Kapitel 22: Die Entstehung der Arten
Werbung

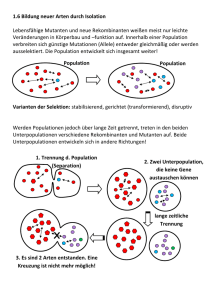
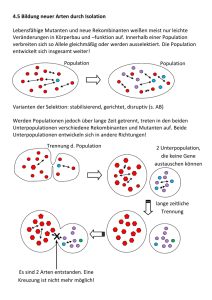
Kapitel 22: Die Entstehung der Arten (englische Version: Kap. 24) Der biologische Artbegriff betont die reproduktive Isolation Zwei Muster der Artbildung: Anagenese: aus einer Art entsteht eine neue Art Kladogenese oder Divergenz: aus einer Art entstehen zwei Schwesterarten Der biologische Artbegriff definiert eine Art als eine Population oder Gruppe von Populationen, deren Mitglieder sich unter natürlichen Bedingungen kreuzen können und dabei lebensfähige, fruchtbare Nachkommen hervorbringen. Eine biologische Art (Biospezies) stellt also die grösste Einheit einer Population, in der genetischer Austausch möglich und die genetisch von anderen solchen Populationen isoliert ist. Folglich ist jede Art von einer Fortpflanzungsbarriere umgeben, die ihre Eigenständigkeit als Art erhält, indem sie die genetische Vermischung mit anderen Arten verhindert. Mitglieder einer werden als conspezifisch bezeichnet, d. h. sie können sich untereinander – zumindest rein hypothetisch – fortpflanzen. Bei Arten, die sich nicht geschlechtlich fortpflanzen, ist der biologische Artbegriff unbrauchbar. Dazu gehören alle Prokaryoten, einige Protisten und Pilze sowie manche Pflanzen und Tiere. Solche Organismenarten bezeichnet man als Agamospezies. Ungeschlechtliche Lebewesen kann man nur Arten zuordnen, indem man sie zu Klonen mit gleicher Biochemie und Morphologie gruppiert. Der biologische Artbegriff ist auch ein schlechtes Kriterium für die Charakterisierung von ausgestorbenen Arten, deren Fossilien nur anhand ihrer Morphologie voneinander unterschieden werden können. Der biologische Artbegriff lässt sich ebenfalls nicht anwenden, wenn es trotz der Überlappung von Verbreitungsgebieten bestimmter Unterarten nicht zur Kreuzung kommt zwischen den Unterarten. Weil der biologische Artbegriff so unbefriedigend ist, sind andere Definitionen vorgeschlagen worden. Es ist jedoch unwahrscheinlich, dass jemals eine Definition gefunden wird, die alle Fälle abdeckt. Eine alternative zum Biospezieskonzept stellt die Einteilung aufgrund morphologischer Kriterien. Diese führen häufig zu derselben Arteneinteilung 1 Fortpflanzungsbarrieren trennen Arten Eine reproduktive Isolation von zwei Arten kann nicht mit einer geographischen Barriere gleichgesetzt werden, weil diese nicht von den Tieren selbst entspringt. Biologische Barrieren für die Reproduktion verhindern sogar eine Fortpflanzung, wenn sich die Verbreitungsgebiete zweier Arten überlappen. Fortpflanzungsbarrieren, welche die Genpools von Arten isolieren, lassen sich in präzygotische und postzygotische unterteilen. Präzygotische Barrieren Habitatisolation: Zwei arten, die in demselben Gebiet in unterschiedlichen Habitaten leben, treffen möglicherweise selten, wenn überhaupt, aufeinander, selbst wenn sie rein formal nicht geographisch isoliert sind. Bsp.: Zwei Arten von Strumpfbandnattern der Gattung Thamnophis kommen in demselben Gebiet vor, doch eine lebt hauptsächlich im Wasser, und die andere ist überwiegend Landbewohner. Verhaltensisolation: Artspezifische Signale und Verhaltensmuster durch welche eine Fortpflanzung überhaupt ermöglicht wird, sind eine wichtige Fortpflanzungsbarriere unter nahe verwandten Tierarten. Bsp.: - Männliche Leuchtkäfer verschiedener Arten machen sich bemerkbar, indem sie Lichtsignale geben, die Artspezifisch sind. Diese Signale werden also von den Weibchen derselben Art erwidert, wodurch die Männchen