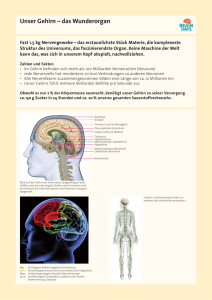
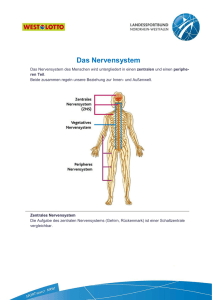
Ein Fenster zum ICH - Teil 1
Werbung

Ein Fenster zum ICH - Teil 1 Grundbegriffe der Psychologie. Ein Lehrbuch in fünf Teilen von Herbert Paukert. Version 9.0 © 2014. Inhalt von Teil 1 Biologische Grundlagen, Modell der Psyche, Methoden der Gehirnforschung. Inhalt von Teil 2 Psychische Funktionen: Wahrnehmung, Gedächtnis, Denken, Sprechen. Psychische Kräfte: Instinkte, Triebe, Interessen, Gefühle und Wille. Inhalt von Teil 3 Psychologische Tests, Intelligenz, Persönlichkeit. Inhalt von Teil 4 Psychoanalyse, Lerntheorie, sozial-kognitive Theorie, Entwicklungspsychologie und Sozialpsychologie. Inhalt von Teil 5 Stress, Psycho-Neuro-Immunologie, Selbstheilung, Psychosomatik, Psychopathologie und Psychotherapie. Hinweis: Das hier vorliegende Lehrbuch ist eine Neufassung des gleichnamigen Buches, welches 1998 erstmalig im öbv/hpt-Verlag erschienen ist. Dort befinden sich auch die Quellen-Nachweise. Paukert: Ein Fenster zum Ich – Teil 1 2 Inhalt von Teil 1: Vorwort (03) [1.1] Biologische Grundlagen (04) [1.1.1] Das biologische System "Mensch" [1.1.2] Das Nervensystem und seine Bausteine [1.1.3] Gliederung des zentralen Nervensystems [1.1.4] Gehirn und Großhirn [1.1.5] Sensorisches und motorisches System [1.1.6] Das vegetative Nervensystem [1.1.7] Das Hormonsystem [1.1.8] Informationsflüsse im Nervensystem [1.1.9] Der Weg zum Bewusstsein [1.1.10] Das Erkenntnisproblem "Bewusstsein" (04) (17) (27) (32) (37) (40) (42) (47) (49) (51) [1.2] Ein Basismodell der Psyche (58) [1.3] Methoden der Gehirnforschung (61) [1.3.1] Stereotaktische Eingriffe [1.3.2] Enzephalogramme [1.3.3] Bildgebende Verfahren (61) (62) (64) Paukert: Ein Fenster zum Ich – Teil 1 3 Vorwort Dieses Buch unternimmt den Versuch, das Phänomen der menschlichen Psyche im Rahmen einer allgemeinen, naturwissenschaftlich orientierten Psychologie zu beschreiben. Dabei werden auch wichtige Gebiete an den Schnittstellen zu Biologie, Medizin und Philosophie erörtert. Der erste Teil vermittelt die erkenntnistheoretische Position der Naturwissenschaften und die allgemeine, biologische Basis des menschlichen Organismus. Es werden die wichtigsten Grundlagen des peripheren und zentralen Nervensystems, des vegetativen Nervensystems, des Hormonsystems und des Immunsystems beschrieben. Im zweiten Teil erfolgt eine ausführliche Darstellung der psychischen Leistungen des menschlichen Gehirns (Wahrnehmen, Erinnern, Lernen, Denken, Sprechen und Instinkte, Triebe, Interessen, Gefühle, Wille). Im dritten Teil werden die Grundlagen des Messens und Testens und der statistischen Datenauswertung erklärt. Eine ordentliche Versuchsplanung und die Verwendung statistischer Hilfsmittel sind unerlässliche Voraussetzungen für jede wissenschaftliche Arbeit. In diesem Abschnitt erfolgt auch eine detaillierte Beschreibung von Persönlichkeit und Intelligenz. Im vierten Teil werden psychodynamische Entwicklungsmodelle und die Lerntheorie beschrieben. Ein eigener Abschnitt ist verschiedenen sozialpsychologischen, insbesonders gruppendynamischen Phänomenen gewidmet. Der fünfteTeil beginnt mit den Themen „Stress“, „Psychoimmunologie“, „Selbstheilung“ und „Psychosomatik“ und bringt dann eine Übersicht über die wichtigsten psychopathologische Erscheinungsformen und über verschiedene Schulen der Psychotherapie. Unser Wissen gleicht einer Kugel im Raum. Ihre Oberfläche ist die Grenze zum Unbekannten und Unerforschten. Mit jeder Erweiterung unserer Wissenskugel, vergrößert sich auch diese Grenzfläche. So ergibt sich der paradoxe Sachverhalt, dass je mehr wir zu wissen vermeinen, wir umso schmerzlicher erkennen müssen, wie wenig wir eigentlich wissen. Dieses Bild einer Wissenskugel erlaubt aber noch eine zweite Interpretation. Nicht die wechselhaften Erscheinungen an der vordergründigen Oberfläche sind von Bedeutung, sondern die hintergründigen, verborgenen Strukturgesetze. Erst das tiefere Verständnis der versteckten Beziehungen und Bindungen ermöglicht dem Schachspieler die richtige Einschätzung der sichtbaren Figurenstellung am Spielbrett. Genauso kann das Wissen um die Gesetzmäßigkeiten der allgemeinen Biologie und Psychologie zu einem besseren Verständnis und einer differenzierteren Beurteilung menschlicher (Lebens-) Situationen führen. Vielleicht kann dadurch auch unser Verhalten am Spielbrett des Lebens in günstiger Art und Weise beeinflusst werden. Der Autor hofft, mit diesem Buch dem interessierten Leser einen möglichst umfassenden Einblick in die Hintergründe der menschlichen Psyche zu vermitteln. Paukert: Ein Fenster zum Ich – Teil 1 4 [1.1] BIOLOGISCHE GRUNDLAGEN Die Psyche ist eine Systemfunktion des Gehirns. Sie ist das Resultat informationsverarbeitender Prozesse in komplexen Netzwerken aus Milliarden von Nervenzellen (Neuronen). Durch Interaktion des Gehirns mit seiner Umwelt hat sich die menschliche Psyche evolutionär entwickelt - von einfachen Reflexen für die Reizbeantwortung über unbewusste instinktive Handlungen für die Lebenserhaltung bis zu den begriffsbildenden neuronalen Netzen in der Großhirnrinde, wodurch Denken als Probehandeln möglich wird. Das bewusste Denken erzeugt im Gehirn ein Modell der Umwelt. Mit Hilfe der Sprache kann dieses Wissen an andere Gehirne weitergegeben und auch auf materielle Datenträger gespeichert werden. Dadurch wird das subjektive Wissen zum objektiven Geist und zur Grundlage der Kultur. Die höchste Entwicklung ist die Selbstwahrnehmung (Reflexion) des Gehirns, d.h. der Mensch konstruiert ein Modell von sich SELBST (Ichbewusstsein). [1.1.1] Das biologische System "Mensch" Die Psyche, als Ergebnis informationsverarbeitender Prozesse im zentralen Nervensystem, unterliegt der biologischen Evolution. Die Psyche kann ohne Biologie nicht hinreichend erfasst werden. Die Biologie ihrerseits stützt sich vor allem auf die Chemie. Ein tieferes Verständnis für das System "Mensch" ist nur möglich auf dem Wissen um die grundlegenden chemischen Vorgänge in den Zellen, im Organismus und in unserer Umwelt. Zur Erklärung vieler biologischer Prozesse sind die Begriffe Information und Regulation von zentraler Bedeutung. Die Ereigniskette A, B, C, D, in welcher das Ereignis A das Ereignis B verursacht, B dann C, und C dann D verursacht, ist ein lineares, determiniertes System. Wenn aber D auf A zurückwirken kann (Rückkoppelung, feed back), dann ist das System zirkulär und hat die Möglichkeit der Selbstregulation. In solchen Regelkreisen werden reale Ist-Werte durch Messfühler gemessen und mit einprogrammierten Soll-Werten verglichen. Bei Abweichungen werden Regelungsprozesse ausgeführt, welche die Ist-Werte an die Soll-Werte anpassen. Damit können lebenswichtige Gleichgewichtszustände (Homöostasen) erhalten werden. Das vorliegende Buch versucht der Verwurzelung der Psychologie in Biologie und Chemie Rechnung zu tragen und beschreibt daher im ersten Teil wichtige biologische Grundlagen. Der daran nicht interessierte Leser kann diesen Teil des Buches durchaus überspringen. Paukert: Ein Fenster zum Ich – Teil 1 5 Die Entwicklung des Lebens Die unbelebte Welt ist der Bereich von Zeit und Raum, in dem Anhäufungen von Materie vorkommen, welche ihrerseits aus Atomen und Molekülen bestehen. Zwischen den Materieteilchen wirken Kräfte, die durch Physik und Chemie beschrieben werden. Durch die gegenseitigen Wechselwirkungen verschiedener Moleküle bilden sich immer komplexere Stoffklassen. So bildeten sich in der Uratmosphäre aus einfachen Molekülen wie Wasser (H2O), Methan (CH4) und Ammoniak (NH3) bei Energiezufuhr (Sonne, Blitz) neue Moleküle wie Aminosäuren (z.B. Glycin H2N-CH2-COOH). Diese wurden in die mineralsalzreichen Urozeane geschwemmt und sie bildeten dort noch komplexere Moleküle; z.B. werden aus den einfachen Aminosäuren längere Polypeptidketten, die sich zu Proteinen (Eiweißen) zusammenfalten. Eine andere Klasse sind die so genannten Phospholipide (komplexere Verbindungen aus Fettsäuren, Glycerin-Alkohol und Phosphorsäure), welche aus einem wasserabstoßenden und einem wasseranziehenden Ende bestehen. Sie ordnen sich im Wasser kugelförmig an und bilden dort doppelschichtige Membrane. Durch Einlagerung von Polypeptiden in diese Membrane können Kanäle erzeugt werden, welche die Verbindung zwischen Außen und Innen herstellen. Damit ist die "Pore" als erstes Organ der Evolution erfunden. Im Innern dieser Gebilde sammeln sich auf Grund einseitiger Membrandurchlässigkeit bestimmte Makromoleküle gehäuft an, die in enge Wechselwirkung miteinander treten können. Solche, in der Ursuppe schwimmende Gebilde (Präzellen) sind die Vorläufer der lebendigen Zellen. In den Präzellen entwickelt sich über die aus Vulkanen stammenden Phosphate ein einfacher Stoffwechsel. Anorganische Polyphosphate sind zwar stabil gegen Wasser, aber instabil gegen kohlenstoffhaltige Hydroxylgruppen. Sie verbinden sich solcherart leicht mit anderen geeigneten Stoffen zu reaktionsfreudigen Molekülen. Die nunmehr organischen Polyphosphatverbindungen beteiligen sich rege am Aufbau weiterer Makromoleküle, unter anderem auch an der Synthese von Nukleinsäuren. Die verdrillten Doppelstränge der Desoxyribonukleinsäure (DNS) bestehen aus Desoxy-Ribose-Zucker, Phosphatresten und genau vier stickstoffhaltigen Basen Adenin, Thymin, Guanin und Cytosin. Aus elektrochemischen Gründen können nur bestimmte basische Stoffe einander gegenüber liegen (komplementäre Basenpaarung: A-T und G-C). Wird durch äußere Einflüsse ein solcher DNS-Doppelstrang aufgetrennt, so können sich an die entsprechenden Basen eines Einzelstranges frei herumschwimmende, komplementäre Bruchstücke anlagern. Damit bildet sich wieder ein neuer DNS-Doppelstrang, der identisch mit dem Ausgangsmolekül ist. Durch diese identische Reduplikation sind solche Makromoleküle in der Lage, gleichartig aufgebaute Tochtermoleküle zu erzeugen. Paukert: Ein Fenster zum Ich – Teil 1 6 Die Abbildung zeigt die Reduplikation eines DNS-Doppelstranges: Aus dem bereits komplexen chemischen Geschehen in der Präzelle entwickelt sich eine neue Struktur, nämlich die Zelle. Diese weist nunmehr völlig neue Funktionsmerkmale auf: einen Energiestoffwechsel, die identische Reduplikation und einen Baustoffwechsel. Für diese Arbeitsleistungen haben sich auch verschiedene Unterstrukturen innerhalb der Zelle gebildet, die in Wechselwirkung zueinander stehen (Kern, Plasma, spezialisierte Zellorganellen, Membran). Die einzelnen Vorgänge in den verschiedenen Zellabteilungen (Zellorganellen) regulieren sich gegenseitig derart, dass immer eine optimale Anpassung an die jeweiligen chemisch-physikalischen Situationen erfolgt. Die im Vergleich zur unbelebten Präzelle neuen strukturellen und funktionellen Systemmerkmale werden unter dem Sammelbegriff "Leben" zusammengefasst. Dadurch unterscheidet sich die belebte von der unbelebten Materie. Natürlich sind die Übergänge fließend und kontinuierlich, z.B. bei den Viren. In weiterer Folge entstehen dann kleinere Mikroorganismen und schließlich höhere Lebensformen mit spezialisierten Zellverbänden (Organen). Das, der stammesgeschichtlichen Entwicklung (Phylogenese) zu Grunde liegende Prinzip formulierte Charles Darwin (1850) in seiner Evolutionstheorie, wonach nur jene Strukturen überleben, welche am besten an die jeweiligen Umweltbedingungen (den Außenraum) angepasst sind. Der Motor unserer Stammesgeschichte sind die Mutationen (Veränderungen des genetischen Codes) und die Selektionen (natürliche Auslesen). Paukert: Ein Fenster zum Ich – Teil 1 7 Die wichtigste Funktion der DNS-Moleküle ist ihre steuernde Mitwirkung bei der Bildung von Eiweißen (Proteinen) durch gezielte Verknüpfung von Aminosäuren. Die DNS-Doppelstränge befinden sich in 46 Kernschleifen (Chromosomen) der Zellkerne. Beim Vorgang der Transkription werden sie ab einer Startposition bis zu einer Endposition mithilfe bestimmter Enzyme aufgetrennt. Von diesem Abschnitt wird auf Grund der komplementären Basenpaarung aus Nukleotidstücken eine Kopie erzeugt, die Messenger-Ribonukleinsäure (mRNS). Die RNS (Ribonukleinsäure) unterscheidet sich von der DNS (Desoxyribonukleinsäure) in ihrer Struktur nur geringfügig. Dann schließt sich der DNS-Doppelstrang wieder und die mRNS wandert aus dem Zellkern in das Zellplasma. Dort erfolgt an eigenen Einrichtungen (Ribosomen) der Vorgang der Translation. Dabei steuern jeweils drei Basen der mRNS (Basentriplett, Codon) die Anheftung einer bestimmten Aminosäure. Der eigentliche Aminosäuretransport erfolgt mithilfe der TransferRNS (tRNS), die auf der einen Seite über komplementäre Basenpaarungen an die mRNS andockt und auf der anderen Seite mit einer Aminosäure beladen ist. In der Basensequenz der Nukleinsäuren ist also die Information für die Verknüpfung der verschiedenen Aminosäuren zu Eiweißmolekülen (Proteinen) verschlüsselt. In diesem Zusammenhang spricht man auch vom genetischen Code. Unter einem Gen versteht man einen Abschnitt des DNS-Moleküls, also eine bestimmte Sequenz von Basentripletts, welche für die Biosynthese eines Proteins verantwortlich ist. Transkription und Translation sind biochemische Vorgänge, die durchaus mit komplizierten feinmechanischen Bearbeitungsmaschinen vergleichbar sind. Paukert: Ein Fenster zum Ich – Teil 1 8 Die Proteine ihrerseits werden als Baustoffe oder als Enzyme (Vermittlerstoffe, Katalysatoren) für weitere chemische Reaktionen verwendet. Die Enzym-Proteine bestehen aus langen Ketten von Aminosäuren, welche sich durch Faltungen zu komplizierten räumlichen Strukturen anordnen. Dabei bilden die für die spezifische Enzymleistung wirksamen Aminosäuren eine Vertiefung in der Oberfläche. An dieses aktive Zentrum des Enzym-Proteins wird dann das passende Substratmolekül angelagert. Nun können dort bestimmte Ionen (Coenzyme) auf das Substratmolekül einwirken, sodass dieses beispielsweise chemisch verändert oder überhaupt zerlegt wird. Die Enzyme wirken so als Biokatalysatoren auf die Stoffwechselvorgänge in der Zelle, wodurch bestimmte Merkmalsstrukturen aufgebaut werden. Die Gene enthalten somit die Erbinformationen für die Entwicklung äußerer Bau- und Leistungsmerkmale des Körpers (Phäne). Die gesamte genetische Steuerung der Biosynthese von Proteinen heißt Genexpression. Eine wesentliche Leistung der Zellen besteht in ihrer Fähigkeit sich in zwei Tochterzellen zu teilen. Der zentrale Mechanismus ist dabei die identische Reduplikation der DNS, wodurch die Erbinformation weitergegeben wird. Durch die Zellteilung wird erst Wachstum und Regeneration (Ersatz von Zellen) möglich. Zellverbände, welche aus überwiegend gleichartig strukturierten Zellen mit bestimmten Funktionen bestehen, werden als Gewebe bezeichnet. Man unterscheidet Epithelgewebe (Oberflächenschutz), Binde- und Stützgewebe, Muskelgewebe und Nervengewebe. Ein interessantes Phänomen in der Entwicklung eines Individuums (Ontogenese) ist die so genannte Zelldifferenzierung. Sämtliche Baupläne eines Lebewesens sind in den DNS-Strängen im Zellkern enthalten. Die Ursachen für die Spezialisierung der Zellen liegen einerseits in der Genaktivtät und andererseits in der Zellaktivität. Die so genannte Genregulation bewirkt, dass nur bestimmte Gene aktiv werden und die Biosynthese spezifischer Proteine steuern. Zur Genregulation werden verschiedene Verfahren angewendet, z.B. die Substratinduktion, welche als echter Regelkreismechanismus angesehen werden kann. Eigene Regulationsgene erzeugen bestimmte Repressorproteine. Diese