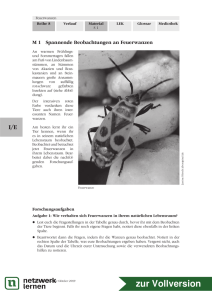
die färbung des feuersalamanders
Werbung

Patrik Matter DIE FÄRBUNG DES FEUERSALAMANDERS sowie eine allgemeine Literaturübersicht zu den Themen Färbung, Farbwirkung und Farbsehen Diplomarbeit eingereicht von Patrik Matter Am Zoologischen Institut der Universität Basel Unter der Leitung von Prof. Dr. David G. Senn Basel, Oktober 2003 Seite 1 Patrik Matter INHALT I.EINLEITUNG............................................................................................................ S.9 1. Zusammenfassung (Abstract) 2. Hintergrund 3. Ein einheimisches Beispiel ....................................................................................... S.10 4. Zielsetzung II. EVOLUTION VON SIGNALEN.......................................................................... S.11 1. Allgemeine Signale 1.1 Aspekte, die durch die Umgebung bedingt sind 1.2 Aspekte, die vom Sender ausgehen 1.2.1 Übermittlungsart 1.2.2 Ursache des Signals ............................................................................................. S.12 1.3 Aspekte, die durch den Empfänger bedingt sind 1.3.1 Sinne 1.3.2 „sensory biases“ (sensorisches Fenster) 1.3.3 Psychologie (Erkennungsmuster) 1.3.4 Lernen von Empfängern beeinflussen 1.4 Aspekte, die vom Empfänger und dem Sender eines Signals abhängen 1.4.1 Qualität des Signals (Fälschbarkeit) 1.4.2 Beziehung zwischen Sender und Empfänger (Kooperation – Konflikt) ................ S.14 1.4.3 Ritualisierung 1.5 Aspekte, die durch zufällige Empfänger bedingt sind 1.5.1 Auffälligkeit 2. Spezielle Signale 2.2 Aspekte, die vom Sender ausgehen 2.2.2 Ursache des Signals 2.3 Aspekte, die durch den Empfänger bedingt sind...................................................... S.15 2.3.3 Psychologie (Erkennungsmuster) 2.3.4 Lernvermögen von Empfängern beeinflussen(vgl. M. Speed (2001)) 2.5 Aspekte, die durch zufällige Empfänger bedingt sind 2.5.1 Auffälligkeit Seite 2 Patrik Matter III. MÖGLICHE FUNKTIONEN DER FÄRBUNG ................................................ S.16 1. Tarnung (kryptische Färbung) (vgl. H.Cott 1940 oder J. Hailman 1977) 1.1. Farbliche Anpassung an den Lebensraum 1.2 Gegenschattierung (Tayersches Prinzip) ................................................................. S.17 1.3 Gestaltauflösende Färbung (Somatolyse) ................................................................ S.18 1.3.1 Kontrastreiche Farbflächen am Rand der Silhouette ............................................. S.19 1.4 Verbergung der Augen 1.5 Mimikry, Mimese, Akinese (vgl. K. Lunau (2002)) 1.6 Besonderheiten von getarnten Tieren 2. Schreck..................................................................................................................... S.20 3. Warnung (aposematische Färbung) ........................................................................... S.21 3.1 Besonderheiten von aposematisch gefärbten Tieren 4. Drohung (aposematische Färbung)............................................................................ S.22 4.1 Statussymbole 4.2 Betonende Färbung 5. Balzfärbung .............................................................................................................. S.23 5.1 Statussymbole 5.2 Betonende Färbung 5.3 Symmetrie 5.4 Augenmuster 5.5 Färbung, die den körperlichen Zustand betont 5.6 Sichtbarkeit ............................................................................................................ S.24 5.6.1 Dunkle Färbung 5.6.2 Helle Färbung 5.6.3 Reflektierende Färbung 5.6.4 Gemischte Färbung 5.7 Sensorische und „mentale“ Fenster 5.8 Fishers Hypothese über die sexuelle Selektion ........................................................ S.25 6. Arterkennung (episematische Färbung) 7. Physikalische Eigenschaften (vgl. (E.Burtt(1977)) 7.1 Thermoregulation 7.2 Schutzfunktion 8. Keine selektive Funktion .......................................................................................... S.26 8.1. Ausprägung einer pleiotropischen Wechselwirkung 8.2 Historisches Relikt 8.3. Zufällige Färbung Seite 3 Patrik Matter IV. FARBWIRKUNG (VGL J.HAILMAN (1977))................................................... S.27 1. Licht 1.1 Sonnenstand 1.2 Höhe über Meer 1.3 Dunkelheit .............................................................................................................. S.28 2. Übermittlungsmedium 2.1 Streuung an Partikeln 2.1.1 Rayleight-Streuung ( Partikeldurchmesser < 1/10 λ ) 2.1.2 Mie-Streuung ( 1/10 λ< Partikeldurchmesser < 25 λ ) 2.1.3 Normale Spiegelung (Partikeldurchmesser > 25 λ) 2.2 Streuungen in versch. Medien................................................................................. S.29 2.2.1 Luft (Nebel, Aerosol, etc) 2.2.2 Wasser................................................................................................................. S.30 2.2.3 Vegetationsreiche Medien................................................................................... S.31 2.2.4 Optische Hindernisse (Pflanzen, Riffblöcke, etc.) 3. Hintergrund 3.1 Offene Lebensräume (Wüsten, Himmel, Polarzonen, Strände, Tiefsee, etc.) ........... S.32 3.2 Wasserflächen (See, Ozean, etc) 3.3 Stark strukturierte Lebensräume.............................................................................. S.33 4. Verhalten V. SEHVERMÖGEN DER WIRBELTIERE (VGL. G.WALLS 1942) ................... S.34 1. Hell-/ Dunkelsehen mit den Stäbchen 1.1 Ursachen für die grössere Sensitivität von Stäbchen 1.1 Ursachen für die grössere Lichtsensitivität von Stäbchen 1.1.1 Form 1.1.2 Verschaltung 1.1.3 Rhodopsin............................................................................................................ S.35 1.2 Doppel-Stäbchen 2.1. Ursachen für die höhere Auflösung der Zapfen verglichen mit den Stäbchen ......... S.36 2.1.1 Verschaltung 3. Farbwahrnehmung des Menschen 3.1 Absorption durch die Zapfen 3.2 Verarbeitung der Signale ........................................................................................ S.38 3.2.1 Verarbeitung der Signale in der Netzhaut............................................................. S.39 3.2.1.1 Bipolarzellen (rezeptive Felder) 3.2.1.2 Ganglienzellen (Farbwerte) 3.2.2 Verarbeitung der Signale im Gehirn 3.3 Farbqualitäten beim Mensch 3.3.1 Primäre (physikalische) Farbqualitäten (Intensität, Wellenlänge, Reinheit) 3.3.2 Sekundäre (psychologische) Farbqualitäten (Helligkeit, Farbton, Sättigung) ........ S.40 3.3.2.1 Helligkeit 3.3.2.2 Farbton ............................................................................................................. S.41 Seite 4 Patrik Matter 3.3.2.3 Sättigung........................................................................................................... S.41 4. Farbwahrnehmung von Wirbeltieren......................................................................... S.42 4.1 Farbqualitäten bei Tieren 4.1.1 Helligkeit 4.1.2 Farbton 4.1.3 Sättigung ............................................................................................................. S.43 4.2. Spezielle Anpassungen an die Umwelt 4.2.1 Gelbgefärbte Linsen 4.2.2 Öltröpfchen 4.2.2.1 Öltröpfchen als Filter gegen die chromatische Abberation................................. S.44 4.2.2.2 Öltröpfchen erhöhen die Farbauflösung. 4.2.2.3 Öltröpfchen als Anpassung an die Lichtbedingungen 4.2.3 Doppel-Zapfen 4.3. Verbreitung des Farbsehens bei Wirbeltieren 4.3.1 Weiterführende Daten.......................................................................................... S.45 6. Spezielle Arten von Farbsehen.................................................................................. S.46 6.1 Farbsehen von Monochromaten 6.2 Farbsehen im Dunkeln VI. FEUERSALAMANDER ...................................................................................... S.47 1. Lebensweise 1.1 Lebenszyklus 1.2 Lebensraum (vgl. B.Tiesmeier & R.Günther (1996)) 1.3 Tagesrhythmus (vgl. R. Klewen ( 1991)) ................................................................ S.48 1.4 Jahresrhythmus 1.5 Paarung (vgl. R. Klewen ( 1991)) 1.6 Rivalenkämpfe des Feuersalamanders..................................................................... S.49 1.7 Nahrung des Feuersalamanders 1.8 Feinde des Feuersalamanders 2. Färbung der verschiedenen Unterarten ...................................................................... S.50 3. Farbsehen des Feuersalamanders 4. Experiment (Störungsverhalten) 4.1 Einleitung 4.2 Material und Methoden........................................................................................... S.51 4.3 Resultate 4.4 Diskussion 4.4.1 Mögliche Fehlerquellen ....................................................................................... S.52 5. Experiment (Giftspritzen) ......................................................................................... S.53 5.1 Einleitung 5.2 Material und Methoden 5.3 Resultate 5.4 Diskussion 5.4.1 Mögliche Fehlerquellen Seite 5 Patrik Matter VII. ALPENSALAMANDER..................................................................................... S.55 1. Lebensweise 1.1 Lebenszyklus (vgl. K.Grossenbacher & R. Günther (1996)) 1.2 Lebensraum (vgl. K.Grossenbacher & R.Günther (1996)) 1.3 Tagesrhythmus (vgl. R. Klewen ( 1991)) 1.4 Jahresrhythmus (vgl. K. Grossenbacher & R.Günther (1996))................................. S.56 1.5 Paarung 1.6. Nahrung des Alpensalamanders 1.7 Feinde des Alpensalamanders 2. Färbung der drei Unterarten ...................................................................................... S.57 VIII. DISKUSSION .................................................................................................... S.58 1. Färbung 1.1.1 Die Färbung des Feuersalamanders – ein Tarnfärbung? 1.1.2 Die Färbung des Feuersalamanders – eine Schreckfärbung? 1.1.3 Die Färbung des Feuersalamanders – eine Warnfärbung? 1.1.3.1 Abklärung, der für Tiere mit Warnfärbung typischen Eigenschaften 1.1.3.2 Vorhandene Eigenschaften 1.1.3.3 Voraussetzung für eine Funktion als Warnfärbung 1.1.4. Die Färbung des Feuersalamanders – eine Drohfärbung? .................................... S.60 1.1.5 Die Färbung des Feuersalamanders – eine Balzfärbung 1.1.6. Die Färbung des Feuersalamanders – eine Episematische Färbung? .................... S.61 1.1.7. Die Färbung des Feuersalamanders – als Färbung mit physikalischer Funktion ? 1.1.8 Die Färbung des Feuersalamanders – ohne selektive Funktion? 1.2 Zusammenfassung 1.3 Unterschiedlich gefärbte Unterarten des Feuersalamanders..................................... S.62 1.4 Evolution von Warnfärbung via Tarnfärbung? 2. Mögliche weiterführende klärende Experimente 2.1 Tarnung? 2.2 Warnung? ............................................................................................................... S.63 3. Vergleich mit dem Alpensalamander (hypothetische Überlegungen) 3.1 Färbung 3.1.1 Gefleckte Unterarten 3.1.2 Ausschliesslich schwarz gefärbte Unterarten 4. Farbwirkung ............................................................................................................. S.65 5. Farbsehen 6. Evolution von Signalen............................................................................................. S.66 Seite 6 Patrik Matter ANHANG: .................................................................................................................. S.67 IX. APOSEMATISCHE MECHANISMEN: (DROH- UND WARNMECHANISMEN) 1.Mechanismen, die auf den Sehsinn wirken................................................................. S.68 1.1. Färbung 1.1.1 Auffällige Dauerfärbung Beispiel für Warnen: (Pfeilgiftfrösche (Dendrobates spec.)) 1.1.2 Physiologischer Farbwechsel Beispiel für Warnen: Blauringkraken (Hapalochlaena spec.)...................................... S.69 Beispiel für Drohung: Chamäleon (Chamaeleo spec.) 1.1.3 Auffällig gefärbter Körperteile Beispiel für Warnung: Unke (Bombina spec.) ............................................................. S.70 Beispiel für Drohung: Saumfinger (Anolis spec.) 1.2. Verändern der Körperform 1.2.1 Mechanismen die den Körper grösser erscheinen lassen....................................... S.71 Beispiel für Warnung :Königskobra (Ophiophagus hannah ) ...................................... S.72 Beispiel für Drohung:Wolf (Canis lupus) 1.2.2 Schutzmechanismen Beispiel für Warnung: (Kleinohr-Igel (Erinaceus spec.)) ............................................ S.73 1.3. Bewegungen Beispiel für Warnung (Grauer Riffhai (Charcharinus amblyrhynchos)) Beispiel für Drohung: Kampffische (Betta spec.)........................................................ S.74 2. Mechanismen, die auf das Gehör wirken 2.1.Lauterzeugung mittels der Stimme Beispiel für Warnung (Warnruf der Graudrosslinge (Turdoides squamiceps))............. S.75 Beispiel für Drohen (Erdkröte (Bufo bufo)) 2.2 Lauterzeugung mit anderen Mitteln ........................................................................ S.76 Beispiel für Warnung (Klapperschlangen (Crotalus spec.)) 3. Mechanismen, die auf Geruchsinn, Geschmacksinn oder auf andere chemische Sinne wirken 3.1. Geruch und Geschmack Beispiel für Warnung (Ringelnatter (Natrix natrix)).................................................... S.77 3.2. Ausspritzen von Körperflüssigkeiten Beispiel für Warnung: Stinktier (Familie: Mephitinae) Beispiel für Drohen: (Rotfuchs (Vulpes vulpes)) ......................................................... S.78 4. Mechanismen, die auf den Tastsinn wirken Beispiel für Warnung (Schimpanse (Pan troglodytes)) Beispiel für Drohung (Hornträger (Familie:Bovidae)) 5. Mechanismen, die auf eine Mehrzahl von Sinne wirken............................................ S.79 Seite 7 Patrik Matter X.LITERATURVERZEICHNIS................................................................................... S.79 XI.GLOSSAR .............................................................................................................. S.85 XII.TABELLEN........................................................................................................... S.88 XIII. DANKSAGUNG ................................................................................................. S.97 Seite 8 Patrik Matter I.EINLEITUNG 1. Zusammenfassung (Abstract) Die gelb-schwarze Färbung des Feuersalamanders wird in der Literatur als typisches Beispiel für eine aposematische Färbung (Warnfärbung) genannt. Auch gibt es Hinweise auf ein Warnverhalten des Feuersalamanders. Meine eigenen Experimente zeigten jedoch kein solches Verhalten. Auch ist der Feuersalamander ein nachtaktives Tier, dessen Färbung nur sehr selten sichtbar ist. Die Literaturübersicht und die Tatsache, dass ein Warnverhalten experimentell nicht nachgewiesen werden konnte, lassen vermuten, dass die Färbung des nachtaktiven Feuersalamanders in erster Linie eine tarnende Funktion hat. Schlüsselwörter: Feuersalamander, Salamandra salamandra, Tarn- und Warnfärbung, Färbung, Evolution, Giftspritzen, Farbsehen The yellow-black coloration of the fire salamander is known in literature as a typical example for aposematic coloration. Also there is some evidence for an aposematic behaviour of the fire salamander. My own experiences don’t show such behaviour. Also is the fire salamander a nocturnal animal, therefore the coloration is only seen on rare occasion. The literature review and the fact that behavioural experiments don’t show aposematic behaviour lead to the conclusion, that the coloration of the nocturnal fire salamander has mainly a somatolytic function. Key words: Fire salamander, Salamandra salamandra, coloration, aposematic, somatolytic, evolution, venom spraying, color vision 2. Hintergrund Charles Darwins Buch „ The Origin of Species“ legt sozusagen den Grundstein für die neuere Biologie. Darwin war der erste, der erkannte, dass nicht die Schöpfung die Ursache für die heutige Artenvielfalt ist, sondern, dass auch heute noch neue Arten entstehen (Evolution). Seiner Meinung nach entwickeln sich durch natürliche Selektion („Surviving of the fittest“) immer neue Arten, bis alle ökologischen Nischen besetzt sind. Es überleben also jene Tiere, die am besten an ihre Umgebung angepasst („fit in“) sind. Eine der vielen Bestätigungen dieser Theorie ist die Tarnfärbung vieler Tiere. Damit Tiere nicht von Raubtieren gefressen werden, passen sie ihre Färbung (teilweise sogar ihre Form) ihrer Umgebung an. Auch Raubtiere sind oft getarnt, um sich besser an ihre Beute anschleichen zu können. In der Natur gibt es jedoch auch das Gegenteil; Tiere mit bunten, auffälligen („conspicious“) Färbungen und Verhalten, die es geradezu darauf anzulegen scheinen, von anderen gesehen zu werden. Daher ist es nicht erstaunlich, dass Warnfarben in der Biologie zu verschiedensten Theorien und hitzigen Diskussionen geführt haben. Bis heute ist noch nicht endgültig geklärt, wie Warnfarben funktionieren und noch weniger, wie sie sich entwickeln konnten. Ich wurde darauf aufmerksam, als ich in einer Vorlesung von einem Gifttier-Experten erfuhr, dass sich die Signalfärbung der gebänderten Schlange (Micrurus corallinus) in ihrem natürlichen Lebensraum (Grassteppe) in eine Tarnfarbe verwandelt. Die mögliche Tarnung in Seite 9 Patrik Matter bestimmten Gebieten wäre eine einfache Erklärung für das schwer zu verstehende Anfangsstadium der Evolution von Warnfärbung! 3. Ein einheimisches Beispiel Für meine Diplomarbeit begab ich mich auf die Suche nach einem ähnlichen Beispiel in der Schweizer Fauna. Schliesslich wurde ich in Form des hier typischen Beispiels für Warnfarben, des Feuersalamanders (Salamandra salamandra) fündig. Dieser gelbgefleckte Salamander ist hauptsächlich dämmerungs- und nachtaktiv. Daher scheint eine Warnfärbung auf den ersten Blick nicht sehr sinnvoll zu sein. Besonders interessant ist dieser Salamander auch in einer weiteren Hinsicht. Ebenfalls in der Schweiz heimisch ist noch eine weitere Salamanderart: der Alpensalamander (Salamandra atra).Dieser ist jedoch, obwohl mit dem Feuersalamander nahe verwandt, mattschwarz. Ausserdem ist kürzlich auch eine teilweise gelbgefärbte Unterart (S.s.aurora) beschrieben worden. Dadurch könnten möglicherweise weitere wichtige Hinweise auf die Ursachen der gelben Färbung gefunden werden. 4. Zielsetzung Das Ziel meiner Diplomarbeit ist es, in einem Literaturstudium zuerst einen allgemeinen Überblick über das Thema zu erarbeiten. Auf diesen gestützt möchte ich schliesslich, ergänzt durch die Resultate eigener Experimente, klären, welches die wahrscheinlichste Funktion der Färbung des Feuersalamanders ist. Im allgemeinen Teil (Kapitel II-V) wird zuerst versucht die Frage zu klären , wie sich Signale in spezielle Warnsignale entwickeln. Danach wird abgeklärt, inwiefern heute das Anfangsstadium der Warnfärbung verstanden ist. Anschliessend wird detailliert auf die Problematik Färbung (Funktion, Farbwirkung, Farbsehen) eingegangen. Ziel ist es, eine möglichst allgemeine Basis für Fragestellungen, welche die Färbung betreffen, zu schaffen, damit für ähnliche Fragestellungen auf diesem Gebiet nur noch der spezifische Teil erarbeitet werden muss. Im spezifischen Teil (Kapitel VI+VII) werden Daten über Feuer- und Alpensalamander gesammelt. Es werden zwei Experimente mit Feuersalamandern durchgeführt, die klären sollen, ob diese ein Warnverhalten besitzen. In der Diskussion (Kapitel VIII) wird exemplarisch am Beispiel „Feuersalamander“ gezeigt, wie die Daten vom allgemeinen Teil verknüpft mit den spezifischen Daten zu einer Hypothese über die Funktion der Färbung des Feuersalamanders führen. Nachdem diese Hypothese mittels eines Vergleichs mit dem Alpensalamander auf ihre Konsistenz überprüft worden ist, wird auf mögliche weiterführende Experimente hingewiesen. Zum Schluss wird kurz auf Probleme und Fragen eingegangen, die in den einzelnen Gebieten aufgeworfen wurden. Seite 10 Patrik Matter II. EVOLUTION VON SIGNALEN 1.Allgemeine Signale Umgebung (enviroment) Sender Beziehung - Übermittlungsart - Ursache Qualität Ritualisierung Empfänger (receiver) - „sensory biases“ - Psychologie - Lernverhalten Auffälligkeit Andere Empfänger(other receivers) Abbildung 1: Grafik zur Verdeutlichung der unterschiedlichen Aspekte der Selektion Eine allgemeine Signal-Theorie liefert eine Erklärung für die Evolution der beobachteten Vielfalt in Form und Stärke von tierischen Signalen. Da jeder Aspekt in der Selektion den entscheidenden Faktor spielen könnte, ist es für das Verständnis sinnvoll, die einzelnen Aspekte, die ein Signal hat (Abbildung 1), getrennt zu betrachten. 1.1 Aspekte, die durch die Umgebung bedingt sind Ein sehr wichtiger Faktor für die Evolution von Signalen ist natürlich die Umgebung, in der ein Signal wirken muss. Damit ein Signal wirken kann, muss es zuerst einmal wahrgenommen werden können, was bedeutet, dass es sich seiner Umgebung (Störsignale) anpassen muss. So sind zum Beispiel die Gesänge von Vögeln an ihren jeweiligen Lebensraum angepasst (C. Chappuis (1971)). 1.2 Aspekte, die vom Sender ausgehen 1.2.1 Übermittlungsart Die Übermittlungsart hängt vor allem von den Voraussetzungen ab, die ein Sender hat, bevor sich das Signalgeben evoluiert. Wenn zum ersten Mal ein Signal gesendet wird, muss der Sender eine der ihm möglichen Übermittlungsarten nutzen. Hat er Erfolg, wird er mit grosser Wahrscheinlichkeit auch in Zukunft dieselbe Art von Signal benützen. Bei einem Misserfolg wird der Signalgeber, falls er überlebt, ein anderes Signal ausprobieren. Seite 11 Patrik Matter 1.2.2 Ursache des Signals Der Grund weshalb ein Signal gegeben wird, hat ebenfalls wichtige Auswirkungen auf Form und Stärke eines Signals. Je wichtiger der Zweck eines Signals für einen Sender ist, desto höhere Kosten (Energie, negative Konsequenzen) darf das Signal haben. Signale sind zudem meistens so gestaltet, dass sie einen Bezug zu der übermittelten Botschaft haben. 1.3 Aspekte, die durch den Empfänger bedingt sind 1.3.1 Sinne Natürlich kann ein Signal nur dann auf einen Empfänger wirken, wenn dieser es wahrnehmen kann. Die Signale, die auf die Hauptsinne von Empfängern wirken, haben oft grösseren Erfolg. 1.3.2 „sensory biases“ (sensorisches Fenster) Sinnesorgane nehmen nicht alle Reize gleich stark wahr. Es gibt gewisse Frequenzen, die besonders gut wahrgenommen werden. Trifft ein Sender ein solches Fenster, wird sein Signal verstärkt wahrgenommen, ohne dass sich der Sender besonders anstrengen muss (Ryan (1985)). 