angelockt werden. - Balzrituale können auch eine Verhaltensisolation darstellen oder bei Vögeln können es unterschiedliche Gesänge sein Zeitliche Isolation: Die Paarung erfolgt in unterschiedliche Zeiten. Bsp.: Die geographischen Verbreitungsgebiete des Westlichen Fleckenskunks (Spilogale gracilis) und des Östlichen Fleckenskunks (Spilogale putorius) überlappen sich, aber diese beiden sehr ähnlichen Arten kreuzen sich nicht, weil sich S. gracilis im Spätsommer und S. putorius im zeitigen Frühjahr. Mechanische Isolation: Anatomische Unterschiede stellen auch eine Barriere dar. Bsp.: - Oft ist die Blütenanatomie an bestimmte Bestäuber angepasst, die den Pollen nur zwischen Pflanzen der gleichen Art übertragen. 2 - Spinnen besitzen hochkomplizierte Begattungsorgane, die bei artgleichen Partnern wie Schlüssel und Schloss zueinander passen. Gametische Isolation: Wenn die Befruchtung im weiblichen Körper zustande kommt (innere Befruchtung), kann es sein, dass die Spermien nicht überlebensfähig sind im weiblichen Körper einer anderen Art. Viele aquatische Tiere setzen ihre Gameten ins Wasser frei, wo es zur Befruchtung kommt (äussere Befruchtung). Dabei kommt eine Kreuzung selten zu Stande, weil sowohl Ei- als auch Samenzellen sich an artspezifischen Makromolekülen auf der Plasmamembran erkennen können. Postzygotische Barrieren Verringerte Lebensfähigkeit der Hybriden (Bastardsterblichkeit) Wenn präzygotische Barrieren überschritten werden, kommt es meistens nicht zu einer vollständigen Ausbildung des Embryos, weil infolge genetischer Unverträglichkeit (Inkompatibilität) zwischen den beiden Arten die Entwicklung der Hybride in irgendeinem Stadium abgebrochen wird. Die Bastarde, welche die Entwicklung doch beenden können, sind schwächlich. Verringerte Fruchtbarkeit der Hybriden (Bastardsterilität) Bei Kreuzungen zwischen verschiedenen Tierarten (z. B. Esel und Pferd) entstehen Bastarde, die völlig oder grösstenteils steril sind. → reproduktive Isolation beider Arten, da kein freier Genfluss möglich ist. Der Grund für die Sterilität ist die abnormale Gametenbildung der Hybriden, wenn die Chromosomen beider Elternarten sich in ihrer Anzahl oder Form unterscheiden. Hybridenzusammenbruch (Bastardzusammenbruch) Manche Arten sind im Stande, sich mit anderen Arten fortzupflanzen und dabei lebensfähige und fruchtbare Nachkommen zu produzieren. Wenn sich aber die Hybriden untereinander oder mit einer der Elternarten fortpflanzen, erzeugen sie schwache oder sterile Nachkommen. Introgression und Bastardschwärme Introgression: Ab und zu kommt es – trotz aller Barrieren – zu fruchtbaren und lebensfähigen Nachkommen zwischen nahe verwandten Arten, die fähig sind, sich erfolgreich mit einer der Elternarten zu paaren. → Erhöhung der genetischen Variabilität. Da aber die Introgression nur in seltenen Fällen passiert, kann man sagen, dass die Artenverschiedenheit erhalten bleibt. → Beide Genpools bleiben praktisch unversehrt. Durch Introgression können Bastardschwärme (Hybridschwärme) entstehen. Diese sind fertil und aufgrund ihrer Mischeigenschaften individuell sehr Anpassungsmöglichkeiten an unterschiedliche Standorte. 