blockieren die Startposition eines Gens. Das die Aktivität dieses Gens auslösende Substrat heftet sich an das Repressormolekül, wodurch sich dieses räumlich umlagert und seine blockierende Bindung zum Gen verliert. So können bestimmte Enzyme (RNSPolymerasen) angreifen und die Transkription starten und kontrollieren. Das wiederum führt im Zellplasma zur Biosynthese des entsprechenden EnzymProteins. Falls dieses das Substrat spaltet, so sinkt im Plasma die Konzentration des Substrates. Der nunmehr wieder unbeladene Repressor kann den Genabschnitt neuerlich blockieren und die Biosynthese wird eingestellt. Neben dieser Genregulation wirken auch die Aktivitäten der Zellen als Ganzes steuernd auf die Spezialisierung der Zellen. Eine Zelle kann sich bewegen, sich teilen, sich an andere anheften (Adhäsion), loslassen und auch sterben. Paukert: Ein Fenster zum Ich – Teil 1 9 Bei der Befruchtung verschmelzen Eizelle und Samenzelle und damit auch ihre genetischen Informationen. Durch fortwährende Zellteilung bildet sich daraufhin das Blastoderm, ein Zellhaufen mit über 100 000 Zellen. Durch Wanderung und Adhäsion der Zellen kommt es zur Einstülpung des Blastoderms, zur Gastrulation, wobei sich drei Zellplatten herausbilden (Keimblätter): Ektoderm, Entoderm und Mesoderm. In diesem Stadium der Keimesentwicklung setzt nun der Mechanismus der embryonalen Induktion ein. Darunter versteht man die Koordination der ortsabhängigen Differenzierung der Zellen (Topobiologie). Diese erfolgt durch Zellaktivitäten und durch Genregulationen, wobei Signalstoffe erzeugt werden, die ihrerseits wiederum auf andere Zellen einwirken können. Durch diese Prozesse entwickeln sich aus den Keimblättern die Primitivorgane. Am Ende der Entwicklung steht das individuelle Lebewesen mit seinen Organsystemen. Das Altern ist ein unausweichlicher Prozess, dem jedes Lebewesen unterworfen ist. Zwei große Theorien versuchen diesen Vorgang zu erklären: die Fehler- und die Programmtheorie. Für die Vertreter der Fehlertheorie ist das Altern das unvermeidbare Resultat des Verschleißes von Zellen und ihrer Erbsubstanz auf Grund schädigender Einflüsse, denen sie während ihres Lebens ausgesetzt sind. Der kritische Punkt ist dann erreicht, wenn die zelleigenen Reparaturmechanismen nicht mehr ausreichen, um die Fehler zu beheben. Die Befürworter der Programmtheorie sind überzeugt, dass Altern und Tod ein ureigener Teil des Lebens sind: Von Anfang an in jeder Zelle installiert, läuft ein genetisches Alterungsprogramm nach einem arttypischen Muster ab. Unstrittig jedenfalls ist, dass die Zellen altern - ob nun auf Grund sich häufender Fehler oder eines vorprogrammierten genetischen Mechanismus. Eine wichtige biologische Uhr, die das individuelle Lebensalter mitbestimmt, ist der chemische Stoffwechsel. Der amerikanische Mediziner Richard Weindruch in Wisconsin ernährte Mäuse und Ratten nur mit zwei Drittel der normalen täglichen Kalorienzufuhr, jedoch mit allen notwendigen Vitaminen und Spurenelementen. Weil in den Zellen dieser Tiere weniger Nahrung verarbeitet wird, bilden sich auch weniger schädliche Stoffwechselprodukte, wie beispielsweise die freien Radikale (endogene Oxidantien). Diese hochreaktiven, aggressiven Moleküle sind verantwortlich für Oxidationsprozesse, welche Membranproteine und Enzymproteine, die Energie liefernden Mitochondrien im Zellplasma, und sogar Genmaterial im Zellkern zerstören können. Glücklicherweise hat die Natur Schutzmittel dagegen entwickelt: Einerseits wurden Gene entdeckt, welche die DNS-Schäden wieder reparieren bzw. ein Enzym produzieren, welches die freien Radikale abbaut (Superoxid-Dismutase). Andererseits hemmt auch eine gesunde Ernährung, die so genannte Antioxidantien enthält (z.B. in Karfiol und Broccoli), die zerstörerische Wirkung der freien Radikale. Allgemein erweist sich die Stoffwechselgeschwindigkeit als indirekt proportional zur Lebensdauer. Langsame und schlanke Tiere leben länger, bei Ratten bis zu einem Drittel der normalen Lebensspanne. Paukert: Ein Fenster zum Ich – Teil 1 Das biologische System "Mensch" Nervensystem: Kommunikation mit Umwelt und Systemkontrolle. Magen, Darm: Aufnahme der Nahrungsstoffe. Lunge: Einatmen von Sauerstoff und Ausatmen von Kohlendioxid. Blutkreislauf, Herz: Transport der Stoffe im Organismus. Leber: Abbau, Umbau, Aufbau von Stoffen und Entgiftung. Darm, Niere, Lunge, Haut: Ausscheiden von Stoffen. 10 Paukert: Ein Fenster zum Ich – Teil 1 11 Die Bausteine des Organismus sind die Zellen. Diese haben im Laufe der Entwicklung hoch spezialisierte Funktionen übernommen (Muskelzellen zur Bewegung, Nervenzellen zur Signalübertragung, Drüsenzellen zur Sekretion, usw.). In der Grundstruktur bestehen sie aus einer Zellmembran, die sie von der Umgebung abgrenzt, und im Inneren aus dem Zellplasma mit verschiedenen Einrichtungen (Ribosomen, Mitochondrien usw.) und dem Zellkern mit seinen Chromosomen, die aus Desoxyribo-Nukleinsäure-Molekülen (DNS) bestehen. Im Zellplasma läuft der biochemische Stoffwechsel ab. Beim so genannten Baustoffwechsel werden Moleküle zerlegt und aus ihren Bestandteilen wieder neue aufgebaut. Im Grunde sind nur wenige Stoffklassen für den Aufbau der belebten Natur wichtig: Wasser, Salze bzw. Ionen, Kohlehydrate, Fette bzw. Lipide, Proteine (Eiweiße), Ribonukleinsäuren (RNS) und Desoxyribonukleinsäuren (DNS). Zerlegt man diese Moleküle weiter, dann erhält man ebenfalls nur wenige typische Bestandteile wie die Aminosäuren der Eiweiße oder die Fettsäuren der Fette oder den Traubenzucker der Kohlehydrate. Damit der Baustoffwechsel reibungslos funktioniert, muss ihm Energie zugeführt werden. Diese Energie liefert der Betriebsstoffwechsel (Energiestoffwechsel) der Zelle. Die Aufnahme der Betriebsstoffe (vor allem von Zucker aus dem Blut) erfolgt durch die Zellmembran. Ihre Verbrennung mit Sauerstoff in den Mitochondrien des Zellplasmas wird als biologische Oxidation bezeichnet und liefert erstens weiter verwertbare chemische Bestandteile und zweitens freiwerdende Energie, welche zur Synthese von Adenosintriphosphat (ATP) aus Adenosindiphosphat (ADP) und Phosphorsäure (P) verwendet wird. Die biologische Oxidation besteht aus einer mehrstufigen Kette von vielen Einzelreaktionen, wo verschiedene Enzyme mitwirken. Glykolyse (Zuckerabbau zur einfacheren Brenztraubensäure), Oxidationen der Fettsäuren und Aminosäurenabbau münden in den so genannten Zitronensäurezyklus, wo eine stufenweise Umformung und Zerlegung von kohlenstoffhaltigen Säuren unter Abspaltung von Kohlendioxid CO2 erfolgt. In der Atmungskette schließlich kommt es zum Endabbau mit Hilfe von Sauerstoff unter Abspaltung von Wasser H2O. Ein zentrales Zwischenprodukt dabei ist die aktivierte Essigsäure (Acetyl- Coenzym-A), welche einerseits als Ausgangspunkt für verschiedene Molekülsynthesen dient (Fettsäuren, Transmitterstoffe, Steroidhormone, Gallensäuren usw.); andererseits wird sie mit Hilfe von Sauerstoff unter Abgabe von Energie in die Endprodukte Kohlendioxid und Wasser zerlegt, welche dann ausgeschieden werden. Die Eiweißzerlegung (Abbau der Aminosäuren) führt zu dem Endprodukt Harnstoff und der Nukleinsäureabbau liefert noch zusätzlich Harnsäure. Aufnahme, Zerlegung und Ausscheidung von Substanzen kennzeichnen den Betriebsstoffwechsel. Paukert: Ein Fenster zum Ich – Teil 1 Gemeinsame Wege des Stoffwechsels der Nahrungsstoffe: 12 Paukert: Ein Fenster zum Ich – Teil 1 Nährstoffumwandlung bei der Verdauung: Die Umgebung der Zelle: Das Innere der Zelle: 13 Paukert: Ein Fenster zum Ich – Teil 1 Der Stoffwechsel in der Zelle: Die biologische Oxidation: Die chemische Energiegewinnung: 14 Paukert: Ein Fenster zum Ich – Teil 1 15 Das ATP-Molekül als Zwischenspeicher biochemisch freigesetzter Energie kommt in vielfältiger Weise zum Einsatz. Durch eine hydrolytische Spaltung (d.h. mittels Wassermolekülen) von Adenosintriphosphat (ATP) in Adenosindiphosphat (ADP) und Phosphorsäure wird die gespeicherte Energie wieder abgegeben und für unterschiedliche Aufgaben verwendet: für den aktiven Stofftransport durch die Zellmembran (Ionenpumpe), für die Synthese von Eiweißmolekülen aus Aminosäuren, für die Bildung verschiedener Aufbaustoffe und vieles mehr. Alle diese biochemischen Reaktionen laufen unter Mitwirkung von spezifischen Katalysatoren ab. Diese Biokatalysatoren nennt man auch Enzyme, von denen über 2000 bekannt sind. Dem menschlichen Organismus liegt der komplexe Mikrokosmos seiner Zellen zu Grunde. Zellen sind offene Systeme, in denen ständig Materie umgeformt, Energie freigesetzt und gebunden wird. Dabei ist die Erhaltung stationärer Gleichgewichte der Stoffkonzentrationen lebensnotwendig, beispielsweise das Säure-Base-Gleichgewicht oder der Zuckergehalt im Blut. Die ständig auftretenden Störungen dieser Gleichgewichtszustände durch äußere Einflüsse werden durch komplizierte Regulations-Mechanismen ausgeglichen. Leben besteht daher in einer dauernden Erhaltung von Fließgleichgewichten. Der zelluläre Stoffwechsel eines einzelnen Organismus ist eingebettet in den globalen Stoffwechsel der Natur. Dieser kann als Kreisprozess aufgefasst werden: Durch die, unter Lichtenergie in den Pflanzen ablaufende Photosynthese wird Zucker (C6H12O6) aus Wasser (H2O) und Kohlendioxid (CO2) gewonnen, wobei auch Sauerstoff (O2) freigesetzt wird. Zur Lichtabsorption ist dabei der grüne Blattfarbstoff (Chlorophyll) unentbehrlich. Die Photosynthese besteht aus einer Abfolge komplizierter chemischer Reaktionen, die unter Mitwirkung mehrerer Katalysatoren abläuft. Mit der Nahrungsaufnahme gelangt der Zucker in den tierischen Organismus. Durch die biologische Oxidation wird der Zucker mit Hilfe von Sauerstoff in den Zellen der Tiere verbrannt. Die freiwerdende Energie wird im ATP-Molekül gespeichert und für die verschiedenen Formen der Zellarbeit verwendet (Nervenerregung, Muskelkontraktion, Stoffsynthese, Transport, Zellteilung usw.). Als Endprodukte des zellulären Stoffwechsels werden wiederum Wasser und Kohlendioxid und einfache Stickstoffverbindungen in die Natur ausgeschieden, und der globale Kreislauf der Energie kann von neuem mit der Photosynthese beginnen. Paukert: Ein Fenster zum Ich – Teil 1 Der Kreislauf der Energie Die Abbildung zeigt eine schematische Darstellung des energetischen Zusammenspiels von individuellem Organismus und umgebender Natur. 16 Paukert: Ein Fenster zum Ich – Teil 1 17 Eine zentrale Frage lautet: Was ist "Lebensenergie" ? Unter Energie versteht man ganz allgemein die Fähigkeit Arbeit zu verrichten. Verzichtet man auf religiöse oder mythologische Spekulationen, dann kann die Quelle unserer Lebensenergie nur in den oben beschriebenen, Energie liefernden biochemischen Prozessen liegen. Die lebende Zelle arbeitet wie eine Energie transformierende Fabrik. Die synchrone Arbeit vieler Zellen eines Gewebes bewirkt eine nach außen hin beobachtbare und messbare Aktivität. Direkt beobachtbar ist der Aktivitätszustand des Organismus vor allem in vier Bereichen: Atemtätigkeit (Frequenz, Tiefe) Herz-Kreislauf-System (Puls, Blutdruck) Peristaltik des Darmes Spannungszustand der Skelettmuskulatur Die Lebensenergie ist keine eigenständige mystische Kraft, sondern sie ist der Sammelbegriff für die synchrone Arbeit unserer Zellen. Synchronisation und Regulation erfolgen durch die Signalsysteme von Nerven- und Hormonsystem. Mit Lebensenergie wird auch oft die Selbstheilungskraft des Organismus bezeichnet. In der Menschheitsgeschichte taucht dieser Begriff der Lebensenergie in verschiedenen Formen auf: QI (China), Prana (Indien), Num (Afrika), Vis vitalis (Europa), Libido (Sigmund Freud), Orgon (Wilhelm Reich). [1.1.2] Das Nervensystem und seine Bausteine Das Nervensystem besteht aus dem somatischen und dem vegetativen System. Das somatische Nervensystem kontrolliert die Kommunikation mit der Umwelt und gliedert sich in zwei Teilbereiche: Das sensorische System besteht aus den Rezeptoren (Sinnesorganen) und den von der Peripherie zur Zentrale führenden Nerven (afferent). Es dient somit zur Aufnahme und Verarbeitung von äußeren Reizen. Das motorische System besteht aus den von der Zentrale zur Peripherie führenden Nerven (efferent) und den Effektoren (Muskeln). Es dient somit zur Steuerung der Körpermotorik. Das vegetative Nervensystem steuert und koordiniert die Funktionen der inneren Organe (Verdauung, Atmung, Herztätigkeit, usw.) Zusätzlich wird zwischen zentraler Informationsverarbeitung (Gehirn, Rückenmark) und peripherer Informationsverarbeitung unterschieden. Paukert: Ein Fenster zum Ich – Teil 1 18 Die Nervenzellen mit ihren Fortsätzen Die Bausteine des Nervensystems sind die Neuronen. Ein Neuron enthält die eigentliche Nervenzelle, viele zuleitende Fortsätze (Dendriten) und immer nur einen wegleitenden Fortsatz (Neurit, Axon). Die Nervenzelle besteht aus dem Zellkern mit seinen Chromosomen und dem Zellplasma, wo in bestimmten Teilen (Zellorganellen) wichtige chemische Stoffwechselvorgänge ablaufen. Die Kontaktstellen zwischen zwei Neuronen heißen Synapsen. Diese unterteilt man in die präsynaptische Membran, den synaptischen Spalt und die postsynaptische Membran. In der Abbildung sind im Inneren der Zelle einzelne Strukturen (Zellkern, GolgiApparat, Mitochondrien usw.) dargestellt. Die Blasen (Vesikel) an den synaptischen Endköpfen enthalten Transmitterstoffe, welche die Übertragung elektrischer Signale von einem Neuron auf ein anderes ermöglichen. Paukert: Ein Fenster zum Ich – Teil 1 19 Die Nervenzelle ist von ihrer Umgebung durch eine Membran abgegrenzt. Auf Grund einer bestimmten Verteilung von Ionen (vor allem Natrium und Kalium) besteht zwischen Innenraum und Außenraum eine elektrische Spannung (Ruhepotential, ca. -70 mV). Die Membran enthält Kanäle von verschiedener Breite, durch welche selektiv bestimmte Moleküle hinein oder hinaus wandern können. Solche Ionenverschiebungen werden durch Transmitterstoffe bewirkt. Dadurch ändert sich die elektrische Spannung zwischen Innen- und Außenraum der Zelle. Diese Spannungsänderung (Aktionspotential, ca. +30 mV) wird nun entlang der Nervenfortsätze weitergeleitet. Dann werden die Ionen wieder in umgekehrter Richtung bewegt (Ionenpumpe) und das Ruhepotential hergestellt. Die Energie für die Ionenpumpe liefert das ATP-Molekül. Den ganzen Vorgang nennt man eine elektrochemische Erregung. Mit ihren Fortsätzen und Synapsen verbundene Neuronen bilden ein lokales Netz. Die im Netz weitergeleitete Information ist in der zeitlichen Aufeinanderfolge der Aktionspotentiale verschlüsselt (Frequenzmodulation). Paukert: Ein Fenster zum Ich – Teil 1 20 Erreicht ein Aktionspotential die präsynaptische Membran, dann werden dort so genannte Transmitter-Moleküle freigesetzt, welche den Spalt überqueren und sich an so genannte Rezeptor-Moleküle an der postsynaptischen Membran anheften. Die Rezeptoren öffnen oder schließen nun selektiv Membrankanäle, wodurch der Ioneneinstrom gesteuert wird. Somit steuern die Transmitter die Weiterleitung der elektrischen Signale, entweder erregend (exzitatorisch) oder hemmend (inhibitorisch). Bestimmte Transmitter werden in bestimmten Hirnregionen erzeugt und steuern dort die Aktivität dieser Regionen. Eine Beeinflussung der Transmitter beeinflusst somit auch die Gehirnfunktionen. Die Wirkung vieler Medikamente beruht darauf, dass sie den natürlichen Botenstoffen nachgebaut werden und daher Rezeptormoleküle besetzen. Die agonistischen Medikamente erzielen die gleiche Wirkung wie die Botenstoffe. Antagonisten