is” (Selye, 1973). Dieses Zitat von Hans Selye verdeutlicht treffend die Schwierigkeiten, die mit dem Begriff „Stress“ verbunden sind. Obwohl der Stressbegriff auch im Alltag weit verbreitet und sehr geläufig ist, fällt eine klare und eindeutige Definition des Konzepts schwer. Entsprechend kommentierten McEwen und Wingfield (2003), das Problem mit dem Wort Stress sei, dass es zu viele Bedeutungen habe. Während im allgemeinen Sprachgebrauch der Begriff Stress häufig gleichbedeutend für Situationen und Reaktionen verwendet wird (McEwen, 2000), herrscht im wissenschaftlichen Sprachgebrauch eine klare Trennung zwischen Auslösern und Reaktionen. Der Begriff „Stress“ bezeichnet im wissenschaftlichen Sprachgebrauch den resultierenden Zustand bzw. die Reaktion des Organismus, wohingegen die stressauslösenden Faktoren mit dem Begriff „Stressor“ bezeichnet werden (McEwen und Wingfield, 2003). Im Folgenden sollen deshalb die Begriffe entsprechend dieser Bedeutung verwendet werden. Zur Definition des Begriffs „Stress“ existieren verschiedene Konzepte, die im Folgenden exemplarisch dargestellt werden. Im Anschluss wird die Physiologie der körperlichen Stressreaktion näher betrachtet, sowie Auswirkungen von Stress auf Funktionen und Strukturen des Gehirns beschrieben. Abschließend werden in diesem Kapitel stressauslösende Faktoren von Laborstressoren vorgestellt. 2.2.1 Stresskonzepte Der erste Abschnitt des Kapitels soll der Darstellung verschiedener Stresskonzepte dienen und die Unterschiedlichkeit der theoretischen Ansätze veranschaulichen. Die hier exemplarisch dargestellten Stresskonzepte lassen sich in zwei Kategorien unterteilen. Zunächst werden psychologisch orientierte Konzepte anhand des transaktionalen Stresskonzepts dargestellt. Eine zweite Kategorie der Stresskonzepte stellen die biologisch orientierten Stresskonzepte dar, die im Gegensatz zu den psychologischen Konzepten den Fokus auf die körperliche Stressreaktion legen. Hierbei werden reaktionsorientierte Stresskonzepte sowie das Konzept der Homöostase vorgestellt. 2.2.1.1 Psychologisch orientierte Stresskonzepte Psychologisch orientierte Stresskonzepte betrachten Stress in Abhängigkeit von individuellen und stimulusspezifischen Aspekten. Anders als die biologischen Stresskonzepte betrachten sie dabei das Konzept Stress unabhängig von physiologischen Reaktionen. Das transaktio- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 15 nale Stresskonzept stellt ein Beispiel für psychologisch orientierte Stresskonzepte dar, da es die Bedeutung von individueller Wahrnehmung, Bewertung und Bewältigung betont. Das transaktionale Stresskonzept Dieses psychologisch orientierte Stresskonzept von Lazarus (Lazarus und Folkman, 1984) geht davon aus, dass einer Wahrnehmung von Stress eine subjektive Bewertung der Situation vorangeht. Das Auftreten eines potenziellen Stressors führt also nicht automatisch zu einem Stresserleben. Vielmehr werden diejenigen Reize als stressauslösend wahrgenommen, die in einer internen Bewertung als potenziell bedrohlich und relevant eingeschätzt werden. Die Interaktion zwischen Reiz und psychologischer Bewertung führt somit zu individuell unterschiedlichen Einschätzungen der Situation. Nur wenn diese Einschätzung negativ ausfällt und die Anforderungen der Situation die subjektiv wahrgenommenen Ressourcen überschreiten kommt es zu einem Stressempfinden. Aus diesen Überlegungen heraus entwickelte Lazarus das transaktionale Stresskonzept, wonach Stress das Resultat interner Bewertungsprozesse ist und damit eine Transaktion zwischen Reiz und Umwelt darstellt (Lazarus und Folkman, 1984). Die Bewertung einer Situation geschieht auf zwei Wegen. Zum einen findet eine primäre Bewertung der Situation statt. Hierbei wird der wahrgenommene Reiz entsprechend seiner Relevanz eingeschätzt. Reize werden entweder als positiv, irrelevant oder potenziell gefährlich eingestuft, wobei nur potenziell gefährliche Reize relevant sind. In diesem Fall findet eine Sekundärbewertung statt, in der das Individuum seine subjektiv wahrgenommenen Ressourcen zur Bewältigung der Situation abschätzt. Fällt diese Bewertung negativ aus, kommt es zu einem Stresserleben. Dieses Stresserleben führt zu einer Suche nach neuen Bewältigungsstrategien (Coping). Diese neuen Bewältigungsstrategien beeinflussen die Interaktion zwischen Person und Situation, womit eine neue Bewertung der Situation notwendig wird. Sobald der Vergleich zwischen Primär- und Sekundärbewertung, also zwischen Situation und Bewältigungsressourcen nicht mehr negativ ausfällt, endet dieser Kreislauf. Das transaktionale Stresskonzept nach Lazarus (Lazarus und Folkman, 1984) postuliert folglich individuelle Unterschiede bei der Einschätzung von Stressoren. Nur wenn Primär- und Sekundärbewertung entsprechend ausfallen, wird ein Ereignis als stressauslösend wahrgenommen. Diese Überlegung ist auch für die laborexperimentelle Stressforschung von Bedeutung, da dem transaktionalen Stresskonzept folgend nicht davon ausgegangen werden kann, dass jeder eingesetzte Stressor zu einem subjektiven Stresserleben führt. Obwohl die psychologisch orientierten Stresskonzepte die körperliche Stressreaktion als einen wichtigen Aspekt nicht betrachten, machen sie deutlich, dass Stress eine wichtige psy- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 16 chologische Facette hat und kein rein biologisches Phänomen ist. Vielmehr entsteht Stress als psychische Reaktion auf bestimmte Reize, die vom Individuum als gefährdend und ressourcenüberlastend wahrgenommen werden. Im Folgenden werden nun biologisch orientierte Stresskonzepte vorgestellt. 2.2.1.2 Biologisch orientierte Stresskonzepte Biologisch orientierte Stresskonzepte betrachten Stress aus der physiologischen Perspektive. Bei diesen Konzepten liegt der Fokus auf der körperlichen Komponente. Hierbei können zwei Ansätze unterschieden werden. Dies sind zum einem die reaktionsorientieren Stresskonzepte und zum anderen das Konzept zur Homöostase, welches ein neueres Modell darstellt. Reaktionsorientierte Stresskonzepte Die reaktionsorientierten Stresskonzepte definieren Stress nicht durch das Vorhandensein eines Stressors, sondern allein durch die körperliche Stressreaktion. Von Stress spricht man in diesem Zusammenhang ausschließlich, wenn der Organismus eine körperliche Stressreaktion zeigt. Einer der ersten Vertreter dieses Ansatzes war Walter Cannon (1914), der bei seinen Untersuchungen zu Stress den Fokus auf die sympathische Aktivierung und die damit einhergehende Ausschüttung von Adrenalin und Noradrenalin legte. Das Ziel der Stressreaktion nach Cannon ist die Bereitstellung von Energie zur Initiierung einer adäquaten Reaktion. Dabei kann der Organismus die bereitgestellte Energie nutzen, um zu kämpfen, oder um zu fliehen (die sogenannte Fight-or-Flight-Antwort). Ein weiterer Vertreter der biologisch orientierten Stresskonzepte ist die Stresstheorie nach Hans Selye (Selye und Fortier, 1950). Anders als Cannon (1914) legte Selye den Fokus jedoch nicht auf das sympathische Nervensystem, sondern auf die Aktivierung der Hypothalamus-Hypophysen-Nebennierenrinden-Achse (HHNA). Selye postuliert die Stressreaktion des Organismus als allgemeines Adaptationssyndrom (Selye und Fortier, 1950). Aus dieser Bezeichnung wird deutlich, dass die physiologische Stressantwort zwei Hauptmerkmale aufweist. Zum einen postuliert Selye die Stressreaktion als allgemein und unspezifisch, das heißt als unabhängig von der Art des Stressors. Diese Schlussfolgerung zog er aus tierexperimentel- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 17 len Studien, in denen die Versuchstiere auf verschiedenste Arten von Stressoren mit denselben und identischen Reaktionen antwortet, nämlich der verstärkten Aktivierung der HHNA. Das zweite Merkmal der Stressreaktion nach Selye ist ihre adaptive Natur. Die körperliche Stressreaktion wird mit dem Ziel initiiert Energiereserven zu mobilisieren und so einen Widerstand gegen den Stressor zu ermöglichen. Gemäß diesem Modell durchläuft der Organismus dabei drei Phasen. Nach einer ersten Alarmreaktion auf den Stressor, in der es zu kurzfristigen und reversiblen Organ- und Funktionsveränderungen kommt, gerät der Organismus in eine Widerstandsphase. Diese Phase zeichnet sich durch eine Rückkehr zu normalisierten Körperfunktionen aus, in der kaum noch Veränderungen der Organe zu erkennen sind und der Organismus erfolgreich Widerstand gegen den Stressor leistet. Hält die Stressbelastung jedoch an, kommt es zur Erschöpfungsphase, in der die Widerstandsenergie des Organismus abnimmt und es zu vergleichbaren Reaktionen und Veränderungen wie in der Alarmphase kommt. Diese Veränderungen der Organe führen in der dritten Phase jedoch zu irreversiblen Organschädigungen wie z.B. eine Vergrößerung der Nebennieren, eine Atrophie der lymphatischen Organe oder Geschwürbildung im Magen-Darm-Trakt. Diese irreversiblen Organschädigungen können letztendlich zum Tode führen (Pritzel, Brand, Markowitsch, 2003). Trotz der unterschiedlichen Fokussierung auf verschiedene körperliche Aspekte der Stressreaktion, verbindet die beiden genannten Vertreter der reaktionsorientierten Stresskonzepte die Tatsache, dass die Stressreaktion als unspezifisch postuliert wird. Sowohl Cannon, als auch Selye arbeiteten dabei rein tierexperimentell, so dass die Befunde nicht ohne weiteres auf den Menschen übertragen werden können. Obwohl sowohl Selye, als auch Cannon bereits die Wichtigkeit psychologischer Aspekte von Stress betonten, nutzen sie vorwiegend physische Stressoren und spezifizierten die Bedeutung psychologischer Aspekte nicht weiter. Im Rahmen der biologisch orientierten Stresskonzepte stellt das Konzept der Homöostase einen Brückenschlag zwischen rein psychologischen und rein biologischen Stresskonzepten her. Das Konzept der Homöostase Während die oben dargestellten biologisch orientierten Stresskonzepte hauptsächlich physische Stressore wie Hitze, Lärm oder Bewegungseinschränkung betrachtet haben, geht das Konzept der Homöostase von McEwen darüber hinaus (McEwen, 1998; McEwen und Wingfield, 2003). In diesem Konzept wird Stress als ein Ereignis definiert, das als bedrohlich wahrgenommen wird und das eine physiologische Stressreaktion hervorruft (McEwen und Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 18 Wingfield, 2003). Damit bildet dieses Konzept einen Konsens zwischen rein psychologisch und rein biologisch orientierten Konzepten. In Übereinstimmung mit den psychologisch orientierten Stresskonzepten misst McEwen individuellen Unterschieden in der Stressreaktion eine große Bedeutung zu. Diese Unterschiede haben zwei Ursachen. Zum einen hängt die körperliche Reaktion auf einen Stressor von der individuellen Interpretation und Einschätzung ab. Nur Reize, die als Bedrohung wahrgenommen werden führen zu physiologischen und verhaltensbezogenen Reaktionen. Eine zweite Ursache für individuelle Unterschiede wird in der Körperverfassung gesehen. Körperliche Faktoren, wie zum Beispiel die allgemeine physische Verfassung oder auch Diabetes oder starkes Übergewicht können die Reaktion auf einen Stressor und dessen Folgen beeinflussen (McEwen, 1998). McEwen (McEwen und Wingfield, 2003) unterscheidet zwischen psychischen Stressoren, die keine direkte körperliche Bedrohung darstellen und physischen Stressoren, die die Homöostase bedrohen und den Organismus in einen Zustand der Allostase versetzen. Der Begriff der Homöostase beschreibt in diesem Zusammenhang einen Zustand des Organismus, in dem er sich im Gleichgewicht befindet. Dieses Gleichgewicht bezieht sich auf die Aktivität der lebenswichtigen Funktionen wie z.B. pH-Wert, Körpertemperatur oder Sauerstoffgehalt des Bluts zur Versorgung der Organe. Die Aktivität dieser lebenswichtigen Funktionen bewegt sich in einem eng begrenzten Funktionsbereich, wobei der Organismus bestrebt ist Schwankungen zu vermeiden. Um die Homöostase aufrechterhalten zu können nutzt der Körper Systeme, die flexibel auf Veränderungen und Anforderungen reagieren können, wie z.B. Veränderungen im Blutdruck während des Aufstehens oder auch die Aktivierung der physiologischen Stressantwort in Reaktion auf einen Stressor. Die Aktivität dieser Systeme wird als Allostase bezeichnet. Schwankungen in der Aktivität dieser Systeme stellen also adaptive Reaktionen zur Sicherstellung des optimalen Funktionsniveaus der lebenswichtigen Funktionen dar. Allostase ist dementsprechend für den Organismus von großem Nutzen, um Bedrohungen bewältigen zu können. Der Nutzen dieser Reaktion ist allerdings mit Kosten für den Organismus verbunden. Die negativen Folgen einer anhaltenden Reaktion werden dementsprechend als allostatische Last bezeichnet (McEwen, 1998). Allostatische Last kann aus drei unterschiedlichen, sich unter Umständen gegenseitig bedingenden Ursachen resultieren. Eine Ursache allostatischer Last kann anhaltender Stress sein. Hierbei hält die Stressreaktion an oder wird in hoher Frequenz aktiviert. Dadurch verliert die Stressreaktion ihren adaptiven Charakter und hat negative Folgen für den Organismus. Eine zweite Ursache allostatischer Last ist eine Fehlregulierung Stressantwort in Form einer chronischen Aktivierung und ausbleibenden Beendigung. Als dritte Ursache lässt sich eine Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 19 Fehlregulation der Stressreaktion in Form einer abgestumpften Reaktion aufführen. Der Organismus initiiert im Falle einer Bedrohung die physiologische Stressantwort nicht oder zu schwach und kann somit nicht adäquat auf den Stressor reagieren (McEwen, 1998). McEwen (McEwen und Wingfield, 2003) führt zusätzlich zu dem Begriff der allostatischen Last noch den Begriff der allostatischen Überladung an, die sich durch schwerwiegende krankhafte Veränderungen und Funktionsbeeinträchtigungen auszeichnet. Allostatische Überladung lässt sich in zwei Formen unterteilen. Während sich Typ 1 der allostatischen Überladung aufgrund physischer Stressoren entwickelt, die dazu führen, dass der Körper mehr Energie verbraucht, als er aufnehmen kann und dadurch eine anhaltende Stressreaktion zeigt, rufen psychische Stressoren Typ 2 der allostatischen Überladung hervor. Hierbei kommt es zu keiner Energieunterversorgung, da weiterhin ausreichend oder mehr Energie als benötigt aufgenommen wird. Die Ursache sind durch die soziale Struktur bedingte psychische Stressoren, die eine andauernde Stressreaktion zur Folge haben und so den Organismus auf Dauer ernsthaft schädigen können (McEwen und Wingfield, 2003). Das Konzept der Homöostase betrachtet Stress als mögliches Resultat psychischer und physischer Stressoren. Dabei unterscheidet es zwischen adaptiven und maladaptiven Folgen der Reaktion und bietet ein Modell zur Entstehung dieser Folgen. Da es biologische und psychologische Aspekte verbindet, stellt es ein umfassendes Stresskonzept dar. 2.2.2 Die physiologische Stressreaktion Nachdem im vorangegangen Abschnitt verschiedene Stresskonzepte dargestellt wurden, soll in diesem Abschnitt näher auf die bereits erwähnte physiologische Stressreaktion eingegangen werden. Dafür wird zunächst die Stressreaktion dargestellt und im Anschluss näher auf das primäre und für die vorliegende Arbeit relevante Stresshormon Cortisol eingegangen. Die körperliche Stressreaktion wird in Situationen initiiert in denen der Organismus einen stressauslösenden Reiz wahrnimmt, um diese Bedrohung zu bewältigen. Somit ist die körperliche Stressreaktion ein Beispiel für Allostase, um die Homöostase aufrecht zu erhalten (de Kloet, Joels, Holsboer, 2005). Diese Reaktion ist durch die Aktivierung zweier Systeme gekennzeichnet. Zum einen reagiert das sympathische Nervensystem (SNS) mit einer verstärkten Aktivität, zum anderen kommt es unter Stress zu einer Aktivierung der Hypothalamus-Hypophysen-Nebennierenrinden-Achse (HHNA). Im Folgenden werden die beiden Systeme näher dargestellt. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 2.2.2.1 20 Das sympathische Nervensystem Die Aktivierung des sympathischen Nervensystems stellt eine erste, schnelle Reaktion des Organismus auf einen wahrgenommenen Stressor dar. Sie dient vornehmlich dazu Energiereserven für eine schnelle Handlungsbereitschaft zur Verfügung zu stellen. Walter Cannon (1914) bezeichnete diese Reaktion des Organismus dementsprechend als Notfallreaktion, die den Körper in einen „Fight-or-Flight“ Zustand versetzt. Das Ziel dieser Notfallreaktion ist eine adäquate Reaktion auf den Stressor. Die aufgrund der sympathischen Aktivierung von der Nebennierenmark ausgeschütteten Katecholamine Adrenalin und Noradrenalin können die Blut-Hirn-Schranke zwar nicht passieren, aktivieren jedoch den Vagusnerv und führen so zu einer verstärkten Ausschüttung von Noradrenalin aus Synapsen den zentralen Nervensystems (de Kloet et al., 2005). Dies geschieht über Projektionen des Vagusnerv in den Kern des Tractus Soletarius (NTS) sowie den Lokus Coeruleus (Roosevelt, Smith, Clough, Jensen, Browning, 2006). Der NTS wiederum projiziert über noradrenerge Zellgruppen zum einen direkt und zum anderen indirekt über den Lokus Coeruleus in die Amygdala, wodurch dort eine verstärkte Ausschüttung von Noradrenalin initiiert wird (Roozendaal, Okuda, de Quervain, McGaugh, 2006a; van Stegeren, 2009). Die Aktivierung des SNS hat eine gesteigerte Blutzirkulation zur Folge, die dazu dient das Herz, die Muskeln und das Gehirn mit mehr Sauerstoff und Glukose zu versorgen. Als Folge kommt es zu den bekannten Stresssymptomen wie beschleunigter Herzschlag oder verstärktes Schwitzen. 2.2.2.2 Die Hypothalamus-Hypophysen-Nebennierenrinden-Achse Während die Aktivierung des sympathischen Nervensystems eine schnelle und handlungsorientierte Reaktion darstellt, handelt es sich bei dem zweiten System, der HypothalamusHypophysen-Nebennierenrinden-Achse (HHNA), um ein relativ gesehen langsamer agierendes System. Nimmt der Organismus eine Bedrohung der Homöostase wahr, wird die HHNA aktiviert. Dies geschieht durch stressorspezifische Projektionen in den Hypothalamus (Jacobson, 2005). Physische Stressoren projizieren dabei über die entsprechenden somatosensorischen, viscerosensorischen oder osmosensorischen afferenten Pfade in den Hirnstamm oder das Diencephalon von wo aus die Signale den Hypothalamus erreichen (Herman, Figueiredo, Mueller, Ulrich-Lai, Ostrander, Choi, Cullinan, 2003). Psychische Stressoren scheinen im Gegensatz dazu eher im prefrontalen Cortex verarbeitet zu werden, von wo aus die Signale den Hypothalamus erreichen (Herman, Prewitt, Cullinan, 1996). Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 21 Dieser reagiert auf die eintreffenden Signale mit einer Ausschüttung der Neuropeptide Vasopressin und Corticotrophin Releasing Hormon (CRH) aus dem paraventriculären Nucleus. Durch das Blutpfortensystem gelangt CRH in die Hypophyse und stimuliert dort die Ausschüttung von Adrenocorticotrophin (ACTH). Über das periphere Blutsystem gelangt ACTH schließlich in die Nebennierenrinde und führt dort zu einer verstärkten Ausschüttung von Glucocorticoiden (GCs; Cortisol beim Menschen sowie Corticosteron bei Nagetieren; Charney, 2004; de Kloet et al., 2005). Eine adäquate Stressreaktion umfasst neben der Initiierung der Reaktion auch die angemessene Beendigung dieser Aktivität, sobald ausreichend Glucocorticoide ausgeschüttet sind. Um diese adäquate Regulierung der HHNA zu gewährleisten, wird ihre Aktivität über ein negatives Feedbacksystem gesteuert. Ansteigende Cortisolkonzentrationen beeinflussen dabei die Hypophyse und den Hypothalamus sowie den Hippocampus. Cortisol inhibiert die Ausschüttung von CRH aus dem Hypothalamus sowie die Ausschüttung von ACTH aus der Hypophyse (Jacobson, 2005). Zusätzliches negatives Feedback wird über ACTH selbst vermittelt, so dass erhöhte Konzentrationen die Ausschüttung von CRH aus dem Hypothalamus hemmen (Lupien und Lepage, 2001). Die Aktivität der HHNA stellt ein komplexes System dar, dessen Funktion von verschiedenen Faktoren beeinflusst wird. Als Einflussfaktoren werden verschiedene Variablen diskutiert, unter anderem das Alter der Versuchspersonen, die Tageszeit, Schwangerschaft, Rauchen, Koffein und Alkohol (Kudielka, Hellhammer, Wust, 2009). Besondere Aufmerksamkeit hat dabei das Geschlecht der Versuchspersonen erhalten, da sich in einer Vielzahl von Studien Geschlechtsunterschiede bei der Cortisolantwort auf Stressoren zeigten (Kudielka und Kirschbaum, 2005). Dabei wurden häufig stärkere Reaktionen bei Männern, als bei Frauen beobachtet (Earle, Linden, Weinberg, 1999; Kirschbaum, Wust, Hellhammer, 1992; Kumsta, Entringer, Koper, van Rossum, Hellhammer, Wust, 2007; Lovallo, Farag, Vincent, Thomas, Wilson, 2006; Seeman, Singer, Wilkinson, McEwen, 2001; Stroud, Salovey, Epel, 2002). Diese Unterschiede zeigen sich allerdings nur bei einer Cortisolbestimmung über den Speichel, in dem das freie und ungebundene Cortisol zu finden ist und nicht bei einer Cortisolbestimmung über das Blutplasma (Kirschbaum, Kudielka, Gaab, Schommer, Hellhammer, 1999). Obwohl diese Geschlechtsunterschiede in einer Vielzahl von Studien berichtet wurden (Kajantie und Phillips, 2006; Kudielka und Kirschbaum, 2005; Otte, Hart, Neylan, Marmar, Yaffe, Mohr, 2005), existieren auch Studien, die diese Unterschiede nicht berichten (Kelly, Tyrka, Anderson, Price, Carpenter, 2008; van Stegeren, Wolf, Kindt, 2008; Wolf et al., 2001). Neben diesen generellen Geschlechtsunterschieden zeigt sich zudem ein starker Einfluss von Geschlechtshormonen auf die Cortisolantwort. So scheinen sowohl die Cortisolantwort bei Frauen als auch die Geschlechtsunterschiede von der aktuellen Zyklusphase Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 22 abzuhängen. Frauen in der Lutealphase zeigen dabei eine mit Männern vergleichbare Cortisolantwort auf einen psychosozialen Stressor, wohingegen die Cortisolausschüttung bei Frauen in der Follikularphase und bei Frauen, die orale Kontrazeptiva verwenden deutlich geringer ausfällt (Kirschbaum et al., 1999; Rohleder, Schommer, Hellhammer, Engel, Kirschbaum, 2001; Rohleder, Wolf, Piel, Kirschbaum, 2003; Wolf et al., 2001). Als ursächlich für diese Unterschiede in der Cortisolstressantwort wird das Korticosteroid-bindende Globulin (CBG) angesehen, da die im Blut zirkulierenden Steroid-bindenden Globuline durch syntetische Östradiolkomponenten verändert werden (Wiegratz, Kutschera, Lee, Moore, Mellinger, Winkler, Kuhl, 2003). Somit wird deutlich, dass das Geschlecht sowie Geschlechtshormone einen bedeutenden Einfluss auf die Aktivität der HHNA nehmen und in experimentellen Studien als Einflussfaktoren mit berücksichtigt werden müssen. Die verstärkte Ausschüttung des primären Stresshormons Cortisol kennzeichnet das Endprodukt der physiologischen Stressantwort. Bei der physiologischen Stressantwort handelt es sich allerdings nicht um eine Reaktion, die ausschließlich im Falle eines wahrgenommenen Stressors aktiviert wird und anschließend wieder vollständig beendet wird, sondern vielmehr um eine ereignisabhängige Verstärkung der basalen Aktivität der HHNA. Die basale Ausschüttung von Cortisol unter normalen, nicht-stressigen Bedingungen geschieht nicht kontinuierlich, sondern in mehreren Phasen über den Tag verteilt. Während einer Sekretionsphase zeigt die Nebennierenrinde eine starke Aktivität, die nach der Beendigung der Phase wieder von Inaktivität abgelöst wird (Van Cauter, 1990). Die Ausschüttung von Cortisol unterliegt einem circadianen Rhythmus, wodurch die Cortisollevel über den Tag hinweg variieren. Die höchsten Cortisolkonzentrationen sind am frühen Morgen zu beobachten, während am Abend die geringsten Konzentrationen zu finden sind (Jacobson, 2005; Kirschbaum und Hellhammer, 1989). Während des Tages zeigen sich Änderungen in Amplitude und Frequenz der Cortisolausschüttung, wobei der circadiane Rhythmus auf Änderungen der Frequenz zurückzuführen ist. Während der Sekretionsphasen am Morgen wird dabei häufiger Cortisol ausgeschüttet, als während der Sekretionsphasen am Abend (Herbert, Goodyer, Grossman, Hastings, de Kloet, Lightman, Lupien, Roozendaal, Seckl, 2006). Der Organismus initiiert im Falle eines wahrgenommenen Stressors eine Stressreaktion, die mit vielfältigen Veränderungen des Hormonhaushalts einhergeht. Die physiologische Stressreaktion stellt somit eine komplexe und aufeinander abgestimmte Reaktion dar, deren Gleichgewicht für die optimale Funktionsfähigkeit notwendig ist. Störungen oder Fehlfunktionen haben hohe Kosten für den Organismus und stellen somit eine allostatische Last dar. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 23 Ursachen für eine auftretende allostatische Last können, wie in Kapitel 1.1.2 dargestellt wurde entweder ein andauernder Stressor, die Unfähigkeit des Organismus zur Beendigung der Stressreaktion oder eine inadäquate Initiierung der Stressreaktion sein. Während die ersten beiden Ursachen zu dauerhaft erhöhten Cortisolspiegeln führen, hat eine inadäquate Initiierung der Stressreaktion niedrige Cortisolspiel zur Folge. Ist dies der Fall, werden diejenigen Systeme, wie z.B. das Immunsystem, die unter normalen Umständen während der Stressreaktion durch Cortisol reguliert werden, nicht adäquat gesteuert (McEwen, 1998). 2.2.3 Stressoren Nachdem im vorangegangenen Abschnitt die physiologische Stressreaktion näher betrachtet wurde, soll im folgenden Abschnitt der Frage nachgegangen werden, welche Stressoren sich im Labor und außerhalb des experimentellen Laborsettings als zuverlässige stressinduzierende Situationen herausgestellt haben und durch welche Faktoren sich diese Ereignisse auszeichnen. Diese Frage ist zum einen für die experimentelle Forschung von Interesse, da im Labor eine Vielzahl von Stressoren eingesetzt wird. Zum anderen stellt sich die Frage, in wie weit die im Labor eingesetzten Stressoren denjenigen Stressoren ähneln, die außerhalb des Labors im menschlichen Alltag zu finden sind. Im Folgenden sollen daher zunächst diejenigen Faktoren dargestellt werden, die sich im Labor als besonders geeignet herausgestellt haben, um eine Stressreaktion hervorzurufen. Anschließend wird auf Stressoren außerhalb des Laborkontextes eingegangen. In laborexperimentellen Studien werden verschiedene Stressoren eingesetzt. Diese Stressoren wurden bezüglich der Faktoren analysiert, die mit einer Stressreaktion einhergehen. Eine erste vergleichende Betrachtung verschiedener psychischer Stressoren im human- und tierexperimentellen Bereich nahm Mason (1968) vor. Dabei beschrieb er drei Faktoren, die eine verstärkte Stressantwort hervorrufen. Zum einen stellte sich der Faktor „Neuheit der Situation“ als bedeutsam für die Initiierung der Stressreaktion heraus. Situationen, die von Probanden oder Experimentaltieren als unvorhersagbar erlebt werden, führen zu einer verstärkten Aktivität der HHNA. Als dritten Faktor neben der „Neuheit der Situation“ und der „Unvorhersagbarkeit“ nennt Mason die „persönliche Involviertheit“ der Probanden (Mason, 1968). Eine vergleichende Analyse von verschiedener Laborstressoren im humanexperimentellen Bereich nahmen Dickerson und Kemeny (2004) vor. Aus dieser Analyse entwickelten sie ein Modell, das dem Faktor „Bedrohung des sozialen Selbst“ eine besondere Bedeutung beimisst. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 24 Der Kern des Modells geht davon aus, dass der Mensch als soziales Wesen danach strebt sein soziales Selbst zu erhalten. Das soziale Selbst eines Menschen definiert seine Stellung und Wertschätzung in der Gesellschaft. Es beinhaltet Fähigkeiten und Eigenschaften, die einem Individuum durch die Bewertung durch andere Mitglieder der Gesellschaft zugeschrieben werden. Eine positive Bewertung der Fähigkeiten und Eigenschaften ist mit Wertschätzung und sozialem Status verbunden. Eine negative Bewertung der für die Gesellschaft relevanten Eigenschaften geht mit einer Reduzierung dieser Wertschätzung und sozialen Stellung einher. Die Erhaltung des sozialen Selbst stellt somit ein zentrales Ziel dar, das vergleichbar mit der Erhaltung der physischen Unversehrtheit ist. Situationen, in denen das Individuum seine Eigenschaften und Fähigkeiten zeigen muss und offenkundig bewertet wird, stellen eine Bedrohung des sozialen Selbst dar. Diese Bedrohung des sozialen Selbst aktiviert ein System, das danach strebt diese Bedrohung zu bewältigen und angemessene psychische, physische und verhaltensbezogene Reaktionen zu initiieren. Zu diesem System gehört auch die Aktivierung der HHNA als adaptive Reaktion zur Reduzierung oder Bewältigung der Bedrohung. Neben der Bedrohung des sozialen Selbst führen die Autoren mit der Unkontrollierbarkeit der Situation einen weiteren Faktor an, die eine Situation zu einem stressauslösenden Ereignis werden lässt. Dieser Faktor verstärkt die wahrgenommene Bedrohung des sozialen Selbst. In Situationen, in denen das Verhalten der Person keine Auswirkung auf den Verlauf hat wird die Bedrohung verstärkt wahrgenommen. Da das Verhalten der Person keinen Einfluss auf den Verlauf oder den Ausgang der Situation hat, nimmt die Bedrohung des sozialen Selbst zu und eine verstärkte Aktivierung der HHNA findet statt. In der genannten Metaanalyse untersuchten Dickerson und Kemeny (2004) 208 Laborstudien, die psychische Stressoren eingesetzt haben. Die Stressoren wurden dabei in 5 Kategorien eingeteilt. Neben Lärmexposition, Filmen, freien Redesituationen und kognitiven Aufgaben wurden Stressoren betrachtet, die eine Kombination aus freier Redesituation und kognitiver Aufgabe darstellten. Alle Stressoren führten zu einer Aktivierung der HHNA. Entsprechend des Modells fanden sich aber die stärksten Stressreaktionen bei denjenigen Stressoren, die aus einer Kombination aus freier Redesituation und kognitiver Aufgabe vor einem Gremium bestanden. Diese Stressoren stellen motivierte Leistungsaufgaben dar, bei denen der Proband von anderen Personen bewertet wird. Dadurch nimmt der Proband eine Bedrohung des sozialen Selbst wahr. Durch diese Kombination der Faktoren werden diese Stressoren besonders effektiv zur Initiierung einer physiologischen Stressreaktion im Laborkontext. Ein Beispiel für solch einen effektiven Stressor stellt der Trier Sozial Stress Test (TSST, Kirschbaum, Pirke, Hellhammer, 1993) dar. Bei diesem Stressor handelt es sich um eine Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 25 Kombination aus freier Redesituation und kognitiver Aufgabe. Der Proband erhält die Aufgabe in einem fiktiven Vorstellungsgespräch eine fünfminütige freie Rede über seine positiven Eigenschaften zu halten und anschließend eine mathematische Aufgabe zu lösen. Dabei wird seine Leistung von einem Gremium bewertet. Dieses Gremium agiert in einer neutralen Weise und vermeidet positive soziale Interaktion. In dieser Situation nimmt der Proband eine Bedrohung des sozialen Selbst wahr, da er persönliche Eigenschaften und Fähigkeiten präsentiert, die bewertet werden. Da sich das Gremium unerwartet distanziert verhält und der Proband mit seinem Verhalten die Reaktion des Gremiums nicht beeinflussen kann, nimmt der Proband zusätzlich eine Unkontrollierbarkeit der Situation wahr. Durch diese Kombination stellt der TSST einen sehr effektiven Stressor in laborexperimentalen Situationen dar. Außerhalb des experimentellen Laborsettings, in denen Stresssituationen künstlich geschaffen werden, existiert eine Vielzahl an potenziellen Stressoren. Neben beruflichen, persönlichen und psychosozialen Stressauslösern ist prinzipiell jede Situation, die die Homöostase einer Person gefährdet ein potenzieller Stressor. Um Aussagen über die Stressreaktion auf naturalistische Stressoren, also Stressoren außerhalb des Laborsettings, treffen zu können, werden Situationen untersucht, die für eine Vielzahl an Menschen stressauslösend sein können. Dazu gehören z.B. Turniersituationen (Rohleder, Beulen, Chen, Wolf, Kirschbaum, 2007), Seefahrten (Liberzon, Abelson, King, Liberzon, 2008), Überforderung im Beruf (Schlotz, Hellhammer, Schulz, Stone, 2004) oder Prüfungssituationen (Stowell, 2003). Letztere wurden häufig in Studien untersucht, um die physiologische Stressreaktion auf einen naturalistischen Stressor zu beobachten. Prüfungssituationen bieten sich für diese Untersuchungen an, da die Probanden in solchen Situationen persönlich involviert sind und eine hohe Motivation mitbringen die Prüfung zu bestehen. In den meisten Fällen haben Prüfungen eine direkte oder indirekte Auswirkung auf die berufliche Zukunft der Person und die Ergebnisse der Prüfungen sind auch im sozialen Bezugsraum relevant. Je nach Bedeutung und Art der Prüfung stellt sie eine Bedrohung des sozialen Selbst dar, da relevantes Wissen von einer Person abgefragt und bewertet wird. Diese Bewertung ist für eine Vielzahl an sozialen Kontakten zugänglich, da über Benotungen sowohl in der Schule, als auch im universitären Kontext gesprochen wird. Da vor einer Prüfung die Prüfungsaufgaben oder –fragen nicht bekannt sind, sind Prüfungssituationen in gewissem Maße auch unvorhersagbar, obwohl sich die Person auf das zu prüfende Thema vorbereitet. Prüfungssituationen weisen also die selben Faktoren auf, die auch bei Laborstressoren effektiv sind. Bedrohung des sozialen Selbst, Unvorhersagbarkeit/Unkontrollierbarkeit und persönliche Involviertheit machen auch Prüfungssituationen zu stressinduzierenden Ereignissen. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 26 Die Ergebnisse aus Prüfungsstudien zeigen allerdings unterschiedliche Ergebnisse. Während einige Autoren von erhöhten Cortisolspiegeln berichten (Herbert, Moore, de la Riva, Watts, 1986; Lacey, Zaharia, Griffiths, Ravindran, Merali, Anisman, 2000; Loft, Thomas, Petrie, Booth, Miles, Vedhara, 2007; Lovallo, Pincomb, Edwards, Brackett, Wilson, 1986; Schoofs, Hartmann, Wolf, 2008), zeigen andere Studien reduzierte Cortisolspiegel oder keine Veränderung (Spangler, 1997). Diese uneinheitlichen Befunde lassen darauf schließen, dass sich die untersuchten Prüfungssituationen teilweise stark von einander unterscheiden (Stowell, 2003). So wurden schriftliche (Gaab, Sonderegger, Scherrer, Ehlert, 2006; Ng, Koh, Mok, Chia, Lim, 2003) und mündliche Prüfungen (Schoofs et al., 2008), einzelne Prüfungen (Frankenhaeuser, von Wright, Collins, von Wright, Sedvall, Swahn, 1978) und Prüfungsphasen (Loft et al., 2007),Immatrikulations- (Frankenhaeuser et al., 1978) und Abschlussprüfungen (Zeller, Handschin, Gyr, Martina, Battegay, 2004) untersucht. Diese Prüfungen unterscheiden sich nicht nur durch das jeweilige Setting, sondern auch durch das Ausmaß der stressauslösenden Faktoren. Während z.B. in mündlichen Prüfungen die Bedrohung des sozialen Selbst als groß anzunehmen ist, scheint sie in schriftlichen Prüfungen, die sich durch ihre Anonymität auszeichnen, geringer zu sein. Es bleibt allerdings offen, welchen Einfluss die im Laborkontext identifizierten Faktoren bei naturalistischen Stressoren haben, da diese Faktoren bisher noch nicht gezielt untersucht wurden. Zusammenfassend lässt sich sagen, dass sowohl im Labor, als auch außerhalb des Labors verschiedenste stressaulösende Situationen existieren, die sich jedoch in ihrer Effektivität unterscheiden. Während für Laborstressoren die kritischen Faktoren bereits definiert werden konnten, sind für Stressoren außerhalb des Laborsettings diese Faktoren bisher nicht systematisch untersucht wurden. In wie weit die Befunde aus laborexperimentellen Stressstudien auf den Alltag übertragbar sind, bleibt daher noch offen. 2.2.4 Cortisoleffekte auf das Gehirn Unabhängig von der Art des Stressors übt das primäre Stresshormon und Endprodukt der Stressreaktion, Cortisol, vielfältige Effekte auf den Organismus aus. Aufgrund seiner Lipidlöslichkeit hat Cortisol, im Gegensatz zu den Katecholaminen Adrenalin und Noradrenalin, die Eigenschaft die Blut-Hirnschanke uneingeschränkt passieren zu können und nimmt so direkt Einfluss auf Strukturen und Prozesse im Gehirn. Im Gehirn selber finden sich zwei unterschiedliche Rezeptorarten für Cortisol, die sich hinsichtlich ihrer Affinität für Cortisol und ihrer Verbreitung im Gehirn unterscheiden lassen (de Kloet, Vreugdenhil, Oitzl, Joels, 1998; Herbert et al., 2006). Während sich der Mineralokorticoidrezeptor (MR oder Typ I-Rezeptor) Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 27 durch eine hohe Affinität für Cortisol auszeichnet und bereits durch geringe Mengen an Cortisol aktiviert wird, weist der Glucocorticoidrezeptor (GR oder Typ II-Rezeptor) eine etwa 10fach geringere Affinität für Cortisol auf (Arriza, Simerly, Swanson, Evans, 1988). Dieser Rezeptor wird erst durch hohe Cortisolkonzentrationen aktiviert, z.B. bei einer verstärkten Ausschüttung von Cortisol bei Stress oder auch während des circadianen Peaks von Cortisol. Im Gegensatz dazu zeigen die MRs eine Sättigung bereits während der basalen Aktivität der HHNA (de Kloet et al., 1998; Herbert et al., 2006). Die Rezeptoren sind weit über das Gehirn verteilt, wobei sie sich deutlich in ihrer Verbreitung unterscheiden (Herbert et al., 2006). Die größte Dichte an GRs findet sich im Nucleus paraventricularis des Hypothalamus sowie in Neuronen des limbischen Systems. Limbische Neurone weisen ebenfalls eine große Anzahl an MRs auf. MRs und GRs finden sich auch vermehrt in Pyramidenzellen des Hippocampus. Hiervon ausgenommen werden muss die CA3 Schicht, dort lassen sich kaum GRs aufweisen. Beide Rezeptorarten befinden sich auch im Gyrus dentatus, dem Nucleus Amygdaloidis und dem lateralem Septum (de Kloet et al., 2005). Insgesamt lässt sich festhalten, dass GRs weit im Gehirn verbreitet sind, wohingegen die Verbreitung von MRs deutlich eingeschränkter ist. Im limbischen System lassen sich beide Rezeptorarten in großer Menge finden, außerhalb des limbischen Systems ist die Verbreitung von MRs jedoch deutlich geringer (de Kloet et al., 1998). Die hohe Konzentration von MRs und GRs im Hippocampus macht diese Formation, bestehend aus Gyrus dentatus, Ammonshorn und Subiculum, besonders anfällig für Cortisoleffekte. Dies ist von besonderer Bedeutung, da der Hippocampus eng mit Gedächtnisfunktionen verknüpft ist (Lynch, 2004; Thompson, 2005). In diesem Zusammenhang sind zwei Funktionen des Hippocampus von besonderer Relevanz. Dies sind zum einen die Neurogenese und zum anderen die Langzeitpotenzierung (LTP), die beide mit dem Gedächtnis assoziiert werden (siehe Kapitel 1.1.2). So zeigte sich, dass Stress zu einer reduzierten Produktion neuer Neurone im Hippocampus führt. Diese reduzierte Neurogenese wurde bei verschiedenen psychischen und physischen Stressoren beobachtet (Kozorovitskiy und Gould, 2004; Malberg und Duman, 2003; Mirescu, Peters, Gould, 2004; Tanapat, Hastings, Rydel, Galea, Gould, 2001). Neben der reduzierten Neurogenese kommt es in Reaktion auf Stress zu einer erhöhten Atrophie der Dendriten der Pyramidenzellen in der CA3-Region des Hippocampus (Magarinos, Orchinik, McEwen, 1998; McKittrick, Magarinos, Blanchard, Blanchard, McEwen, Sakai, 2000). Die reduzierte Neurogenese und eine erhöhte Atrophie werden daher als Ursache der häufig mit chronischem Stress einhergehenden beobachteten Reduzierung des Hippocampusvolumens betrachtet. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 28 Ein Einfluss von Stress und Cortisol wurde ebenfalls auf die LTP im Hippocampus beobachtet. Dabei müssen jedoch kurzzeitige und langfristige Effekte unterschieden werden. Innerhalb weniger Minuten kommt es aufgrund gestiegener Cortisollevel in der CA1 Region des Hippocampus zu einer erhöhten Frequenz der postsynaptischen Potenziale (Karst, Berger, Turiault, Tronche, Schutz, Joels, 2005; Venero und Borrell, 1999). Zudem konnte demonstriert werden, dass Corticosterone die synaptische Plastizität in der CA1 Region erleichtert, sofern die Verabreichung zeitlich nah und nicht länger als 30 Minuten vor der Induzierung der LTP liegt (Wiegert, Joels, Krugers, 2006). Im Gegensatz zu diesen kurzzeitigen verstärkenden Effekten findet sich einige Stunden nach dem Stressor eine reduzierte LTP (Diamond, Bennett, Fleshner, Rose, 1992; Kim und Diamond, 2002; Mesches, Fleshner, Heman, Rose, Diamond, 1999; Pavlides, Ogawa, Kimura, McEwen, 1996; Wiegert, Pu, Shor, Joels, Krugers, 2005). Während Stress auf der einen Seite zu kurzfristiger Verstärkung von LTP und synaptischer Erregung führt, zeigen sich im Anschluss an diese kurzfristigen Effekte langfristige Hemmungen der LTP und synaptischen Erregbarkeit. Während also die kurzfristigen Effekte eine verbesserte Verarbeitung neu eingehender Informationen zur Folge haben, werden Informationen im Anschluss an die akute Stressphase weniger stark verarbeitet oder sogar gehemmt, so dass diejenigen Informationen aus der akuten Stressphase konsolidiert werden können (Joels, Krugers, Karst, 2008). Die adaptive Stressreaktion des Organismus dient einerseits der adäquaten Reaktion auf einen Stressor. Diese adaptive Reaktion geht allerdings auch mit Kosten für den Organismus einher. Während die Effekte auf die Funktionen und Strukturen des Gehirns bei akuten Stressoren reversibel sind, kann eine chronische Stressbelastung hingegen zu bleibenden Veränderungen einhergehen, die den Organismus in seiner Funktion schädigen (Wolf, 2006). In diesem Abschnitt der Arbeit wurde das Thema Stress genauer betrachtet. Zur Definition und Betrachtung von Stress existieren verschiedene Konzepte. Zum einen definiert das psychologisch orientierte Stresskonzept von Lazarus (Lazarus und Folkman, 1984) Stress als Interaktion zwischen Person und Situation. Biologisch orientierte Stresskonzepte wie Cannon (1914) und Selye (Selye und Fortier, 1950) sie vertreten, definieren Stress hingegen über die Initiierung der körperlichen Stressreaktion. Das Konzept der Homöostase (McEwen, 1998; McEwen und Wingfield, 2003) schließlich betrachtet Stress als Reaktion auf eine wahrgenommene Bedrohung dieser Homöostase. Weiterhin wurde in diesem Kapitel die physiologische Stressreaktion näher betrachtet und ihre zwei Komponenten, die Aktivierung des sym- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 29 pathischen Nervensystems und der HHNA, sowie die Effekte von Stress auf Funktion und Struktur des Gehirns dargestellt, wobei besonderes Interesse denjenigen Strukturen und Funktionen galt, die auch für das Gedächtnis von Relevanz sind. Abschließend wurde eine Betrachtung verschiedener stressauslösender Faktoren von Laborstressoren vorgenommen. Im folgenden Abschnitt der Arbeit sollen nun die beiden Themen „Stress“ und „Gedächtnis“ verknüpfend betrachtet werden und Einflüsse von Stress auf das Gedächtnis dargestellt werden. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 2.3 30 Der Einfluss von Stress auf das Gedächtnis Nachdem in den vorangegangenen Abschnitten das Gedächtnis und das Konzept Stress getrennt voneinander betrachtet wurden, geht das folgende Kapitel auf die Zusammenhänge zwischen Stress und Gedächtnis ein. Dabei werden die Auswirkungen von Stress auf die verschiedenen Gedächtnisphasen näher betrachtet. Betrachtet man den Einfluss von Stress und Stresshormonen auf das Gedächtnis, so hängen die gefundenen Effekte von der spezifischen Gedächtnisphase ab, die untersucht wird. Dabei sind drei Phasen zu unterscheiden. Die erste Wahrnehmung und Analyse eines Reizes wird als Enkodierung bezeichnet. Dabei wird der Reiz aufgenommen und verarbeitet. Ein Stressor oder pharmakologisch erhöhte GC-Level zu diesem Zeitpunkt beeinflussen also die Enkodierung des neu zu lernenden Materials. Da sich an die Phase der Enkodierung jedoch direkt die Konsolidierung des Reizmaterials anschließt, sind Effekte auf die Konsolidierung bei derartigen Versuchsdesigns nicht auszuschließen. Bei der Konsolidierung der Information handelt es sich um die Festigung der Inhalte und Integration in bereits bestehende Strukturen. Ein Stressor oder pharmakologische Behandlung nach der Enkodierung des Lernmaterials beeinflusst somit die Phase der Konsolidierung. Um hierbei einen Effekt auf den Abruf der Informationen, der dritten Phase des Gedächtnisprozesses, auszuschließen sollte der Abruf der Informationen zeitlich so verzögert geschehen, dass die GC-Level wieder auf ein Level gesunken sind, die mit den Leveln vor dem Stressor oder der pharmakologischen Behandlung vergleichbar sind (Baselinewert). Die Einflussmöglichkeiten von Stress und Cortisol sind in Abbildung 3 veranschaulicht. Abbildung 3: Einflussmöglichkeiten von Cortisol und Stress auf die verschiedenen Gedächtnisphasen. Stress/Cortisol vor der Enkodierung übt Einflüsse auf die Enkodierung und Konsolidierung aus und bei zeitlicher Nähe auch auf den Abruf. Stress/Cortisol nach der Enkodierung wirkt auf die Konsolidierung und bei zeitlicher Nähe auf den Abruf. Stress/Cortisol vor dem Abruf wirkt auf die Phase des Gedächtnisabrufs. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 31 Während sich die Forschung weitgehend einig darüber ist, dass Stress vorwiegend negative Effekte auf den Abruf von Gedächtnisinhalten hat, ist die Befundlage für die Bedeutung von Stress auf die Phasen der Enkodierung und Konsolidierung weniger eindeutig (Roozendaal et al., 2006a; Wolf, 2008). Daher sollen im folgenden Kapitel die Befunde zum Einfluss von Stress auf die Gedächtnisenkodierung und -konsolidierung dargestellt werden. 2.3.1 Stresseffekte auf die Enkodierung Bei Untersuchungen zum Einfluss von Stress auf die Gedächtnisenkodierung, muss zwischen Stressoren unterschieden werden, die innerhalb des Lernkontextes stattfinden und Stressoren, die außerhalb des Lernkontextes vor der Enkodierung stattfinden (Joels, Pu, Wiegert, Oitzl, Krugers, 2006). Beiden Arten von Studien ist gemeinsam, dass die Enkodierung der Stimuli unter erhöhten GC-Leveln stattfindet. Allerdings können hierbei Einflüsse auf die Konsolidierung nicht ausgeschlossen werden, da sich diese direkt an die Enkodierung anschließt und sich die Effekte der GCs auch auf diese Gedächtnisphase auswirken können (Lupien und McEwen, 1997). Untersuchungen zu Stress innerhalb des Lernkontextes werden zumeist tierexperimentell durchgeführt. Dabei wird die Lernsituation für die Versuchstiere so variiert, dass sie mehr oder weniger stressauslösend ist. Die Ergebnisse dieser Studien zeigen, dass eine stressvolle Lernsituation die Bildung des Gedächtnisses begünstigen kann. So testeten Sandi et al. (1997) das räumliche Lernen von Ratten in einer Wasserlabyrinthaufgabe. Dabei fanden die Lerndurchgänge entweder in kaltem Wasser (19°C) oder in warmem Wasser (25°C) statt. Die Autoren fanden, dass das kalte Wasser bei den Ratten zu erhöhten Corticosteronspiegeln sowie einem schnelleren Erlernen und einer besseren Aufrechterhaltung der Lerninhalte führte, als das warme Wasser. Die zusätzliche Injektion von Corticosteron direkt im Anschluss an die Lernepisoden führte in der Gruppe der Ratten, die bei 25°C trainiert wurden zu einer verbesserten Gedächtnisleistung. Dieser Effekt zeigte sich nicht bei Ratten, die in 19°C kalten Wasser trainiert wurden. In einer weiteren Studie konnten die Autoren zeigen, dass dieser Effekt durch die Ausschüttung von GCs verursacht wird (Akirav, Kozenicky, Tal, Sandi, Venero, Richter-Levin, 2004) Des weiteren konnten Conrad et al. (Conrad, Lupien, McEwen, 1999) demonstrieren, dass die Stresseffekte außerhalb des Lernkontextes auf das Gedächtnis durch die GR-Rezeptoren, nicht aber durch die MRRezeptoren moduliert werden. Dass nicht nur pharmakologische Behandlung einen Einfluss auf die Gedächtnisleistung hat, sondern auch Stress vor der Lernaufgabe zeigten Conrad et al. (2004). Männliche und weib- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 32 liche Ratten wurden eine Stunde lang Immobilisationsstress („restraint stress“) unterzogen. Zwei Stunden nach Beginn des Stressors wurden die Ratten in einer Wasserlabyrinthaufgabe trainiert. Vier Stunden nach der Lernaufgabe zeigte sich, dass der Stress die Gedächtnisleistung bei männlichen Ratten beeinträchtigte, bei weiblichen Ratten jedoch verbesserte, wobei gezeigt werden konnte, dass die Zyklusphase bei weiblichen Ratten keinen Einfluss hatte. Diese tierexperimentellen Studien zeigen, dass pharmakologische Behandlung und Stress die Gedächtnisleistung beeinflussen. Dabei wurde offensichtlich, dass die Effekte von verschiedenen Faktoren wie dem Geschlecht der Versuchstiere und der Dosis der pharmakologischen Behandlung abhängen. Im Humanbereichen finden sich gemischte Effekte von Stress und Cortisol auf die Gedächtnisleitung für Stimuli, die unter erhöhten Cortisolspiegeln enkodiert werden. Pharmakologische Studien zeigten auf der einen Seite negative Effekte auf die Gedächtnisleistung für neutrale Reize wie Objekte und Gesichter (Monk und Nelson, 2002) oder neutrale Wörter (Kuhlmann und Wolf, 2006b), auf der anderen Seite aber positive Effekte auf das Langzeitgedächtnis für emotionale Bilder (Buchanan und Lovallo, 2001; Kuhlmann und Wolf, 2006b). Diese Befunde sind jedoch nicht immer zu beobachten. So zeigen weitere Studien positive Effekte von Cortisol auf das Gedächtnis für sowohl emotionale, als auch neutrale Reize, wobei diese Effekte eine umgekehrt U-förmige Beziehung zeigten und nur bei der niedrigen Dosierung der Cortisolgabe zu beobachten waren (Abercrombie, Kalin, Thurow, Rosenkranz, Davidson, 2003). Im Gegensatz zu diesen positiven Effekten vor allem auf emotional erregendes Material zeigten sich auch negative Effekt von Cortisol auf die Gedächtnisleistung für emotional erregendes Material (Tops, van der Pompe, Baas, Mulder, Den Boer, Meijman, Korf, 2003) bzw. die Details dieser Stimuli (Rimmele, Domes, Mathiak, Hautzinger, 2003) und keine Effekte (Tops et al., 2003) oder positive Effekte auf die Details (Rimmele et al., 2003) von neutralen Stimuli. Neben diesen uneinheitlichen Befunden existieren auch Studien, die keine Effekte von Cortisol auf die Gedächtnisleistung berichten konnten (Hsu, Garside, Massey, McAllister-Williams, 2003; Lupien, Gillin, Hauger, 1999). Die Befunde pharmakologischer Studien berichten somit uneinheitliche Befunde für die Effekte von Cortisol auf die Enkodierung. Auch beim Einsatz von Stressoren vor der Enkodierung finden sich heterogene Befunde zu den Stresseffekten auf die Gedächtnisleistung. Kirschbaum et al. (1996) verglichen die Effekte eines psychosozialen Stressors (TSST) mit den Effekten von pharmakologisch verabreichtem Cortisol auf die Gedächtnisleistung. In der ersten Studie wurden die Probanden zunächst dem Stressor oder der Kontrollbedingung un- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 33 terzogen und lernten im Anschluss eine Wortliste. Das Gedächtnis für diese Wörter wurde in einem direkten Abruftest 5 Minuten nach der Enkodierung getestet. In der zweiten Studie wurde den Probanden 10 mg Cortisol oder Placebo verabreicht, bevor sie eine Stunde später die selbe Aufgabe bearbeiteten. Die Ergebnisse zeigen für beide Studien einen negativen Einfluss auf die Gedächtnisleistung. In einer weiteren Studie mit diesem Design, in der als Stressor ebenfalls der TSST eingesetzt wurde, zeigten sich starke Geschlechtsunterschiede für die Effekte von Stress auf die Gedächtnisleistung (Wolf et al., 2001). In dieser Studie zeigte sich kein genereller Effekt des Stressor, jedoch eine negative Korrelation zwischen der Cortisolantwort und der Gedächtnisleistung. Diese Korrelation wurde ausschließlich durch die starken Zusammenhänge bei Männern getragen, wohingegen sich für Frauen keine Zusammenhänge zeigten. Elzinga et al. (2005) testeten den Einfluss eines Stressors auf Wörter, die vor dem Stressor und Wörter, die nach dem Stressor enkodiert wurden. Der Stressor in dieser Studie bestand dabei aus kognitiven Aufgaben, die unter zwischenmenschlichen Druck gelöst werden mussten. Als Maße für die Gedächtnisleistung wurden ein direkter Abruftest und ein 24-stunden verzögerter Abruftest eingesetzt. Die Autoren fanden, dass sich Stress und die damit einhergehenden Cortisolanstiege negativ auf das Langzeitgedächtnis für Informationen, die nach dem Stressor enkodiert wurden auswirkte, nicht aber auf die Leistung in der direkten Abrufaufgabe. In dieser Studie wurde allerdings keine Kontrollgruppe untersucht, so dass alle Probanden den psychosozialen Stressor durchliefen und die Stresseffekte auf die Enkodierung mit der Leistung für die Wörter verglichen wurde, die vor dem Stressor präsentiert wurden. Dieses Versuchsdesign lässt jedoch Fragen nach dem Einfluss des Stressors auf die Konsolidierung der vorher präsentierten Wörter offen, so dass die Aussagekraft dieser Studie eingeschränkt ist. Eine weitere Studie, die den Einfluss eines psychosozialen Stressors auf die Gedächtnisenkodierung untersuchte, wurde von Domes et al. (2002) durchgeführt. Die Autoren setzten eine modifizierte Version des Tier Social Stress Test (TSST; Kirschbaum et al., 1993) ein. Dabei lösten die Probanden die verbale und arithmetische Aufgabe nicht vor einem Gremium, sondern vor einem Einwegspiegel. Das Gedächtnis für eine Wortliste wurde 5 Minuten nach der Enkodierung getestet. Es zeigten sich in dieser Studie keine Unterschiede in der Gedächtnisleistung zwischen Stress- und Kontrollgruppe. Allerdings konnten die Autoren zeigen, dass unabhängig von der Experimentalgruppe Probanden mit hohen Cortisolanstiegen eine verbesserte Gedächtnisleistung aufwiesen, als Probanden mit niedrigen Cortisolanstiegen. Einen anderen Befund berichten Jelicic et al. (2004). Die Autoren unterzogen Probanden entweder dem TSST oder einer Kontrollbedingung und präsentierten im Anschluss eine Liste Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 34 mit positiven, negativen und neutralen Wörtern. Probanden der Stressgruppe zeigten eine schlechtere Gedächtnisleistung für neutrales Material und gleichzeitig eine Verbesserung für emotional erregendes Material. Da sich in dieser Studie kein Effekt des Stressors auf die Cortisolspiegel fand, scheinen die Stresseffekte unabhängig von der Cortisolantwort auf den Stressor zu sein. Im Gegensatz zu dieser Studie zeigten Nater et al. (2007), dass die Cortisolantwort hauptverantwortlich für Stresseffekte auf das Gedächtnis zu sein scheint. In dieser Untersuchung zeigte sich kein genereller Effekt des TSST auf die Gedächtnisleistung einer verbalen Lernaufgabe. Eine Unterteilung der Probanden in Probanden, die mit einer starken Cortisolausschüttung auf den Stressor reagieren („high Responder“) und Probanden, die mit einer schwachen Cortisolausschüttung auf den Stressor („low Responder“) reagierten, ergab jedoch, dass „high Responder“ eine verbesserte Gedächtnisleistung zeigten, „low Responder“ hingegen nicht. In Übereinstimmung mit den Ergebnissen von Nater et al. (2007) konnten Schwabe et al (2008) nach einem modifizierten „Cold Pressure Test“ (CPS), bei dem Probanden die Hand in Eiswasser halten, ein verbessertes Gedächtnis für neutrale und negative Wörter im Vergleich zu einer Kontrollgruppe zeigen. Ein Effekt auf positive Wörter zeigte sich nicht. Während die Verbesserung des Gedächtnisses für neutrale Items unabhängig von der Cortisolantwort der Probanden auftrat, zeigte sich, dass die positiven Stresseffekte auf die negativen Items abhängig von der Cortisolantwort waren und sich lediglich für „high Responder“ zeigten. Im Gegensatz zu den positiven Befunden für neutrale Items, die von Nater et al. (2007) und Schwabe (2008) berichtet wurden, zeigten Smeets et al. (2006) negative Effekte des TSST auf die Gedächtnisleistung in einem 30 Minuten verzögerten Gedächtnistest für neutrale Wörter. In dieser Studie zeigte sich kein Effekt auf emotional erregende positive und negative Wörter. Entgegen dieser Befunde berichten Payne et al (2006) Effekte sowohl auf emotional neutrale, als auch auf emotional erregende Stimuli. Die Autoren setzten als Stressor eine modifizierte Version des TSST ein, bei dem die Versuchspersonen nicht vor einem Gremium, sondern vor einem Einwegspiegel die freie Rede halten mussten und verglichen die Leistung der Probanden mit der Leistung einer Kontrollgruppe. In dieser Studie zeigte sich, dass Probanden der Stressgruppe ein beeinträchtigtes Gedächtnis für neutrale Informationen einer Diashow zeigten, wohingegen emotionale Informationen nicht beeinträchtigt waren. Frauen der Stressgruppe zeigten eine Woche später sogar ein verbessertes Gedächtnis für emotionale Informationen. Es zeigte sich also, dass psychosozialer Stress das Gedächtnis für neutrale Reize verschlechtert, wohingegen das Gedächtnis für emotional erregende Reize erhalten Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 35 bzw. verbessert wird. Allerdings wurden in dieser Studie keine physiologischen Marker der Stressreaktion erhoben, sondern lediglich subjektive Stressratings. In einer weiteren Studie erhoben Payne et al. (2007) zusätzlich zu den subjektiven Stressratings auch die physiologische Stressantwort auf den modifizierten TSST in Form von Cortisol und Alpha-Amylase. Auch in dieser Studie zeigte sich, dass Stress vor der Enkodierung einen negativen Effekt auf das Langzeitgedächtnis für neutrale Informationen der Diashow und einen positiven Effekt das Langzeitgedächtnis für emotional erregende Inhalte ausübte. Die Befunde zum Einfluss von Stress und erhöhten Cortisolspiegeln auf die Gedächtnisenkodierung zeigen unterschiede Ergebnisse. Allerdings finden viele Studien, die die Emotionalität der Reize als Faktor mit einbeziehen, dass emotional erregende Items vornehmlich positiv und neutrale Items nicht oder negativ beeinflusst werden. 2.3.2 Stresseffekte auf die Konsolidierung Während die Befunde zum Einfluss von Stress und GCs auf die Enkodierung uneinheitlich sind und sowohl positive, als auch negative Effekte zeigen, findet sich für die Konsolidierungsphase ein einheitlicheres Bild. Tierexperimentelle Studien demonstrierten wiederholt, dass die Konsolidierungsphase von erhöhten GC-Leveln profitieren kann. So führte eine Verabreichung von GCs direkt im Anschluss an eine Lernaufgabe zu einer Verbesserung des Gedächtnisses bei Ratten. Roozendaal et al. (1999) testeten den Einfluss verschiedener Dosen eines GC- Rezeptoragonisten (3, 10 oder 30ng) auf das Vermeidungslernen bei Ratten. Die Ergebnisse zeigen eine verlängerte Aufrechterhaltung der Vermeidungsreaktion bei Tieren, die mit dem Agonisten behandelt wurden. Die Zusammenhänge waren hierbei dosisabhängig, da lediglich bei einer Dosierung von 3ng und 10ng Effekte zu beobachten waren, nicht aber bei 30ng. Eine Injektion eines beta-adrenergen-Rezeptorantagonisten in die basolaterale Amygdala 10 Minuten vor der Lernaufgabe führte dazu, dass die positiven Effekte auf die Konsolidierung verhindert wurden. Die Ergebnisse zeigen zum einen, dass die Erhöhung der GCLevel im Anschluss an eine Lernaufgabe das Gedächtnis für dieses Material im Sinne einer umgekehrten U-Funktion verbessern kann und zum anderen, dass für diese Verbesserung eine beta-adrenerge Aktivität der Amygdala notwendig ist. Während Roozendaal et al. (1999) die Bedeutung der Dosis der pharmakologischen Behandlung demonstrierten, untersuchten Sandi und Rose (1994) die Bedeutung verschiedener Zeitintervalle, in denen Hühnerkücken Corticosteron verabreicht wurde. Den Versuchstie- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 36 ren wurde entweder 15 Minuten vor dem Erlernen einer Vermeidungsreaktion, oder 5, 30, 60, 120, 180 oder 360 Minuten nach der Lernaufgabe Corticosteron injiziert. In einem Gedächtnistest 24 Stunden nach dem Training zeigte sich ein positiver Effekt von 1µg Corticosteron auf die Aufrechterhaltung der Vermeidungsreaktion, wenn es vor dem Training sowie 5, 30 und 60 Minuten nach dem Training injiziert wurde. Für die längeren Zeitintervalle zeigte sich kein Effekt. Zusätzlich konnten die Autoren demonstrieren, dass der Effekt über Glucocorticoidrezeptoren vermittelt wird. Auch für die Effekte auf die Gedächtniskonsolidierung konnte im tierexperimentellen Bereich der Einfluss von Stress auf das Gedächtnis demonstriert werden. Diamond et al. (1996) testeten den Einfluss von Stress auf eine hippocampusabhängige und eine hippocampusunabhängige Gedächtnisaufgabe bei Ratten. Dabei wurde ein Labyrinth mit 14 Armen verwendet, in dem in 7 Armen Futter zu finden war und in den verbleibenden 7 Armen niemals Futter war. Nachdem die Versuchstiere 4 Wochen lang gelernt hatten, in welchen Armen niemals Futter und in einem einmal geleerten Arm in einem Durchgang nicht erneut Futter zu erwarten war, fand der Testdurchgang statt. Die Versuchstiere wurden, nachdem sie in drei von den sieben gefüllten Armen Futter zu sich genommen hatten für vier Stunden einer unbekannten Umgebung ausgesetzt oder wurden nicht gestresst. Die gestressten Ratten zeigten im anschließenden Gedächtnistest ein schlechteres Gedächtnis für die bereits geleerten Arme. Auf das Gedächtnis für die Arme, in denen niemals Futter war fand sich kein Effekt. Daraus leiten die Autoren ab, dass Stress zu einer Beeinträchtigung der neu erlernten Informationen führte, das hippocampusunabhängige Referenzgedächtnis aber nicht beeinflusste. Während pharmakologische Studien Cortisol vor der Enkodierung der Stimuli verabreichen, werden bei Stressstudien sowohl Stressoren vor, als auch nach der Enkodierung eingesetzt. Dementsprechend liegen im Humanbereich Stressstudien, aber keine pharmakologischen Studien vor, die die Effekt von Cortisol nach der Enkodierung auf die Gedächtnisleistung untersuchen. Andreano und Cahill (2006) konnten zeigen, dass der Zusammenhang zwischen stressinduzierten Cortisolleveln und Gedächtnisleistung eine umgekehrt U-förmige Beziehung darstellt. Die Autoren teilten männliche und weibliche Probanden entweder einer Kontrollbedingung oder dem CPS zu, nachdem sie eine neutrale Geschichte gelesen hatten. Im Abruftest eine Woche später zeigte sich, dass Männer in der Stressgruppe ein besseres Gedächtnis für die Geschichte zeigten, als die Kontrollgruppe. Für Frauen zeigte sich dieser Effekt nicht. Für Männer der Stressgruppe zeigte sich zudem, dass sowohl hohe als auch niedrige Cortisolanstiege mit einer schlechteren Gedächtnisleistung einhergingen, als mäßige Cortisolanstiege. Dieser Befund demonstriert somit eine umgekehrt U-förmige Beziehung. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 37 Zudem zeigte sich in dieser Studie, dass die Effekte für Männer stärker waren, als für Frauen. Um die häufig beobachteten nicht-signifikanten oder schwächeren Effekte bei Frauen näher zu untersuchen, führten Andreano et al. (2008) eine Studie durch, bei der sie die Zusammenhänge zwischen Cortisol und Gedächtnisleistung bei Frauen in verschiedenen Zyklusphasen untersuchten. Die Frauen befanden sich entweder in der frühen Follikularphase, der späten Folikularphase oder in der Mitte der Lutealphase. Nach der Präsentation einer Textgeschichte wurden die Probandinnen entweder dem CPS oder einer Kontrollbedingung unterzogen. Es zeigte sich weder ein genereller Effekt des Stressors auf die Gedächtnisleistung in einem Gedächtnistest eine Woche nach der Enkodierung, noch eine Interaktion mit der Zyklusphase. Für die Zusammenhänge zwischen Cortisol und der Gedächtnisleistung konnte jedoch gezeigt werden, dass positive korrelative Zusammenhänge nur in der Mitte der Lutealphase, nicht aber in den anderen beiden Zyklusphasen zu beobachten waren. Cahill, Gorski und Lee (2003) gingen der Frage nach, inwieweit emotionale Erregung einen Einfluss auf die Konsolidierung hat. Dafür teilten sie die Probanden in eine Kontrollgruppe und eine Stressgruppe, die den CPS nach dem Erlernen von emotionalen und neutralen Bildern durchführte. Die Probanden der Stressgruppe zeigten eine Woche später ein besseres Gedächtnis als die Probanden der Kontrollgruppe. Diese Gedächtnisverbesserung beschränkte sich auf die emotionalen Bilder und zeigte sich nicht für die neutralen Bilder. Daraus schlossen die Autoren, dass emotionale Erregung zum Zeitpunkt der Enkodierung mit stressinduzierten Cortisolleveln interagiert und so die Konsolidierung emotional erregender Informationen verbessert. Smeets, Otgaar, Candel und Wolf (2008) testeten ebenfalls den Einfluss des CPS auf das Gedächtnis für neutrale und emotionale Wörter. Dabei verglichen sie die Effekte auf Enkodierung, Konsolidierung und Abruf, indem sie Probanden entweder vor der Enkodierung, während der Konsolidierungsphase oder vor dem verzögerten Abruf 24 Stunden nach der Enkodierung einer Wortliste mit neutralen und emotional erregenden Wörtern dem CPS unterzogen. Die Ergebnisse dieser Studie zeigen, dass Stress die Konsolidierung verbesserte, den Abruf jedoch verschlechterte. Dieser Effekt zeigte sich besonders für das emotionale Gedächtnismaterial. Zudem waren diese Effekte eng mit den stressinduzierten Cortisolleveln sowie der sympathischen Aktivierung assoziiert. Auch Studien, die psychischen Laborstress einsetzten, konnten Stresseffekte auf die Konsolidierung zeigen. Eine Studie, die den Einfluss eins psychischen Stressors auf die Konsolidierung untersuchte, stellt die Untersuchung von Beckner et al. (2006) dar. Die Autoren verglichen die Effekte von Stress auf die Konsolidierung und den Abruf von verbalen und visuellen Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 38 Informationen aus einem kurzen Film. Dafür wurden die Probanden entweder nach dem Betrachten des Films oder vor dem Abruf der Informationen gestresst. Als Stressor setzten die Autoren eine antizipierte freie Redesituation ein, die jedoch nicht durchgeführt wurde. Stress verbesserte die Konsolidierung von verbalen Informationen, hatte aber keinen Einfluss auf den Abruf der Informationen. Zudem zeigte sich ein signifikanter Zusammenhang zwischen den Cortisolleveln und dem Gedächtnis für verbale Informationen. Abercrombie et al. (2006) gingen der Frage nach, inwieweit subjektive emotionale Erregung der Versuchspersonen einen Einfluss auf die cortisolvermittelten Gedächtniseffekte hat. Dabei durchliefen Probanden nach der Enkodierung von neutralen und emotionalen Bildern eine modifizierte Version des TSST, bei der sie eine 15minütige freie Rede über ihr subjektives Empfinden während der Bilderpräsentation halten sollten. Zusätzlich wurde der negative Affekt der Versuchspersonen mittels Fragebogen erfasst. Die Ergebnisse zeigten, dass der Cortisolanstieg und der Anstieg der negativen Stimmung während des Stressors in Interaktion die Gedächtnisleistung in einem verzögerten Gedächtnistest zwei Tage später vorhersagten. Eine stärkere Cortisolausschüttung führte nur bei denjenigen Versuchspersonen zu einer besseren Gedächtnisleistung besonders für negative Bilder, die auch einen Anstieg der negativen Stimmung berichteten. Zusammenfassung Die Darstellung der Befunde zum Einfluss von Stress und Cortisol vor und nach der Enkodierung zeigt vorwiegend positive Effekte auf die Konsolidierung, während die Effekte auf die Enkodierung gemischter ausfallen. Hier stellt sich bei genauerer Betrachtung heraus, dass das Gedächtnis für neutrales Material häufig negativ beeinflusst wird, wohingegen das Gedächtnis für emotional erregendes Material positiv beeinflusst wird. Auch für die Effekte auf die Konsolidierung finden sich stärkere Effekte für emotional erregendes Material, als für neutrales Material. Aus diesen Befunden leitet sich die Annahme ab, dass GCs und erregungsinduzierte noradrenerge Aktivität in der Amygdala in Interaktion das Gedächtnis für emotional erregendes Material beeinflussen, wohingegen die Amygdala keine Rolle für die Effekte auf neutrales Material spielt (Roozendaal, Barsegyan, Lee, 2008; Roozendaal et al., 2006a; Wolf, 2008). Zudem zeigten sich in verschiedenen Studien (Andreano et al., 2008; Andreano und Cahill, 2006; Wolf et al., 2001) Geschlechtsunterschiede für die Stresseffekte auf die Gedächtnisleistung. Diese Befunde sind nicht überraschend, wenn man bedenkt, dass sich sowohl für die Gedächtnisleistung unabhängig von Stress (siehe Kapitel 1.1.3), als auch für die Stressantwort (siehe Kapitel 1.2.2.2) Geschlechtsunterschiede finden lassen. Diese Befunde verdeutlichen umso mehr die Notwendigkeit einer Berücksichtigung des Geschlechts als Ein- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 39 flussfaktor auf potenzielle Zusammenhänge zwischen Stress und verschiedenen Gedächtnisphasen. Diamond et al. (Diamond, Campbell, Park, Halonen, Zoladz, 2007) postulieren ein Modell, welches die stärkeren Effekte von Stress auf emotional erregendes Material, und hier speziell die verbesserte Konsolidierung, mit LTP-Prozessen in Hippocampus und Amygdala erklärt. Während im Hippocampus die Dauer einer verstärkte LTP direkt im Anschluss an einen Stressor zeitlich sehr gegrenzt ist und lediglich Sekunden bis Minuten andauert, bis es in einer zweiten Phase zu einer verminderten LTP kommt, dauert die Phase der verstärkten LTP in der Amygdala länger an und umfasst Minuten bis möglicherweise Stunden. Aufgrund der Interaktion zwischen Amygdala und Hippocampus bei der Verarbeitung emotional erregender Stimuli lassen sich die im Vergleich zu neutralen Stimuli verstärkten Konsolidierungseffekte für emotionale Reize erklären. Der dritte Abschnitt der Arbeit beschäftigte sich mit den Effekten von Stress und erhöhten GC-Leveln auf die Enkodierung und Konsolidierung von neutralen und emotional erregenden Stimuli. Hierfür wurde ein Überblick über die aktuelle Befundlage laborexperimenteller Studien gegeben. Eine Übersicht über die hier besprochenen humanexperimentellen Stressstudien ist in Tabelle 1 dargestellt. Das nun folgende Kapitel stellt die für die durchgeführten Untersuchungen relevanten Fragestellungen dar, die sich aus den vorliegenden Studien zu Stresseffekten auf die Gedächtnisbildung ergeben. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 40 Tabelle 1: Zusammenfassung der Stressstudien im humanexperimentellen Bereich. (Erläuterung der Symbole: „–” = negativer Stresseffekt, „+“ = positiver Stresseffekt, „+/-„ = kein Effekt) Autoren Stichprobe Treatment Abrufzeitpunkt nach Enkodierung Stimuli Ergebnis Enkodierung (Stress vor der Enkodierung) Kirschbaum et al. 1996 ♀5 ♂8 (40) TSST (Cortisol) Ca. 5 Minuten Wortliste - Wolf et al. 2001 ♀25 ♂33 TSST Ca. 5 Minuten Wortliste Negative Korrelation bei Männern Elzinga et al. 2005 ♀16 Kognitive Aufgabe Direkt 24 Stunden Wortliste - Domes et al. 2002 ♀32 TSST 5 Minuten Wortlisten Jelicic et al. 2004 ♀31 ♂9 TSST direkt Wortliste + für emotional - für neutral Nater et al. 2007 ♂20 TSST Direkt 20 Minuten Wortliste Positive Korrelation bei High-Respondern ♀48 ♂48 CPT Wortliste Wortliste + für neutral, nur bei Smeets et al. 2006 ♀30 ♂30 TSST 30 Minuten Wortliste - für neutral +/- für emotional Payne et al. 2006 ♀64 ♂53 TSST Direkt 1 Woche Bilder in einer Diashow - für neutral + für emotional bei Frauen Payne et al. 2007 ♀44 ♂32 TSST 1 Woche Bilder in einer Diashow - für neutral Smeets et al. 2008 ♀84 ♂6 CPT 24 Stunden Wortliste Schwabe 2008 et al. Positive Korrelation High-Respondern +/- für positiv + für emotional +/- Konsolidierung (Stress nach der Enkodierung) 1 Woche Neutrale schichte Ge- - bei Männern Andreano & Cahill 2006 ♀47 ♂39 CPT Andreano et al. 2008 ♀64 CPT 1 Woche Textgeschichte Cahill et al. 2003 ♀/♂ 59 CPT 1 Woche Bilder + für emotional +/- für neutral Smeets et al. 2008 ♀84 ♂6 CPT 24 Stunden Wörter + für emotional Beckner et al. 2006 ♀101 ♂56 Antizipierte freie Redesituation 48 Stunden Film Abercrombie et al. 2006 ♂34 TSST 48 Stunden Bilder umgekehrte UFunktion Positive Korrelation nur in der Lutealphase + + für emotional Abhängig von Stimmungsreaktion Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 2.4 41 Herleitung der Fragestellung Zahlreiche Studien haben sich bisher mit der Stressreaktion und ihrer Bedeutung für das Gedächtnis beschäftigt. Für den Bereich der Gedächtnisbildung sind die Befunde bisher uneinheitlich. Die Arbeiten der vorliegenden Promotionsarbeit hatten daher zum Ziel die Zusammenhänge zwischen Stress, Cortisol und der Gedächtniskonsolidierung zu untersuchen und weitere Einflussfaktoren auf diese Zusammenhänge zu betrachten. Darüber hinaus sollte die Bedeutung laborexperimenteller Untersuchungen für Alltagssituationen betrachtet werden. In einer ersten Studie wurde daher der Zusammenhang zwischen basalen Cortisolspiegeln und der Gedächtnisleistung untersucht. In der zweiten Studie wurde darauf aufbauend untersucht welchen Einfluss psychosozialer Stress auf die Gedächtniskonsolidierung hat. In der dritten Studie schließlich wurde betrachtet, welche Situationen außerhalb des Laborkontextes Stressreaktionen auslösen und durch welche Faktoren sie sich auszeichnen. Dabei wurde in einer ersten Studie zunächst der Frage nachgegangen, welchen Einfluss basale, natürlich zirkulierende Cortisolspiegel und interindividuelle Unterschiede in den Cortisolspiegeln auf das Gedächtnis haben. Zusätzlich wurde untersucht, welchen Einfluss die Emotionalität des Reizmaterials auf diese Zusammenhänge hat. Der Fragestellung, welchen Einfluss basale Cortisolspiegel auf das Gedächtnis haben, sind bisher nur wenige Studien nachgegangen. Den Einfluss von basalen Cortisolspiegeln auf die selektive Aufmerksamkeit für negative und neutrale Gesichter untersuchten van Honk et al. (2003) in einer StroopAufgabe, bei der die Probanden die Farbe nennen sollten, in der die Gesichter dargestellt waren. In dieser Studie zeigte sich, dass Probanden mit höheren basalen Cortisolspiegeln schneller die Farbe der negativen Gesichter nannten, als Probanden mit niedrigen Cortisolspiegeln. Dieser Befund deutet darauf hin, dass Probanden mit hohen basalen Cortisolspiegeln die Aufmerksamkeit von den negativen Gesichtern abwendeten und so eine schnellere Benennung der Farbe stattfinden konnte. Cortisol scheint also die Aufmerksamkeit für negative Gesichter zu modulieren. Den Zusammenhang zwischen basalen Cortisolspiegeln und dem räumlichen Gedächtnis für emotionale und neutrale Gesichter untersuchten Putman et al. (2004) in einem direkten und einem verzögerten Gedächtnistest 20 Minuten nach der Enkodierung. Die Autoren fanden ein besseres Gedächtnis für emotionale Gesichter im verzögerten Gedächtnistest bei Probanden mit höheren basalen Cortisolspiegeln. In dieser Studie hatte Cortisol demnach einen positiven und förderlichen Effekt auf die Gedächtnisbildung. Die beiden genannten Studien setzten als Reizmaterial Gesichter ein. Der Einfluss von basalen Cortisolspiegeln auf das Gedächtnis für komplexe Reize ist bislang noch nicht untersucht worden. Aus diesem Grund wurden in der ersten Studie neutrale und Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 42 emotional erregende Bilder als Reizmaterial eingesetzt, die jeweils von einem gesprochenen Satz begleitet wurden und die auf diese Weise komplexe Episoden bilden (Buchanan, Denburg, Tranel, Adolphs, 2001; Buchanan et al., 2003). Als zusätzlicher Faktor wurde zudem die Enkodierungsstrategie der Probanden manipuliert. In experimentellen Studien zum Einfluss von Cortisol auf das Gedächtnis, wurden bisher entweder explizite (Kirschbaum et al., 1996; Kuhlmann, Piel, Wolf, 2005; Wolf et al., 2001) oder implizite (Beckner et al., 2006; Buchanan und Lovallo, 2001; Cahill, 2003; Payne et al., 2007) Gedächtnistests eingesetzt. Die Probanden lernten dabei das Reizmaterial entweder in dem Wissen, dass es darauf folgend einen Gedächtnistest geben würde (intentionales Lernen) oder die Probanden wurden nicht über den folgenden Gedächtnistest informiert (implizites Lernen). Bisher wurde die Bedeutung der Enkodierungsstrategien für die Stresseffekte noch nicht untersucht, so dass deren Einfluss nicht bekannt ist. Aus diesem Grund befasste sich die erste Studie der Arbeit mit dem Einfluss basaler Cortisolspiegel auf das Gedächtnis für emotional erregende und neutrale Reize und erfasste zudem mögliche Einflüsse der Enkodierungsstrategie. Aus den Befunden laborexperimenteller Studien, die vorwiegend positive Effekte erhöhter Cortisolspiegel auf die Enkodierung und Konsolidierung von emotional erregendem Reizmaterial und keine bzw. negative Effekte auf neutrales Material finden (Buchanan und Lovallo, 2001; Cahill et al., 2003; Kuhlmann und Wolf, 2006b; Payne et al., 2007) wurde abgeleitet, dass sich dieser Zusammenhang auch bei basalen Cortisolspiegeln zeigen sollte. Für die erste Studie wurde ein positiver Zusammenhang zwischen der Höhe der basalen Cortisolspiegeln und der Gedächtnisleistung für emotional erregende Stimuli, nicht aber für neutrale Stimuli erwartet. Die zweite Studie beschäftigte sich anschließend mit dem Effekt eines psychosozialen Stressors auf die Konsolidierung von neutralen und emotional erregenden Stimuli. Als Reizmaterial wurden dabei dieselben Bilder mit Begleitsätzen verwendet, wie in Studie 1 (Buchanan et al., 2001; Buchanan et al., 2003). In Studie 2 wurde den Probanden zunächst das Reizmaterial präsentiert, bevor sie im Anschluss daran entweder den psychosozialen Stressor oder die Kontrollbedingung durchliefen. Als Stressor wurde wie in Studie 2 der TSST verwendet. Humanexperimentelle Stressstudien, die den Einfluss von Stress auf die Konsolidierung untersuchten, haben als Stressoren entweder den „Cold Pressure Test“ (CPS) eingesetzt, einen physischen Stressor bei dem die Probanden die Hand in Eiswasser halten (Andreano und Cahill, 2006; Cahill et al., 2003; Smeets et al., 2008) oder eine antizipierte freie Redesituation verwendet (Beckner et al., 2006). In dieser Studie wurde den Probanden angekündigt, dass sie vor einer Gruppe von anderen Versuchspersonen eine freie Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 43 Rede halten würden, diese freie Rede wurde allerdings nicht durchgeführt. Die Befunde dieser Studien zeigen einen positiven Effekt von Stress auf die Konsolidierung und eine damit einhergehende verbesserte Gedächtnisleistung. Da der Effekt eines psychosozialen Stressors bisher dementsprechend noch nicht untersucht wurde, beschäftigt sich die dritte Studie daher mit dem Einfluss eines durchgeführten psychosozialen Stressors auf die Konsolidierung von neutralen und emotionalen Reizen. Aufgrund der Befunde bisheriger Studien wurde ein positiver Effekt des Stressors auf die Gedächtnisleistung 24 Stunden nach der Enkodierung erwartet (Andreano und Cahill, 2006; Beckner et al., 2006; Cahill et al., 2003; Smeets et al., 2008). Dieser Effekt sollte sich verstärkt für emotional erregte Items zeigen und bei neutralen Items weniger stark oder gar nicht ausgeprägt sein (Cahill et al., 2003; Smeets et al., 2008). Studie 2 wurden mit Frauen und Männern durchgeführt. Es wurden gemischte Stichproben verwendet, um zusätzlich den Einfluss des Geschlechts der Probanden auf die Zusammenhänge zwischen Stress und Gedächtnisbildung zu untersuchen. Aus laborexperimentellen Studien ist bekannt, dass Frauen und Männer mit unterschiedlich starken physiologischen Reaktionen auf Stressexpositionen antworten. Dabei zeigen Männer häufig eine stärkere Reaktion als Frauen (Kirschbaum et al., 1999; Kirschbaum et al., 1992; Stroud et al., 2002). Dieser Befund ist jedoch nicht unumstritten, da auch Befunde vorliegen, die keinen Unterschied zwischen der Stressreaktion bei Männern und Frauen zeigen (Kelly et al., 2008). Zudem zeigte sich in verschiedenen Studien, dass Männer deutlichere Stresseffekte auf das Gedächtnis aufweisen, als Frauen (Andreano und Cahill, 2006; Cahill, 2003; Wolf et al., 2001). Daher wurde in den Studien 2 zusätzlich der Faktor Geschlecht betrachtet. Es wurde erwartet, dass die Befunde für Männer stärker ausgeprägt sind, als für Frauen. Die dritte Studie der vorliegenden Promotionsarbeit beschäftigte sich mit Stressoren außerhalb des Laborkontextes. Dabei wurde der Frage nachgegangen, welche Situationen im Alltag Stress verursachen und zu einer physiologischen Stressantwort führen. Aus laborexperimentellen Untersuchungen sind verschiedene Faktoren bekannt, die im Labor signifikante Stressreaktionen bei Versuchspersonen auslösen. In einer Metaanalyse (Dickerson und Kemeny, 2004) zeigte sich, dass die Bedrohung des sozialen Selbst ein wichtiger Faktor ist. In Laborsituationen in denen dieser Faktor in Kombination mit der Unkontrollierbarkeit der Situation zu signifikant stärkeren Stressreaktionen führte, als Laborsituationen, in denen diese Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 44 Faktoren nicht existierten. Die dritte Studie ging darauf aufbauend der Frage nach, ob diese Faktoren auch außerhalb von laborexperimentellen Settings