3 variabel. → Erweiterung der Geographische Separation kann zur Entstehung von Arten führen: Allopatrische Artbildung Die Artbildung kann man aufgrund der geographischen Beziehung einer neuen Art zu ihrer Ausgangsart in zwei Formen aufteilen. Das anfängliche Hindernis für einen Genfluss kann eine geographische Barriere sein, welche die Population physisch isoliert. Diese Form der Artbildung bezeichnet man als allopatrische Artbildung. Bei der zweiten Form der Artbildung wird eine Teilpopulation inmitten ihrer Ausgangspopulation reproduktiv isoliert; dies ist die sympatrische Artbildung. Populationen bezeichnet man als sympatrisch, wenn sich ihre Verbreitungsgebiete überlappen. Geographische Barrieren Welches Ausmass eine geographische Barriere erreichen muss, um allopatrische Populationen aufrechtzuerhalten, hängt von der Ausbreitungsgeschwindigkeit bzw. der Ausbreitungsfähigkeit der betreffenden Spezies. Bussarde können bspw. den Grand Canyon problemlos überfliegen, was für kleine Nagetiere eine unüberwindliche Barriere darstellt. Folglich leben auf beiden Seiten des Canyons gleiche Vogelarten, aber unterschiedliche Erdhörnchenarten. Bsp.: Der Death Valley bestand vor ca. 50'000 Jahren aus einem Netzwerk von Flüssen und Seen, das vor ungefähr 10'000 Jahren auszutrocknen begann. Vor etwa 4'000 Jahren wurde der Death Valley zur Wüste. Aus dem Netzwerk von Flüssen und Seen blieben nur noch vereinzelte Quellen. Zwischen den einzelnen Quellen variieren die Lebensbedingungen stark, wie z. B. Temperatur und Salzgehalt. Das hat dazu geführt, dass die dort lebenden Zahnkärpflinge der Gattung Cyprinodon sich zu unterschiedlichen Arten entwickelten als sich deren Verbreitungsgebiet durch die Trockenheit aufsplitterte. Bedingungen, die eine allopatrische Artbildung begünstigen Bei Populationen, die allopatrisch werden, besteht immer die Möglichkeit der Artbildung, da sich durch Mikroevolution die genetischen Unterschiede im Genpool anhäufen. Je kleiner eine Population ist, desto grösser sind die Chancen der Artbildung. Die geographische Isolation erfolgt in der Regel am Rande des Verbreitungsgebiets. Das periphere Isolat (Splitterpopulation) ist aus drei Gründen ein guter Kandidat für die Artbildung: 4 1. Periphere Isolate weisen wahrscheinlich schon vor der Isolierung Merkmalunterschiede, da sie wegen ihrer Lage extremen Lebensbedingungen ausgesetzt sind (Cline). Ist das periphere Isolat zudem eine kleine Population, dann ist der Gründereffekt praktisch schon vorprogrammiert. Gründereffekt: Eine Ursache der genetischen Drift, die auf die Kolonisierung eines neuen Lebensraumes durch eine Zahl von Individuen aus einer Ausgangspopulation, zurückzuführen ist. 2. Bis aus dem peripheren Isolat eine grosse Population entsteht, kommt es nach dem Zufallsprinzip zu Neumutationen und Kombinationen der Allele, die in ihrer Anpassungsfähigkeit neutral sind. Das führt zu einer Veränderung des Genpools, was wiederum zu einer geno- und phänotypischen Abwandlung führt. 3. Ihre Umwelt divergiert also bis zu einem gewissen Grad von derjenigen der Ausgangspopulation, da sie die Verbreitungsgrenze bewohnen. → abweichende Selektionsfaktoren, die meistens härter sind. Die Überlebenschancen eines peripheren Isolats sind gering, und nicht immer sind die Veränderungen gross genug für die Bildung einer neuen Art. Adaptive Radiation auf Inselketten Die Evolution vieler unterschiedlich angepasster Arten aus einem gemeinsamen Vorfahren bezeichnet man als adaptive Radiation. Ein gutes Beispiel für adaptive Radiation stellen die unterschiedlichen Arten von Darwinfinken auf den Galapagos-Inseln. Auf der vom südamerikanischen Festland abgedrifteten Inselkette entstand möglicherweise ein peripheres Isolat von Darwinfinken und daraus evolvierte eine neue Art. Diese neue Art besiedelte benachbarte Inseln, die weit genug waren, um die Entstehung von weiteren peripheren Isolaten zu ermöglichen. