(ME-Virus) in L-Zellen
Werbung
 in L-Zellen")
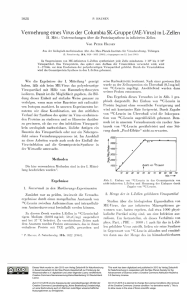
P. HAUSEN 1028 Vermehrung eines Virus der C olum bia SK-Gruppe (ME-Virus) in L-Zellen I I . M itt.: Untersuchungen über die Proteinsynthese in infizierten Zellen Von P e te r H ausen Aus der biologisch-medizinischen Abt. des Max-Planck-Instituts für Virusforschung, Tübingen (Z. Naturforschg. 18 b, 1028— 1031 [1963] ; eingegangen am 12. Juni 1963) In Suspensionen von ME-infizierten L-Zellen synthetisiert jede Zelle mindestens 1-105 bis 3 -IO5 Viruspartikel. Das Virusprotein, das später zum Aufbau der Virusteilchen verwendet wird, wird 1 —2 Stdn. vor dem Erscheinen der polyederartigen Viruspartikel gebildet. Durch die Virusinfektion wird die Gesamtprotein-Synthese in den L-Zellen gehemmt. Wie die Ergebnisse der I. Mitteilung 1 gezeigt haben, läßt sich beim ME-Virus das polyederartige Viruspartikel mit Hilfe von Hammelerythrocyten isolieren. Damit ist die Möglichkeit gegeben, die Bil­ dung dieser Einheit auf einfache Weise genauer zu verfolgen, wenn man seine Bausteine mit radioakti­ ven Isotopen markiert. In unseren Experimenten be­ nutzten wir diese Kombination, um den zeitlichen Verlauf der Synthese des später im Virus erscheinen­ den Proteins zu studieren und so Hinweise darüber zu gewinnen, ob das vor den infektiösen Virusparti­ keln serologisch nachweisbare, lösliche Antigen ein Baustein des Viruspartikels oder nur ein Nebenpro­ dukt seines Vermehrungsprozesses ist. Im Anschluß an diese Arbeiten wurde auch noch der Einfluß der Virusinfektion auf die Gesamtprotein-Synthese in der Wirtszelle untersucht. seine Radioaktivität bestimmt. Nach einer gewissen Zeit wurde zu der Zellsuspension ein Überschuß (0,5 mg/ml) von 12C-Leucin zugefügt. Anschließend wurden dann weitere Proben entnommen. Das Ergebnis dieses Versuches ist in Abb. 1 gra­ phisch dargestellt. Der Einbau von 14C-Leucin in Protein beginnt ohne wesentliche Verzögerung und wird mit konstanter Rate fortgesetzt. Durch Zugabe von 12C-Leucin im Überschuß wird die Inkorpora­ tion von 14C-Leucin augenblicklich gehemmt. Dem­ nach ist in unserem Versuchsansatz ein rascher Aus­ tausch von 14C-Leucin gewährleistet und eine Stö­ rung durch „Pool-Effekte“ nicht zu erwarten. Methoden Die hier verwendeten Methoden sind in der I. Mittei­ lung beschrieben worden 1. Min.— Ergebnisse 1. Vorversuch zu den Markierungs-Experimenten Zunächst war zu prüfen, inwieweit die Versuchs­ ergebnisse durch einen mangelhaften Austausch von 14C-Leucin zwischen Außenmedium und intrazellulä­ rem Aminosäurevorrat beeinflußt werden können. Zu diesem Zweck wurden L-Zellen in 14C-Leucin-haltigem Medium (0,001 mg/ml; 50 /<C/mg) suspendiert und bei 37 °C bebrütet. Zu verschiedenen Zeiten nach dem Ansetzen wurden Proben entnommen, das in ihnen enthaltene Protein mit TCE gefällt, gewaschen und 1 P. H ausen, Z. Naturforschg. 18b. 1022 [1963]. Abb. 1. Einbau von 14C-Leucin in das Gesamtprotein von nicht-infizierten L-Zellen und Hemmung des Einbaues durch Zugabe von 12C-Leucin. 2. Menge der in L-Zellen gebildeten Viruspartikel Studien über die biologischen Eigenschaften von ME-Virus, das aus infizierten Mäusegehirnen ge­ wonnen war. hatten ergeben, daß etwa 1000 physi­ kalische Partikel nötig sind, um eine Infektion aus­ zulösen. Um festzustellen, ob dieses Verhältnis von phvs. Part. : PBE = 1000 : 1 auch für das in L-Zel­ len gebildete Virus zutrifft, ließen wir seine Synthese in Gegenwart von 14C-Leucin ablaufen und ermittel­ ten dann aus der Menge des im hämadsorbierbaren Unauthenticated Download Date | 2/14/17 3:17 AM VERMEHRUNG DES ME-VIRUS IN L-ZELLEN II Material enthaltenen Isotops die ungefähre Zahl der Virusteildien. Infizierte L-Zellen wurden unmittelbar nach der In­ fektion 1-mal mit leucinfreiem Gewebekulturmedium gewaschen, um intrazelluläres 12C-Leucin möglichst weitgehend zu entfernen, und dann mit 14C-Leucin-haltigem Medium inkubiert. 