Stressreaktionen auslösen, welche Faktoren also bei naturalistischen Stressoren wirken. Im Bereich der naturalistischen Stressoren bieten sich Prüfungssituationen als Stresssituationen an und eine Vielzahl an Studien hat die Stressreaktion in Prüfungssituationen beobachtet (Stowell, 2003). Dabei zeigten sich allerdings gemischte Befunde. Während einige Studien erhöhte Cortisolspiegel in Reaktion auf eine Prüfungssituation berichteten (Herbert et al., 1986; Lacey et al., 2000; Loft et al., 2007; Lovallo et al., 1986; Schoofs et al., 2008), fanden andere Studien reduzierte Cortisolspiegel (Spangler, 1997) . Diese Studien haben allerdings sehr unterschiedliche Prüfungssituationen, wie schriftliche (Gaab et al., 2006; Ng, Koh, Chia, 2003) oder mündliche (Herbert et al., 1986; Lacey et al., 2000; Schoofs et al., 2008) Prüfungen, einzelne (Frankenhaeuser et al., 1978) oder Prüfungsphasen (Loft et al., 2007), Matrikulationsprüfungen (Frankenhaeuser et al., 1978) oder Abschlussprüfungen an der Universität (Zeller et al., 2004) untersucht. Eine mögliche Ursache für die uneinheitlichen Befunde könnte daher sein, dass sich die verschiedenen Prüfungssituationen stark voneinander unterscheiden. Hierbei ist eine Vielzahl von Faktoren denkbar. Aus dem Bereich der Laborstressoren sind die Faktoren Bedrohung des sozialen Selbst, Unkontrollierbarkeit und persönliche Involviertheit bekannt. Es ist denkbar, dass auch bei naturalistischen Stressoren diese Faktoren einen starken Einfluss auf die Stressreaktion haben. Daher könnten sich die heterogenen Befunde der Prüfungsstudien mit Unterschieden in der Stärke der Bedrohung des sozialen Selbst sowie der Unkontrollierbarkeit der Situation erklären lassen. Um den Einfluss dieser Faktoren näher zu betrachten, wurden in der dritten Studie zum einen eine schriftliche Prüfungssituation und zum anderen eine Referatssituation untersucht. Diese beiden Situationen sollten sich in Bezug auf die Stärke der sozialen Bedrohung und der Unkontrollierbarkeit unterscheiden. Es wurde erwartet, dass die Referatssituation zu einer starken Stressreaktion führen, die anonymere schriftliche Prüfungssituation jedoch eine weniger starke Reaktion auslösen sollte. Zudem wurde für die schriftliche Prüfung eine antizipatorische Stressreaktion erwartet, die sich in erhöhten Cortisolspiegeln an dem Tag vor der Prüfung zeigen sollte. Studie 1 2 STUDIE 1: ASSOCIATIONS 45 BET WEEN ENDOGENOUS CORTISOL LEVELS AND M EM ORY IN Y OUNG WOM EN: INFLUENCE OF ENCODING INST RUCT IONS Studie 1: Associations between endogenous cortisol levels and emotional memory: influence of encoding instructions Diana Preuß, Daniela Schoofs, Oliver T. Wolf Stress, 2008; iFirst: 1–9 Associations between endogenous cortisol levels and emotional memory in young women: Influence of encoding instructions DIANA PREUß, DANIELA SCHOOFS, & OLIVER T. WOLF Department of Cognitive Psychology, Ruhr-University Bochum, Bochum, Germany Downloaded By: [Wolf, Oliver T.] At: 10:36 13 November 2008 (Received 26 May 2008; revised 1 August 2008; accepted 2 October 2008) Abstract The stress hormone cortisol is known to influence memory. Elevated cortisol levels as a consequence of stress or as a consequence of cortisol administration have been repeatedly shown to enhance encoding and consolidation of (emotional) memory. Whether similar associations exist between basal cortisol levels and emotional memory remains to be established. The present study therefore evaluated if resting cortisol levels are correlated with memory for emotionally arousing and neutral pictures in a sample of young healthy females (n ¼ 56). A second aim of the study was to explore if the relationship between basal cortisol levels and memory might be modulated by encoding instructions (intentional vs. incidental encoding). A significant positive correlation between basal salivary cortisol levels and memory for emotionally arousing pictures in a 24 h delayed free recall test was found. Further analyses revealed that this association only occurred in the group receiving intentional encoding instructions. Results indicate that basal cortisol levels, similarly to stress induced cortisol levels, are associated with emotional memory formation. Moreover this effect seems to be modulated by encoding instructions, suggesting a role of focussed attention or arousal induced by testing in this relationship. Keywords: Arousal, cortisol, emotional memory, emotional enhancement effect, encoding instructions, salivary samples Introduction In response to stress the hypothalamic-pituitaryadrenal axis is activated which leads to an increased secretion of cortisol (McEwen 2000; Sapolsky et al. 2000). Studies investigating the influence of elevated cortisol levels on memory have found enhancing as well as impairing effects (Wolf 2008). For example increased cortisol secretion, either in response to stress or after pharmacological cortisol administration, impairs memory retrieval (de Quervain et al. 1998; Kuhlmann and Wolf 2005; Kuhlmann et al. 2005a; Buchanan et al. 2006). In contrast (emotional) memory encoding and consolidation appears to be enhanced. Several studies have found beneficial effects of elevated cortisol levels at times of encoding or consolidation (Cahill et al. 2003; Andreano and Cahill 2006; Beckner et al. 2006; Kuhlmann and Wolf 2006b; Payne et al. 2007). This enhancing effect of cortisol on memory was often more pronounced for emotional arousing material (Buchanan and Lovallo 2001; Cahill et al. 2003; Kuhlmann and Wolf 2006b; Payne et al. 2006, 2007; Smeets et al. 2006). Cortisol thus appears to potentiate the emotional enhancement effect (LaBar and Cabeza 2006). Studies in rodents have revealed that glucocorticoids interact with arousal-induced noradrenergic activation in the amygdala, thereby strengthening memory consolidation in the adjacent hippocampus (Roozendaal et al. 2006a). Cortisol levels not only increase in response to stress, but also show a strong circadian rhythm, characterized by a continuous decline over the course of the day (Kirschbaum and Hellhammer 1989). In addition, the interindividual variance in endogenous cortisol levels is substantial. In contrast to the experimental studies summarized above, studies that examine the relationship between basal cortisol levels and emotional memory in young healthy subjects are rare to date and have not shown consistent results (van Honk et al. Correspondence: O. T. Wolf, Department of Cognitive Psychology, Ruhr-University Bochum, Universitätsstr. 150, D-44780 Bochum, Germany. Tel: 49 234 32 22670. Fax: 49 234 32 14308. E-mail: [email protected] ISSN 1025-3890 print/ISSN 1607-8888 online q 2008 Informa USA, Inc. DOI: 10.1080/10253890802524592 Downloaded By: [Wolf, Oliver T.] At: 10:36 13 November 2008 2 D. Preuß et al. 2003; Putman et al. 2004). The question of whether cortisol and arousal interact not only in cases of stress or pharmaco-induced cortisol elevations, but also under resting conditions remains unanswered. The current study therefore was conducted to test the associations between emotional memory and basal salivary cortisol levels. Previous research has shown enhancing effects of elevated cortisol levels on encoding and consolidation especially for emotional arousing items (Buchanan and Lovallo 2001; Cahill et al. 2003; Kuhlmann and Wolf 2006b; Payne et al. 2007). Therefore a positive correlation was expected between cortisol level and emotional memory. Previous studies on the topic of cortisol and emotional memory have used either incidental (Buchanan and Lovallo 2001; Cahill et al. 2003; Beckner et al. 2006; Payne et al. 2007) or intentional (Kirschbaum et al. 1996; Wolf et al. 2001; Kuhlmann and Wolf 2005; Kuhlmann et al. 2005b) encoding instructions. Differences in encoding instructions might lead to differences in attention or to altered task-induced arousal as a consequence of an increased motivation to remember the items. In addition, different brain regions might be involved in incidental vs. intentional encoding (Grady et al. 1998; Bernstein et al. 2002; Stark and Okado 2003). Whether the associations between cortisol levels and memory might differ between incidental and intentional encoding instructions was a second exploratory aim of the study. Materials and method Sixty young female subjects aged between 19 and 35 years (23.7 ^ 0.44 years; mean ^ SEM) participated in this study. Four participants were excluded because of an insufficient amount of saliva for cortisol measurement. No information about the use of hormonal contraceptives or the stage of the natural menstrual cycle was collected. All participants were recruited from the university campus and written informed consent was collected from each subject. The study was approved by the national ethic committee of the German Psychological Association (Deutsche Gesellschaft für Psychologie). Participants were tested individually on two consecutive days 24 h apart. Testing took place between 9 am and 4.30 pm Fifty five percent of the subjects were tested before 12 pm. On the first day participants filled out a mood questionnaire (for description see below). Afterwards the memory material was presented. Participants were randomly assigned to one of the two encoding conditions. One-half of the participants were told to memorize the pictures and narratives as well as possible (intentional encoding), whereas the other half was not instructed to memorize (incidental encoding). Additionally, subjects under both conditions were informed that we were interested in their physiological reaction to the stimuli and that they therefore should empathise strongly to the individual scenes. Immediately after watching the pictures, participants filled out the mood questionnaire again. On the second day the memory tests occurred, which were unexpected for the participants in the incidental encoding condition, and were expected for the participants who received the intentional encoding instruction. After completing the memory tests on the second day, participants were asked to rate the stimuli on a 5-point Likert scale for emotional arousal. For arousal, 1 refers to the less arousing picture and 5 to the most arousing picture. Stimuli The stimuli and memory tests used in the present study were developed and validated in previous studies (Buchanan et al. 2001, 2003). The stimuli consisted of 5 positive (e.g. two happy girls eating ice-cream), 5 negative (e.g. a diseased child from Africa with bandages and cannulae) and 5 neutral (e.g. people leaving or entering a building) pictures, which were presented in a fixed order for 10 s each on a computer screen. Several of the pictures were chosen from the International Affective Picture System (IAPS; Lang et al. 1997) and the rest were drawn from print media sources. Each picture was accompanied by a single narrative sentence presented via ear-phones which consisted of information that was not obvious in the picture. For example, the picture with the little girls eating ice-cream was accompanied by a sentence in which the girls’ names and the special kind of ice-cream they liked were mentioned. Based on the finding that amygdala activity is linked to emotional arousal rather than valence (Kensinger 2004), and based on previous stress/cortisol studies indicating that emotional arousal rather than valence determines the strength of the cortisol effects (Buchanan and Lovallo 2001; Cahill et al. 2003; Kuhlmann and Wolf 2006b; Payne et al. 2006, 2007; Smeets et al. 2006), the two emotional valence categories (positive and negative pictures) were combined and data averaged, thereby creating a single category of emotionally arousing items. Memory tasks The memory tests took place 24 h after the encoding session. Firstly, the participants performed a free-recall test, for which they were told that they would have 5 min to write down everything they could remember from the pictures and narratives. In addition to the verbal instruction, participants received this instruction in written form at the top of the answer sheet. Answers were evaluated by two independent judges, who were blind to the group membership of the subjects. The agreement between the two raters was evaluated with an intra-class correlation for the total free recall Downloaded By: [Wolf, Oliver T.] At: 10:36 13 November 2008 Cortisol and emotional memory score. The intra-class correlation coefficient was high (ricc ¼ 0.912, p , 0.001) indicating good reliability. Therefore, the average score of the two raters was used. Participants scored 2 points, if the information they wrote down could be associated clearly to one of the pictures and was correct in details. One point was given for information that could be associated clearly to one of the pictures but consisted of some incorrect details. If the information was completely incorrect or could not be linked to one picture, participants got 0 points. In total the participants could score a maximum of 30 points. The second test was a four-alternative multiplechoice test with six questions for each stimulus, which asked for information from pictures and narratives. In this task, every correct answer scored one point, so that a total of 90 points was possible. This test also allowed differentiating between memory for gist and for detail information. Gist is defined as “information which could not be changed or excluded without changing the basic story line” (Heuer and Reisberg 1990). According to Buchanan et al. (2003), 47 of the questions referred to gist information and 37 to detail information. The remaining six items could not be categorized clearly and were thus not included in the gist analysis (Buchanan et al. 2003). Here again every correct answer scored one point, so that 47 points for the gist items and 37 points for detail items were achievable. In order to allow comparisons between memory for gist and memory for detail memory performance was expressed in percentages. Questionnaires As an indicator of mood the “Positive and Negative Affective Scale” (PANAS, Watson et al. 1988) was used. The questionnaire consists of 20 adjectives, which are summarized into one scale for positive and one scale for negative affect. For each adjective participants had to mark on a five point Likert scale, how far the adjective described their actual state. Sums of the scale marks were made by addition of the answers to the respective items (resulting in a minimum sum of 10 per scale). Participants filled out the questionnaire twice, once before presentation of the stimuli and for the second time after the presentation. Saliva sample Saliva was collected using Salivette collection devices (Sarstedt, Nümbrecht, Germany). Cortisol concentrations were measured using an immunoassay (IBL, Hamburg, Germany). Inter- and intra assay variations were below 15%. Two samples were taken, one before presentation of the stimuli and the second one immediately after presentation was completed (approximately 5 min after the first saliva sample). Two samples were taken in order to obtain a more reliable and valid basal cortisol measure. Testing and 3 thus salivary sampling was spread over the day (ranging from 9 am to 4:30 pm). Statistical analysis Data were analyzed with Spearman t-tests or ANOVAs for repeated measurements and post hoc adjusted paired t-tests. Greenhouse-Geisser corrected p values were used when indicated. Because cortisol measures did not show a normal distribution, data were log 10 transformed to approximate them to a Gaussian distribution. After transformation all data were normally distributed. Results Arousal ratings The data of the arousal ratings for the pictures that were made after completion of the memory tests on the second day were analyzed using ANOVA with the within subject factor arousal (arousing vs. neutral items) and the between subject factor encoding condition (incidental vs. intentional encoding). A significant main effect of arousal was detected [F(1,54) ¼ 148.041, p , 0.001]. No main effect for encoding condition was detected [F(1,54) ¼ 2.240, p ¼ 0.140]. The interaction with encoding condition did not reach significance [F(1,54) ¼ 0.458, p ¼ 0.501]. Participants rated the arousing items with values of 3.80 (^0.06) and the neutral items as 2.55 (^0.09); post hoc t-tests showed that this difference was significant [t(55) ¼ 212.211, p , 0.001]. Influence of arousal and encoding condition on memory retrieval An ANOVA with the within subject factor arousal (arousing vs. neutral items) and the between subject factor encoding condition (incidental vs. intentional encoding) was conducted for each memory test separately. There was a significant main effect of arousal for the free recall test [F(1,54) ¼ 71.784, p , 0.001] and the multiple choice test [F(1,54) ¼ 16.996, p , 0.001]. Analyses did not show main effects of encoding condition [free recall: F(1,54) ¼ 0.009, p ¼ 0.925; multiple choice: F(1,54) ¼ 0.617, p ¼ 0.435] or interaction effects between arousal and encoding condition [free recall: F(1,54) ¼ 1.616, p ¼ 0.209; multiple choice: F(1,54) ¼ 0.002, p ¼ 0.961]. To investigate the significant main effect of arousal further, we conducted paired t-tests for both memory tests. In both tests, neutral items were recalled significantly less than arousing items [free recall: t(55) ¼ 2 8.386, p , 0.001; multiple choice: t(55) ¼ 24.165, p , 0.001]. Results are displayed in Figure 1(a),(b) respectively. Additionally an ANOVA with the within subject factors gist (gist vs. detail), arousal (arousing vs. neutral items) and the between subject factor encoding condition (incidental vs. intentional encoding) was Downloaded By: [Wolf, Oliver T.] At: 10:36 13 November 2008 4 D. Preuß et al. Figure 1. Mean memory scores for arousing and neutral items in the free recall test (a), the multiple choice test (b) and for gist and detail analysis of the multiple choice test (c). All comparisons are with paired t-tests. Participants reached a higher memory score for arousing items compared to neutral ones in the free recall test (**p , 0.001) and in the multiple choice test (**p , 0.001). Participants reached higher memory scores (expressed in percentages) for arousing (**p , 0.001) and neutral (*p , 0.05) gist information than for detail information. For the gist of the stimuli the participants remembered more emotional than neutral information (**p , 0.001). Values are mean ^ SEM. conducted. Results revealed a main effect for arousal [F(1,54) ¼ 22.044, p , 0.001] and gist [F(1,54) ¼ 66.819, p , 0.001] and a significant interaction effect between arousal and gist [F(1,54) ¼ 18.005, p , 0.05]. No main effect of encoding condition [F(1,54) ¼ 0.770, p ¼ 0.384] was detected and none of the possible interactions with this factor were significant (all p . 0.20). Post hoc t-tests showed that participants remembered more gist than detail information for emotionally arousing [t(55) ¼ 11.983, p , 0.001] and neutral items [t(55) ¼ 2.426, p , 0.05]. In addition for the gist of the stimuli participants remembered more emotional than neutral information [t(55) ¼ 29.971, p , 0.001] while no such effect was observed for details [t(55) ¼ 20.647, p ¼ 0.521]. Results are displayed in Figure 1(c). Salivary cortisol Salivary cortisol concentrations decreased slightly but significantly between the two sampling points from 7.02 (^ 0.54) to 5.83 (^ 0.44) nmol/l [t(55) ¼ 4.033, p , 0.001]; the t-test was conducted with logtransformed data. In order to create a single measure indicative of the basal cortisol levels during the memory task the average of the two measures was taken. For nine participants, only one of the two saliva samples contained enough fluid for the analysis. In those cases, the available measure of the participant was used as the best estimator. Mean cortisol levels were 6.43 ^ 0.47 nmol/l. Associations between cortisol and emotional memory Analysis for the entire sample. Bivariate Pearsons’s correlations between the mean salivary cortisol concentration and the memory scores revealed a significant positive correlation between cortisol concentration and memory for arousing items in the free recall task (r ¼ 0.295, p , 0.05). A smaller and nonsignificant correlation emerged for the neutral items (r ¼ 0.122, p ¼ 0.372). The scatter plots are presented in Figure 2. For the multiple choice test no significant correlation was observed (arousing items r ¼ 20.054, p ¼ 0.693; neutral items r ¼ 0.163, p ¼ 0.231). Because of the well-known circadian rhythm of cortisol secretion (Kirschbaum and Hellhammer 1989), we additionally conducted a partial correlation analysis which controlled for time of day. This analysis ascertained that the association between salivary cortisol concentration and emotional memory was not secondary to an unspecific effect of the circadian rhythm. The correlation coefficient between salivary cortisol concentration and free recall of arousing items became only slightly smaller when time of day was partialed out (r ¼ 0.263, p ¼ 0.052). Fisher’s z-test indicated that the correlations between arousing items and cortisol concentration Cortisol and emotional memory 5 Downloaded By: [Wolf, Oliver T.] At: 10:36 13 November 2008 Figure 2. Scatter plots for the correlations between log transformed salivary cortisol concentration data and memory performance in the free recall test (n ¼ 56) for (a) emotionally arousing items (r ¼ 0.295; p , 0.05), (b) neutral items, not significant (r ¼ 0.112, p ¼ 0.372). differed significantly from the correlation between neutral items and cortisol concentration. This was true for the correlations without (z ¼ 0.375, p , 0.05) and with the control for time of day (z ¼ 0.382, p , 0.05). Influence of encoding condition To evaluate whether intentional vs. incidental encoding influenced the observed associations between cortisol and emotional memory Pearson’s correlations were conducted for the intentional and incidental group separately. Results revealed significant correlations between salivary cortisol concentration and memory for the arousing items only in the intentional encoding group (see Table I and Figure 3). A similar significant correlation was obtained when time of day was controlled for. In contrast, no association was observed between cortisol concentration and memory in the incidental encoding group (see Table I and Figure 3). Additionally Fisher z-values were calculated to evaluate if the strength of the correlation differed between the intentional and incidental encoding group. The correlations between arousing items and cortisol concentration did significantly differ between the two groups (r ¼ 0.533, p , 0.01). The significant difference remained when controlling for time of day (z ¼ 0.626, p , 0.01). Exploratory analysis Finally, for the significant correlation between emotional memory and cortisol concentration within the intentional learning group the potential influence of two modulating factors were explored, namely time of day and gist vs. details. Influence of time of day Previous work has suggested that the effects of stress or cortisol treatment might differ depending on the time of day (morning vs. afternoon; Het et al. 2005; Maheu et al. 2005). So far our analysis had revealed that the associations between cortisol and emotional memory persisted, when time of day was controlled for. In order to investigate this issue further, we conducted two separate correlations for those subjects from the intentional encoding group who were tested in the morning and those subjects who were tested in the afternoon. In the morning, the correlation was r ¼ 0.542, n ¼ 16, p , 0.05. In the afternoon, the correlation was still sizable but non-significant r ¼ 0.385, n ¼ 11, p ¼ 0.242. Table I. Correlations between basal salivary cortisol concentrations and memory for arousing and neutral items, computed separately for the intentional and incidental encoding group. Cortisol Memory for arousing items Intentional encoding (n ¼ 27) Incidental encoding (n ¼ 29) Memory for neutral items Intentional encoding (n ¼ 27) Incidental encoding (n ¼ 29) Without time of day as a covariate With time of day as a covariate r ¼ 0.482 p , 0.05 r ¼ 20.007 p ¼ 0.969 r ¼ 0.485 p , 0.05 r ¼ 20.096 p ¼ 0.626 r ¼ 0.339 p ¼ 0.083 r ¼ 20.223 p ¼ 0.245 r ¼ 0.313 p ¼ 0.120 r ¼ 20.199 p ¼ 0.310 Results are presented with and without time of day as a covariate. 6 D. Preuß et al. Figure 3. Scatter plots for the correlations between log transformed cortisol data and memory performance in the free recall test for emotionally arousing items in a: the intentional encoding group, significant (r ¼ 0.482, n ¼ 27, p , 0.05), b: the incidental encoding group, not significant (r ¼ 20.007, n ¼ 29, p ¼ 0.969). Downloaded By: [Wolf, Oliver T.] At: 10:36 13 November 2008 Gist vs. details In the incidental encoding group cortisol concentration was correlated with emotional memory when tested with free recall, but not when tested with cued recall (see above). To investigate whether in the cued recall test cortisol level might be specifically related to memory for gist or details a bivariate Pearson’s correlation was conducted between cortisol concentration and memory scores for arousing gist and details. However, the correlation did not reach significance for gists (r ¼ 0.159, n ¼ 27, p ¼ 0.429) or for details (r ¼ 0.141, n ¼ 27, p ¼ 0.484). Associations between mood and cortisol To assess changes in mood an ANOVA with the inner subject factors time (pre- vs. post-measurement) and the between subject factor encoding condition (incidental vs. intentional encoding) was conducted for each of the two scales separately. For positive mood there was a significant main effect of time [F(1,54) ¼ 96.529, p , 0.001]. No main effect of encoding condition [F(1,54) ¼ 0.246, p ¼ 0.622] and no interaction effect between time and encoding condition [F(1,54) ¼ 1.215, p ¼ 0.275] was detected. For negative mood again a significant main effect of time occurred [F(1,54) ¼ 17.123, p , 0.001]. No main effect of encoding condition [F(1,54) ¼ 0.937, p ¼ 0.337] and no interaction effect between time and encoding condition [F(1,54) ¼ 2.954, p ¼ 0.091] was detected. Post hoc t-tests revealed that participants reported an increase in negative mood [t(55) ¼ 24.009, p , 0.001] and a decrease in positive mood [t(55) ¼ 9.851, p , 0.001] after the slide presentation. To investigate possible associations between emotional reactivity and basal cortisol levels bivariate Pearson’s correlation were conducted. To examine the influence of changes in mood as a reaction to the stimuli, we computed a delta value for negative emotional reactivity (negative mood after presentation—negative mood before presentation of the stimuli) and positiveemotional reactivity (positive mood after presentation— positive mood before presentation of the stimuli). No significant correlations were found between subject’s mood changes and average cortisol levels ( p . 0.05). Discussion The present study was conducted to evaluate the relationship between basal salivary cortisol concentrations and memory for emotional arousing and neutral pictures. A second aim of the study was to evaluate possible influences of encoding strategies on this effect. The results revealed a positive relationship between basal cortisol levels and memory for arousing items in the free recall test. Subjects with higher endogenous cortisol levels during the encoding of emotional arousing material showed superior memory for these items 24 h later. This finding fits the assumption that arousal and cortisol interact to modulate memory (Roozendaal 2002; Wolf 2008). Our finding is in line with previous human studies which found that stress prior to encoding (Payne et al. 2007) or directly after encoding (Cahill et al. 2003; Andreano and Cahill 2006) leads to enhanced emotional memory consolidation. Similarly, pharmacological studies found enhanced emotional memory after cortisol treatment (Buchanan and Lovallo 2001; Kuhlmann and Wolf 2006b). The current results also fit well to a study by Putman et al. (2004), who observed that higher endogenous cortisol levels were associated with better memory for emotional faces tested 20 min after encoding. Our observations indicate that these facilitating effects of cortisol on encoding/consolidation can also be observed for basal cortisol levels in a 24 h recall paradigm. The present finding of a positive correlation between basal cortisol levels and memory for arousing items expands findings from stress studies to situations of endogenous Downloaded By: [Wolf, Oliver T.] At: 10:36 13 November 2008 Cortisol and emotional memory cortisol fluctuations. Studies in rodents have revealed that glucocorticoids interact with noradrenergic activation in the amygdala to enhance memory consolidation (Roozendaal et al. 2006b). In support of these findings is a recent human neuroimaging study which observed that participants with higher endogenous cortisol levels showed a stronger amygdala response to emotionally arousing slides (van Stegeren et al. 2007). A similar scenario might underlie our present results. A second aim of the study was to evaluate if the relationships between basal salivary cortisol levels and emotional memory differ depending on the encoding strategies that were used. This analysis indicated that only participants with explicit encoding instruction (intentional encoding) showed a positive correlation between cortisol level and emotional memory. Thus the results discussed above were exclusively driven by this group. No association between cortisol and emotional memory was observed in the incidental encoding group and the correlations differed significantly between the two conditions. Previous studies investigating the effects of stress on emotional memory encoding or consolidation have used intentional (Kirschbaum et al. 1996; Wolf et al. 2001) as well as incidental (Cahill et al. 2003; Beckner et al. 2006) encoding instructions. Similarly previous pharmacological studies used intentional (Kuhlmann and Wolf 2005; Kuhlmann et al. 2005a) as well as incidental (Buchanan and Lovallo 2001) instructions. Our study looking at basal cortisol levels suggest that cortisol influences emotional memory encoding/ consolidation only in situations of intended learning and directed attention. Emotional items are known to enhance activation of the amygdala (Cahill et al. 1996; Hamann et al. 1999) and influences of cortisol rely on its interaction with noradrenergic activation in the basolateral amygdala (Cahill and McGaugh 1996; Roozendaal et al. 2006a). Recent neuroimaging evidence suggests that attention and emotional arousal sometimes interact in an additive fashion (Vuilleumier et al. 2001). Our findings might suggest that basal cortisol levels can only modulate memory when both factors (emotional arousal and attention) are present. Another reason for the finding of an enhanced emotional memory in participants with higher basal cortisol levels and intentional encoding might be the enhanced testing induced arousal associated with the announced memory test. In a previous study, we observed that a non-arousing test situation abolished the effects of cortisol treatment on memory (Kuhlmann and Wolf 2006a). The effects on memory were only observed in test situations that induced arousal. This result was highly similar to observations made in rodents (Okuda et al. 2004). Test-induced arousal therefore seems to be a prerequisite for cortisol to influence memory, at least in pharmacological studies and possibly also in studies investigating associations 7 between basal cortisol levels and memory. The incidental encoding group might have lacked the necessary test-induced arousal. The present results therefore suggest that heightened attention and/or test-induced arousal might strengthen the relationship between basal cortisol levels and emotional memory formation. Cortisol is known to have a strong circadian rhythm which is characterized by a continuous decline over the course of the day in humans. It is therefore important to note that the correlation between cortisol and emotional memory persisted when time of day was controlled for. Some experimental stress studies (Maheu et al. 2005) and some pharmacological studies (Het et al. 2005) suggest that the effects of cortisol might vary depending on the time of day. Negative effects might occur in the morning, when basal cortisol levels are already relatively high. These effects might reflect an inverted U-shaped dose response curve between cortisol and memory. Indeed an inverted U-shaped relationship has been observed in some pharmacological studies (Lupien et al. 1999; Abercrombie et al. 2003; Domes et al. 2005) and a recent stress study (Andreano and Cahill 2006), but results are far from consistent. Our study suggests that at least for basal cortisol levels the relationship between cortisol and emotional memory formation appears to be linear. In our study, we only found (within the intentional encoding group) significant associations between cortisol and free recall performance, but not between cortisol and a multiple choice cued recall task. This is in line with several previous studies (de Quervain et al. 2003; Kuhlmann et al. 2005b; Kuhlmann and Wolf 2006b). Within the cued recall condition cortisol was neither related to emotional memory for gist nor for details. Studies in patients with amygdala lesions suggest that the amygdala is especially important for the gist memory of an emotional episode (Adolphs et al. 2005). Based on this clinical finding one could have hypothesized that cortisol is related to the gist of the emotional material. In the current study, we did not find any association between cortisol and cued recall in general, suggesting that this measure, at least in this specific task, is not sensitive to cortisol effects. Moreover cued recall memory for emotional gist was evidently good, which might have reduced the possibility to find associations with the cortisol measure. There are several limitations of the current study which need to be considered. First of all, we cannot differentiate associations with emotional memory encoding from associations with emotional memory consolidation. The use of an immediate retrieval test similar to Kuhlmann and Wolf (2006b) would have allowed distinction of immediate from delayed effects. However, since we were interested in differentiating between intentional and incidental encoding, two separate repeated retrieval tests were not feasible. Downloaded By: [Wolf, Oliver T.] At: 10:36 13 November 2008 8 D. Preuß et al. Based on our previous findings (Kuhlmann and Wolf 2006b) and those of others (Cahill et al. 2003), we speculate that the association is driven by an effect of cortisol on emotional memory consolidation, but this remains to be firmly established. Secondly, the present study included only a female sample. Conclusions from this study therefore cannot be extended to males. Some previous studies have reported sex differences for the relationship between cortisol and memory (Wolf et al. 2001; Cahill 2003; Andreano and Cahill 2006) and between cortisol and emotional learning (Jackson et al. 2006; Stark et al. 2006; Zorawski et al. 2006). Therefore it still has to be shown whether basal cortisol levels also relate to emotional long-term memory in males. A third limitation of the study is the fact that we did not collect information about the menstrual cycle phase or use of oral contraceptives. There are reports suggesting that associations between cortisol and memory might only occur at a specific menstrual cycle phase (Andreano et al. 2008). Moreover, oral contraceptives appear to lead to a reduced sensitivity to cortisol, at least when it is given pharmacologically (Kuhlmann and Wolf 2005). Future studies are needed in order to explore the potential impact of sex steroids on the association between endogenous cortisol levels and emotional memory. A fourth restriction to the present study is the fact that we did not use psychophysiological measures of arousal. It would have been important to test whether or not the intentional and incidental encoding instructions were associated with different levels of physiological arousal. In sum we report a positive correlation between basal cortisol levels and memory for arousing items in a sample of healthy women. Interestingly this association was only apparent in those subjects who had received explicit encoding instructions. Thus, focused attention or a stronger test-induced arousal might be a prerequisite for the occurrence of beneficial cortisol effects on emotional memory, at least when tested under resting (non-stress) conditions. Our findings illustrate that cortisol is related to emotional memory not only under circumstances of stress or pharmacological glucocorticoid treatment, but also in situations when cortisol levels vary within the basal range. Acknowledgements This study was supported by a grant from the German Research Foundation (DFG WO 733/7-1). We wish to thank Tony Buchanan (Department of Psychology Saint Louis University) for providing us with the emotional memory task used in this study. Declaration of interest: The authors report no conflicts of interest. The authors alone are responsible for the content and writing of the paper. References Abercrombie HC, Kalin NH, Thurow ME, Rosenkranz MA, Davidson RJ. 2003. Cortisol variation in humans affects memory for emotionally laden and neutral information. Behav Neurosci 117:505–516. Adolphs R, Tranel D, Buchanan TW. 2005. Amygdala damage impairs emotional memory for gist but not details of complex stimuli. Nat Neurosci 8:512–518. Andreano JM, Cahill L. 2006. Glucocorticoid release and memory consolidation in men and women. Psychol Sci 17:466– 470. Andreano JM, Arjomandi H, Cahill L. 2008. Menstrual cycle modulation of the relationship between cortisol and long-term memory. Psychoneuroendocrinology 33:874–882. Beckner VE, Tucker DM, Delville Y, Mohr DC. 2006. Stress facilitates consolidation of verbal memory for a film but does not affect retrieval. Behav Neurosci 120:518–527. Bernstein LJ, Beig S, Siegenthaler AL, Grady CL. 2002. The effect of encoding strategy on the neural correlates of memory for faces. Neuropsychologia 40:86– 98. Buchanan TW, Lovallo WR. 2001. Enhanced memory for emotional material following stress-level cortisol treatment in humans. Psychoneuroendocrinology 26:307–317. Buchanan TW, Denburg NL, Tranel D, Adolphs R. 2001. Verbal and nonverbal emotional memory following unilateral amygdala damage. Learn Mem 8:326–335. Buchanan TW, Karafin MS, Adolphs R. 2003. Selective effects of triazolam on memory for emotional, relative to neutral, stimuli: Differential effects on gist versus detail. Behav Neurosci 117: 517 –525. Buchanan TW, Tranel D, Adolphs R. 2006. Impaired memory retrieval correlates with individual differences in cortisol response but not autonomic response. Learn Mem 13:382–387. Cahill L. 2003. Sex- and hemisphere-related influences on the neurobiology of emotionally influenced memory. Prog Neuropsychopharmacol Biol Psychiatry 27:1235–1241. Cahill L, McGaugh JL. 1996. Modulation of memory storage. Curr Opin Neurobiol 6:237–242. Cahill L, Haier RJ, Fallon J, Alkire MT, Tang C, Keator D, et al. 1996. Amygdala activity at encoding correlated with long-term, free recall of emotional information. Proc Natl Acad Sci USA 93: 8016–8021. Cahill L, Gorski L, Le K. 2003. Enhanced human memory consolidation with post-learning stress: Interaction with the degree of arousal at encoding. Learn Mem 10:270–274. Domes G, Rothfischer J, Reichwald U, Hautzinger M. 2005. Inverted-U function between salivary cortisol and retrieval of verbal memory after hydrocortisone treatment. Behav Neurosci 119:512–517. Grady CL, McIntosh AR, Rajah MN, Craik FI. 1998. Neural correlates of the episodic encoding of pictures and words. Proc Natl Acad Sci USA 95:2703–2708. Hamann SB, Ely TD, Grafton ST, Kilts CD. 1999. Amygdala activity related to enhanced memory for pleasant and aversive stimuli. Nat Neurosci 2:289–293. Het S, Ramlow G, Wolf OT. 2005. A meta-analytic review of the effects of acute cortisol administration on human memory. Psychoneuroendocrinology 30:771–784. Heuer F, Reisberg D. 1990. Vivid memories of emotional events: The accuracy of remembered minutiae. Mem Cognit 18: 496 –506. van Honk J, Kessels RP, Putman P, Jager G, Koppeschaar HP, Postma A. 2003. Attentionally modulated effects of cortisol and mood on memory for emotional faces in healthy young males. Psychoneuroendocrinology 28:941–948. Jackson ED, Payne JD, Nadel L, Jacobs WJ. 2006. Stress differentially modulates fear conditioning in healthy men and women. Biol Psychiatry 59:516–522. Downloaded By: [Wolf, Oliver T.] At: 10:36 13 November 2008 Cortisol and emotional memory Kensinger EA. 2004. Remembering emotional experiences: The contribution of valence and arousal. Rev Neurosci 15:241–251. Kirschbaum C, Hellhammer DH. 1989. Salivary cortisol in psychobiological research: An overview. Neuropsychobiology 22:150–169. Kirschbaum C, Wolf OT, May M, Wippich W, Hellhammer DH. 1996. Stress- and treatment-induced elevations of cortisol levels associated with impaired declarative memory in healthy adults. Life Sci 58:1475–1483. Kuhlmann S, Wolf OT. 2005. Cortisol and memory retrieval in women: Influence of menstrual cycle and oral contraceptives. Psychopharmacology (Berl) 183:65–71. Kuhlmann S, Wolf OT. 2006a. A non-arousing test situation abolishes the impairing effects of cortisol on delayed memory retrieval in healthy women. Neurosci Lett 399:268–272. Kuhlmann S, Wolf OT. 2006b. Arousal and cortisol interact in modulating memory consolidation in healthy young men. Behav Neurosci 120:217–223. Kuhlmann S, Kirschbaum C, Wolf OT. 2005a. Effects of oral cortisol treatment in healthy young women on memory retrieval of negative and neutral words. Neurobiol Learn Mem 83: 158– 162. Kuhlmann S, Piel M, Wolf OT. 2005b. Impaired memory retrieval after psychosocial stress in healthy young men. J Neurosci 25: 2977–2982. LaBar KS, Cabeza R. 2006. Cognitive neuroscience of emotional memory. Nat Rev Neurosci 7:54 –64. Lang PJ, Bradley MM, Cuthbert BN. 1997. International Affective Picture System (IAPS): Technical Manual and Affective Ratings. Lupien SJ, Gillin CJ, Hauger RL. 1999. Working memory is more sensitive than declarative memory to the acute effects of corticosteroids: A dose-response study in humans. Behav Neurosci 113:420–430. Maheu FS, Collicutt P, Kornik R, Moszkowski R, Lupien SJ. 2005. The perfect time to be stressed: A differential modulation of human memory by stress applied in the morning or in the afternoon. Prog Neuropsychopharmacol Biol Psychiatry 29:1281– 1288. McEwen BS. 2000. The neurobiology of stress: From serendipity to clinical relevance. Brain Res 886:172–189. Okuda S, Roozendaal B, McGaugh JL. 2004. Glucocorticoid effects on object recognition memory require training-associated emotional arousal. Proc Natl Acad Sci USA 101:853–858. Payne JD, Jackson ED, Ryan L, Hoscheidt S, Jacobs JW, Nadel L. 2006. The impact of stress on neutral and emotional aspects of episodic memory. Memory 14:1–16. Payne JD, Jackson ED, Hoscheidt S, Ryan L, Jacobs WJ, Nadel L. 2007. Stress administered prior to encoding impairs neutral but enhances emotional long-term episodic memories. Learn Mem 14:861–868. Putman P, van Honk J, Kessels RP, Mulder M, Koppeschaar HP. 2004. Salivary cortisol and short and long-term memory for emotional faces in healthy young women. Psychoneuroendocrinology 29:953–960. 9 de Quervain DJ, Roozendaal B, McGaugh JL. 1998. Stress and glucocorticoids impair retrieval of long-term spatial memory. Nature 394:787–790. de Quervain DJ, Henke K, Aerni A, Treyer V, McGaugh JL, Berthold T, et al. 2003. Glucocorticoid-induced impairment of declarative memory retrieval is associated with reduced blood flow in the medial temporal lobe. Eur J Neurosci 17:1296–1302. Roozendaal B. 2002. Stress and memory: Opposing effects of glucocorticoids on memory consolidation and memory retrieval. Neurobiol Learn Mem 78:578– 595. Roozendaal B, Okuda S, de Quervain DJ, McGaugh JL. 2006a. Glucocorticoids interact with emotion-induced noradrenergic activation in influencing different memory functions. Neuroscience 138:901– 910. Roozendaal B, Okuda S, Van der Zee EA, McGaugh JL. 2006b. Glucocorticoid enhancement of memory requires arousalinduced noradrenergic activation in the basolateral amygdala. Proc Natl Acad Sci USA 103:6741–6746. Sapolsky RM, Romero LM, Munck AU. 2000. How do glucocorticoids influence stress responses? Integrating permissive, suppressive, stimulatory, and preparative actions. Endocr Rev 21:55–89. Smeets T, Jelicic M, Merckelbach H. 2006. The effect of acute stress on memory depends on word valence. Int J Psychophysiol 62: 30–37. Stark CE, Okado Y. 2003. Making memories without trying: Medial temporal lobe activity associated with incidental memory formation during recognition. J Neurosci 23:6748–6753. Stark R, Wolf OT, Tabbert K, Kagerer S, Zimmermann M, Kirsch P, et al. 2006. Influence of the stress hormone cortisol on fear conditioning in humans: Evidence for sex differences in the response of the prefrontal cortex. Neuroimage 32:1290–1298. van Stegeren AH, Wolf OT, Everaerd W, Scheltens P, Barkhof F, Rombouts SARB. 2007. Endogenous cortisol level interacts with noradrenergic activation in the human amygdala. Neurobiol Learn Mem 87:57– 66. Vuilleumier P, Armony JL, Driver J, Dolan RJ. 2001. Effects of attention and emotion on face processing in the human brain: An event-related fMRI study. Neuron 30:829–841. Watson D, Clark LA, Tellegen A. 1988. Development and validation of brief measures of positive and negative affect: The PANAS scales. J Pers Soc Psychol 54:1063–1070. Wolf OT. 2008. The influence of stress hormones on emotional memory: Relevance for psychopathology. Acta Psychol (Amst) 127:513–531. Wolf OT, Schommer NC, Hellhammer DH, McEwen BS, Kirschbaum C. 2001. The relationship between stress induced cortisol levels and memory differs between men and women. Psychoneuroendocrinology 26:711–720. Zorawski M, Blanding NQ, Kuhn CM, LaBar KS. 2006. Effects of stress and sex on acquisition and consolidation of human fear conditioning. Learn Mem 13:441–450. Studie 2 3 55 ST UDIE 2: POST -LEARNING PSY CHO SOCI AL ST RESS ENHANCES CONSOL IDAT ION OF NEUT RAL ST IMULI Studie 2: Post-learning psychosocial stress enhances consolidation of neutral stimuli Diana Preuß, Oliver T. Wolf Author's personal copy Neurobiology of Learning and Memory 92 (2009) 318–326 Contents lists available at ScienceDirect Neurobiology of Learning and Memory journal homepage: www.elsevier.com/locate/ynlme Post-learning psychosocial stress enhances consolidation of neutral stimuli Diana Preuß, Oliver T. Wolf * Department of Cognitive Psychology, Ruhr-University Bochum, Universitatsstr. 150, D-44780 Bochum, Germany a r t i c l e i n f o Article history: Received 23 October 2008 Revised 23 February 2009 Accepted 26 March 2009 Available online 10 April 2009 Keywords: Psychosocial stress Salivary cortisol Salivary alpha-amylase Memory Arousal Consolidation Sex differences a b s t r a c t Post-learning stress has been reported to enhance memory consolidation in humans. This effect was observed in studies using physical stressors or an anticipatory speech task. In the present study 58 participants (28 females and 30 males) were exposed to a psychosocial stressor (Trier Social Stress Test) or a control condition following the presentation of neutral and emotionally arousing positive and negative pictures, which were accompanied by a brief narrative. The stressor induced a significant neuroendocrine stress response in men and women. In a 24 h delayed free recall test the stress group showed an enhanced memory for neutral but not for emotionally arousing positive and negative items. Additionally, a significant correlation between the cortisol stress response and memory for neutral items was evident. Thus, in contrast to previous studies, post-learning stress primarily enhanced consolidation of neutral material. Several theoretical and methodological explanations for the observed effects are discussed. Ó 2009 Elsevier Inc. All rights reserved. 1. Introduction We know from everyday experiences that stressful events are well remembered and experimental laboratory research has shown that stress influences memory (Wolf, 2008). The modulatory effects of stress on memory are caused by the release of stress hormones. The activation of the sympathetic nervous system (SNS) in response to stress results in a release of catecholamines. Additionally, the hypothalamic-pituitary (HPA) axis is activated, which results in the release of glucocorticoids (GCs; de Kloet, Joels, & Holsboer, 2005). The effects of stress on memory depend on the particular memory phase influenced by stress (Roozendaal, Okuda, de Quervain, & McGaugh, 2006; Wolf, 2008). Additionally, it became apparent that stress differs depending on whether it is related to the learning situation or is outside the learning context (Joels, Pu, Wiegert, Oitzl, & Krugers, 2006). Increasing the stressfulness of a learning episode was found to enhance memory in rodents (Akirav et al., 2004; Akirav, Sandi, & Richter-Levin, 2001; Sandi, Loscertales, & Guaza, 1997). Akirav et al. (2004) for example observed that rats performed better in a spatial task (Morris Water Maze) when the situation was so designed as to be more stressful (colder water temperature). The authors could demonstrate that this memory enhancement was due to the release of corticosterone. Similarly, glucocorticoids injected immediately after acquisition (post-learn- * Corresponding author. Fax: +49 (0)234 32 14308. E-mail address: [email protected] (O.T. Wolf). 1074-7427/$ - see front matter Ó 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.nlm.2009.03.009 ing) and thereby influencing memory consolidation were found to enhance consolidation of newly learned material (Roozendaal, de Quervain, Ferry, Setlow, & McGaugh, 2001; Roozendaal, Nguyen, Power, & McGaugh, 1999). The empirical picture becomes more complex when stressor and learning task are not directly associated and the stressor is detached from the learning episode. This is the case when the animal receives foot shocks or is exposed to a predator before or after learning a maze task (e.g. Park, Zoladz, Conrad, Fleshner, & Diamond, 2008). Similarly, in human studies, the stressor (e.g. cold water immersion or a public speech) is typically unrelated to the memory tests conducted (e.g. Beckner, Tucker, Delville, & Mohr, 2006; Cahill, Gorski, & Le, 2003; Wolf, Schommer, Hellhammer, McEwen, & Kirschbaum, 2001). When reviewing previous human studies using this approach pre-learning and post-learning stress exposure need to be differentiated. With respect to pre-learning stress, enhancing as well as impairing effects have been observed. The direction of the effects appears to depend on several variables. The delay between stress exposure and the learning episode (Diamond, Campbell, Park, Halonen, & Zoladz, 2007) and the delay between initial learning and recall (immediate vs. delayed recall; Elzinga, Bakker, & Bremner, 2005) have turned out to be important variables. Moreover the emotionality of the learning material has been reported to influence the outcome in that, although pre-learning stress impaired neutral memory, it often enhanced emotional memory (Jelici, Geraerts, Merckelbach, & Guerrieri, 2004; Payne, Jackson, Hoscheidt, Ryan, Jacobs, et al., 2007; Schwabe, Bohringer, Chatterjee, & Schachinger, 2008). Author's personal copy D. Preuß, O.T. Wolf / Neurobiology of Learning and Memory 92 (2009) 318–326 For post-learning stress exposure, the empirical picture is more homogenous and reveals mostly enhancing effects. Several human studies observed that immediate post-learning stress either with the cold pressure test (CPT; Andreano & Cahill, 2006; Cahill et al., 2003; Smeets, Otgaar, Candel, Wolf, 2008) or with an anticipatory speech stressor (Beckner et al., 2006) led to enhanced memory consolidation. In those studies which used negative as well as neutral learning material, the effect was only found for the emotionally arousing negative items (Cahill et al., 2003; Smeets, Otgaar, Candel, & Wolf, 2008). In line with these stress studies we recently reported that basal cortisol levels were specifically associated with enhanced memory for emotional items (Preuss, Schoofs, Wolf, & emotional memory: influence of encoding instructions. Stress, in press). However, the beneficial effects of post-learning stress on memory consolidation have also been reported in studies where only neutral learning material was employed (Andreano & Cahill, 2006; Beckner et al., 2006). An impact of sex on the relationship between stress and memory has been reported in previous research. One aspect is that the HPA stress response to performance based laboratory stressors is influenced by sex. Men often show a more salient response than women (Kirschbaum, Kudielka, Gaab, Schommer, & Hellhammer, 1999; Kirschbaum, Wust, & Hellhammer, 1992; Stroud, Salovey, & Epel, 2002) but contrary results have also been reported (Kelly, Tyrka, Anderson, Price, & Carpenter, 2008). Additionally there are not only sex differences in the HPA reactivity to psychosocial stress but also sex differences for the influence of stress on memory or emotional learning. The effects of GCs here were repeatedly found to be more pronounced for men than for women (Andreano & Cahill, 2006; Cahill, 2003; Jackson, Payne, Nadel, & Jacobs, 2006; Stark, Wolf, Tabbert, Kagerer, Zimmermann, et al., 2006; Wolf et al., 2001; Zorawski, Blanding, Kuhn, & LaBar, 2006). Possible sex differences should therefore be considered when exploring the influence of stress on memory (Cahill, 2006; Wolf, 2008). Building up on recent findings in the field the present study was conducted to further clarify the influence of stress on consolidation. Existing studies evaluating the influence of stress on consolidation processes have either used a physical stressor (Andreano & Cahill, 2006; Cahill et al., 2003; Smeets et al., 2008) or an anticipatory speech stressor where the speech itself had not to be performed (Beckner et al., 2006). To date, no experiment has systematically assessed the influence of the Trier Social Stress Test (TSST, Kirschbaum, Pirke, & Hellhammer, 1993) on memory consolidation. This stressor typically leads to a more pronounced cortisol stress response compared to the CPS or anticipatory speech stressors used in previous consolidation studies (Beckner et al., 2006; Cahill et al., 2003; Kirschbaum et al., 1993; Kuhlmann, Piel, & Wolf, 2005; van Stegeren, Wolf, & Kindt, 2008). In addition it has a stronger ‘cognitive load’, since the subjects have to deliver a speech and work on a mathematical task. The aim of the present study was to evaluate the effects of post-learning stress on memory consolidation. Based on previous observations, as summarized above, the factors emotional arousal of the learning material as well as sex of the participants was taken into account. 2. Materials and methods 319 excluded and the data of 58 participants (28 female, 30 male) was analyzed. Women were between the age of 19 and 28 (mean age 23.68 ± 0.45). Men were between the age of 20 and 29 (mean age 23.53 ± 0.48). Mean body mass index for the men was 24.12 (±0.45) and for the women 21.56 (±0.49). Participants were excluded if they reported any use of medicaments that could have influenced the hormonal stress response (e.g. antibiotics, and antihistamines). Women were free of hormonal contraception. All participants were recruited at the university campus and written informed consent was collected from each subject. The study was approved by the national ethic committee of the German Psychological Association (Deutsche Gesellschaft für Psychologie). 2.2. Materials 2.2.1. Stimuli The stimuli and memory tests used in the present study were developed and validated by Buchanan, Karafin, and Adolphs (2003). The material, recently translated by our group, had been used in a first study testing the associations between basal cortisol levels and emotional memory (Preuss et al., in press). The stimuli consisted of five positive (e.g. two happy girls eating ice-cream), five negative (e.g. a diseased child from Africa with bandages and cannulae) and five neutral (e.g. people leaving or entering a building) pictures, each presented in a random order for a duration of 10 sec on a computer screen. Several of these pictures were chosen from the International Affective Picture System (IAPS; Lang, Bradley, & Cuthbert, 1997) and the remaining drawn from print media sources. Each picture was accompanied by a single narrative sentence which consisted information that was not obvious in the picture. For example the picture with the little girls eating ice-cream was accompanied by a sentence in which the girls’ names and the special kind of ice-cream they preferred being mentioned. 2.2.2. Memory tasks Participants solved several written memory tests. 2.2.3. Immediate recall test The immediate free recall test took place immediately after the presentation of the pictures. Participants were asked to write down everything they remembered from the pictures and narratives. Time was restricted to 5 min. Answers were evaluated by two independent judges. Differences in test scores were discussed and were solved by a third judge. A participant received three points, if the information noted could be clearly associated to one of the pictures and was correct in details. Two points were given for information that could be clearly associated to one of the pictures but consisted of some wrong details. If the information was completely wrong or could not be linked to one picture, participants got one point. A total of 45 points could be achieved. 2.2.4. Delayed recall test On the second day, 24 h after presentation of the pictures, the delayed free recall test was conducted. Again, participants were given 5 min to write down everything they remembered from the pictures and narratives. Answers were evaluated in the same manner as in the immediate recall test. 2.1. Participants Participants were 30 healthy men and 30 healthy free cycling women. Women were tested during the whole menstrual cycle except menses. Two outliers with data above or below 2, 5 standard deviations in immediate memory recall scores had to be 2.2.5. Multiple choice test This task consisted of six questions asking for information pertaining to pictures and narratives for each stimulus. In this task every correct answer scored a point, so that a total of 90 points was possible. Author's personal copy 320 D. Preuß, O.T. Wolf / Neurobiology of Learning and Memory 92 (2009) 318–326 2.2.6. Memory for gist and detail The multiple choice test also permits a differentiation between memory for gist and detail information. Gist is defined as ‘‘an information which could not be changed or excluded without changing the basic story line” (Heuer & Reisberg, 1990). According to Buchanan et al. (2003) 47 questions referred to gist information and 37 to detail information. The remaining six items could not be categorized clearly and were thus not included in the gist analysis (Buchanan et al., 2003). Here again every correct answer scored one point, so that 47 points for the gist items and 37 points for detail items were achievable. In order to allow comparisons between memory for gist and memory for detail memory performance was expressed in percentages. 2.2.7. Mood assessment 2.2.7.1. Positive and negative affective schedule (PANAS; Watson, Clark, and Tellegen, 1988). This questionnaire consists of ten items for negative and ten items for positive mood. For the present study, only the negative mood scale was used. Participants filled out the PANAS three times, the first time before the presentation of the pictures, the second time after the stress or control condition and for the third time at the beginning of the memory tests on the second day. 2.2.8. Stressor and control condition In the present study a psychosocial stressor, the Trier Social Stress Test (TSST; Kirschbaum et al., 1993), was used. This stress protocol consisted of a video-taped oral presentation and an arithmetic task before a panel (one woman and one man) whose attitude was very reserved. This psychosocial stressor, with a total duration of 15 min, reliably elicits a response of the HPA and SNS (Dickerson & Kemeny, 2004; Kuhlmann et al., 2005). The nonstressful control condition also consists of an oral presentation and an arithmetic task but participants did not perform in front of an audience and were not video-taped. The control condition therefore lacks the stressful components of the TSST and did not elicit a cortisol stress response (Dickerson & Kemeny, 2004; Kuhlmann et al., 2005). 2.2.9. Saliva samples Saliva was collected using Salivette collection devices (Sarstedt, Nümbrecht, Germany). Totally, seven saliva samples were collected, five on the first day and two on the second day. Cortisol (Kirschbaum & Hellhammer, 1989) and Alpha-Amylase (sAA) as a measure of SNS activity (Chatterton, Vogelsong, Lu, Ellman, & Hudgens, 1996; Rohleder, Nater, Wolf, Ehlert, & Kirschbaum, 2004; van Stegeren, Rohleder, Everaerd, & Wolf, 2006) were assessed. 2.3. Procedure The experimental protocol is illustrated in Fig. 1. Participants were tested on 2 days 24 h apart. The testing started between 2 p.m. and 4 p.m. After arrival on the first day, participants filled out the PANAS for the first time (PANAS pre-treatment). Subsequently, the first saliva sample was collected (base 1), followed by presentation of the pictures and narratives. Participants then solved the immediate recall test and collected the second saliva sample (base 2) after finishing the test. This was followed by the TSST or the control condition. Only after entering the TSST or control condition room were participants aware of whether or not they would be part of the stress or control condition. Afterwards the third saliva sample was collected (+01) and subjects filled out the PANAS for the second time (PANAS post-treatment). Then the fourth saliva sample was assessed (+10). The last saliva sample (+25) on the first day was collected 25 min after the respective treatment. Participants were debriefed about the TSST at the end of the first day. On the next day, 24 h after the encoding of the pictures on the first day, participants returned to the laboratory and the first saliva sample (pre) was collected before they filled out the PANAS for the third time (PANAS day 2). Subsequently, they solved the memory tests and the last saliva sample (post; approximately 45 min after the first saliva sample) was collected. 2.4. Statistical analyzes Data were analyzed with t-tests or ANOVAs for repeated measurements and post-hoc paired t-tests. Greenhouse–Geisser corrected p values were used when indicated. Cortisol baseline levels were normally distributed. Because saliva alpha-amylase baseline measure did not show a normal distribution, data were log 10 transformed to approximate them to Gaussian distribution. After transformation all data were normally distributed. 3. Results 3.1. Cortisol response The cortisol responses to the TSST for the entire sample, as well as for women and men separately, are displayed in Fig. 2. To analyze the cortisol response an ANOVA with the inner subject factor time (base 1 vs. base 2 vs. +01 vs. +10 vs. +25) and the between subject factors stress (TSST vs. control condition) and sex (male vs. female) was conducted. A main effect of time occurred (F(4, 216) = 8.990, p < 0.01). Additionally, main effects for sex (F(1, 54) = 8.429, p < 0.05) and stress occurred (F(1, 54) = 21.111, p < 0.001). Also the interactions between time and stress (F(4, 216) = 27.990, p < 0.001) reached significance. The interaction between sex and stress (F(4, 216) = 3.483, p = 0.05), sex and time (F(4, 216) = 3.483, p = 0.05) and sex and stress and time (F(4, 216) = 3.464, p = 0.05) just fell short of significance. For further evaluation t-tests were conducted for the comparison between the conditions and additionally for the sexes. The TSST group had higher cortisol levels for the measurements +01 (t(56) = 4.534, p < 0.001), +10 (t(56) = 5.565, p < 0.001) and +25 (t(56) = 5.388, p < 0.001). Fig. 1. Procedure of the study. Author's personal copy D. Preuß, O.T. Wolf / Neurobiology of Learning and Memory 92 (2009) 318–326 321 Fig. 2. Cortisol response to the TSST and the control condition. A significant interaction between stress and time occurred in the ANOVA. Follow-up t-tests revealed significant differences between TSST and control group for (a) the whole sample, (b) females and (c) males at measurements +01, +10 and +25 (**p < 0.001; *p < 0.05). No difference was observed for the remaining measurements (all p > 0.10). Post-hoc t-test for the comparison between males and females were conducted for the baseline measurements and the rise in cortisol (measurement + 10 baseline measurement) in response to the TSST. No difference was found for the baseline measurements (all p > 0.05). For women, a rise in cortisol (measurement + 10 baseline measurement) of 3.76 nmol/l (±1.66) was detected while cortisol levels in men showed a significantly stronger rise of 10.72 nmol/l (±2.16; t(27) = 2.369, p < 0.05). Additionally, ttests were conducted for the two sexes separately. For both sexes higher cortisol values were found for the TSST group for the measurements +01 (males: t(28) = 3.404, p < 0.01; females: t(26) = 4.233, p < 0.001), +10 (males: t(28) = 4.424, p < 0.001; females: t(26) = 5.443, p < 0.001) and +25 (males: t(28) = 4.057, p < 0.001; females: t(26) = 4.760, p < 0.001) but not for the remaining time points (all p > 0.05). For the second day, an ANOVA with the inner subject factor time (pre- vs. post-) and the between subject factors stress (TSST vs. control condition) and sex (male vs. female) was conducted. Results revealed a main effect of time (F(1, 54) = 31.913, p < 0.001) and sex (F(1, 54) = 7.022, p < 0.001). The remaining effects did not reach significance (all p > 0.05). To assess the significant main effects of time and sex further, additional t-tests were conducted. Results revealed that males had higher cortisol levels than women at both measurements (pre: t(56) = 2.330, p < 0.05; post: t(56) = 2.975, p < 0.01). Analysis of the significant main effect of time showed that cortisol levels on the second day decreased slightly but significantly during testing (t(57) = 5.746, p < 0.001) from 6.25 (±0.45) to 4.58 (±0.24) nmol/l (males: from 7.24 (±0.66) to 5.22 (±0.34) nmol/l; females: from 5.21 (±0.56) to 3.90 (±0.30) nmol/l) reflecting the well known circadian decline of the hormone. the sAA stress response an ANOVA with the inner subject factor time (base 1 vs. base 2 vs. +01 vs. +10 vs. +25) and the between subject factors stress (TSST vs. control condition) and sex (male vs. female) was conducted. The main effects of time (F(4, 204) = 27.639, p < 0.001) and stress (F(1, 54) = 5.530, p < 0.05) were significant. Additionally, a significant interaction effect between time and stress occurred (F(4, 204) = 4.573, p < 0.05). The remaining effects did not reach significance (all p > 0.10). Post-hoc t-test revealed higher values in the TSST group at measurements +01 (t(56) = 3.826, p < 0.001), +10 (t(56) = 2.555, p < 0.05) and +25 (t(56) = 2.101, p < 0.05) but not at the other measurements (all p > 0.10). For the second day an ANOVA with the inner subject factor time (pre vs. post) and the between subject factors stress (TSST vs. control condition) and sex (male vs. female) was conducted. No significant results were detected (all p > 0.05). 3.2. Alpha-amylase response (sAA) ANOVAs with the inner subject factors valence (positive vs. negative vs. neutral) and the between subject factors sex (male vs. female) and stress (TSST vs. control condition) were conducted for the ratings of arousal and valence of the pictures, conducted on The sAA responses to the TSST for the entire sample as well as for women and men are separately displayed in Fig. 3. To analyze 3.3. Mood To evaluate changes in mood in reaction to the TSST an ANOVA with the factors time (pre-treatment vs. post-treatment), sex (male vs. female) and stress (TSST vs. control condition) was conducted for negative mood. A significant main effect of stress (F(1, 51) = 14.089, p < 0.001) and a significant interaction between time and stress (F(1, 51) = 11.006, p < 0.01) occurred. The remaining effects did not reach significance (all p > 0.05). To investigate the significant interaction effect further t-tests were conducted. The TSST group reported more negative mood after the TSST (t(55) = 4.257, p < 0.001) but not before the TSST (t(54) = 1.429, p = 0.159) compared to the control group. 3.4. Ratings of the pictures Fig. 3. sAA response (log transformed data) to the TSST and control condition. A significant interaction between stress and time occurred in the ANOVA. Follow-up t-tests showed significant differences between TSST and control group for (a) the whole sample, (b) females and (c) males at measurements +01, +10 and +25 (**p < 0.001; *p < 0.05). Author's personal copy 322 D. Preuß, O.T. Wolf / Neurobiology of Learning and Memory 92 (2009) 318–326 day two. For valence a significant main effect of valence occurred (F(2, 108) = 450.637, p < 0.001). The remaining effects did not reach significance (all p > 0.10). Participants rated the positive items more positive than the neutral (t(57) = 13.363, p < 0.001) and negative ones (t(57) = 24.078, p < 0.001). Neutral items were rated as more positive than negative items (t(57) = 23.621, p < 0.001). For arousal a significant main effect of valence occurred (F(2, 108) = 197.486, p < 0.001). Again, the remaining effects did not reach significance (all p > 0.10). Participants rated the negative items as more arousing than the positive (t(57) = 10.867, p < 0.001) and neutral ones (t(57) = 19.330, p < 0.001). Positive items were rated as more arousing than the neutral ones (t(57) = 9.314, p < 0.001). 3.5. Effect of valence on immediate recall To evaluate the influence of valence on initial acquisition an ANOVA with the factors valence (positive vs. negative vs. neutral), sex (male vs. female) and stress (TSST vs. control condition) was conducted for the immediate recall test, which occurred before the TSST or control condition. Results revealed a significant main effect of valence (F(2, 108) = 39.313, p < 0.001) and a significant main effect of sex (F(1, 54) = 9.205, p < 0.01). The remaining main and interaction effects did not reach significance (all p > 0.05). Paired t-tests showed that participants significantly recalled more negative than positive (t(57) = 2.359, p < 0.05) and neutral items (t(57) = 8.067, p < 0.001). Additionally more positive items were recalled than neutral items (t(57) = 6.098, p < 0.001). Women overall recalled more pictures than men (t(56) = 3.086, p < 0.05). Results are presented in Fig. 4. 3.6. Effects of post-learning stress on delayed free recall For analyzes of delayed free recall we created a value which accounts for possible within and between subject variance in initial learning. Therefore free recall performance on the second day was expressed as the percentage of memory score in relation to the immediate recall score ((memory score on the second day/ memory score on the first day) 100; Kuhlmann et al., 2005; Kuhlmann & Wolf, 2005). Values above 100% thereby show an increase of memory over time (higher memory score on the second day), while values below 100% show decrease of memory (higher mem- ory scores on the first day). This value was computed for positive, negative and neutral items separately. To evaluate the influence of stress on memory an ANOVA was conducted with the factors valence (positive vs. negative vs. neutral), stress (TSST vs. control condition) and sex (male vs. female). A significant interaction effect between valence and stress occurred (F(2, 108) = 3.910, p < 0.05). No other significant effects could be observed (all p > 0.10). A post-hoc t-test was conducted to evaluate the significant interaction effect further. The TSST group showed better memory for the neutral items (t(56) = 2.183, p < 0.05) but not for the positive (t(56) = 0.411, p = 0.682) and negative ones (t(56) = 1.135, p = 0.261). Additionally we conducted the ANOVA for each sex separately. A significant interaction effect between valence and stress was found in men (F(2, 56) = 4.603, p < 0.05) but not in women (F(2, 52) = 0.585, p = 0.561). The remaining effects did not reach significance (all p > 0.10). Additional t-tests were conducted for the comparison between stress and control group for males and females separately. No significant difference occurred for positive (males: t(28) = 0.462, p = 0.647, females: t(26) = 0.040, p = 0.969) or negative items (males: t(28) = 0.805, p = 0.427; females: t(26) = 0.813, p = 0.424). However, for males a trend for a better recall of neutral items in the TSST group (t(56) = 1.824, p = 0.079) occurred. In females this effect was non-significant (t(56) = 1.190, p = 0.245). The results for the entire sample as well as for women and men are separately presented in Fig. 5. 3.7. Effects of post-learning stress on delayed recall assessed with a multiple choice test For the multiple choice test an ANOVA with the factors valence (positive vs. negative vs. neutral), sex (male vs. female) and stress (TSST vs. control condition) was conducted. Results revealed significant main effects of valence (F(2, 108) = 17.472, p < 0.001) and a significant interaction effect between sex and valence (F(2, 108) = 8.509, p < 0.001). No main effect of stress (F(1, 54) = 1.702, p = 0.198) and no interaction effects between stress and valence (F(2, 108) = 1.539, p = 0.219) were detected. The remaining effects did not reach significance (all p > 0.10) as well. Participants showed a better memory for negative than for positive (t(57) = 4.838, p < 0.001) and neutral items (t(57) = 5.005, p < 0.001). No differences emerged between positive and neutral items (t(57) = 0.577, p = 0.566). Women remembered significantly more positive items than men (t(56) = 4.158, p < 0.001). No differences emerged for negative and neutral items (all p > 0.10). 3.8. Effects of post-learning stress on memory for gist and detail Fig. 4. Memory score for the immediate recall test for positive, negative and neutral items for the whole sample. An ANOVA revealed a significant main effect of valence and sex. Post-hoc t-tests showed that participants learned significantly more negative items than positive items (*p < 0.05) and neutral items (**p < 0.001). Positive items were better learned than neutral items (**p < 0.001). Overall women recalled more pictures than men (p < 0.05). To evaluate memory for gist and detail information an ANOVA with the inner subject factors valence (positive vs. negative vs. neutral) and gist (gist vs. detail) and the between subject factors sex (male vs. female) and stress (TSST vs. control condition) was conducted. Analysis revealed significant main effects of valence (F(2, 108) = 19.250, p < 0.001), gist (F(1, 54) = 179.992, p < 0.001) and a significant interaction effect between valence and gist (F(2, 108) = 22.555, p < 0.001). Additionally a significant main effect of sex was detected (F(1, 54) = 5.958, p < 0.05). No main effect and no interactions with stress could be observed (all p > 0.10). Results of t-tests revealed that overall women remembered more items than men (t(56) = 2.454, p < 0.05). Results also revealed that more gist information was remembered for positive (t(57) = 5.199, p < 0.001), negative (t(57) = 14.233, p < 0.001) and neutral items (t(57) = 4.959, p < 0.001). Author's personal copy 323 D. Preuß, O.T. Wolf / Neurobiology of Learning and Memory 92 (2009) 318–326 Fig. 5. Mean memory score for positive, negative and neutral items in the TSST and control group for (a) the whole sample, (b) females and (c) males. An ANOVA revealed a significant interaction between stress and valence in the whole sample and for the males. Follow-up t-tests showed a significant difference for the neutral items (*p < 0.05) in the whole sample and a trend for the males (§p < 0.10). Memory scores reflect the percentage of words recalled on the second day in relationship to the immediate recall assessed on the first day. Memory scores above 100% reflect an increase in memory, whereas memory scores below 100% reflect a decrease in memory. 3.9. Relationship between memory, mood, cortisol and alpha-amylase To evaluate possible associations between the neuroendocrine and affective stress markers and delayed memory retrieval, bivariate Pearson’s correlations were conducted. We computed a measure for the cortisol response (cortisol + 10 cortisol baseline; Kirschbaum, Wolf, May, Wippich, Hellhammer, 1996; Wolf et al., 2001). Higher values indicate a stronger response to the TSST. Similarly, we also created a value for the response between baseline and the highest value (sAA + 01 sAA baseline) for alpha-amylase. Additionally a measure for negative mood increase (negative mood after TSST negative mood before TSST) was computed. Again higher values indicate a stronger increase of negative mood. For the second day a mean value was conducted for the two cortisol samples ((pre + post/2)). This was done to create a single measure indicative of the basal cortisol level on the day of retrieval testing. The cortisol response was significantly correlated with memory for neutral items. This was the case for the entire group as well as for males and females separately. For negative items a trend (0.10 < p < 0.05) emerged for the analysis including the entire group, while no association was observed when the two sexes were analyzed separately. Results are presented in Table 1. Cortisol levels during retrieval (day 2) were not significantly (p > 0.10) associated with any of the memory measures (data not shown). In order to ascertain that the observed correlations are not simply reflective of the group differences between the stressed group and the control group, partial correlations were conducted controlling for the grouping factor stress (TSST vs. control). For the entire sample, the correlation between the cortisol response and memory for neutral items was still significant (r = 406, p > 0.01). The correlations between cortisol and memory for positive (r = 0.093, p = 0.491) and negative (r = 0.210, p = 0.118) items were non-significant. When the sample was split according to sex the relationship between cortisol and memory for neutral items was still significant in men (r = 0.391, p < 0.05) but turned into a non-significant trend in women (r = 0.343, p = 0.08). The correlations be- tween cortisol and memory for positive (males: r = 0.044, p = 0.822; females: r = 0.205, p = 0.306) and negative (males: r = 0.216, p = 0.260; females: r = 0.081, p = 0.687) items also remained non-significant. No significant correlations were detected between mood and memory performance or sAA and memory performance (all p > 0.10). 