besetzen die Rezeptoren und blockieren sie ohne eine Wirkung zu erzielen. In der folgenden Abbildung sind diese Mechanismen schematisch dargestellt. Paukert: Ein Fenster zum Ich – Teil 1 21 Gelangt ein Aktionspotential in der Nervenfaser an eine Synapse, dann kommt es dort zu einer Erregungsübertragung auf das benachbarte Neuron. Das elektrochemische Geschehen soll am Beispiel einer Synapse mit Acetylcholin (ACh) als Transmitter beschrieben werden, wobei das Acetylcholin in der Nervenendigung aus Acetyl-Coenzym-A (Acetyl-CoA) und Cholin gebildet und in den Vesikeln (Bläschen) gespeichert wird. [a] Wenn ein Aktionspotential die Endigung erreicht, öffnen sich die zuvor geschlossenen Calciumkanäle und lassen Calciumionen Ca2+ einströmen. [b] Dies bewirkt, dass die Vesikel mit der präsynaptischen Membran verschmelzen und ACh-Moleküle in den synaptischen Spalt entlassen. [c] Der freigesetzte Transmitter bindet sich an spezifische ACh-Rezeptoren auf der postsynaptischen Membran des benachbarten Neurons und löst dort die Öffnung der Natriumkanäle aus. Viele Na+ -Ionen strömen ins Zelleninnere und wenige K+ -Ionen nach außen. Dadurch kann ein postsynaptisches Aktionspotential aufgebaut und die Erregungsleitung fortgesetzt werden. [d] Der Transmitter ACh wird dann an den Membranrezeptoren durch das Enzym Acetylcholinesterase (AChE) sofort zu Acetat (A) und Cholin (Ch) abgebaut. [e] Diese Produkte werden von der präsynaptischen Nervenendigung aufgenommen und anschließend zur neuerlichen Synthese von Acetylcholin verwendet. Paukert: Ein Fenster zum Ich – Teil 1 22 Transmitterstoffe und Membranrezeptoren Wie das Gehirn funktioniert, versteht man am besten, wenn man die synaptische Übertragung betrachtet, also die Weise, in der Nervenzellen Information an andere Nerven-, Muskel- oder Drüsenzellen übermitteln. In der Zusammenschau ist dieser Prozess unkompliziert: Sobald ein Aktionspotential eine Nervenfaserendigung (den präsynaptischen Teil der Synapse) erreicht, öffnet es normalerweise geschlossene, elektrisch gesteuerte Calcium-Kanäle in der präsynaptischen Membran. Calcium strömt ein und setzt die Ausschüttung von Neurotransmittersubstanzen in Gang, die durch den synaptischen Spalt diffundieren, auf Rezeptoren in der postsynaptischen Membran einwirken und so Reaktionen in der postsynaptischen Nerven-, Muskel- oder Drüsenzelle auslösen. Die durch den präsynaptischen Strom freigesetzte Neurotransmittersubstanz heftet sich an spezifische Rezeptormoleküle in der postsynaptischen Membran. Diese Rezeptoren bewirken dann Änderungen in der postsynaptischen Zelle. Bei der schnellen (weniger als eine Millisekunde benötigenden) synaptischen Übertragung steuern die Rezeptoren unmittelbar Ionenkanäle. Hierbei handelt es sich nicht um einen elektrisch gesteuerten, sondern um einen chemisch gesteuerten Ionenkanal. Er ist nur zu öffnen, indem sich der Transmitter an jene Rezeptoren heftet, die die Kontrolle über den Kanal ausüben. Daneben gibt es auch seltene, rein elektrisch gesteuerte Synapsen, die ohne Transmitter auskommen. Bei der schnellen erregenden synaptischen Übertragung (exzitatorisch) öffnen die Rezeptoren Natrium-Kanäle. Natrium strömt ein und verursacht eine gewisse Depolarisation (das heißt, das lokale Membranpotential wird weniger negativ). Diese erregende Reaktion (EPSP, exzitatorisches postsynaptisches Potential) ist abgestuft und in ihrer Stärke davon abhängig, wie viele Rezeptoren bzw. Kanäle aktiviert wurden. Sind ausreichend viele Rezeptoren in Tätigkeit versetzt (Summationseffekt), dann wird das Membranpotential im Anfangsteil der Nervenfaser, wo sie den Zellkörper verlässt, depolarisiert. Sobald die Aktionspotentialschwelle der hier befindlichen Natriumkanäle erreicht ist, entsteht ein postsynaptisches Aktionspotential, welches sich dann entlang der Nervenfaser weiter fortpflanzt. Bei der schnellen hemmenden synaptischen Übertragung (inhibitorisch) steuern die Rezeptoren gewöhnlich geschlossene Chlorid-Kanäle. Sie öffnen diese, Chloridionen strömen ein und hyperpolarisieren die Zellmembran (ihr Potential wird also negativer als das Ruhepotential). Dieses inhibitorische postsynaptische Potential (IPSP, ca. -75 mV) verhindert das Erreichen des Schwellwertes zur Erzeugung eines Aktionspotentials, sodass keine Signalfortpflanzung erfolgen kann. Paukert: Ein Fenster zum Ich – Teil 1 23 Heute hat die moderne Molekularbiologie bereits das Wissen und die Instrumente entwickelt, um die Neurotransmitter-Rezeptoren (die komplexe Proteinmoleküle sind) zu charakterisieren. In zunehmendem Maß wird klar, dass es weniger die bloße, als Transmitter benutzte Substanz ist, sondern der Rezeptor, der die "Botschaft" bei der synaptischen Übertragung in sich trägt. So kann ACh sowohl als erregender wie als hemmender Neurotransmitter fungieren, je nachdem auf welchen Rezeptor es einwirkt. Ein Typ von ACh-Rezeptor (nämlich der erregend wirkende Rezeptor an der neuromuskulären Endplatte) hat ein hohes Molekulargewicht und besteht aus vier Typen von Untereinheiten. Der Grund, weshalb so viele Wirkstoffe einen derart mächtigen Einfluss auf das Gehirn haben, liegt darin, dass sie den spezifischen Rezeptoren der Nervenzellen vortäuschen sie wären Neurotransmitter. Dies gelingt ihnen, indem sie sich auf Grund ihrer chemischen Struktur an den Rezeptor anlagern und ihn entweder aktivieren (agonistische Wirkstoffe) oder das normale Transmittermolekül daran hindern, sich an den Rezeptor zu heften (antagonistische Stoffe). Bekannte klassische Transmitter sind: Acetylcholin, die Aminosäuren Glycin, Glutamat und GABA, die Monoamine Dopamin, Noradrenalin, Adrenalin und Serotonin. Daneben gibt es noch einige modulierend wirkende Neuropeptide wie beispielsweise Endorphine und Enkephaline. Diese bewirken unmittelbar keine Leitfähigkeitsänderungen in den synaptischen Membranen, sondern beeinflussen Intensität und Dauer der Wirkung der klassischen Transmitter. Glutamat, eine Aminosäure, die sowohl in der Nahrung als auch in allen Zellen vorkommt, ist das Arbeitspferd unter den schnellen erregenden Neurotransmittern im Gehirn. Auch bei der Speicherung von Gedächtnisinhalten scheint es eine Schlüsselrolle zu spielen. Ein Beispiel hiefür ist die Langzeitpotenzierung (LTP). Wird eine Leitungsbahn, die Glutamat als Transmitter benutzt (etwa im Hippocampus), in rascher Folge gereizt, ruft dies eine anhaltend gesteigerte Erregbarkeit der aktivierten Synapsen hervor. Dieser Verstärkereffekt (Potenzierung) wird durch Anregung eines bestimmten Typs von Glutamatrezeptor eingeleitet, dem so genannten NMDA-Rezeptor (NMDA=N-Methyl-D-Aspartat). LTP wurde erstmals 1973 im Hippocampus entdeckt. Auch ein anderer synaptischer Wirkungsmechanismus, die Langzeitdepression (LTD), kann an Glutamatsynapsen (beispielsweise im Kleinhirn) vorkommen. Sie geht mit einer langdauernden abgeschwächten Erregbarkeit von Glutamatrezeptoren des AMPA-Typs einher (AMPA= -Amino-3-Hydroxy-5-Methyl-4Isoxazol-Propionsäure). Die LTD wurde erst 1981 von dem japanischen Neurobiologen Masao Ito an den Purkinje-Zellen im Kleinhirn entdeckt. Paukert: Ein Fenster zum Ich – Teil 1 24 Gamma-Amino-Buttersäure (GABA) ist das Arbeitspferd unter den schnellen hemmenden Neurotransmittern im Gehirn. Seine Rezeptoren steuern Chloridkanäle in Nervenzellmembranen. Die Tranquilizer (Benzodiazepine) wirken auf GABA-Rezeptoren, indem sie deren Wirkungen steigern und so die Hemmung verstärken. Mit Benzodiazepinen werden panischen Angstzustände behandelt. Acetylcholin (ACh) ist der Transmitter an der neuromuskulären Endplatte zwischen Nervenfaser und Muskelzelle und an bestimmten anderen peripheren Synapsen des autonomen Nervensystems (zum Beispiel im Herzen). Er kann als der am gründlichsten untersuchte und wohl bekannteste Transmitter gelten. 1924 entdeckte Otto Loewi in einem klassischen Experiment der Neurobiologie das Acetylcholin und klärte damit die Frage, ob die synaptische Übertragung vom Vagusnerv zum Herzmuskel (und auch an anderen Synapsen) elektrischer oder chemischer Natur ist. Loewis Experiment - ein Modell dafür, wie einfach ein Versuch sein kann - verdient es, genauer beschrieben zu werden. Der Vagusnerv ist einer der größeren Nerven, die das Herz kontrollieren. Aus einem Frosch kann man ihn und das Herz herauspräparieren und in einer Schale mit so genannter Ringerlösung am Leben halten. Diese Lösung ähnelt in ihrer Salzzusammensetzung dem Blut. Eine elektrische Reizung des Vagus, ob im lebenden Tier oder an einem isolierten Herzen in einem Gefäß, verlangsamt den Herzschlag. Loewi reizte den Vagus eines in Ringerlösung überführten Herzens viele Male und löste jedesmal eine Senkung der Herzfrequenz aus. Anschließend entnahm er der Schale mit dem stimulierten Herzen etwas Lösung und gab sie in eine andere Schale mit einem zweiten Froschherzen. Auch dieses Herz schlug daraufhin langsamer. Das Experiment bewies, dass die synaptische Übertragung mit Hilfe eines chemischen Transmitterstoffes abläuft. Das Acetylcholin ist die vielleicht am besten erforschte Neurotransmittersubstanz, denn sie wirkt an neuromuskulären Endplatten und lässt sich dort sehr gut untersuchen. Es gibt zwei Haupttypen von ACh-Rezeptoren, von denen die einen erregend auf die Skelettmuskulatur einwirken (nicotinerge Rezeptoren) und die anderen beispielsweise den Herzmuskel hemmend beeinflussen (muscarinerge Rezeptoren). Über ACh-Bahnen im Gehirn weiß man weitaus weniger. Die Zellkörper der wichtigsten ACh-Bahn des Gehirns befinden sich im Nucleus basalis (ihre Fasern ziehen in weite Teile der Großhirnrinde) und in den Septumkernen (deren Fasern zum Hippocampus ziehen). In den Gehirnbahnen übt ACh anscheinend keine schnellen synaptischen Wirkungen aus (wie es dies an den Muskeln tut), sondern ruft eher langsame synaptische Effekte über zusätzliche Vermittlerstoffe (second messenger) hervor. Bei der langsamen und schnellen synaptischen Übertragung gleichen sich die ersten Schritte bis einschließlich zu dem Moment, in dem sich der Überträgerstoff an die Rezeptormoleküle der postsynaptischen Membran anlagert; doch dann trennen sich ihre Wege. Paukert: Ein Fenster zum Ich – Teil 1 25 Langsame Rezeptoren sind nicht direkt an Ionenkanäle gekoppelt. Sie aktivieren statt dessen so genannte G-Proteine, die ihrerseits Vermittlerstoffe im Inneren der Zelle aktivieren. Beispielhaft für ein solches System ist die (durch das G-Protein vermittelte) Umwandlung von ATP in cAMP (zyklisches Adenosinmonophosphat). cAMP wirkt dann als second messenger für die Phosphorylierung eines Proteins, d.h. die Verbindung eines Proteinmoleküls mit einem Phosphatmolekül. Dadurch wird das Protein als Enzym aktiv und beschleunigt einen bestimmten biochemischen Prozess im intermediären Stoffwechsel der Zelle. Weitere gut untersuchte Neurotransmitter des Gehirns sind die zwei Monoamine Dopamin und Noradrenalin. Sie werden in den Zellen aus Tyrosin hergestellt, einer Aminosäure, die gewöhnlich in der Nahrung vorkommt. Tyrosin wird zunächst in L-Dopa, dann in Dopamin, schließlich in Noradrenalin und zuletzt in Adrenalin umgewandelt. Welches Endprodukt - ob Dopamin oder Noradrenalin entsteht, hängt davon ab, welche Enzyme in der Zelle vorliegen. Es gibt drei wichtige dopaminerge Nervenbahnen im Gehirn. Eine befindet sich im Hypothalamus im Zwischenhirn, eine andere erstreckt sich von der "Substantia nigra" im Mittelhirn zu den Basalganglien des Großhirns, eine weitere verläuft vom Hirnstamm zur Großhirnrinde und zu anderen Vorderhirnstrukturen. Bei der Parkinson-Krankheit (Schüttellähmung) gehen dopaminhaltige Zellen in der "Substantia nigra" zu Grunde. Die Symptome der Erkrankung sind Ausdruck der daraus resultierenden verminderten Dopaminübertragung in den Basalganglien. Injiziert man den Patienten L-Dopa (welches sich im Gehirn in Dopamin umwandelt), verbessert sich ihr Zustand. Das dopaminerge System, welches auf das Vorderhirn hinzielt (projiziert), scheint eine Rolle bei der schweren Geisteskrankheit Schizophrenie zu spielen. Im Allgemeinen blocken Substanzen, die schizophrene Symptome lindern, dopaminerge Synapsen im Gehirn. Viele Forscher vertreten so die Ansicht, Ursache der Schizophrenie sei eine Überaktivität des mesolimbischen Dopaminsystems in der Tiefe des Großhirns und eine kompensatorische Unteraktivität im Vorderhirn. Fast alle noradrenergen Bahnen des Gehirns entspringen im "Locus coeruleus", einer kleinen Nervenzellansammlung im Hirnstamm, und entsenden ihre Fasern zu praktisch allen Vorderhirnstrukturen. Das Noradrenalinsystem soll das Aktivierungsniveau regulieren (ARAS, aufsteigendes retikuläres System) und möglicherweise an der Konsolidierung des Gedächtnisses mitwirken (Langzeitspeicherung von Gedächtnisinhalten). Paukert: Ein Fenster zum Ich – Teil 1 26 Serotonin ist ein weiterer Neurotransmitter, der in den Zellen aus einer in der Nahrung vorkommenden Aminosäure hergestellt wird, dem Tryptophan (es ist in Bananen reichlich vorhanden). Serotonin wird wie Dopamin und Noradrenalin zur chemischen Klasse der Monoamine gezählt. Die Zellkörper der serotonergen Bahnen im Gehirn befinden sich hauptsächlich in den so genannten RapheKernen des Hirnstammes. Ihre Fasern ziehen zum Hypothalamus im Zwischenhirn und zu Vorderhirnstrukturen des Großhirns. Bei schwerer Depression scheinen die noradrenergen und serotonergen Bahnen eine Rolle zu spielen. Es gibt zwei Formen der Depression: Bei der einen handelt es sich um eine anhaltende schwere Verstimmung (Major Depression), bei der anderen tritt neben der schweren depressiven Verstimmung mindestens eine manische Episode auf (Bipolare Störung, früher als manisch-depressive Erkrankung bezeichnet). Im Allgemeinen lassen sich schwere depressive Zustände durch Substanzen günstig beeinflussen, welche die Aktivität noradrenerger und serotonerger Bahnen im Gehirn erhöhen oder verstärken. Allerdings gelingt es diesen Substanzen kaum, die Symptome der bipolaren Störung zu lindern. Jedoch sprechen Patienten mit bipolarer Störung auf Lithium gut an. Alle diese für das psychische Wohlbefinden offenbar entscheidenden Neurotransmittersysteme des Gehirns scheinen über den langsamen Mechanismus der "second messenger" zu wirken. Interessanterweise machen sie nur wenige Prozent der gesamten Nervenüberträgerstoffe im Gehirn aus. Die schnellen Transmitter wie Glutamat und GABA kommen sehr viel häufiger vor und sind viel weiter verbreitet. Die langsamen synaptischen Wirkungen dauern einige Zehntelsekunden und dienen häufig der Modulation der schnellen Transmitterwirkungen. So dämpfen beispielsweise so genannte Opiate an den Synapsen im Rückenmark die Schmerzübertragung mittels Glutamat. Der Nachweis von Nervenzellrezeptoren im Gehirn, die auf Opium und seine Abkömmlinge - Morphin und Heroin - ansprechen, ist ein verblüffendes Forschungsergebnis der neueren Zeit. In der Folge stieß man auf Hirnopiate, von Nervenzellen und Hypophyse (Hirnanhangdrüse) hergestellte Substanzen, die auf eben diese Rezeptoren einwirken und sehr ähnliche Effekte wie Morphin hervorrufen: Sie lindern Schmerzen und lösen angenehme Empfindungen aus. Die Hirnopiate sind allesamt Peptide (Ketten von Aminosäuren) und entstammen drei Superhormonfamilien, deren Aufbau von den Genen der entsprechenden Zellen gesteuert wird. Diese drei riesigen Eiweißmoleküle werden gespalten, um die viel kleineren opiumähnlichen Peptide hervorzubringen: Endorphine, Enkephaline und Dynorphine. Enkephaline findet man in Nervenzellen, die zum langsamen Schmerzsystem gehören. Endorphin wird von der Hypophyse freigesetzt. Paukert: Ein Fenster zum Ich – Teil 1 27 Die grundsätzliche Arbeitsweise von Neuronen im Netz Ein Neuron ist idealisiert ein einfaches Schaltelement, das viele Eingangssignale Xi (Inputs) in ein Ausgangssignal Y (Output) umwandelt. Die Signale sind digitalisiert, d.h. 0 = KEIN Signal und 1 = EIN Signal. Der Index i bezeichnet die i-te Synapse von insgesamt n Inputleitungen. Die Verbindungsstärke einer Synapse mit dem Neuron ist durch die Anzahl der chemischen Transmitterspeicher und Rezeptor-Moleküle gegeben. Man nennt sie Synapsengewicht Wi. Das Neuron bildet die gewichtete Summe aller Inputs S = (Wi * Xi). Dann wird diese Summe mit einem internen Schwellwert G verglichen. Ist S < G, dann bleibt das Neuron stumm, sein Outputsignal ist 0. Ist S = G oder S > G, dann feuert das Neuron, sein Outputsignal ist 1, d.h. an seiner Membran entsteht ein Aktionspotential. Lernen bedeutet eine Änderung des neuronalen Netzes, d.h. es ändern sich nachhaltig die Synapsenstärken Wi der beteiligten Neuronen. Damit ändert sich auch die gesamte Outputleistung des Netzes. [1.1.3] Gliederung des zentralen Nervensystems (ZNS) Der überwiegende Teil der ungefähr 200 Milliarden Neuronen des zentralen Nervensystems befindet sich im Gehirn. Dabei kann eine Nervenzelle bis zu 10000 Synapsen aufweisen, sodass ein vielschichtiges Netzwerk entsteht. Die Zellen selbst erscheinen als graue, ihre Fortsätze als weiße Substanz. Das ZNS besteht oben aus dem Gehirn in der Schädelhöhle und setzt sich nach unten durch das Hinterhauptsloch der Schädelbasis in das Rückenmark im Wirbelkanal der Wirbelsäule fort. Im interzellulären Raum befinden sich neben Blutgefäßen noch so genannte Gliazellen, die für bestimmte Nervenfasern isolierende Hüllschichten (Markscheiden) produzieren. Die Gliazellen umkleiden auch die Blutgefäße und tragen so zur Blut-Hirn-Schranke bei, die verhindert, dass viele Substanzen (z.B. Antibiotika) weiter ins Gehirn gelangen. Das Nervengewebe wird geschützt durch drei Hirnhäute. Im Spalt zwischen den beiden inneren Häuten befindet sich die Hirnflüssigkeit (Liquor cerebrospinalis), die in den vier Hirnkammern (Ventrikeln) gebildet wird und als Stoßdämpfer dient. Die Flüssigkeit wird durch die venösen Blutgefäße resorbiert. Die Blutversorgung des Gehirns erfolgt frontal über die zwei Kopfarterien (a. carotis) und dorsal über die zwei Wirbelarterien (a. vertebralis). Paukert: Ein Fenster zum Ich – Teil 1 28 Von oben nach unten wird das ZNS grob in folgende Abschnitte gegliedert: Großhirn - Hirnstamm und Kleinhirn - Verlängertes Mark - Rückenmark Das Gehirn ist grundsätzlich in zwei Hälften geteilt, wobei jede für die gegengleiche Körperhälfte zuständig ist. Alle afferenten (zuleitenden) und efferenten (wegleitenden) Bahnen kreuzen auf die Gegenseite. Paukert: Ein Fenster zum Ich – Teil 1 29 Der entwicklungsgeschichtlich ältere Hirnstamm wird in Zwischenhirn, Mittelhirn und Brücke unterteilt. Das dahinter liegende Kleinhirn dient der Koordination der Körpermotorik. Das jüngere Großhirn gliedert sich in das weiße Mark innen und die außen liegende graue Rinde (Cortex). In der Tiefe des Großhirns erstreckt sich das ältere limbische System, das auch Anteile am Hirnstamm hat. Unter einem Kern (nucleus) versteht man eine abgrenzbare Ansammlung von Nervenzellen, welcher eine bestimmte Funktion zugeordnet ist. Im gesamten Gehirn hat man viele solche Kerngebiete lokalisiert. Dies geschieht meistens durch elektrische Reizung mit dünnen, vorsichtig eingeführten Stahlelektroden. Das Großhirn (Telencephalon) Außen befindet sich die graue, aus Nervenzellen bestehende Großhirnrinde und innen das weiße, aus Nervenfasern bestehende Großhirnmark. Ohne Hirnrinde (Cortex) ist kein bewusstes Erleben möglich. Eine ausführliche Beschreibung des Großhirns erfolgt weiter unten. Das Zwischenhirn (Diencephalon) Unterhalb des Großhirns befindet sich das Zwischenhirn. In der Mitte liegt die dritte Hirnkammer (Ventrikel), seitlich dorsal befinden sich die beiden Thalami (Sehhügel). In diese strahlen die afferenten sensorischen Fasern von den Sinnesorganen ein und werden in den Thalamus-Kernen umgeschaltet. Entweder erfolgt die Weiterleitung zur Cortex (bewusstes Erleben) oder die Umschaltung zu ventralen Steuerkernen im Hirnstamm, welche die Körpermotorik regulieren. Der Thalamus wird auch als das Vorzimmer zum Bewusstsein bezeichnet. In einer Schleife zwischen Großhirn und Thalamus erfolgt über absteigende und aufsteigende, hemmende und erregende Impulse die Kontrolle der bewussten Aufmerksamkeit. Am Boden der dritten Hirnkammer liegt der so genannte Hypothalamus mit seinen Steuerzentren für Temperaturregelung, Sexualität, Lust, Hunger und Durst. An den Hypothalamus schließen die zwei Hormondrüsen Epiphyse und Hypophyse. Das Zwischenhirn ist die Schnittstelle zwischen dem zentralen somatischen Nervensystem, dem vegetativen Nervensystem und dem Hormonsystem. Das Mittelhirn (Mesencephalon) Die dritte Hirnkammer setzt sich hier nach unten als schmaler Gang fort. Ventral (vorne) liegt das Tegmentum (Haube) mit seinen motorischen Ursprungskernen von einigen Hirnnerven und dorsal (hinten) liegt das Tectum (Vierhügelplatte). Paukert: Ein Fenster zum Ich – Teil 1 30 In das Tectum strahlen Seitenäste von afferenten Bahnen ein und dessen Kerne sind für optische und akustische Reflexe verantwortlich. Weitere sehr bedeutsame Areale sind die motorischen Zentren, vor allem die Stammganglien des schwarzen und roten Kerns (Nucleus niger und Nucleus ruber). Sie sind den Basalganglien des Großhirns (Striatum und Pallidum) nachgeschaltet und steuern über absteigende Bahnen die unbewusste Körpermotorik. Dieses System wird extrapyramidal motorisch genannt (EPM) - im Gegensatz zur willkürlichen Motorik, welche von motorischen Zentren der Cortex und über die absteigende Pyramidenbahn kontrolliert wird. Die Brücke (Pons) Der schmale Gang im Mittelhirn erweitert sich nach unten zur vierten Hirnkammer. Auf deren Boden liegt die Rautengrube, an die sich zur Hirnbasis hin eine kräftige Auftreibung anschließt. Diese Brücke enthält wichtige Kerne, welche der Verbindung von Großhirn und Kleinhirn dienen (Großhirn - Kleinhirn - Bahnen). Unterhalb der Brücke liegt das verlängerte Mark. Das Kleinhirn (Cerebellum) Das Kleinhirn liegt hinter der Brücke unter dem Hinterhauptslappen des Großhirns und gliedert sich außen in eine graue Rinde und innen in ein weißes Mark. Die Kleinhirnschenkel verbinden das Kleinhirn mit dem Mittelhirn, der Brücke und dem verlängerten Mark. Die Neuronen des Kleinhirns erhalten von afferenten Fasern sensorische Meldungen vom benachbarten Gleichgewichts-Sinnesorgan und von den Sehnen-Rezeptoren der Körpermuskulatur. Nach deren Verarbeitung werden motorische Erregungen erzeugt, die zur Regulation des Muskeltonus und der Feinregulation der gesamten Körpermotorik dienen. Beinträchtigungen oder Ausfälle von Kleinhirnkernen führen zu so genannten Kleinhirnataxien (Bewegungsstörungen). Das verlängerte Mark (Medulla oblongata) Neben den zum Hirnstamm aufsteigenden und zum Rückenmark absteigenden Bahnen enthält das verlängerte Mark wichtige Kerngebiete von Hirnnerven, aber auch Steuerzentren für das vegetative Nervensystem (Atemzentrum und Kreislaufzentrum). Ein sehr interessantes Gebiet ist die formatio reticularis (ARAS, aufsteigendes, retikuläres Aktivierungssystem). Dieses verstreute Netzwerk versorgt die Großhirnrinde mit unspezifischen Erregungen und steuert so den Wachheitszustand des Individuums (von tiefster Bewusstlosigkeit bis zur hellwachen Aufmerksamkeit). Paukert: Ein Fenster zum Ich – Teil 1 31 Aus dem Gehirn treten im Gebiet des Hirnstammes und des verlängerten Markes 12 Hirnnerven-Paare, welche sensorische, motorische und vegetative Anteile aufweisen (für jede Körperhälfte ein Nerv). Diese innervieren vor allem verschiedene Bereiche des Gesichtsschädels. Beispiele dafür sind: II. Hirnnerv (Sehnerv, N. opticus): Netzhaut des Auges - Sehnervenkreuzung Zwischenhirn - Thalamus - Großhirnrinde. V. Hirnnerv (Drillingsnerv, N. trigeminus): Austrittsstelle bzw. Eintrittsstelle ist die Brücke (Pons). Motorische Versorgung: Kaumuskeln. Sensorische Versorgung von Gesichts- und Kopfhaut, Nasen-, Mund- und Augenhöhle. X. Hirnnerv (herumschweifender Nerv, N. vagus): Austrittsstelle bzw. Eintrittsstelle ist das verlängerte Mark. Parasympathische Versorgung der inneren Organe in Kopf-, Hals-, Brust- und Bauchraum. Sensorische Versorgung von Kehlkopf und inneren Organen. Aus dem Rückenmark treten beidseitig durch die Zwischenwirbellöcher in der Wirbelsäule 31 Körpernerven-Paare. Jeder dieser Spinalnerven versorgt ein bestimmtes Körpersegment, bestehend aus einem Hautbereich (Dermatom), einem Muskelbereich (Myotom) und einem inneren Organbereich (Enterotom). Im Wirbelkanal liegt in Form einer Schmetterlingsfigur die graue Substanz der Nervenzellen, umgeben von der weißen Substanz ihrer Fortsätze. Hinten strahlen die sensorischen afferenten Fasern der Körpernerven ein, vorne liegen die motorischen efferenten Leitungsbahnen. Die Nervenzellen der grauen Substanz dienen erstens der direkten Umschaltung von sensorischen auf motorische Leitungen (Reflexschaltungen zur automatischen Bewegungskoordination). Zweitens dienen sie entweder der Weiterleitung der sensorischen Signale von den Rezeptoren in der Peripherie zum Gehirn (Hinterstrang) oder der Weiterleitung von motorischen Signalen von der Zentrale zu den Muskeln in der Peripherie. Dabei sind die Bahnen der unwillkürlichen Motorik (EPM-System, Vorderstrang) und jene der willkürlichen Motorik (Pyramidenbahn, Seitenstrang) zu unterscheiden. Die drei Hauptfunktionen des Nervensystems Verbindung mit der Welt: Aufnahme, Verarbeitung, Beantwortung von Reizen. Das entspricht der sensorischen, zentralen und motorischen Informationsverarbeitung. Bemerkenswert ist, dass die sensorischen Funktionen zumeist dorsal (hinten), jedoch motorische Funktionen zumeist ventral (vorne) zu finden sind. Regulation der Organtätigkeit: vor allem durch das vegetative Nervensystem. Sitz des Bewusstseins: im Gehirn, insbesonders in der Großhirnrinde. Paukert: Ein Fenster zum Ich – Teil 1 32 [1.1.4] Gehirn und Großhirn Von außen betrachtet scheint die Hauptmasse des Gehirns aus den beiden Großhirnhemisphären (linke und rechte Großhirnhalbkugel) zu bestehen, welche die übrigen Teile so überwölben, dass diese von oben und von der Seite kaum sichtbar sind. Beide Halbkugeln sind durch die Längsfurche (Fissura longitudinalis cerebri) getrennt, die bis zu den querlaufenden Fasermassen des Balkens (Corpus callosum) herunterreicht. Der Balken enthält die Verbindungsbahnen der beiden Hirnhälften. Die Oberfläche des Großhirns zeigt erhabene Windungen (Gyrus). Dazwischen liegen Furchen (Sulcus). Diese Faltung bewirkt eine deutliche Vergrößerung der aktiven Oberfläche. Die großen Lappen der Großhirnrinde heißen nach ihrer Lage: Stirnlappen (Lobus frontalis) Scheitellappen (Lobus parietalis) Schläfenlappen (Lobus temporalis) Hinterhauptslappen (Lobus occipitalis) Paukert: Ein Fenster zum Ich – Teil 1 33 Die Lappengrenzen werden teilweise durch fest und tief ausgebildete Furchen markiert. So ist der Schläfenlappen durch die tiefe Seitenfurche (Sulcus lateralis oder Sylvius-Spalte) gegenüber Stirn- und Scheitellappen abgegrenzt. Zwischen Stirn- und Scheitellappen verläuft die Zentralfurche (Sulcus centralis). Dahinter liegt die hintere Zentralwindung (Gyrus postcentralis), davor die vordere Zentralwindung (Gyrus präcentralis). Der stark entwickelte Hinterhauptslappen grenzt sich mit dem Sulcus parietooccipitalis vom Scheitellappen ab. Drängt man linken und rechten Hinterhauptslappen auseinander, so kommt die Region der Calcarinafurche (sulcus calcarinus) zum Vorschein. Der Schläfenlappen lässt von außen eine obere, mittlere und untere Schläfenwindung erkennen. Einen ähnlichen Verlauf dreier übereinander gelegener Windungen zeigt der Stirnlappen. In der Tiefe der Seitenfurche liegt die so genannte Insel (Lobus insularis). Unter dem Stirnlappen liegt das Riechhirn (Riechkolben, bulbus olfactorius). Die beiden Großhirnhemisphären besitzen mit der Hirnrinde (Cortex cerebri) eine gleichmäßig dicke Randschicht grauer Substanz (ca. 5 mm), die allen Windungen und Furchen der Oberfläche folgt. Die Cortex allein enthält ungefähr 20 Milliarden Neuronen. Im Innern jeder Hemisphäre erstreckt sich eine geräumige, mit Hirn-Rückenmarks-Flüssigkeit (Liquor cerebrospinalis) gefüllte Hirnkammer (linke und rechte Seitenventrikel). Zwischen diesen Hohlräumen und der Hirnrinde dehnt sich eine große Masse weißer Substanz, in die mehrere Kerngebiete, die so genannten Basalganglien, eingelagert sind. Die weiße Substanz (Hirnmark) setzt sich aus Faserbündeln (Bahnen) zusammen. Es handelt sich dabei um Assoziationsbahnen, Kommissurenbahnen und Projektionsbahnen. Die Assoziationsbahnen sind Verbindungszüge, welche verschiedene Teile der gleichen Großhirnhemisphäre verknüpfen. Kommissurenbahnen verbinden einander entsprechende Teile beider Hemisphären. Sie sind unter der Längsfurche in der Mitte zwischen den Hemisphären zu einer Nervenfaserplatte, dem Balken (Corpus callosum), zusammengedrängt. Das Großhirn steht durch seine rindenwärts (afferent) und rückenmarkwärts (efferent) ziehenden Fernbahnen oder Projektionsbahnen mit dem ganzen Organismus in wechselseitiger Verbindung. Die Projektionsbahnen durchlaufen ziemlich geschlossen die innere Kapsel (Capsula interna). Die großen subcortikal (unter der Hirnrinde) gelegenen Kerngebiete der Hemisphären heißen Basalganglien. Sie grenzen an die Seitenventrikeln. Man unterscheidet verschiedene Komponenten: den bogenförmig mit dem Seitenventrikel verlaufenden Schweifkern (Nucleus caudatus), den keilförmigen Linsenkern (Nucleus lentiformis) und den lateralen Schalenkern (Putamen). Diese Kerngebiete werden auch als Streifenkörper (Striatum) bezeichnet. Paukert: Ein Fenster zum Ich – Teil 1 34 Schließlich muss noch der mediale bleiche Kern (Pallidum) erwähnt werden, der auch schon zum Zwischenhirn gezählt wird. Auch die Vormauer (Claustrum) wird zu den Basalganglien gerechnet. Zusammen mit Kerngebieten des Hirnstamms (den Stammganglien im Mittelhirn) sind die Basalganglien vor allem für die unbewusst ablaufende Bewegungskontrolle sehr wichtig. Zwischen dem Linsenkern, dem Schweifkern sowie dem Thalamus des Zwischenhirns verlaufen die Nervenfaserbündel der inneren Kapsel (Capsula interna). Hierbei handelt es sich um Projektionsbahnen, wie die Pyramidenbahn und die extrapyramidalen Bahnen. Ein sehr interessantes Gebiet an der Basis des Großhirns an der Grenze zum Zwischenhirn ist das limbische System, welches den Balken saumförmig umgibt (limbus = Saum). Die wichtigsten Strukturen davon sind der Mandelkern (Amygdala), der Hippocampus und der Gyrus cinguli. Der Fornix ist ein dickes Nervenbündel, das die Teile des limbischen Systems miteinander verknüpft. Es bestehen starke Verbindungen mit Bereichen des Thalamus, des Hypothalamus und der Großhirnrinde. Das limbische System spielt eine entscheidende Rolle bei der Entstehung von Gefühlen (emotionale Bewertung von sensorischen Erregungen). So konnten dort Lust-, Furcht- und Wut-Zentren lokalisiert werden. Forschungen haben gezeigt, dass der Hippocampus (Seepferdchen) für die Einspeicherung von Erregungen (Lernen, Gedächtnis) eine wichtige Funktion ausübt. Der komplexe Bau des Zentralnervensystems ist am Beispiel der Großhirnrinde besonders eindrucksvoll zu veranschaulichen. Die Rindensubstanz besteht aus sechs oberflächenparallelen, gut abgrenzbaren Schichten, die sich durch Art und Anordnung der Nervenzellen und Nervenzellfortsätze unterscheiden (z.B. die großen Pyramidenzellen). Die Anteile dieser Schichten in den einzelnen Arealen der Hirnrinde sind unterschiedlich, was auch verschiedenen Funktionen der Rinde entspricht. So heben sich Rindenfelder strukturell und funktionell voneinander ab. Es muss zwischen sensorischen und motorischen Arealen unterschieden werden. Erstere liegen dorsal hinter der Zentralfurche (z.B. in der hinteren Zentralwindung). Sie verarbeiten die, von den Rezeptoren über aufsteigende Bahnen einlangenden sensorischen Erregungen zu bewussten Wahrnehmungen. Zweitere liegen ventral vor der Zentralfurche (z.B. in der vorderen Zentralwindung). Sie erzeugen für eine willkürlich beabsichtigte Bewegung jene Erregungsmuster, die über absteigende Leitungsbahnen (Pyramidenbahn) die entsprechenden Muskeln steuern. In den beiden Rindenbereichen sind sämtliche Regionen des Körpers repräsentiert (Homunculus-Projektion). Außerdem muss noch zwischen primären und sekundären Arealen unterschieden werden. Letztere enthalten so genannte Erinnerungsspuren von den primären sensorischen und motorischen Erregungsmustern und dienen auch zu deren Verknüpfung (Assoziationsfelder). Paukert: Ein Fenster zum Ich – Teil 1 35 Einige wichtige primäre sensorische und motorische Rindenfelder: 1. Primäres Sehzentrum Ort : Sulcus calcarinus im Hinterhauptslappen Funktion : Umsetzung von Erregungen des Sehnervs in bewusste, optische Empfindungen Ausfall : Rindenblindheit (trotz Funktionstüchtigkeit von Auge und Sehnerv) 2. Primäres Hörzentrum Ort : Heschlsche Querwindung im Schläfenlappen Funktion : Umsetzung von Erregungen des Hörnervs in bewusste, auditive Empfindungen Ausfall : Rindentaubheit (trotz Funktionstüchtigkeit von Ohr und Hörnerv) 3. Primäre Hautsensibilität Ort : Abschnitte des Gyrus postcentralis hinter der Zentralfurche Funktion : Umsetzung von sensorischen Erregungen in bewusste Hautempfindungen Ausfall : Empfindungslosigkeit in den entsprechenden Projektionsgebieten 4. Primäre willkürliche Körpermotorik Ort : Abschnitte des Gyrus präcentralis vor der Zentralfurche Funktion : Bewusste Erzeugung von motorischen Erregungen für Muskelbewegungen Ausfall : Bewegungsstörungen bestimmter Muskelgruppen (Apraxien) Einige wichtige sekundäre Rindenfelder (Assoziationsfelder): 1. Optisches Assoziationsfeld Ort : Gyrus angularis, hinten im Schläfenlappen Funktion : Engramme (Erinnerungsspuren) von visuellen Signalmustern Ausfall : Alexie, Unfähigkeit zu Lesen 2. Auditives Ort Funktion Ausfall Assoziationsfeld : Wernikesche Sprachregion, hinten im Schläfenlappen : Engramme von akustischen Signalmustern : Sensorische Aphasie, Unfähigkeit zum Wortverstehen 3. Sprachmotorisches Assoziationsfeld Ort : Brocasche Sprachregion, seitlich hinten im Stirnlappen Funktion : Engramme von motorischen Wort- und Satzmustern Ausfall : Motorische Aphasie, Unfähigkeit zum sinnvollen Sprechen Paukert: Ein Fenster zum Ich – Teil 1 36 4. Schreibmotorisches Assoziationsfeld Ort : Vor dem Gyrus präcentralis Funktion : Engramme von koordinierten Schreibbewegungen der Hände Ausfall : Agraphie, Unfähigkeit zum sinnvollen Schreiben Mittels verschiedener moderner Lokalisationstechniken wurde die Kartierung des Cortex in den letzten Jahrzehnten sehr verfeinert. Interessant sind die Areale im Stirnlappen. Dieser präfrontale Cortex erhält seine Zuleitungen (Afferenzen) hauptsächlich von unspezifischen Thalamus-Kernen und hat ausgedehnte reziproke Verbindungen mit verschiedenen Teilen des limbischen Systems (Hippocampus, Amygdala) und Hypothalamus. Während Hypothalamus und limbisches System für Triebe und Gefühle verantwortlich sind, erweist sich der Stirnlappen als die oberste cortikale Kontrollinstanz für triebhafte und emotionale Verhaltensweisen. Individuen mit Stirnhirnläsionen zeigen dementsprechend auch auffällige Störungen im Sozialverhalten und oftmals eine allgemeine Antriebslosigkeit. Paukert: Ein Fenster zum Ich – Teil 1 37 [1.1.5] Sensorisches und motorisches System In der Abbildung ist die hintere Zentralwindung dargestellt, wo die Körperzonen des Menschen repräsentiert sind. Das sensorische Nervensystem leitet Signale von den Sinnesorganen in der Körperperipherie über das Rückenmark und den Hirnstamm bis in die primär sensorischen Felder der Großhirnrinde (Cortex). Dort entstehen bewusste Sinnesempfindungen. Eine wichtige Zwischenstation vor der Cortex ist der Thalamus (Sehhügel) im Zwischenhirn, der mit vielen anderen Hirnteilen verbunden ist. Grundsätzlich gibt es drei Arten von Neuronen: (a) Die sensorischen Neuronen, deren Zellen direkt außerhalb des Rückenmarks in den hinteren Nervenwurzeln liegen. Sie erhalten Signale von den Sinnesorganen und leiten sie weiter ins ZNS. (b) Die motorischen Neuronen (Motoneuronen) befinden sich in verschiedenen Regionen von Gehirn und Rückenmark. Ihre Axone ziehen zu den Muskeln und steuern dort über die motorischen Endplatten die Kontraktion der Muskelfasern. (c) Die Interneuronen sind zwischen sensorischen und motorischen Neuronen geschaltet und machen 90 % aller Nervenzellen aus. In ihnen kommt es zur spezifischen Verarbeitung der Signale (entweder verstärkend oder abschwächend). Sie sind als Zwischenschichten in neuronalen Netzen zu finden, beispielsweise in der Netzhaut des Auges, wo sie u.a. die so genannte laterale Inhibition bewirken. Paukert: Ein Fenster zum Ich – Teil 1 38 In der Abbildung ist die vordere Zentralwindung dargestellt, wo die Körperzonen des Menschen repräsentiert sind. Das motorische Nervensystem mit seinen Motoneuronen steuert die Kontraktion der Muskelfasern der quergestreiften Skelettmuskulatur. Die unbewusste Stützund Haltemotorik wird mit Reflexen über Rückenmark und Kleinhirn reguliert. Die bewusste Zielmotorik hingegen hat ihren Ursprung in der vorderen Zentralwindung der Großhirnrinde, wo die einzelnen Körperbereiche des Menschen repräsentiert sind, wo die dicke Pyramidenbahn beginnt, welche über das Rückenmark zur Muskulatur zieht. Die unwillkürliche Körpermotorik wird über extrapyramidale Bahnen (EPM) gesteuert. Neben der quergestreiften Skelettmuskulatur gibt es noch die glatte Muskulatur als Wandauskleidung von Blutgefäßen und inneren Hohlorganen. Ihre Steuerung erfolgt autonom über das vegetative Nervensystem. Das gilt auch für die Spezialmuskulatur der Herzens. Paukert: Ein Fenster zum Ich – Teil 1 39 Die obere Abbildung zeigt das Rückenmark mit den Nervenwurzeln der Körpernerven. Die untere Abbildung zeigt einen typischen Reflexbogen. Paukert: Ein Fenster zum Ich – Teil 1 40 [1.1.6] Das vegetative Nervensystem Das vegetative Nervensystem reguliert autonom die Tätigkeit der inneren Organe. Oberste Steuerungszentrale ist der Hypothalamus. Das vegetative Nervensystem besteht aus zwei Gegenspielern. Der Sympathikus ist der Nerv der Spannung und Unruhe (fördert Kreislauf-Funktionen, "fight or flight"). Wichtige Ursprungszellen liegen im Rückenmark und seine Fasern verlaufen über den Grenzstrang links und rechts vom Rückenmark. Der Parasympathikus ist der Nerv der Entspannung und Ruhe (fördert Verdauungs-Funktionen, "feed or breed"). Wichtige Ursprungszellen liegen im Hirnstamm und im craniosakralen Rückenmark. Die Abbildung zeigt die Wirkungen des vegetativen Systems auf einzelne Organe. Paukert: Ein Fenster zum Ich – Teil 1 41 Die vegetativen Reflexbögen beginnen in den Rezeptoren (Mechano-, Thermo-, Chemo- und Schmerzsensoren) der glatten Muskulatur in der Wand eines inneren Hohlorgans. Der afferente Neurit führt zum sensorischen Neuron im Spinalganglion. Die Erregung wird weitergeführt über die Hinterwurzel in das Hinterhorn des Rückenmarkes. Dort erfolgt die Umschaltung auf das erste vegetative Neuron im Seitenhorn. Dessen efferenter Neurit (präganglionär) zieht über die Vorderwurzel aus dem Rückenmark zu einem vegetativen Ganglion (Nervenzellengruppe), wo die Weiterschaltung auf das zweite vegetative Neuron erfolgt (postganglionär). Sein efferenter Neurit zieht nun direkt zur Zielzelle des Erfolgsorganes (glatte Muskelfaser, Spezialmuskulatur des Herzens, Drüsenzelle). Im Gegensatz zu den Motoneuronen des zentralen Nervensystems können vegetative Signale in den Zielzellen sowohl hemmend (inhibitorisch) als auch anregend (exzitatorisch) wirken. Die Reaktion der Zielzelle hängt vom jeweiligen Erfolgsorgan ab, ist also weitgehend organspezifisch. Zur Erregungsübertragung vom postganglionären Nervenende auf die Zielzelle werden bestimmte Transmitterstoffe benötigt, die durch chemische Bindung an spezifischen Membranrezeptoren der Zielzelle die entsprechende Aktion auslösen (z.B. Aktionspotentiale zur Muskelfaserverkürzung). Damit ist der vegetative Reflexbogen abgeschlossen. Der Transmitter im präganglionären Neuron ist Acetylcholin. Die Transmitter im postganglionären Neuron sind Noradrenalin oder Adrenalin. Paukert: Ein Fenster zum Ich – Teil 1 42 [1.1.7] Das Hormonsystem Hypophyse und untergeordnete Hormondrüsen Hormone sind chemische Botenstoffe, welche in Senderzellen erzeugt werden und dann über den Blutweg auf Empfängerzellen einwirken. Die Steuerung erfolgt nach dem Regelkreisprinzip (feed back): Das Hormon regt die Empfängerzelle zur Produktion eines bestimmten Wirkstoffes an. Dieser gelangt über das Blut zurück zur Senderzelle und hemmt dort die weitere Erzeugung des Hormons. Das führt wiederum dazu, dass die Empfängerzelle weniger Wirkstoff produziert. Paukert: Ein Fenster zum Ich – Teil 1 43 Die Übersichttafel enthält alle wichtigen Hormone – ausgenommen der Sexualhormone. Paukert: Ein Fenster zum Ich – Teil 1 44 Die hormonale Regelung der Geschlechtsfunktionen Hypothalamus und Hypophyse regeln die Spermatogenese (Samenzellenreifung) in den Hoden des Mannes und die Oogenese (Eizellenreifung) in den Eierstöcken der Frau. Das so genannte follikelstimulierende Hormon (FSH) des Hypophysenvorderlappens fördert direkt Keimzellenbildung und -reifung. Es ist, wie auch die anderen hypophysären Hormone, bei beiden Geschlechtern gleich. Ein weiteres auf die Keimdrüsen wirkendes Hormon des Hypophysenvorderlappens ist das luteinisierende Hormon (LH). LH und FSH werden als Gonadotropine bezeichnet, weil sie auf die Geschlechtsdrüsen (Gonaden) wirken. Das GonadotropinReleasing-Hormon (GnRH) des übergeordneten Hypothalamus steigert Produktion und Freisetzung der Gonadotropine in der Hypophyse. Paukert: Ein Fenster zum Ich – Teil 1 45 Beim Mann verstärkt das LH die Testosteronbildung in den Leydig-Zwischenzellen im Nebenhoden, welche als Zellgruppen zwischen den Samenkanälchen liegen. Die Produktion und Freisetzung von Testosteron durch den Hoden erfolgt mithilfe eines Regelkreises, an dem Hypothalamus und Hypophyse beteiligt sind. Dabei werden durch einen Abfall von Testosteron im Blut zunächst das Gonadotropin-Releasing-Hormon im Hypthalamus und dann die Gonadotropine (LH, FSH) des Hypophysenvorderlappens vermehrt ausgeschüttet, was dann zur Steigerung der Testosteronbildung im Hoden führt. Testosteron wird außer im Hoden auch in der Nebennierenrinde sowie im Eierstock der Frau und in der Leber gebildet. Es steuert entscheidend die Entwicklung der männlichen Geschlechtsmerkmale, beeinflusst die sexuelle Aktivität und hat darüber hinaus anabolische Stoffwechselwirkungen (Proteinaufbau und damit Zunahme der Muskulatur). Bei der Frau stimulieren FSH und LH gemeinsam die Produktion der beiden Hormone Östrogen und Progesteron durch den Eierstock. Östrogen beeinflusst maßgeblich die Ausbildung der sekundären Geschlechtsmerkmale in der Pubertät. Dazu gehören Wachstum der Brust und die geschlechtsspezifische Verteilung von Unterhautfettgewebe. Beginnend mit der Pubertät (12. bis 15. Lebensjahr) reifen in den Eierstöcken die ersten Eizellen. Nach dem ersten Eisprung kommt es zur ersten Regelblutung, der Menarche. Danach stellt sich allmählich ein regelmäßiger Menstrualzyklus von ungefähr 28 Tagen ein. Der erste Tag der monatlichen Regelblutung (Menstruation) ist als erster Tag des Zyklus festgelegt. Der Zyklus entsteht durch ein kompliziertes Zusammenspiel verschiedener Hormone und Organe. Er besteht aus: Follikelphase (l. bis 12. Tag) Ovulationsphase (l3. bis 15. Tag) Lutealphase (l6. bis 28. Tag) Jede dieser Phasen ist durch charakteristische Hormonspiegel im Blut und Veränderungen in verschiedenen Organen (insbesonders in Ovar und Uterus, also in Eierstock und Gebärmutter) gekennzeichnet. Follikelphase: Zu Beginn der Follikelphase kommt es zur Menstruation. Diese beruht auf einer Abstoßung (Desquamation) eines großen Teils der Gebärmutterschleimhaut (Endometrium). Sie tritt immer dann ein, wenn die aus dem Eierstock freigesetzte Eizelle nicht befruchtet wird. Zu dieser Zeit steigt die FSHAusschüttung der Hypophyse an. Dies führt zu einer beschleunigten Follikelreifung und der Oogenese im Ovar mit gleichzeitiger Erhöhung der Östrogenproduktion durch die Granulosazellen der Follikel. Dabei reift jener Follikel, der am meisten FSH bindet und am meisten Östrogen produziert, zum sprungreifen Follikel (dominanter Follikel) heran. Paukert: Ein Fenster zum Ich – Teil 1 46 Unter Einfluss von Östrogen regeneriert die Schleimhaut der Gebärmutter durch Wucherung (Proliferation) von Bindegewebe, Drüsenschläuchen und Gefäßen (Proliferationsphase). Ovulationsphase: Der in der Follikelphase steigende Östrogenspiegel unterdrückt die FSH-Freisetzung der Hypophyse (negatives Feed-back) und fördert andererseits dort die LH-Produktion (positives Feed-back). Bei einem bestimmten Konzentrationsverhältnis von FSH zu LH erfolgt dann der Eisprung, wobei der Follikel platzt und die reife Eizelle im Eileiter abwärts zur Gebärmutter wandert. In dieser Phase beginnt die Erzeugung von Progesteron (Gestagen) durch den geplatzten Follikel, während die Östrogenausschüttung absinkt. Lutealphase: Nach dem Eisprung wandelt sich der zurückbleibende Follikelrest unter Einfluss von LH zum Gelbkörper (Corpus luteum). Er setzt steigende Mengen von Progesteron frei. Dieses Hormon verändert die Uterusschleimhaut. Die Drüsenschläuche verlängern sich und beginnen zu sezernieren (Sekretionsphase). Die Schleimhaut wird damit für die Einnistung einer befruchteten Eizelle vorbereitet. Progesteron führt in dieser Phase auch zu einem Anstieg der Körpertemperatur um etwa 0,5°C (Basaltemperatur) sowie durch Wassereinlagerungen zu einer Erhöhung des Körpergewichts. Bleibt eine Befruchtung aus, so kommt es gegen Ende der Lutealphase zu einer Rückentwicklung des Gelbkörpers und zu einer Einstellung der Progesteronausschüttung. In der Gebärmutter wird die äußere Schicht der Schleimhaut abgestoßen (Menstrualblutung). Durch anhaltende Erhöhung der Östrogen- und Progesteronkonzentration lässt sich die Freisetzung von GnRH des Hypothalamus und Gonadotropinen (LH, FSH) der Hypophyse hemmen und damit auch ein Eisprung im Eierstock unterbinden. Durch diese hormonale Ovulationshemmung kann eine Empfängnis (Konzeption) verhindert werden. Dazu nimmt die Frau über einen meist 28tägigen Zyklus Östrogen und Progesteron täglich in Form von Pillen ein (Antibaby-Pille). Zwischendurch wird die Hormoneinnahme kurzzeitig unterbrochen, sodass es zu einer Abstoßung der aufgebauten Gebärmutterschleimhaut (Abbruchblutung) kommt. Danach wird die künstliche Hormonzufuhr wieder fortgesetzt. Paukert: Ein Fenster zum Ich – Teil 1 [1.1.8] Informationsflüsse im Nervensystem (1) Reflexe und Instinktbewegungen: Rezeptoren - unbewusste Zentren (Hirnstamm, Rückenmark) - Effektoren. (2) Wahrnehmung und willkürliches Handeln: Rezeptoren - bewusste Zentren (Großhirnrinde) - Effektoren (Muskeln). (3) Gedächtnis, Denken und Motivation: Informationsfluss zwischen unbewussten und bewussten Zentren. 47 Paukert: Ein Fenster zum Ich – Teil 1 Wichtige Stationen und Bahnen zwischen Peripherie und Zentrum Reizaufnahme und Signalerzeugung in den Sinnesorganen. Reflexe zur unbewussten, schnellen Reizbeantwortung im Rückenmark. Aufsteigende sensorische Nervenbahnen im Rückenmark. Triebzentren zur Lebenserhaltung im Stammhirn. Emotionale Reizbewertung im Zwischenhirn und limbischen System. Filterung der sensorischen Erregungen durch Motive (Triebe, Emotionen). Bewusste Informationsverarbeitung in der Großhirnrinde. Leitung der bewussten, willkürlichen Motorik über die Pyramidenbahn. Leitung der unbewussten Motorik über das extrapyramidale System. Motorische Reaktion (Kontraktion) der Muskeln. 48 Paukert: Ein Fenster zum Ich – Teil 1 49 [1.1.9] Der Weg zum Bewusstsein Nur adäquate Reize lösen in den Sinnesorganen elektrochemische Erregungen aus (erster Filter). Diese sensorischen Signale gelangen über aufsteigende Nervenbahnen in den Thalamus im Zwischenhirn. Zugleich fließen die sensorischen Signale auch zum Hypothalamus und zum limbischen System und erregen dort jene Nervenzentren, in denen Triebe und Gefühle entstehen. Damit kommt es zu einer unbewussten (bzw. vorbewussten) emotionalen Bewertung der sensorischen Inputs. Diese Zentren senden ihrerseits Steuersignale zum Thalamus, wobei es zu einer neuerlichen Auswahl kommen kann (zweiter Filter). Die nunmehr gefilterten und emotional bewerteten sensorischen Inputs fließen vom Thalamus aufwärts in die entsprechenden Wahrnehmungszentren in der Großhirnrinde (Cortex). Gleichzeitig werden Erinnerungsspuren aktiviert. Erst hier in der Rinde des Großhirns entsteht Bewusstsein, d.h. ein bewusstes Wahrnehmungserlebnis. Unspezifische Wachheit Bewusste Erlebnisse in der Cortex sind nur dann möglich, wenn unspezifische Erregungen die Cortex aktivieren. Diese beginnen in einem netzartigen Nervengeflecht im verlängerten Rückenmark (formatio reticularis) und steigen über Mittelhirn und Thalamus zur Cortex auf (ARAS, aufsteigendes retikuläres Aktivierungssystem). Das ARAS wird durch sensorische Inputs eingeschaltet und bewirkt eine unspezifische Wachheit des Gehirns. Erst durch diese Aktivierung kann die Cortex spezifische Sinnesqualitäten (Qualia) bewusst erleben. Eine durch einoperierte Sonden erfolgte Ausschaltung des ARAS führt bei wachen Versuchstieren zur sofortigen Bewusstlosigkeit. Eine künstliche Elektrostimulation des ARAS führt bei schlafenden Versuchstieren zum sofortigen Aufwachen. Der Thalamus im Zwischenhirn ist ein außerordentlich wichtiger Teil des Gehirns. Einerseits strömen dort alle sensorischen Inputs von der Körperperipherie ein, aber auch Signale vom limbischen System und vom Hypothalamus. Auf diese Weise erfolgt eine unbewusste emotionale Bewertung der Inputs. Der Thalamus kann als das Vorzimmer zum Bewusstsein angesehen werden. Paukert: Ein Fenster zum Ich – Teil 1 50 Selektive Aufmerksamkeit und Bewusstsein als Systemfunktionen In den Thalamus als Vorzimmer der Hirnrinde münden auch absteigende Bahnen vom Stirnlappen der Hirnrinde. Über diese Leitungen erfolgt die Steuerung der selektiven Aufmerksamkeit, welche auf die zu den primären Rindenzentren aufsteigenden sensorischen Inputs einwirkt. Unter der selektiven Aufmerksamkeit versteht man die bewusste Einengung der Wahrnehmung auf bestimmte Inhalte. Die meisten Prozesse der Informationsverarbeitung im Nervensystem sind nicht bewusst (unbewusst), z.B. Reizaufnahme und Signalerzeugung in Sinnesorganen, Erkennen von Mustern (Gestalten), die emotionale Bewertung der sensorischen Signale, direkte und einfache motorische Reizbeantwortungen, usw. Bewusst hingegen sind die gefilterten Wahrnehmungen, neu zu erlernendes Verhalten, das Nachdenken und Entscheiden bei schwierigen Handlungsalternativen, usw. Wirft man drei Holzstäbe (z.B. von einem Mikadospiel) in die Luft, so fallen sie in einer zufälligen Anordnung zurück auf den Tisch. Ordnen sie sich dabei in der Gestalt eines Dreiecks an, dann treten neue Strukturmerkmale auf, die vorher nicht zu bemerken waren: z.B. Winkel oder Fläche. Jeder muss wohl zugeben, dass es völlig unsinnig ist, von einem Winkel eines einzelnen Stabes zu sprechen. Das System (Ganzheit), in unserem Beispiel das Dreieck, ist mehr als die Summe seiner Einzelteile (Übersummativität). Aus den Interaktionen der Teile resultieren neue Systemmerkmale. In diesem Sinne kann das Bewusstsein als ein ganzheitliches Funktionsmerkmal des zentralen Nervensystems verstanden werden. Bewusste Erlebnisse entstehen erst dadurch, dass sich Milliarden von Nervenzellen des Gehirns im Laufe der Evolution in einer besonderen Weise anordnen, vernetzen und interagieren. Über dieses primäre Bewusstsein hinausgehend hat das Gehirn noch die Möglichkeit der Selbstbewusstheit, d.h. es kann ein Protokoll über die in ihm laufenden bewussten Prozesse der Informationsverarbeitung führen, also ein Modell des eigenen Bewusstseins entwerfen (Reflexivität). Dieses sekundäre Bewusstsein bildet sich aber erst im Dialog mit anderen Gehirnen ("Ich weiß, dass Du weißt, dass Ich fühle .......... "). Das Bewusstsein kann sich nur durch Wechselwirkung mit anderen Gehirnen entwickeln. Damit wird aber Bewusstsein zu einem Teil des sozialen Miteinanders. Und mehr noch: Weil die am Dialog mit dem werdenden Gehirn teilhabenden Bezugspersonen (Eltern, Lehrer usw.) ihrerseits wieder stark von der Gesellschaft und der Kultur, in der sie leben, geprägt sind, erhält das Bewusstsein zur sozialen noch zusätzlich eine historische Dimension. Höheres Bewusstsein (Wahrnehmen, Analysieren und Bewerten des eigenen Bewusstseins) wird in dieser Sicht zu einem Entwicklungsprodukt nicht nur der biologischen, sondern auch der kulturellen Evolution. Paukert: Ein Fenster zum Ich – Teil 1 51 [1.1.10] Das Erkenntnisproblem "Bewusstsein" Eine Grunderfahrung ist, dass wir uns selbst als ein wahrnehmendes, bewertendes, entscheidendes, handelndes ICH erleben, dem ein freier Wille zugesprochen wird, der allen neuronalen Verarbeitungsprozessen vorangeht. Wir glauben an einer immateriellen (geistigen) Dimension teilzuhaben, die von den Phänomenen der dinglichen Welt gänzlich verschieden ist. Die immateriellen Erlebnisse unseres Gehirns scheinen uns genau so real zu sein wie die Phänomene der materiellen Außenwelt. Einerseits begreifen wir uns selbst als beseelte Wesen - andererseits erkennen wir, dass wir ein Teil der materiellen Natur sind, die sich evolutionär entwickelt, und welche mit einer objektiven, naturwissenschaftlichen Beschreibungssprache erklärt werden kann. Der so genannte Dualismus ist eine philosophische Theorie, welche die Existenz von zwei einander ausschließenden Erscheinungsformen annimmt, eine nicht materielle und eine materielle Welt. Sein wichtigster Vertreter war wohl René Descartes mit seinen "res cogitans" und "res extensa". Von ihm stammt auch der berühmte Satz "cogito, ergo sum" (ich denke, daher bin ich). Das ist die Meinung des Rationalismus, welcher der geistigen Welt den Vorrang gibt vor der realen materiellen Welt. Der Dualismus hat ein Hauptproblem, nämlich das Zusammenwirken von Geist und Materie zu erklären. Wie, wo und wann wird der Körper beseelt? Ist das bei der Befruchtung, bei der Geburt oder in späteren Entwicklungsphasen? Empirische Naturwissenschaften gehen zunächst von einer materiellen Welt von objektiv beobachtbaren und messbaren Dingen aus. Im Gegensatz zu René Descartes lautet der Leitsatz jetzt "sum, ergo cogito" (ich bin, daher denke ich). Die Biologie, besonders die klassische Verhaltensforschung, beobachtet das Verhalten von Lebewesen und versteht es als determiniert durch die genetische Organisation des jeweiligen Nervensystems und durch die individuelle Lerngeschichte, d.h. durch die Reizkonstellationen der jeweiligen Umwelt. Die verfeinerten Messmethoden der modernen Neurobiologie ermöglichen es, auch die höheren kognitiven Leistungen komplexer Gehirne objektiv zu analysieren. Das sind vor allem: Reize wahrnehmen und erinnern, mit selektiver Aufmerksamkeit bestimmte Reize filtern und andere unterdrücken, zwischen verschiedenen Reaktionsoptionen entscheiden, Belohnungen und Bestrafungen erkennen, soziale Bindungen herstellen und diese mit Affekten aufladen, Emotionen erleben, usw. Paukert: Ein Fenster zum Ich – Teil 1 52 Alle diese kognitiven Leistungen werden als emergente Funktionen von komplexen neuronalen Vorgängen verstanden, d.h. sie werden mit den physikalischchemischen Interaktionen in den Nervennetzen zwar nicht gleichgesetzt, aber sie gehen kausal erklärbar aus diesen hervor. Die modernen Neurowissenschaften sehen sich drei Hauptfragen gegenüber, von denen der Dualismus behauptet, dass sie aus naturwissenschaftlicher Sicht nicht beantwortet werden können. (1) Wie ist die so genannte Qualia erklärbar? Darunter versteht man die spezifische Qualität subjektiver Sinnesempfindungen, beispielsweise die Farbqualität von "Blau" oder den Geschmack von "Süß" oder den unverwechselbaren Klang eines Saxophons. (2) Wie ist die Selbstwahrnehmung (Ichbewusstsein, Selbstreflexion) möglich? Darunter versteht man die Grunderfahrung, dass wir uns selbst als ein wahrnehmendes, bewertendes, entscheidendes und handelndes ICH wahrnehmen. Unser Bewusstsein konstruiert ein Modell von sich selbst. (3) Wie kann der intuitive Glaube an den eigenen freien Willen erklärt werden? Darunter versteht man die grundsätzlich freie Entscheidungsmöglichkeit für verschiedene Handlungsalternativen. Im Folgenden werden verschiedene wissenschaftliche Erklärungsversuche dieser drei Phänomene (Qualia, Selbstwahrnehmung und Willensfreiheit) vorgestellt. QUALIA Der Neurobiologe Francisco Varela führte folgendes Experiment durch: Er bat Versuchspersonen (Vpn) Schwarz-Weiß-Bilder anzuschauen, von denen einige Profilansichten von Gesichtern darstellten. Während die Vpn versuchten, in diesen Bildern Gestalten zu erkennen, wurden mit einem dichten Netz von Elektroden Hirnströme gemessen. Die Vpn mussten durch Drücken einer Taste angeben, ob sie ein Gesicht erkannt hatten. Jedes Mal, wenn dies der Fall war, wurden über den Hirnrindenarealen, welche sich mit dem Sehen befassen, kurzfristige (einige Zehntel-Sekunden andauernde) hochsynchrone Wellen mit einer Frequenz von etwa vierzig Hertz registriert. Dies war nicht der Fall, wenn die Vpn die vorgelegten Bilder nicht als Gesichter identifizieren konnten. Paukert: Ein Fenster zum Ich – Teil 1 53 Wenn sensorische Inputs zu einer bewussten Wahrnehmung zusammengefügt (rekonstruiert) werden, dann synchronisieren jene Neuronen, welche sich in der Hirnrinde mit der Verarbeitung dieser Inputs befassen, ihre Entladungen über kurze Zeitspannen. Die zeitlich koordinierte und synchrone Aktivität einer sehr großen Anzahl von räumlich verteilten Nervenzellen charakterisiert somit eine bewusste Sinnesempfindung und ist damit das neurophysiologische Korrelat der Qualia. Unsere Sinne liefern oft nur unvollständige und lückenhafte Informationen an die Großhirnrinde. In diesen Fällen vervollständigt und ergänzt unser Gehirn mit Hilfe von gespeichertem Vorwissen die Wahrnehmung. Es rekonstruiert die Wahrnehmung. Das Gleiche gilt auch für unsere Denkprozesse. Parallel zu den neurobiologischen Untersuchungen über Wahrnehmungsprozesse wurden auch Untersuchungen über die Willkürmotorik durchgeführt. In einem Experiment von Benjamin Libet wurden die Vpn aufgefordert, zu beliebigen Zeitpunkten innerhalb der Versuchszeit ihre Handgelenke zu bewegen während sie einen schnell laufenden Lichtzeiger beobachteten. Dieser durchlief in 2.56 Sekunden einen Kreis. Sofort wenn eine Vp sich bewusst für eine Bewegung entschied, musste sie die Stellung des Zeigers notieren. Dadurch wurde der Zeitpunkt einer Entscheidung genau registriert. Gleichzeitig wurden die Gehirnaktivtäten mittels EEG aufgezeichnet. Die Muskelbewegungen wurden mittels EMG (Elektromyogramm) aufgezeichnet. Es wurde nun festgestellt, dass die für die Bewegungen verantwortlichen Gehirnaktivitäten etwa eine halbe Sekunde VOR jenen Zeitpunkten einsetzten, in denen sich eine Vp bewusst entschied ein Handgelenk zu bewegen. Dieses und auch viele ähnliche Experimente beweisen, dass einer bewussten Entscheidung für eine bestimmte Bewegung bereits eine dafür verantwortliche unbewusste Gehirnaktivtät zeitlich vorangeht (Bereitschaftspotential). Damit ist aber auch bewiesen, dass die bewusste Willensentscheidung für eine Bewegung nicht die alleinige Ursache für die dann ausgeführte Bewegung sein kann. Eine freie Willensentscheidung im engeren Sinne scheint daher eine Illusion zu sein. SELBSTWAHRNEHMUNG Die neuronalen Netze können trainiert werden oder sie können auch selbstorganisierend sein. In den selbstorganisierenden Neuronennetzen passen sich die Synapsenstärken schrittweise den einlangenden Inputmustern an. Die stummen und feuernden Neuronengruppen werden zu einem Abbild der Inputmuster, so dass gleiche Inputreize immer dieselben Neuronengruppen erregen. Paukert: Ein Fenster zum Ich – Teil 1 54 Das neuronale Netz ist dann zu einer Landkarte (map) der Reizumwelt geworden. Zusätzlich kommt es auf der neuronalen Eigenschaftskarte zu einer Reduzierung der Dimensionalität. Aus der Reizvielfalt werden einige wenige Haupteigenschaften extrahiert. Das sind diejenigen, in denen die Inputmuster am deutlichsten variieren. So können auch ähnliche Inputreize dieselben Neuronen erregen. Diese Abstraktionsleistung ermöglicht erst die Bildung von Kategorien und ist wesentlich für das begriffliche Denken. Mit ihrer Hilfe werden beispielsweise einfache Kategorien (Begriffe) wie "rund", "eckig", "schwer" oder "leicht" gebildet. Es gibt keinen Grund gegen die Annahme, dass Selbstwahrnehmung und Selbsterfahrung auf neuronalen Vorgängen beruhen. Ein erstes wichtiges Faktum ist, dass die meisten neuronalen Informationsverarbeitungen unbewusst ablaufen. Wir haben beispielsweise keinen bewussten Zugriff zu Informationen über unseren Blutdruck oder über unseren Blutzuckerspiegel, obgleich diese Variablen sehr sorgfältig gemessen, vom Gehirn ausgewertet und in Regulationsprozesse umgewandelt werden. Der wahrscheinliche Grund hierfür ist, dass diese Informationen ohne Beteiligung der Großhirnrinde verarbeitet werden. Aber auch von den in der Großhirnrinde ablaufenden Prozessen wird uns immer nur ein kleiner Ausschnitt bewusst - das sind jene Aspekte, denen wir unsere Aufmerksamkeit schenken und die dann unser Handeln steuern. Auch die unbewussten Prozesse hinterlassen Gedächtnisspuren und beeinflussen unser zukünftiges Handeln. Aber wir werden uns dieser Handlungsdeterminanten nicht bewusst und können sie deshalb nicht als Gründe für unser Tun anführen. Neben diesen unbewussten Verarbeitungsprozessen im Gehirn ist eine zweite Voraussetzung für die Konstitution eines Selbst (ICH) die sogenannte soziale Interaktion. Für die Entwicklung des Selbstmodells scheint der Dialog mit den Anderen wesentlich. Dialoge der Form "Ich weiß, dass Du weißt, dass Ich fühle" führen dazu, dass wir uns in der Wahrnehmung des Anderen spiegeln. Mit Hilfe seiner Reaktionen (re)konstruiert dann das Gehirn das eigene Selbst. Damit ein effektiver zwischenmenschlicher Dialog stattfinden kann, müssen aber zwei Prämissen erfüllt sein: Erstens muss das abstrakte Denken entsprechend hoch entwickelt sein. Zweitens muss die sprachliche Codierung von Vorstellungen und Denkinhalten möglich sein. Mit Ausnahme der großen Menschenaffen fehlen den Tieren diese Fähigkeiten. Auch kleine Menschenkinder besitzen sie noch nicht, weil für diese höchsten kognitiven Leistungen bestimmte Hirnstrukturen vor allem in der Großhirnrinde erforderlich sind, die erst beim Menschen im Laufe der ersten Lebensjahre ihre volle Ausprägung erfahren. Paukert: Ein Fenster zum Ich – Teil 1 55 Somit kann die Selbstwahrnehmung (eben unser Ichbewusstsein) als eine höchst komplexe kognitive Leistung von selbstorganisierenden neuronalen Netzen verstanden werden, wobei viele unbewusste Verarbeitungsprozesse mitspielen und die soziale Kommunikation ein wesentlicher Faktor ist. Zur Erklärung der Selbstwahrnehmung ist es nicht nötig, eine Transzendenz in immaterielle Sphären zu postulieren. Auch wäre der Schluss falsch, dass hinter den kognitiven Leistungen des Gehirns ein Dirigent, eine Führungsinstanz steht, welche die neuronalen Prozesse steuert und lenkt. Die neuronalen Verarbeitungsprozesse laufen parallel und dezentral ab und erzeugen in ihrer Gesamtheit kohärente Wahrnehmungen und Handlungen. Zwar gibt es verschiedene hierarchische Gliederungsebenen im Nervensystem mit reziproken Kopplungen, aber es gibt kein eigenes übergeordnetes Koordinationszentrum der parallelen Prozesse. An dieser Stelle sei auch noch ein Querverweis auf das biochemische Geschehen in der Zelle erlaubt. Schon auf molekularer Ebene gibt es so etwas wie Selbstanalyse und Selbstreparatur. Wenn bei der identischen Reduplikation eines DNS-Moleküls (des Trägermoleküls der Gene) Fehler passieren, dann gibt es Hilfsmoleküle (spezialisierte Enzyme), welche das DNS-Molekül analysieren und etwaige Fehler reparieren. Schon hier liegt eine elementare Form der Selbstwahrnehmung der einzelnen Zellen vor. WILLENSFREIHEIT Wie aber kommen wir nun zu der unerschütterlichen Überzeugung, dass unser Selbst (ICH) freie Entscheidungen treffen und über Prozesse in unserem Gehirn verfügen kann, welche dann unser Handeln steuern ? Eine erste und vermutlich entscheidende Erfahrung mit der Zuschreibung von Autonomie und Freiheit machen wir schon als Kleinkinder. Eltern sagen ihren Kindern fortwährend sie sollten dies tun und jenes lassen, weil andernfalls diese oder jene Konsequenzen eintreten würden. Diese Verweise und die mit ihnen verbundenen Sanktionen legen den Schluss nahe, man könne sich auch anders verhalten und müsse dazu nur wollen. Wir erfahren (erleiden) also schon sehr früh eine Behandlung, welche sich durch die Annahme rechtfertigt, wir seien in unseren Entscheidungen frei - eine Annahme, die mit Hilfe der Erziehung verlässlich von Generation zu Generation übermittelt wird. Wir internalisieren diese Annahme der Willensfreiheit und handeln dann in unserem Leben auch nach ihr. Paukert: Ein Fenster zum Ich – Teil 1 56 Die Dialoge, die ein Wertesystem vermitteln, beginnen in einer Entwicklungsphase, in der Kleinkinder noch kaum ein deklaratives Gedächtnis haben. Sie lernen, machen sich das Gelernte zu Eigen. Sie können aber nicht angeben, woher sie wissen, was sie wissen, d.h. sie erinnern sich zwar an das Gelernte, nicht aber an den Lernprozess. Diese Unfähigkeit, den Kontext von Erlebnisinhalten bewusst zu erinnern, bezeichnet man als frühkindliche Amnesie. So erscheint den Kleinkindern das, was sie wissen als nicht verursacht, als immer schon gewusst. Diese frühkindliche Amnesie könnte der Grund dafür sein, dass uns später, wenn wir beginnen über uns nachzudenken, die Inhalte des frühkindlichen Lernens als nicht verursacht erscheinen. Die Annahme einer Willensfreiheit erscheint als eine durch die Erziehung vermittelte Attribution. Die fehlende Erinnerung an diese frühen sozialen Lernprozesse könnte somit der Grund für die eigentümliche Transzendenz unseres Selbstmodells sein, für den Glauben an einen freien Willen, welcher unverursacht allen materiellen Prozessen vorausgeht. Bemerkenswert ist, dass wir - trotz aller Überzeugung grundsätzlich frei zu sein in der Bewertung des eigenen und fremden Verhaltens sehr wohl zwischen freien und unfreien Akten unterscheiden. Für erstere übernehmen wir Verantwortung, für zweitere fordern wir Nachsicht. Dieser Unterschied wird bewirkt durch den Bewusstheitsgrad der Motive. Nur jene Motive erscheinen dem freien Willen unterworfen, die wir auch bewusst erkennen. Jene Motive aber, die unbewusst wirken, unterliegen offenbar nicht dem freien Willen. Aus der Sicht der modernen Neurowissenschaft sind die Annahme einer Willensfreiheit und die Unterscheidung von freien und weniger freien Handlungen nicht haltbar. Allen kognitiven Leistungen des Gehirns, so auch dem Entscheiden, gehen Verarbeitungsprozesse in neuronalen Netzen voran. Diese determinieren unseren Handlungsspielraum. Die Experimente von Benjamin Libet bestätigen dies sehr eindrucksvoll. Das Gehirn besteht aus Milliarden von Nervenzellen. Eine Nervenzelle kann mit tausenden anderen Nervenzellen verbunden sein. In hoch organisierten Gehirnen machen die Eingänge von den sensorischen Rezeptoren und die Ausgänge zu den motorischen Effektoren nur einen verschwindend kleinen Prozentsatz der Verbindungen aus. Die meisten Verbindungen kommen von anderen Nervenzellen. Das Gehirn beschäftigt sich also vorwiegend mit sich selbst. Das Gehirn kann die eigene Tätigkeit, seine eigenen kognitiven Prozesse analysieren und bewerten. Paukert: Ein Fenster zum Ich – Teil 1 57 Aus der wiederholten Anwendung der Kognition auf sich selbst resultiert dann die Selbstwahrnehmung (Ichbewusstsein). Willensentscheidungen sind mehrstufige Prozesse, bei denen sich immer das stärkste Motiv durchsetzt. Soll ein Motiv einem anderen vorgezogen werden, dann muss es auch gestärkt werden - beispielsweise durch überzeugende Argumente in einem sozialen Dialog. Wir sind zwar im neurobiologischen Mechanismus des Entscheidungsprozesses determiniert, aber sowohl durch äußere Einflüsse als auch durch innere Denkakte kann es zu einer Umordnung der Motivstärken, zu einer Neuorientierung des erlernten Wertesystems kommen. Aber damit verschiebt sich nur die Determiniertheit. Ein nicht determiniertes, freies Ich ist eine Illusion. Trotz aller Determiniertheit unseres Wollens und Tuns haben wir einen gewissen Spielraum. Wir können lernen unser Handeln und unsere Motive zu hinterfragen. Wir können lernen selbstanalytisch und selbstkritisch zu agieren und gewinnen damit Selbstdistanz, d.h. eine Distanz zu unseren Trieben und Zwängen. Dadurch verlieren diese ihre unmittelbare Macht. Eine kritische Selbsterkenntnis wird nur mühsam errungen, kann aber zu jenem Spielraum führen, der ein verantwortungsvolles Entscheidungsverhalten ermöglicht. Soziale Verantwortung erscheint somit durchaus kompatibel mit der Determiniertheit des Menschen. Das trainierte, sich selbst reflektierende Bewusstsein erarbeitet sich ein Vetorecht, welches ihm erlaubt, sich dem Zwang der subkortikalen Triebe und Gefühle zumindest teilweise zu entziehen. Befriedigungsaufschub und auch Frustrationstoleranz sind trainierbar. Paukert: Ein Fenster zum Ich – Teil 1 58 [1.2] EIN BASISMODELL DER PSYCHE Der Wiener Psychologe Hubert Rohracher (1903 - 1972) gliedert die Inhalte der Psyche in zwei Erlebnisklassen: Psychische Kräfte sind die angeborenen Instinkte und Triebe, die erlernten Interessen, die Gefühle (Emotionen) und die Willenserlebnisse. Sie werden subjektiv als drängend und zielsetzend erlebt. Psychische Funktionen sind die Wahrnehmung, das Gedächtnis (Lernen) und das Denken und Sprechen. Sie sind Werkzeuge zur Erreichung der gesetzten Ziele. Der Zusammenhang wird durch das Prinzip der funktionalen Aktivierung erklärt: es gibt keine funktionale Aktivität ohne einen inneren Antrieb oder einen äußeren Anreiz. Psychische Funktionen Wahrnehmung: Gedächtnis (Lernen): Denken: Sprechen: Aufnehmen von Informationen. Speichern und Abrufen von Informationen. Verknüpfen von Informationen zur Problemlösung. Weitergeben von Informationen durch phonetische Artikulation von bewussten Erlebnisinhalten. Psychische Kräfte Instinkte und Triebe: Angeborene Drangzustände, die überwiegend zu lebenserhaltenden Aktionen führen. Interessen: Zumeist erlernte Drangzustände, die zur Ausführung kultureller Aktionen streben. Gefühle (Emotionen): Reaktive Erlebniszustände auf äußere oder innere Reize, die angenehm oder unangenehm erlebt werden. Wollen: Bewusste Entscheidungserlebnisse. Paukert: Ein Fenster zum Ich – Teil 1 59 Als Beispiel sei die Situation nach intensiver Sportbetätigung angeführt. Durch den Flüssigkeitsverlust beginnt über einen Regelkreismechanismus ein Nervenzentrum (Durstzentrum) im tiefer gelegenen Stammhirn zu feuern, d.h. elektrochemische Erregungen zu produzieren. Diese steigen höher in das Großhirn und erzeugen dort das unlustvolle Trieberlebnis des Durstes. Dadurch werden die Wahrnehmung, das Gedächtnis und das Denken aktiviert, um in der Umwelt nach durstlöschenden Objekten zu suchen. Nach deren Auffindung kommt es zur lustvoll erlebten Triebbefriedigung (Trinken). Dabei werden über entsprechende motorische Steuerungen passende Verhaltensweisen ausgeführt. Die Emotionen Lust und Unlust dienen als sinnvolle Triebverstärkungen. Ursprünglich sind die gestellten Handlungsziele auf Lebens- und Arterhaltung gerichtet. Mit Hilfe des psychischen Apparates, insbesondere seiner Denkleistungen hat sich das menschliche Gehirn einen entscheidenden Leistungsvorteil im täglichen Daseinskampf geschaffen. Die bewussten Erlebnisse sind Systemfunktionen von komplex vernetzten, gegenseitig miteinander gekoppelten und hierarchisch gegliederten Teilbereichen des zentralen Nervensystems, insbesondere der Großhirnrinde (Cortex). Zentrales Nervensystem und Bewusstsein haben sich evolutionär entwickelt und dienen letztendlich der optimalen Anpassung an die Umwelt. Unter Psyche versteht man die Gesamtheit der bewussten Erlebnisse, aber auch der nicht bewussten (unbewussten) Vorgänge im zentralen Nervensystem. Der Begriff Seele wird hier ausschließlich als Synonym für eine so verstandene Psyche verwendet. Entsprechend der stammesgeschichtlichen Entwicklung des Gehirns und des zentralen Nervensystems durchläuft die Psyche unterschiedliche Ausbildungsstufen. Immer dann, wenn einfachere Formen sensomotorischer Informationsverarbeitung (z.B. Reflexe) zur Steuerung und Kontrolle des Organismus nicht mehr ausreichen, hat sich eine höhere und leistungsfähigere Funktionsebene entwickelt. Bei einfachen Reflexen wird ein Reiz von den peripheren Sensoren (Sinnesorganen) aufgenommen und in eine spezifische Folge von elektrischen Spannungsschwankungen verschlüsselt. Diese wird entlang von Nervenfasern über das Rückenmark oder das Stammhirn zu den Effektoren (Muskeln, Drüsen) weitergeleitet, wo der Reiz durch entsprechende motorische Reaktionen beantwortet wird. Etwas komplexer gestaltet sich der Ablauf einer Instinktreaktion. Hier fließt der Informationsstrom über höher gelegene Stammhirn-Zentren im zentralen Nervensystem. Als Beispiel sei das Hinaustreten aus einem dunklen Raum in das helle Sonnenlicht genannt. Zunächst erfolgen reflektorische Reaktionen (Pupillenreflex), dann instinktive Schutzreaktionen (Heben der Hände zum Augenschutz) und schließlich noch komplexere Verhaltensweisen (Aufsetzen einer Sonnenbrille). Paukert: Ein Fenster zum Ich – Teil 1 60 Reichen reflektorisches und instinktives Verhalten zur Problemlösung nicht aus, dann erfolgt die Informationsverarbeitung in den noch höher gelegenen Zentren in der äußeren Rinde des Großhirns (Cortex). Hier werden unsere Wahrnehmungen bewusst erlebt und Handlungen bewusst veranlasst. Allen diesen bewussten Erlebnissen liegen spezifische Erregungskonstellationen zugrunde, welche in wechselseitig gekoppelten Gruppen von Nervenzellen (neuronale Ensembles) ablaufen. Schließlich ziehen dann die entsprechenden elektrischen Signalfolgen über die so genannte Pyramidenbahn abwärts zu den Muskeln und steuern dort die willkürlichen Handlungen. Damit ist die höchste Stufe sensomotorischer Regelkreise erreicht: Reize werden selektiv wahrgenommen und mit motorischen Reaktionen bewusst beantwortet. Das Ergebnis des Verhaltens wird wiederum wahrgenommen (Feed-back) und führt zu neuerlichen Reaktionen, usw. Ein sensomotorischer Prozess wird bewertet (z.B. eine erfolgreiche Nahrungssuche), wenn Signale zu jenen tiefer gelegenen Nervenzentren an der Basis des Großhirns und im Zwischenhirn übermittelt werden, wo Gefühle und Triebe entstehen (limbisches System, Hypothalamus). Diese bewirken eine Bewertung (z.B. Lust - Unlust) und eine Selektion der Information. Die Filterung erfolgt im so genannten Thalamus im Zwischenhirn, das zwischen Stammhirn und Großhirn liegt. So wird beispielsweise beim sehnsüchtigen Warten auf das Eintreffen eines geliebten Menschen der Wahrnehmungsfilter durch psychische Kräfte wirksam. Zum Zeitpunkt des Erscheinens der erwarteten Person werden andere Reize (z.B. irgendein Vorfall in der näheren Umgebung) kaum wahrgenommen - die Wahrnehmung engt sich auf das entsprechende Objekt ein. Auf dem Weg vom Sensor an der Körperperipherie bis zur Rinde des Großhirns, also vom Reiz bis zu seiner bewussten Wahrnehmung, wird die Informationsmenge drastisch reduziert. Erstens werden durch die Bauart des Sensors nur adäquate Reize aufgenommen und zweitens erfolgt eine Selektion der aufgenommenen Information durch die psychischen Kräfte, d.h. eine Sensibilisierung der Wahrnehmung. Weil jeder sensorische Input emotional bewertet wird, kommt es immer zu einer Filterung der Information. Daher haben verschiedene Individuen auch unterschiedliche bewusste Bilder von ein- und demselben Bereich der Außenwelt. Was subjektiv für wirklich gehalten wird, hängt somit von den individuellen Gefühlen, Trieben und Interessen ab. Die daraus resultierende Divergenz der subjektiven Weltbilder führt nicht selten zu Konflikten. Starke triebhafte oder emotionale Bewertungen von Inhalten und wiederholte Rückkopplungen (Feed-back) von motorischem Verhalten und Sinnesrezeptionen führen zur Ausbildung von so genannten synaptischen Verstärkungen in den beteiligten Nervenbahnen. Dadurch werden die molekularen Grundlagen für das Gedächtnis geschaffen. Paukert: Ein Fenster zum Ich – Teil 1 61 [1.3] METHODEN DER GEHIRNFORSCHUNG [1.3.1] Stereotaktische Eingriffe (Läsion und Stimulation) Mit Hilfe eines dreidimensionalen Hirnatlas und einem stereotaktischen (räumlich ausgerichteten) Apparat werden Elektroden oder Kanülen, die auf bestimmte Zielgebiete in der Tiefe des Gehirns gerichtet sind, durch eine Operation eingesetzt. Über Elektroden kann elektrischer Strom in das Zielgebiet geschickt werden, durch die Kanülen werden bestimmte chemische Substanzen eingebracht. Das Zielgebiet kann irreversibel oder reversibel ausgeschaltet werden. Bei der irreversiblen Läsion wird durch die isolierte Elektrode hochfrequenter Wechselstrom geschickt, der durch Hitzeentwicklung an der Spitze das umgebende Gewebe zerstört (Elektrokoagulation). Selektiver wirken chemische Läsionen, die nur die Zellkörper zerstören, aber nicht deren Ausläufer. Verwendet werden dazu Kainsäure und auch