8 Stdn. p.i. wurden die Zellen lysiert, die Viruspartikel an Erythrocyten adsorbiert und ihre Radioaktivität bestimmt (s. Methoden I. Mitt.). Für die Berechnung der Virusmenge aus der gefunde­ nen Radioaktivität und der spezifischen Aktivität des angebotenen Leucins wurde angenommen, daß das 14CLeucin in den Zellen nur unwesentlich durch 12C-Leucin verdünnt wird, und daß weiterhin der für das EMCVirus gefundene Wert von 8,7 g Leucin/100 g Virus­ protein 2 auch für das ME-Virus gültig ist. Legt man diese Annahmen zugrunde, so ergibt sich aus den Meßwerten (s. Tab. 1 ), daß mindestens 1 bis 3'10° Viruspartikel von der einzelnen L-Zelle synthetisiert werden. Da nach den Ergebnissen der I. Mitteilung (s. dort Legende Abb. 1) jede Zelle 50 —200 PBE bildet, sind etwa 1000 polyederartige Viruspartikel für die Bildung eines Plaque erforder­ lich. Danach verhält sich das in L-Zellen produzierte ME-Virus in dieser Hinsicht ähnlich wie das in Mäusegehirnen gebildete. Experi­ ment Nr. I y-Leucin im hämads. Virus aus 1 ml Zellsuspension II Viruspart./ml Zellsuspension [• 10u ] III Viruspart./ Zelle 1 2 3 4 5 0,30 0,30 0.15 0,29 0,33 5,2 5,2 2,6 5,0 5,7 2,6 • 105 2,6 • 105 1,3-105 2,5 • 105 2,9 • 105 1029 lichkeit und Genauigkeit durchzuführen. Deshalb wurden sowohl die Synthese des Virusproteins als auch das Auftreten der Viruspartikel unter Verwen­ dung von 14C-Leucin ermittelt. Eine Suspension infizierter L-Zellen in Leucin-freiem Kulturmedium wurde in zwei Teile geteilt. Zu einer Hälfte wurde sofort nach der Infektion (Zeit 0 ) 14CLeucin (5 juClmg; 0,02 mg/ml) zugesetzt und davon stündlich Proben von 10 ml entnommen. Um den Ein­ bau von weiterem 14C-Leucin zu verhindern, wurden diese Proben sofort bei —70" eingefroren (Reihe I; Viruspartikel). Aus der anderen Hälfte der infizierten Kultur wurden jede Stde. 10 ml einer Zellsuspension ent­ nommen und diese dann in einem besonderen Gefäß mit 14C-Leucin inkubiert. Diese Proben wurden erst 8 Stdn. p.i. eingefroren (Reihe II; Virusprotein) . Später wur­ den die Zellen in allen Proben durch 3-maliges Einfrie­ ren und Auftauen lysiert. Nachdem Material für die HA- und Plaque-Teste sowie für die Bestimmung der Radioaktivität im Gesamtprotein entnommen war, wurdas Lysat 1-mal mit Frigen extrahiert und dann mit Ery­ throcyten behandelt. Die Radioaktivität der an die Blut­ zellen adsorbierten Viruspartikel wurde im Ge i g e r M ü l l e r - Zähler gemessen. In Abb. 2 ist das Ergebnis dieses Versuches zu­ sammenfassend graphisch dargestellt. Die zur Kon­ struktion der Kurven dienenden Werte sind in Tab. 2 enthalten. Die Werte für das Virusprotein wurden aus den Differenzen der Proben 0 —8 und X — 8 (Reihe II) errechnet. Aus dem Vergleich der Kurven geht hervor, daß das Virusprotein 1 —2 Stdn. vor dem Auftreten der polyederartigen Viruspartikel synthetisiert wird. Tab. 1. Ermittlung der Anzahl der in L-Zellen gebildeten Viruspartikel. Die Werte in Spalte I wurden aus der Radio­ aktivität des in die Viruspartikel eingebauten 14C-Leucins be­ stimmt. Zur Berechnung der Anzahl der Viruspartikel (II) wurde ein Verhältnis von 8,7 g Leucin/100 g Virusprotein 2 angenommen. Die Zellkonzentration in diesen Versuchen be­ trug 2 -IO6 Zellen/ml (s. Methodik d. 1. Mitteilung). 