4. Discussion The aim of the present study was to assess the influence of postlearning psychosocial stress on memory consolidation of positive, negative and neutral pictures. Additionally, possible sex differences were evaluated. Results demonstrate that psychosocial stress elicited a significant neuroendocrine stress response in men and women. In both sexes cortisol and alpha-amylase levels were elevated after the TSST. However, the cortisol stress response was more pronounced in men. Additionally, an affective reaction to the TSST was observed with both sexes reporting more negative mood after the TSST. With respect to memory post-learning, stress enhanced the consolidation of neutral items but did not significantly affect memory for positive or negative items. This interaction effect was significant in the whole sample as well as in the male group. In support of these group comparisons, correlations were found between the cortisol stress response and memory for neutral items in the free recall test for the entire sample, as well as for both sexes separately. Additionally, a trend for a correlation between cortisol response and memory for negative items occurred for the whole sample. No effect of stress was found for the multiple choice test or the separate analysis of memory for gist and details assessed with the multiple choice test. The finding of a neuroendocrine response to the TSST is well in line with the literature reporting an enhanced release of cortisol and salivary alpha-amylase in response to psychosocial stress (Dickerson & Kemeny, 2004; Kudielka, Schommer, Hellhammer, & Kirschbaum, 2004; Nater et al., 2006; Rohleder et al., 2004). In Table 1 Correlation coefficients and p-values for the correlations between memory scores in the delayed free recall test and cortisol response on day 1 (learning session). Cortisol response (day 1) * ** p < 0.05. p < 0.001. Whole sample n = 58 Males n = 30 Females n = 28 Positive items delayed free recall Negative items delayed free recall Neutral items delayed free recall r = 0.042 p = 0.755 r = 0.030 p = 0.874 r = 0.173 p = 0.379 r = 0.256 p = 0.053 r = 0.258 p = 0.169 r = 0.149 p = 0.450 r = 0.480 p < 0.001** r = 0.492 p < 0.05* r = 0.403 p < 0.05* Author's personal copy 324 D. Preuß, O.T. Wolf / Neurobiology of Learning and Memory 92 (2009) 318–326 addition to the physical response to the stressor, an increase of negative mood was observed which indicates an emotional arousal in response to the TSST (Abercrombie, Speck, & Monticelli, 2006). In the present experiment women showed a weaker cortisol response to the TSST than men. This is in agreement with several studies that demonstrate a weaker salivary cortisol response to performance based stressors in women than in men (Kirschbaum, Pirke, & Hellhammer, 1995; Kirschbaum et al., 1996; Kirschbaum et al., 1992; Stroud et al., 2002, but see Kelly et al., 2008). No sex differences were detected for salivary alpha-amylase, which is in contrast to a recent report of overall higher sAA levels in men (van Stegeren et al., 2008). In sum, the present results demonstrate that the TSST induced a robust neuroendocrine and affective stress response. For cortisol, but not for the other two stress markers (sAA and mood), the response was more pronounced in men. The present study reports an enhanced memory consolidation after psychosocial stress treatment. Post-encoding stress is known to enhance memory consolidation in rodents (Akirav et al., 2001; Akirav et al., 2004; Roozendaal et al., 1999; Roozendaal et al., 2001; Sandi et al., 1997) and humans (Andreano & Cahill, 2006; Beckner et al., 2006; Cahill et al., 2003; Smeets et al., 2008). However, in the present study, this beneficial effect was restricted to neutral items and no significant effect was found for positive or negative items. This result was somewhat unexpected, as the beneficial effects of cortisol have often been found to be more pronounced for emotional items. An enhanced memory for emotional items was observed after cortisol treatment (Buchanan & Lovallo, 2001; Kuhlmann & Wolf, 2006b) or post-learning stress (Abercrombie et al., 2006; Cahill et al., 2003; Smeets et al., 2008). It has therefore been assumed that arousal caused by the learning material potentiates the effects of stress and elevated GCs on memory (Roozendaal et al., 2006). However, in our study the opposite picture emerged. We found stress effects on the non-arousing neutral items, but no significant effect on the arousing positive and negative ones, even though a numeric trend could be observed for negative items. Moreover, the correlational analysis revealed a trend between the stress induced cortisol rise and memory in the free recall test for negative items. This association would be in line with previous reports on a post-learning stress induced consolidation enhancement for negative items (Cahill et al., 2003; Smeets et al., 2008). It suggests that the failure to find an effect on negative items at the between group level might be caused by specifics of the used task, which might not have been sensitive in detecting the beneficial effects on the negative items. Several studies observed an enhanced memory for both kinds of stimuli after pre-learning cortisol treatment (Abercrombie, Kalin, Thurow, Rosenkranz, & Davidson, 2003) or for neutral stimuli after post-learning stress, when only neutral stimuli were presented (Andreano & Cahill, 2006; Beckner et al., 2006). The present finding of an effect on the neutral stimuli therefore, is in line with these studies but the unanswered question is why we did not detect an effect on emotional items. One explanation could be differences in acquisition. At the time of immediate recall we found, as expected, an emotional enhancement effect. Positive and negative pictures were significantly better remembered than neutral ones. This emotional enhancement effect did not further increase between the immediate and delayed recall, which is in contrast to some previous studies (e.g. Quevedo et al., 2003), but in line with previous studies from our group (Kuhlmann & Wolf, 2006b). The stress group recalled more neutral items in the delayed recall test, compared to the immediate recall test. Thus, in this task, post-learning stress not only prevented forgetting but actually boosted memory consolidation leading to a superior memory performance 24 h after the original presentation of the slides. One interpretation of our findings could be that stress especially enhances weak or fragile memory traces. Having said this it must be acknowledged that due to their better initial encoding, emotional memories had less room for a further improvement due to post-learning stress (possible ceiling effect). In the present study we used pictures which were accompanied by a narrative. Here, the stimuli formed a complex episode and are somewhat comparable to the stimuli used by Andreano and Cahill (2006) who also found an effect of stress on neutral material. The specific effects of post-learning stress or pre-learning cortisol treatment on emotional memory were obtained in studies using a larger number of slides, which were presented without the additional presentation of verbal information (Cahill et al., 2003; Kuhlmann & Wolf, 2006b). This rather long presentation interval might lead to a deeper processing. The absence of forgetting during the 24 h delay strengthens the assumption of a deep encoding. In most studies participants perform better in the immediate recall test compared to the delayed recall test (Kuhlmann & Wolf, 2005; Kuhlmann et al., 2005). In the present study, no forgetting took place which might be due to deep encoding and the relatively small number of slides used. Our overall findings are somewhat similar to a recent study from Buchanan and Tranel (2008), where the authors used a slightly modified version of this task containing 20 pictures (ten neutral and ten negative). Similar to our findings the authors of this study observed little to no forgetting over a 24 h delay. In addition pre-retrieval stress had an impact on memory, which however was not specific to the emotionally arousing pictures. A different explanation might be that the observed effects are due to an inverted U-shape relationship between cortisol and arousal (Baldi & Bucherelli, 2005). Arousal is associated with noradrenergic activation in the basolateral amygdala and this activation is thought to be a prerequisite for the effects of GCs on memory (de Quervain, Aerni, & Roozendaal, 2007; Kuhlmann & Wolf, 2006a; Roozendaal et al., 2006). A recent study of ours using the identical learning material observed that basal cortisol levels were positively associated with memory for arousing pictures but not for neutral ones (Preuss et al., in press). In this study no stressor was used and therefore effects rely on the arousal, which is induced by the learning material. Our present finding could suggest that cortisol can exert its positive effects on memory consolidation only at an optimal arousal level. This optimal level might reflect the interaction of arousal induced by the learning material with arousal induced by the experimental manipulation. In situations where there is no stressor, cortisol enhances memory for items that induce arousal but not for items which lack this arousal (Buchanan & Lovallo, 2001; Kuhlmann & Wolf, 2006b; Preuss et al., in press). In situations with a mild stressor (e.g. cold pressure stressor) this scenario might still take place. However in the case of a strong psychological stressor (e.g. the TSST) neutral items might have the optimal arousal level, while the arousing positive and negative items are shifted to the right side of the inverted U-shape. Anticipation of and, even more, participating in a strong stressor might therefore cause the effects observed on neutral items in the present study. Probably cortisol affects consolidation of non-arousing neutral items only under circumstances of strong external induced arousal. The assumption of an inverted U-shape function for arousal might explain some of the inconsistent results in the field of cortisol and arousal but this hypothesis needs further evaluation and is not consistent with all previously published findings. Effects of post-learning stress were restricted to the free recall test. No influence of stress or GCs on memory performance in the multiple choice test was detected. The finding of an unaffected performance in cued recall tests is in line with several previous stress or cortisol studies (e.g. (de Quervain et al., 2003; Kuhlmann & Wolf, 2006b; Kuhlmann et al., 2005). Evidence has been provided that recollection and recognition are mediated by different brain structures (hippocampus vs. perirhinal cortex; Aggleton & Brown, Author's personal copy D. Preuß, O.T. Wolf / Neurobiology of Learning and Memory 92 (2009) 318–326 2006). Knowing that the hippocampus is especially sensitive to the effects of stress, it is not surprising to find the strongest effects for free recall. Additionally, in the cued recall test, we observed no effect on gist or detail aspects of this task. This is in contrast to previous pre-learning stress studies which reported that stress impaired memory for the gist but not for details of a story or even increased the number of details remembered (Cahill et al., 2003; Payne et al., 2006). This might suggest that the task used to differentiate between gist and details in the present study was not sensitive to stress effects. Thus more research is needed in order to understand whether or not stress differentially affects information for details. An additional aim of the present study was to evaluate possible sex differences in the effects of stress on memory consolidation. Results indicate slight differences between men and women. While the overall ANOVA did not reveal a significant sex by treatment interaction, separate analyzes for men and women indicated a significant effect of stress on memory for men but not for women. For men we found a beneficial effect of stress on memory for neutral items. In women the effects were smaller and non-significant. It has been demonstrated in several studies that sex differences exist for the influence of stress on memory or emotional learning. The effects of stress were often more pronounced for men than for women (Andreano & Cahill, 2006; Cahill, 2003; Jackson et al., 2006; Stark et al., 2006; Wolf et al., 2001; Zorawski et al., 2006). In our study, the smaller cortisol response to the stressor is the most likely mechanism behind the observed sex differences, since the effects in women were descriptively rather similar to the results obtained in men. In support of this conclusion we observed, in both sexes, an association between the cortisol response and memory consolidation for neutral pictures. Due to pragmatic reasons women were tested at every phase of the menstrual cycle in our current study with the exception of menses. A recent report suggests that only during the luteal phase correlations between cortisol and memory might be detectable (Andreano, Arjomandi, & Cahill, 2008). However, the literature on this topic is heterogeneous and opposing results (no associations in the luteal phase) have been reported as well (Wolf et al., 2001). Moreover Smeets et al. (2008) as well as Preuss et al. (in press) reported an association between cortisol and memory consolidation in a sample of women not further characterized with respect to their hormonal status. Finally, in the present study, the cortisol stress response was associated with memory consolidation in women, even though not as strong as it was in men. In sum, even though the empirical situation on the impact of the menstrual cycle on stress induced memory changes is far from clear, menstrual cycle associated alterations might have contributed to the smaller effects of the stressor on memory consolidation in the women group. There are some limitations of the current study. First, our sample size of 28 women and 30 men is certainly not large enough to exclude the possibility that non-significant findings might be secondary to a lack of power. Second, as mentioned above, we did not control for menstrual cycle stage (except of menses), which might have increased the variance in our group of women. In order to evaluate the influence of menstrual cycle associated changes in gonadal steroids on the observed effects, sex hormones should be measured in future studies on this topic whenever possible (Andreano et al., 2008). Third, more objective psychophysiological measures (e.g. skin conductance) of arousal could be helpful in clarifying the obviously quite complex association between stress and the emotional arousal of the learning material (Cahill et al., 2003). Fourth, the number of items presented (five per valence category), together with the choice of a delay of 24 h between immediate and delayed recall might have resulted in a task which was almost too easy for the subjects. This problem is illustrated in 325 the failure to find forgetting during the delay period. Especially for emotionally arousing items there might not have been enough room in order to detect a stress induced consolidation enhancement as observed in previous studies (Cahill et al., 2003; Smeets et al., 2008). Future studies on this topic should use memory tasks with more items and/or a longer retention interval. In sum, the current study reports on the beneficial effect of post-learning stress on the consolidation of neutral stimuli. In contrast, positive and negative stimuli were not affected. Thus, our study illustrates that the effects of stress on memory are not always stronger for emotional material. We have discussed several methodological and theoretical explanations for the specificity of the effects for neutral material observed in this study. Additional experiments are needed to disentangle the factors determining the consolidation of memories acquired directly prior to stress. Acknowledgments This study was supported by a Grant from the German Research Foundation (DFG WO 733/7-1). The authors wish to thank Angelika Baumeister for helping with the data acquisition and wish to thank Lily-Maria Silny for language editing. Finally we wish to thank Tony Buchanan (Department of Psychology Saint Louis University) for providing us with the emotional memory task used in this study. References Abercrombie, H. C., Kalin, N. H., Thurow, M. E., Rosenkranz, M. A., & Davidson, R. J. (2003). Cortisol variation in humans affects memory for emotionally laden and neutral information. Behavioral Neuroscience, 117, 505–516. Abercrombie, H. C., Speck, N. S., & Monticelli, R. M. (2006). Endogenous cortisol elevations are related to memory facilitation only in individuals who are emotionally aroused. Psychoneuroendocrinology, 31, 187–196. Aggleton, J. P., & Brown, M. W. (2006). Interleaving brain systems for episodic and recognition memory. Trends in Cognitive Sciences, 10, 455–463. Akirav, I., Kozenicky, M., Tal, D., Sandi, C., Venero, C., & Richter-Levin, G. (2004). A facilitative role for corticosterone in the acquisition of a spatial task under moderate stress. Learning and Memory, 11, 188–195. Akirav, I., Sandi, C., & Richter-Levin, G. (2001). Differential activation of hippocampus and amygdala following spatial learning under stress. European Journal of Neuroscience, 14, 719–725. Andreano, J. M., Arjomandi, H., & Cahill, L. (2008). Menstrual cycle modulation of the relationship between cortisol and long-term memory. Psychoneuroendocrinology, 33, 874–882. Andreano, J. M., & Cahill, L. (2006). Glucocorticoid release and memory consolidation in men and women. Psychological Science, 17, 466–470. Baldi, E., & Bucherelli, C. (2005). The inverted ‘‘U-shaped” dose–effect relationships in learning and memory: Modulation of arousal and consolidation. Nonlinearity in Biology, Toxicology, and Medicine, 3, 9–25. Beckner, V. E., Tucker, D. M., Delville, Y., & Mohr, D. C. (2006). Stress facilitates consolidation of verbal memory for a film but does not affect retrieval. Behavioral Neuroscience, 120, 518–527. Buchanan, T. W., Karafin, M. S., & Adolphs, R. (2003). Selective effects of triazolam on memory for emotional, relative to neutral, stimuli: Differential effects on gist versus detail. Behavioral Neuroscience, 117, 517–525. Buchanan, T. W., & Lovallo, W. R. (2001). Enhanced memory for emotional material following stress-level cortisol treatment in humans. Psychoneuroendocrinology, 26, 307–317. Buchanan, T. W., & Tranel, D. (2008). Stress and emotional memory retrieval: Effects of sex and cortisol response. Neurobiology of Learning and Memory, 89, 134–141. Cahill, L. (2003). Sex- and hemisphere-related influences on the neurobiology of emotionally influenced memory. Progress in Neuropsychopharmacology and Biological Psychiatry, 27, 1235–1241. Cahill, L. (2006). Why sex matters for neuroscience. Nature Reviews Neuroscience, 7, 477–484. Cahill, L., Gorski, L., & Le, K. (2003). Enhanced human memory consolidation with post-learning stress: Interaction with the degree of arousal at encoding. Learning and Memory, 10, 270–274. Chatterton, R. T., Jr., Vogelsong, K. M., Lu, Y. C., Ellman, A. B., & Hudgens, G. A. (1996). Salivary alpha-amylase as a measure of endogenous adrenergic activity. Clinical Physiology, 16, 433–448. de Kloet, E. R., Joels, M., & Holsboer, F. (2005). Stress and the brain: From adaptation to disease. Nature Reviews Neuroscience, 6, 463–475. de Quervain, D. J., Aerni, A., & Roozendaal, B. (2007). Preventive effect of betaadrenoceptor blockade on glucocorticoid-induced memory retrieval deficits. American Journal of Psychiatry, 164, 967–969. Author's personal copy 326 D. Preuß, O.T. Wolf / Neurobiology of Learning and Memory 92 (2009) 318–326 de Quervain, D. J., Henke, K., Aerni, A., Treyer, V., McGaugh, J. L., Berthold, T., et al. (2003). Glucocorticoid-induced impairment of declarative memory retrieval is associated with reduced blood flow in the medial temporal lobe. European Journal of Neuroscience, 17, 1296–1302. Diamond, D. M., Campbell, A. M., Park, C. R., Halonen, J., & Zoladz, P. R. (2007). The temporal dynamics model of emotional memory processing: A synthesis on the neurobiological basis of stress-induced amnesia, flashbulb and traumatic memories, and the Yerkes–Dodson law. Neural Plasticity, 2007, 1–33. Dickerson, S. S., & Kemeny, M. E. (2004). Acute stressors and cortisol responses: A theoretical integration and synthesis of laboratory research. Psychological Bulletin, 130, 355–391. Elzinga, B. M., Bakker, A., & Bremner, J. D. (2005). Stress-induced cortisol elevations are associated with impaired delayed, but not immediate recall. Psychiatry Research, 134, 211–223. Heuer, F., & Reisberg, D. (1990). Vivid memories of emotional events: The accuracy of remembered minutiae. Memory and Cognition, 18, 496–506. Jackson, E. D., Payne, J. D., Nadel, L., & Jacobs, W. J. (2006). Stress differentially modulates fear conditioning in healthy men and women. Biological Psychiatry, 59, 516–522. Jelici, M., Geraerts, E., Merckelbach, H., & Guerrieri, R. (2004). Acute stress enhances memory for emotional words, but impairs memory for neutral words. International Journal of Neuroscience, 114, 1343–1351. Joels, M., Pu, Z., Wiegert, O., Oitzl, M. S., & Krugers, H. J. (2006). Learning under stress: How does it work? Trends in Cognition Science, 10, 152–158. Kelly, M. M., Tyrka, A. R., Anderson, G. M., Price, L. H., & Carpenter, L. L. (2008). Sex differences in emotional and physiological responses to the Trier Social Stress Test. Journal of Behavior Therapy and Experimental Psychiatry, 39, 87–98. Kirschbaum, C., & Hellhammer, D. H. (1989). Salivary cortisol in psychobiological research: An overview. Neuropsychobiology, 22, 150–169. Kirschbaum, C., Kudielka, B. M., Gaab, J., Schommer, N. C., & Hellhammer, D. H. (1999). Impact of gender, menstrual cycle phase, and oral contraceptives on the activity of the hypothalamus-pituitary-adrenal axis. Psychosomatic Medicine, 61, 154–162. Kirschbaum, C., Pirke, K. M., & Hellhammer, D. H. (1993). The ‘Trier Social Stress Test’ – a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology, 28, 76–81. Kirschbaum, C., Pirke, K. M., & Hellhammer, D. H. (1995). Preliminary evidence for reduced cortisol responsivity to psychological stress in women using oral contraceptive medication. Psychoneuroendocrinology, 20, 509–514. Kirschbaum, C., Schommer, N., Federenko, I., Gaab, J., Neumann, O., Oellers, M., et al. (1996). Short-term estradiol treatment enhances pituitary-adrenal axis and sympathetic responses to psychosocial stress in healthy young men. Journal of Clinics in Endocrinology and Metabolism, 81, 3639–3643. Kirschbaum, C., Wolf, O. T., May, M., Wippich, W., & Hellhammer, D. H. (1996). Stress- and treatment-induced elevations of cortisol levels associated with impaired declarative memory in healthy adults. Life Sciences, 58, 1475–1483. Kirschbaum, C., Wust, S., & Hellhammer, D. (1992). Consistent sex differences in cortisol responses to psychological stress. Psychosomatic Medicine, 54, 648–657. Kudielka, B. M., Schommer, N. C., Hellhammer, D. H., & Kirschbaum, C. (2004). Acute HPA axis responses, heart rate, and mood changes to psychosocial stress (TSST) in humans at different times of day. Psychoneuroendocrinology, 29, 983–992. Kuhlmann, S., Piel, M., & Wolf, O. T. (2005). Impaired memory retrieval after psychosocial stress in healthy young men. Journal of Neuroscience, 25, 2977–2982. Kuhlmann, S., & Wolf, O. T. (2005). Cortisol and memory retrieval in women: Influence of menstrual cycle and oral contraceptives. Psychopharmacology (Berlin), 183, 65–71. Kuhlmann, S., & Wolf, O. T. (2006a). A non-arousing test situation abolishes the impairing effects of cortisol on delayed memory retrieval in healthy women. Neuroscience Letters, 399, 268–272. Kuhlmann, S., & Wolf, O. T. (2006b). Arousal and cortisol interact in modulating memory consolidation in healthy young men. Behavioral Neuroscience, 120, 217–223. Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (1997). International Affective Picture System (IAPS): Technical manual and affective ratings. Gainesville, FL: Center for Research in Psychophysiology, University of Florida. Nater, U. M., La Marca, R., Florin, L., Moses, A., Langhans, W., Koller, M. M., et al. (2006). Stress-induced changes in human salivary alpha-amylase activity – associations with adrenergic activity. Psychoneuroendocrinology, 31, 49–58. Park, C. R., Zoladz, P. R., Conrad, C. D., Fleshner, M., & Diamond, D. M. (2008). Acute predator stress impairs the consolidation and retrieval of hippocampusdependent memory in male and female rats. Learning and Memory, 15, 271–280. Payne, J. D., Jackson, E. D., Hoscheidt, S., Ryan, L., Jacobs, W. J., & Nadel, L. (2007). Stress administered prior to encoding impairs neutral but enhances emotional long-term episodic memories. Learning and Memory, 14, 861–868. Payne, J. D., Jackson, E. D., Ryan, L., Hoscheidt, S., Jacobs, J. W., & Nadel, L. (2006). The impact of stress on neutral and emotional aspects of episodic memory. Memory, 14, 1–16. Preuss, D., Schoofs, D., & Wolf, O. T. (in press). Associations between endogenous cortisol levels and emotional memory: Influence of encoding instructions. Stress. Quevedo, J., Sant’ Anna, M. K., Madruga, M., Lovato, I., de Paris, F., Kapczinski, F., et al. (2003). Differential effects of emotional arousal in short- and long-term memory in healthy adults. Neurobiology of Learning and Memory, 79, 132–135. Rohleder, N., Nater, U. M., Wolf, J. M., Ehlert, U., & Kirschbaum, C. (2004). Psychosocial stress-induced activation of salivary alpha-amylase: An indicator of sympathetic activity? Abdominal Aortic Aneurysm: Genetics, Pathophysiology, and Molecular Biology, 1032, 258–263. Roozendaal, B., de Quervain, D. J., Ferry, B., Setlow, B., & McGaugh, J. L. (2001). Basolateral amygdala–nucleus accumbens interactions in mediating glucocorticoid enhancement of memory consolidation. Journal of Neuroscience, 21, 2518–2525. Roozendaal, B., Nguyen, B. T., Power, A. E., & McGaugh, J. L. (1999). Basolateral amygdala noradrenergic influence enables enhancement of memory consolidation induced by hippocampal glucocorticoid receptor activation. Proceedings of the National Academy of Sciences of the United States of America, 96, 11642–11647. Roozendaal, B., Okuda, S., de Quervain, D. J., & McGaugh, J. L. (2006). Glucocorticoids interact with emotion-induced noradrenergic activation in influencing different memory functions. Neuroscience, 138, 901–910. Sandi, C., Loscertales, M., & Guaza, C. (1997). Experience-dependent facilitating effect of corticosterone on spatial memory formation in the water maze. European Journal of Neuroscience, 9, 637–642. Schwabe, L., Bohringer, A., Chatterjee, M., & Schachinger, H. (2008). Effects of prelearning stress on memory for neutral, positive and negative words: Different roles of cortisol and autonomic arousal. Neurobiology of Learning and Memory, 90, 44–53. Smeets, T., Otgaar, H., Candel, I., & Wolf, O. T. (2008). True or false? Memory is differentially affected by stress-induced cortisol elevations and sympathetic activity at consolidation and retrieval. Psychoneuroendocrinology. Stark, R., Wolf, O. T., Tabbert, K., Kagerer, S., Zimmermann, M., Kirsch, P., et al. (2006). Influence of the stress hormone cortisol on fear conditioning in humans: Evidence for sex differences in the response of the prefrontal cortex. Neuroimage, 32, 1290–1298. Stroud, L. R., Salovey, P., & Epel, E. S. (2002). Sex differences in stress responses: Social rejection versus achievement stress. Biological Psychiatry, 52, 318–327. van Stegeren, A., Rohleder, N., Everaerd, W., & Wolf, O. T. (2006). Salivary alpha amylase as marker for adrenergic activity during stress: Effect of betablockade. Psychoneuroendocrinology, 31, 137–141. van Stegeren, A. H., Wolf, O. T., & Kindt, M. (2008). Salivary alpha amylase and cortisol responses to different stress tasks: Impact of sex. International Journal of Psychophysiology, 69, 33–40. Watson, D., Clark, L. A., & Tellegen, A. (1988). Development and validation of brief measures of positive and negative affect: The PANAS scales. Journal of Personality and Social Psychology, 54, 1063–1070. Wolf, O. T. (2008). The influence of stress hormones on emotional memory: Relevance for psychopathology. Acta Psychologica, 127, 513–531. Wolf, O. T., Schommer, N. C., Hellhammer, D. H., McEwen, B. S., & Kirschbaum, C. (2001). The relationship between stress induced cortisol levels and memory differs between men and women. Psychoneuroendocrinology, 26, 711–720. Zorawski, M., Blanding, N. Q., Kuhn, C. M., & LaBar, K. S. (2006). Effects of stress and sex on acquisition and consolidation of human fear conditioning. Learning and Memory, 13, 441–450. Studie 3 4 STUDIE 3: T HE 65 STRESSED ST UDENT: UNIVERSITY ST UDENTS INFLUENCE OF WRITTEN EXAM S AND ORAL PRESENT AT IONS ON SALIVARY CORTISOL CONCENTRATIONS IN Studie 3: The stressed student: Influence of written exams and oral presentations on salivary cortisol concentrations in university students Diana Preuß, Daniela Schoofs, Wolff Schlotz, Oliver T. Wolf The stressed student: Influence of written exams and oral presentations on salivary cortisol concentrations in university students Diana Preuß a (MSc), Daniela Schoofs a (MSc), Wolff Schlotz b (PhD), Oliver T. Wolf a (PhD) * a) Department of Cognitive Psychology, Ruhr-University Bochum, Germany b) School of Psychology, University of Southampton Running title: Cortisol response to university stressors Address for correspondence Oliver T. Wolf Department of Cognitive Psychology Ruhr-University Bochum Universitätsstr. 150 D-44780 Bochum Germany Phone: +49 (0)234 32 22670 Fax: +49 (0) 234 32 14308 e-mail: [email protected] Manuscript pages: 17; Figures = 3; Tables = 0. Key words: Cortisol, salivary samples, socio-evaluative threat, stress, university students ABSTRACT Laboratory research has demonstrated that social-evaluative threat has an influence on the hypothalamus pituitary adrenal axis (HPA). In two studies using independent samples we evaluated the anticipatory cortisol response to a written university examination (n = 35) and to an oral presentation (n = 34). Saliva samples were collected before and after the examinations and on a control day. Additionally saliva samples were collected on the day before the written examination and a control day. Results revealed significantly elevated cortisol concentrations on the day prior to the exam; however this effect occurred only in those participants who had their control day after the exam. Cortisol concentrations were elevated on the exam day, with elevated concentrations before but not after the examination. For the oral presentation study results revealed substantially elevated cortisol concentrations before and after the oral presentation. Taken together results indicate that written exams cause a mild anticipatory HPA response while oral presentations induce a strong HPA response. These findings appear to support the idea that social-evaluative threat is an important factor determining the size of the HPA response to laboratory stressors as well as to real-life stressors. 2 INTRODUCTION In the case of threat or challenge the body answers with an adaptive reaction to cope with the situation. This stress response includes an enhanced activity of the sympathetic nervous system (SNS) and the hypothalamic-pituitary- adrenal (HPA) axis. Activation of the HPA axis leads to an increased secretion of glucocorticoids (GCs; particularly cortisol in humans) from the adrenal cortex (de Kloet, Joels, & Holsboer, 2005). This hormone can be measured in saliva (Kirschbaum & Hellhammer, 1989). It has been repeatedly shown that the two stress systems influence cognitive and affective processes (de Kloet et al., 2005; Lupien, Maheu, Tu, Fiocco, & Schramek, 2007; Wolf, 2008). In addition multiple target systems in the periphery (cardiovascular system, immune system, glucoregulatory system) are influenced by these stress mediators (McEwen, 1998). It long has been conceptualised that in humans the situational factors novelty and uncontrollability in combination with ego involvement lead to psychological stress (Mason, 1968). More recently it has been hypothesized that a threat to the social self (e.g. status, reputation) is especially stressful (Dickerson, Gruenewald, & Kemeny, 2004) for humans as social individuals. Indeed by analysing over 100 laboratory studies Dickerson and Kemeny (2004) were able to show that social evaluative threat was a potent predictor for the stress response of the HPA axis (Dickerson & Kemeny, 2004). Thus social-evaluative threat seems to be an important prerequisite to cause an HPA response in the laboratory. However, it remains controversial whether the findings from studies with controlled experimental stressors can be predictive of cortisol reactions to real life stressors (Cohen & Hamrick, 2003; Kamarck & Lovallo, 2003; van Eck, Nicolson, Berkhof, & Sulon, 1996). To heighten ecological validity the physiological response to naturalistic stressors can be assessed. University life contains several different stressors for students and examinations are often used as real-life stressors (Stowell, 2003). There are different forms of examinations and two kinds are very popular for studies evaluating university stress. On the one hand there are oral examinations (Schoofs, Hartmann, & Wolf, 2008) and on the other 3 hand written examinations (Gaab, Sonderegger, Scherrer, & Ehlert, 2006; Ng, Koh, Mok, Chia, & Lim, 2003). While the results for oral examinations are mostly homogenous, the empirical picture is less clear for written examinations. Oral examinations were found to be associated with increases in cortisol (Herbert, Moore, de la Riva, & Watts, 1986; Lacey et al., 2000; Schoofs et al., 2008). For example we recently observed that cortisol concentrations of university students were increased by more two fold immediately before undergoing an oral exam (Schoofs et al., 2008). In written examinations results are less consistent. While some studies report stress effect (Loft et al., 2007; Lovallo, Pincomb, Edwards, Brackett, & Wilson, 1986), other studies failed to find effects on cortisol (Spangler, 1997). It is obvious that the results in the field of examination stress are heterogenic and a consensus about the influence of exam stress on the neuroendocrine stress response has not been reached until now. There are several possible reasons for these inhomogeneous results. First there are large differences in the written examinations used in the different studies (Stowell, 2003). While some authors investigated a single written examination (Frankenhaeuser et al., 1978), other authors assessed the hormonal stress response during a whole examination period (Loft et al., 2007). During a period of examinations anticipatory effects might exert additional influences on the stress response. Additionally the impact of the evaluated examination on the students’ life also differs between the studies. While some studies evaluate matriculation examinations (Frankenhaeuser et al., 1978), other studies evaluate the impact of final examinations in medical school (Zeller, Handschin, Gyr, Martina, & Battegay, 2004) or written examinations in undergraduate students (Gaab et al., 2006; Ng et al., 2003). These differences might account for the heterogeneity in the field of written examinations but there is still need for further investigation. Especially the impact of examination stress on the anticipatory stress response needs additional attention. A reason for the fact that the results for oral examinations are more homogeneous than the results for oral examinations might be that these two kinds of examinations differ in socialevaluative threat. While oral examinations always contain a social-evaluative threat because of the listening and judging audience, written examinations lack this component. This lack of 4 social-evaluative threat, which is an important factor in laboratory research (Dickerson & Kemeny, 2004), might also influence the magnitude of the stress response in real-life stress. Although examinations are popular real-life stressors other stressors in university context exist. Examples are oral presentations in university courses. These situations contain performance pressure and social-evaluative threat induced by the listening audience and therefore can be compared with oral examinations. However, the impact on the students’ overall grade level is typically smaller for oral presentations. Moreover the factor “uncontrollability” (Dickerson & Kemeny, 2004) appears to be less present during oral presentations, since the students can determine most of the action, in contrast to exams, where the students have to respond to the raised questions. However, the impact of socialevaluative threat in university situations with mild performance pressure, low uncontrollability but high social-evaluative threat has not been investigated yet. Thus the characterisation of the HPA response to oral presentations is of interest. In light of the heterogeneity of previous findings it becomes clear that the influence of university stress on the HPA axis needs further investigation. The aim of the present study therefore was to evaluate the impact of stress on the cortisol response in university students. Therefore two studies were conducted. In a first study we assessed the influence of a single written examination on the cortisol stress response. Additionally daytime cortisol concentrations were assessed on the day before the examination and the day before the control day. A second study was conducted to evaluate the influence of an oral presentation on the cortisol stress response. For the written examination we hypothesized an anticipatory stress response with higher cortisol concentrations on the day before the examination compared to the day before the control day. We expected both the written examination and the oral presentation to elicit a significant stress response because both types of performance tests are potential stressors for students. Because of the stronger impact of social evaluative threat in oral presentations we hypothesized that in the oral presentation study a stronger cortisol response would be observable compared to the written examination study. 