Ibotensäure, die über eine Kanüle in das Zielgebiet eingeschleust werden. Reversible Läsionen erfolgen durch Kühlung mit Hilfe kleiner Mengen von Kaliumchlorid-Lösungen. Sinn der Läsion eines umgrenzten Hirngebietes ist es, jene Veränderungen zu registrieren, die nach der Läsion auftreten. Paukert: Ein Fenster zum Ich – Teil 1 62 Im Gegensatz zu Läsionen werden bei Stimulationen die Zielgebiete nicht ausgeschaltet sondern künstlich gereizt. Die Reizung erfolgt über eine isolierte Elektrode mittels genau abgestimmter elektrischer Stromzufuhr. Es können aber auch winzige Mengen von chemischen Substanzen über Kanülen oder Mikropipetten an die Synapsen transportiert werden, die dort als Agonisten oder Antagonisten von bestimmten Neurotransmittern wirken. Sinn der Stimulation eines umgrenzten Hirngebietes ist es, jene Veränderungen zu registrieren, welche bei der Stimulation auftreten. Durch solche gezielte Läsionen und Stimulationen kann der kausale Zusammenhang von bestimmten Hirnregionen und bestimmten Verhaltensweisen bzw. physiologischen Funktionen sehr gut erforscht werden. [1.3.2] Enzephalogramme (EEG und MEG) Das Elektroenzephalogramm (EEG) 1902 begann Hans Berger mit Experimenten an Hunden und Katzen, die ihn soweit führten, daß er 20 Jahre später über außen an der Schädeldecke befestigte Elektroden an einem siebenjährigen Patienten mit Hilfe eines empfindlichen Saitengalvanometers erstmals spontane elektrische Spannungsschwankungen der Hirnrinde registrierte. Mindestens 2 Elektroden aus Silberlegierung werden an die Schädeldecke mit einer Paste angeklebt. Sie sind mit einem Verstärker verbunden, der die elektrische Potentialdifferenz der beiden Orte verstärkt und an ein Aufzeichnungsgerät weitergibt. Diese Messungen und Aufzeichnungen erfolgen in bestimmten vorgegebenen zeitlichen Abständen. Dadurch ist es möglich, den elektrischen Spannungsverlauf zwischen den beiden Elektroden zeitlich zu erfassen. Zusätzlich werden sehr langsame Schwankungen und auch die Gleichspannungsanteile herausgefiltert, so dass nur Wechselspannungen in einem Amplitudenbereich von 1 - 200 mV und in einem Frequenzbereich von 1 - 50 Hz registriert werden. Anstelle von nur 2 Elektroden werden zumeist die zeitlichen Spannungsänderungen zwischen mehreren Elektroden aufgezeichnet, die an typischen Schädelpositionen angebracht sind. (Polygraph mit bis zu 20 Ableitungspunkten). Das mit Elektroden abgenommene elektrische Potential entsteht durch die Summierung der exzitatorischen postsynaptischen Potentiale in der obersten Rindenschicht des Großhirns. In Wirklichkeit werden die cortikalen Neuronen durch vom Thalamus aufsteigende Erregungen angetrieben. Der Thalamus wirkt somit als Tor für sensorische Impulsströme. Die Durchlässigkeit dieses Tores hängt von den inhibitorischen Signalen ab, welche über absteigende Leitungen vor allem vom präfrontalen Cortex (Stirnlappen) ankommen. Paukert: Ein Fenster zum Ich – Teil 1 63 Durch den thalamo-cortikalen Feedback wird die selektive Aufmerksamkeit reguliert. Je größer die Hemmung des Thalamus ist, umso langsamer und synchronisierter sind die EEG-Wellen. Je ungehemmter der Thalamus ist, umso desynchronisierter und heftiger verlaufen die EEG-Schwankungen. Je nach Wachheitsgrad des Bewußtseins treten verschiedene Wellenformen im EEG auf. Im entspannten Wachzustand herrscht der Alpha-Rhythmus (8-13 Hz) vor. Bei selektiver Aufmerksamkeit (visuell und auditiv) wird er blockiert und geht in den höher frequenten Beta-Rhythmus (13-30 Hz) über. Frequenzen über 30 Hz werden als Gamma-Wellen bezeichnet. Sie haben eine sehr kleine Amplitude (1-10 mV) und sehr hohe lokale Spezifität. Sie werden mit sich lokal entladenden Zellenensembles (vernetzten Neuronengruppen) in Verbindung gebracht. Die Theta-Wellen (4-8 Hz) und Delta-Wellen (unter 4 Hz) sind typisch für den Schlaf. Beim Träumen werden die langsamen Delta-Wellen durch Thetaund Beta-Phasen unterbrochen (REM-Phasen: rapid eye movement, schnelle Augenbewegungen). Im klinischen Bereich wird das EEG zur Diagnose und zur Lokalisation von Funktionsstörungen der Gehirntätigkeit verwendet: bei epileptischen Anfallsleiden, bei Tumoren, bei Durchblutungsstörungen und zur Abschätzung von Pharmakawirkungen und Narkosetiefen. Unter evozierten, ereigniskorrelierten Hirnpotentialen (EKP) versteht man alle elektrocortikalen Potentialschwankungen, welche kurz vor, während und kurz nach einem sensorischen, motorischen oder psychischen Ereignis im EEG messbar sind. Diese EKPs sind von wesentlich kleinerer Amplitude als das Standard-EEG. Durch spezielle mathematische Mittelungstechniken werden die EKP-Signale aus der Hintergrundaktivität (Rauschen) hervorgehoben und graphisch dargestellt. Diese Registrierungs- und Berechnungstechniken werden von Computern schnell und zuverlässig durchgeführt. Das Magnetenzephalogramm (MEG) Jede elektrische Ladungsdifferenz zwischen zwei Orten (Dipol) erzeugt eine Bewegung von elektrischer Ladung (Stromfluss). Jeder solcher Stromfluss ruft in seiner Umgebung ein magnetisches Kraftfeld hervor, dessen Feldlinien kreisförmig um die Stromrichtung verlaufen. Die hintereinander geschalteten Neuronen in der äußeren Rindenschicht wirken wie elektrische Dipole und erzeugen daher auch schwache Magnetfelder, die mit hochempfindlichen Detektoren, so genannten SQUIDS (superconducting quantum interference device) nachgewiesen werden können. Paukert: Ein Fenster zum Ich – Teil 1 64 Die SQUIDS werden ca. 10 mm über der Schädeldecke angebracht und registrieren magnetische Feldstärkenveränderungen von weniger als einem Hundertmillionstel der erdmagnetischen Feldstärke (also kleiner als ein Pico Tesla). Mit den auf der Temperatur des flüssigen Helium gehaltenen SQUIDS wurde 1968 erstmals Alpha-Aktivität und ab 1975 auch ereigniskorrelierte Aktivität registriert. Da mit dem MEG aus technischen Gründen nur zur Schädeldecke parallele elektrische Ströme und mit dem EEG meist nur zur Schädeldecke vertikal laufende Ströme gemessen werden, lassen sich durch Kombination von MEG und EEG Quellen elektrischer Aktivität in der Cortex bis auf 2 mm Genauigkeit lokalisieren. Dadurch wird eine räumliche und zeitliche Bestimmung der gehirnelektrischen Aktivität sehr genau ermöglicht. [1.3.3] Bildgebende Verfahren (CT, PET und MRT) Zur Ergänzung von EEG und MEG, die elektrische und magnetische Aktivitäten in der Gehirnrinde registrieren, werden auch Bild gebende Verfahren in der Gehirnforschung eingesetzt. Dabei werden verschiedene Hirnschichten röntgenologisch gescannt (CT, Computertomographie), oder die regionale Hirndurchblutung mit Hilfe von radioaktiven Markierungssubstanzen gemessen (PET, Positronen-Emmissions-Tomographie), oder der Blutdurchfluss in bestimmten Arealen über die dabei verstärkten kernmagnetischen Resonanzen von Wasserstoffionen registriert (MRT, Magnetische Resonanz-Tomographie). Die Messergebnisse werden durch einen Computer ausgewertet und dann schattierte oder verschieden gefärbte grafische Bilder der Hirnregionen hergestellt. Mit diesen Bildern lassen sich neuronale Aktivitäten auch in tieferen Hirngebieten sehr gut darstellen. Der Nachteil liegt in ihrer trägen zeitlichen Auflösung (meist werden nur Prozesse, die Minuten dauern, sichtbar). Zum Abschluss sollen diese Bild gebenden Verfahren näher beschrieben werden. Röntgen-Computertomographie (CT) Röntgenstrahlen entstehen beim Aufprall hoch beschleunigter Elektronen auf einer Schwermetall-Anode. In der so genannten Röntgenröhre emittiert eine Glühkathode Elektronen, die durch eine angelegte Spannung von etwa 30 kV zur gegenüberliegenden Anode fliegen. Durch umgebende negativ geladene Platten werden die Elektronen gebündelt (Wehnelt-Zylinder). Beim Aufprall auf der Anode erreichen sie Geschwindigkeiten von rund 100 000 km/sec.. Durch die Abbremsung beim Aufprall wird Energie in Form von kurzwelligen Röntgenstrahlen freigesetzt (10 -10 m Wellenlänge). Paukert: Ein Fenster zum Ich – Teil 1 65 Eine der wichtigsten Eigenschaften von Röntgenstrahlen ist ihr hohes Durchdringungsvermögen. Sie durchdringen viele Stoffe (Papier, Holz, Fleisch) fast ungeschwächt und werden von Materialien absorbiert, welche chemische Elemente mit hoher Ordnungszahl enthalten. Dazu zählt das Kalzium in den Knochen. Auf der anderen Seite des durchstrahlten Körpers wird eine fotografische Platte angebracht, welche je nach Intensität der einlangenden Röntgenstrahlen mehr oder minder stark geschwärzt wird. Durch die Anordnung der Schwärzungspunkte kann auf die Struktur des durchdrungenen Körpers rückgeschlossen werden. Die Röntgendiagnostik ist ein sehr wichtiges Hilfsmittel in der Medizin; sie ist aber nicht ungefährlich, weil sie wie jede kurzwellige elektromagnetische Strahlung ionisierend wirkt und so die Zellen schädigen kann. Zur Darstellung von Weichteilen eignet sich ein röntgen-technisches Verfahren, die Computertomographie (CT), mit welcher kleinste Dichteunterschiede im Gewebe ermittelt werden können (bis zu 0,5 %). Dabei wird der Patient schichtweise mittels einer um ihn rotierenden Röntgenröhre mit niedriger Intensität durchstrahlt, wobei in den verschiedenen Richtungen die Strahlung verschieden stark absorbiert wird. Die Strahlung wird in ringförmig um den Patienten angeordneten Zählern (Detektoren) gemessen. Aus den erhaltenen Daten wird mittels Computer ein kontrastreiches Bild einer Schichtebene rekonstruiert. Danach wird der Patient in seiner Lage etwas verschoben und die nächste Gewebeschicht durchgescannt. Die Abbildung zeigt das Schema eines CT-Scans, wobei der Röntgenemittor schrittweise um 3° in jeder Schicht gedreht wird. Paukert: Ein Fenster zum Ich – Teil 1 66 Radioisotopen-Tomographie Ein Maß für die Aktivität einer bestimmten Körperregion ist der dortige Verbrauch von Sauerstoff und Zucker. Durch den erhöhten chemischen Stoffwechsel kommt es zu vermehrter Bildung von sauren Zwischenprodukten bei der Zuckerverbrennung. Diese sauren Stoffwechselprodukte bewirken eine Erweiterung der arteriellen Blutgefäße, was eine Erhöhung der lokalen Durchblutung zufolge hat. Radioaktive Stoffe senden die so genannte Gamma-Strahlung aus, die noch kurzwelliger und energiereicher und daher auch gefährlicher als die Röntgenstrahlung ist. Diese radioaktive Strahlung kann nur durch dicke Blei- und Betonplatten abgeschirmt werden. Durch den hohen Energiegehalt wirkt die radioaktive Strahlung sehr stark ionisierend auf die Stoffatome der jeweiligen Umgebung. Auf der ionisierenden Wirkung beruht auch der Strahlungsnachweis durch das Geiger-Müller-Zählrohr. Bei der Szintigraphie wird ein radioaktiver Markierungsstoff (tracer) in die Blutbahn gebracht. Seine Ausbreitung und Verteilung in bestimmten Körpergebieten kann nun an Hand der von ihm ausgesendeten radioaktiven Strahlung mit Hilfe von Geigerzählern gemessen werden. Die Messdaten werden von einem angeschlossenen Computer zu grafischen Bildern verarbeitet, an denen die Verteilung des Radioisotops in der Körperregion ersichtlich ist. Dadurch können Rückschlüsse auf die jeweilige lokale Durchblutung des Gewebes gezogen werden. Die dargestellte Abbildung zeigt die prinzipielle Messanordnung zur Feststellung der regionalen Gehirndurchblutung mit Hilfe von intraarteriell injiziertem radioaktivem Xenon (Xe). Paukert: Ein Fenster zum Ich – Teil 1 67 Bei der Positronen-Emissions-Tomographie (PET) werden die radioaktiven Isotope biologisch wichtiger Atome (F, O, N, C) verwendet, die Positronen (ßStrahlung) freisetzen. Direkt am Ort ihrer Freisetzung treffen diese Positronen auf die den Atomkern umgebenden Elektronen. Eine solche Kollision führt zu einer Vernichtung beider Teilchen. Dabei wird die vernichtete Masse in Energie umgewandelt, welche in Form von radioaktiver Gamma-Strahlung auftritt. Diese Strahlung wird nun von rund um den Kopf des Patienten angeordneten Detektoren registriert. Aus Richtung und Intensität der Strahlung kann die räumliche Verteilung der, die Positronen emittierenden Atome, ermittelt und dann graphisch dargestellt werden. In der Nuklearmedizin werden vor allem Substanzen wie Wasser, Glukose oder Aminosäuren mit den genannten Radioisotopen markiert. Aus dem graphischen Verteilungsbild radioaktiv markierter Glukose kann dann auf die lokale Durchblutung einer Region und somit auch auf ihre Aktivität rückgeschlossen werden. Die Abbildungen zeigen verschieden stark durchblutete Gehirnregionen (sehr gut ersichtlich an den Schwärzungen) bei unterschiedlichen sensorischen und motorischen sprachlichen Tätigkeiten. Die PET erlaubt auf diese Art und Weise die Darstellung von Bildern der geistigen Aktivität des Gehirns. Paukert: Ein Fenster zum Ich – Teil 1 68 Magnetresonanztomographie (MRT) Wie die Elektronen der Atomhülle haben auch die einzelnen Bausteine des Atomkerns (Nukleonen) einen Eigendrehimpuls (Spin) und verhalten sich wie winzige Magnete. Bei Atomkernen mit gerader Anzahl von Nukleonen können sich die magnetischen Kraftmomente der Kernbausteine kompensieren, bei ungerader Nukleonenanzahl kann dies nicht erfolgen. Der Kern besitzt dann einen resultierenden Gesamtspin und ein magnetisches Kraftmoment. Der Atomkern von Wasserstoff (H) besteht aus einem einzigen Proton und hat daher ein magnetisches Moment. Beim Anlegen eines starken äußeren Magnetfeldes beginnen die Atomkerne eine kreiselförmige Bewegung um ihre Gleichgewichtslage auszuführen (Auslenkung, Präzession). Die Geschwindigkeit dieser Kreiselbewegung (Präzessionsfrequenz) ist proportional zur angelegten magnetischen Feldstärke. Wenn die Atomkerne anschließend wieder in ihre energetisch günstigere Ausgangslage zurückkehren, senden sie genau jene elektromagnetische Frequenz aus, welche sie zuvor absorbiert haben. Diese Resonanzfrequenz des Atomkerns hängt einerseits vom angelegten äußeren magnetischen Feld, andererseits auch von den lokalen Magnetfeldern ab, die durch die Elektronen der Atomhülle erzeugt werden. Dadurch ist die Resonanz auch stoffspezifisch und kann in der Chemie zur Strukturanalyse von unterschiedlichen organischen Verbindungen benutzt werden (nuclear magnetic resonance spectroscopy, NRMS). In der Nuklearmedizin werden die oben beschriebenen atomphysikalischen Effekte in der so genannten Kernspintomographie verwendet (NMR-Tomographie oder auch MRT genannt). Der Großteil des menschlichen Körpers besteht aus Wasser, dessen Verteilung mit Hilfe der MRT in Schnittbildern graphisch dargestellt wird. Dadurch können Dichteunterschiede im Gewebe deutlich sichtbar gemacht werden. Bei der Untersuchung befindet sich der Patient im Inneren einer großen, von Strom durchflossenen Spule, die ein starkes, völlig homogenes Magnetfeld erzeugt. Dadurch werden die Wasserstoffkerne zuerst entsprechend diesem Felde ausgerichtet. Mit zusätzlichen Hochfrequenz-Spulen lässt sich die Stärke des Magnetfeldes räumlich variieren. Durch die kurzfristigen Zusatzimpulse werden die Wasserstoffkerne in einer bestimmten Schnittebene zur Kernspinresonanz angeregt. Paukert: Ein Fenster zum Ich – Teil 1 69 Die von den Kernen emittierte Resonanzstrahlung wird von Antennen empfangen, digital gespeichert und im Computer mit komplizierten Rechenverfahren zu einem grafischen Bild verarbeitet. Nach Fertigstellung eines solchen Schichtbildes wird der Patient in der Spule weiterbewegt, so dass ein Bild von der nächsten Schnittebene erzeugt werden kann. Medizinische Risken der Magnetresonanztomographie sind nicht bekannt. Paukert: Ein Fenster zum Ich – Teil 1 70