Folien: 4,6 MB - Institut für Biologie
Werbung

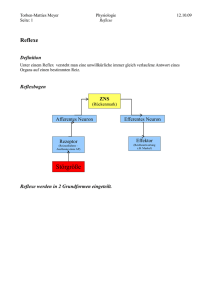
Organisatorisches • Klausur: – 24.01.08 – grosser Hörsaal der Zoologie – 12:50 Uhr Angeborene und Erlernte Verhaltenskontrolle 1. Reiz – Reaktion 2. Spontanverhalten 3. Rhythmisches Verhalten 1. Reiz - Reaktion ? Reflexe • Reflexe sind automatische, beliebig wiederholbare, zweckgerichtete Reaktionen. • Eigen- und Fremdreflexe; mono -, di -, tri -, polysynaptische, (non-synaptische) Reflexe; motorische vegetative Reflexe; angeborene und erlernte (bedingte) Reflexe; tonische und phasische Reflexe; T- und H-Reflexe. Eigen- und Fremdreflexe Monosynaptischer Eigenreflex Polysynaptischer Fremdreflex Beispiel für einen polysynaptischen Reflex bei einem Insekt Reflexe – Reaktionen – angeborene Auslösemechanismen 2. Spontanverhalten • Kaum Mechanismen bekannt („Rauschen“, „Triebe“) • Lange vernachlässigt oder ignoriert („response“) • Erst vor kurzem neue Studien zur Biologie: Drosophila am Drehmoment-Kompensator Variables Verhalten bei konstanter Reizsituation 3. Rhythmische Motorische Aktivität A A 1s B B C C Stemmphase Rhythmische Bewegungsmuster • • • Bewegungen wie Laufen, Rennen, Fliegen, Schwimmen, Kaubewegungen, Atmung etc. werden durch neuronale Netzwerke im ZNS erzeugt und durch sensorische Rückkopplung den Umwelterfordernissen angepasst. Der grundlegende alternierende Rhythmus wird dabei von einem Netzwerk von Neuronen im ZNS erzeugt (zentraler Mustergenerator „CPG“, zentrales Programm, Oszillator). Typischerweise werden nur der Beginn und das Ende der Bewegungsfolge bewusst kontrolliert, während der Ablauf mehr oder weniger automatisiert ist. ein kontinuierlicher Erregungseingang wird in einen rhythmischen Ausgang übersetzt (Brown Half Center Model) Alternierende Rhythmen sind oft das Ergebnis von Zentralen Rhythmusgeneratoren (CPG = Central Pattern Generator) im ZNS: Schwimmbewegungen von Neunaugen Das lokomotorische Netzwerk im Rückenmark des Neunauges Motorische Aktivität im intakten Neunauge Motorische Aktivität im isolierten Rückenmark nach Grillner et al. Laufen im Menschen Wie steuert unser Gehirn Bewegungen ? Windungen (Gyri) und Täler (Sulci) als anatomische Landmarken „Karten“ im Gehirn: Topographische Organisation Funktionen derHemisphäre Großhirnrinde, linke Hemisphäre Funktionen der linken Nach Karl Kleist, Nach 1920-1930 Karl Kleist, 1920-1930 „Phrenologie“ des 19. und 20. Jahrhunderts Karten im Gehirn (heute) Bewegungen Körpergefühl Wie steuert unser Gehirn Verhalten? • Willkür- und Zielbewegungen erfordern kortikale Kontrolle (primärer motorischer Kortex, supplementäres motorisches Areal, prämotorischer Kortex): – Auswahl und Zahl der beteiligten Muskeln – ausgeübte Muskelkraft und zeitliche Modulation – Bewegungsverlauf (Zielrichtung, Geschwindigkeit der Bewegung, Gelenkstellung) • Absteigende Bahnen von Kortex zum Rückenmark: – Pyramidenbahn (tractus corticospinalis; 60% Zellkörper im motorischen Kortex, 40% and. Kortex; insg. ca. 106 Fasern in das Rückenmark). – Extrapyramidale Bahnen (T. rubrospinalis, T. vestibulospinalis, T. reticulospinalis, T. tectospinalis): Kontrolle der Körperhaltung, Koordination des normalen Ablaufs (Harmonie) der Willkürmotorik und Absicherung gegen die Einflüsse der Schwerkraft. Aminerge Fasern aus dem Hirnstamm (z. B. Schmerz) • Absteigende Bahnen projizieren Kollaterale in Basalganglien und Kleinhirn: – „motorischen Programmgestaltung“, Erstellung der Zeitstruktur, „Feinkontrolle“ – Basalganglien wirken vorwiegend „dämpfend“: Lassen nur die vom Kortex „beabsichtigten“ Bewegungen ohne grössere Hemmungen zu, damit Akzentuierung der gewollten Bewegungen (wirken stabilisierend auf gewollte Bewegungsabläufe) – Kleinhirn wirkt vorwiegend „erregend“. Extrapyramidalen Bahnen Pyramidenbahnen Lernen und Gedächtnis Lernen ist der Gedächtnis-bildende Prozess Memory Declarative (explicit) Priming Nondeclarative (implicit) Nonassociative Associative (predictive) Sensitization Classical conditioning Habituation Operant conditioning Classical conditioning Assoziatives Lernen: Klassische Konditionierung (nicht-deklarativ) Klassisches Konditionieren Wo im Gehirn findet lernen statt? CSCS-US KonvergenzKonvergenz-Punkt! Wichtig! CS: conditioned stimulus US: unconditioned stimulus CR: conditioned response UR: unconditioned response Klassisches Konditionieren Aplysia californica (See-Hase) Neuronales Netzwerk, das dem Rückziehreflex der Kiemen bei der Schnecke Aplysia zugrunde liegt Gill withdrawal circuit US UR CS Klassisches Konditionieren Aplysia californica (See-Hase) Sensori-motorisches Netwerk: Mechanismus: Synaptische Fazilitation Was passiert am CS-US Konvergenzpunkt? Fazilitation von sensori-motorischen Synapsen Vorher: Nachher: Prä-Synapse Post-Synapse Spike Verbreiterung: Prä-synaptischer Mechanismus (To be) Conditioned schwache Reaktion stimulus Vor dem Lernen: CS löst nur schwache Reaktion aus US CS UR US CS Unconditioned stimulus Vor dem Lernen: der US löst starke Reaktion aus US Starke Reaktion CS im CS Neuron aber nur wenig Wirkung UR US CS Während des Lernens (Paarung) Starke Reaktion K Nach dem Lernen: der CS löst alleine eine starke Reaktion aus CR after conditioning US Starke Reaktion CS UR US CS Drosophila Lern- und Gedächtnis Mutanten Klassisches Konditionieren • • • • • Olfaktorisches Lernen in Drosophila Furchtlernen in Ratten und Mäusen Lidschlussreflex in Kaninchen PER Konditionierung bei Bienen usw. Mittlerweile sind die molekularen, neuronalen und Netzwerk-Mechanismen des klassischen Konditionierens sehr weit erforscht. Diese Mechanismen scheinen in allen Tierarten von der Schnecke bis zum Menschen prinzipiell gleich abzulaufen (evolutionär konserviert) Lehrbuch!!! Operantes Konditionieren (nicht deklarativ) Skinner-Box • Basalganglien (Striatum) • unbekannte molekulare Gundlagen (versch. v. KK!) • Interagiert mit klassischen Komponenten •Gewohnheitslernen / Drogenabhängigkeit Medial temporal lobe Medialer temporaler Lobus (deklaratives Lernen) Die Rolle des Hippokampus beim deklarativen Lernen Die Geschichte des Patienten HM (Brenda Milner) Patient H.M. (*1926) 1953: •Schwere anterograde Amnesie •Partielle retrograde Amnesie •Intaktes Kurzzeit-Gedächtnis •Intaktes motorisches Lernen: Motorisches Lernen des Patienten HM Normales motorisches Lernverhalten des Patienten HM Hippocampus: LTP Bewertungsneuron Hebb´sche Regel: Wenn immer ein Neuron ein anderes treibt und dieses zur gleichen Zeit aktiv ist verstärkt sich die Synapse zwischen diesen Neuronen. (Assoziation, Koinzidenzdetektion) Neuron des Hinweissignals Dies geschieht, wenn die die Bedingungen dafür geeignet sind (das Tier gestimmt ist, aufmerksam ist, …) Modulator Neuron Effektor Neuron Synaptische Phänomene der langzeitigen Plastizität: Langzeit-Potenzierung LTP Langzeit-Depression LTD Der NMDA Rezeptor ist ein molekularer Koinzidenzdetektor: nur Glu-Ausschüttung der Kanal öffnet sich bleibt aber durch Mg verstopft Glu-Ausschüttung und Depolarisation des postsynaptischen Neurons Ca2+ und Na+ strömen ein. Der intrazelluläre Anstieg des Ca2+ führt zu einer lang anhaltenden Verstärkung der Synapse (Langzeit-Potenzierung; LTP) NO ist ein gasförmigere Transmitter, der auch zur Präsynapse diffundiert (retrograder Transmitter) und dort über cGMP abhängige Prozesse zu einer anhaltenden Verstärkung der Transmitterausschüttung führt. metapotroper Glu-Rezeptor Langzeit-Depression LTD tritt ein wenn häufig die präsynaptische Seite aktiv ist zu einer Zeit wenn die postsynaptische Seite nicht aktiv ist bewertendes Neuron (US) Kurzzeit-Gedächtnis: Verstellung der synaptischen Übertragung mit den vorhandenen Strukturen und Molekülen Langzeit-Gedächtnis: spezifische Aktivierierung von Genen, Neubildung von Strukturen Adenylatcyclase Strukturelle Veränderungen