5 MATERIALS AND METHODS Written examination study Thirty five undergraduate psychology students (4 males: mean age = 25.0 ± 1.12; 31 females: mean age = 23.42 ± 1.11 (mean ± SEM)) participated in this study. The averaged BMI was 20.97 ± 0.39 kg/m2 for the females and 21.59 ± 1.92 kg/m2 for the males. Twenty three female participants used oral contraceptives. No information about the stage of the menstrual cycle was collected. Four participants suffered from hypothyroidism but were under stable medicament substitution. Eight participants were smokers. Smokers reported to smoke between 0.5 and 18 (7.5 ±2.36) cigarettes per day. Subjects gave written informed consent and were paid for participating. The study was approved by the national ethic committee of the German Psychological Association (DGPs). Oral presentation study 37 students (28 females, 9 males) participated in the study. Three participants showed cortisol concentrations above 100nmol/liter (indicative of sample contamination and/or acute disease) and therefore were excluded from the analyses. Data of 34 students (7 males, 27 females) were analysed. The mean age of males was 25.28 ± 1.79 years. Mean age of females was 24.96 ± 1.25 years. One female participant suffered from hypothyroidism but was under stable medicament substitution. Sixteen of the 28 female participants used oral contraceptives. The averaged body-mass-index for males was 24.67 ± 2.02 and for the females 22.06 ± 0.68. Experimental procedure Written examination study Subjects took part in written exams at the end of the winter semester (February or March; n = 20) or at the end of the following summer semester (July; n = 15). All exams started in the morning at 9am or 10am and lasted about two and a half hours. Participants took part in a control day taking place between 4-8 days after or before the exam. For practical reasons 6 participants could choose on which day they would participate in the control day. Nine participants took part in the control session before the examination and the remaining 26 after the examination. This session started at the same time as the exams did (9am or 10am respectively) and participants solved some filler tasks in a seminar room of the university. Time of awakening was recorded on both days. Saliva samples In total participants collected ten saliva samples. Participants were told not to smoke, eat or drink anything (except water) for at least 30 minutes prior to each saliva collection. Two salivettes were handed out to the participants in front of the exam room. One sample was collected immediately before the students entered the exam room and the second one was given to the students with the instruction to collect saliva immediately after finishing the exam. Students wrote down the time, when they collected the second saliva sample. On the control day the saliva samples were collected at the same times as on the exam day. The first saliva sample was collected at 9am or 10am and the second one at the same time as students collected the sample at the exam day. Students who took part in the control day before the examination were asked to collect the second saliva sample on the control day at the time when the exam would probably end. Saliva samples were kept refrigerated and brought to the exam or control day respectively. Additionally to the saliva samples on the control and examination day, participants were asked to collect three saliva samples on the day before examination and control day at 10am, 4pm and 9pm respectively. This was done to evaluate the daytime cortisol concentrations on the day before a written examination in order to detect possible early anticipatory cortisol increases. Saliva samples were kept refrigerated and were brought by the participants to the university on the exam or control day respectively. Saliva was collected using salivette collection devices (Sarstedt, Nuembrecht, Germany). Cortisol was measured using a commercially available immunoassay (IBL, Hamburg, 7 Germany). Inter- and intra-assay variations were below 10%. Analyses were carried out at Professor Kirschbaums laboratory at the Technical University of Dresden, Germany. Oral presentation study All participants gave an oral presentation in a university course. Saliva was collected before the beginning of the presentation and immediately after finishing it. Proper sampling was controlled by one of the authors (OTW). Additionally participants collected data after their presentation at the identical times during listening to a different presentation in the same university course. Saliva was collected and cortisol was analysed identical to the previous study. Mean duration of the oral presentation was 43:58 (± 2:54) minutes. The oral presentations started between 10 a.m. and 4 p.m. (mean 11:54 a.m. ± 0:20) Statistical analysis Differences in cortisol levels between academic assessments and control days were tested by mixed models using a repeated measures design with an unstructured error covariance matrix. Effect of interest were: average cortisol differences between academic assessment and control day (main effect day), average differences between assessment times within days (main effect time) and the difference in the time course of cortisol between academic assessment and control day (interaction day x time). Analyses of written exams were adjusted for gender, smoking status, season, oral contraceptive (OC) use and order of measurement days (exam day or control day first), and time of getting up was included as a time-varying covariate. Analyses of oral presentations were adjusted for sex, OC use, time of day when the presentation was given and duration of the presentation. All analyses were done using SPSS Statistics v17 (SPSS Inc., Chicago, USA). We used an alpha level of .05 for all statistical tests. 8 RESULTS Written examination study Cortisol concentrations on the day prior to the written exam (daytime cortisol concentrations) Participants reported waking up at 8:28am (± 0:11) on the day prior to the examination day and at 8:42 am (± 0:12) on the day prior to the control day. The wake up times were not significantly different from each other (t(31) = -0.775, p = 0.44). Cortisol measurements in the morning, afternoon and late evening on the day preceding the exam (or the control day) are presented in Figure 1a and b. Results revealed the expected decline of cortisol levels between 10am and 9pm (p < .001). Average cortisol levels were higher on the day before the exam (p = .022). Although the test of differential cortisol courses fell short of significance (p = .106), the differences in average cortisol levels seemed to be largely due to differences in the 4pm cortisol levels (4pm diff = 1.4 nmol/L; p = .002; 10am diff = 0.9 nmol/L, p = .313; 9pm diff = 0.4 nmol/L; p = .147). In contrast to the cortisol concentrations on the exam day (see below), overall cortisol concentrations were higher in participants who did the control and exam measurements during winter (p = .009), and there was a trend towards higher cortisol levels in women (p = .064). OC usage and smoking had no significant effect. Participants had higher cortisol levels when they reported getting up later on that day (p = .011). Overall cortisol levels were higher in participants who did the control measurements before the exam measurements (p < .001). Including the interaction of order with exam in the model showed a borderline significant effect (p = .053; see Figure 1a and b). Mean cortisol levels on the day before the exam were higher than those on the control day only if the control day measurements were done after the exam (diff = 1.2 nmol/L; p = .005) but not when they were done before the exam (diff = 0.3 nmol/L; p = .617). 9 Fig. 1: Salivary cortisol concentrations before the day before the written exam and the day before the control day for 10am, 4pm and 9pm for a) participants participating in the control day after the examination (N=26) and b) participants participating in the control day before the examination (N=9; results show mean ± SEM). Mixed model analysis revealed that cortisol concentrations were significantly elevated on the day preceding the exam. Further analyses showed that this effect was due to differences at 4pm (4pm diff = 1.4 nmol/L; p = .002). However this effect was only detected in those participants who took part in the control day after the written exam (interaction between exam day and order). Cortisol concentrations on the exam day 10 Participants reported to wake up at approx. 6:52am (± 0:07) on the examination day and at approx. 7:10am (± 0:06) on the control day. This difference (although relatively small) revealed to be statistically significant (t(33) = 2.075, p < 0.05). Cortisol concentrations on the day of the exam and the control day are shown in Figure 2. Mean cortisol concentrations were slightly higher on the day of the exam (p = .030). Cortisol concentrations on average were smaller at the second sampling point (post exam or post control day; p < .001), and this circadian decline tended to be more pronounced on the exam day (p = .067) due to a higher cortisol level before the exam (diff = 3.5 nmol/L; p = .019). Similar to the analysis of the daytime cortisol concentrations, participants tended to have higher overall cortisol concentrations when they reported getting up later on that day (p = .085) and when they did the control day before the exam day (both p = .084). However, the order of the measurement day did not interact with day and time, indicating that the cortisol differences between exam and control day were similar in both conditions (in contrast to results obtained for the daytime cortisol concentrations). In this analysis season, smoking, gender and OC usage had no significant effect. Fig. 2: Salivary cortisol concentrations before and after a written exam and a control day (N=35; results show mean ± SEM). The exams lasted on average about 2 ½ hours and started in the morning between 9 and 10 a.m. Mixed model analysis revealed that cortisol concentrations were significantly elevated on the exam day, with the effect being most pronounced before the exam (diff = 3.5 nmol/L; p = .019). 11 Oral presentation study Results for the cortisol measurements before and after the oral presentation and the control day are shown in Figure 3. The comparison of cortisol changes on these days showed opposite trends, resulting in a negligible overall time effect (p = .764) but a highly significant interaction day x time (p = .010). In addition, overall cortisol levels were clearly higher on the day of the oral presentation (p < .001). While there was already a significant difference before the oral presentation (diff pre = 5.1 nmol/L; p = .001), the difference increased until after the presentation (diff post = 12.0 nmol/L; p < .001). This was due to a significant decrease on the control day (diff pre-post = 3.1 nmol/L; p = .002) and a marked increase, with higher variability, on the day of the oral presentation (diff pre-post = -3.8 nmol/L; p = .089). There was no significant influence of the time or duration of the presentation. Men showed higher average cortisol levels (p = .035), but OC usage had no significant effect. The time of day when the presentation was given as well as the duration of the presentation had no significant influence on the measured cortisol concentrations. Fig. 3: Salivary cortisol concentrations before and after an oral presentation and a control day (N=34; results show mean ± SEM). Mixed model analysis revealed that cortisol concentrations were significantly elevated before (diff pre = 5.1 nmol/L; p = .001) and after the oral presentation (diff post = 12.0 nmol/L; p < .001). 12 DISCUSSION The aim of the present study was to characterize the cortisol stress response to different forms of university stress. Therefore two studies were conducted and the cortisol stress response to a written examination and an oral presentation in two samples of undergraduate students was assessed. Additionally we assessed the cortisol concentrations on the day before the written examination and the day before the control day to evaluate possible anticipatory stress effects to the written examination. For the written examination results revealed significantly higher cortisol concentrations on the day preceding the exam. This effect was modulated by order, since it could only be detected in participants who participated in the control day after the written exam. With respect to the exam day itself participants showed again higher cortisol concentrations and this was most pronounced before the exam. For the oral presentation results revealed substantial differences before the presentation which further increased until the post presentation measurement. The finding of a hormonal stress response to academic stressors is in line with findings of several studies investigating the influence of examinations on cortisol release (Al-Ayadhi, 2005; Frankenhaeuser et al., 1978; Lindahl, Theorell, & Lindblad, 2005; Lovallo et al., 1986; Schoofs et al., 2008). However there are some studies reporting no significant influence of examinations on cortisol release (Frankenhaeuser et al., 1978; Malarkey, Pearl, Demers, Kiecolt-Glaser, & Glaser, 1995; Spangler, 1997) or even a decrease (Loft et al., 2007). These inhomogenous effects might be in part due to the differences in the experimental designs. As Stowell (2003) pointed out are there some problems in comparing different studies in the field of examination stress. Studies reporting significant elevated cortisol responses to examination stress use different designs. While some studies assess the hormonal response to one discrete examination (Schoofs et al., 2008; Spangler, 1997) other studies evaluate the cortisol concentrations during longer periods of examinations (Loft et al., 2007; Weekes et al., 2006). Comparability of the studies is furthermore limited because of the fact that some studies do not report if the examinations were oral or written examinations (Al-Ayadhi, 2005; 13 Lovallo et al., 1986; Malarkey et al., 1995). While results for written examinations are inhomogeneous oral examinations have been repeatedly found to produce a strong stress response (Herbert et al., 1986; Schoofs et al., 2008). For example a recent study of ours evaluated the impact of an oral examination on the acute cortisol release and found substantially (two to three fold) elevated mean cortisol concentrations on the exam day (Schoofs et al., 2008). A characteristic of oral examinations is the experience of social threat. In laboratory studies social threat is known to be a strong factor in determining the size of the HPA response (Dickerson & Kemeny, 2004) but its impact on naturalistic stressors has not been evaluated sufficiently yet. In our oral examination study grade pressure was combined with social-evaluative threat triggered by the presence of an auditor (the professor as the main examiner present at the oral exam) and a co-auditor (a co-worker taking notes; (Schoofs et al., 2008). Written examinations on the other hand also induce grade pressure, but the experience of social threat is much weaker. Students in written examinations have to show their knowledge about a special topic but the written examination is graded afterwards without the students being present. Therefore the experience of social threat is weaker compared to an oral examination where the performance is graded and assessed directly when the students are present. In line with these arguments the present written examination study revealed a significant but small effect of the exam on salivary cortisol concentrations. An interesting finding was observed by evaluating cortisol concentrations one day before the written examination. Cortisol concentrations of the students were already elevated on the day prior to the exam, indicative of an anticipatory HPA response. A recent study in ballroom dancers (Rohleder, Beulen, Chen, Wolf, & Kirschbaum, 2007) observed anticipatory HPA responses several hours before the start of the tournament. Our findings replicate and extend these observations by demonstrating elevated cortisol concentrations on the day preceding the exam. It has to be noted that an anticipatory HPA response on the day preceding the exam could only be detected in those participants in whom the control day took part after the exam day. Thus students who were approx. one week away from a written exam might already show elevated cortisol concentrations, which would support the idea that 14 exam periods are characterized by elevated cortisol concentrations (e.g. see Loft et al., 2007; Weekes et al., 2006). In contrast students who have completed an exam about one week ago appear to show already a normal (lower) HPA activity when measured during the course of the day. This finding emphasizes the need to pay close attention to the issue of order as well as temporal distance between the exam and control condition in future studies on this issue (see Stowell, 2003). An interesting side finding was the fact that cortisol levels were overall higher when the sampling took place during the winter months. This is in line with other studies suggesting that HPA activity is stronger in winter months (King et al., 2000; Walker, Best, Noon, Watt, & Webb, 1997) and indicates that seasonal factors are able to influence the HPA response in humans. Another important modulating variable was wake up time. Participants who reported to wake up later displayed in both parts of the written exam study higher cortisol concentrations. This highlights the need to pay close attention to inter as well as intra-individual alterations in wake up time when conducting studies investigating HPA activity/reactivity. In order to evaluate another form of university stress in students, the second study was conducted. Students had to perform oral presentations in university courses in front of the course. Results of the oral presentation study show that the oral presentations elicited a strong cortisol stress response which resulted in elevated cortisol concentrations before and after the oral presentation. The cortisol elevations were much more pronounced when compared to the written exam. However, compared to our previous oral examination study (Schoofs et al., 2008) the effects were smaller. While a direct comparison of these different studies is problematic, the three studies discussed were highly similar with respect to sample size, gender distribution, and OC use. It seems that the social-evaluative threat in oral presentations makes these situations more stressful (with respect to an HPA activation) than written examinations. Oral presentations on the other hand seem to be less stressful than oral examinations, maybe because students experience an oral presentation as less uncontrollable as an oral exam. It is known from 15 laboratory research that social-evaluative threat in combination with uncontrollability are high potent stressors (Dickerson & Kemeny, 2004). From the present results this also seems to be the case for real-life. Nevertheless, it has to be mentioned that of course social-evaluative threat is not the only factor that differs between oral presentations, oral examinations and written examinations. Other factors may be novelty, preparedness or predictability (Mason, 1968). We found some evidence for sex differences in the two current studies, but the effects were not consistent. Moreover they reflected higher overall cortisol concentrations in women (exam study) or men (oral presentation study) rather than differences in cortisol reactivity. Because of the small number of male participants in the current studies these findings should be interpreted with caution. The present results reveal no influence of oral contraceptives in the written examination study and the oral presentation study. In laboratory stress studies the use of oral contraceptives is known to alter the cortisol stress response in women, which might be in part mediated by an increased production of the cortisol binding globulin CBG. Women using oral contraceptives show a blunted free salivary stress response to psychosocial laboratory stressors like the Trier Social Stress Test (Kirschbaum, Kudielka, Gaab, Schommer, & Hellhammer, 1999). Our recent oral exam study found no influence of oral contraceptives on the cortisol stress response (Schoofs et al., 2008). Similarly in the present studies no impact of OC use was detected. It appears that the inability of OC using women to mount a free cortisol response is restricted to moderate and surprising stressful events induced in the laboratory, which most likely reflect a single brief HPA activation. In contrast the anticipatory HPA response to social evaluative threat appears to start hours (Rohleder et al., 2007) or days (see daytime cortisol concentrations of the written exam study) ahead of the event and might thus allow the organism to assure a robust free cortisol increase via a feedback based repeated activation of the axis. 16 There are several limitations of our two studies which need to be acknowledged. We investigated the effects of a written exam and an oral presentation in two different samples of students. This limits the comparability between the two situations. Ideally we would have tested the same students in those two different real life stressors. In the context of this study such an approach was not feasible due to organisational issues, since most students we had access to had to take part in a written exam or an oral presentation. However least at a descriptive level (sample size, male female ratio, percentages of women using OCs) the two study groups were quite similar. The sample size was medium (n=35 for the written examination study and n = 34 for the oral presentation study) and was restricted to psychology students. Although we conducted analysis for the influence of oral contraceptives it was not possible to control for the phase of menstrual cycle. Additionally only four males participated in the written examination study and seven males in the oral presentation study but this mirrors the fact that much more females study psychology in Germany and male students are thus harder to find. Thus the potential influence of sex (or gender) could not be addressed with an adequate power in this study. The HPA response to academic stressors might be influenced by anxiety and preparedness. It thus would have been desirable to measure these constructs with the current study. Another aspect is the fact that we did not have any information about the performance of the participants in the written examination. Although it would be interesting to assess possible relationships between performance and the cortisol response, we did not collect information about performance because of privacy reasons. For the assessment of daytime cortisol levels participants were instructed to collect saliva during the day on their own, without any control of compliance. An ambulatory compliance assessment as described by Broderick et al. (2004) would have been desirable, but was not feasible in the present study. At least the proper sampling of saliva before and after the exam as well as before and after the oral presentation and the respective control days were controlled by the investigators. 17 Another limitation is the fact that we did not assign participants randomly to the control condition before or after the written examination. While such an approach would have been desirable from a methodological point of view this was not feasible in the framework of the current study since otherwise participation would have decreased further. This is an example of the sort of compromises a field study has to make. A possible anticipatory stress effect might have influenced the cortisol concentrations on the control day, when it took place before the examination. In fact this is exactly what we observed. Daytime cortisol concentrations were only elevated on the day preceding the exam in the participants who had their control day after the exam day. An alternative explanation could be that participants with a better preparation for the exam or with lower anxiety choose the control day before the examination, while participants with higher anxiety and a worse stress management choose the day after the examination. Since we did not randomise the order of exam and control days we can not decide between these different explanations for our empirical findings. We did not control for additional examinations around the time of the assessed written examination or the control day. An exam period could be associated with chronic stress leading to altered HPA activity which might influence the stress response to an acute stressor (Loft et al., 2007). Additionally chronic examination stress might influence cortisol concentrations on the control day as well (Weekes et al., 2006), which in our study was about one week apart from the exam itself. Thus in future studies an additional assessment of baseline cortisol concentrations several weeks before the exam period would be desirable. In addition measurements of chronic stress should be included. Cortisol is known to shown a strong circadian rhythm and the HPA reactivity might differ between the morning and the afternoon (Dickerson & Kemeny, 2004), but see Kudielka and Kirschbaum (2005). The written exams always took place in the morning, while the oral presentations took part in the morning, at noon or in the afternoon. Even though we found no evidence that the response to the oral presentation was modulated through time of day we can nevertheless not exclude the possibility that the more pronounced response to the oral 18 presentations is in part mediated by the fact that they differed in their starting time from the written exams. An additional factor that can influence HPA reactivity is the individual’s chrono type. Morning types were found to show higher cortisol morning levels than evening types (Bailey & Heitkemper, 1991; Bailey & Heitkemper, 2001; Griefahn & Robens, 2008; Kudielka, Bellingrath, & Hellhammer, 2007; Kudielka, Federenko, Hellhammer, & Wust, 2006). Moreover an exam in the morning may be more stressful for evening types than for morning types because the conditions are less optimal for the evening type (May & Hasher, 1998). Therefore future studies on this topic might want to collect information about the chronotype of their participants. In sum the present studies report an anticipatory stress response to a written examination. Cortisol concentrations on the day before the examination were elevated indicative of an anticipatory HPA response. Similarly cortisol concentrations were elevated immediately prior to the exam. Additionally in a second sample a cortisol stress response was observed to an oral presentation. Cortisol concentrations were elevated before and after the presentation and the effects were larger than those observed in response to the written exam. When compared to our previous oral examination study (Schoofs et al., 2008) the current data appear to suggest that written examinations are weaker stressors than oral examinations at least for their impact on the HPA axis. Oral presentations on the other hand seem to have less impact on the HPA axis than oral examinations but more impact than written examinations. A reason for that might be that oral examinations and oral presentations contain social-evaluative threat but that written examinations lack this factor. This interpretation of the current findings would support the notion that social evaluative threat is a major determent of the acute HPA response to challenge. However, this hypothesis needs to be substantiated with additional studies comparing cortisol responses to different real life stressors, ideally within the same participants. 19 Reference List Al-Ayadhi LY 2005. Neurohormonal changes in medical students during academic stress. Ann.Saudi.Med., 25, 36-40. Bailey SL and Heitkemper MM 1991. Morningness-eveningness and early-morning salivary cortisol levels. Biol.Psychol., 32, 181-192. Bailey SL and Heitkemper MM 2001. Circadian rhythmicity of cortisol and body temperature: morningness-eveningness effects. Chronobiol.Int., 18, 249-261. Broderick JE, Arnold D, Kudielka BM, and Kirschbaum C 2004. Salivary cortisol sampling compliance: comparison of patients and healthy volunteers. Psychoneuroendocrinology, 29, 636-650. Cohen S and Hamrick N 2003. Stable individual differences in physiological response to stressors: implications for stress-elicited changes in immune related health. Brain Behav.Immun., 17, 407-414. de Kloet ER, Joels M, and Holsboer F 2005. Stress and the brain: from adaptation to disease. Nat.Rev.Neurosci., 6, 463-475. Dickerson SS, Gruenewald TL, and Kemeny ME 2004. When the social self is threatened: shame, physiology, and health. J.Pers., 72, 1191-1216. Dickerson SS and Kemeny ME 2004. Acute stressors and cortisol responses: a theoretical integration and synthesis of laboratory research. Psychol.Bull., 130, 355-391. Frankenhaeuser M, von Wright MR, Collins A, von Wright J, Sedvall G, and Swahn CG 1978. Sex differences in psychoneuroendocrine reactions to examination stress. Psychosom.Med., 40, 334-343. Gaab J, Sonderegger L, Scherrer S, and Ehlert U 2006. Psychoneuroendocrine effects of cognitive-behavioral stress management in a naturalistic setting-a randomized controlled trial. Psychoneuroendocrinology, 31, 428-438. Griefahn B and Robens S 2008. The cortisol awakening response: a pilot study on the effects of shift work, morningness and sleep duration. Psychoneuroendocrinology., 33, 981-988. Herbert J, Moore GF, de la Riva C, and Watts FN 1986. Endocrine responses and examination anxiety. Biol.Psychol., 22, 215-226. Kamarck TW and Lovallo WR 2003. Cardiovascular reactivity to psychological challenge: conceptual and measurement considerations. Psychosom.Med., 65, 9-21. King JA, Rosal MC, Ma Y, Reed G, Kelly TA, Stanek EJ, III et al. 2000. Sequence and seasonal effects of salivary cortisol. Behav.Med., 26, 67-73. 20 Kirschbaum C and Hellhammer DH 1989. Salivary cortisol in psychobiological research: an overview. Neuropsychobiology, 22, 150-169. Kirschbaum C, Kudielka BM, Gaab J, Schommer NC, and Hellhammer DH 1999. Impact of gender, menstrual cycle phase, and oral contraceptives on the activity of the hypothalamus-pituitary-adrenal axis. Psychosom.Med., 61, 154-162. Kudielka BM, Bellingrath S, and Hellhammer DH 2007. Further support for higher salivary cortisol levels in "morning" compared to "evening" persons. J.Psychosom.Res., 62, 595-596. Kudielka BM, Federenko IS, Hellhammer DH, and Wust S 2006. Morningness and eveningness: the free cortisol rise after awakening in "early birds" and "night owls". Biol.Psychol., 72, 141-146. Kudielka BM and Kirschbaum C 2005. Sex differences in HPA axis responses to stress: a review. Biol.Psychol., 69, 113-132. Lacey K, Zaharia MD, Griffiths J, Ravindran AV, Merali Z, and Anisman H 2000. A prospective study of neuroendocrine and immune alterations associated with the stress of an oral academic examination among graduate students. Psychoneuroendocrinology, 25, 339356. Lindahl M, Theorell T, and Lindblad F 2005. Test performance and self-esteem in relation to experienced stress in Swedish sixth and ninth graders--saliva cortisol levels and psychological reactions to demands. Acta Paediatr., 94, 489-495. Loft P, Thomas MG, Petrie KJ, Booth RJ, Miles J, and Vedhara K 2007. Examination stress results in altered cardiovascular responses to acute challenge and lower cortisol. Psychoneuroendocrinology, 32, 367-375. Lovallo WR, Pincomb GA, Edwards GL, Brackett DJ, and Wilson MF 1986. Work pressure and the type A behavior pattern exam stress in male medical students. Psychosom.Med., 48, 125-133. Lupien SJ, Maheu F, Tu M, Fiocco A, and Schramek TE 2007. The effects of stress and stress hormones on human cognition: Implications for the field of brain and cognition. Brain Cogn., 65, 209-237. Malarkey WB, Pearl DK, Demers LM, Kiecolt-Glaser JK, and Glaser R 1995. Influence of academic stress and season on 24-hour mean concentrations of ACTH, cortisol, and beta-endorphin. Psychoneuroendocrinology, 20, 499-508. Mason JW 1968. A review of psychoendocrine research on the pituitary-adrenal cortical system. Psychosom.Med., 30, Suppl-607. May CP and Hasher L 1998. Synchrony effects in inhibitory control over thought and action. J.Exp.Psychol.Hum.Percept.Perform., 24, 363-379. 21 McEwen BS 1998. Stress, adaptation, and disease. Allostasis and allostatic load. Ann.N.Y.Acad.Sci., 840, 33-44. Ng V, Koh D, Mok BY, Chia SE, and Lim LP 2003. Salivary biomarkers associated with academic assessment stress among dental undergraduates. J.Dent.Educ., 67, 10911094. Rohleder N, Beulen SE, Chen E, Wolf JM, and Kirschbaum C 2007. Stress on the dance floor: the cortisol stress response to social-evaluative threat in competitive ballroom dancers. Personality and Social Psychology Bulletin, 33, 69-84. Schoofs D, Hartmann R, and Wolf OT 2008. Neuroendocrine stress responses to an oral academic examination: no strong influence of sex, repeated participation and personality traits. Stress., 11, 52-61. Spangler G 1997. Psychological and physiological responses during an exam and their relation to personality characteristics. Psychoneuroendocrinology, 22, 423-441. Stowell JR 2003. Use and abuse of academic examinations in stress research. Psychosom.Med., 65, 1055-1057. van Eck MM, Nicolson NA, Berkhof H, and Sulon J 1996. Individual differences in cortisol responses to a laboratory speech task and their relationship to responses to stressful daily events. Biol.Psychol., 43, 69-84. Walker BR, Best R, Noon JP, Watt GC, and Webb DJ 1997. Seasonal variation in glucocorticoid activity in healthy men. J.Clin.Endocrinol.Metab., 82, 4015-4019. Weekes N, Lewis R, Patel F, Garrison-Jakel J, Berger DE, and Lupien SJ 2006. Examination stress as an ecological inducer of cortisol and psychological responses to stress in undergraduate students. Stress, 9, 199-206. Wolf OT 2008. The influence of stress hormones on emotional memory: Relevance for psychopathology. Acta Psychol.(Amst)., 127, 513-531. Zeller A, Handschin D, Gyr N, Martina B, and Battegay E 2004. Blood pressure and heart rate of students undergoing a medical licensing examination. Blood Press, 13, 20-24. 22 Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 6 D I S K U S S I ON UND 87 AU S BLIC K 6.1 Zusammenfassung der Studien Das Ziel der drei durchgeführten Studien war die Untersuchung und Spezifizierung des Einflusses basaler Cortisolspiegel (Studie 1) und psychosozialen Stresses (Studie 2) auf die Gedächtnisleistung sowie die Spezifizierung von Faktoren und stressauslösenden Situationen außerhalb des Laborkontextes (Studie 3). Für diese Fragestellung wurden drei Studien durchgeführt. In den ersten beiden Studien, den Gedächtnisstudien, wurde jeweils dasselbe Gedächtnismaterial verwendet. Dabei handelte es sich um Bilder, die mit einem Begleitsatz, welcher zusätzliche Informationen erhielt, präsentiert wurden. Es wurden jeweils 5 emotional nicht erregende neutrale, 5 emotional erregende positive und 5 emotional erregende negative Bilder verwendet, wobei die positiven und negativen Bilder in der ersten Studie zu der Kategorie „emotional erregend“ zusammengefasst wurden. In den beiden ersten Studien wurde ein verzögerter Gedächtnistest eingesetzt, der aus einer freien Abrufaufgabe und einer Multiple-Choice Aufgabe bestand. In Studie 2 wurde zusätzlich eine direkte freie Abrufaufgabe verwendet, die unmittelbar nach der Enkodierung durchgeführt wurde. In der ersten Studie wurde zunächst der Einfluss basaler, also natürlich zirkulierender Cortisolspiegel auf die Gedächtnisleistung in einer weiblichen Stichprobe (N=56) untersucht. Zusätzlich wurde in dieser Studie der Einfluss der Enkodierungsstrategie betrachtet. Die Probanden erhielten entweder eine explizite Enkodierungsinstruktion, bei der die Versuchspersonen über den folgenden Gedächtnistest informiert wurden oder eine implizite Enkodierungsinstruktion, bei der die Probanden keine Information über den Gedächtnistest erhielten. Der Faktor der Enkodierungsstrategie wurde untersucht, um hieraus Implikationen für die nachfolgenden Studien zu erhalten. In der zweiten Studie wurde aufgrund der Ergebnisse der ersten Studie eine explizite Lerninstruktion verwendet. Die zweite Studie wurde durchgeführt, um die Auswirkungen eines psychosozialen Stressors (TSST) auf die Gedächtniskonsolidierung näher zu betrachten. Die zweite Studie wurde an einer für das Geschlecht gemischten Stichprobe (N=58; 28 Frauen und 30 Männer) durchgeführt, um den Einfluss dieses Faktors auf eventuelle Stresseffekte zu kontrollieren. In der dritten Studie schließlich wurde an zwei unabhängigen Stichproben untersucht, welchen Einfluss zwei universitäre Stresssituationen (schriftliche Klausur, N= 35 und mündliches Referat, N=34) auf die hormonelle Stressreaktion haben. Die Ergebnisse der ersten Studie zeigten keinen Effekt der Enkodierungsstrategie auf die Leistung der Probandinnen in dem durchgeführten verzögerten Gedächtnistest. Es fand sich Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 88 kein Unterschied in der Gedächtnisleistung zwischen der impliziten und der expliziten Enkodierungsinstruktion. Jedoch wurde ein positiver Zusammenhang zwischen der Höhe der basalen Cortisolspiegel und der Gedächtnisleistung für emotional erregende, nicht aber für emotional neutrale Items berichtet. Des Weiteren zeigte sich, dass die gefundenen Zusammenhänge lediglich in der Gruppe der expliziten Enkodierungsstrategie auftraten. Die Ergebnisse der Studie deuten somit darauf hin, dass höhere natürlich zirkulierende und nicht durch Stress erhöhte Cortisolspiegel das explizite Gedächtnis für emotional erregende Bilder verbessern können. Die Ergebnisse der zweiten Studie zeigten eine deutliche hormonelle Reaktion auf den Stressor, die sich in höheren Cortisol- und Alpha-Amylasespiegeln der Stressgruppe nach dem Stressor äußerte. In dieser Studie zeigten sowohl männliche, als auch weibliche Probanden zudem eine affektive Stressreaktion. Zusätzlich zu dieser hormonellen und affektiven Reaktion zeigte sich ein positiver Effekt des Stressors, der nach der Enkodierung eingesetzt wurde, auf die Gedächtnisleistung für neutrale Bilder. Bei einer nach den Geschlechtern getrennten Analyse zeigte sich dieser gedächtnisfördernde Effekt des Stressors nur bei männlichen Probanden. Das Gedächtnis für positive und negative Bilder wurde nicht durch den Stressor beeinflusst. Zusätzlich zeigte sich ein positiver korrelativer Zusammenhang zwischen der Cortisolantwort und der Gedächtnisleistung für neutrale Items, der sich für beide Geschlechter finden lies, sowie ein Trend für einen positiven Zusammenhang zwischen der Cortisolantwort und der Gedächtnisleistung für negative Bilder in der Gesamtstichprobe. Die Ergebnisse der zweiten Studie deuten darauf hin, dass Stress nach der Enkodierung das Gedächtnis für neutrale Items verbessern kann, ohne einen signifikanten Einfluss auf negative und positive Bilder zu haben. In der dritten Studie schließlich wurden nicht die Zusammenhänge zwischen Stress und Gedächtnis untersucht, sondern zwei unterschiedliche naturalistische Stressoren verglichen, um Aussagen darüber treffen zu können, welche Faktoren außerhalb des Laborsettings zu einer Stressreaktion führen. Dazu wurden die Cortisolreaktionen auf eine schriftliche Klausur sowie eine mündliche Referatssituation bei männlichen und weiblichen Studierenden untersucht. Es zeigte sich für die schriftliche Prüfung, dass die Cortisolspiegel einen Tag vor der Prüfung erhöht waren, dies allerdings lediglich bei Probanden, die den Kontrolltag nach der Prüfung hatten. Am Prüfungstag selber waren ebenfalls im Vergleich zum Kontrolltag erhöhte Cortisolspiegel zu beobachten, wobei diese lediglich vor der Prüfung, nicht aber nach der Prüfung signifikant erhöht waren. Bei der mündlichen Referatssituation zeigten sich deutlich erhöhte Cortisolkonzentrationen vor dem Referat, die während der Referatssituation weiter anstiegen. Aus dieser Studie lässt sich ableiten, dass beide beobachteten Situationen signi- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 89 fikante Stresssituationen im Leben der Studierenden darstellen, die mündlichen Referate aber zu einer deutlicheren Cortisolausschüttung führen, als die schriftlichen Klausuren. 