1.3.3 Psychologie (Erkennungsmuster) Ein wahrgenommener Reiz wird vom Hirn verarbeitet. Dabei führt das Gehirn eine Mustererkennung durch, um einige Schlüsselreize sofort anhand weniger Informationen zu erkennen. So haben Empfänger (ähnlich wie ein sensorisches Fenster) auch ein „mentales Fenster“ durch welches Signale verstärkt wahrgenommen werden können (Arak&Enquist (1993)). 1.3.4 Lernen von Empfängern beeinflussen Viele Signale beeinflussen auch das Lernverhalten des Empfängers. Einige sind einprägsamer, andere wirken so, dass sie unmöglich mit anderen Signalen verwechselt werden können (Speed MP (2001)). 1.4 Aspekte, die sowohl vom Empfänger, wie auch vom Sender eines Signals abhängen 1.4.1 Qualität des Signals (Fälschbarkeit) In der menschlichen Sprache sind Lügen gang und gäbe. In der Tierwelt hingegen sind sie selten. Das Handicap-Prinzip (A.Zahavi (1997)) sorgt dafür, dass in der Tierwelt nur selten getäuscht wird. Nach dieser heute weitgehend akzeptierten Theorie müssen Signale drei Bedingungen erfüllen: Seite 12 Patrik Matter 1. Sie müssen für das Individuum, das sie sendet, kostspielig sein. 2. Der Preis muss für einen Betrüger höher sein, als für den ehrlichen Sender. 3. Es muss eine Beziehung zwischen den Kosten für das Signal und der vom Signal vermittelten Botschaft geben. Thomson-Gazellen (Gazella thomsoni) flüchten nicht, wenn sich ihnen ein Raubfeind nähert, sondern springen auf der Stelle hoch, um diesem zu demonstrieren, dass sie keine lohnende Beute darstellen. 1. Das auf der Stelle Hochspringen erfordert viel Energie. Energie, die bei einer späteren Flucht von grossem Nutzen ist. 2. Das Hochspringen auf der Stelle soll demonstrieren, dass die Gazelle über genügend Reserven verfügt und sie dem Raubfeind in einer für beide anstrengenden Flucht entkommen würde. Falls der Raubfeind jedoch sehr hungrig ist, wird er trotz dieser Warnung eine anstrengende Jagd riskieren müssen. Wenn nun eine ausgemergelte Gazelle ihre gute Kondition nur vorgetäuscht hat, muss sie ihren Betrug mit dem Tod bezahlen, während die ehrliche Senderin eine gute Chance hat, dem Wolf zu entkommen. 3. Die hohen Prellsprünge brauchen viel Kraft und sind eine grosse Energieverschwendung. Energie verschwenden, kann nur, wer auch genügend Energie hat. Hier ist also der Bezug offensichtlich! Interessanterweise springen Gazellen, die vor einem Jeep mit menschlichen Jägern flüchten, nicht auf der Stelle hoch, sondern fliehen - die örtlichen Gegebenheiten als Deckung nutzend. Denn bei einem so überlegenen Jäger ist eine unsichtbare Flucht die einzige Rettung (Fitzgibbon & Fanshawe (1988)). Da in der Natur trotzdem einige Täuschungen vorkommen, haben Maynard Smith & Harper(1988) vorgeschlagen, Signale in zwei Kategorien einzuteilen: „assessment“ Signale, die zuverlässig mit ihrer zugrunde liegenden Qualität verknüpft sind (z.B. Kampffähigkeit) und konventionale Signale. Weil diese nur bedingt mit den ihnen zugrunde liegenden Qualitäten verknüpft sind, können sie gefälscht werden. Allerdings können Signale und Fälschung nur koexistieren, solange zwei Bedingungen erfüllt sind: a) Der Wert der umstrittenen Ressource ist klein. b) Individuen, die Signale fälschen, haben im Verhältnis zu dem Nutzen nicht zu hohe Kosten (Energieaufwand, Strafe von Artgenossen, etc.). Ein Signal kann zwei verschiedene Arten von Kosten haben: Direkte Kosten, welche durch die Produktion des Signals verursacht werden (z.B. das Röhren der Hirsche) und indirekte Kosten, welche durch die Folgen des Sendens entstehen (z.B. Warnfarben, die ein Tier auffälliger für Fressfeinde erscheinen lassen). Da indirekte Kosten vom Verhalten von Empfängern abhängig sind, kann es bei solchen Kosten oft dazu kommen, dass diese Kosten durch einen „faulen“ Empfänger herabgesetzt werden (bei Warnfärbung kann es zu Mimikry kommen, wenn Raubfeinde nicht genügend oft „proben“). Seite 13 Patrik Matter 1.4.2 Beziehung zwischen Sender und Empfänger (Kooperation – Konflikt) Handelt es sich um Signale zwischen kooperierenden Gegnern, ist es möglich, dass die Kosten nur aus indirekten Kosten bestehen. Diese sind dadurch zustande kommen, dass ein gefälschtes Signal die sofortige Kündigung der Kooperation zur Folge hat. Darüber hinaus entwickeln sich in kooperativen Systemen oft nicht teure Signale, damit nicht unnötige Energie verschwendet wird. In Konfliktverhältnissen hingegen tendieren die Signale dazu, sehr auffällig zu sein. Sie haben oft direkte Kosten, um den Gegner zu beeinflussen, da die Glaubhaftigkeit des Signals nur durch die hohen Kosten gewährleistet ist. 1.4.3 Ritualisierung Eine weitere Möglichkeit der Entstehung von Signalen ist, dass ein Verhalten eines Senders unbewusste Bewegungen voraussetzt, die für den Empfänger eine Signalwirkung entfalten können. Nun ist es für den Sender möglich dieses Verhalten oder einige Aspekte davon, bewusst einzusetzen, um dem Empfänger dadurch ein Signal zu geben (Hinde RA (1970)). Oft sind solche ursprüngliche Signale Handlungen, die einen Motivationskonflikt widerspiegeln, also den Wechsel von freundlichem zu aggressivem Verhalten (z. B Schwanzwedeln bei der Hauskatze). 1.5 Aspekte, die durch zufällige Empfänger bedingt sind 1.5.1 Auffälligkeit Signale haben den Nachteil, dass sie auch von zufälligen Empfängern wahrgenommen werden können. Falls es sich bei den zufälligen Empfängern um Raubfeinde handelt, können auffällige Signale sehr gefährlich sein. Es könnte natürlich auch sein, dass einige Signale gerade durch die indirekten Kosten, die durch Raubfeinde entstehen, eine sichere Qualität haben. Dies könnte zum Beispiel bei dem Prachtkleid der Entenerpel der Fall sein, welches sich auf kleinen Inseln ohne Raubfeinde zurückentwickelt (A. Zahavi (1981)). Um die Gefahr einer erhöhten Auffälligkeit zu mindern, sind Alarmrufe einiger Vögel so konzipiert, dass sie das Orten der Rufer erschweren (P.Marler (1995)). 2. Spezielle Signale Hier wird auf besondere Aspekte eingegangen, die Warn-, Droh-, Balzsignale sowie das Senden von keinem Signal (Tarnung) haben können. 2.2 Aspekte, die vom Sender ausgehen 2.2.2 Ursache des Signals Bei der natürlichen Selektion überlebt nur „the fittest“, also das Individuum welches sich optimal fortpflanzen und am längsten überleben kann. Die Tarnung und Warnung dienen dem Überleben, die Drohung und Balz der Chance der Fortpflanzung. Ein so hoher Einsatz rechtfertigt auch hohe Kosten. Daher sind solche Signale teilweise sehr kostspielig! Seite 14 Patrik Matter 2.3 Aspekte, die durch den Empfänger bedingt sind 2.3.2 „sensory biases“ (sensorisches Fenster) 2.3.3 Psychologie (Erkennungsmuster) Bei Balzsignalen ist es besonders wichtig, dass das stärkste Signal aussendet wird. Daher sind in diesem Zusammenhang oft Anpassungen an ein sensorisches Fenster oder an die Mustererkennung der Empfänger zu beobachten. Tarnsignale wirken zudem oft so, dass sie durch die Mustererkennung von Feinden nicht wahrgenommen werden. 2.3.4 Lernvermögen von Empfängern beeinflussen(vgl. M. Speed (2001)) Von Warnsignalen ist bekannt, dass sie das Lernvermögen auf mindestes vier verschiedene Arten beeinflussen: 1.Verstärkung der den meisten Tieren angeborenen Angst vor Neuem (Neophobia) 2.Beschleunigte Ausbildung einer Abneigung gegen spezifisch gefärbte Beute. 3. Erhöhung der Entdeckungsrate durch erfahrene Raubfeinde und somit eine Reduktion der Chance zufälligerweise erbeutet zu werden 4.Verminderung oder Verhinderung des Vergessens von unangenehmen Konsequenzen eines Angriffs des Raubfeinds 2.5 Aspekte, die durch zufällige Empfänger bedingt sind 1.5.1 Auffälligkeit „Tarnsignale“ dürfen überhaupt nicht auffällig sein. Warnsignale, müssen hingegen möglichst auffällig sein, um wirken zu können. 3. Problem der Evolution von Warnsignalen Ein grosses Problem bei der Evolution von Warnverhalten stellt das ursprüngliche, erste Signal dar. Falls der Angreifer auf das Signal nicht reagiert, überlebt der Sender oft nicht. Daher vermutete man früher, dass Warnsignale nur durch Verwandtenselektion oder Ansammlungen („aggregation“) entstehen können. Heute gibt es jedoch einige Theorien, die das Entstehen von Warnsignalen ohne Verwandtenselektion erklären können (M.Speed 2001, M. Riipi 2001, S.Yachi 1998). Eine Evolution über synergistische Wechselwirkungen mit anderen Funktionen von Signalen wurde bis heute nicht speziell berücksichtigt und wäre eventuell eine weitere, interessante Möglichkeit für die Entwicklung von aposematischen Signalen(vgl. VIII. Diskussion). Seite 15 Patrik Matter III. MÖGLICHE FUNKTIONEN DER FÄRBUNG 1. Tarnung (kryptische Färbung) (vgl. H.Cott 1940 oder J. Hailman 1977) 1.1. Farbliche Anpassung an den Lebensraum Wie wichtig die Tarnung in der Natur ist, sieht man an der weiten Verbreitung der farblichen Anpassung an den eigenen Lebensraum: In der Wüste sind viele Tiere sandfarben, im tropischen Regenwald sind grüne Tiere sehr häufig, in der verschneiten nördlichen Tundra überwiegen weisse Tiere und in den endlosen Weiten des Ozeans ist blau die vorherrschende Färbung (Abbildung 2). Abbildung 2: Farbliche Anpassung an den Lebensraum im ganzen Tierreich Ein ähnliches Bild ergibt sich, wenn man nur eine ausgewählte Tierordnung (bzw. Gattungen) anschaut. So sind zum Beispiel Echsen (Sauria) je nach Lebensraum unterschiedlich gefärbt. Waldbewohnende Arten sind oft grün oder braun (Baumrinde), in der Wüste lebende Echsen sind oft sandfarbig mit staubähnlichen Flecken und Echsen die auf felsigem Untergrund leben, sind gräulich und haben oft eine gestaltauflösende Färbung. Im durch viele Lichtflecken erhellten Unterholz des Waldes haben die Echsen ebenfalls helle Flecken, während die Echsen des Graslandes gestreift sind (Abbildung 3). Abbildung 3: Farbliche Anpassung der Echsen an ihren Lebensraum Ein weiteres gutes Beispiel sind lokale Rassen von verschiedenen Kleinlebewesen, die sich durch ihre dem Lebensraum angepasste Färbung in verschiedene Unterarten aufteilen (z.B. Dice and Blossom (1930)). Seite 16 Patrik Matter 1.2 Gegenschattierung (Tayersches Prinzip) Ein Tier kann farblich noch so gut an seine Umgebung angepasst sein, in freier Natur kann es jedoch durch Beleuchtung verraten werden. Das Sonnenlicht hellt die Oberseite eines Körpers optisch auf, während die Unterseite des Körpers vom eigenen Schatten optisch dunkler erscheint. Dadurch wird selbst der Effekt der besten Tarnfärbung aufgehoben. Um diesem Phänomen entgegenzuwirken, sind viele Tiere auf ihrer Oberseite dunkler gefärbt als auf ihrer Unterseite. Dieses weit verbreitete Prinzip der Gegenschattierung wurde erstmals von Abott Tayer (1897) beschrieben(Abbildung 4). Abbildung 5: Abbildung 4: Das Prinzip der Gegenschattierung: Oben: Ein einfarbig gefärbter Raubfisch mit natürlicher Belichtung von oben Mitte: Ein gegenschattiert gefärbter Raubfisch gleichmässig beleuchtet Unten: Ein gegenschattiert gefärbter Raubfisch mit natürlicher Belichtung von oben Seite 17 Gegenschattiert gefärbte Tiere aus verschiedenen offenen Lebensräumen Patrik Matter Besonders in offenen Lebensräumen, wo Tiere schon von weitem sichtbar sind, ist Tarnung durch Gegenschattierung weit verbreitet. So sind praktisch alle Lebewesen, die im offenen Ozean jagen (Pinguine, Delphine, Haie, etc.), ebenso wie ihre Beute (Makrelen, Füsiliere, Sardinen, Meeresschildkröte, Kalmare, etc.) nach diesem Prinzip gefärbt. Dasselbe gilt aber auch für viele Landraubtiere (Löwen, Geparde, Leoparden, Wölfe, Füchse, etc.) wie auch für ihre Beute (Antilopen, Gazellen, Hirsche, etc.). Bei kleineren Tieren (Singvögel, Schlangen, Nager, Molche, etc.) ist dieses Prinzip ebenfalls weit verbreitet (Abbildung 5). Der Farbgradient für die Gegenschattierung kann zudem durch ein sich auf der Oberseite verdichtendes Muster erreicht werden (Abbildung 6). Abbildung 6: Gegenschattierte Musterung von Helmperlhuhn (Numida meleagris) und Gingsterkatze (Genetta tigrina) 1.3 Gestaltauflösende Färbung (Somatolyse) In offenen einheitlich gefärbten Lebensräumen ist ein Tier welches farblich an seine Umgebung angepasst ist und nach dem tayerschen Prinzip gefärbt ist, praktisch unsichtbar. Solche idealen Bedingungen sind in der Natur jedoch selten. Die meisten Lebensräume sind nicht einheitlich gefärbt, sondern bieten ein wirres Muster von Farben. Tiere die in solchen Lebensräumen überleben, sind nach dem Prinzip der Somatolyse gefärbt. Dieses Prinzip beruht darauf, dass Raubtiere gleich gefärbte Flächen als zusammenhängende Formen interpretieren .So passen die somatolytisch gemusterten Beutetiere nicht mehr in ihr Suchschema. Nach A. Thayer (1897) kann diese Wirkung auch durch sehr bunte Färbung erreicht werden (Abbildung 7). Abbildung 7: Prinzip der somatolytischen (gestaltauflösenden) Färbung Seite 18 Patrik Matter Je vielfältiger ein Muster ist, in desto mehr Teile zerfällt das Objekt und desto geringer ist die Chance, dass die wahre Form wahrgenommen wird. Die Form ist zudem besser aufgelöst, wenn das Muster einen möglichst grossen Kontrast bietet (z.B. helle Muster auf einem dunklen Objekt). Ein grösserer Grenzflächenkontrast erhöht ebenso die somatolytische Wirkung des Musters. Teilweise haben die Muster sogar einen Stereo-Effekt; sie schaffen die Illusion einer Dreidimensionalität. Bei vielen getarnten Tieren ist das somatolytische Muster so optimiert, dass es in der Ruhestellung den grössten Schutz bietet. Eines der berühmtesten Beispiele dafür ist der Baumfrosch (Megalixalus fornasinii). 1.3.1 Kontrastreiche Farbflächen am Rand der Silhouette Die Wirkung der besten Tarnung wird aufgehoben, wenn sich ein Tier bewegt. Die Mustererkennung des Auges kann Bewegungen wahrnehmen, selbst wenn eine Form noch nicht sichtbar ist. Ein aufmerksames Tier würde daher durch die Bewegung aufschrecken, seine Aufmerksamkeit auf das „unsichtbare“ Objekt fixieren und die Tarnung schon bald durchschauen. Um diesem Problem entgegenzuwirken, haben gut getarnte Tiere oft am Rande ihrer Silhouette eine kontrastreiche Färbung. So nimmt ein Beobachter von weitem nur kleine, sich bewegende Formen wahr. Als alternative Hypothesen kommt teilweise auch eine Signaloder thermoregulatorische Funktion in Frage. (z.B. Schneehase (Leptus timidus), Riffhaie (Familie:Charcharhinidae), Hermelin (Mustela erminea), etc.). 1.4 Verbergung der Augen Die Augen eines Tieres sind oft die auffälligsten Teile des Tieres. Daher zielen viele somatolytische Färbungen darauf ab, die Augen zu verbergen (z.B. dunkle Augenstreifen der Rifffische). Einige Rifffische haben zusätzlich noch falsche Augenflecken, um mögliche Angreifer irre zu führen (Abbildung 8). 1.5 Mimikry, Mimese, Akinese (vgl. K. Lunau (2002)) Abbildung 8: Falterfisch (Chaetodon capistratus) mit Scheinauge und Augenbinde Eine andere Möglichkeit der Tarnung ist die Mimikry von Tieren, die für den Raubfeind nicht lohnend sind (z.B. giftige Tiere). Auch die Mimese von toten Materialien ist weit verbreitet (z.B. Blätter, Äste, Seegräser, etc.). Da viele Raubtiere nur sich bewegende Beute wahrnehmen, ist auch Akinese (Sicht-TotStellen) eine Möglichkeit, sich vor Feinden zu schützen. 1.6 Besonderheiten von getarnten Tieren Tarnung wirkt am besten, wenn sich ein Tier nicht bewegt. Daher fliehen sehr gut getarnte Tiere erst, wenn kein Zweifel mehr besteht, dass ein Raubfeind die Tarnung durchschaut hat (z.B. am Boden brütende Vögel). Seite 19 Patrik Matter Getarnte Raubfeinde sind hingegen still verharrende Lauerräuber (z.B. Anglerfische ( Attenarius spec.)) oder sie pirschen sich langsam an eine Beute, damit ihre Tarnung nicht durch schnelle Bewegungen durchbrochen wird. Meistens wird die Tarnung jedoch ab einer gewissen Nähe sowieso durchschaut. Dann beginnt ein Wettlauf zwischen Räuber und Beute. Es könnte sein, dass die somatolytische Tarnfärbung sich ab einer gewissen Nähe ins Gegenteil umkehrt und somit zu einer aposematischen Signalfärbung wird. Diese lässt den Raubfeind auf nahe Distanz besser sichtbar werden. Die Beute wird sich also der Nähe des Feindes bewusst und dieser könnte aus dem daraus resultierenden Verhalten wichtige Schlüsse auf die körperliche Leistungsfähigkeit der einzelnen Beutetiere ziehen. (I. Eshel (1978)) Eine mögliche zweite Funktion von somatolytisch gefärbten Jungtieren könnte sein, dass sie durch die Wahl einer unpassenden Umgebung die Aufmerksamkeit ihrer Eltern erzwingen können. In diesem Fall könnte die somatolytische Färbung sehr auffallend sein. (A.Zahavi (1997)). 2. Schreck Einige kryptisch gefärbte Tiere haben in der Ruhestellung verdeckte Körperteile, die sehr grell gefärbt sind. Werden sie nun von einem Feind angegriffen, der ihre Tarnung durchschaut hat, zeigen sie plötzlich diese grell gefärbten Körperteile. Diese Zuschaustellung wird oft mit Schrecklauten verstärkt. Der Moment, in dem der erschreckte Feind zögert, kann reichen, dass die Beute entkommen kann. Das berühmteste Beispiel für die Schreckfärbung sind Schmetterlinge (Abbildung 9). Besonders Ordensbänder (Catocola spec.), aber auch viele andere Insekten (z.B. Schnarrheuschrecke (Psophus stridulus)) machen sich dieses Prinzip zu Nutze: Einige Echsen (z.B. Saumfinger (Anolis spec.)) spreizen ihre auffällig gefärbten Hautsäcke, einige Fische (z.B. Knurrhahn ( Trigla spec.)) spreizen auffällig gefärbten Flossen. Die Schreckwirkung von grell gefärbten Körperteilen wurde von D.Schlenoff (1985) experimentell nachgewiesen. Eine andere mögliche Funktion ist, dass die grellen Farben bei einer Flucht den Focus eines Raubfeindes auf sich ziehen. Dies vermittelt dem Feind ein falsches Suchbild und erschwert so das Widerauffinden der Beute. Abbildung 9: Schreckfärbung des Tagpfauenauges (Inachis io) Seite 20 Patrik Matter 3. Warnung (aposematische Färbung) Während die kryptische Färbung die Auffälligkeit eines Tieres auf ein Minimum reduziert, erhöht eine aposematische Warnfärbung die Auffälligkeit auf ein Maximum. Eine hohe Auffälligkeit ist in der Natur mit Selbstmord gleichzusetzen. Daher können sich nur Tiere, die besondere Schutzvorrichtungen (Gift, Ungeniessbarkeit, Stacheln, etc.) haben, leisten, auffällig zu sein. Die aposematische Färbung beeinflusst das Lernvermögen von Raubfeinden auf unterschiedlichste Weise (Siehe II.2.1.2). Eine aposematische Wirkung wird meistens durch ein schwarzes, weisses, rotes, oranges oder gelbes Muster (Flecken, Ringe, Streifen, etc.) auf einem Hintergrundton mit einem möglichst grossen Kontrast, erreicht (Abbildung 10). Abbildung 10: Typische Aposematische Muster 3.1 Besonderheiten von aposematisch gefärbten Tieren Damit die aposematische Färbung ihre Funktion erfüllen kann, müssen die Tiere sichtbar sein. Daher verstecken sich aposematische Tiere selten und sind meistens tagaktiv (z.B. Kalifornische Salamander ( M.E. Hubbard (1903)). Es gibt jedoch auch aposematische Tiere, die hauptsächlich nachtaktiv sind, da ihre Beute ebenfalls nachtaktiv ist. Wenn ein kryptisches Tier entdeckt wird, versucht es oft zu flüchten oder sich zu verstecken. Ein aposematisches Tier hingegen macht sich gemächlich davon oder zeigt möglicherweise ein Warnverhalten. Da aposematische Tiere es nicht nötig haben, vor Feinden zu fliehen, zeigen sie oft ein sehr träges („sluggish“)Verhalten. Häufig haben sie einen schwerfälligen Gang und sind auf Beute spezialisiert, die einfach zu fangen ist (Bsp. Skunk, Gilatier, etc.). Um versehentliche Attacken zu überleben, sind aposematische Tiere oft sehr robust gebaut (z. B aposematische Schmetterlinge (R. Trimen (1869)). Aposematische Tiere treten oft in grösseren Gruppen auf (M. Speed (2001), G.A. Sword et al. (2000), M. Riipi et al. (2001)). Auffällig ist ausserdem, dass viele aposematische Tiere während des Winters (oder in Dürrezeiten) nicht aktiv sind. Dies könnte damit zusammenhängen, dass Raubfeinde, die dem Hungertod nahe sind, alles fressen, was sie erbeuten können – auch aposematisch gefärbte Tiere (E. Poulton (1908)). Seite 21 Patrik Matter 4. Drohung (aposematische Färbung) Drohungen müssen verlässliche Informationen über Motivation und/oder Kampfstärke eines Gegners geben. Die Motivation hängt vom momentanen Zustand ab, daher kann sie nicht mit permanenter Färbung signalisiert werden. Auch Kampfstärke wird nur in den seltensten Fällen mit der Färbung korrelieren. Daher ist die Funktion der Drohfärbung meistens nicht eine Drohung an sich, sondern eine Unterstützung der Drohung. Da Drohsignale einen ernsten Kampf meistens verhindern, ist es wichtig, dass sie nicht zu fälschen sind. Ausser den so genannten Übersprungshandlungen (unbewusste Signale, die einen Stimmungswechsel anzeigen (II.1.4.3) kommen daher vor allem Handicaps (II.1.4.1) in Frage. 4.1 Statussymbole Statussymbole sind Signale, die bei adulten Männchen deutlicher sind als bei Weibchen oder Jungtieren (Abbildung 11). Abbildung 11: Unterschiedliche Beispiele von Statussymbolen der adulten Männchen - Der unversehrte Kamm eines Gockels (Gallus gallus) zeigt, dass er im Rivalenkampf die Oberhand hat - Je grösser der Brustlatz eines Sperlings (Passer spec.), desto stärker ist sein Träger (Fugle et all 1984) - Der silberne Rücken eines „weisen“ Gorillas (Gorilla gorilla) zeigt Alter (Erfahrung). (Roper 1986) 4.2 Betonende Färbung Viele Gesichtszeichnungen verraten die Blickrichtung schon von weitem. Ein motivierter Gegner schaut seinem Rivalen fest in die Augen, wobei er bewusst darauf verzichtet, andere Informationen (Ausschauhalten nach Feinden, etc.) aus der Umgebung aufzunehmen (A.Zahavi (1997)). Streifen können die Länge bzw. Breite einer Körperform betonen, was wiederum ein wichtiger Hinweis auf die Grösse eines Rivalen sein kann. Farben können zudem einen Körperteil betonen, der Ausschlag gebend für eine Übersprunghandlung ist. Seite 22 Patrik Matter 5. Balz (epigamische Färbung) Bei der Balz geht es den Weibchen darum, einen möglichst optimalen Partner auszusuchen. Dabei ist von Art zu Art verschieden, welche Eigenschaften (Fütterung des Weibchens und seiner Nachkommen, gute Gene, etc.) diesen Partner optimal erscheinen lassen. 5.1 Statussymbole Statussymbole sind auch epigamische Signale, da die männlichen Nachkommen denselben Vorteil haben wie ihre Väter. 5.2 Betonende Färbung Auch als Balzfärbung kann eine Färbung alle möglichen positiven Eigenschaften betonen. 5.3 Symmetrie Oft führen Stress oder genetische Mängel zu asymmetrischer Entwicklung von Körperteilen. (P. Parson (1990)). Es könnte daher gut sein, dass die vielen symmetrischen Muster und Formen im Tierreich ein verlässliches Indiz für die genetische Qualität eines Trägers sind. Zusätzlich wirken sie auch als Handicap, da eine symmetrische Form auf einem asymmetrischen Körper die Asymmetrie des Körpers betont. Ein weiterer Grund für symmetrische Formen könnte auch sein, dass sich diese sehr gut vom meist asymmetrisch gemusterten Hintergrund abheben. 5.4 Augenmuster Viele Tiere haben kreisartige, komplizierte Zeichnungen, die als Augenmuster bezeichnet werden. Diese könnten als Mimikry von Tieraugen funktionieren, als Schreckfärbung dienen (D.Schlenoff (1985)) oder eine epigamische Funktion haben, in dem sie entweder die Symmetrie betonen oder eine Mimese von Eiern (W.Wickler (1962)) oder Samenkörnern (R.Schenkel (1956)) sind. 5.5 Färbung, die den körperlichen Zustand betont Bei einigen Vögeln und Säugern gibt es Körperstellen, an denen die nackte Haut sichtbar ist (z.B. Hahnenkamm, menschliche Lippen, etc.). Bei diesen ist die Färbung ein direkter Indikator für die Durchblutung jener Stellen. Bei Stress, Krankheit oder Kälte verengen sich die Blutgefässe. Somit ist die Durchblutung ist vermindert. Auch haben verschiedene Studien gezeigt, dass bunte Farben ein Indiz für wenig Parasiten sein können. (W.Hamilton und M.Zuk (1982), Milinski&Bakker (1990)). A.Zahavi (1997) verallgemeinert diese Aussage, indem er die schlechtere Färbung auf den durch Parasiten bedingten Stress zurückführt. Daraus folgert er, dass jegliche Art von Stress (Krankheit, Hunger, etc.) diese Symptome verursachen kann. Glanz kann ebenso ein Indiz für die Gesundheit sein. So haben kranke Amphibien oft eine trockenere, weniger glänzende Haut als Gesunde. Glänzende Federn nutzen sich zudem schneller ab, als normal gefärbte, daher müssen sie besonders oft erneuert werden, was sich nur gesunde Tiere leisten können. Seite 23 Patrik Matter 5.6 Sichtbarkeit (vgl. A. Zahavi 1997) Eine wichtige Funktion der epigamischen Färbung ist, potentielle Partner schon von weitem sichtbar zu machen. Da die erhöhte Sichtbarkeit auch für Feinde und Rivalen gilt, funktioniert eine auffällige Färbung zusätzlich auch noch als ein Handicap. Die Farbwahl hängt ausserdem sehr stark von der Umgebung ab (vgl. IV.2). 5.6.1 Dunkle Färbung Dunkle Farben absorbieren den grössten Teil des Lichtes, daher grenzen sie sich deutlich von ihrer Umgebung ab. Durch diesen hohen Kontrast werden die Konturen des Körpers betont, was wiederum zur Folge hat, dass Grösse und Form einfacher zu erkennen sind. Helle Farben sind jedoch aus weiterer Distanz sichtbar als dunkle. 5.6.2 Helle Färbung Helle Farben reflektieren den grössten Teil des Lichtes. Daher sind sie auch auf weite Distanz sichtbar. Allerdings haben sie den Nachteil, dass durch die Reflektion der Rand des Körpers verzerrt wird. Da bei sich bewegenden Körpern nur verzerrte Konturen zu erkennen sind, eignen sich helle Farben zur Betonung von Bewegungen. Bei kleinen Tieren können zu helle Farben dazu führen, dass sich deren Umrisse ins Nichts verzerren. Deswegen haben kleine Tiere oft eine dunkle Färbung. 5.6.3 Reflektierende Färbung Glänzende Farben streuen das Licht in eine einzige Richtung. Daher sind sie ebenfalls geeignet, Bewegung zu zeigen. Allerdings haben sie nicht den Nachteil, die Form zu verzerren. (Bsp. Kolibri). Zusätzlich haben glänzende Farben in heissen Gegenden auch den Vorteil einer erhöhten Reflektion. Daraus resultiert eine verminderte Aufnahme von Hitzestrahlung. 5.6.4 Gemischte Färbung Oft werden helle und dunkle Farben kombiniert, um einerseits die Form, andererseits die Bewegung zu betonen (z.B. Vogelflügel). Der grosse Kontrast zwischen Schwarz und Weiss führt zu einer optischen Verkleinerung des Körpers. Deswegen wird häufig eine der Farben durch eine Kompromissfarbe ersetzt. Die Wirkung von kombinierten Farben hängt ausserdem stark von der Betrachtungsdistanz ab. Ähnliche Farben verschwimmen schon ab geringer Distanz zu einer einfarbigen Mischfarbe. Selbst das auffällige Muster der Zebras wandelt sich ab genügender Distanz in ein tarnendes Grau. 5.7 Sensorische und „mentale“ Fenster Eine weitere mögliche Funktion von Balzfärbung ist die Stimulierung eines sensorischen oder mentalen Fensters des Partners (T. Bakker (1993)). Seite 24 Patrik Matter 5.8 Fishers Hypothese über die sexuelle Selektion Einige Männchen mit guter Fitness haben ein zufälliges optisches Merkmal, Weibchen eine zufällige Präferenz für dieses. Die Nachkommen erben Fitness und Präferenz. Dadurch verbreiten sich beide Merkmale in der Population. Ein Männchen mit „besserem“ optischem Merkmal hätte wiederum einen Vorteil, und so beginnt eine Spirale, die zu immer extravaganteren optischen Merkmalen führt (R.Fisher (1930)). 6. Arterkennung (episematische Färbung) Die auffällige Zeichnung von Fischen im Korallenriff, könnte dazu dienen, dass Artgenossen sich untereinander erkennen, was bedeutet, dass sie nicht fälschlicherweise gegen andere Arten ihr Revier verteidigen oder um Fortpflanzungsgelegenheiten balzen (K. Lorenz (1962)). Smith (1966) bestätigte diese Ansicht mit einem Versuch, bei dem er die Färbung nahe verwandter Möwenarten manipulierte, indem er einer Möwenart die artspezifischen Merkmale der anderen aufmalte. Dadurch wurden die manipulierten Möwen wie arteigene behandelt. 7. Physikalische Eigenschaften (vgl. (E.Burtt(1977)) 7.1 Thermoregulation Je dunkler eine Fläche ist, desto mehr Strahlung absorbiert sie. Deswegen wärmen sich durch Sonnenstrahlung dunkle Flächen schneller auf als helle. Auch Glanz kann durch die Reflektion von Strahlung eine thermoregulatorische Funktion haben. Wechselwarme Tiere mit Farbwechselfähigkeit nutzen dieses Prinzip, um sich schneller aufzuwärmen. Chamäleons (Chamaeleon spec.) nehmen beim Sonnentanken morgens eine sehr dunkle Färbung an. (Grzimeks Tierleben ( 1971) H. Schifter)) Vielfach sind die ansonst weissen Winterfelle von Tieren aus thermoregulatorischen Gründen an den Spitzen von Ohren und Schwänzen schwarz gefärbt. Von dort wird die aufgenommene Wärme durch die starke Durchblutung auf den ganzen Körper verteilt. 7.2 Schutzfunktion Die Farbstoffe Melanin und Karotin schützten gegen die schädliche ultraviolette Strahlung. Melaninhaltige Federn sind zudem besser vor Abnutzung geschützt, die resultiert, wenn Vögel im Luft mit kleinsten Teile zusammenstossen (E.Burtt (1979)). Nach der gloger’schen Regel (C. Gloger (1833)) sind in wärmeren Gebieten (Gebieten mit höherer Sonneneinstrahlung) die Tiere aus Gründen des UV-Schutzes dunkler gefärbt als verwandte Arten in kalten Regionen. Seite 25 Patrik Matter 8. Keine selektive Funktion Natürlich könnte es auch sein, dass die Färbung gar keine Funktion hat. Eine solche Erklärung sollte aber nur in Betracht gezogen werden, wenn alle Alternativen ausgeschlossen wurden. 8.1. Ausprägung einer pleiotropischen Wechselwirkung Theoretisch wäre es denkbar, dass die Färbung das Resultat einer pleiotropischen Wechselwirkung ist, dass also die Gene, die für ein wichtiges selektives Merkmal zuständig sind, ebenfalls für die Ausprägung der Färbung verantwortlich sind. 8.2 Historisches Relikt Es kann auch sein, dass früher ein Selektionsdruck vorhanden war, der jedoch unter den heutigen Lebensbedingungen abgeschwächt ist oder nicht mehr existiert. In diesem Fall stellt die heutige Färbung daher ein historisches Relikt dar. 8.3. Zufällige Färbung Falls alle anderen Erklärungen sich als nicht plausibel erweisen, ist es möglich, dass die Färbung rein zufällig entstanden ist, und dass somit kein oder nur geringer Selektionsdruck ausgeübt wird. Seite 26 Patrik Matter IV. FARBWIRKUNG (VGL J.HAILMAN (1977)) Die Wirkung von Farben hängt sehr stark von der Umgebung (Licht, Übermittlungsmedium, Hintergrund), sowie vom Verhalten der Träger ab. Um die Färbung von Tieren zu verstehen, muss man demzufolge wissen, welches Verhalten unter welchen Bedingungen die Tiere zeigen. 1. Licht Die Farbwirkung ist in erster Hinsicht vom Licht abhängig, in dem die Farben betrachtet werden. Sowohl die Lichtmenge (Quantität) wie auch die Lichtzusammensetzung (Qualität) spielen eine wichtige Rolle. 1.1 Sonnenstand Bei Sonnenaufgang und Sonnenuntergang ist das flach einfallende Licht orange (Abbildung 12), denn die Sonnenstrahlung legt in diesem Moment einen weiteren Weg zurück. Somit wird durch die RayleighStreuung an Luftpartikeln kurzwelliges Licht (-550 nm (gelb)) seitlich gestreut, so dass hauptsächlich das langwellige Licht (ab 590 (orange)) auf die Erde trifft. Abbildung 12: Oranger Sonnenuntergang Rayleight-Streuung färbt das Licht orange Am Mittag fällt das Licht steiler auf die Erde. Daher wird hauptsächlichkurzwelliges blaues Licht (-450 nm) gestreut, was bewirkt, dass der Himmel blau und das Licht gelb scheint. Der Sonnestand variiert natürlich auch mit dem Breitengrad, wobei auch hier das einfallende Licht umso kleinere kurzwellige Anteile hat, je flacher es einfällt. Tiere, die unter solchen Bedingungen Signalfarben präsentieren wollen, sollten daher speziell rote, orange oder gelbe Signalfarben benutzen (Bsp. Die nördlichen Zeisige (Acanthis spec.)) 1.2 Höhe über Meer Da der Druck in der Höhe abnimmt, wird mit zunehmender Höhe die Dichte der Atmosphäre ebenfalls kleiner. Je dichter die Atmosphäre ist, desto grösser ist der Effekt der Rayleigh-Streuung. Schon relativ kleine Höhenunterschiede können bemerkbare Verschiebungen des Strahlungsspektrums ergeben. Daher kommen im Gebirge auch kurzwellige Wellenlängen (400nm (violettpurpur)) vor. (Carduelis cannabina) Abbildung 13: purpurne Signalfarben im Gebirge Im Gebirge können aus diesem Grund auch „purpurne“ Signalfarben vorkommen (Abbildung 13). Seite 27 Patrik Matter 1.3 Dunkelheit Im Dunkeln sind Farben nur noch als Helligkeitsunterschiede erkennbar. Als Signalfarben kommen deshalb nur sehr helle Farben in Frage, am besten Weiss. Zum Beispiel die weissen Blüten von Pflanzen, die durch nachtaktive Tiere (Fledermäuse, Motten, etc.) bestäubt werden. Die Alternative dazu ist Biolumineszenz. Besonders in den lichtlosen Tiefen der Ozeane ist Biolumineszenz weit verbreitet und übernimmt verschiedenste Funktionen der Färbung (Gegenschattierung, epigamische Färbung, Schreckfärbung, etc.). 2. Übermittlungsmedium Das Übermittlungsmedium beeinflusst Signale in zweierlei Weise. Einerseits bestimmt es das Spektrum, das von Signalfarben reflektiert werden kann, andererseits „filtert“ es auch das vom Objekt reflektierte Licht erneut. 2.1 Streuung an Partikeln Wenn sich Licht nicht durch ein Vakuum bewegt, wird es an kleinsten Partikeln des Mediums gestreut. Die Art der Streuung hängt dabei stark von der Partikelgrösse ab. Teilchendurchmesser X < 1/10 λ 1/10 λ < X <25 λ 25 λ < X Streuungsart Rayleight-Streuung Mie-Streuung Spiegelung, Brechung 2.1.1 Rayleight-Streuung ( Partikeldurchmesser < 1/10 λ ) Bei kleinsten Teilchen ist die Streuung etwa umgekehrt proportional zur 4. Potenz der Wellenlänge (λ). Sr = (n-1)2 / λ4) λ= Wellenlänge V = Geschwindigkeit n = Brechungsindex = V(Vakuum) / V (Medium) Seite 28 Patrik Matter 2.1.2 Mie-Streuung ( 1/10 λ< Partikeldurchmesser < 25λ λ) Bei kleinen Teilchen hängt die Streuung vom Teilchenradius und in komplexer Weise von der Wellenlänge und dem Partikelradius (y) ab (Abbildung 14). Die meisten natürlichen Medien sind Stoffgemische. Da schon kleine Unterschiede in der Partikelgrösse sehr unterschiedliche Effekte auf die Streuung haben können, ist die MieStreuung in der Natur nur sehr schwer berechenbar. In Lebensräumen in denen Mie-Streuung die Hauptursache der Streuung ist, müssen daher zuerst quantitative Daten gesammelt werden, um die Form der Streuung zu erfassen. Mie –Streuung: Sm = y2 * M (n, y/λ) y = Partikelradius λ= Wellenlänge n = Brechungsindex M= Siehe (Abbildung 14) Abbildung 14: Mie-Streuung (Funktion M) 2.1.3 Normale Spiegelung (Partikeldurchmesser > 25λ λ) Bei so grossen Teilchen gelten die bekannten, optischen Reflektions- und Brechungsgesetzte, wobei kurzwelliges Licht weniger gebrochen wird als langwelliges. Reflektion (Einfallswinkel = Ausfallswinkel): α0 = α1 Brechung beim Übergang von n1 nach n2 (n1<n2): sin(α0) / sin(α2) = n2/n1 Seite 29 Patrik Matter 2.2 Streuungen in versch. Medien 2. 2.1 Luft (Nebel, Aerosol, etc) Transparente Filter verändern die Wirkung von Farbe enorm, in dem sie einerseits den Farbton verändern und andererseits den Kontrast schwächen oder verstärken. Dies kann durch komplizierte Modelle berechnet werden. (V.Chen et al. (1998). Nebel und Aerosole können als solche transparente Filter behandelt werden. (J. Hagedorn (2000)). Farbtöne, die eine gelbe Komponente enthalten erscheinen am hellsten und solche mit blauer Komponente am blassesten (Takamatsu M et al. (2001)). 2.2.2 Wasser Wasser streut Licht umso mehr, je länger dessen Wellenlänge ist. Lichtstreuung im Wasser ~λ Je tiefer man taucht, desto mehr verschiebt sich das Spektrum gegen die kurzwellige (blaue) Seite und die Kurve wird gleichmässiger. (Abbildung 15) Wenn Licht auf Wasser trifft, hat das angrenzende Medium einen weiteren grossen Einfluss auf die Lichtqualität. So ist zum Beispiel die Intensität des Lichtes bei Bewölkung deutlich verringert. a) Strahlung von oben oberhalb eines Ozeans bei bewölktem und unbewölktem Himmel b) Strahlung von unten und von oben drei Meter unterhalb der Wasseroberfläche. Einheit der y-Achse: Lichtquanten pro (cm2 * s * nm) Abbildung 15: Strahlungsintensität im Ozean (McFarland & Munz ( 1975)) Allerdings gilt dies nur in klarem Wasser, da Trübungen im Wasser einen gegenteiligen Effekt haben, der bewirkt, dass sie kurzwelligeres Licht mehr streuen (Luria et al. (1970)). Lichtstreuung durch Trübung ~ 1/λ Noch komplizierter wird es in flachem Wasser, da hier die Reflektion des Untergrundes einen grossen Teil des Spektrums ausmacht (Mc Farland et al. (1975)). Seite 30 Patrik Matter 2.2.3 Vegetationsreiche Medien Rotkehlchen Da Chlorophyll hauptsächlich grünes Licht reflektiert, hat das Spektrum in vegetationsreichen Medien oft einen hohen Grünanteil. Eine rote oder orange Signalfärbung könnte hier den grössten Kontrast bieten (Abbildung 16). 2.2.4 Optische Hindernisse (Pflanzen, Riffblöcke, etc.) (Erithacus rubecula) Abbildung 16: orange Färbung im Wald Optische Hindernisse verdecken meistens einen Teil des Senders. Daher gibt es zwei alternative Strategien für die Signalfärbung. Entweder sollten sich grosse Signale herausbilden, welche, selbst wenn nur ein Teil des Signals sichtbar ist, erkennbar sind oder sie sollten so klein sein, dass sie zwischen den Lücken der Hindernisse sichtbar sind. Eine weitere Möglichkeit könnte die Ausbildung von symmetrischen Signalen sein, die sich von asymmetrischen Hindernissen abheben (Abbildung 17). Abbildung 13: Mit vielfältigen Mustern stechen Tiere auch hinter optischen Hindernissen hervor. 3. Hintergrund Entscheidend für die Wirkung von Signalfärbung ist der Hintergrund, von welchem sie sich abheben muss. Um einen groben Überblick zu geben, wurden die Lebensräume in drei Gruppen unterteilt, die verschiedenste Ansprüche an die Signalfärbung ihrer Bewohner stellen. Natürlich sind auch Zwischenstufen und Kombinationen von Lebensräumen möglich, die wiederum eine besondere Betrachtung erfordern würden. Seite 31 Patrik Matter 3.1 Offene Lebensräume (Wüsten, Himmel, Polarzonen, Strände, Tiefsee, etc.) Um sich von einem hellen Lebensraum abzuheben, eignen sich dunkle Färbungen besonders gut. Die Alternative dazu sind glänzende Farben, welche zusätzlich noch als Strahlungsschutz wirken können. Dunkle Lebensräume hingegen fordern eine helle Färbung (Abbildung 18). Abbildung 14: helle und dunkle Färbungen sind in offenen Lebensräumen Ideal (Rabenkrähen (Corvus Corone) und Möwen (Larus spec.)) 3.2 Wasserflächen (See, Ozean, etc) Wasserflächen haben besondere Ansprüche an Signalfärbung, da sie je nach Betrachtungswinkel verschiedenste optische Wirkung haben. Blickt man gegen die Sonne, erscheint das reflektierende Wasser sehr hell. Hat man hingegen die Sonne im Rücken erscheint das Wasser in dunklen Farbtönen (blau, grün, schwarz-blau, etc). Eine Signalfärbung müsste sich daher von allen Bedingungen genügend abheben. Daher sind zweifarbig dunkel –hell gefärbte Tiere häufig, die oft besondere Stellen durch Kontrastfarben von blau, also grün, gelb oder orange betonen, welche oft glänzend sind (Abbildung 19). Abbildung 19:Glänzende und blaukontrastierende Signalfärbung auf Wasserflächen (Papageientaucher (Fratercula arctica), Stockente (Anas platyrhynchos), Kaiserpinguin (A, forsteri) Seite 32 Patrik Matter 3.3 Stark strukturierte Lebensräume Stark strukturierte Lebensräume zeichnen sich durch ein fleckiges Muster von Licht und Schatten aus. Teilweise sind sie sehr homogen gemustert (Seegraswiesen, BraunalgenTeppiche, Grasländer, Riede, etc). Da sich weiss und schwarz ungenügend vom Licht bzw. Schatten abheben, können sie in stark strukturierten Lebensräumen nicht als Signalfarben verwendet werden. Dies bewirkt, dass sie oft durch dunkle (braun, rot oder blau) und helle (gelb, grau) oder glänzende Kompromissfarben ersetzt werden (Abbildung 20). Abbildung 20: Buntheit als Anpassung an den stark strukturierten Lebensraum: „Tropenwald“ (Quetzal(Pharomachrus mocinno), Ara(Ara spec.), Tukan(Tockus spec.), Paradiesvogel (Paradisaea apoda) In homogen gemusterten Lebensräumen kann auch ein kontrastierendes Muster (z.B Querstreifung im Ried) als Signalfärbung eingesetzt werden. 4. Verhalten Einen riesigen Einfluss auf die Wirkung ihrer Signalfärbung hat natürlich auch das Verhalten der Tiere. So kommt es, dass männliche Vögel oft von erhöhten Warten aus balzen, damit sie von Weibchen besser gesehen werden. Andere Tiere bewegen sich während der Balz schnell, was zu einem weiteren Kontrast mit der Umgebung führt. Dies sind nur einige Beispiele der unendlichen Vielfalt von Einflüssen des Verhaltens auf die Evolution der Signalfärbung. Der hellgelbe Säulengärtner (Giardiniere dorato) baut seine Laube auf dem Boden des Regenwaldes. Hellgelb ist eine Färbung, die im grünlichen Licht nicht gut zur Geltung kommt. Das Balzverhalten des Laubenvogels erklärt jedoch diese Unstimmigkeit. Dieser Vogel zupft das Blätterwerk oberhalb der Laube so ab, dass das Sonnenlicht auf die Laube fällt und so sein gelbes Gefieder hervorragend zur Geltung bringt (A. Zahavi (1997)). Seite 33 Patrik Matter V. SEHVERMÖGEN DER WIRBELTIERE (VGL. G.WALLS 1942) In seinem 15 jährigen Studium des Aufbaus der Retina der Wirbeltiere fiel Schultze (1866) auf, dass tagaktive Tiere viel mehr Zapfen in ihrer Retina haben als nachtaktive. Daher formulierte er die Duplizitätstheorie, die postulierte, dass Stäbchen für das Hell-/Dunkelsehen und Zapfen für das Farbsehen verantwortlich sind. Heute ist diese Theorie durch viele empirische Befunde bestätigt. 1. Hell-/ Dunkelsehen mit den Stäbchen Die optisch sensitiveren Sehzellen werden aufgrund ihrer Gestalt Stäbchen genannt und sehen bei allen Tieren ungefähr gleich aus. Sie haben durch das Rhodopsin ein Absorptionsmaximum von 500 nm (Abbildung 21). Legende: a) systematisches Stäbchen b) Protopterus aethiopicus c) Golfisch (Carassius. auratus) d) Leopardenfrosches (Rana pipiens) e) Rhodopsinloses Stäbchen von (Rana pipiens) f) Flughörnchen (Glaucomys volans) g) Mensch (Homo sapiens sapiens) 1.1 Ursachen für die grössere Sensitivität Abbildung 21: Unterschiedliche Stäbchentypen von Stäbchen (Quelle: G.Walls (1942)) 1.1 Ursachen für die grössere Lichtsensitivität von Stäbchen 1.1.1 Form Ein zylinderförmiges Stäbchen hat ein dreimal grösseres Volumen als ein kegelförmiges Stäbchen mit derselben Grundfläche und Höhe. Ein Lichtstrahl, der auf ein Stäbchen fällt, stimuliert daher mehr photosensitives Material, als einer, der auf einen Zapfen fällt. Deshalb sind Stäbchen nur schon aufgrund ihrer Gestalt besser geeignet, schwaches Licht wahrzunehmen, als Zapfen. 1.1.2 Verschaltung Ein weiterer grosser Unterschied in Bezug auf die Sensitivität liegt in der Verschaltung mit den Nervenzellen. Meistens sind viele Stäbchen gemeinsam mit einem Bipolar verknüpft. Somit genügt es, wenn nur einige Stäbchen Licht bekommen, um den Impuls an das Gehirn weiterzuleiten. Zapfen hingegen sind meist nur einzeln mit einem Bipolar verknüpft. Dadurch kann bei Schwachlicht nur ein Teil der Zapfen gereizt werden. Seite 34 Patrik Matter 1.1.3 Rhodopsin Der wichtigste Faktor für die grosse Sensitivität der Stäbchen ist jedoch die in ihnen enthaltene chemische Substanz Rhodopsin. Das rote Rhodopsin absorbiert alle Wellenlängen des visiblen Lichtes ausser rot. Durch diese unspezifische Lichtwahrnehmung hat es eine höhere Lichtsensitivität, als die rot-, grün-, oder blau spezifischen Zapfen. Allerdings zerfällt Rhodopsin durch eine Transformationsänderung in eine optisch inaktive Form, wenn es Licht ausgesetzt ist. Deswegen kann Rhodopsin nur bei Schwachlicht als Verstärker der Sensitivität eingesetzt werden. Am Tag ist es inaktiv und wandelt sich erst, wenn die Lichtintensität unter einen gewissen Schwellenwert fällt, erneut in die optisch aktive Form um. Allerdings dauert es etwa eine Stunde bis die maximale Konzentration erreicht wird. (Dies ist der Grund, weshalb wir uns zuerst an die Dunkelheit gewöhnen müssen, um etwas zu sehen (Dunkeladaption)). 1.2 Doppel-Stäbchen Ebenso wie Doppel-Zapfen kommen bei den Wirbeltieren auch Doppel-Stäbchen vor. Bisher hat man sie bei Geckos sowie bei verschiedenen Schlangen nachgewiesen (G.Walls (1942)). Experimente mit Ameisen haben gezeigt, dass Doppel-Stäbchen dort zur Wahrnehmung von polarisiertem Licht dienen (Mitteilung: Prof. J. Flammer, Augenspital). 2. Farbsehen mit Zapfen (vgl. Gegenfurter et al. (1999)) Die Sehzellen, die für grosses Auflösungsvermögen und Farbsehen verantwortlich sind, werden aufgrund ihrer Gestalt Zapfen genannt. Allerdings sehen Zapfen je nach Tierart sehr unterschiedlich aus (Abbildung 22). a) Stör (Acipenser fulvescens) b) Goldfisch (Carassius auratus) c) Leopardenfrosch (Rana pipiens) d) Schildkröte ( Chelydra serpentine) e) Falke (Circus hudsonius) f) Mensch (Homo sapiens sapiens) (Quelle: G.Walls (1942)) Abbildung 22: Unterschiedliche Zapfentypen Seite 35 Patrik Matter In unserer Netzhaut gibt es drei verschiedene Arten von Zapfen, die sich durch ihre verschiedenen Absorptionsmaxima (440nm, 535nm, 570 nm) unterscheiden. Je nach Tierart kommen bis zu vier verschiedene Arten von Zapfen vor, deren Absorptionsmaxima von Art zu Art verschieden sein kann. 2.1. Ursachen für die höhere Auflösung der Zapfen verglichen mit den Stäbchen 2.1.1 Verschaltung Die bessere Sehschärfe der Zapfen ist durch ihre Verschaltung mit den Nervenzellen bedingt. Oft sind einzelne Zapfen direkt mit einem Bipolar verknüpft. Somit kann die Information von einzelnen Zapfen ausgewertet werden, was zu einem Bild mit viel grösserer Auflösung führt. Theoretisch könnte man auch die Auflösung der Stäbchen durch eine geringere Verschaltung heraufsetzen, nur wäre es sinnlos, die durch die geringe Lichtmenge geforderte Sensitivität der Stäbchen durch geringere Verschaltungen herabzusetzen, wenn in der Netzhaut schon Zapfen enthalten sind, deren Farbsehen eine geringere Verschaltung erfordert. 