3. Zeitlicher Verlauf der Synthese des Virusproteins In den folgenden Experimenten sollte der Verlauf der Bildung desjenigen Proteins, das später in den Virusteilchen erscheint, mit dem Auftreten der poly­ ederartigen Viruspartikel verglichen werden. Letzte­ res läßt sich leicht mit Hilfe der HA-Reaktion und des Plaque-Tests verfolgen. Es war aber wünschens­ wert, den Vergleich mit Methoden gleicher Empfind2 P. T. E. M . M a r t i n , S. S ved , R. C. Biochem. J. 80, 597 [1961]. F a u lk n e r , S. W o r k , V a le n tin e u. Abb. 2. Bestimmung des zeitlichen Verlaufs der Synthese des Virusproteins und der Viruspartikel in L-Zellen. (Zusammen­ fassung der Daten aus Tab. 2.) In Abb. 2 ist außerdem noch die Kurve, die den Einbau von 14C-Leucin in das Gesamtprotein der Zelle wiedergibt, eingetragen. Angegeben sind die Meßwerte aus den beiden angewendeten Markie­ rungsverfahren (0 —X und X — 8 Stdn. p.i.). Die Kurve zeigt, daß die Rate der Gesamtprotein-Synthese in Abhängigkeit von der Zeit nach der Infek- Unauthenticated Download Date | 2/14/17 3:17 AM P. HAUSEN 1030 Zeit X (Stdn. p. i.) 0 2 3 4 5 6 7 8 Imp./Min. im hämads. Virus 0 0 0 7,5 60 120 240 242 H AE/m l PBE/m l 160 640 1280 1280 2560 5,0 • 103 2,9 • 104 5,6 • 104 7,4 • 106 3,2 • 107 5,5 • 107 1.1 • 10« 1,0 • 108 a) Bildung der polyederartigen Viruspartikel (Reihe I; Markierung von Zeit 0 bis Zeit X ) . Zeit X (Stdn. p. i.) 0 2 3 4 5 6 7 8 Imp./Min. im hämads. Virus 294 340 235 161 96.5 28.2 28,2 — HAE/ml 2560 2560 2560 1280 2560 2560 2560 2560 b) Bildung des Virusproteins (Reihe II; Markierung von Zeit X bis 8 Stdn. p.i.). Tab. 2. Ermittlung des zeitlichen Verlaufs der Bildung der Viruspartikel (Reihe I) und des Virusproteins (Reihe II) durch Markierung mit 14C-Leucin. tion geringer wird. Der Einfluß der Virusinfektion auf die Gesamtprotein-Synthese wurde im folgenden Experiment noch näher untersucht. 4. Einfluß der ME-lnfektion auf die Gesamtprotein- dann aber scharf ab. Zwischen 3 und 6 Stdn. p.i., d. h. zu der Zeit, zu der das Virusprotein gebildet wird (s. Abb. 3), wurde bei allen Versuchen dieser Art ein Abflachen der entsprechenden Kurve beob­ achtet. Synthese der L-Zellen Von einer Zellsuspension wurde die eine Hälfte mit Virus infiziert, die andere mit einem Extrakt aus nor­ malem Mäusegehirn, der in gleicher Weise wie das Impfvirus hergestellt war, behandelt. Die Zellen wur­ den danach gewaschen und im Gewebekulturmedium bei 37 cC inkubiert. Zu verschiedenen Zeiten wurden der Zellsuspension jeweils 2 ml entnommen und mit 14C-Leucin (5//C/mg; 0,02 mg/ml) weitere 15 Min. in­ kubiert. Nach dieser Zeit wurde das Protein mit 10-proz. Trichloressigsäure gefällt und seine Radioaktivität be­ stimmt. In Abb. 3 ist der Einbau von 14C-Leucin in das Protein infizierter Zellen als Prozent des Einbaues bei nicht-infizierten Kontrollzellen dargestellt. Kurz nach der Infektion steigt der Einbau etwas an, fällt Besprechung der Ergebnisse Die in der vorliegenden Arbeit beschriebenen Ex­ perimente deuten darauf hin, daß die Synthese des Proteins des ME-Virus etwa 1^2 Stdn. vor dem Auf­ treten der polyederartigen Virusteilchen einsetzt. Das Ergebnis des in dieser Hinsicht wichtigen Versuches (s. Abb. 3) kann nicht durch „Pool“-Effekte bestimmt worden sein. Vorversuche hatten gezeigt, daß bei den L-Zellen 14C-Leucin ohne Verzögerung aufge­ nommen und eingebaut wird, und daß deshalb ein erheblicher intrazellulärer Vorrat an freiem Leucin nicht existieren kann. Ein „Pool-Effekt“ würde sich nicht nur auf die Virusprotein-, sondern auch auf die Gesamtprotein-Synthese auswirken. Das ist aber nach Abb. 3 nicht der Fall. Die Gesamtprotein-Syn­ these zeigt bei den zwei verschiedenen Markierungs­ verfahren nahezu den gleichen Verlauf. Etwa zur gleichen Zeit, zu der nach den Markie­ rungs-Experimenten das Virusprotein synthetisiert wird, ist in den infizierten Zellen ein nichthämadsorbierbares, lösliches Antigen nachzuweisen (s. I. M itt.1). Es liegt nahe anzunehmen, daß dieses Antigen mit dem vor den polyederartigen Viruspar­ tikeln erscheinenden Virusprotein identisch ist. 1 2 3 1 5 6 7 Stdn. p .i — 8 ► Abb. 3. Veränderung der Proteinsynthese-Rate in infizierten L-Zellen. Es sind 2 Experimente eingetragen ( x; • ) . Faßt man diese und frühere Befunde zusammen, so ergibt sich folgendes Bild von der Produktion des ME-Virus: Zunächst werden Virus-RNS und-Protein gebildet. Liegen ausreichende Mengen von diesen Unauthenticated Download Date | 2/14/17 3:17 AM VERMEHRUNG DES ME-VIRUS IN L-ZELLEN II 1031 Viruskomponenten vor, so treten sie zu den poly­ ederartigen Viruspartikeln zusammen. In diesem Augenblick gewinnt das Virusprotein seine häm­ adsorbierende bzw. hämagglutinierende Fähigkeit, wahrscheinlich durch Änderung seiner elektrostati­ schen Eigenschaften. Für diese Vorstellung spricht weiterhin die Feststellung, daß die Viruspartikel schon unter schonenden Bedingungen im schwach sauren Milieu in Untereinheiten zerfallen, die unge­ fähr die Größe des löslichen Antigens besitzen und im Komplementbindungs-Test, nicht aber im Hämagglutinations-Test, positiv reagieren. Ebenso wie beim ME-Virus wird auch beim Polio­ myelitis- 3 und EMC-Virus4 zunächst Virusprotein synthetisiert, ehe die reifen Viruspartikel erscheinen. Der zeitliche Abstand ist aber hier geringer als beim ME-Virus. Womöglich beruht das darauf, daß bei diesen Virusstämmen die einzelnen Prozesse besser koordiniert sind als bei unserem Modell-Virus. Dies könnte auch die Ursache dafür sein, daß beim MEVirus in relativ großer Menge nicht-infektiöse, „feh­ lerhafte“ Viruspartikel entstehen. Auch im Hinblick auf die Hemmung der Gesamtprotein-Synthese bestehen zwischen Polio- 5, EMC- 6 und ME-Virus weitgehende Parallelen. Unsere Ver­ suche lassen erkennen, daß im Zeitraum von 3 bis 6 Stdn. nach der Infektion die unter der Wirkung des Virus einsetzende Hemmung der GesamtproteinSynthese durch einen gegenläufigen Prozeß beein­ flußt wird. Die zeitlichen Verhältnisse lassen vermu­ ten, daß es sich dabei um die Synthese des Virus­ proteins handelt. Die Hemmung der GesamtproteinSynthese im infizierten System läuft annähernd parallel mit der Hemmung der RNS-Synthese im Zellkern7, die früher auf autoradiographischem Wege festgestellt wurde. Damit ist aber noch nicht erwiesen, daß zwischen beiden Prozessen ein unmit­ telbarer Zusammenhang besteht. Untersuchungen, die sich mit diesem Problem befassen, werden Ge­ genstand einer weiteren Mitteilung sein 8. 3 J . E. D a rn ell [ I9 6 0 ] . 2 3 5, 74 6 Virology 19, 8 u. L . L evintow , J . biol. Chemistry 4 E. M. M a r t i n , persönliche Mitteilung. 5 E. F. Z im m e r m a n n , M . H e e t e r u . J. E. Ich bin Herrn Professor Dr. W . S c h ä f e r für seine Unterstützung und anregende Diskussionen zu diesen Arbeiten zu großem Dank verpflichtet. Herrn U. W a n ­ d e l danke ich für seine Mitarbeit. Die Arbeiten wurden von der D e u t s c h e n F o r ­ s c h u n g s g e m e i n s c h a f t gefördert. 7 D a r n e ll, E. M . M a r t i n , J. M a l e c , S. S ved u . T. S. W o r k , Biochem. J. 80, 585 [1961]. H . H a u s e n , Z. Naturforschg. 17 b , 158 [1962]. P. H a u s e n u . D. W . V e r u w o e r d , Virology, im Druck. 400 [1963]. Unauthenticated Download Date | 2/14/17 3:17 AM