6.2 Diskussion der Ergebnisse 6.2.1 Die hormonelle Stressreaktion In der zweiten Studie wurde als psychosozialer Stressor der Trier Sozial Stress Test (TSST; Kirschbaum et al. 1993) eingesetzt. Es zeigte sich, dass dieser Stressor zu einer hormonellen Stressreaktion führte, die sich in erhöhten Cortisol und Alpha-Amylasewerten in der Stressgruppe nach dem Stressor äußerte, was die Validität des TSST und die Befunde verschiedener Stressstudien (Dickerson und Kemeny, 2004; Kudielka und Kirschbaum, 2003; Nater, La Marca, Florin, Moses, Langhans, Koller, Ehlert, 2006; Rohleder, Nater, Wolf, Ehlert, Kirschbaum, 2004) unterstützt. Die Befunde der zweiten Studie zeigen somit, dass der TSST erfolgreich eine hormonelle Stressreaktion in der Stressgruppe erzeugte und werden daher durch die aktuelle Forschungslage zum Einfluss von Stress auf die hormonelle Stressreaktion gestützt. In der dritten Studie wurde die Cortisolreaktion auf zwei verschiedene naturalistische Stressoren untersucht. Dabei zeigte sich für die schriftliche Klausur eine antizipatorische Stressreaktion, die sich in erhöhten Cortisolspiegeln am Tag vor der Prüfung äußerte. Zudem wurden erhöhte Cortisolspiegel am Prüfungstag selber gefunden, hier allerdings nur vor der Prüfung nicht aber danach. Für das mündliche Referat zeigte sich eine deutliche Stressreaktion mit erhöhten Cortisolspiegeln vor und nach dem Referat, wobei die Cortisolausschüttung während des Referats weiter anstieg. Die dritte Studie konnte somit zeigen, dass naturalistische Stressoren im universitären Prüfungskontext in der Lage sind hormonelle Stressreaktionen auszulösen. Dieser Befund deckt sich mit Befunden anderer Studien aus diesem Bereich, die erhöhte Cortisolausschüttungen in Reaktion auf Prüfungssituationen zeigen konnten (Al-Ayadhi, 2005; Frankenhaeuser et al., 1978; Lindahl, Theorell, Lindblad, 2005; Lovallo et al., 1986; Schoofs et al., 2008). Nichtsdestotrotz herrscht im Rahmen der naturalistischen Prüfungsstressoren kein einheitliches Befundbild, da auch Studien existieren, die keinen Effekt von Prüfungen auf die Cortisolantwort (Frankenhaeuser et al., 1978; Malarkey, Pearl, Demers, Kiecolt-Glaser, Glaser, 1995; Spangler, 1997) oder sogar eine reduzierte Cortisolauschüttung (Loft et al., 2007) berichten. Als Ursache für diese heterogene Befundlage kann die Unterschiedlichkeit der untersuchten Prüfungssituationen angenommen werden (Stowell, 2003). Bisherige Studien untersuchten entweder mündliche (Herbert et al., 1986; Schoofs et al., 2008) oder schriftliche (Frankenhaeuser et al., 1978; Lindahl et al., 2005) Prüfungen, erhoben die Stressreaktion zu einem Prüfungszeitpunkt (Schoofs et al., 2008; Spangler, 1997) Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 90 oder während einer längeren Prüfungsperiode (Loft et al., 2007; Weekes, Lewis, Patel, Garrison-Jakel, Berger, Lupien, 2006) oder berichten gar nicht, um welche Art Prüfung es sich handelte (Al-Ayadhi, 2005; Lovallo et al., 1986; Malarkey et al., 1995). Eine Vergleichbarkeit der Studien ist daher schwierig. Dennoch lässt sich im Bereich der Prüfungsstudien festhalten, dass die Befundlage für mündliche Prüfungen ein einheitlicheres Bild mit erhöhten Cortisolspiegeln zeigt, als schriftliche Prüfungen (Stowell, 2003). Betrachtet man mündliche und schriftliche Leistungssituationen im universitären Kontext, so wird deutlich, dass sich diese Situationen durch verschiedene Faktoren unterscheiden. Während im laborexperimentellen Kontext die stressauslösenden Faktoren für Laborstressoren bekannt sind, wurden diese bei naturalistischen Prüfungssituationen bisher nicht untersucht. Einen starken Einfluss auf die Höhe der Cortisolausschüttung im Labor übt der Faktor der Bedrohung des sozialen Selbst aus (Dickerson und Kemeny, 2004). Aus den Befunden der dritten Studie dieser Arbeit lässt sich der Schluss ziehen, dass dieser Faktor auch im naturalistischen Stressbereich eine große Bedeutung hat. Während die betrachtete schriftliche Prüfung in einem großen Hörsaal stattfindet, in dem die Studierenden die Arbeit alleine und ohne direktes Feedback lösen, ist die Situation bei einem Referat sehr viel weniger anonym. Hier stellen die Studierenden ihr Können und Wissen im Beisein ihrer Kommilitonen und Dozenten dar und erhalten direkten verbales und nonverbales Feedback. Die Bedrohung des sozialen Selbst ist also bei Referaten deutlich stärker ausgeprägt, als bei schriftlichen Prüfungen, wohingegen der Leistungsdruck vergleichbar ist. Da sich in der dritten Studie für die schriftliche Prüfung eine moderate, für das Referat jedoch eine deutliche und starke Stressreaktion zeigte, scheint auch bei naturalistischen Stressoren die Bedrohung des sozialen Selbst eine zentrale Rolle einzunehmen. Diese Interpretation ist natürlich mit einer gewissen Vorsicht zu betrachten, da sich schriftliche und mündliche Leistungssituationen an der Universität durch mehrere Faktoren unterscheiden, wobei die Bedrohung des sozialen Selbst nur einer davon ist. Neben diesen Unterschieden stellt sich auch die Frage, welchen Einfluss die individuelle Vorbereitung und Ängstlichkeit der Probanden auf die Stressantwort hatten und ob die Stärke der Stressreaktion mit den tatsächlichen Leistungen zusammenhing, da diese Faktoren nicht erhoben wurden. Somit bleiben einge Fragen unbeantwortet und benötigen weitere Forschungsbemühungen. Die Ergebnisse der dritten Studie zeigen zusätzlich weitere interessante Aspekte der hormonellen Stressreaktion bei naturalistischen Stressoren. Zum einen zeigte sich am Tag vor der schriftlichen Klausur eine antizipatorische Stressreaktion, die sich in erhöhten Cortisolspiegeln äußerte. Daraus und aus weiteren Befunden zu antizipatorischen Stressreaktionen (Rohleder et al., 2007) lässt sich schlussfolgern, dass im naturalistischen Setting angekündigte und bekannte Stresssituationen nicht nur in der akuten Stressphase, sondern bereits vorher zu einer verstärkten Aktivität der HHNA führen. Darüber hinaus beobachtete die dritte Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 91 Studie diese antizipatorische Stressreaktion nur bei Probanden, die den Kontrolltag nach der Prüfung hatten. Daraus wird deutlich, dass nicht nur am Tag des Stressors selber, sondern bereits mehrere Tage vorher die Aktivität der HHNA stärker zu sein scheint, als an Tagen ohne kommende Prüfung. Daraus lässt sich ableiten, dass die HHNA während längeren Prüfungsphasen und bei einzelnen Prüfungen bereits mehrere Tage vorher eine verstärkte Aktivität zeigt (Loft et al., 2007; Weekes et al., 2006). Die Tatsache, dass die Probanden nicht randomisiert den Kontrollbedingungen zugeordnet wurden, sondern frei wählen konnten, ob sie vor oder nach der Klausur an der Kontrollsituation teilnehmen wollten, stellt allerdings eine Schwäche der Studie dar. Somit kann nicht ausgeschlossen werden, dass weitere Faktoren, wie z.B. die Vorbereitung der Probanden oder eine geringe allgemeine Ängstlichkeit die Effekte beeinflusst haben. Zudem wurden weitere Klausuren um den Zeitpunkt der erfassten Klausur und weitere Referate um den Zeitpunkt des erfassten Referats nicht erhoben, so dass die Möglichkeit einer chronischen Stressreaktion bei einigen Probanden nicht ausgeschlossen werden kann. Neben dem beobachteten Einfluss der Reihenfolge des Kontrolltags zeigten sich in der der dritten Studie zusätzlich noch Einflüsse der Jahreszeit sowie der Aufwachzeit der Probanden. Während die beobachteten und zu weiteren Befunden (King, Rosal, Ma, Reed, Kelly, Stanek, III, Ockene, 2000; Walker, Best, Noon, Watt, Webb, 1997) passenden höheren Cortisolspiegel in den Wintermonaten den Einfluss der Jahreszeit verdeutlichen, zeigen die höheren Cortisolspiegel von Probanden, die später aufgestanden sind die Bedeutung inter- und intraindividueller Unterschiede, die häufig bei Betrachtungen der Stressreaktion nicht kontrolliert werden. 6.2.1.1 Geschlechtsunterschiede Bei den durchgeführten Stressstudien zeigten sich sowohl für den Laborstressor, als auch für den naturalistischen Stressor Geschlechtsunterschiede. Diese Stressreaktion auf den TSST war bei Männern ausgeprägter, als bei Frauen. Dieser Befund spiegelt die Ergebnisse anderer Studien wieder, die ebenfalls zeigen, dass Frauen weniger stark auf psychosoziale Laborstressoren reagieren, als Männer (Kirschbaum et al., 1992; Stroud et al., 2002). Während die Geschlechtsunterschiede bei Laborstressoren deutlich und homogen sind, zeigt sich bei den in der dritten Studie erfassten Prüfungssituationen ein weniger deutliches und gegenläufiges Bild. Für die Referatssituation zeigten sich wie auch beim TSST höhere Cortisollevel bei Männern, als bei Frauen, wohingegen Frauen bei der schriftlichen Prüfung höhere Cortisolspiegel zeigten, als Männer. Die männlichen Stichproben in der dritten Studie waren allerdings sehr klein (4 Männer bei der schriftlichen Klausur und 9 Männer bei dem Referat), so dass Aussagen über Geschlechtsunterschiede nur mit großer Vorsicht getroffen Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 92 werden können. Ein weiterer Einflussfaktor auf die hormonelle Stressreaktion zeigt sich in der Zusammensetzung der weiblichen Stichprobe. Hier wurden Frauen mit und ohne hormonelle Verhütung eingeschlossen. Aus Laborstudien ist bekannt, dass hormonelle Kontrazeptiva die Stressantwort modulieren, was durch die vermehrte Produktion des Cortisol Bindungsglobulin vermittelt wird (Kirschbaum et al., 1999). In der dritten Studie hingegen zeigte sich jedoch keine Einfluss von oralen Kontrazeptiva, was mit Befunden anderer Prüfungsstudien übereinstimmt (Schoofs et al., 2008). Es scheint also, dass hormonelle Kontrazeptiva zwar die Stressantwort auf akute und überraschende Stresssituationen im Labor beeinflussen, nicht aber die hormonelle Stressreaktion auf bekante und mit einer antizipatorischen Stressantwort einhergehenden naturalistische Stressoren. Insgesamt lässt sich in Bezug auf die hormonelle Stressreaktion festhalten, dass sowohl der TSST als Laborstressor, als auch Leistungssituationen im universitären Kontext valide Stressoren zur Initiierung der HHNA darstellen. Für den Bereich der naturalistischen Stressoren konnten verschiedene Faktoren identifiziert werden, welche die Stressreaktion modulierend beeinflussen. Im Bereich des Laborstresses konnten die bekannten und in der hier durchgeführten Laborstudie betrachteten Einflussfaktoren bestätigt werden. Abbildung 4 zeigt eine Zusammenfassung derjenigen Einflussfaktoren, die im Rahmen dieser Promotionsarbeit betrachtet wurden. Abbildung 4: Einflussfaktoren auf die Stressantwort in Reaktion auf naturalistische Stressoren und Laborstressoren. Für Laborstressoren zeigte sich ein Einfluss des Geschlechts, hormoneller Verhütung sowie der Bedrohung des sozialen Selbst. Die Bedrohung des sozialen Selbst sowie das Geschlecht beeinflussen ebenfalls die Reaktion auf naturalistischen Stressor. Zudem zeigte sich ein Einfluss von Aufwachzeit und Jahreszeit. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 6.2.2 93 Die Zusammenhänge zwischen Cortisol, Stress und freier Abrufaufgabe Die erste Studie wurde durchgeführt, um mögliche Zusammenhänge zwischen basalen Cortisolspiegeln und dem Gedächtnis für emotional neutrale und emotional erregende Bilder aufzudecken. Zusätzlich wurde der Einfluss der Enkodierungsstrategie erhoben. In der zweiten Studie wurde darauf aufbauend der Einfluss eines psychosozialen Stressors auf die Gedächtniskonsolidierung dieser Stimuli untersucht. Der in der ersten Studie gefundene positive Zusammenhang zwischen der Höhe der Cortisolspiegel und der Gedächtnisleistung für emotional erregende Stimuli wird durch Studien unterstützt, die den Effekt von Cortisol oder Stress auf die Gedächtnisbildung untersucht haben. Dabei zeigten sich positive Effekte eines Stressors sowohl auf die Enkodierung emotional erregender Items (Jackson, Payne, Nadel, Jacobs, 2006; Jelicic et al., 2004; Payne et al., 2007; Schwabe et al., 2008) als auch auf die Konsolidierung (Andreano und Cahill, 2006; Beckner et al., 2006; Cahill et al., 2003; Smeets et al., 2006). Auch pharmakologische Studien zeigten positive Effekte auf die Gedächtnisbildung für emotional erregende Stimuli (Buchanan und Lovallo, 2001; Kuhlmann und Wolf, 2006b). Die in der ersten Studie berichteten positiven Zusammenhänge zwischen der Höhe basaler Cortisolspiegel und dem Gedächtnis für emotional erregende Bilder unterstützen diese Befunde. Zudem weiten sie die Befunde zu erhöhten Cortisolwerten und Stress aus, indem sie die Bedeutung von basalen Cortisolspiegeln für das emotionale Gedächtnis zeigen. Wie in verschiedenen Stress- (Abercrombie et al., 2006; Cahill et al., 2003; Jelicic et al., 2004; Payne et al., 2007; Payne et al., 2006; Schwabe et al., 2008; Smeets et al., 2008) und pharmakologischen Studien (Buchanan und Lovallo, 2001; Kuhlmann und Wolf, 2006b) berichtet wurde, wurde auch in der ersten Studie ein stärkerer Effekt für emotional erregende, als für neutrale Stimuli beobachtet. Als verantwortlich für diese stärkeren Effekte auf emotional erregendes Material wird eine Interaktion zwischen Cortisol und Amygdalaaktivität angenommen (Roozendaal, Okuda, Van der Zee, McGaugh, 2006b). Für die gedächtnisfördernden Effekte der Emotionalität der Stimuli ist hierbei nicht die Valenz, sondern die ausgelöste Erregung bestimmend. Während die Valenz der Stimuli präfrontal verarbeitet wird, geht mit der Verarbeitung emotional erregender Stimuli eine Aktivierung der Amygdala einher (Kensinger und Corkin, 2004; LaBar und Cabeza, 2006). Die noradrenerge Aktivierung der Amygdala übt eine modulierende Wirkung auf die Aktivität des Hippocampus aus, so dass eine Interaktion dieser beiden Hirnstrukturen für die stärkeren Effekte von Cortisol und Stress auf emotional erregende Items angenommen wird (Roozendaal et al., 2006b). Diese Interaktion scheint den Befunden der ersten Studie folgend nicht nur bei erhöhten Cortisolspiegeln, sondern auch bei basalen Cortisolspiegeln bedeutsam zu sein. Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 94 Die zweite Studie wurde durchgeführt, um den Einfluss erhöhter Cortislspiegel auf die Gedächtniskonsolidierung zu untersuchen. Es zeigte sich für die Stressgruppe ein verbessertes Gedächtnis für neutrale Items. Für positive und negative Bilder zeigte sich kein Effekt. Zusätzlich wurden positive Korrelationen zwischen der Cortisolreaktion auf den Stressor und der Gedächtnisleistung für neutrale Bilder berichtet. Zudem zeigte sich ein Trend für einen positiven Zusammenhang zwischen der Cortisolantwort und der Gedächtnisleistung für negative Bilder. Der positive Effekt des Stressors auf die Gedächtniskonsolidierung spiegelt Befunde aus human- und tierexperimentellen Stressstudien wieder. Sowohl im Human(Andreano und Cahill, 2006; Beckner et al., 2006; Cahill et al., 2003; Smeets et al., 2008), als auch im Tierbereich (Akirav et al., 2004; Akirav, Sandi, Richter-Levin, 2001; Roozendaal, de Quervain, Ferry, Setlow, McGaugh, 2001; Roozendaal et al., 1999; Sandi et al., 1997) zeigten sich positive Effekte von psychosozialem Stress, der nach der Enkodierung stattfand auf die Gedächtnisleistung. Allerdings zeigten humanexperimentelle Stressstudien (Abercrombie et al., 2006; Smeets et al., 2008; Cahill et al., 2003) einen stärkeren Effekt auf emotional erregende Items, als auf neutrale Items. Die Befunde der zweiten Studien werden somit nicht durch diese Studien gestützt. Allerdings berichten einige Studien auch Effekte auf sowohl emotional erregende, als auch neutrale Items (Abercrombie et al., 2003) sowie positive Effekt auf die Konsolidierung von neutralen Items in Designs, in denen keine emotional erregenden Stimuli verwendet wurden (Andreano und Cahill, 2006; Beckner et al., 2006). In der zweiten Studie fand sich lediglich ein Trend für einen positiven korrelativen Zusammenhang zwischen der Cortisolantwort auf den Stressor und der Gedächtnisleistung für negative Bilder, der jedoch nicht signifikant wurde. Somit zeigten sich in den beiden durchgeführten Gedächtnisstudien unterschiedliche Ergebnisse. Während in Studie eins ein positiver Zusammenhang zwischen den basalen Cortisolspiegeln und der Gedächtnisleistung beobachtet wurde, zeigte sich in Studie zwei ein positiver Einfluss des Stressors und erhöhter Cortisolspiegel auf die Gedächtnisleistung für neutrale Items. Diese auf den ersten Blick gegenläufigen Ergebnisse lassen jedoch bei genauerer Betrachtung auf einen wichtigen modulierenden Faktor, nämlich die Interaktion zwischen Erregung durch die Stimuli und die Erregung durch die Testsituation, schließen. Die Zusammenhänge zwischen basalen Cortisolspiegeln und der Gedächtnisleistung für emotional erregende Bilder wurden in der ersten Studie lediglich in der Gruppe der Probanden beobachtet, die eine explizite Enkodierungsinstruktion erhalten hatten. In der Gruppe der Probanden, die die Bilder enkodierten, ohne über den folgenden Gedächtnistest informiert zu werden, zeigte sich kein Zusammenhang. Aus diesem Befund kann abgeleitet werden, dass basale Cortisolspiegel nur dann einen positiven Effekt auf die Gedächtnisbildung ausüben, wenn die Stimuli bewusst und zielgerichtet enkodiert werden. Dieses zielgerichtete und be- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 95 wusste Lernen der Stimuli ist mit einer hohen Motivation und daher mit verstärkter Aufmerksamkeit verbunden, die auf das zu lernende Material gerichtet ist. Von dieser gerichteten Aufmerksamkeit kann angenommen werden, dass sie die Verarbeitung der Stimuli vertieft. Die verstärkte Aufmerksamkeit bei der Betrachtung der Bilder könnte zudem eine zusätzlich verstärkte Aktivität der Amygdala bei der Verarbeitung dieser Bilder verursacht haben. Es ist deshalb möglich, dass die durch die Aufmerksamkeit erhöhte Amygdalaaktivität die Interaktion zwischen Cortisol und noradrenerger Aktivität erleichterte und somit die positiven Effekte von basalen Cortisolspiegeln auf die emotional erregenden Stimuli bei einer expliziten Enkodierung ermöglichte. Da mit der Verarbeitung der emotional neutralen Reize keine Amygdalaaktivierung einherging, stellte sich in keiner der beiden Enkodierungsgruppen ein positiver Effekt von Cortisol auf die Gedächtnisleistung für diese Items ein. Die neutralen Items konnten somit nicht von der verstärkten Aufmerksamkeit, die durch eine explizite Enkodierungsinstruktion ausgelöst wurde profitieren. Zusätzlich zu einer verstärkten Amygdalaaktivität aufgrund einer erhöhten Aufmerksamkeit, kann zudem angenommen werden, dass das Wissen an einem Gedächtnistest teilzunehmen die Erregung der Probanden ebenfalls verstärkte. Eine gesteigerte Erregung durch die Testsituation lässt sich als weitere Ursache für die Zusammenhänge zwischen basalen Cortisolspiegeln und der Gedächtnisleistung für emotional erregende Stimuli, die nur bei der expliziten Enkodierung zu beobachten waren, annehmen. Die Bedeutung der Testsituation für die Effekte von Cortisol auf das Gedächtnis wurde bereits von Kuhlmann und Wolf (2006a) demonstriert. Diese Studie zeigt, dass eine nicht erregende Testsituation die Effekte einer pharmakologischen Cortisolbehandlung auf den Gedächtnisabruf aufheben kann. Die Interaktion zwischen Cortisol und Amygdalaaktivität bei der Verarbeitung emotional erregender Bilder scheint demnach durch die Erregung, die durch die Testsituation ausgelöst wurde, verstärkt worden zu sein. Die Befunde der ersten Studie zeigen somit eine Bedeutung von Aufmerksamkeit und durch die Testsituation erzeugte Erregung für den Zusammenhang zwischen basalen Cortisolspiegeln und dem Gedächtnis für emotional erregendes Material. Die Ergebnisse der zweiten Studie weiten die in Studie eins beobachtete Bedeutung der durch die Testsituation erzeugten Erregung auf einen umgekehrt U-förmigen Zusammenhang zwischen Cortisol und Erregung aus. In der ersten Studie zeigten sich positive Zusammenhänge zwischen Cortisol und der Gedächtnisleistung für emotional erregende Bilder. Diese Zusammenhänge wurden nur für die explizite Enkodierungsstrategie beobachtet, so dass die Vermutung naheliegt, dass Cortisol und die durch die Testsituation erzeugte Erregung und Aufmerksamkeit das Gedächtnis für emotional erregende Items beeinflussen. In der zweiten Studie wurde ebenfalls eine explizite Enkodierungsstrategie verwendet, so dass die durch die Lerninstruktion erzeugte Aufmerksamkeit ähnlich hoch war, wie in der ersten Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 96 Studie. Im Gegensatz zur ersten Studie wurde in der zweiten Studie jedoch ein psychosozialer Stressor eingesetzt, der die Cortisolspiegel signifikant erhöhte und auch die durch die Testsituation erzeugte Erregung durch das Wissen über die mögliche Teilnahme an einer stressvollen Situation teilzunehmen, verstärkte. Während in der ersten Studie, in der die Testsituation weniger erregend und die Cortisolspiegel nicht erhöht waren positive Zusammenhänge zwischen Cortisol und dem Gedächtnis für negative Bilder beobachtet wurden, zeigten sich in der zweiten Studie, in der die Testsituation eine stärkere Erregung auslöste und die Cortisolspiegel durch den psychosozialen Stressor erhöht waren, positive Effekte auf neutrale Bilder. Eine umgekehrt U-förmige Beziehung zwischen Cortisol und Erregung für die Effekte auf das Gedächtnis ist diesen Befunden nach denkbar. Der Effekt auf emotional erregende und neutrale Stimuli scheint von der ausgelösten Erregung der Testsituation abzuhängen. Die Befunde der ersten und zweiten Studie implizieren, dass es ein optimales Erregungsniveau von Gedächtnisstimuli für Stresseffekte zu geben scheint. Cortisoleffekte auf das Gedächtnis hängen von einem optimalen Erregungsniveau ab, das sich aus der Erregung, die durch die Stimuli ausgelöst wird und der Erregung, die durch die Situation ausgelöst wird, zusammensetzt. Während in Situationen mit geringer extern ausgelöster Erregung, wie in Experimenten ohne Stressor, in pharmakologischen Studien (Buchanan und Lovallo, 2001; Kuhlmann und Wolf, 2006b) oder schwachen physischen Stressoren emotional erregende Items das optimale Erregungsniveau für Cortisoleffekte haben, ist die ausgelöste Erregung dieser Items in Situationen mit starker extern ausgelöster Erregung zu hoch, um für Cortisoleffekte sensitiv zu sein. In diesen Situationen, wie zum Beispiel in Experimenten, in denen starke psychosoziale Stressoren eingesetzt werden, scheinen vielmehr neutrale Items das optimale Erregungsniveau zu besitzen. Die Befunde der zweiten Studie deuten demnach darauf hin, dass Stresseffekte auf die Gedächtnisleistung von der emotionalen Erregung, die durch die Items und der emotionalen Erregung, die durch die Testsituation ausgelöst wird, abhängen. Die Ergebnisse aus Studie zwei zeigen zusätzlich zu einer möglichen umgekehrt U-förmigen Beziehung einen möglichen Einfluss der verwendeten Stimuli. Betrachtet man die Gedächtnisleistung der Probanden für emotional erregende und emotional neutrale Bilder, so zeigte sich im direkten Abruftest ein besseres Gedächtnis für emotional erregende, als für neutrale Bilder. Dieser „emotional enhancement effect“ (LaBar und Cabeza, 2006) zeigte sich nicht mehr in der Gedächtnisleistung im verzögerten Abruftest. Der Stressor scheint demnach die Gedächtnisbildung für neutrale Bilder verbessert, emotional erregende Bilder aber nicht beeinflusst zu haben. Dabei ist denkbar, dass die emotional erregenden Bilder durch ihre emotionale Erregung besser und tiefer enkodiert wurden und sich damit auch ein verbessertes Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 97 Gedächtnis gegenüber den neutralen Bildern im direkten Abruftest zeigte. Es ist denkbar, dass sich Stress in der zweiten Studie nur auf das Gedächtnis für diejenigen Bilder auswirkte, die weniger gut gelernt wurden und für die eine Gedächtnisverbesserung möglich war. Das Gedächtnis für emotional erregende Bilder hingegen war bereits so gut, dass Stress keine positiven Effekte mehr ausüben konnte. Möglicherweise hätten sich deutlichere und auch signifikante Effekte für emotional erregende Bilder bei einer schwierigeren Aufgabe gezeigt, bei der die Enkodierung der Items nicht unter Bedingungen geschieht, die eine tiefe und gute Verarbeitung der einzelnen Items erlauben und die damit sensitiver für Stresseffekte ist. Durch die gemeinsame Betrachtung der Ergebnisse der beiden durchgeführten Gedächtnisstudien zeigen sich wichtige Einflussfaktoren auf den Zusammenhang zwischen Stress, Cortisol und der Gedächtnisleistung. Studie eins macht deutlich, dass für den Zusammenhang zwischen basalen Cortisolspiegeln und der Gedächtnisleistung die durch die Testsituation induzierte Erregung sowie die durch die Enkodierungsinstruktion verstärkte Aufmerksamkeit auf die Gedächtnisstimuli einen bedeutsamen Einfluss auf die Zusammenhänge zwischen basalen Cortisolspiegeln und der Gedächtnisleistung haben. Aus dieser Studie wurde die Verwendung einer expliziten Enkodierungsstrategie für die nachfolgende Studie abgeleitet. Die Ergebnisse aus Studie zwei deuten unter Berücksichtigung der Ergebnisse der ersten Studie darauf hin, dass für Stresseffekte auf das Gedächtnis nicht nur die Güte des Erlernens der Stimuli von Bedeutung ist, sondern auch die Interaktion zwischen der durch die Gedächtnisstimuli und der durch die Testssituation ausgelösten Erregung. Mit diesem Befund weitet die zweite Studie die Befunde zum Einfluss der Testsituation aus Studie eins auf eine umgekehrt U-förmige Beziehung zwischen Cortisol und emotionaler Erregung aus. Insgesamt verdeutlichen die Befunde der durchgeführten Studien, dass die Effekte von Cortisol auf das Gedächtnis stark von der emotionalen Erregung der Stimuli abhängen. Allerdings zeigen die Ergebnisse der Studien auch, dass die Zusammenhänge und Einflüsse von Stress auf das emotionale Gedächtnis sehr komplex sind, so dass weitere Studien notwendig sind, um die hier beschriebenen Einflussfaktoren weiter zu untersuchen. 6.2.3 Die Cortisoleffekte auf hippocampusunabhängige Gedächtnisleistungen In den beiden durchgeführten Gedächtnisstudien wurde neben den freien Abrufaufgaben auch eine Multiple-Choice Aufgabe verwendet, welche am zweiten Tag der Testung durchgeführt wurde. In keiner der Studien fanden sich Effekte des Stressors oder Zusammenhänge mit Cortisol auf die Leistung der Probanden in dieser Aufgabe. Dieser Befund wird sowohl Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 98 durch pharmakologische, als auch durch Stressstudien gestützt, die ebenfalls keine Stressoder Cortisoleffekte auf diese Art Aufgabe finden (de Quervain, Henke, Aerni, Treyer, McGaugh, Berthold, Nitsch, Buck, Roozendaal, Hock, 2003; Kuhlmann et al., 2005; Kuhlmann und Wolf, 2006b). Als Ursache für ausbleibende Effekte auf die Gedächtnisleistung für diesen Aufgabentyp wird angenommen, dass es sich hierbei nicht um einen freien, sondern um eine Rekognitionsaufgabe handelt, bei der die Probanden aus vier Möglichkeiten die richtige auswählen. Während der freie Abruf von Gedächtnisinhalten den Hippocampus involviert, findet Wiedererkennen eher im perirhinalen Cortex statt (Aggleton und Brown, 2006). Da der Hippocampus eine Gehirnstruktur darstellt, in der Cortisolrezeptoren vermehrt zu finden sind, ist auch zu erwarten, dass sich Stress- oder Cortisoleffekte vornehmlich auf Aufgaben zeigen, die mit dieser Struktur assoziiert sind. Allerdings existieren auch Studien, welche Stresseffekte auf die Leistung der Probanden in Wiedererkennungsaufgaben berichten. In diesen Studien wurden allerdings keine weiteren Gedächtnisaufgaben eingesetzt (Beckner et al., 2006; Monk und Nelson, 2002). Daher könnte es möglich sein, dass Stresseffekte auf diese Aufgabenart dann zu finden sind, wenn die relevanten Gedächtnisinhalte vorher nicht durch eine freie Abrufaufgabe aktiviert wurden. Zukünftige Studien sollten daher die Effekte von Stress und Cortisol auf verschiedene Aufgabentypen näher untersuchen. 6.2.4 Der Einfluss des Geschlechts auf die Gedächtnisleistung In der zweiten Studie wurde zusätzlich zu allgemeinen Stress- und Cortisoleffekten auf das Gedächtnis der Einfluss des Geschlechts der Versuchspersonen auf diese Effekte untersucht. Verschiedene Studien zeigten stärkere Effekte von Stress und Cortisol auf das Gedächtnis bei Männern, als bei Frauen (Andreano und Cahill, 2006; Cahill, 2003; Wolf et al., 2001). Dabei hat sich der hormonelle Zyklus bei Frauen als potentieller Einflussfaktor auf die Zusammenhänge zwischen Cortisol und Gedächtnisleistung herausgestellt. Allerdings ist die Forschungslage bisher heterogen. So zeigen einige Studien einen klaren Zusammenhang zwischen Stress und Gedächtnisleistung bei Frauen nur in der Lutealphase (Andreano et al., 2008), wohingegen andere Studien in dieser Phase keine Zusammenhänge berichten (Wolf et al., 2001) oder Stresseffekte auf das Gedächtnis bei weiblichen Stichproben finden, deren Zyklusphase nicht kontrolliert wurde (Smeets et al., 2008). In den beiden durchgeführten Gedächtnisstudien wurden Frauen untersucht, deren Zyklusphase nicht kontrolliert wurde. Dennoch zeigten sich in der ersten Studie Zusammenhänge zwischen basalen Cortisolspiegeln und der Gedächtnisleistung. In der zweiten Studie wurde für diese Zusammenhänge ebenfalls kein Unterschied zwischen Männern und Frauen beobachtet, wohingegen in der zweiten Studie ein Stresseffekt nur für Männer berichtet wurde. Allerdings zeigten sich hier Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 99 die Zusammenhänge zwischen dem Cortisolanstieg und der Gedächtnisleistung für neutrale Items ebenfalls für beide Geschlechter. Ein Einfluss der Zyklusphase kann aus den Ergebnissen der durchgeführten Studien weder eindeutig bestätigt, noch ausgeschlossen werden. Die Bedeutung der Zyklusphase für Stresseffekte auf das Gedächtnis bleibt daher weiterhin unklar, so dass weitere Forschungsbemühungen zur Klärung dieser Fragestellung notwendig sind. 6.3 Kritische Betrachtung der Studien Bei einer kritischen Betrachtung der durchgeführten Studien offenbaren sich einige Schwächen. Als erster Punkt ist die Stichprobengröße anzumerken. In Studie eins wurden 60 Frauen, in Studie zwei 30 Männer und 28 Frauen und in Studie drei 35 Probanden für die schriftliche Klausur sowie 34 Probanden für das Referat getestet. Für die dritte Studie wäre es für die Vergleichbarkeit und zur Steigerung der Aussagekraft vorteilhaft gewesen die Effekte einer schriftlichen Klausur und einer eines Referats innerhalb einer Stichprobe zu untersuchen. Dies war jedoch aus praktischen Gründen nicht möglich. Für die Gedächtnisstudien stellt sich aufgrund der nichtsignifikanten Ergebnisse, vor allem in Studie zwei die Frage, ob eine größere Stichprobe und eine damit einhergehende größere Power der Studien eventuell zusätzliche Effekte sichtbar gemacht hätten. Allerdings konnten vorherige Studien mit einer ähnlichen Stichprobengröße Stresseffekte auf das Gedächtnis demonstrieren (Payne et al., 2007; Smeets et al., 2006), weshalb die Power der durchgeführten Studien als ausreichend anzunehmen ist und sich die berichteten Effekte ebenso wie die nicht signifikanten Effekte nicht allein auf die Stichprobengröße zurückführen lassen. Nichtsdestotrotz stellen die Charakteristika der Stichproben eine potenzielle Schwierigkeit dar. Wie bereits in der Diskussion dargestellt, ist das Geschlecht ein wichtiger Einflussfaktor zum einen für die Hormonreaktion selber, wobei Frauen häufig mit einer schwächeren hormonellen Stressreaktion auf psychosoziale Laborstressoren reagieren (Kirschbaum et al., 1999; Kirschbaum et al., 1992; Stroud et al., 2002). Zum anderen werden Stresseffekte auf das Gedächtnis ebenfalls vom Geschlecht der Versuchsperson beeinflusst, wobei Männer häufig stärkere Stresseffekte zeigen, als Frauen (Andreano und Cahill, 2006; Cahill, 2003; Wolf et al., 2001). Studie eins sowie die weiblichen Stichproben in den Studien zwei und drei bestanden aus Frauen, deren Zyklusphase nicht kontrolliert wurde. Da die Zyklusphase ein potentieller Einflussfaktor für die Effekte bei Frauen darstellt (Andreano et al., 2008), ist es denkbar, dass dies zu einer Maskierung von Effekten in der weiblichen Stichprobe geführt hat. Gegen diese Annahme spricht allerdings, dass auch Studien existieren, die Stresseffekte in weiblichen Stichproben berichten, deren Zyklusphase nicht kontrolliert wurde (Smeets et al., 2008). Die Ergebnisse Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 100 der korrelativen Analysen in den Studien eins und zwei zeigen zudem signifikante korrelative Zusammenhänge zwischen Cortisol und der Gedächtnisleistung auch bei den weiblichen Stichproben, was gegen die Annahme spricht, dass Cortisoleffekte auf das Gedächtnis lediglich in der Lutealphase zu beobachten sind (Andreano et al., 2008). Nichtsdestotrotz kann ein Einfluss der Zyklusphase auf die Ergebnisse nicht ausgeschlossen werden. Eine weitere Schwäche der Studien zeigt sich bei der Betrachtung der Gedächtnisleistung. Die Probanden der Studie zeigen in Studie zwei, in denen zusätzlich zu einem direkten Abruftest auch ein verzögerter Gedächtnistest eingesetzt wurde, kein Vergessen. Die Gedächtnisleistung der Probanden für das Gedächtnismaterial nimmt von Tag 1 zu Tag 2 nicht nennenswert ab. Verschiedene Studien, die das Gedächtnis über einen längeren Zeitraum untersuchten zeigen, dass die Leistung von Probanden mit der Zeit abnimmt (Kuhlmann et al., 2005; Kuhlmann und Wolf, 2005). Da dies in den durchgeführten Studien nicht zu beobachten war, liegt die Vermutung nahe, dass das Gedächtnismaterial vergleichsweise einfach zu behalten war. Die Aufgabe bestand aus insgesamt 15 Bildern, wobei jeweils 5 Bilder emotional neutral, positiv und negativ waren. Die Bilder wurden auf einem Computerbildschirm und die Begleitsätze über Kopfhörer präsentiert. Die Präsentationsdauer betrug pro Bild 10 Sekunden. Es ist aufgrund der Ergebnisse anzunehmen, dass diese Art der Präsentation dazu führte, dass die Probanden die Bilder tief enkodiert und somit gut erlernt haben. Der Zeitabstand von 24 Stunden zum zweiten Gedächtnistest war möglicherweise nicht lang genug, um ein Verfallen der Gedächtnisspuren zu ermöglichen. Hinzu kommt, dass für die zweite Studie eine explizite Enkodierungsstrategie eingesetzt wurde, das heißt die Probanden waren sich über den folgenden Gedächtnistest bewusst. Die explizite Enkodierungsstrategie wurde auf Grund der Ergebnisse aus Studie eins verwendet, die keinen generellen Unterschied in der Gedächtnisleistung zwischen Probanden, denen der Gedächtnistest bewusst war und Probanden, denen er nicht bewusst war, zeigte, jedoch offenbarte, dass die positiven Zusammenhänge zwischen Cortisol und dem Gedächtnis für emotionale erregende Stimuli nur unter der expliziten Lerninstruktion zu finden waren. Da in dieser Studie jedoch der Zusammenhang mit basalen Cortisolspiegeln und nicht mit Stress betrachtet wurde, kann nicht ausgeschlossen werden, dass ein impliziter Gedächtnistest, bei dem die Probanden die Bilder nicht bewusst und somit eventuell schlechter lernen und der zudem die durch die Testsituation ausgelöste Erregung reduziert, Stresseffekte sichtbar gemacht hätte. Eine weitere Schwierigkeit für die Vergleichbarkeit der beiden Gedächtnisstudien ist die Zeit der Versuchsdurchführung. Aufgrund des circadianen Rhythmus von Cortisol ist der Cortiolspiegel in den Morgenstunden am höchsten und nimmt über den Tag ab, bis er am Abend seinen Tiefpunkt erreicht sind (Jacobson, 2005; Kirschbaum und Hellhammer, 1989). Bei Stressstudien ist deshalb die Tageszeit der Versuchsdurchführung von großer Bedeutung Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 101 (Het, Ramlow, Wolf, 2005; Maheu, Collicutt, Kornik, Moszkowski, Lupien, 2005). Studie zwei wurde am Nachmittag durchgeführt, die Probanden der ersten Studie wurden allerdings zu verschiedenen Tageszeiten getestet. Der Einfluss der Tageszeit wurde mit einer partiellen Korrelation kontrolliert, allerdings wäre es für die Vergleichbarkeit der Studien und zum Ausschluss eines Einflusses des circadianen Rhythmus von Vorteil gewesen, wenn die Studien zu gleichen Tageszeiten durchgeführt worden wären. Auch in Studie drei wurde der circadiane Rhythmus nicht optimal kontrolliert. Während dei Klausur immer am Morgen stattfand, fanden die Referate am Morgen, am Mittag oder am Nachmittag statt. Dies macht es schwierig die Prüfungssituationen miteinander zu vergleichen. Zwar ergaben sich keine Hinweise auf einen Einfluss der Tageszeit, jedoch kann nicht ausgeschlossen werden, dass die Unterschiede in der Stärke der Cortisolantworten auf die Klausur und die Referate zum Teil durch die unterschiedlichen Startzeiten bedingt waren. Ein Ziel der Gedächtnisstudenstudien war es, den Einfluss von Cortisol und psychosozialem Stress auf emotional neutrale und emotional erregende positive und negative Items zu untersuchen. In diesem Zusammenhang wurden in den Studien subjektive Ratingskalen eingesetzt, auf denen die Probanden die subjektiv wahrgenommene Valenz und die durch die Bilder ausgelöste Erregung beurteilten. Die Analysen dieser Ratings zeigten, dass die Probanden die theoretische Kategorisierung, wie sie von den Autoren der Aufgabe vorgegeben war (Buchanan et al., 2001; Buchanan et al., 2003), unterstützten. Diese subjektive Einschätzung durch die Probanden wurde in den Studien nicht durch ein objektives Maß der Erregung ergänzt. Der Einsatz eines objektiven Maßes wie z.B. Hautleitfähigkeit hätte Informationen zur ausgelösten Erregung bereitgestellt. Zwar zeigten Befunde, dass subjektive Erregungsmaße mit objektiven Erregungsmaßen korrelieren (Bradley, Greenwald, Petry, Lang, 1992), dennoch bieten objektive Maße eine höhere Zuverlässigkeit. 6.4 Ausblick Die im Rahmen der vorliegenden Dissertation durchgeführten Studien dienten der weiteren Erkenntnisgewinnung zu Cortisol- und Stresseffekten auf das Gedächtnis für neutrale und emotional erregende positive und negative Informationen und der Spezifizierung der Stressreaktion auf naturalistische Stresssituationen. Aus den diskutierten Schwächen und Schwierigkeiten der durchgeführten Studien sowie durch die Ergebnisse der Studien lassen sich Implikationen für zukünftige Studien ableiten. Die Hauptaufgabe zukünftiger Studien wird es sein, den Einfluss von Stress auf die Konsolidierung von neutralen und emotional erregenden Stimuli weiter herauszustellen und die den Stresseffekten zu Grunde liegenden Mechanismen und Einflussfaktoren weiter zu spezifizieren. Hierfür bietet es sich in erster Linie an, andere und zudem unterschiedliche Stimuli ein- Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 102 zusetzen. Die Bilder, die für die hier berichteten Studien verwendet wurden stellten sich als nicht optimal zur Untersuchung von Stresseffekten auf die Gedächtnisbildung heraus. Da Bilder in Kombination mit Begleitsätzen aber nichtsdestotrotz interessante Stimuli darstellen, da sie komplexe und realistische Episoden bilden, sollte das Ziel sein diese Aufgabe so zu modifizieren, dass Stresseffekte oder auch ausbleibende Effekte nicht auf die Schwierigkeit oder andere Charakteristika der Aufgabe zurückzuführen sind. Dazu sollte die Menge der Bilder erhöht werden, so dass nicht nur 15 Bilder und damit 5 Bilder pro emotionaler Valenz dargeboten werden. Dies würde die Schwierigkeit der Aufgabe deutlich erhöhen. Einen weiteren Aspekt stellt der Abstand zwischen Erlernen und Abruf der Stimuli dar. Die hier verwendeten 24 Stunden könnten auf eine Woche erhöht werden. Für diesen Zeitabstand konnten bereits andere Studien Stresseffekte auf die Gedächtnisbildung zeigen (Payne et al., 2007; Payne et al., 2006). Für zukünftige Studien ist es ebenfalls notwendig die Stichprobengröße zu erhöhen und den Einfluss des hormonellen Zyklus bei Frauen zu kontrollieren. So könnten zum einen nur Frauen untersucht werden, die sich in der Lutealphase befinden, da sich für die Zyklusphase Zusammenhänge zwischen Stress und Gedächtnis zeigen ließen (Andreano et al., 2008). Um den Einfluss des hormonellen Zyklus nicht nur zu kontrollieren, sondern auch dessen Einfluss näher zu beschreiben, bietet es sich zudem an, weibliche Stichproben zu untersuchen, bei denen die Stresseffekte auf die Enkodierung und Konsolidierung in den verschiedenen Zyklusphasen verglichen wird. Ergänzend hierzu sollten auch Männer in die Untersuchung eingeschlossen werden, um den Einfluss des Geschlechts der Probanden auf die Zusammenhänge zu spezifizieren. Die durchgeführten Gedächtnisstudien haben sich mit dem Einfluss basaler Cortisolspiegel auf die Gedächtnisleistung und dem Einfluss von Stress auf die Gedächtniskonsolidierung beschäftigt. Neben der Gedächtniskonsolidierung ist der Einfluss von Stress auf die Enkodierung ebenfalls von Interesse und stellt einen weiteren Forschungsbereich dar, der bisher keine homogenen und eindeutigen Befunde aufweisen kann (Wolf, 2008). Daher sollte in weiteren Studien der Einfluss des TSST auf die Gedächtnisenkodierung untersucht werden. Dabei sollte das selbe Gedächtnismaterial eingesetzt werden, wie in den hier durchgeführten Studien (Buchanan et al., 2001; Buchanan et al., 2003), um einen Vergleich zu ermöglichen. In der dritten Studie wurde der Einfluss naturalistischer Stressoren auf die Stressreaktion erhoben. Darauf aufbauend wäre es interessant zu untersuchen, welchen Einfluss naturalistische Stressoren auf die Gedächtnisleistung haben. Dazu sollte ebenfalls das hier eingesetzte Gedächtnismaterial verwendet werden (Buchanan et al., 2001; Buchanan et al., 2003). Wichtig bei einer solchen Studie ist dabei, dass potenzielle Einflussfaktoren wie die Uhrzeit, Der Einfluss von psychosozialem Stress und Cortisol auf das emotionale Gedächtnis 103 weitere Klausuren, die das Geschlecht und die Zyklusphase der weiblichen Probanden bestmöglich kontrolliert werden. Fazit Zusammenfassend berichtet diese Promotionsarbeit von Cortisol- und Stresseffekten auf die Gedächtnisleistung für emotional neutrale und emotional erregende Stimuli und zeigt, dass auch außerhalb des Laborkontextes die Bedrohung des sozialen Selbst die Stärke der Stressantwort zu beeinflussen scheint. Prüfungssituationen stellten sich dabei als geeignete naturalistische Stressoren heraus, wobei Referate eine stärkere Stressreaktion auslösten, als schriftliche Klausuren. Bei den Gedächtnisstudien wurden positive Effekte basaler Cortisolspiegel auf die Gedächtnisleistung für emotional erregende Bilder in einem 24 Stunden verzögerten freien Abruftest beobachtet. Dieser Effekt zeigte sich jedoch nur bei einer expliziten Lerninstruktion. Für die Auswirkungen eines psychosozialen Stressors auf die Konsolidierung fand sich ein positiver Effekt. Dieser positive Effekt zeigte sich allerdings lediglich für neutrale Bilder, nicht aber für emotional erregende positive und negative Bilder. Zudem zeigte sich in der zweiten Studie ein positiver Zusammenhang zwischen den Cortisolspiegeln und der Gedächtnisleistung für neutrale Stimuli. Die Ergebnisse werden durch die aktuelle Forschungslage nur teilweise gestützt. Aus den durchgeführten Studien wird deutlich, dass verschiedene Einflussfaktoren die Zusammenhänge modulieren. Zu diesen Faktoren gehören neben dem Geschlecht der Probanden auch die emotionale Erregung, die durch die Stimuli sowie durch die Testsituation ausgelöst wird. Des weiteren zeigte sich, dass die Art der Gedächtnistests einen starken Einfluss auf die Stress- und Cortisoleffekte ausübt. Die Aufgabe zukünftiger Studien wird es sein diese Einflussfaktoren näher zu betrachten und weiteren Aufschluss über Cortisol- und Stresseffekte auf die Gedächtnisbildung zu geben. 