3. Farbwahrnehmung des Menschen Da das Farbsehen hauptsächlich beim Menschen erforscht wurde, wird dieses zuerst erklärt, um dann auf die Farbwahrnehmung von Tieren schliessen zu können. 3.1 Absorption durch die Zapfen In der menschlichen Netzhaut kommen drei verschiedene Zapfentypen vor, die aufgrund ihrer Absorptionsmaxima in S(shortwave)-Zapfen (440nm(blau)), M-Zapfen (535nm(grün)) und LZapfen (570nm(rot)) unterteilt werden. Diese Zapfen übermitteln an die Bipolarzellen, wie viele Photonen sie absorbiert haben. Wie die Zapfen durch ein Objekt, das sie wahrnehmen, angeregt werden, hängt von der Beleuchtung sowie vom Reflektionsspektrum des Objekts ab (Grafik 23). Das Verhältnis der verschiedenen Zapfentypen ist von Individuum zu Individuum sehr unterschiedlich, wobei die S-Zapfen nur etwa 10% der Zapfen ausmachen. Dies hat jedoch nur geringen Einfluss auf die Farbwahrnehmung. Seite 36 Patrik Matter Abbildung 23: grafische Darstellung der Absorption der Zapfen Seite 37 Patrik Matter 3.2 Verarbeitung der Signale Die Signale welche, die verschiedenen Zapfen empfangen, werden auf komplizierte – noch nicht vollständige erforschte – Art weiterverarbeitet. Abbildung 24: Aufbau der Netzhaut (Krey et al. (1987)) Seite 38 Patrik Matter 3.2.1 Verschaltung in der Netzhaut 3.2.1.1 Bipolarzellen (Rezeptive Felder) Die erste Stufe bildet die Verschaltung der einzelnen Zapfen zu rezeptiven Feldern in den Bipolarzellen (Abbildung 24) der Netzhaut. Durch diese Verschaltung wird der relative Input von einem zentralen Zapfen mit dem relativen Input von den umgebenden Zapfen verglichen. Je nach zentralem Zapfen, kann man so L, M und SFelder unterscheiden (Abbildung 25). Abbildung 25: Grafische Darstellung eines rezeoptorischen Feldes (Quelle: P. Lennie (2000)) 3.2.1.2 Ganglienzellen (Farbwerte) In den Ganglienzellen (Abbildung 24) werden die Signale der Bipolarzellen durch Addition und Subtraktion so weiterverarbeitet, dass drei unterschiedliche Farbwerte unterschieden werden können. Schwarz-Weiss-Wert := L-Feld + M-Feld Rot-Grün-Wert := L-Feld - M-Feld Blau-Gelb-Wert := S-Feld – (L-Feld + M-Feld) 3.2.2 Verschaltung im Gehirn In einem weiteren noch ungenügend erforschten Verarbeitungsschritt werden diese Farbwerte zu den uns bekannten Farbqualitäten (Helligkeit, Farbton, Sättigung) weiterverarbeitet (B.Conway (2003), R. De Valois (1993)). 3.3 Farbqualitäten beim Mensch 3.3.1 Primäre (physikalische) Farbqualitäten (Intensität, Wellenlänge, Reinheit) Die Intensität des Lichtes ist ein Mass für die gesamte Energie – also die Anzahl Photonen des Lichtes. Sie hat die Einheit Joule. Je kürzer die Wellenlänge des Lichtes ist, desto kleiner ist der Energielevel der einzelnen Photonen, was aus der Gleichung für die Photonenenergie (E= h*v) folgt. Die Reinheit von farbigem Licht ist das Verhältnis zwischen dem reinen Licht der dominanten Wellenlänge und dem Total des einfallenden Lichtes („weisses Licht“). Seite 39 Patrik Matter 3.3.2 Sekundäre (psychologische) Farbqualitäten (Helligkeit, Farbton, Sättigung) Ohne technische Hilfsmittel ist es uns nicht möglich, die primären Qualitäten des Lichtes zu bestimmen. Daher sind für meine Fragestellung nicht die primären, sondern die sekundären Qualitäten von Licht ausschlaggebend, welche jedoch in komplexer Weise mit den primären Qualitäten zusammenhängen. 3.3.2.1 Helligkeit Helligkeit Wir können etwa 500 verschiedenen Helligkeiten unterscheiden. Die Helligkeit des Lichtes ist die Summe der Anregung der verschiedenen Zapfentypen (resp. Feldtypen). Allerdings ist noch nicht endgültig geklärt, ob die Information aller Zapfentypen summiert wird oder nur die der L- und M-Felder (Abbildung 26). Sättigung Abbildung 26: optische Darstellung von Helligkeit und Sättigung Seite 40 Patrik Matter 3.3.2.2 Farbton Abbildung 27: Menschliches Farbtonspektrum Wir können etwa 200 Farbtöne unterscheiden, die sich in der Anregung des Rot-Grün- und des Blau-Gelb-Kanals unterscheiden (Abbildung 27) (Hurwich et. all. (1957)). Rot-Grün-Kanal = Rot-Grün-Wert / (Rot-Grün-Wert + Blau-Gelb-Wert) Blau-Gelb-Kanal = Blau-Gelb-Wert / (Rot-Grün-Wert + Blau-Gelb-Wert) Allerdings weicht der errechnete Wert des Farbtons insbesondere bei kleiner Reinheit teilweise stark vom tatsächlich beobachteten Wert ab (T. Kulp et. all. (1995)). 3.3.2.3 Sättigung Wir können etwa 20 verschiedene Sättigungsgrade unterscheiden (Abbildung 26). Sättigung ist der Grad der Buntheit einer Farbe, verglichen mit dem gleichhellen „Unbunt“ (Grauton). Die Sättigung wird vom Cortex als Verhältnis zwischen der chromalen Reizung in Bezug zur Summe von chromaler und achromaler Reizung repräsentiert (Hurwich et. all. (1957)). (Rot-Grün-Wert + Blau-Gelb-Wert) / (Rot-Grün-Wert + Blau-Gelb-Wert + „Weissanteil“) Die Sättigung ist daher in komplizierter Weise abhängig von der Wellenlänge sowie von der Reinheit des Lichtes (Abbildung 28).Allerdings weicht auch hier der errechnete Wert des Farbtons (besondere bei kleiner Reinheit) stark vom tatsächlich beobachteten Wert ab (T. Kulp et al. (1995)). Quelle: (Uchikawa et al (1984)) Abbildung 28: Abhängigkeit der Sättigung von Reinheit und Wellenlänge. Seite 41 Patrik Matter 4. Farbwahrnehmung von Wirbeltieren 4.1 Farbqualitäten bei Tieren Mit dem heutigen Stand der Wissenschaft können keine objektiven Aussagen über das Farbsehvermögen von Tieren gemacht werden. Das wird auch in Zukunft eher unwahrscheinlich sein, weil ein grosser Teil der Farbwahrnehmung im Gehirn der Tiere stattfindet. In diesem Abschnitt wird eine Extrapolation der bekannten Fakten des menschlichen Farbsehen auf das mögliche Farbsehen von tierischen Di, Tri-, Tetra- und Pentachromaten versucht. In Ermangelung eines wahrscheinlicheren Ansatzes wird davon ausgegangen, dass auch Tiere ihre Welt mittels der drei Farbqualitäten Helligkeit, Farbton und Sättigung wahrnehmen. 4.1.1 Helligkeit Da die Lichtabsorption von L- und M-Zapfen sehr nahe bei einander liegt, haben dichromate Vertebraten ein sehr ähnliches Helligkeitsempfinden von Farben wie wir. Je nach Absorptionsspektren der weiteren Zapfentypen kann sich das Helligkeitsempfinden von anderen Tri-, Tetra oder Pentachromaten sehr von unserem unterscheiden. 4.1.2 Farbton Im Farbspektrum der Dichromaten sind nur Unterscheidungen von Blau bis Gelb möglich (Abbildung 29). Abbildung 29: Farbspektrum von Dichromaten (Keine Rot-Grün-felder) Farbspektren von anderen Trichromaten sind dem unseren sehr ähnlich, einzig die quantitative Unterscheidung zwischen zwei Absorptionsmaxima würde sich unterscheiden (Abbildung 30). Abbildung 30: Farbspektrum eines Trichromaten mit anderen Absorptionsmaxima (vgl. Abbildung 25) Seite 42 Patrik Matter Das Farbspektrum eines Tetrachromaten hätte noch eine weiteres Farbwertpaar (FarbeXFarbeY-Wert) und würde sich, im gleichen Rahmen vom unserem unterscheiden, wie sich das unsere von dem eines Dichromaten unterscheidet (Abbildung 31). Abbildung 31: Nicht farblich darstellbares Farbspektrum eines Tetrachromaten (X, Y stehen für ein weiteres für uns nicht vorstellbares Farbpaar) 4.1.3 Sättigung Die Sättigung für Dichromaten könnte durch Farbenblinde beschrieben werden. Trotz intensiver Recherche wurden jedoch noch keine entsprechenden Daten gefunden. Aufgrund der Komplexität der Sättigung ist es nicht vorhersagbar, Sättigungswahrnehmung für andere Tri-, Tetra- oder Pentachromaten ist. wie die 4.2. Spezielle Anpassungen an die Umwelt Zusätzlich zu der unterschiedlichen Zahl an Zapfentypen haben einige Tiere noch spezielle Anpassungen an ihren Lebensraum entwickelt, welche eine vernünftige Hypothese für deren Farbwahrnehmung noch weiter erschweren (G.L. Walls (1942)). 4.2.1 Gelbgefärbte Linsen Viele tagaktive Tiere sowie wir Menschen haben gelbgefärbte Linsen, deren Hauptfunktion darin besteht, die chromatische Abberation zu minimieren. Chromatische Aberration entsteht dadurch, dass, die verschiedenen Wellenlängen ungleich stark gebrochen werden, wenn Licht gebrochen wird. Das kurzwellige Licht fällt daher nicht mehr exakt auf den Brennpunkt der Linse und sorgt somit für ein verschwommenes Signal. Gelbe Linsen absorbieren jedoch das kurzwellige (violett) Licht und verhindern so ein verschwommenes Signal. Dies hat jedoch den Nachteil, dass das absorbierte, violette Licht zu einem Verlust von Farbinformationen führt. 4.2.2 Öltröpfchen Die ältesten Fische (Lungenfische) sowie moderne Reptilien, Schildkröten und Vögel besitzen auf ihren Zapfen gefärbte Öltröpfchen (S. Robinson (1994)). Es kommen meistens mehrere Farben von Öltröpfchen mit Farben von gelb bis rot vor. Die Farbe ändert sich als Anpassung an variierende Umweltbedingungen laufend. Seite 43 Patrik Matter 4.2.2.1 Öltröpfchen als Filter gegen die chromatische Abberation Ebenso wie gelbe Linsen wirken die Öltröpfchen als Filter für die chromatische Abberation. Da aber nicht alle Zäpfchen einen solchen Farbfilter besitzen, führt dies nur zu einem minimalen Verlust an Farbinformation. Je dunkler der Farbton der Tröpfchen ist, desto besser ist die Filterwirkung. Allerdings können dunkle Tröpfchen nur realisiert werden, wenn das gefilterte Licht genügend hell ist (z.B. auf spiegelnden Wasseroberflächen). 4.2.2.2 Öltröpfchen erhöhen die Farbauflösung. Wie aus Mischexperimenten bekannt ist, sehen Menschen bei gleichem Anregungsverhältnis der drei Zapfen den gleichen Farbeindruck. Solche Farben, die trotz unterschiedlicher Zusammensetzung den gleichen Farbeindruck verursachen, nennt man metamere Farben. Eine weitere Funktion der Öltröpfchen ist die Verhinderung von Metamerie. Durch die unterschiedliche Verteilung der Öltröpfchen wird es sehr unwahrscheinlich, dass unterschiedliche Wellenlängen einen gleichen Farbeindruck verursachen (M. Vorobyev (2003)). 4.2.2.3 Öltröpfchen als Anpassung an die Lichtbedingungen Eine weitere mögliche Funktion der Öltröpfchen könnte die Anpassung an Lichtbedingung sein. Dies würde erklären, wieso Vögel am frühen Morgen oder am späten Abend, wenn die grössere Rayleight-Streuung das Licht „rot“ färbt, den grössten Prozentsatz an roten Öltröpfchen haben. Im Verlauf des Tages sind die Tröpfchen zusehends gelber (an bewölkten Tagen sogar farblos) (G.L Walls (1942)). Auch unter nebligen Bedingungen sind gelbe Filter sehr hilfreich, da sie den Kontrast verstärken (G.Rieger (1992)). 4.2.3 Doppel-Zapfen Doppel-Zapfen sind im Tierreich weit verbreitet und kommen bei allen Wirbeltieren ausser den Säugern vor. Die Funktion der Doppel-Zapfen ist noch nicht vollständig geklärt. Mögliche Funktionen scheinen jedoch das Sehen von polarisiertem Licht (Cameron et al. (1991), I.Flamarique (1998)) oder eine Anpassung für die Erkennung von Bewegungen zu sein (Campenhausen et al. (1998)). 4.3. Verbreitung des Farbsehens bei Wirbeltieren Fische, Reptilien und Vögel haben bis zu vier unterschiedliche Zapfentypen, Primaten und Beuteltiere bis zu drei und die restlichen Säuger bis zu zwei. Da jedoch (wie oben erwähnt) eine Anpassung an eine nachtaktive Lebensweise mehr Sensibilität und somit weniger Zapfen erfordert, gibt es viele Ausnahmen, die Zapfentypen zugunsten von Stäbchen reduziert haben. Seite 44 Patrik Matter Um das Farbsehen von Tieren zu erforschen, gibt es zwei grundsätzliche Ansätze. Beim quantitativen Ansatz werden die Absorptionsmaxima der unterschiedlichen Zapfen gemessen. Der qualitative Ansatz hingegen geht von Verhaltensexperimenten mit Tieren aus, wobei die Tiere aus verschiedenen hellen achromatischen Grautönen einen Farbton heraussuchen müssen. Falls ein Tier farbenblind ist, kann es nicht zwischen dem gleichhellen Grau und dem Farbton unterscheiden. Deshalb besitzen die Tiere Farbsehen, die diese Tests erfolgreich absolvieren. (Siehe Anhang: XII. Tabellen). 4.3.1 Weiterführende Daten H. Walls (1940) war der erste, der sich eingehender mit diesem Thema befasste. G. Jacobs (1993) verfasste einen sehr detaillierten Artikel über das Farbsehen der Säuger. Cuthill et al. (2000) sammelte detailliertere Daten über das Farbsehen von Vögeln. C.Arese et al. (2002) formulierte die interessante Hypothese, dass australische Beuteltiere grundsätzlich Trichromaten sind. L.Peichl et al. (2001) fand heraus, dass für Wale und Robben der Ozean nicht blau ist, da sie ihre S-Zapfen reduziert haben. 5.Evolution des Farbsehens bei Landwirbeltieren 5.1 Anzahl verschieden Zapfentypen Es gilt heute als wahrscheinlich, dass der Vorfahre der Landwirbeltiere ein Tetrachromat war. Viele Gruppen haben jedoch durch Anpassung an eine nachtaktive Lebensweise Zapfen zugunsten der sensiblen Stäbchen zurückgebildet. Deshalb haben viele Amphibien weniger als vier Zapfentypen. Der Grund der allgemeinen Dichromatie der Säuger liegt vermutlich an der nachtaktiven Lebensweise unserer kleinen insektenfresserähnlichen Vorfahren im Schatten der Dinosaurier. Die Vorfahren der australischen Beuteltiere scheinen entweder nur einen Zapfentyp zurückgebildet zu haben oder dieser wurde durch eine Mutation zurück gewonnen. Dies ist zumindest bei den Primaten passiert, bei welchen sich aus dem ursprünglich langwelligen Zapfentyp die heute verbreiteten L- und M-Zapfen entwickelt haben. 5.2 Öltröpfchen Der Australische Lungenfisch (Neoceratodus forsteri), einer unserer ältesten Vorfahren, besitzt farbige Öltröpfchen. Auch hier ist eine Anpassung an eine nachtaktive Lebensweise ein Verlust der Öltröpfchen (zumindest ein Verlust derer Farbe, mit welchem sie ihre Funktion verlieren). Daher haben heute nur noch Echsen, Schildkröten und Vögel die Farbigkeit ihrer Öltröpfchen erhalten (S.Robinson (1994)). Jacobs (1993) vermutet, dass hier der Grund liegt, weshalb noch keine Säuger sekundär ein tetrachromatisches Sehvermögen entwickelt haben, denn die Absorptionsmaxima der drei Zapfentypen überlappen sich ohne Öltröpfchen zu sehr. Dies könnte bewirken, dass aus einem vierten Zapfentypen nur unwesentliche und zusätzliche Informationen gewonnen werden. Auch durch die gelbe Linse vieler tagaktiver Säuger (G.Walls (1940)) kann der ultraviolette Bereich nicht erschlossen werden. Seite 45 Patrik Matter 6. Spezielle Arten von Farbsehen 6.1 Farbsehen von Monochromaten Experimente mit menschlichen Monochromaten haben gezeigt, dass diese mittels des Vergleiches von Stäbchen und Zäpfchen Farbtöne im Bereich von 440 bis 500 nm unterscheiden können. ( A.Reitner et al. (1991)). Es ist daher auch möglich, dass Tiere einen reicheren Farbeindruck besitzen, als ihre Zapfenzahl vorhersagt. 6.2 Farbsehen im Dunkeln Sobald die Helligkeit zu gering wird, verlieren die menschlichen Zapfen ihre Funktion und die Stäbchen repräsentieren unsere Umwelt mit Grautönen. Bisher wurde angenommen, dass dies für das ganze Tierreich gilt. Kürzlich fand jedoch A.Kelber et al. (2002) heraus, dass dies nicht für alle Tiere gelten muss, zumindest verliert die Motte (Deilephila elpenor) ihr Farbsehen im Dunkeln nicht. Das Farbsehen im Dunkeln wird durch den speziellen AugenBauplan dieser Insektengruppe ermöglicht, indem über 500 Facetten die Farbinformation für einen Punkt liefern. Es ist daher sehr unwahrscheinlich, dass auch Wirbeltiere im Dunkeln Farbe wahrnehmen können. Seite 46 Patrik Matter VI. FEUERSALAMANDER Abbildung 32: Adulter und juveniler Feuersalamander (Salamandra salamandra) 1. Lebensweise 1.1 Lebenszyklus Nach über halbjähriger Entwicklung der Embryonen im Mutterleib werden die Kaulquappen im Frühling in einem geeigneten fischfreiem Gewässer abgesetzt. Die Metamorphose erfolgt je nach Wassertemperatur in 3-5 Monaten. Sehr kaltes Wasser oder Hungerstress können die Larven jedoch dazu zwingen, im Gewässer zu überwintern. Nach abgeschlossener Metamorphose sind die Salamander landlebend. Sie verstecken sich die meiste Zeit in feuchten Tagverstecken und gehen hauptsächlich in der Nacht bei grosser Luftfeuchtigkeit auf Jagd. Im Winter ziehen sie sich in geeignete, frostsichere Winterverstecke zurück. Nach drei Jahren sind die Salamander geschlechtsreif. Feuersalamander können in Gefangenschaft bis zu 50 Jahre alt werden (Böhme (1979)). In freier Wildbahn liegt der bisherige erfasste Rekord bei 20 Jahren (Feldmann (1987)), obwohl sicherlich auch diese Tiere deutlich älter werden können. 1.2 Lebensraum (vgl. B.Tiesmeier & R.Günther (1996)) Da der Feuersalamander eine zweiteilige Entwicklung durchläuft, muss sein Lebensraum sowohl für die Larven als auch für die Adulttiere geeignet sein. Der Lebensraum der Larven ist ein fischfreies Gewässer. Hierbei kann es sich um ein von einer Quelle gespeistes Fliessgewässer (75%) oder um ein stehendes Gewässer (25%) Handeln. Seite 47 Patrik Matter Der typische Lebensraum der Adulttiere sind feuchte Laubmischwälder der collinen bis submontanen Stufe. Nadelwälder sind normalerweise zu trocken. Wenn jedoch eine genügend dichte Krautschicht aus Moosen und Farnen vorhanden ist, welche die Feuchtigkeit speichert, kann sich der Feuersalamander auch hier etablieren. Grundsätzlich kann der Feuersalamander überall dort überleben, wo es genügend feucht ist und genügend Tagverstecke vorhanden sind. 1.3 Tagesrhythmus (vgl. R. Klewen ( 1991)) Der Tagesrhythmus des Feuersalamanders wird vor allem von den Lichtverhältnissen und ferner durch die relative Luftfeuchtigkeit bestimmt. Erst unter 2°C und über 30°C wird die Temperatur zum limitierenden Faktor. Bei hinreichender Luftfeuchtigkeit (85% relative Luftfeuchtigkeit) verlassen die Salamander kurz nach der Dämmerung ihre Tagverstecke und suchen sie in der Morgendämmerung wieder auf. Der Feuersalamander kann daher als streng nachtaktive Art bezeichnet werden. Vereinzelt werden nachts auch Exemplare bei sehr niedriger Luftfeuchtigkeit beobachtet. Bei heftigen Regenfällen nach langer Trockenheit wird der Feuersalamander bereits in der Dämmerung, manchmal sogar tagsüber, in grösserer Zahl beobachtet (B.Tiesmeier & R.Günther (1996)). 1.4 Jahresrhythmus Ein erstes Aktivitätsmaximum ist die Eiablage der Weibchen. Diese erfolgt in den Monaten März-April, in denen Weibchen 60% der beobachteten Tiere ausmachen. Danach sind Weibchen für einige Wochen nicht mehr im Gelände anzutreffen. Es scheint, dass sie diese Zeit zur Erholung brauchen. Ein zweites Aktivitätsmaximum wird zwischen Juli und Oktober erreicht. Zu dieser Paarungszeit sind allerdings zu 75% Männchen zu beobachten. Ein drittes Aktivitätsmaximum fällt in den Zeitraum zwischen August und September. Hier verlassen die juvenilen Feuersalamander das Wasser und machen somit den grössten Teil der Beobachtungen aus. Spätestens wenn der erste Schnee fällt, ziehen sich die Salamander in ihre frostsicheren Winterverstecke zurück, was bewirkt, dass Beobachtungen in Januar und Februar selten sind. 1.5 Paarung (vgl. R. Klewen ( 1991)) Die Paarung findet nachts von März bis September (mit einem deutlichen Höhepunkt im Juni) statt. Es ist noch nicht geklärt, ob die Männchen dabei durch Duftstoff von den Weibchen angelockt werden (Melsheimer (1886)). Die eigentliche Paarung besteht aus den gleichen fünf Phasen wie beim Alpensalamander. Wie bei diesem genügt eine Salamander ähnliche Silhouette als Auslöser für die Verfolgung. 1. Verfolgung 2. Aufsteigen und Kopfreiben 3. Unterkriechen, Klammern und Kehlreiben 4. Schwanzwurzelreiben und Absetzen der Spermatophore 5. Aufnahme der Spermatophore durch das Weibchen Im Zusammenhang mit der Paarung sind auch territoriale Verhaltensweisen ( W.Duellman et al.(1986)) sowie Rivalenkämpfe beschrieben worden (W. Kästle (1986)). Seite 48 Patrik Matter 1.6 Rivalenkämpfe des Feuersalamanders Bei Rivalenkämpfen der Feuersalamander gehen grössere (ältere) Männchen meistens als Sieger hervor. (Abbildung 33) Es könnte sich dabei jedoch auch um eine Fehldeutung von Fehlpaarungen handeln (R.Klewen (1991)). 1.7 Nahrung des Feuersalamanders Abbildung 33: Rivalenkampf (Kästle (1986)) Wie die meisten Schwanzlurche sind auch Feuersalamander Nahrungsgeneralisten, die keine spezielle Beute bevorzugen. Alles, was der Grösse nach in ihr Suchschema passt, wird gefressen. Die Nahrung der Salamander Larven spiegelt daher die Bachbiozönose wieder. Adulttiere fressen vor allem Schnecken und Regenwürmer, aber auch Spinnentiere und Insekten stehen auf ihrem Speiseplan (Tiesmeier B, Günther R (1996), B.Tiesmeier (1992)). 1.8 Feinde des Feuersalamanders Beobachtungen im Labor ergaben, dass Ratten, Hühner und Enten juvenile Feuersalamander fressen. Über natürliche Feinde des Feuersalamanders gibt es nur wenig Berichte und seltene Beobachtungen. Igel, Iltis, Dachse, Wildschweine, Ratten, verschiedene Vogelarten (Graureiher , Weissstorch, Grosse Rohrdommel), aber auch Schlangen (Ringelnatter) werden als Raubfeinde genannt (Joly (1968), Kabisch and Belter (1968), Sauer and Weisbecker (1994), Dietrich (1999)). Als weitere potentielle Feinde kämen Raubtiere (Fuchs, Wolf, Marder etc.), Eulen, Greifvögel, Rabenvögel und Blindschleichen in Frage. Seite 49 Patrik Matter 2. Färbung der verschiedenen Unterarten Abbildung 34: Färbung der unterschiedlichen Unterarten des Feuersalamanders (R. Klewen (1991)) Momentan sind 13 Unterarten des Feuersalamanders bekannt, die sich teilweise aufgrund ihrer Färbung unterscheiden. Eine bebilderte Beschreibung der einzelnen Unterarten findet sich bei R. Klewen (1991). Das Farbkleid aller Arten enthält sowohl schwarze (olivebrauneschwarze) und gelbe (selten orange-rote) Flächen. Einzelexemplare können auch nur schwarz oder gelb gefärbt sein. Allgemein besitzen alle Unterarten je einen gelben Fleck am Ursprung der Extremitäten und auch über den Augen. Zudem sind auf den Paratoiden gelbe Muster vorhanden (Abbildung 34). 3. Farbsehen des Feuersalamanders Feuersalamander haben wie wir Menschen ein trichromatisches Farbsehen mit den Absorptionsmaxima der Zapfen bei ca. 450, 500 und 570nm (C.Przyrembel et al. (1995)). 