104 L I T E R AT U R V E R ZE I C H N I S Abercrombie, H. C., Kalin, N. H., Thurow, M. E., Rosenkranz, M. A., Davidson, R. J. (2003). Cortisol variation in humans affects memory for emotionally laden and neutral information. Behav.Neurosci. 117, 505-516. Abercrombie, H. C., Speck, N. S., Monticelli, R. M. (2006). Endogenous cortisol elevations are related to memory facilitation only in individuals who are emotionally aroused. Psychoneuroendocrinology 31, 187-196. Adolphs, R., Denburg, N. L., Tranel, D. (2001). The amygdala's role in long-term declarative memory for gist and detail. Behav.Neurosci. 115, 983-992. Aggleton, J. P. and Brown, M. W. (2006). Interleaving brain systems for episodic and recognition memory. Trends Cogn Sci. 10, 455-463. Akirav, I., Kozenicky, M., Tal, D., Sandi, C., Venero, C., Richter-Levin, G. (2004). A facilitative role for corticosterone in the acquisition of a spatial task under moderate stress. Learn.Mem. 11, 188-195. Akirav, I., Sandi, C., Richter-Levin, G. (2001). Differential activation of hippocampus and amygdala following spatial learning under stress. Eur.J.Neurosci. 14, 719-725. Al-Ayadhi, L. Y. (2005). Neurohormonal changes in medical students during academic stress. Ann.Saudi.Med. 25, 36-40. Altman, J. and Das, G. D. (1965). Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J.Comp Neurol. 124, 319-335. Andreano, J. M., Arjomandi, H., Cahill, L. (2008). Menstrual cycle modulation of the relationship between cortisol and long-term memory. Psychoneuroendocrinology. 33, 874-882. Andreano, J. M. and Cahill, L. (2006). Glucocorticoid release and memory consolidation in men and women. Psychol.Sci. 17, 466-470. Arriza, J. L., Simerly, R. B., Swanson, L. W., Evans, R. M. (1988). The neuronal mineralocorticoid receptor as a mediator of glucocorticoid response. Neuron. 1, 887-900. Atkinson, R. C. & Shiffrin, R. M., 1968. Human memory: a proposed system and its control processes. In K.W.Spence & J. T. Spence (Eds.), The Psychology of Learning and Motivation: Advances in Research and Theory (). Academic Press, New York, pp. 89-195. Barnea, A. and Nottebohm, F. (1994). Seasonal recruitment of hippocampal neurons in adult freeranging black-capped chickadees. Proc.Natl.Acad.Sci.U.S.A. 91, 11217-11221. Beckner, V. E., Tucker, D. M., Delville, Y., Mohr, D. C. (2006). Stress facilitates consolidation of verbal memory for a film but does not affect retrieval. Behav.Neurosci. 120, 518-527. Bliss, T. V. and Gardner-Medwin, A. R. (1973). Long-lasting potentiation of synaptic transmission in the dentate area of the unanaestetized rabbit following stimulation of the perforant path. J.Physiol. 232, 357-374. 105 Bradley, M. M., Greenwald, M. K., Petry, M. C., Lang, P. J. (1992). Remembering pictures: pleasure and arousal in memory. J.Exp.Psychol.Learn.Mem.Cogn. 18, 379-390. Buchanan, T. W., Denburg, N. L., Tranel, D., Adolphs, R. (2001). Verbal and nonverbal emotional memory following unilateral amygdala damage. Learn Mem 8, 326-335. Buchanan, T. W., Karafin, M. S., Adolphs, R. (2003). Selective effects of triazolam on memory for emotional, relative to neutral, stimuli: differential effects on gist versus detail. Behav.Neurosci. 117, 517-525. Buchanan, T. W. and Lovallo, W. R. (2001). Enhanced memory for emotional material following stress-level cortisol treatment in humans. Psychoneuroendocrinology 26, 307-317. Burke, A., Heuer, F., Reisberg, D. (1992). Remembering emotional events. Mem.Cognit. 20, 277-290. Cahill, L. (2003). Sex- and hemisphere-related influences on the neurobiology of emotionally influenced memory. Prog.Neuropsychopharmacol.Biol.Psychiatry 27, 1235-1241. Cahill, L. and Alkire, M. T. (2003). Epinephrine enhancement of human memory consolidation: interaction with arousal at encoding. Neurobiol.Learn.Mem. 79, 194-198. Cahill, L., Gorski, L., Belcher, A., Huynh, Q. (2004). The influence of sex versus sex-related traits on long-term memory for gist and detail from an emotional story. Conscious.Cogn. 13, 391-400. Cahill, L., Gorski, L., Le, K. (2003). Enhanced human memory consolidation with post-learning stress: interaction with the degree of arousal at encoding. Learn.Mem. 10, 270-274. Cahill, L., Haier, R. J., Fallon, J., Alkire, M. T., Tang, C., Keator, D. et al. (1996). Amygdala activity at encoding correlated with long-term, free recall of emotional information. Proc.Natl.Acad.Sci.U.S.A 93, 8016-8021. Cahill, L., Haier, R. J., White, N. S., Fallon, J., Kilpatrick, L., Lawrence, C. et al. (2001). Sex-related difference in amygdala activity during emotionally influenced memory storage. Neurobiol.Learn.Mem. 75, 1-9. Cahill, L. and McGaugh, J. L. (1998). Mechanisms of emotional arousal and lasting declarative memory. Trends Neurosci. 21, 294-299. Cahill, L., Prins, B., Weber, M., McGaugh, J. L. (1994). Beta-adrenergic activation and memory for emotional events. Nature 371, 702-704. Cahill, L., Uncapher, M., Kilpatrick, L., Alkire, M. T., Turner, J. (2004). Sex-related hemispheric lateralization of amygdala function in emotionally influenced memory: an FMRI investigation. Learn Mem 11, 261-266. Cahill, L. and van Stegeren, A. (2003). Sex-related impairment of memory for emotional events with beta-adrenergic blockade. Neurobiol Learn Mem 79, 81-88. Canli, T., Desmond, J. E., Zhao, Z., Gabrieli, J. D. (2002). Sex differences in the neural basis of emotional memories. Proc.Natl.Acad.Sci.U.S.A 99, 10789-10794. 106 Canli, T., Zhao, Z., Brewer, J., Gabrieli, J. D., Cahill, L. (2000). Event-related activation in the human amygdala associates with later memory for individual emotional experience. J.Neurosci. 20, RC99. Cannon, W. B., 1914. The emergency function of the adrenal medulla in pain and the major emotions. In ( Charney, D. S. (2004). Psychobiological mechanisms of resilience and vulnerability: implications for successful adaptation to extreme stress. Am.J.Psychiatry 161, 195-216. Conrad, C. D., Jackson, J. L., Wieczorek, L., Baran, S. E., Harman, J. S., Wright, R. L. et al. (2004). Acute stress impairs spatial memory in male but not female rats: influence of estrous cycle. Pharmacol.Biochem.Behav. 78, 569-579. Conrad, C. D., Lupien, S. J., McEwen, B. S. (1999). Support for a bimodal role for type II adrenal steroid receptors in spatial memory. Neurobiol Learn Mem 72, 39-46. Dayer, A. G., Ford, A. A., Cleaver, K. M., Yassaee, M., Cameron, H. A. (2003). Short-term and longterm survival of new neurons in the rat dentate gyrus. J.Comp Neurol. 460, 563-572. de Kloet, E. R., Joels, M., Holsboer, F. (2005). Stress and the brain: from adaptation to disease. Nat.Rev.Neurosci. 6, 463-475. de Kloet, E. R., Vreugdenhil, E., Oitzl, M. S., Joels, M. (1998). Brain corticosteroid receptor balance in health and disease. Endocr.Rev. 19, 269-301. de Quervain, D. J., Henke, K., Aerni, A., Treyer, V., McGaugh, J. L., Berthold, T. et al. (2003). Glucocorticoid-induced impairment of declarative memory retrieval is associated with reduced blood flow in the medial temporal lobe. Eur.J.Neurosci. 17, 1296-1302. Diamond, D. M., Bennett, M. C., Fleshner, M., Rose, G. M. (1992). Inverted-U relationship between the level of peripheral corticosterone and the magnitude of hippocampal primed burst potentiation. Hippocampus. 2, 421-430. Diamond, D. M., Campbell, A. M., Park, C. R., Halonen, J., Zoladz, P. R. (2007). The temporal dynamics model of emotional memory processing: a synthesis on the neurobiological basis of stress-induced amnesia, flashbulb and traumatic memories, and the Yerkes-Dodson law. Neural Plast. 2007, 1-33. Diamond, D. M., Fleshner, M., Ingersoll, N., Rose, G. M. (1996). Psychological stress impairs spatial working memory: relevance to electrophysiological studies of hippocampal function. Behav.Neurosci. 110, 661-672. Dickerson, S. S. and Kemeny, M. E. (2004). Acute stressors and cortisol responses: a theoretical integration and synthesis of laboratory research. Psychol.Bull. 130, 355-391. Domes, G., Heinrichs, M., Reichwald, U., Hautzinger, M. (2002). Hypothalamic-pituitary-adrenal axis reactivity to psychological stress and memory in middle-aged women: high responders exhibit enhanced declarative memory performance. Psychoneuroendocrinology 27, 843-853. Dudai, Y. (2006). Reconsolidation: the advantage of being refocused. Curr.Opin.Neurobiol. 16, 174178. 107 Earle, T. L., Linden, W., Weinberg, J. (1999). Differential effects of harassment on cardiovascular and salivary cortisol stress reactivity and recovery in women and men. J.Psychosom.Res. 46, 125-141. Easterbrook, J. (1959). The effect of emotion on cue utilization and the organization of behavior. Psychol.Rev. 66, 183-201. Elzinga, B. M., Bakker, A., Bremner, J. D. (2005). Stress-induced cortisol elevations are associated with impaired delayed, but not immediate recall. Psychiatry Res. 134, 211-223. Frankenhaeuser, M., von Wright, M. R., Collins, A., von Wright, J., Sedvall, G., Swahn, C. G. (1978). Sex differences in psychoneuroendocrine reactions to examination stress. Psychosom.Med. 40, 334343. Gaab, J., Sonderegger, L., Scherrer, S., Ehlert, U. (2006). Psychoneuroendocrine effects of cognitivebehavioral stress management in a naturalistic setting-a randomized controlled trial. Psychoneuroendocrinology 31, 428-438. Gould, E., Beylin, A., Tanapat, P., Reeves, A., Shors, T. J. (1999). Learning enhances adult neurogenesis in the hippocampal formation. Nat.Neurosci. 2, 260-265. Gould, E., Tanapat, P., Hastings, N. B., Shors, T. J. (1999). Neurogenesis in adulthood: a possible role in learning. Trends Cogn Sci. 3, 186-192. Gustafsson, B. and Wigstrom, H. (1988). Physiological mechanisms underlying long-term potentiation. Trends Neurosci. 11, 156-162. Hamann, S. (2001). Cognitive and neural mechanisms of emotional memory. Trends Cogn Sci. 5, 394-400. Hamann, S. B., Ely, T. D., Grafton, S. T., Kilts, C. D. (1999). Amygdala activity related to enhanced memory for pleasant and aversive stimuli. Nat.Neurosci. 2, 289-293. Herbert, J., Goodyer, I. M., Grossman, A. B., Hastings, M. H., de Kloet, E. R., Lightman, S. L. et al. (2006). Do corticosteroids damage the brain? J.Neuroendocrinol. 18, 393-411. Herbert, J., Moore, G. F., de la Riva, C., Watts, F. N. (1986). Endocrine responses and examination anxiety. Biol.Psychol. 22, 215-226. Herman, J. P., Figueiredo, H., Mueller, N. K., Ulrich-Lai, Y., Ostrander, M. M., Choi, D. C. et al. (2003). Central mechanisms of stress integration: hierarchical circuitry controlling hypothalamo-pituitaryadrenocortical responsiveness. Front Neuroendocrinol. 24, 151-180. Herman, J. P., Prewitt, C. M., Cullinan, W. E. (1996). Neuronal circuit regulation of the hypothalamopituitary-adrenocortical stress axis. Crit Rev.Neurobiol 10, 371-394. Het, S., Ramlow, G., Wolf, O. T. (2005). A meta-analytic review of the effects of acute cortisol administration on human memory. Psychoneuroendocrinology 30, 771-784. Heuer, F. and Reisberg, D. (1990). Vivid memories of emotional events: the accuracy of remembered minutiae. Mem.Cognit. 18, 496-506. 108 Hsu, F. C., Garside, M. J., Massey, A. E., McAllister-Williams, R. H. (2003). Effects of a single dose of cortisol on the neural correlates of episodic memory and error processing in healthy volunteers. Psychopharmacology (Berl). 167, 431-442. Jackson, E. D., Payne, J. D., Nadel, L., Jacobs, W. J. (2006). Stress differentially modulates fear conditioning in healthy men and women. Biol.Psychiatry 59, 516-522. Jacobson, L. (2005). Hypothalamic-pituitary-adrenocortical axis regulation. Endocrinol.Metab Clin.North Am. 34, 271-92, vii. Jelicic, M., Geraerts, E., Merckelbach, H., Guerrieri, R. (2004). Acute stress enhances memory for emotional words, but impairs memory for neutral words. Int.J.Neurosci. 114, 1343-1351. Joels, M., Krugers, H., Karst, H. (2008). Stress-induced changes in hippocampal function. Prog.Brain Res. 167:3-15., 3-15. Joels, M., Pu, Z., Wiegert, O., Oitzl, M. S., Krugers, H. J. (2006). Learning under stress: how does it work? Trends Cogn Sci. 10, 152-158. Kajantie, E. and Phillips, D. I. (2006). The effects of sex and hormonal status on the physiological response to acute psychosocial stress. Psychoneuroendocrinology 31, 151-178. Karst, H., Berger, S., Turiault, M., Tronche, F., Schutz, G., Joels, M. (2005). Mineralocorticoid receptors are indispensable for nongenomic modulation of hippocampal glutamate transmission by corticosterone. Proc.Natl.Acad.Sci.U.S.A. 102, 19204-19207. Kelly, M. M., Tyrka, A. R., Anderson, G. M., Price, L. H., Carpenter, L. L. (2008). Sex differences in emotional and physiological responses to the Trier Social Stress Test. J.Behav.Ther.Exp.Psychiatry. 39, 87-98. Kensinger, E. A. and Corkin, S. (2004). Two routes to emotional memory: distinct neural processes for valence and arousal. Proc.Natl.Acad.Sci.U.S.A. 101, 3310-3315. Kim, J. J. and Diamond, D. M. (2002). The stressed hippocampus, synaptic plasticity and lost memories. Nat.Rev.Neurosci. 3, 453-462. King, J. A., Rosal, M. C., Ma, Y., Reed, G., Kelly, T. A., Stanek, E. J., III et al. (2000). Sequence and seasonal effects of salivary cortisol. Behav.Med. 26, 67-73. Kirschbaum, C. and Hellhammer, D. H. (1989). Salivary cortisol in psychobiological research: an overview. Neuropsychobiology 22, 150-169. Kirschbaum, C., Kudielka, B. M., Gaab, J., Schommer, N. C., Hellhammer, D. H. (1999). Impact of gender, menstrual cycle phase, and oral contraceptives on the activity of the hypothalamus-pituitaryadrenal axis. Psychosom.Med. 61, 154-162. Kirschbaum, C., Pirke, K. M., Hellhammer, D. H. (1993). The 'Trier Social Stress Test'--a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology 28, 76-81. Kirschbaum, C., Wolf, O. T., May, M., Wippich, W., Hellhammer, D. H. (1996). Stress- and treatmentinduced elevations of cortisol levels associated with impaired declarative memory in healthy adults. Life Sci. 58, 1475-1483. 109 Kirschbaum, C., Wust, S., Hellhammer, D. (1992). Consistent sex differences in cortisol responses to psychological stress. Psychosom.Med. 54, 648-657. Kozorovitskiy, Y. and Gould, E. (2004). Dominance hierarchy influences adult neurogenesis in the dentate gyrus. J.Neurosci. 24, 6755-6759. Kudielka, B. M., Hellhammer, D. H., Wust, S. (2009). Why do we respond so differently? Reviewing determinants of human salivary cortisol responses to challenge. Psychoneuroendocrinology. 34, 2-18. Kudielka, B. M. and Kirschbaum, C. (2003). Awakening cortisol responses are influenced by health status and awakening time but not by menstrual cycle phase. Psychoneuroendocrinology 28, 35-47. Kudielka, B. M. and Kirschbaum, C. (2005). Sex differences in HPA axis responses to stress: a review. Biol.Psychol. 69, 113-132. Kuhlmann, S., Kirschbaum, C., Wolf, O. T. (2005). Effects of oral cortisol treatment in healthy young women on memory retrieval of negative and neutral words. Neurobiol.Learn.Mem. 83, 158-162. Kuhlmann, S., Piel, M., Wolf, O. T. (2005). Impaired memory retrieval after psychosocial stress in healthy young men. J.Neurosci. 25, 2977-2982. Kuhlmann, S. and Wolf, O. T. (2005). Cortisol and memory retrieval in women: influence of menstrual cycle and oral contraceptives. Psychopharmacology (Berl) 183, 65-71. Kuhlmann, S. and Wolf, O. T. (2006a). A non-arousing test situation abolishes the impairing effects of cortisol on delayed memory retrieval in healthy women. Neurosci.Lett. 399, 268-272. Kuhlmann, S. and Wolf, O. T. (2006b). Arousal and cortisol interact in modulating memory consolidation in healthy young men. Behav.Neurosci. 120, 217-223. Kumsta, R., Entringer, S., Koper, J. W., van Rossum, E. F., Hellhammer, D. H., Wust, S. (2007). Sex specific associations between common glucocorticoid receptor gene variants and hypothalamuspituitary-adrenal axis responses to psychosocial stress. Biol.Psychiatry. 62, 863-869. LaBar, K. S. and Cabeza, R. (2006). Cognitive neuroscience of emotional memory. Nat.Rev.Neurosci. 7, 54-64. Lacey, K., Zaharia, M. D., Griffiths, J., Ravindran, A. V., Merali, Z., Anisman, H. (2000). A prospective study of neuroendocrine and immune alterations associated with the stress of an oral academic examination among graduate students. Psychoneuroendocrinology 25, 339-356. Lazarus, R. S. & Folkman, S. (1984). Stress, appraisal and coping. New York: Springer Publishing Company, Inc. Lemaire, V., Koehl, M., Le, M. M., Abrous, D. N. (2000). Prenatal stress produces learning deficits associated with an inhibition of neurogenesis in the hippocampus. Proc.Natl.Acad.Sci.U.S.A. 97, 1103211037. Liberzon, J., Abelson, J. L., King, A., Liberzon, I. (2008). Naturalistic stress and cortisol response to awakening: adaptation to seafaring. Psychoneuroendocrinology. 33, 1023-1026. 110 Lindahl, M., Theorell, T., Lindblad, F. (2005). Test performance and self-esteem in relation to experienced stress in Swedish sixth and ninth graders--saliva cortisol levels and psychological reactions to demands. Acta Paediatr. 94, 489-495. Loft, P., Thomas, M. G., Petrie, K. J., Booth, R. J., Miles, J., Vedhara, K. (2007). Examination stress results in altered cardiovascular responses to acute challenge and lower cortisol. Psychoneuroendocrinology 32, 367-375. Lovallo, W. R., Farag, N. H., Vincent, A. S., Thomas, T. L., Wilson, M. F. (2006). Cortisol responses to mental stress, exercise, and meals following caffeine intake in men and women. Pharmacol.Biochem.Behav. 83, 441-447. Lovallo, W. R., Pincomb, G. A., Edwards, G. L., Brackett, D. J., Wilson, M. F. (1986). Work pressure and the type A behavior pattern exam stress in male medical students. Psychosom.Med. 48, 125-133. Lupien, S. J., Gillin, C. J., Hauger, R. L. (1999). Working memory is more sensitive than declarative memory to the acute effects of corticosteroids: a dose-response study in humans. Behav.Neurosci. 113, 420-430. Lupien, S. J. and Lepage, M. (2001). Stress, memory, and the hippocampus: can't live with it, can't live without it. Behav.Brain Res. 127, 137-158. Lupien, S. J. and McEwen, B. S. (1997). The acute effects of corticosteroids on cognition: integration of animal and human model studies. Brain Res.Brain Res.Rev. 24, 1-27. Lynch, M. A. (2004). Long-term potentiation and memory. Physiol Rev. 84, 87-136. Madsen, T. M., Kristjansen, P. E., Bolwig, T. G., Wortwein, G. (2003). Arrested neuronal proliferation and impaired hippocampal function following fractionated brain irradiation in the adult rat. Neuroscience. 119, 635-642. Magarinos, A. M., Orchinik, M., McEwen, B. S. (1998). Morphological changes in the hippocampal CA3 region induced by non-invasive glucocorticoid administration: a paradox. Brain Res. 809, 314318. Maheu, F. S., Collicutt, P., Kornik, R., Moszkowski, R., Lupien, S. J. (2005). The perfect time to be stressed: a differential modulation of human memory by stress applied in the morning or in the afternoon. Prog.Neuropsychopharmacol.Biol.Psychiatry 29, 1281-1288. Malarkey, W. B., Pearl, D. K., Demers, L. M., Kiecolt-Glaser, J. K., Glaser, R. (1995). Influence of academic stress and season on 24-hour mean concentrations of ACTH, cortisol, and beta-endorphin. Psychoneuroendocrinology 20, 499-508. Malberg, J. E. and Duman, R. S. (2003). Cell proliferation in adult hippocampus is decreased by inescapable stress: reversal by fluoxetine treatment. Neuropsychopharmacology. 28, 1562-1571. Markowitsch, H. J. (2002). Dem Gedächtnis auf der Spur: Vom Erinnern und Vergessen. Darmstadt: Primus Verlag. Mason, J. W. (1968). A review of psychoendocrine research on the pituitary-adrenal cortical system. Psychosom.Med. 30, Suppl-607. 111 McEwen, B. S. (1998). Stress, adaptation, and disease. Allostasis and allostatic load. Ann.N.Y.Acad.Sci. 840, 33-44. McEwen, B. S. (2000). The neurobiology of stress: from serendipity to clinical relevance. Brain Res. 886, 172-189. McEwen, B. S. and Wingfield, J. C. (2003). The concept of allostasis in biology and biomedicine. Horm.Behav. 43, 2-15. McGaugh, J. L., Cahill, L., Roozendaal, B. (1996). Involvement of the amygdala in memory storage: interaction with other brain systems. Proc.Natl.Acad.Sci.U.S A 93, 13508-13514. McKittrick, C. R., Magarinos, A. M., Blanchard, D. C., Blanchard, R. J., McEwen, B. S., Sakai, R. R. (2000). Chronic social stress reduces dendritic arbors in CA3 of hippocampus and decreases binding to serotonin transporter sites. Synapse. 36, 85-94. Mesches, M. H., Fleshner, M., Heman, K. L., Rose, G. M., Diamond, D. M. (1999). Exposing rats to a predator blocks primed burst potentiation in the hippocampus in vitro. J.Neurosci. 19, RC18. Miller, G. A. (1994). The magical number seven, plus or minus two: some limits on our capacity for processing information. Psychol.Rev. 101, 343-352. Mirescu, C., Peters, J. D., Gould, E. (2004). Early life experience alters response of adult neurogenesis to stress. Nat.Neurosci. 7, 841-846. Monk, C. S. and Nelson, C. A. (2002). The effects of hydrocortisone on cognitive and neural function: a behavioral and event-related potential investigation. Neuropsychopharmacology 26, 505-519. Nader, K., Schafe, G. E., LeDoux, J. E. (2000). The labile nature of consolidation theory. Nat.Rev.Neurosci. 1, 216-219. Nater, U. M., La Marca, R., Florin, L., Moses, A., Langhans, W., Koller, M. M. et al. (2006). Stressinduced changes in human salivary alpha-amylase activity-associations with adrenergic activity. Psychoneuroendocrinology 31, 49-58. Nater, U. M., Moor, C., Okere, U., Stallkamp, R., Martin, M., Ehlert, U. et al. (2007). Performance on a declarative memory task is better in high than low cortisol responders to psychosocial stress. Psychoneuroendocrinology 32, 758-763. Ng, V., Koh, D., Chia, S. E. (2003). Examination stress, salivary cortisol, and academic performance. Psychol.Rep. 93, 1133-1134. Ng, V., Koh, D., Mok, B. Y., Chia, S. E., Lim, L. P. (2003). Salivary biomarkers associated with academic assessment stress among dental undergraduates. J.Dent.Educ. 67, 1091-1094. O'Carroll, R. E., Drysdale, E., Cahill, L., Shajahan, P., Ebmeier, K. P. (1999). Stimulation of the noradrenergic system enhances and blockade reduces memory for emotional material in man. Psychol.Med. 29, 1083-1088. Otte, C., Hart, S., Neylan, T. C., Marmar, C. R., Yaffe, K., Mohr, D. C. (2005). A meta-analysis of cortisol response to challenge in human aging: importance of gender. Psychoneuroendocrinology 30, 8091. 112 Packard, M. G. and Teather, L. A. (1998). Amygdala modulation of multiple memory systems: hippocampus and caudate-putamen. Neurobiol.Learn.Mem. 69, 163-203. Pavlides, C., Ogawa, S., Kimura, A., McEwen, B. S. (1996). Role of adrenal steroid mineralocorticoid and glucocorticoid receptors in long-term potentiation in the CA1 field of hippocampal slices. Brain Res. 738, 229-235. Payne, J. D., Jackson, E. D., Hoscheidt, S., Ryan, L., Jacobs, W. J., Nadel, L. (2007). Stress administered prior to encoding impairs neutral but enhances emotional long-term episodic memories. Learn.Mem. 14, 861-868. Payne, J. D., Jackson, E. D., Ryan, L., Hoscheidt, S., Jacobs, J. W., Nadel, L. (2006). The impact of stress on neutral and emotional aspects of episodic memory. Memory. 14, 1-16. Phelps, E. A. (2004). Human emotion and memory: interactions of the amygdala and hippocampal complex. Curr.Opin.Neurobiol. 14, 198-202. Pritzel, M., Brand, M., & Markowitsch, H. J. (2003). Gehirn und Verhalten: EIn Grundkurs der physiologischen Psychologie. Heidelberg, Berlin: Spektrum Akademischer Verlag. Putman, P., van Honk, J., Kessels, R. P., Mulder, M., Koppeschaar, H. P. (2004). Salivary cortisol and short and long-term memory for emotional faces in healthy young women. Psychoneuroendocrinology 29, 953-960. Raber, J., Fan, Y., Matsumori, Y., Liu, Z., Weinstein, P. R., Fike, J. R. et al. (2004). Irradiation attenuates neurogenesis and exacerbates ischemia-induced deficits. Ann.Neurol. 55, 381-389. Raber, J., Rola, R., LeFevour, A., Morhardt, D., Curley, J., Mizumatsu, S. et al. (2004). Radiationinduced cognitive impairments are associated with changes in indicators of hippocampal neurogenesis. Radiat.Res. 162, 39-47. Rimmele, U., Domes, G., Mathiak, K., Hautzinger, M. (2003). Cortisol has different effects on human memory for emotional and neutral stimuli. Neuroreport 14, 2485-2488. Rohleder, N., Beulen, S. E., Chen, E., Wolf, J. M., Kirschbaum, C. (2007). Stress on the dance floor: the cortisol stress response to social-evaluative threat in competitive ballroom dancers. Personality and Social Psychology Bulletin 33, 69-84. Rohleder, N., Nater, U. M., Wolf, J. M., Ehlert, U., Kirschbaum, C. (2004). Psychosocial stress-induced activation of salivary alpha-amylase: an indicator of sympathetic activity? Ann.N.Y.Acad.Sci. 1032, 258-263. Rohleder, N., Schommer, N. C., Hellhammer, D. H., Engel, R., Kirschbaum, C. (2001). Sex differences in glucocorticoid sensitivity of proinflammatory cytokine production after psychosocial stress. Psychosom.Med. 63, 966-972. Rohleder, N., Wolf, J. M., Piel, M., Kirschbaum, C. (2003). Impact of oral contraceptive use on glucocorticoid sensitivity of pro-inflammatory cytokine production after psychosocial stress. Psychoneuroendocrinology 28, 261-273. 113 Roosevelt, R. W., Smith, D. C., Clough, R. W., Jensen, R. A., Browning, R. A. (2006). Increased extracellular concentrations of norepinephrine in cortex and hippocampus following vagus nerve stimulation in the rat. Brain Res. 1119, 124-132. Roozendaal, B., Barsegyan, A., Lee, S. (2008). Adrenal stress hormones, amygdala activation, and memory for emotionally arousing experiences. Prog.Brain Res. 167:79-97., 79-97. Roozendaal, B., de Quervain, D. J., Ferry, B., Setlow, B., McGaugh, J. L. (2001). Basolateral amygdala-nucleus accumbens interactions in mediating glucocorticoid enhancement of memory consolidation. J.Neurosci. 21, 2518-2525. Roozendaal, B., Nguyen, B. T., Power, A. E., McGaugh, J. L. (1999). Basolateral amygdala noradrenergic influence enables enhancement of memory consolidation induced by hippocampal glucocorticoid receptor activation. Proc.Natl.Acad.Sci.U.S A 96, 11642-11647. Roozendaal, B., Okuda, S., de Quervain, D. J., McGaugh, J. L. (2006a). Glucocorticoids interact with emotion-induced noradrenergic activation in influencing different memory functions. Neuroscience. 138, 901-910. Roozendaal, B., Okuda, S., Van der Zee, E. A., McGaugh, J. L. (2006b). Glucocorticoid enhancement of memory requires arousal-induced noradrenergic activation in the basolateral amygdala. Proc.Natl.Acad.Sci.U.S.A. 103, 6741-6746. Sandi, C., Loscertales, M., Guaza, C. (1997). Experience-dependent facilitating effect of corticosterone on spatial memory formation in the water maze. Eur.J.Neurosci. 9, 637-642. Sandi, C. and Rose, S. P. (1994). Corticosterone enhances long-term retention in one-day-old chicks trained in a weak passive avoidance learning paradigm. Brain Res. 647, 106-112. Schlotz, W., Hellhammer, J., Schulz, P., Stone, A. A. (2004). Perceived work overload and chronic worrying predict weekend-weekday differences in the cortisol awakening response. Psychosom.Med. 66, 207-214. Schoofs, D., Hartmann, R., Wolf, O. T. (2008). Neuroendocrine stress responses to an oral academic examination: no strong influence of sex, repeated participation and personality traits. Stress. 11, 5261. Schwabe, L., Bohringer, A., Chatterjee, M., Schachinger, H. (2008). Effects of pre-learning stress on memory for neutral, positive and negative words: different roles of cortisol and autonomic arousal. Neurobiol.Learn.Mem. 90, 44-53. Scoville, W. B. and Milner, B. (1957). Loss of recent memory after bilateral hippocampal lesions. J.Neurol.Neurosurg.Psychiatry. 20, 11-21. Seeman, T. E., Singer, B., Wilkinson, C. W., McEwen, B. (2001). Gender differences in age-related changes in HPA axis reactivity. Psychoneuroendocrinology 26, 225-240. Selye, H. (1973). The Evolution of the Stress concept. American Scientist 61, 692-699. Selye, H. and Fortier, C. (1950). Adaptive reaction to stress. Psychosom.Med. 12, 149-157. 114 Shors, T. J., Miesegaes, G., Beylin, A., Zhao, M., Rydel, T., Gould, E. (2001). Neurogenesis in the adult is involved in the formation of trace memories. Nature. 410, 372-376. Shors, T. J., Townsend, D. A., Zhao, M., Kozorovitskiy, Y., Gould, E. (2002). Neurogenesis may relate to some but not all types of hippocampal-dependent learning. Hippocampus. 12, 578-584. Smeets, T., Jelicic, M., Merckelbach, H. (2006). The effect of acute stress on memory depends on word valence. Int.J.Psychophysiol. 62, 30-37. Smeets, T., Otgaar, H., Candel, I., Wolf, O. T. (2008). True or false? Memory is differentially affected by stress-induced cortisol elevations and sympathetic activity at consolidation and retrieval. Psychoneuroendocrinology 33, 1378-1386. Snyder, J. S., Hong, N. S., McDonald, R. J., Wojtowicz, J. M. (2005). A role for adult neurogenesis in spatial long-term memory. Neuroscience. 130, 843-852. Spangler, G. (1997). Psychological and physiological responses during an exam and their relation to personality characteristics. Psychoneuroendocrinology 22, 423-441. Squire, L. R. (1992). Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychol.Rev. 99, 195-231. Stowell, J. R. (2003). Use and abuse of academic examinations in stress research. Psychosom.Med. 65, 1055-1057. Strange, B. A. and Dolan, R. J. (2004). Beta-adrenergic modulation of emotional memory-evoked human amygdala and hippocampal responses. Proc.Natl.Acad.Sci.U.S.A 101, 11454-11458. Stroud, L. R., Salovey, P., Epel, E. S. (2002). Sex differences in stress responses: social rejection versus achievement stress. Biol.Psychiatry 52, 318-327. Tanapat, P., Hastings, N. B., Rydel, T. A., Galea, L. A., Gould, E. (2001). Exposure to fox odor inhibits cell proliferation in the hippocampus of adult rats via an adrenal hormone-dependent mechanism. J.Comp Neurol. 437, 496-504. Thompson, R. F. (2005). In search of memory traces. Annu.Rev.Psychol. 56, 1-23. Tops, M., van der Pompe, G., Baas, D., Mulder, L. J., Den Boer, J. A., Meijman, T. F. et al. (2003). Acute cortisol effects on immediate free recall and recognition of nouns depend on stimulus valence. Psychophysiology 40, 167-173. Tulving, E., 1995. Organization of memory: Quo vadis? In M.S.Gazzaniga (Ed.), The cognitive neurosciences (). MIT Press, Cambridge, pp. 839-847. Van Cauter, E. (1990). Diurnal and ultradian rhythms in human endocrine function: a minireview. Horm.Res. 34, 45-53. van Honk, J., Kessels, R. P., Putman, P., Jager, G., Koppeschaar, H. P., Postma, A. (2003). Attentionally modulated effects of cortisol and mood on memory for emotional faces in healthy young males. Psychoneuroendocrinology. 28, 941-948. 115 van Stegeren, A. H. (2009). Imaging stress effects on memory: a review of neuroimaging studies. Can.J.Psychiatry. 54, 16-27. van Stegeren, A. H., Goekoop, R., Everaerd, W., Scheltens, P., Barkhof, F., Kuijer, J. P. et al. (2005). Noradrenaline mediates amygdala activation in men and women during encoding of emotional material. Neuroimage. 24, 898-909. van Stegeren, A. H., Wolf, O. T., Kindt, M. (2008). Salivary alpha amylase and cortisol responses to different stress tasks: impact of sex. Int.J.Psychophysiol. 69, 33-40. Venero, C. and Borrell, J. (1999). Rapid glucocorticoid effects on excitatory amino acid levels in the hippocampus: a microdialysis study in freely moving rats. Eur.J.Neurosci. 11, 2465-2473. Walker, B. R., Best, R., Noon, J. P., Watt, G. C., Webb, D. J. (1997). Seasonal variation in glucocorticoid activity in healthy men. J.Clin.Endocrinol.Metab. 82, 4015-4019. Weekes, N., Lewis, R., Patel, F., Garrison-Jakel, J., Berger, D. E., Lupien, S. J. (2006). Examination stress as an ecological inducer of cortisol and psychological responses to stress in undergraduate students. Stress 9, 199-206. Wiegert, O., Joels, M., Krugers, H. (2006). Timing is essential for rapid effects of corticosterone on synaptic potentiation in the mouse hippocampus. Learn.Mem. 13, 110-113. Wiegert, O., Pu, Z., Shor, S., Joels, M., Krugers, H. (2005). Glucocorticoid receptor activation selectively hampers N-methyl-D-aspartate receptor dependent hippocampal synaptic plasticity in vitro. Neuroscience. 135, 403-411. Wiegratz, I., Kutschera, E., Lee, J. H., Moore, C., Mellinger, U., Winkler, U. H. et al. (2003). Effect of four different oral contraceptives on various sex hormones and serum-binding globulins. Contraception 67, 25-32. Wolf, O. T. (2006). Effects of stress hormones on the structure and function of the human brain. Expert Review of Endocrinology & Metabolism 1, 623-632. Wolf, O. T. (2008). The influence of stress hormones on emotional memory: relevance for psychopathology. Acta Psychol.(Amst). 127, 513-531. Wolf, O. T., Schommer, N. C., Hellhammer, D. H., McEwen, B. S., Kirschbaum, C. (2001). The relationship between stress induced cortisol levels and memory differs between men and women. Psychoneuroendocrinology 26, 711-720. Zeller, A., Handschin, D., Gyr, N., Martina, B., Battegay, E. (2004). Blood pressure and heart rate of students undergoing a medical licensing examination. Blood Press 13, 20-24. Zola-Morgan, S. and Squire, L. R. (1986). Memory impairment in monkeys following lesions limited to the hippocampus. Behav.Neurosci. 100, 155-160. I A B K Ü R Z U N GS V E R ZE I C H N I S AAS Allgemeines Adaptationssyndrom ACTH Adrenocorticotropes Hormon ANS Autonomes Nervensystem BLA Basolaterale Amygdala CBG Cortisolbindendes Globulin CPT Cold Pressure Test CRH Corticotropin Releasing Hormon GC Glucocorticoide GR Glucocorticoidrezeptor HHNA Hypothalamus-Hypophysen-Nebennierenrinden-Achse KZG Kurzzeitgedächtnis LTP Long term potentiation/ Langzeitpotenzierung LZG Langzeitgedächtnis MR Mineralocorticoidrezeptor SNS Sympathisches Nervensystem TSST Trier Social Stress Test ZNS Zentrales Nervensystem II A B B I LD U N GS V E R ZE I C H N I S 1.1.1.2 Abbildung 1: Schematische Einteilung des Gedächtnisses nach zeitlichen und inhaltlichen Aspekten 1.1.3 Abbildung 2: Die mit dem Gedächtnis assoziierten Hirngebiet. (aus: Markowitsch, 2002) 1.3 Abbildung 3: Einflussmöglichkeiten von Cortisol und Stress auf die verschiedenen Gedächtnisphasen. 5.2.1 Abbildung 4: Einflussfaktoren auf die Stressantwort in Reaktion auf naturalistische Stressoren und Laborstressoren. III T AB E L LE N V E R ZE I C H N I S 1.3.2 Tabelle1: Zusammenfassung der Stressstudien im humanexperimentellen Bereich. IV C U R R I C U L U M V I T AE Persönliche Daten Geburtsdatum Geburtsort Familienstand Staatsangehörigkeit 09.08.1982 Leverkusen ledig deutsch Schulischer Werdegang 1989 – 1993 1993 – 2002 Gemeinschaftsgrundschule Dönhoffstraße, Leverkusen Marienschule Opladen, Gymnasium Allgemeine Hochschulreife (Note 1,5) 10/ 02 – 03/ 07 Studium der Psychologie an der Universität Bielefeld mit den Schwerpunkten Arbeits- und Organisationspsychologie sowie klinische Psychologie 03/ 07 Abschluss des Psychologie- Studiums (Dipl.-Psych.) nach 9 Semestern mit der Note „Gut“ (1,7) Titel der Diplomarbeit: „Stress und semantisches Gedächtnis“ (Note 1,0) 04/ 07 – 09/ 2007 Promotionsstudium Psychologie an der Universität Bielefeld Promotionsstudium Psychologie an der Ruhr-Universität Bochum Seit 10/ 2007 Berufspraxis 12/ 05 - 02/ 07 04/ 07 – 09/ 07 10/07 – 09/ 09 studentische Hilfskraft der Arbeitseinheit allgemeine Psychologie 2 der psychologischen Fakultät der Universität Bielefeld Wissenschaftliche Mitarbeiterin an der Fakultät für Psychologie (Abt. Psychologie, Allgemeine Psychologie 2) an der Universität Bielefeld Wissenschaftliche Mitarbeiterin an der Fakultät für Psychologie (Kognitionspsychologie) an der Ruhr- Universität Bochum V Veröffentlichungen Preuß, D, Schoofs, D, Schlotz, W, Wolf, OT (2009): The stressed student: Influence of a written examination and oral presentation on salivary cortisol concentrations in university students Stress, in press Preuß, D & Wolf, OT (2009): Post-learning psychosocial stress enhances consolidation of neutral stimuli. Neurobiology of learning and memory; 92(3): 318-326. Preuß D, Schoofs D, Wolf OT (2008): Associations between endogenous cortisol levels and emotional memory in young women: Influence of encoding instructions. Stress, in press. Schoofs D, Preuß D, Wolf OT (2008): Psychosocial stress induces working memory impairments in an n-back paradigm. Psychoneuroendocrinology; 33(5): 643-653. Posterbeiträge Preuß, D &Wolf, OT(2009): Der Einfluss von psychosozialem Stress auf die Konsolidierung emotionaler und neutraler Stimuli. 35. Arbeitstagung Psychophysiologie und Methodik, APM Preuß D, Schoofs D, Wolf OT (2008): Neuroendocrine responses to academic stress International Congress of Psychology (ICP), Berlin. Preuß D, Schoofs D, Wolf OT (2008): Neuroendocrine responses to academic stress 39th Annual Conference of the International Society of Psychoneuroendocrinology (ISPNE), Dresden. Preuß D, Schoofs D, Wolf OT (2007): The influence of psychosocial Stress on semantic memory. Fachtagung "Psychologie und Gehirn" der Deutsche Gesellschaft für Psychophysiologie und ihre Anwendung (DGPA) zusammen mit der Fachgruppe Biologische Psy- VI chologie und Neuropsychologie der Deutschen Gesellschaft für Psychologie (DGPs), Dortmund. Wolf OT, Preuß D, Schoofs D (2007): Psychosocial stress influences working memory and semantic memory in humans. Neuroscience 2007, the Society's 37th annual meeting. Lehre SS 2007: Universität Bielefeld/ Psychologie Diplom Seminar Stress und Gedächtnis WS 2007/2008: Ruhr- Universität Bochum/ Bachelor Psychologie Seminar Kognition I, Wahrnehmung, Aufmerksamkeit und Gedächtnis SS 2008: Ruhr- Universität Bochum/ Bachelor Psychologie Seminar Kognition II, Sprache, Denken und Problemlösen WS 2008/2009: Ruhr- Universität Bochum/ Bachelor Psychologie Seminar Kognition I, Wahrnehmung, Aufmerksamkeit und Gedächtnis SS 2009: Ruhr- Universität Bochum/ Bachelor Psychologie Seminar Kognition II, Sprache, Denken und Problemlösen ERKLÄRUNG Bochum, den 11.09.2009 Ich versichere hiermit, dass die vorliegende Dissertationsschrift eigenständig und ausschließlich von meiner Person verfasst wurde, ich keine anderen als die angegebenen Hilfsmittel und Quellen verwendet habe und sie weder in der vorliegenden noch in einer anderen Fassung in einer anderen Fakultät bisher vorgelegen hat Diana Preuß