4. Experiment ( Störungsverhalten) 4.1 Einleitung Um einen Beitrag zur Beantwortung der Funktion der Färbung des Feuersalamanders zu leisten, wird das Störungsverhalten des Feuersalamanders untersucht. Dieses müsste sich insbesondere in bezug auf eine Warnfärbung (Warnverhalten) oder eine Tarnfärbung (Flucht oder Bewegungslosigkeit) unterscheiden. Seite 50 Patrik Matter 4.2 Material und Methoden Die Daten wurden auf dem steilen Fussweg, der von Rümlingen (CH-4444) nach Wittinsburg führt, erhoben. Anfangs wurde einige Male erfolglos versucht, die Tiere tagsüber bei Niederschlag und nachts bei Trockenheit aufzusuchen. Aufgrund dieser negativen Erfahrungen wurden die restlichen Daten ausschliesslich in niederschlagsreichen Nächten (Dämmerung, vor Mitternacht, nach Mitternacht, Morgen) gesammelt. Da es leider nicht gelang ein Nachtsichtgerät aufzutreiben, blieb nichts anderes übrig, als die Reaktion der Tiere mittels künstlichem Licht zu beobachten. Das Störungsverhalten wurde untersucht, indem die Tiere mit einem kleinen Ast (Länge ca. 1m) leicht angetippt wurden. Dabei wurde keine Reaktion festgestellt. Das Antippen wurde also wiederholt, bis eine Reaktion zu erkennen war. Meistens ergab sich eine sichtbare Reaktion nach 1-3-maligem Antippen. 4.3 Resultate 71.1% der 45 Feuersalamander zeigten bei Störung ein Fluchtverhalten, 28.9% verharrten einen Augenblick ruhig. Die Extremwerte der einzelnen Nächte mit 66.6% - 75% weichen nur wenig von den totalen Werten ab. In diese Aufzählung sind auch diverse Zwischenformen eingeflossen, die nach der ersten erkennbaren Reaktion gewertet wurden. - Verharrende Feuersalamander, die nach weiterem Antippen trotzdem flüchteten = Unbeweglichkeit - Flüchtende Feuersalamander, die nach einem Meter ruhig verharrten = Flucht - Einzelne Feuersalamander, die bei einem versehentlich zweimaligen Versuch das zweite Mal ein anderes Verhalten zeigten (wobei beide Arten des Verhaltenswechsels vorkamen) = wurden nicht nochmals gewertet. 4.4 Diskussion Die vielen Zwischenverhalten legen nahe, dass es sich bei dem beobachteten Verhalten (Flucht/ Unbeweglichkeit) um zwei Aspekte eines komplexen Fluchtverhaltens handelt, wobei Phasen der Unbeweglichkeit sich mit schnellen Fluchtphasen abwechseln. Einerseits erhöht die Unbeweglichkeit die Chance, dass ein Feind den mit einem gestaltauflösenden Muster gefärbten Feuersalamander nicht von der Umgebung unterscheiden kann. Andererseits entkommt er so Raubfeinden, die nur bewegende Beute wahrnehmen (Greifvögel). Im weiteren erhöht Unbeweglichkeit die Chance, dass der Räuber die Ungeniessbarkeit seiner Beute erkennt, da sie ihm eine vorsichtigere Attacke ermöglicht. Seite 51 Patrik Matter Ein unbewegliches Beutetier erlaubt dem Räuber eine weniger kraftvolle Attacke, als wenn er ein flüchtendes Beutetier zu Fall bringen muss. Dank seiner grossen Zähigkeit und seinem guten Regenerationsvermögen hat der Feuersalamander gute Chancen eine Attacke, die aufgrund seiner Unbeweglichkeit nicht so kraftvoll ist, mit kleinen Verletzungen zu überstehen. Falls die Unbeweglichkeit von einem Feind durchschaut wird, bleibt als einzige Möglichkeit die rasche Flucht. (Alle unbeweglichen Feuersalamander gingen nach zahlreichem Antippen in eine schnelle Flucht über, um nach kurzer Zeit erneut zu erstarren oder sich ins Unterholz zu verkriechen.) Die Kombination zwischen Flucht und Unbeweglichkeit ist daher sehr gut geeignet, um einem breiten Spektrum von Feinden zu entkommen. Es fehlten alle Anzeichen auf ein spezifisches Warnverhalten. Die Tiere verharrten in Unbeweglichkeit (in der Position, in der sie gestört wurden) und nahmen keine besondere Warnstellung ein. 4.4.1 Mögliche Fehlerquellen Da Kunstlicht keine natürliche Umweltbedingung des Feuersalamanders ist und einige Exemplare eine deutliche Lichtflucht zeigten, ist es nicht auszuschliessen, dass die hier vorgestellten Resultate nur bedingt als natürliche Verhaltensweisen angesehen werden dürfen. Auch ist es möglich, dass die Feuersalamander den Experimentator frühzeitig wahrgenommen haben, und dass das beschriebene Störverhalten ein sekundäres Verhalten ist. Bei den acht Beobachtungen in der Morgendämmerung ergriffen die Tiere nämlich teilweise schon die Flucht, wenn sich der Experimentator auf 5 m näherte. Allerdings zeigte sich auch bei ihnen nach dem Antippen das ganze Verhaltensmuster. Es ist auch denkbar, dass der Feuersalamander zwischen unterschiedlichen Feindkategorien unterscheidet und je nach Kategorie ein unterschiedliches Verhalten zeigt. Somit könnte das zweiphasige Verhalten möglicherweise auf eine Irritation in diesem Mechanismus zurückzuführen sein. ( Antippen = Vogel => Unbeweglichkeit) / ( mein Körper = Grosser Feind => Flucht). Als Reaktion gegenüber einer weiteren nicht simulierten Feindklasse käme dann auch ein Warnverhalten in Frage, welches in dem Experiment nicht zu beobachten war. Es könnte auch möglich sein, dass die experimentelle Situation (Antippen durch einen Stock) für den Feuerssalamander eine so unnatürliche Situation ist, dass er auch eine künstliche (unnatürliche) Reaktion darauf zeigt. Der Vollständigkeit halber soll die sehr unwahrscheinliche Möglichkeit erwähnt sein, dass Feuersalamander nämlich ein genügend hohes Lernvermögen besitzen und gelernt haben könnten, dass ihr ursprüngliches Warnverhalten gegen Menschen nichts nützt. Eine weitere mögliche Fehlerquelle ist, dass als Verhalten bei Störung die erste sichtbare Verhaltensänderung erfasst wurde. Es wäre daher möglich, dass eine erste Berührung von den Feuersalamandern bereits als Angriff wahrgenommen wurde, auf den aber aufgrund des Giftschutzes noch keine Reaktion erfolgte (=> aposematisches Verhalten). Ein zweiter Angriff bedeutet jedoch, dass ein Feind vom Gift nicht abgehalten wird. Folglich ist Flucht bzw. Unbeweglichkeit die einzige Chance. Da jedoch bei einigen Exemplaren schon beim Seite 52 Patrik Matter ersten Antippen eine Reaktion ausgelöst wurde, ist die Erklärung, dass ein erstes Antippen als Zufallsberührung wahrgenommen wird, wahrscheinlicher. Als letzten wichtigen Aspekt muss noch verdeutlicht werden, dass aufgrund der nächtlichen Experimente nur bedingt auf das Verhalten der Feuersalamander bei Tageslicht geschlossen werden darf. 5. Experiment (Giftspritzen) 5.1 Einleitung Als ein mögliches Warnverhalten wäre das nur selten beobachtete aktive Giftspritzen des Feuersalamanders denkbar. Daher wurde in einem zweiten Experiment versucht, das aktive Giftspritzen als Verteidigung gegen Angreifer, welches von Brodie (1990) unter Laborbedingungen erzeugt wurde, auch unter natürlichen Bedingungen zu verursachen. 5.2 Material und Methoden Die Feuersalamander wurden mit unterschiedlichen Gegenständen (Grashalme, spitze Gegenstände, Wattenstäbchen, etc) unterschiedlich stark angetippt. 5.3 Resultate Trotz über 50 Versuchen wurde bei keinem der Tiere ein aktives Giftspritzen beobachtet. 5.4 Diskussion Da das Giftspritzen in freier Natur nur selten beobachtet wurde. Von B. Tiesmeier wurde (1992) die Vermutung geäussert, dass seine Bedeutung vielleicht vor allem in innerartlichen Auseinandersetzungen eingesetzt wird. Demzufolge ist es möglich, dass Feuersalamander unter natürlichen Bedingungen kein solches Verhalten zeigen und daher auch kein solches Verhalten beobachtet werden konnte. Allerdings sprechen mündliche Mitteilungen von Dr. Meeps eher gegen diese Theorie. Dieser streicht frei lebenden Feuersalamander mit Wattenstäbchen über die Parotiden, um so Salamandergiftproben sammeln zu können. Es wäre daher auch denkbar, dass es lokale (genetisch bedingte?) Unterschiede im GiftspritzVermögen der Feuersalamander gibt. So könnte eine solche Spezialisierung bei geringem Feinddruck zurückgebildet werden. 5.4.1 Mögliche Fehlerquellen Die wahrscheinlichste Fehlerquelle ist, dass aus Rücksicht die Tiere zu wenig fest angetippt worden sind, sodass diese sich nur bedingt bedroht gefühlt haben und keine Warnreaktion gezeigt haben. Im Zeitraum, in dem die Salamander untersucht wurden, war es mehrheitlich sehr trocken (Waldbrandgefahr in der ganzen Schweiz). Da Salamanderhaut nur einen schlechten Vertrocknungsschutz bietet (L.Jecklin (1935)), sind die spärlichen Gewitter für die Seite 53 Patrik Matter Salamander überlebenswichtig. Es ist daher denkbar, dass sie einen Warnmechanismus, der so viel lebenswichtige Flüssigkeit verbraucht, nur dann einsetzen, wenn kein Wassermangel herrscht. Eine weitere Fehlerquelle könnte sein, dass das Experiment (Giftspritzen) meistens mit den schon gestesteten Exemplaren des Experimentes (Störungsverhalten) durchgeführt wurde. Allerdings zeigten auch die 10 Exemplare, die nur auf Giftspritzen getestet wurden, keine andere Reaktion. Da beide Experimente einen Angriff darstellen sollten, resultiert aus dem doppelten Testen der Salamander auch keine Verfälschung. Ein längerer Angriff sollte mit erhöhter Wahrscheinlichkeit ein Warnverhalten auslösen. Es wäre auch denkbar gewesen, schon beim ersten Experiment ein Giftspritzen zu beobachten, was jedoch nie der Fall war. Es kann nicht ausgeschlossen werden, dass die experimentelle Situation (Antippen durch einen Gegenstand bei Kunstlicht) für den Feuersalamander eine so unnatürliche Situation ist, dass er eine künstliche (unnatürliche) Reaktion darauf zeigt, also kein Gift spritzt. Seite 54 Patrik Matter VII. ALPENSALAMANDER 1. Lebensweise Abbildung 35: Nominalform (Salamandra atra atra) und die gelbgefleckte Unterart (S. atra. Aurora) 1.1 Lebenszyklus (vgl. K.Grossenbacher & R. Günther (1996)) Nach 2-3 Jahren Entwicklungszeit werden Alpensalamander als vollständig entwickelte Jungtiere geboren. Sie gehen danach hauptsächlich in den frühen Morgenstunden bei hoher Luftfeuchtigkeit auf Jagd und verstecken sich ansonsten in Tagverstecken. Im Winter ziehen sie sich etwas tiefer in den Boden zurück. Nach 2-4 Jahren werden die Jungen geschlechtsreif und erreichen ein verbürgtes Alter von 15 Jahren (Fachbach (1988)). Sie können aber wahrscheinlich, ähnlich wie der Feuersalamander, weitaus älter werden. Die grosse Streuung der Entwicklungsdaten liegt an der unterschiedlichen Länge der genügend warmen Wetterbedingungen (4-6 Monate) in den unterschiedlichen Höhenlagen des Verbreitungsgebietes. 1.2 Lebensraum (vgl. K.Grossenbacher & R.Günther (1996)) Der Alpensalamander ist ein typisches Tier der montanen Stufe. Er kommt in allen Lebensräumen vor, die genügend feucht sind und sichere Tagverstecke bieten können. Da reine Nadelwälder zu trocken sind, kommt der Alpensalamander dort nicht vor. Ebenso meidet er unstabile Geröllhalden, da die Gefahr besteht, von abstürzendem Geröll zerquetscht zu werden. 1.3 Tagesrhythmus (vgl. R. Klewen ( 1991)) Der Tagesrhythmus des Alpensalamanders wird in erster Linie von den Lichtverhältnissen, aber auch durch die relative Luftfeuchtigkeit bestimmt. Erst unter 4°C wird die Temperatur zum limitierenden Faktor. Bei hinreichender Luftfeuchtigkeit (85% relative Luftfeuchtigkeit) verlassen die Salamander in der Nacht ihre Verstecke und suchen sie in der Morgendämmerung wieder auf. Dabei ist in den frühen Morgenstunden zwischen 4 und 8 Uhr ein Aktivitätsmaximum beobachten („Morgentau“). Der Alpensalamander kann daher als nachtaktive Art mit Tendenz zu morgendlichem Aktivitätsmaximum bezeichnet werden. Seite 55 Patrik Matter Bei heftigen Gewittern nach langer Trockenheit ist der Alpensalamander auch tagsüber in grösserer Zahl ausserhalb der Verstecke anzutreffen (K. Grossenbacher & R.Günther (1996)). 1.4 Jahresrhythmus (vgl. K. Grossenbacher & R.Günther (1996)). Alpensalamander werden einige Wochen nach der Schneeschmelze aktiv (Mai-Juni). Die klimatischen Bedingungen zwingen die wechselwarmen Tiere, sich in der Regel im September in ihre Winterverstecke zurückzuziehen. Alpensalamander sind deshalb nur 4-5 Monate aktiv. Die restlichen 7-8 Monate verbringen sie ohne Nahrungsaufnahme in ihren frostsicheren Winterverstecken im Boden. 1.5 Paarung Die Paarung findet während der ganzen Aktivitätsperiode statt, wobei im Sinne von Lorenz (1978) die Silhouette eines vorbeilaufenden Weibchens als Auslöser dient. Ähnliche Formen wie männliche Alpensalamander, Feuersalamander oder Attrappen können ebenfalls als Auslöser fungieren. Die fünf Phasen der Paarung werden bei Klewen (1991) sehr detailliert beschrieben: 1. Verfolgung 2. Aufsteigen und Kopfreiben 3. Unterkriechen, Klammern und Kehlreiben 4. Schwanzwurzelreiben und Absetzen der Spermatophore 5. Aufnahme der Spermatophore durch das Weibchen 1.6. Nahrung des Alpensalamanders Wie alle Schwanzlurche ist auch der Alpensalamander ein Nahrungsgeneralist. Er frisst jegliche Beute, die er aufgrund seiner Grösse überwältigen kann. Somit fallen Regenwürmer, Schnecken, Spinnentiere und diverse Insekten in sein Beutespektrum (R. Kleven (1991)). 1.7 Feinde des Alpensalamanders Über natürliche Feinde des Alpensalamanders gibt es nur wenig Berichte und seltene Beobachtungen (Elstern, Alpendohlen, Kreuzottern, Ringelnattern, Blindschleichen) (K. Grossenbacher & R.Günther (1996), E.Sochurek (1956)). Als weitere potentielle Feinde kämen Raubtiere (Fuchs, Wolf, Marder etc.), Eulen und Greifvögel in Frage. Seite 56 Patrik Matter 2. Färbung der drei Unterarten Salamandra.atra.atra und S.a.prenjensis, sind beide ausschliesslich schwarz (braun) gefärbt. S.a.aurora besitzt einen Gelbanteil von 50% oder mehr, wobei er in erstaunlicher Übereinstimmung mit dem Feuersalamander je einen gelben Fleck am Ursprung der Extremitäten besitzt. Oft sind auch Flecken auf Augen und Paratoiden vorhanden. Am Monte Pasubio wurden weitere Tiere mit einem Gelbanteil bis zu etwa 20% entdeckt. Besitzen diese Salamander gelbe Flecken, so sind diese immer auch auf dem Ursprung der Extremitäten zu finden, allerdings nicht immer auf allen vier Extremitäten. Es wäre denkbar, dass sie schon bald als weitere Unterart beschrieben werden (Grossenbacher, mündliche Mitteilung). 3. Farbsehen des Alpensalamanders Leider liegen noch keine detaillierten Untersuchungen über das Farbsehen des Alpensalamanders vor. Es ist aber wahrscheinlich, dass er, ebenso wie der Feuersalamander, ein trichromatisches Sehvermögen besitzt. Seite 57 Patrik Matter VIII. DISKUSSION 1. Färbung 1.1.1 Die Färbung des Feuersalamanders – ein Tarnfärbung? Obwohl die Feuersalamander tagsüber als sehr auffällig erscheinen, kommt es nach A. Thayer (1897) für eine gestaltauflösende Färbung hauptsächlich auf das Farbmuster und nur bedingt auf den Farbton an. Das kontrastreiche Muster des Feuersalamanders ist geradezu ein Paradebeispiel für eine gestaltauflösende Färbung. Diese Wirkung wird in der Nacht noch verstärkt, weil nun der schwarze Grundton mit dem Dunkel des Waldes verschmilzt und so nur noch ein nicht zusammengehörendes gelbes Muster übrigbleibt. Zusätzlich wird eine tarnende Färbung bei nachtaktiven Tieren noch dadurch unterstützt, dass im Dunklen kein grosses Auflösungsvermögen erreicht werden kann (vgl. V 2.1.1.1). Insbesondere die Flecken an den Extremitätenwurzeln sorgen dafür, dass der Umriss von Körper und Beinen getrennt wird und der Feuersalamander von vielen potentiellen Feinden nicht erkannt wird. Auch die Augen als verräterische Körperteile werden durch das gelbe Muster hervorragend verborgen. Schnelle Bewegung löst die Wirkung der besten Tarnung auf. Dies könnte mit ein Grund sein, weshalb sich Feuersalamander so gemächlich bewegen. Auch passt diese Hypothese sehr gut zu den Resultaten der Experimente, dem zweiphasigen Verhalten. Dieses stellt gleichsam einen Kompromiss zwischen Tarnung durch Gestaltauflösung und Flucht dar. Die Funktion als Tarnfärbung scheint daher eine sehr gute Erklärung für die Färbung des Feuersalamanders zu sein, auch liefert sie als einzige eine Erklärung für die Flecken auf den Ursprüngen der Extremitäten. 1.1.2 Die Färbung des Feuersalamanders – eine Schreckfärbung? Da die Färbung des Feuersalamanders immer sichtbar ist, kommt eine Funktion als Schreckfärbung nicht in Frage. 1.1.3 Die Färbung des Feuersalamanders – eine Warnfärbung? 1.1.3.1 Abklärung, der für Tiere mit Warnfärbung typischen Eigenschaften (vgl. III.3). - spezielle Färbung Das gelbe Muster auf schwarzem Hintergrund ist eine typische Form der aposematischen Färbung. Speziell ist hierbei zu betonen, dass die Parotiden (Giftdrüsen am Kopf) bei allen Unterarten zumindest teilweise gelb gefärbt sind. Seite 58 Patrik Matter - Schutzvorrichtung Die Schutzvorrichtung des Feuersalamanders ist seine aktive Giftigkeit. Tiere bis zur Grösse eines Hundes können nach dem Verzehr eines Feuersalamanders durch dessen in seinen Hautdrüsen vorhandenes Gift sterben. Des Weiteren hat der Feuersalamander die Fähigkeit sein Gift über einen Meter weit zu spritzen. - Warnverhalten In den Störexperimentes zeigen die Feuersalamander kein eigentliches Warnverhalten. Allenfalls könnte die Unbeweglichkeit als eine bedingt aposematische Reaktion betrachtet werden. Allerdings findet man in der Literatur Hinweise auf ein Warnverhalten der Feuersalamander. Brodie (1977) beschreibt in seiner umfassenden Übersicht über das Störverhalten der Salamander ein unter Laborbedingungen beobachtetes Warnverhalten der Feuersalamander: „ Wenn sie am Kopf oder an den Vorderbeinen berührt werden, stellen die Salamander ihre Vorderbeine auf und senken den Kopf, so zeigen die Parotiden in die Richtung des Stimulus. Werden die Hinterbeine berührt, folgt entweder eine Flucht oder der Kopf wird gegen den Stimulus ausgerichtet.“ Dieses Verhalten konnten jedoch nicht durch neuere Beobachtungen bestätigt werden ( M.Horter, H. Greven (1981). Auch die Bemerkung:“adulte Salamander senken den Kopf und präsentieren dem Angreifer ihre giftreichen Parotiden“ von (B.Tiesmeier, R.Günther (1996)) geht auf Brodie (1977) zurück, kann also nicht als schlüssige Bestätigung dieses Sachverhaltes dienen. Auch das Giftspritzen könnte ein Warnverhalten sein. Die wenigen Beobachtungen in freier Natur legen jedoch nahe, dass das Giftspritzen keine normale Abwehrreaktion darstellt. Möglicherweise wird es vor allem in innerartlichen Auseinandersetzungen gebraucht und stellt kein Warnverhalten dar (B.Tiesmeier(1992)). Unter natürlichen Bedingungen ist also bisher kein Warnverhalten beschrieben worden und auch das seltene Giftspritzen stellt nur bedingt ein Solches dar. - Träges Verhalten Feuersalamander zeichnen sich durch ein träges Verhalten aus und erbeuten eher einfach zu fangende Beute wie Schnecken und Würmer. Befinden sie sich jedoch in Gefahr, können sie sich mit erstaunlicher Geschwindigkeit bewegen. - Zähigkeit Salamander sind wie die meisten Amphibien sehr robust gebaut, und können verlorene Gliedmassen durch Regeneration mit der Zeit ersetzen. - Auftreten in grösseren Gruppen Obwohl der Feuersalamander zuweilen in grossen Gruppen auftritt (Eiablage der Weibchen, Verlassen des Gewässers der juvenilen Salamander, Paarungszeit, Regenschauer nach langer Trockenheit) sind solche Beobachtungen eher die Ausnahme als die Regel. Allerdings wäre es denkbar, dass gerade in Zeiten der Eiablage und der Paarung eine Warnfärbung einen grossen Vorteil bringen könnte. Ob sie jedoch einen grossen selektiven Vorteil bildet, bleibt fraglich. Seite 59 Patrik Matter - Aktivität und Jahreszyklus Wenn Feuersalamander trockener Luft ausgesetzt werden, verlieren sie sehr viel Feuchtigkeit. Dies ist auch ein Grund für die nachtaktive Lebensweise des Feuersalamanders (und für die Inaktivität während des Winters. Ist der Winter im Gebirge nicht auch feucht???) 1.1.3.2 Vorhandene typische Eigenschaften Von den typischen Eigenschaften aposematischer Tiere besitzen die Feuersalamander die Giftigkeit, eine aposematisches Musterung, ein träges Verhalten und einen robusten Körperbau. Sie treten jedoch nur zuweilen in Gruppen auf und zeigen kein deutliches Warnverhalten in freier Natur. Obwohl ein deutliches Warnverhalten fehlt, ist es Aufgrund so viel passender Merkmale gut möglich, dass die Färbung des Feuersalamanders eine warnende Funktion hat. Allerdings scheint es merkwürdig, dass sich dann nicht ein den Schutzeffekt verstärkendes deutlicheres Warnverhalten herausgebildet hat, zumal nahe verwandte Arten ein solches Verhalten zeigen (E.D Brodie (1977)). 1.1.3.3. Voraussetzungen für eine Funktion als Warnfärbung Um das zweifarbige Muster zu erkennen, sind nur zwei unterschiedliche Zapfentypen nötig. Deshalb können alle potentiellen Feinde das auffällige Muster wahrnehmen. Feuersalamander entfalten ihre Aktivität, sobald in der Nacht genügend Feuchtigkeit herrscht. Tagsüber kommen sie nur selten bei Regenschauer nach langer Trockenheit zum Vorschein. Da jedoch in der Nacht die Augen der Wirbeltiere keine Farben mehr zu erkennen vermögen und das Nachtsichtsystem der Stäbchen nur verschiedenen Helligkeiten unterscheiden kann, sind Feuersalamander nachts - wenn überhaupt - nur als heller Streifen sichtbar. Daher müsste ein nächtlicher Jäger zuerst die unscheinbaren Streifen als Tier mit Warnfärbung erkennen, damit der Feuersalamander geschützt werden könnte. Wahrscheinlicher ist, dass der Feuersalamander gar nicht erst entdeckt wird, daher könnte die Warnfärbung also nur in seltenen Situationen zum Einsatz kommen. 1.1.4. Die Färbung des Feuersalamanders – eine Drohfärbung? Als betonende Zeichnung im Sinne einer Drohfärbung kämen am ehesten die gelben Flecken am Ansatz der Gliedmassen in Frage, da sie ein Indiz für die Grösse sein könnten. Nach Kästle(1986) ist aber beim Rivalenkampf hauptsächlich der Tastsinn beteiligt, optische Kanäle scheinen, wenn überhaupt, nur eine untergeordnete Rolle zu spielen. Auch die Funktion der Zeichnung als Statussymbol ist eher unwahrscheinlich, da kein Unterschied zwischen dem Muster der Weibchen und dem der Männchen existiert. Aufgrund dieser Überlegungen ist eine Funktion als Drohfärbung nicht anzunehmen. 1.1.5 Die Färbung des Feuersalamanders – eine Balzfärbung Auch bei der Paarung scheint die Färbung eine geringe Rolle zu spielen, da Feuersalamandermännchen allem folgen, was der Silhouette eines Weibchens entfernt ähnelt. Sobald die eigentliche Paarung beginnt, ist auch hier der Tastsinn entscheidend. Seite 60 Patrik Matter Da das Muster des Feuersalamanders nur selten symmetrisch ist, spielt auch Symmetrie als “Kriterium für gutes genetisches Material“ keine Rolle. Kranke Amphibien haben oft eine weniger glänzende Haut als Gesunde, Aus der Gelbfärbung ein Statussymbol für Gesundheit konstruieren zu wollen, scheint jedoch sehr gewagt. Sensorische und „mentale“ Fenster und Fisher’s „Hypothese über die sexuelle Selektion“ überzeugen als mögliche Erklärung ebenfalls nicht. Eine Funktion als Balzfärbung ist deshalb eher unwahrscheinlich. 1.1.6. Die Färbung des Feuersalamanders – eine Episematische Färbung? Da Männchen sich sehr unwählerisch auf Weibchen bzw. den Weibchen ähnliche Objekte stürzen um sich mit ihnen zu paaren ist eine episematische Funktion der Färbung praktisch auszuschliessen. 1.1.7. Die Färbung des Feuersalamanders – als Färbung mit physikalischer Funktion ? Aufgrund des wirr verteilten gelben Muster des Feuersalamanders ist eine physikalische Eigenschaft unwahrscheinlich. Es wäre jedoch denkbar, dass die gelben Flächen über den Augen eine physikalische Schutzfunktion haben könnten. Auch könnte es sein, dass ein Schwarz-weisser Körper ein anderes Temperatur-Optimum besitzt als ein rein schwarzer. Allerdings könnten beide Aspekte nicht die Erklärung sein, für die beiden gefleckten Unterarten des Alpensalamanders, da längst nicht alle oberhalb der Augen gelb sind und sie einen klimatisch sehr ähnlichen Lebensraum haben, wie ihre ungefleckten Verwandten. Physikalische Eigenschaften liefern also keine brauchbare Erklärung für die Färbung des Feuersalamanders. 1.1.8 Die Färbung des Feuersalamanders – ohne selektive Funktion? Da es einige wahrscheinliche Erklärungen für die Färbung des Feuersalamanders gibt, ist es unwahrscheinlich, dass die Färbung des Feuersalamanders zufällig oder das Resultat pleiotropischer Wechselwirkungen ist. 1.2 Zusammenfassung Selbstverständlich erlauben diese Überlegungen, ebenso wie die Resultate aus den Experimenten, keine definitive Stellungnahme zur Bedeutung der Färbung des Feuersalamanders. Dennoch scheint eine Funktion als gestaltauflösende Färbung am plausibelsten, da Färbung und Verhalten des Feuersalamanders alle Kriterien für eine Tarnfärbung erfüllen. Obwohl ein typisches Warnverhalten fehlt, erfüllen Färbung und Verhalten die restlichen Kriterien einer Warnfärbung und daher ist es wahrscheinlich, dass die Färbung des Feuersalamanders unter gewissen Umständen (Licht) auch als Warnfärbung funktionieren kann. Die anderen möglichen Funktionen sind beim Feuersalamander eher unwahrscheinlich. Seite 61 Patrik Matter Es wäre natürlich auch möglich, dass erst die synergetische Wirkung beider Funktionen einen genügend starken selektiven Vorteil bietet. . 1.3 Unterschiedlich gefärbte Unterarten des Feuersalamanders Die Unterarten des Feuersalamanders weisen eine grosse Variabilität in ihrer Färbung auf. Allerdings kommen keine ganz schwarzen Unterarten vor. Dies weisst darauf hin, dass die schwarz-gelbe Färbung einen selektiven Vorteil für Feuersalamander bietet oder zumindest mal geboten hat. Falls die unterschiedlichen Färbungen der Unterarten also nicht auf eine Anpassung an ihren Lebensraum zurückzuführen sind, scheint es, dass für die Funktion die Färbung und nicht das Muster entscheidend ist (war). 1.4 Evolution von Warnfärbung via Tarnfärbung? Die Analyse der Färbung hat also gezeigt, dass eine Färbung sowohl als Tarn- wie auch als Warnfärbung funktionieren kann. Daher wäre es auch denkbar, dass sich im Verlaufe der Evolution eine Tarnfärbung in eine Warnfärbung umwandeln kann, oder zumindest Synergien mit einer Warnfärbung die Evolution einer Tarnfärbung erleichtern könnten. 2. Mögliche weiterführende klärende Experimente und Fragestellungen 2.1 Tarnung? Die Funktion der Tarnung ist die Entdeckung durch einen Raubfeinden zu verhindern. Um eine tarnende Wirkung nachzuweisen müsste man daher Experimente machen, in denen gezeigt wird, dass gelbgefleckte Feuersalamander weniger häufig von Feinden erbeutet wird, als ganzheitlich schwarzgefärbte Zuchtformen. Ähnliche Vergleiche könnten auch zwischen Alpensalamandern und seinen gefleckten Unterarten gemacht werden. Allerdings können solche Experimente nur funktionieren, wenn die Feinde Immun gegen das Gift der Salamander sind, da ansonsten eine aposematische Wirkung nicht ausgeschlossen werden kann. Auch ist es wichtig die Experiment unter verschiedensten Bedingungen (Umgebung, Lichtmenge, Feuchtigkeit, etc.) durchzuführen. Eine andere Möglichkeit, wäre die Experimente so durchzuführen, dass potentielle Feinde so trainiert werden, dass sie eine Belohnung bekommen, wenn sie einen Feuersalamander entdecken. Auch hier würde wieder ein Vergleich der Entdeckungsraten der versch. Unterarten unter versch. Bedingungen durchgeführt. Der Vorteil dieses Experimentes wäre, dass die aposematische Wirkung der Färbung ausgeschlossen wird, da ja die Entdeckung und nicht die Erbeutung gewertet wird. Besonders interessant wäre ein Vergleich zwischen der Erbeutungsrate des ersten Experimentes verglichen mit der Entdeckungsraste des zweiten Experimentes. Da bei gut trainierten Tieren falls es zu grossen Unterschieden kommt, diese nur durch eine aposematische Wirkung der Färbung bezogen auf diese Tierart deuten kann. Seite 62 Patrik Matter 2.2 Warnung? Laborversuche könnten einen Hinweis geben, welche der potentiellen Feinde Feuersalamander erbeuten und ob sich diese teilweise von dessen Warnverhalten (falls vorhanden) beeindrucken lassen? Auch wären Laborversuche interessant, bei denen der Schutz von ganz schwarzgefärbten Zuchten mit dem des Wildtypen verglichen wird. Besonders interessant wäre ein Vergleich zwischen der Erbeutungsrate des ersten Tarnexperimentes verglichen mit der Entdeckungsraste des zweiten Tarnexperimentes. Da bei gut trainierten, hungrigen Tieren falls es zu grossen Unterschieden kommt, diese nur durch eine aposematische Wirkung der Färbung bezogen auf diese Tierart deuten kann. Auch wäre ein Versuchsansatz denkbar in dem die beiden Salamanderzuchten auf einem für gelb und schwarz gleich kontrastreichen Hintergrund, verschiedenen hungrigen potentiellen Feinden präsentiert und ermittelt wird, ob sie unterschiedlich oft erbeutet werden, sowie ob es einen unterschied zwischen erfahrenen und unerfahrenen Feinden gibt. Um die Frage, ob die Färbung des Feuersalamanders eine warnende Funktion hat endgültig zu klären, wäre es wichtig, nächtliche infrarot Beobachtungen vom Verhalten bei Begegnungen mit potentiellen Feinden analysieren zu können. 3. Vergleich mit dem Alpensalamander (hypothetische Überlegungen) 3.1 Färbung 3.1.1 Gefleckte Unterarten Die beiden gefleckten Unterarten des Alpensalamanders haben ebenfalls meistens einen gelben Fleck, der die Extremitäten optisch vom Körper abtrennt, sowie Flecken am Kopf. Da die Färbung mit dem Feuersalamander vergleichbar ist, kommt auch hier eine tarnende Wirkung der Färbung am ehesten in Frage, allerdings wäre auch eine warnende Funktion möglich. Experimente und Fragestellungen: Vgl. 2 3.1.2 Ausschliesslich schwarz gefärbte Unterarten Die andere Unterart und der gemeine Alpensalamander sind hingegen ausschliesslich schwarz gefärbt. Was wären mögliche Gründe dafür, dass diese Unterarten eine schwarze Färbung besitzen und keine Warnfärbung/ Tarnfärbung zu besitzen? Da auch Alpensalamander hauptsächlich nachtaktiv sind, scheint auf den ersten Blick eine Funktion der schwarzen Haut als Strahlungsschutz eher unwahrscheinlich zu sein. Allerdings zeigen Alpensalamander ein morgentliches Aktivitätsmaximum und sind auch häufig bei Regenfällen nach langer Trockenheit aktiv, wodurch es vorkommen kann, dass sie direktem Sonnelicht ausgesetzt werden. Daher wäre es denkbar, dass die schwarze Färbung eine Seite 63 Patrik Matter Strahlungsanpassung ist, zumal die schädliche UV-Strahlung im Gebirge deutlich höher ist, als in den Niederungen. Experimente und Fragestellungen: - Überprüfung wie viel Sonnenstrahlung es braucht, bis kurz- bzw. langfristige Fitnessverluste bei Alpensalamandern erkennbar sind. - Vergleich der Strahlungsempfindlichkeit zwischen Feuer-, gefleckten und ungefleckten Alpensalamandern Zweifarbige gefärbte Tiere wirken optisch kleiner als einfarbige. Es wäre denkbar, dass diese optische Verkleinerung ein zu grosser Nachteil bei der Paarung für die Tiere darstellt, da ja zumindest der Auslöser des Paarungsverhaltens ein optischer Reiz ist. Experimente und Fragestellungen: - Paarungserfolg bei verschieden grossen Individuen - Paarungserfolg zwischen gefleckten und ungefleckten Alpensalamandern - Paarungserfolg von künstlich gefärbten Alpensalamandern Eine andere Möglichkeit wäre auch, dass das gelbe Muster der Salamander in vegetationsarmen Lebensräumen, nicht mehr tarnend wirkt, sondern die Tiere sehr auffällig macht. Falls dem so wäre, müssten diese Lebensräume jedoch von den beiden gefleckten Unterarten gemieden werden. Experimente und Fragestellungen: Siehe: 2.1 Tarnung? Auffällig ist, dass deutlich mehr Daten über Feinde der Alpensalamander verfügbar sind, als über den weiterverbreiteten Feuersalamander. Es ist daher sehr gut möglich, dass die kleinere Grösse auch eine kleiner Giftmenge und somit einen schlechteren Schutz vor Feinden bedeutet. Falls immune Feinde existieren, könnte es einen sehr starken Selektionsdruck in Richtung einer unauffälligen Färbung geben. Wobei auch hier die gestaltauflösende Funktion der Färbung durch die vegetationsarme Umgebung aufgehoben werden müsste. Falls ein solcher Feind existieren würde, müsste es dann natürlich wieder einen Grund (fehlendes Feindvorkommen, höhere Giftigkeit) geben, wieso sich eine gelbe Färbung für die beiden gefleckten Unterarten trotzdem lohnt. Experimente und Fragestellungen: - Giftigkeit der verschiedenen Unterarten - immune Feinde suchen, deren Jagdrevier die erforderlichen Lücken zeigt Eine unwahrscheinliche Erklärung, wäre auch, dass die aposematische Wirkung der Färbung nur verbunden mit der Spritzfähigkeit der Feuersalamander genügen wirksam ist und dass Alpensalamander trotz des vergleichbaren Körperbaus dazu nicht befähigt sind. Weitere sehr unwahrscheinliche Erklärung wären, dass es beim Alpensalamander bisher einfach noch nicht zu den erforderlichen Mutationen gekommen ist, oder dass die Entwicklung der gelben Färbung viel Energie oder eine spezifische Nahrung erfordert, was Aufgrund des Lebensraums oder der speziellen Entwicklung für den Alpensalamander nicht möglich ist. Seite 64 Patrik Matter 4. Farbwirkung Obwohl im der Bereich der Erforschung der Wirkung von Farbe in unterschiedlichen Übermittlungsmedien bzw. Umgebungen noch in den Kinderschuhen steckt, gibt es auch hier noch einige wichtigen Ergänzungen bezogen auf den Feuersalamander. Signale die in der Dunkelheit wirken sollen sind sehr hell (weiss oder gelb), daher könnte die gelbe Signalfarbe des Feuersalamanders eine Anpassung an seine nächtliche Lebensweise sein. Luft die durch kleine Partikel (Aerosole) verschmutzt ist, oder die einen hohen Wasserdampfgehalt enthält (Nebel), wirken wie ein transparenter Filter auf die Farbigkeit der Umgebung. Unter solchen Bedingungen wird gelb zur hellsten und blau zur dunkelsten Färbung. Da Feuersalamander tagsüber nur aktiv sind, wenn eine sehr hohe Luftfeuchtigkeit herrscht, wäre es vielleicht möglich, dass die gelbe Signalfarbe eine Anpassung an diese Bedingungen darstellt. Der Lebensraum des Feuersalamanders sind vor allem vegetationsreiche Buchenwälder. In vegetationsreichen Lebensräumen gibt es zwei unterschiedliche Signal-Strategien: Ganzkörper Signale, so dass auch schon ein Teil des Körpers als Signal wirkt oder sehr kleine Signale, so dass meistens zumindest ein Teil des Signals sichtbar ist. Die wild verstreute Anordnung der gelben Flecken auf dem Körper des Feuersalamanders, könnte daher dazu dienen, dass immer zumindest ein kleiner Teil des unverkennbaren Musters sichtbar ist. Als letztes gilt noch zu bedenken, dass vegetationsreichen Lebensräumen eine Mischung aus Licht und Schatten darstellen, und daher Weiss und Schwarz aufgrund der Verwechslungsgefahr nur bedingt als Signalfarben in Frage kommen. Daher könnte die schwarze Färbung eine Anpassung an ein mit dem Schatten verschmelzen sein, während das gelbe Muster für die gestaltauflösende Wirkung sichtbar sein muss. Auch könnt mit einer ähnlichen Argumentation das gelbe Muster des Feuersalamanders dafür sorgen, dass die Warnfärbung sich von Licht und Schatten abhebt. 5. Farbsehen Auf dem Gebiet des Farbsehens wird bereits seit 50 Jahren intensiv geforscht, trotzdem ist man noch weit von einem Verständnis der Farbwahrnehmung entfernt. Obwohl heute die ersten zwei Schritte (Absorption durch die Zapfen, Verschaltung in der Netzhaut) der Farbnehmung zumindest beim Menschen grundsätzlich verstanden sind, findet der entscheidende Schritt im Hirn statt und es wird daher noch viele Jahre bis auch dieser Schritt erforscht ist. Noch schwieriger stellt sich das ganze bei Tieren dar. Da sich bei vielen Tieren bereits die Grundlagen des Farbsehsystems (Anzahl unterschiedliche Zapfentypen, Verhältnis StäbchenZapfen) von unserem Unterscheidet, kann man nur bedingt Aussagen über das Farbsehen von Tieren machen. Da jedoch Wirbeltiere minimal zwei Unterschiedliche Zapfentypen haben und in der Evolution nicht funktionale Anpassungen durch Selektion zurückgebildet werden darf man davon ausgehen, dass alle Wirbeltiere zumindest zwei unterschiedliche Farbqualitäten unterscheiden können. Daher können Wirbeltiere das zweifarbige Muster des Feuersalamanders wahrnehmen. Seite 65 Patrik Matter Der Feuersalamander scheint für uns nur zweifarbig gefärbt zu sein, um dies jedoch sicher sagen zu können müsste man ein Absorptionsspektrum erstellen. Erst wenn nur zwei Absorptionsmaxima festzustellen wären, wäre der Feuersalamander wirklich zweifarbig! Sollte man mehr Absorptionsmaxima feststellen, wäre es unmöglich vorherzusagen, wie die Färbung von Tieren mit unterschiedlichem Farbsehvermögen wahrgenommen wird! Aufgrund der unterschiedlichen Grundlagen des Farbsehens von Tieren ist es uns also nicht möglich vorherzusagen wie Tiere die Farbigkeit ihrer Welt wahrnehmen und ob dies ebenfalls in den uns bekannten Qualitäten (Farbton, Sättigung, Helligkeit) geschieht. Zusätzlich wird die Interpretation noch durch morphologische Anpassungen (z.B Öltröpfchen) an ihren Lebensraum noch weiter erschwert. Aufgrund dieser Schwierigkeiten wird oft aufgrund von Verhaltensexperimenten auf das Farbsehen von Tieren geschlossen. Solche Experimente finden jedoch immer in künstlicher Umgebung und unter künstlichen Bedingungen statt. Daher könnten erfolglose Experimente auch den Grund haben, dass die Versuchstiere ein ungenügendes Abstraktionsvermögen besitzen, um Farben als unabhängige Qualität zu erkennen. Grundsätzlich ist also immer Vorsicht geboten, wenn wir unsere menschliche Sichtweise von Dingen, mit denen von Tieren gleichsetzen. Dies gilt insbesondere für alle Fragestellungen, welche optische Signale betreffen, aber auch für alle anderen optischen Fragestellungen! 6. Evolution von Signalen Ein weiterer wichtiger Aspekt zur Evolution von Signalen könnte die synergetische Wechselwirkung zwischen unterschiedlichen Funktionen von Signalen sein. Am Feuersalamander kann man sehen, dass es zum Beispiel möglich ist, dass eine Färbung sowohl als Tarnung wirken, wie auch zur Warnung dienen kann. Seite 66 Patrik Matter ANHANG: IX. APOSEMATISCHE MECHANISMEN: (DROH- UND WARNMECHANISMEN) Aposematische Mechanismen dienen dazu, Rivalen oder Feinde abzuschrecken. Hingston (1933) schlägt vor, zwischenartliche Warn- und innerartliche Drohmechanismen zu unterscheiden. Warnmechanismen treten in „Räuber-Beute“-Beziehungen auf. Die Beute gibt durch ihre Warnung dem Raubfeind zu verstehen, dass ihr noch weitere Möglichkeiten (Gift, gute Fluchtchancen, etc) offen stehen. Drohmechanismen wirken in Konkurrenz-Situationen, wenn Individuen um eine Ressource (Revier, Weibchen, Futter, etc) kämpfen. Oft lassen solche Mechanismen in eindeutiger Weise – in physiologischer oder psychologischer Hinsicht - Rückschlüsse auf die ihnen zu Grunde liegende Qualität (Kraft, Ausdauer, Mut, Kampfstärke, etc) zu. Daher ist es nicht erstaunlich, dass der gleiche Mechanismus oftmals sowohl als Drohung, wie auch als Warnung verwendet wird. Drohung: Alle Einrichtungen und Verhaltensweisen, die dazu dienen den Artgenossen abzuweisen. Drohungen müssen verlässliche Informationen über Motivation und/oder Kampfstärke eines Gegners geben. Auch Reviermarkierungen sind Drohungen. Warnung: Alle Einrichtungen und Verhaltensweisen, die dazu dienen, einen artfremden Feind abzuwehren. Nach dieser Definition sind etwa Warnfärbungen, Prellsprünge der Thomsongazelle und das warnende Klappern einer Schlange Warnungen. Zwischen diesen unterschiedlichen Möglichkeiten des Warnens gibt es aber tiefgreifende Unterschiede. Während Warnfärbungen unabhängig von einer Konfrontation vorhandene Einrichtungen sind, tritt Warnverhalten (Warnlaute und Klappern) erst bei Feindkontakt auf. Auch ist aus energetischer Sicht eine Warnfärbung kostengünstiger als etwa Schlangengeklapper oder Prellsprünge. Warnfärbungen, sowie das Rasseln der Klapperschlange scheinen überdies eher vor einem (Notwehr-)Angriff zu wahren, während die Prellsprünge der Thomsongazelle vor einer möglichen Flucht warnen. Man kann und sollte deshalb auch permanente Warneinrichtungen von temporärem Warnverhalten unterscheiden. Permanente Warnungen (z.B Warnfärbung der Wespen) Permanente Warnfarben sind unabhängig von einer Konfrontation vorhanden. Sie verursachen vergleichsweise geringe Kosten und schrecken eine so grosse Zahl von Feinden ab, dass sich die grössere Auffälligkeit, welche damit verbunden ist lohnt. Solche Signale können nur vor der Gefahr für den Raubfeind warnen, die ein Angriff mit sich bringt. Seite 67 Patrik Matter Temporäre Warnungen (z.B Einrollen des Igels) Hier wird durch ein besonderes Verhalten, das erst bei einer Konfrontation mit einem Feind auftritt, gewarnt. Das Verhalten wird auf den jeweiligen Feind abgestimmt. Solche aktiven Signale erfordern Energie. Sie haben aber den Vorteil, dass sie nicht nur vor einer Gefahr für den Raubfeind warnen können, sondern beinhalten auch die Möglichkeit, sich vor einem Feind zu schützen. 1.Mechanismen, die auf den Sehsinn wirken Viele aposematische Mechanismen wirken auf den Sehsinn. Da permanente Warnungen möglichst kostengünstig sein sollten, bietet sich hier eine visuelle Signalisierung gerade zu an. Temporäre Warnungen wirken erst, wenn ein Gegner sie wahrnehmen kann und erkennt, dass sie sich auf ihn beziehen. Dies ist bei einem Blickkontakt automatisch der Fall. Deshalb sind temporäre visuelle Warnungen weit verbreitet. Drohungen finden ebenfalls meistens bei Sichtkontakt statt. 1.1. Färbung Optische Signale haben den Vorteil, dass ihre Aussendung im Gegensatz zu den meisten anderen Signalen keine Energie verbraucht. Deshalb sind sie ideal als permanente Warnung und als „Statussymbole“. Letztere haben die Eigenschaft ein bestimmtes Körpermerkmal zu betonen, das für eine Drohung wichtig ist (Bsp.: Die Augenringe vieler Vögel verdeutlichen die Blickrichtung). 1.1.1 Auffällige Dauerfärbung Oft sind Beutetiere - auf welche sich der Angriff aus einem guten Grund (Gift, Stachel, Ungeniessbarkeit, etc) nicht lohnt - auffallend gefärbt, damit sie rechtzeitig von den Raubfeinden entdeckt und gemieden werden können. Eine solche Färbung hat als Warnung die Aufgabe, Beutegreifer abzuschrecken. Da eine Dauerfärbung nicht den momentanen Zustand widerspiegelt, ist sie für eine Drohung nicht geeignet! Daher gibt es auch keine Beispiele für Drohen durch Dauerfärbung des Körpers. Allerdings können Körperfärbungen eine Drohung unterstützen (z.B Statussymbole). Beispiel für Warnen: (Pfeilgiftfrösche (Dendrobates spec.)) Die grellen Farben der Pfeilgiftfrösche der Familien Dendrobatidae, Epipedobates und Phyllobates sind wohl eines der bekanntesten Beispiele für die aposematische Wirkung von aufälligen Farben (Summers. K et al. 2001). 1.1.2 Physiologischer Farbwechsel Einige Tiere haben die Fähigkeit, ihre Färbung zu verändern. Eines der bekanntesten Beispiele ist das Chamäleon, aber auch viele Echsen und Tintenfische sowie einige Fische können ihre Färbung verändern. Dieser Farbwechsel hat als gestaltauflösende Anpassung an die Umgebung allerdings oft die Funktion der Tarnung. Seite 68 Patrik Matter Beispiel für Warnen: Blauringkraken (Hapalochlaena spec.) Die golfballgrossen Blauringkraken haben nur eine Armlänge von etwa 10 cm. Sie kommen im westlichen Pazifik zwischen der Australischen Küste und den Inseln Südostasiens vor. Der Name Blauringkraken kommt daher, dass diese Kraken, wenn sie sich bedroht fühlen, von ihrer normalen Tarnfärbung auf einen hellen Hintergrund wechseln, auf dem überall blaue Ringe erscheinen. Diese Farbänderung dient als Warnung vor ihrem tödlichen Biss. Der Speichel der Blauring-Kraken enthält Tetrodoxin – ein Gift bakteriellen Ursprungs, das in vielen marinen Organismen vorkommt. Dies ist ein schnell wirkendes Nervengift, das die Acetylcholinrezeptoren der postsynaptischen Membranen hemmt. Das Opfer wird gelähmt und kann nicht flüchten. Bei den kleinen Krebstieren führt das Gift des Blauring-Kraken innerhalb weniger Sekunden zum Tod, beim Menschen dauert es einige Stunden, bis der Tod durch Lähmung der Atemmuskulatureintritt. Das Gift wird vom Muttertier ebenfalls dazu gebraucht um ihr Gelege vor Feinden zu schützen. (D.J. Tranter et al.) Beispiel für Drohung: Chamäleon (Chamaeleo spec.) Die Chamäleons, deren Fähigkeit zum physiologischen Farbwechsel allgemein bekannt ist, nutzen diese unter anderem zur innerartlichen Kommunikation und drohen anderen Artgenossen, indem sie eine Drohfärbung annehmen. Entgegen der allgemeinen Meinung können Chamäleons ihre Farbe nicht beliebig wechseln, sondern jeder Art steht nur ein begrenztes Farbspektrum zur Verfügung. Daher hat jede Art auch ihre eigene Drohgebärde. Eine besonders eindrucksvolle Drohfärbung hat das Bergchamäleon (Chamaeleo montium). Bei ihm färbt sich der Augenstrich tiefschwarz, während auf dem Vorderrücken sehr lebhafte, schwarze und gelbe Flecken auf grünem Grund auftreten. Die meisten Chamäleons drohen jedoch, indem sie eine sehr dunkle Färbung annehmen (z.B: Gewöhnliches Chamäleon Chamaeleo chamaeleon). Ausser zur innerartlichen Kommunikation (Erregungszustand, Balz, Schwangerschaft) dient der Farbwechsel aber auch noch zur Temperaturregulierung (dunkle Färbung zum Aufwärmen und helle Färbung zur Kühlung), spiegelt die körperliche Fitness der Tiere wieder ( kranke oder hungernde Tiere zeigen oft blasse Farbmuster) und ermöglicht den Tieren eine begrenzte Tarnung ( Schattierung, Gestaltauflösen, Anpassung an die Umgebung) (Grzimeks Tierleben ( 1971) H. Schifter). 1.1.3 Auffällig gefärbter Körperteile Da eine auffällige Dauerfärbung die Entdeckung durch Feinde erleichtert, müssen Tiere, die es sich nicht leisten können von all ihren Feinden entdeckt zu werden, eine KompromissLösung finden. Bei einigen Tieren ist nur ein Teil des Körpers auffällig gefärbt, bei anderen sind nur die Organe (Stacheln, Skalpelle der Doktorfische), von denen die Gefahr ausgeht aposematisch gefärbt. Bei manchen Arten werden diese auch erst bei einer Konfrontation präsentiert (versteckte Augen bei verschiedenen Nachtfaltern). Oft werden auffällig gefärbte Körperteile, auch zum Drohen benutzt (Statussymbole). Und vielfach auch zur optischen Reviermarkierung eingesetzt (Nicken vieler Echsen ( z.B. Anolis spec.)). Seite 69 Patrik Matter Beispiel für Warnung: Unke (Bombina spec.) Unken sind eine in Europa und Westasien verbreitete Kröten-Gattung mit 5 Arten. Der englische Namen „fire-bellied toads“ weisst auf die rot-gelb gefärbte Unterseite dieser Lurchen hin, welche sie bei Gefahr ihren Angreifern warnend entgegenstrecken. Zu Hause (Oftringen, AG) haben wir einen kleinen Gartenteich. Schon immer war ich fasziniert von der Vielfalt der Lebewesen, die sich in diesem Biotop ansiedelten. Eines der faszinierendsten war, eine Gelbbauchunke, welche durch ein unverwechselbares glockenhelles Quacken auf sich aufmerksam machte. Als ich sie entdeckte war, ich recht überrascht im Sommer eine schwimmende Kröte zu sehen, da Erd-, Kreuz- und Geburtshelferkröte eigentlich nur in der Paarungs- und Laichzeit im Wasser zu finden sind. Aus kindlicher Neugierde versuchte ich sie mit einem Netz zu fangen. Als ich jedoch mit dem Netz nach ihr fischte, stellte sie sich Tod und warnte mich, mit ihrer gelb-schwarz gesprenkelten Unterseite, vor ihrem Gift, welches selbst für einen Fuchs noch tödlich enden kann. Wenn Unken aber über ein so potentes Gift verfügen, wieso warnen sie dann nicht mit einer permanent exponierten Dauerfärbung davor, sie zu fressen? Einer der Gründe könnte sein, dass das Gift seine Wirkung nicht bei all ihren Feinden zeigt, so sollen schon verschiedentlich Reste von Unken in den Mägen von Vögeln (Amseln, Elstern) gefunden worden sein. In diesem Fall wäre eine permanente rötlich-gelbe Warnfärbung, etwa dorsal, als Attraktionsmerkmal für Fressfeinde besser geeignet. Als weitere Schutzmassnahme stellt sich die Unke noch tot. Dies soll Feinde abhalten, die kein Aas fressen. (Detailierte Erklärung für das Todstellen siehe Abschnitt: Mimik und Mimikri) (Grzimeks Tierleben ( 1971) R. Heusser). Beispiel für Drohung: Saumfinger (Anolis spec.) Saumfinger sind 20 – 50cm grosse Echsen. Sie gehören zur Familie der Leguane. Als hervorragende Kletterer leben sie vor allem auf Büschen und Bäumen in den wärmeren Regionen Amerikas und der Karibik. Bei Balzritualen und bei Revierstreitigkeiten stellen (männliche?) Saumfinger ihren auffällig gefärbten Kehlsack auf. Wird ein Saumfinger bedroht, vermag er mit Hilfe eines Hebelmechanismus des Zungenbeins seinen Kehlsack sehr stark zu spreizen, wodurch dessen volle Pracht sichtbar wird. So zeigen sie auf den Kehlsäcken weisse, gelbe, braune, rote, grüne, blaue und schwarze Farbtöne, die teilweise bunt gemischt sind. Einige Arten haben dünnhäutige Kehlsäcke, die nur im durchfallenden Licht richtig zur Geltung kommen. Bei Revierkämpfen hocken sich die Konkurrenten gegenüber und nicken mit ausgefahrenem Kehlsack um die Wette, meistens kann durch dieses Ritual ein Kampf vermieden werden. Ist eine Auseinandersetzung entschieden, nimmt der Unterlegene eine braune Farbe an, während der Sieger mit hellem grün seine Dominanz demonstriert (R.Mertens (1947)). 1.2. Verändern der Körperform Optische Signale sind auch durch eine Veränderung der Körperform möglich. Solche temporäre Signale werden erst durch eine Konfrontation mit einem Rivalen oder einem Feind ausgelöst. Diese Signale haben den Vorteil, auch auf Tiere zu wirken, die keine oder nur wenige Farben sehen können. Seite 70 Patrik Matter 1.2.1 Mechanismen die den Körper grösser erscheinen lassen Die meisten Tiere fürchten sich vor grösseren Gegnern und haben Hemmungen, diese an zu greifen. Daher haben viele aposematische Veränderungen der Körperform den Zweck, den Signalgeber grösser erscheinen zu lassen. Obwohl diese Erklärung intuitiv richtig erscheint, übt A. Zahavi (1999) in seiner „Handicap-Theorie“ begründete Kritik. Bei Drohungen ist allerdings das Vortäuschen einer imposanteren Grösse meist nur ein erstes Signal in einer langen Signalkette, wobei der Kampf mit jedem Glied wahrscheinlicher wird (Bsp. Kampffische). 1.2.1.1 Schrägstellen des Körpers Da die meisten Tiere länglich sind, wirken sie in der Seitenansicht grösser, als wenn sie sich frontal präsentieren (Bsp. Chamäleon (Chameleo spec.)). 1.2.1.2 Strecken des Körpers Auch das weit verbreitete Strecken des Körpers ermöglicht eine imposantere Erscheinung. Die meisten Revierstreitigkeiten beginnen mit einem Strecken des Körpers, auf welches dann weitere Drohungen folgen. Auch einem Raubfeind gegenüber kann es sich lohnen möglichst gross zu erscheinen. Während einige Tiere sich damit begnügen, nur den Kopf hochzuhalten (z.B Warane (Varanus spec.)), strecken andere den ganzen Körper (z.B „Katzenbuckel“). Manche Landraubtiere stellen sich sogar auf zwei Beinen, um bedrohlicher zu erscheinen. (z.B. „Grizzly“ (Ursus arctos horribilis)) sollte mit dem eingefügtem Satz abgegolten sein, oder? 1.2.1.3 Abspreizen von Gliedmassen Auch das Abspreizen der Gliedmassen kann zur Vortäuschung einer imposanteren Grösse dienen. Fische spreizen ihre Flossen (z.B Riffbarsche ( Pomacentridae)), Vögel stellen ihre Flügel auf (z.B Schwan (Cygnus spec.)). 1.2.1.4 Spezielle Organe Viele Tiere haben speziell zu diesem Zweck ausgebildete Organe, die entweder für einen permanenten Effekt sorgen (Hörner, Höcker, Geweihe, Mähnen, etc) oder aber im Ruhezustand unauffällig sind und erst bei einer Bedrohung sichtbar werden (Halskragen, Rückenkämme, Kehlsack, etc). Auch das Haarsträuben vieler Säugetiere gehört in diese Kategorie. 1.2.1.5 Aufblasen des Körpers Einige Tiere haben eine erstaunliche Art gefunden, ihren Körper zu vergrössern. Sie blasen sich einfach auf. Froschlurchen, Echsen und Schlangen pumpen ihre Lungen mit Luft voll (z. B. Erdkröte (Bufo bufo), Chuckwalla (Sauromalus spec.), Ringelnater (Natrix natrix), Puffotter (Bitis arietans)), wohingegen Kugel- und Igelfische (Tetradontiformes) Wasser in ihren Darm saugen. Seite 71 Patrik Matter 1.2.1.6 Abplatten des Körpers Vor allem Schlangen, aber auch einige Echsen, haben die Fähigkeit ihren Körper seitlich abzuflachen. Meistens dient dieser Mechanismus dazu, sich durch ein günstigeres Oberflächen-Volumenverhältnis („Bergmannsche Regel“) an kalten Morgen schneller aufzuheizen. Hat aber bei vielen Arten zusätzlich auch eine aposematische Funktion in dem ein grösserer Körpergrösse vorgetäuscht wird (Bsp. Kobra (Naja spec.). Beispiel für Warnung :Königskobra (Ophiophagus hannah ) Die Königskobra ist mit bis zu fünf Meter Länge die grösste Giftschlange der Welt, sie kommt in den dichten, feuchten Wälder Indiens, Südostasiens und auf den indoaustralischen Archipel vor. Wird eine dieser Riesinnen bedroht, richtet sie ihren Oberkörper etwa einen Meter auf, stellt ihren Halsschild („Hut“), beginnt laut zu zischen und greift mit gestelltem Hut an, falls sich der Gegner nicht zurückzieht. Dies gilt sowohl für einen Feind, wie auch für einen Konkurrenten. Beispiel für Drohung:Wolf (Canis lupus) Je aggressiver sich ein Wolf in einem innerartlichen Konflikt gebärdet, desto grösser will er wirken. Ein drohender Wolf, zieht seine Leftzen hoch, knurrt, streckt seinen Schwanz senkrecht in die Höhe, versteift seine Schultern und sträubt sein Fehl. Ein unterwürfiger Wolf hingegen duckt sich und klemmt den Schwanz zwischen den Beinen ein (E.Zimen 1997). 1.2.2 Schutzmechanismen Die Grenze zwischen der aposematischen Funktion gewisser Verhaltensweisen und reiner Schutzfunktion ist teilweise fliessend. So wirkt das Aufblasen des Kugelfisches einerseits als Vortäuschung eines stärkeren Gegners. Andererseits kann er mit diesem Grössengewinn so sperrig werden, dass ihn seine Raubfeinde nicht mehr fressen können. Durch eine Ausweitung des aposematischen Begriffes könnte man jegliche Schutzreaktion als aposematische Funktion sehen. Das Erbeuten eines geschützten Tieres erfordert mehr Aufwand, als die Erbeutung des Ungeschützten (In gleicher Hinsicht wirken ja auch die Prellsprünge der Thomsongazelle). Drohungen sollten einen Hinweis auf die Kampfstärke eines Konkurrenten geben. Schutzreaktionen sind dazu vor einem Kampf zu schützen und daher als Drohungen denkbar ungeeignet. 1.2.2.1 Zusammenrollen Igel, Ameisenigel, Schuppentiere, Gürteltiere und Asseln rollen sich bei Gefahr zusammen. Sie bieten so nur noch ihre stachelige bzw. gepanzerte Aussenseite als (vergleichsweise unattraktive) Angriffsfläche dar. Seite 72 Patrik Matter 1.2.2.2 Panzerung Eine Panzerung kann als ein ideales, mobiles Versteck betrachtet werden, in das sich ein Tier bei Gefahr zurückziehen kann. Einige Tiere tragen einen Panzer als integralen Körperbestandteil(Schnecken, Nautilus, Schildkröten, Muscheln,etc.).Andere Tierarten suchen aktiv ein Fremdgehäuse (Bsp. Einsiedlerkrebse) oder bauen sich dieses selbst (Köcherfliegenlarve, etc). Beispiel für Warnung: (Kleinohr-Igel (Erinaceus spec.)) Igel sind mit mehreren Gattungen in der Alten Welt verbreitet. In der Schweiz kommt allerdings nur der Westeuropäische Igel (Erinaceaus europeaus) vor. Er frisst alles, was nicht zu flink und nicht grösser als ein Junghase ist. Zur Not nimmt er auch pflanzliche Kost zu sich. Da er durch seinen stacheligen Panzer sehr gut geschützt ist, hat er nur wenig natürliche Feinde. Einzig grosse Greifvögel mit ihren gepanzerten Krallen können einen eingerollten Igel erbeuten. Auch sollen einige Dachse, Marder und Iltisse eine Methode entwickelt haben, um einen eingerollten Igel aufzurollen. 1.3. Bewegungen Bei Droh- und Warnungen, geht es darum, dem Gegner zu zeigen, dass er bei einem Angriff mit negativen Konsequenzen rechnen muss. Da die verschiedenen Tierarten auf unterschiedliche Arten kämpfen, ist je nach Tierart eine unterschiedliche aposematische Gebärde optimal, um die Kampfstärke zu demonstrieren. Die Thomson-Gazelle springt auf der Stelle hoch, um zu zeigen, dass sie noch Reserven für die Flucht hat. Hirsche laufen lange nebeneinander her, um abschätzen zu können, wer der Stärkere ist. Krokodile und Nilpferde reissen ihr gefährliches Maul auf, um Konkurrenten auf Abstand zu halten. Angesichts ihrer Vielfalt ist es allerdings unmöglich, solche aposematischen Bewegungen in vernünftige Kategorien einzuordnen. Beispiel für Warnung (Grauer Riffhai (Charcharinus amblyrhynchos)) Der über zwei Meter lange Graue Riffhai kommt in mittlerer Tiefe an allen pazifischen Korallenriffen vor. Er jagt bevorzugt in den tiefen Passagen und Kanälen, die das Innenriff mit dem Aussenriff verbinden. Falls man sich diesem Hai nähert, zeigt er ein stereotypisches Verhaltensmuster, welches „das aufgeregte Schwimmen“ genannt wird. Als erstes verringert der Hai sein Tempo, danach klappt er seine Brustflossen nach unten und beginnt seinen nach oben gestreckten Kopf hin und her zu bewegen, was seinem Schwimmen ein s-förmiges Aussehen verleiht. Schliesslich wendet der Hai und greift, teilweise nur zum Schein an. Auch andere Haiarten zeigen Teile dieses Verhaltensmusters, welches entfernt an die Bewegungen erinnert, die ein Hai macht, wenn er ein Stück Fleisch aus einer grossen Beute herausreissen will. Auch andere Haie zeigen teile dieses Verhalten und es wäre denkbar, dass nicht verstandene Warnungen die Ursache von vielen Haiattacken sein könnte. Dies könnte auch die Ergebnisse der Analyse der internationalen Haiangriffs-Statistiken erklären die eine Vielzahl von Angriffen aufweisst, bei denen das Opfer wenig bis kein Fleischverlust erlitten hat (Baldrige, Williams (1969)). Seite 73 Patrik Matter Beispiel für Drohung: Kampffische (Betta spec.) Kampffische sind kleine Fische, welche grosse farbenprächtige Flossen haben. Sie sind eine Gattung der Labyrinthfische die in Südostasien und im Malaischen-Archipel in allen sauberen Süssgewässern vorkommen. Ihr kämpferisches Drohverhalten hat dazu geführt, dass sie vor allem in Tailand für „tierquälerische“ Shows benutzt werde, in der zwei Kampffische -in einem engen Glas, der Möglichkeit der Flucht beraubt, bis zur Vernichtung des Gegners kämpfen. In freier Natur kommt es unter Kampffischen nicht zum Kampf, weil der Unterlegene stets die Möglichkeit zum Rückzug nutzt. Kampffische haben ein sehr ausgeprägtes Territorialverhalten und verteidigen ihr Revier gegenüber Artgenossen. Hierbei handelt es sich um ein mehrstufiges Drohritual. Wobei die nächste Stufe nur erreicht wird, wenn in der vorigen Stufe eine Pattsituation erreicht wurde (Grzimeks Tierleben ( 1971) D.Vogt). I. II. III. IV. Die beiden Männchen stellen sich Kopf an Kopf gegenüber und klappen die Kiemendeckel auf und spreizen ihre Flossen ab. Dann stellen sich die Männchen seitlich gegenüber für einen Grössenvergleich. Die Kontrahenten stellen sich wiederum seitlich gegenüber und erzeugen durch schlängelnde Körperbewegungen starke Druckwellen, die sich entgegenschleudern. Mit ihren empfindlichen Seitenlinien-Organ können sie so die Stärke ihres Gegners abschätzen. Nun versuchen sich die Rivalen gegenseitig in die Flossen zu beissen und Stücke davon rauszureissen. Der Kampf ist nun schon so ernst, dass nicht mehr die Flossen sondern der Körper des Feindes das Ziel der Bissattacken ist. 2. Mechanismen, die auf das Gehör wirken Als permanente Signale sind Laute nicht geeignet, da ihre Erzeugung mit permanentem Aufwand verbunden ist. Laute sind für Temporäre Warnungen grundsätzlich ebenso geeignet, wie optische Signale. Sie haben diesen gegenüber sogar den Vorteil, dass gewarnt werden kann, wenn man einen Raubfeind entdeckt, bevor dieser einem entdeckt hat.Dies muss nicht zwangsweise zur eigenen Entdeckung führen (Bsp. Warnung von Artgenossen: Murmeltier, Surikaten, Mungos etc.) und Warnrufe (siehe Bsp. Graudrosslinge). Drohlaute sind in der Natur weit verbreitet, da sie ein guter Indikator für die Körperspannung sind und zeigen wie nervös ein Gegner ist, also wie zuversichtlich er einem zu erwartenden Kampf gegenüber steht. (A.Zahavi 1997) Ebenfalls in dieses Gebiet, gehören die Reviergesänge der Singvögel. 2.1.Lauterzeugung mittels der Stimme Die meisten Warnungen vor einem Notwehrangriffwerden mit entsprechenden einschüchternden Lauten kombiniert (Bsp. Kobras fauchen zusätzlich zur Körpervergrösserung). Am sinnvollsten scheinen solche akkustische Warnungen gegenüber Raubfeinden, die ausschliesslich mit dem Gehör jagen. So überrascht es nicht, dass Fledermäuse von Motten (Familie: Arctiidae) mit aposematischen Klicklauten im Ultraschallbereich vor deren Ungeniessbarkeit gewarnt werden. (D.C.Dunning und Martin Kruger 1995). Eine Spezialität der akkustischen Warnungen ist das Phänomen der Warnrufe. Seite 74 Patrik Matter Konkurrenten können aufgrund der Lautäusserung ihres Rivalen auf dessen Verfassung schliessen, ebenso wie wir an der Stimme einer uns fremden Person ihre Stimmung ausmachen können (Bsp. Erdkröte (Bufo bufo). Beispiel für Warnung (Warnruf der Graudrosslinge (Turdoides squamiceps)) Graudrosslinge sind amselgrosse Singvögel, die hauptsächlich auf der arabischen Halbinsel, im Sinai und in Israel verbreitet sind. Ihr Lebensraum sind die warmen Wadis (ausgetrocknete Flusstäler, die nur wenige Tage im Jahr Wasser führen). Dort finden sie unter Buschwerk Schutz vor Raubvögeln und genügend Nahrung (Insekten, Skorpione, Schlangen, Eidechsen). Die Graudrosslinge leben in kleinen Familienverbänden und warnen , wenn einer am Himmel der Schatten eines Raubvogels sieht. Interessanterweise sind diese sogenannten Warnrufe jedoch viel lauter, als sie sein müssten um den nahen Familienverband zu warnen. Dies hat den Nachteil, dass auch der Raubfeind den Ruf hört und so gegebenenfalls eine gar noch nicht bemerkte mögliche Beute entdeckt. Wieso konnte sich also ein so lauter Ruf gegenüber der sinnvoller scheinenden leisen Variante in der Evolution durchsetzen? A.Zahavi (1997) vermutet, dass dies Laute vielleicht nur sekundär für die Gruppenmitglieder bestimmt sind und sich in erster Linie an den Raubfeind richten. Ein Raubfeind hat die grösste Chance eine Beute zu ergreifen, wenn er sie weit genug entfernt von einem sicheren Unterschlupf überraschen kann. Indem eine potentielle Beute also ihrem Raubfeind mitteilt, dass sie ihn gesehen hat, warnt sie ihn davor, seine Energie auf die Jagd nach einer geschützten Beute zu verschwenden. Diese Ansicht wird dadurch untermauert, dass auch Vögel, die Einzelgänger sind, Warnrufe ausstossen (z.B solitäre Würger im Winter (Lanius spec.)). Erstaunlich ist auch, dass die Vögel verschieden rufen, wenn die Gefahr vom Boden oder von der Luft aus droht. Auch dies hängt mit dem Empfänger der Warnrufe zusammen, ein Vogel, der einen Raubvogel (bzw. vor einem Raubvogel warnt), ruft meistens von den höchsten Wipfeln aus, damit er auch das weiter Verhalten des Feindes beobachten kann. Daher ist der Warnruf gegen den Raubvogel ein langezogenenes Trillern, das den Aufenthalt des stationären Rufers nicht sofort verrät, während der Warnruf für einen Feind am Boden aus einer Reihe von kurzen, rhytmischen „Tzwicks“ besteht, die durch die hüpfenden Bewegungen entstehen (A.Zahavi 1997). Beispiel für Drohen (Erdkröte (Bufo bufo )) Die Erdkröte ist die häufigste Kröte der Schweiz. Sie kommt in den mittleren Breiten in ganz Eurasien vor. Bemerkenswert sind die weiten Massenwanderungen der Erdkröten, bei denen einen grosser Männchenüberschuss herrscht, da Weibchen nicht jedes Jahr ablaichen. Entsprechend gross ist der Konkurrenzdruck zwischen den Männchen und nur das Männchen mit dem tiefsten Drohruf kann sich auf dem umklammerten Weibchen behaupten. , Je tiefer der Ruf der Kröten ist, desto grösser sind sie. (Davies und Halliday 1978). Damit wird auch im Dunkeln über die Stimme ein Grössenvergleich möglich. Zusätzlich könnte die Tiefe des Rufes auch noch zeigen, wie sicher sich ein Männchen seines Weibchens ist, da der Ruf durch angespanntes Klammern erhöht wird. Nur wer sich seines Weibchens sicher ist, kann es riskieren sich nur lose an ihm festzuklammern, da er es im Falle eines Angriffs zurückerobern kann. (A.Zahavi 1997). Seite 75 Patrik Matter 2.2 Lauterzeugung mit anderen Mitteln Lautäusserungen mit anderen Mitteln haben den Vorteil, dass sie meistens besser geeignet sind, um die Gefährlichkeit (Stärke) eines Tieres zu beurteilen. Dies sieht man besonders gut an dem Drohritual des Gorillas, der zuerst einige spitze Schreie ausstösst, sich dann auf die Hinterbeine stellt und sich als Höhepunkt mit den Händen mehrmals gegen die Brust trommelt. Eine andere beeindruckende Drohgebärde ist das Stampfen einiger Huftiere (z.B Kaffernbüffel (Syncerus caffer)). Beispiel für Warnung (Klapperschlangen (Crotalus spec.)) Viele Schlangen (z.B. Sandrasselottern (Echis spec.)) produzieren warnendende Geräusche, in dem sie Körperpartien gegenläufig aneinander reiben, was zu einem rasselnden Geräusch führt. Auch das aposematische Vibrieren mit dem Schwanzende ist bei Schlangen (z. B Nachtbaumnattern (Boiga spec.)) weit verbreitet. Oft wird das Schwanzende zur Verstärkung der warnenden Wirkung gegen den Boden geschlagen, was ein deutliches Warngeräusch erzeugt. Bei Klapperschlangen hat sich das Schwanzende zu einem eigentlichen Warnorgan entwickelt. Die Häutungsreste bilden mit der Zeit eine mehrgliedrige „Rassel“, welche bei grösseren Arten bis zu 30 m weit zu hören ist. 3. Mechanismen, die auf Geruchsinn, Geschmacksinn oder auf andere chemische Sinne wirken Geruch an Land (bzw. Geschmack im Wasser) ist eine Möglichkeit, permanente aposematische Signale zu geben. Oft wird der spezifische Körpergeruch (bzw. Körpergeschmack) zu einem Warnsignal, welches keine extra Kosten verursacht. Für Drohungen sind chemische Signale nicht sonderlich geeignet, da sie weder als Mass für momentane Stimmung, noch Kampfstärke eines Konkurrenten dienen können. Trotzdem sind sie vor allem als Reviermarkierungen weit verbreitet (Raubtiere (z.B. Rotfuchs (Vulpes vulpes))). 3.1. Geruch und Geschmack Jede Tierart hat einen temporären Körpergeruch, und es wäre denkbar, dass dieser Körpergeruch von Beutegreifern als Warnung gedeutet wird, nach dem sie einmal schlechte Erfahrung mit einer solchen Art gesammelt haben. Experimente haben gezeigt, dass der Nasenbär (Nasua nasua) der ausschliesslich mit dem Geruchssinn jagt, Insekten mit aposematischem Geruch verschmäht (J.Myers 1930). Dieser Mechanismus könnte vor allem für Pflanzen und Pilze die einzige Möglichkeit sein, um auf ihre Giftigkeit hinzuweisen (S.Camazine 1985). Viele Tiere sondern stinkende Stoffe aus, wenn sie attackiert werden. Dies kann auf zwei Arten schützen. Einerseits betäubt der starke Geruch den Geruchssinn des Raubfeindes – was besonders in stockdunkler Nacht oft lebensrettend sein kann. Andererseits wird nur ein sehr hungriger Raubfeind eine stinkende Beute verzehren. Was an Luft der Körpergeruch ist unter Wasser der Körpergeschmack. Es wird angenommen, dass etwa Fische die Giftigkeit von Schwämmen und Weichtieren anhand chemischer Stoffe erkennen könnten. Viele der nicht kryptischen Invertebraten (Wirbellosen) sind giftig für Fische (G.J.Bakus 1981). Eine permanente aposematische Wirkung der Ausdünstungen Seite 76 Patrik Matter konnte jedoch noch nicht nachgewiesen werden. Viele Meerestiere (Seifenbarsch, MosesScholle, Tintenfische, Kofferfisch, Lochschnecken, etc) sondern jedoch übel schmeckenden bis giftigen Schleim ab, der sie vor Angriffen durch ihre Fressfeinde schützt. Auch an Land gibt es aposematische Mechanismen, die auf den Geschmackssinn wirken. Die Haut vieler Amphibien ist so schleimhautreizend, dass diese sofort wieder ausgespieen werden, wenn sie ein unerfahrener Raubfeind versehentlich erbeutet. Auch einige Schlangen und Echsen haben die Fähigkeit entwickelt über die Haut bitterschmeckende Stoffe auszuscheiden ( Bsp: Macropisthodon rhodomelas (Familie: Wassernattern)) oder um den Mund giftigen Schaum zu produzieren (Bsp. SkorpionKrustenechse (Heloderma horridum)) (R.Mertens (1947)). Beispiel für Warnung (Ringelnatter (Natrix natrix)) Wird eine Ringelnatter überrascht, zeigt sie ein komplexes Warnverhalten, um Feinde vor einem Angriff abzuhalten. Zuerst beginnt sie agressiv zu zischen und zu züngeln, plattet den aufgestellten Teil ihres Körpers ab und setzt sich mit Scheinbissen zur Wehr. Falls ein Feind dann noch nicht von ihr ablässt, entleert sie aus ihren Postanaldrüsen eine stinkende Flüssigkeit. Einige Ringelnattern bilden dann ein Knäuel, indem sie ihren Kopf verstecken, andere stellen sich tot. Eine sich tot stellende Ringelnatter lässt aus dem geöffnetem Mund ihre Zunge raushängen und kann teilweise sogar einige Tropfen Blut aus dem geöffneten Mund träufeln lassen. Meistens werden jedoch nur einzelne Aspekte dieses Verhaltens gezeigt (Eckstein HP (1993)). 3.2. Ausspritzen von Körperflüssigkeiten Viele Tiere setzen chemische Stoffe über eine gewisse Entfernung ein. Oft zielen solche Attacken auf den empfindlichen Gesichtsbereich des Feindes. Dies dürfte das lohnendste Ziel sein, da so die Sinne des Feindes vorübergehend betäubt sind und die Beute entkommen kann. Krustenechsen können die giftige Schaummasse um ihren Mund nach dem Angreifer blasen, die Krötenechsen spritzen einige Tropfen ihres Blutes aus ihren Augenliedern und das Stinktier spritzt eine stinkende Flüssigkeit auf Feinde. Beispiel für Warnung: Stinktier (Familie: Mephitinae) Stinktiere sind marderartige Tiere, die durch eine eigentümliche weiss-schwarze aposematische Färbung auffallen. Den Namen „Stinktier“ erhielten sie aufgrund ihres Warnverhaltens, bei dem sie ihren Feinden eine übelriechende Flüssigkeit aus ihren Aftertaschen ins Gesicht spritzen. Wird ein Stinktier bedroht dreht es dem Gegner das erhobene Hinterteil zu und kratzt oder klopft mit den Vorderfüssen auf den Boden. Falls diese Warnung noch nicht genügt, dreht es fauchend den Kopf nach seinem Gegner um, und stülpt den After vor. Zieht sich nun der Feind nicht sofort zurück, spritzt das Stinktier seine Abwehrflüssigkeit auf bis zu vier Meter Entfernung zielsicher ins Gesicht seines Gegners. Falls solche Stinkflüssigkeit in die Augen eines Feindes gelangt, ruft sie heftige Schmerzen empor und soll sogar eine vorübergehende Erblindung hervorrufen. Auf jeden Fall lähmt dieser bestialische Gestank vorübergehend den Geruchssinn des Feindes und schwächt ihn auch Tage später noch. Alle Marder haben die Möglichkeit als Warnung mit ihren Aftertaschen stinkende Flüssigkeit zu spritzen, dies jedoch meistens weniger weit und weniger ausgiebig als die Stinktiere (Grzimeks Tierleben ( 1971) K.Herter). Seite 77 Patrik Matter Beispiel für Drohen: (Rotfuchs (Vulpes vulpes)) Rotfüchse zeigen ein ausgeprägtes Territorialverhalten. Um die Grenzen seines Reviers deutlich zu machen markiert er die Grenze mittels einer speziellen Stinkdrüse. Diese Stinkdrüse welche unterhalb der Schwanzwurzel sitzt wird auch Viole genannt, da sie einen an Veilchen erinnernden Duft verströmen kann. Für andere Füchse die in diese Revier eindringen wollen, bildet sich so eine Geruchsgrenze, deren Überschreitung ein Konflikt mit dem Revierhalter auslöst (F.Labhardt et al. (1990)). 4. Mechanismen, die auf den Tastsinn wirken Um auf den Tastsinn eines Feindes wirken zu können, sollte man diesem sehr nahe sein. Deshalb gibt es nur sehr wenige Warnungen, die auf den Tastsinn eines Feindes zielen. Die meisten innerartlichen Drohgesten sind Bestandteile ganzer Drohketten, wobei jede Stufe dieser Drohketten von den Drohenden mehr Einsatz verlangt und einen anschliessenden Kampf wahrscheinlicher macht. Spätestens vor dem eigentlichen Kampf findet oft ein direkter Kräftevergleich mit Körperkontakt statt. Daher sind Drohungen, die auf den Tastsinn wirken, in der Endphase von Drohketten sehr weit verbreitet. Beispiel für Warnung (Schimpanse (Pan troglodytes)) Schon lange ist bekannt, dass Schimpansen und andere Affen manchmal mit Gegenständen nach ihren Wärtern werfen, oder mit Knüppeln nach ihnen schlagen. Im Zoo Basel werden sogar manchmal Zuschauer von den Schimpansen mit Kot beworfen. Experimente mit einer Leopardenattrappe haben gezeigt, dass Schimpansen vermutlich auch in freier Wildbahn Feinde vertreiben, in dem sie mit Stöcken nach ihnen schlagen oder mit Gegenständen nach ihnen werfen (Grzimeks Tierleben ( 1971) A. Kortlandt). Beispiel für Drohung (Hornträger (Familie:Bovidae)) Das gemeinsame Merkmal der Hornträger sind ihre Hörner, wobei meistens die Männchen viel grössere Hörner haben als Weibchen. Oft haben Weibchen spitzte Waffen die bestens zur Verteidigung gegen Angreifer geeignet sind, während Männchen mächtige Hörner haben, die zusätzlich als Statussymbol und zum Drohen benutzt werden. Hornlose Weibchen (z.B Nilgau-Antilope) tragen Konflikte mit Artgenossinen aus, indem sie mit ihrem Kopf in die Seite ihrer Gegnerin schlagen. Auch die Böcke der Gemsen und Schneeziegen kämpfen noch auf diese ursprüngliche Art, was nicht selten zu schweren Wunden führt. Daher haben viele Horntiere eine Möglichkeit entwickelt, auf ungefährlichere Art zu kämpfen. Sie richten ihren Angriff nicht mehr auf die verletzliche Seite, sondern zielen mit der Wucht ihres Angriffs auf die unempfindlichen Hörner ihres Kontrahenten. Die häufigsten Drohrituale sind heute das Rammen, das Fechten und das Stirndrängen. Beim Rammen rasen die Gegner mit Anlauf aufeinander zu und rammen die Hörner gegeneinander. Beim Fechten treffen die Rivalen mit der Hornmitte aufeinander und es folgt ein Schlagwechsel, wobei die einzelnen Schläge vom Gegner pariert werden müssen, was zu langen lanzenartigen Hörnern geführt hat. Beim Stirndrängen verhacken die Kontrahenten die Hörner ineinander und versuchen den andern zu schieben. Dazu sind stark strukturierte Hörner von Vorteil, da so Verletzungen durch Abrutschen verhindert werden können (Grzimeks Tierleben ( 1971) F.Walther). Seite 78 Patrik Matter 5. Mechanismen, die auf eine Mehrzahl von Sinne wirken Die bisherigen Beschreibungen sind stark vereinfachend. Es gibt nur sehr wenige Tiere, deren Warn- oder Drohrituale nur auf einen Sinn wirken. Die Ursache dafür ist einerseits, dass Warnungen auf alle möglichen Empfänger der Signale abgestimmt sein müssen (Warnfarben nützen nichts gegen farbenblinde Feinde, etc). Andererseits bedeuten die verschiedenen aposematischen Mechanismen unterschiedlichen energetischen Aufwand. Deshalb hängt die Wahl bzw. Stärke des Signals auch von dessen Wichtigkeit für den Signalgebers ab (ein hungriges Tier wird länger um eine Beute streiten, als ein Gesättigtes). X.LITERATURVERZEICHNIS Arak A & Enquist M (1993) Hidden preferences and the evolution of signals Philosophical Transactions: Biological Sciences Serie B Vol 340 Iss 1292 207-213 Arrese CA et all. (2003)Trichromacy in Australian Marsupials Current Biology, Vol. 12, 657–660 Bakker TCM (1993) Positive genetic correlation between female preferences and preferred male ornament in sticklebacks Nature 363 255-257 Baldridg HD, Williams J Shark attack – feeding or fighting Military Medicine Vol 134 130ff Böhme W (1979) Zum Höchstalter des Feuersalamanders Salamandra 15 176-179 Brodie ED (1977) Salamander Antipredator Postures Copeia 2 523-535 Brodie ED (1990) The antipredator arsenal of fire salamanders: Spraying of Secretion…. Herpetologica 46 1 1-7 Burrt EH (1979) The behavioural significance of color Garland STPM New York Burtt EH(1977) The coloration of wood warblers (Parulidae) Ph. D. Thesis, Unive. of Winsconsin-Madison Cameron DA, Pugh ENJ (1991) Double cones as a basis for a new type of polarization vision in vertebrates. Nature 353 161-164 Campenhausen MV, Kirschfeld K (1998) Spectral sensitiy of the accessory optic system of of pigeon Journal of Comperative Physiologie A 159 473-479 Seite 79 Patrik Matter Chappuis C (1971) Un example de l’influence du millieu sur les émission vocales des oiseaux : l’evolution des chants en forêt équatoriale Terre Vie 25, 183-202 Chen VJ, D’Zmura M. (1998) Test of a convergence model for color transparency perception. Perception, 27, 595-608 Conway BR (2003) Colour Vision: A Clue to Hue in V2 Current Biology, Vol. 13, R308–R310 Cott HB (1940) Adaptive Coloration in Animals III. Adaptive Radiation in colour Ressemblances. Methuen London Cuthill IC et all (2000) Ultraviolet Vision in Birds Advances in the Study of Behaviour 29 159-214 De Valois RL, De Valois KK. (1993) A multi-stage color model. Vision Res. 33(8):1053-65. Dice LR and Blossom PM (1930) Studies of Mammalian Distribution Ecology Carnegie Inst. Wash. Pub. 485 129ff Dietrich N. (1999): Jahreszyklus der Feuersalamander (Salamandra salamandra) des Neissetales Landkreis Löbau-Zittau. Elaphe N.F. 7: 62-65. Duellman WE, Trueb L (1986) Biology of Amphibians New York, Mc. Graw Hill Eckstein, H.-P. [1993]. Untersuchungen zur Ökologie der Ringelnatter (Natrix natrix) Jb. für Feldherpetologie, Beiheft 4 Verlag für Ökologie u. Faunistik, Duisburg Eshel I (1978) Prey-predator nonzero-sum game and evolution of gregarious behavior of evasive prey American Naturalist 112 (987): 787-795 Feldmann R (1987) Überwinterungsverhalten des Feuersalamanders Praxis Naturwiss. Biologie 27 246-248 Fisher RA (1930) The genetical theory of natural selection Oxford University Press, Oxford Fitzgibbon CD, Fanshawe JH (1988) Stotting in Thomson Gazelles An honest Signal of Condition-Behaviour Ecology and Sociobiologiy Flamarique IN, Hawryshyn CW, Harosi FI (1998) Double-cone internal reflection as a basis for polarization detection in fish Journal of the optical society of America –Optics Image Science and Vision 15 (2): 349-358 Seite 80 Patrik Matter Fugle et all. (1984) Signals of status in wintering white-crowned sparrows Ainmal Behaviour 32 283-293 G.Fachbach (1988) Röhrenknochenentwicklung und Altersbestimmung bei Salamandra atra Zoologischer Anzeiger Jena 221 (3/4) 188-200 Gegenfurter KR, Sharpe LT (1999) Color Vision: form genes to perceptions Cambridge University Press New York Gloger, CL (1833) Das Abändern der Vögel durch Einfluss des Klimas. Breslau, Deutschland Grossenbacher K, Günther R (1996) Alpensalamander In Günther R. Die Amphibien und Reptilien Deutschlands Gustav Fischer Jena Grzimek B (1971) Grzimeks Tierleben Kindler Verlag AG, Zürich Günther R (1996) Die Amphibien und Reptilien Deutschlands Gustav Fischer Jena Hagedorn J, D' Zmura M (2000) Color appearance of surfaces viewed through fog. Perception 29, 1169-1184 Hailman JP (1977) Optical signals: Animal communication and light Indiana University Press Bloomington Hamilton WD, Zuk M (1982) Heritable true fitness and bright birds: A role for parasites? Science 218 384-387 Hinde RA (1970) Animal Behaviour: A Synthesis of Ethology and Comperative Psychology 2nd Edition Mc. Graw-Hill, New York Hingston, RWG (1933)The meaning of colour and adornement. London Horter M, Greven H (1981) Zur relativen Geniessbarkeit juveniler Feuersalamander Amphibia-Reptilia 2, 15-21 Hubbard ME (1903) Correlated protective devices in some California Salamanders University Cal. Pub. Zool. 4 157-70 Hurwich LM and Jameson D (1957) An opponent-process theory of color vision Psychological Review. 64 384-404 Jacobs GH (1993) The distribution and Nature of Colour vision among Mammals Biological Review 68, 413-471 Jecklin L (1935) Zur Kältebeständigkeit des Feuersalamanders Rev. Suisse Zool. 42 29 731-738 Seite 81 Patrik Matter Joly J. (1968): Données écologiques sur la salamandre tachetée Ann. Sci. Nat. Zool. 12: 301-366. Kabisch K, Belter H (1968) Das Verzehren von Amphibien und Reptilien durch Vögel Zoologische Abhandlung Museeum für Tierkunde Dresden 29 (15) 191-227 Kästle W (1986) Rival combat in Salamandra salamandra In Rocek Z (Ed.) Studies in Herpetology Prag 525-528 Kelber A, Vorobyev M, Osorio D (2003) Animal colour vision ± behavioural tests and physiological concepts Biol. Rev., 78, pp. 81-118 Kelber A., Balkenius A. and Warrant EJ. (2002). Scotopic colour vision in nocturnal hawkmoths. Nature 419, 922–925. Klewen R (1991) Die Landsalamander Europas (Neue Brehmbücherei 584) Ziemsen Wittenberg Lutherstadt Krey, Grunau, Bräuer (1987) Exempla ophthlmologica: Bildatlas zur Physiologie und Pathophysiologie des Auges Höchst-Pharma, Zürich Kulp TD, Fuld K (1995) The Prediction of Hue and Saturation for Non-spectral Lights Vision Research 35 (21) 2967-2983 Labhardt F. (1990) Der Rotfuchs Parey, Berlin Lennie P (2000) Color vision: Putting it together Current Biology 10 589-591 Lorenz K (1962) The function of colour in coral reef fishes Proc. R. Inst. Gr. Br. 39 282-296 Lorez K (1978) Vergleichende Verhaltensforschung. Grundlage der Ethologie Wien Lunau K (2002) Warnen, Tarnen, Täuschen Wissenschaftliche Buchgesellschaft Darmstadt Luria SM, Kinney (1970) Underwater vision Science 167 1454-1461 Marler P (1995) Characteristics of some animals calls Nature 176 6-8 Maynard Smith J and Harper DGC 1988 The evolution of aggression: can selection generate variability Philosophical Transactions: Biological Sciences Serie B Vol 319 557-570 Seite 82 Patrik Matter Mc Farland WN, Munz FW (1975) The photic environment of clear tropical seas Vision Research 15 1063-1070 Melsheimer M (1886) Zur Naturgeschichte des Erdsalamanders Verh. Naturhist. Ver. Preuss Rhein Corr Bl 43 80-86 Mertens R (1947) Die Warn-und Drohreaktionen der Reptilien Abhandlung der Senckenbergischen Naturforschenden Gesellschaft Vol 471 1-108 Milinski M & Pakker TCM (1990) Female sticklebacks use male coloration in mate choice and hence avoid parasitited males Nature 344 330-333 Myers JG (1930) Observations of the insect food of the coati Ibid. 5 69-75 Parson PA (1990) Fluctuating asymmetry: An epigenetic measure of stress Biol.rev. Cambr. Phil. Soc. 65 131-145 Peichl L et all (2001) For whales and seals the ocean is not blue: a visual pigment loss in marine mammals European Journal of Neuroscience, Vol. 13, pp. 1520-1528 Porter WP et all (1973) Behavioral implications of mechanistic ecology Oecologia 13 1-54 Przyrembel C, Keller B, Neumeyer C (1995) Trichomatic color vision in the Salamander (Salamandra salamandra) Journal of Comperative Physiology 176 (4) 575-586 Pulton EB (1908) Essays on Evolution 1889-1907 Oxford pp. xlviii + 479 Reiner A, Shatp LT, Zrenner E (1991) Is colour vision possible with only rods and blue sensitve cones? Nature Vol. 352 pp 798-800 Rieger G (1992)Improvement of contrast sensitivity with yellow filter glasses. Can J Ophthalmol. Apr;27(3):137-8. Riipi M (2001) Multiple benefits of gregariousness cover detectability costs in aposematic aggregations Nature Vol 413 512-514 Riipi M et all. (2001) Multiple benefits of gregariousness cover detectability costs in aposematic aggregations Nature 413 512-514 Robinson SR (1994) Early vertebrate colour vision Nature Vol 367 p.121 Seite 83 Patrik Matter Roper T (1986) Badges of status in avian society New Sci 109 38-40 Ryan M.J. (1985) The tungara frog: a study in sexual selection and communication. University of Chicago Press, Chicago Schenkel R (1956) Zur Deutung der Balzleistung einiger Phasianiden und Tetraonieden Der Ornithologische Beobachter 53: 182-201 Schlenoff DH (1985) The startle reponse of blue jays to Catocala (Lepiidoptera: Noctuidae) prey models. Animal behaviour 33 : 1057-1067 Schultze M (1866) Ueber Stäbchen und Zapfen der Retina Arch. F. mikr. Anat. Bd 2 175-28 Smith NG (1966) Evolution of some arctic gulls (Larus) Ornith. Monogr. (AOU) 4 1-99 Sauer, H. und H. Weisbecker (1994): Einheimische Schlangen als gelegentliche Verfolger des Feuersalamanders (Salamandra salamandra) - zwei Feldbeobachtungen. Natur und Museum, 124: 349-350. Sochurek E (1956) Kreuzottern fressen auch Alpensalamander DATZ 9 195 Speed MP (2001) Can receiver psychology explain the evolution of aposematism?. Animal Behaviour:Vol 61, pp 205-216 Summers E., Clough ME (2001) The evolution of coloration and toxicity in the poison frog family (Dendrobatidae) Proceedings of the Nat. Academy of Science of the USA Vol. 98 6227-6232 Sword GA et all. (2000) Density-depend aposematism in desert locust Proceedings of the Royal Society of London B.267 63-68 Takamatsu M, Nakashima Y (2001) Brightness perception of light source colors in dense fog Optical Review 8 (3): 198-202 Thayer AH (1897) The Law which underlies Protective Coloration Annual Report of the Smithsonian Institution, Washington Thiesmeier B (1992) Ökologie des Feuersalamanders Westarp Wissenschaften Essen Tiesmeier B, Günther R (1996) Feuersalamander In Günther R. Die Amphibien und Reptilien Deutschlands Gustav Fischer Jena Seite 84 Patrik Matter Tranter DJ, Augustin O (1973) Observations on life-history of blue-ringed octopus (Haplochlaena maculosa) Marine Biology Vol 18 115-128 Triemen R (1869) On some remarkable mimetic analogies among African Butterflies Transactions of the Linnean Society Zool. 26 497-522 Uchikawa K et al (1984) Luminance and saturation of equally bright colors Color research and Application 9, 5-14 Vorobyev M (2003) Coloured oil droplets enhance colour discrimination. Proc R Soc Lond B Biol Sci. 2003 Jun 22;270(1521):1255-61. Walls GL (1942) The vertebrate Eye and its adaptive Radiation Hafner Publishing New York Wickler W (1962) Ei-Atrappen und Maulbrüten bei bei afrikanischen Cichliden Z. Tierpsycologie 19 129-164 Yachi S (1998) The evolution of warning signals Nature Vol 394, 882-884 Zahavi A (1997) The Handicap Prinziple. A Missing Piece of Darwin’s Puzzle Oxford University Press, Oxford Zahavi et all (1981) Natural selection, sexual selection and the selection of signals Evolution Today 113-138 Pittsburgh Carnegie-Mellon University Press Zimen E (1997) Der Wolf, Verhalten, Ökologie und Mythos Knesebeck Verlag München XI.GLOSSAR absorbieren Absorbtionsmaximum Abstraktion Adulttiere Aerosol Akinese aposematisch Bachbiozönose Balz Bergmannsche Regel Biolumineszenz. Bipolar Chlorophyll collinen bis submontanen Stufe. aufnehmen Bereich der maximalen Aufnahme Vereinfachung, Zusammenfassung Geschlechtsreife Individuen Schmutzpartikel Regungsloses Verhalten (Sich-Tod-stellen) drohende, warnende Alle Lebewesen des Baches Paarungsverhalten Innerhalb einer Gattung sind Tiere um so kleiner je näher sie am Äquator leben Biochemisch erzeugtes Licht Eine Zelle die Signale weitervearbeiten kann Blattgrünkorn = Teil der Zelle in dem die Pflanzen Licht in Energie umwandeln ca. 450 - 1000 m.ü. M Seite 85 Patrik Matter conspicious Di, Tri-, Tetra- und Penta Embryonen empirische Befunde epigamische Färbung episematische Färbung Evolution. Extremitätenwurzeln Facettenaugen Fisher’s Hypothese über die sexuelle Selektion Metamorphose Mimikry M-Zapfen natürliche Selektion Parotiden Partikel Photonen photosensitives Pleiotroische Wechselwirkung auffällig Zwei, Drei, Vier und Fünf Ungeborenes Leben Resultate von Experimenten Balz-Färbung Färbung zur Arterkennung Entwicklung des Lebens Bereich des Bein, bzw Armansatzes Insektenaugen, die aus vielen einzel Augen bestehen Weibchen bevorzugen gewisse Merkmale und vererben diese Bevorzugung an ihre Nachkommen Licht von oben, erhellt alle Körper um dies auszugleichen sind viele Tiere oben dunkler als unten verwirrende Muster, die mit dem Hintergrund verschmelzen Arme, Beine, etc. Signale kosten und deshalb lohnt es sich nicht zu betrügen. Anpassung die früher nötig war Vermutung Einzeltier innerhalb der Art Stärke Energieeinheit noch nicht fortpflanzungfähige Tiere Vielfältigkeit Teil, Stück Streit Zusammentreffen Bestand Zusammenarbeit tarnende Färbung Lippen der Hundeartigen longwave = rot (570 nm) unterschiedliche Farbreize vermitteln den gleichen Farbeindruck Umwandlung Nachahmung von Tieren middlewave = grün (535nm) natürliche Auslese (nur wer gut angepasst ist überlebt) Giftdrüsen auf dem Kopf der Salamander Schmutzteilchen Lichteilchen auf Licht reagierend Ein Gen bestimmt mehrere Merkmale Präferenz Bevorzugung Gegenschattierung (Tayersches Prinzip) Gestaltauflösende Färbung (Somatolyse) Gliedmassen Handicap-Prinzip historisches Relikt Hypothese Individuen innerartlichen Intensität Joule Juvenile Tiere Komplexität Komponente Konflikt Konfrontation Konsistenz Kooperation kryptische Färbung Leftzen L-Zapfen Metamerie Seite 86 Patrik Matter Reflektion Retina Rodopsin Selektionsdruck Sensitivität sensorisches Fenster sensory biases somatolytische Spermatophore Stäbchen Surviving of the fittest S-Zapfen tayerschen Prinzip territoriales Verhalten thermoregulatorische visiblen Lichtes Wildtypen Zapfen Spiegelung Netzhaut Sehpurpur = Farbstoff der Stäbchen Gefahr, Umstand - so dass man sich anpassen muss Feinfühligkeit sensory biases = Signale, die ein bestimmtes Muster haben werden verstärkt wahrgenommen sensorisches Fenster gestaltauflösende Schutzbehälter für Spermien Sehzellen für das Hell/Dunkelsehen Überleben des Stärkeren (Angepassteren) shortwave = blau (440nm) Gegenschattierung Revier abstecken Temperatur regulierend sichtbares Licht natürliche Formen Sehzellen für das Farbsehen Seite 87 Patrik Matter X I I . TA B E LLE N Seite 88 Patrik Matter Seite 89 Patrik Matter Seite 90 Patrik Matter Seite 91 Patrik Matter Seite 92 Patrik Matter Seite 93 Patrik Matter Seite 94 Patrik Matter Seite 95 Patrik Matter Seite 96 Patrik Matter XIII. DANKSAGUNG In erster Linie möchte ich meinem Betreuer Herrn Prof. Dr. J. Meier für seine grossartige Unterstützung danken, der selbst in seinen Ferien Zeit fand, meine Fragen zu beantworten; sowie Herrn Prof. D.G. Senn, der trotz seinen vielen Auslandreisen immer wieder Zeit fand sich mit mir zu treffen. Ebenfalls möchte ich den vielen Fachexperten herzlich danken, die sich immer wieder die Mühe nahmen meine fachspezifischen Fragen zu beantworten: Prof. K.Grossenbacher, Alpensalamander-Experte des Naturhistorischen Museums Bern Prof. D. Mebs, Gifttierexperte: Uni Frankfurt Prof. Durrer, Amphibienexperte der Uni Basel Prof. B. Tiesmeier, Feuersalamander-Experte PD Dr. P. Meyer, Ophthalmologische Klinik Basel Prof. S. Messmer, technischer Mitarbeiter des Physikalisches Institut Basel Weiterer Dank geht an die lokalen Feuersalamander-Experten, welche mir vor allem im Anfangsstadium meiner Diplomarbeit wertvolle Hinweise zum Thema gaben, insbesondere in Form von Literatur, Empfehlungen und Beobachtungsorten: P. Brodmann F.Labhardt D.Küry M.Kopp Und natürlich ebenfalls den vielen Kollegen und Kolleginnen, die mich mit Rat und Tat unterstützt haben: Florian Spinnler Esther Scherer Simone Zahler Beat Schaub Sarah Pfister Seite 97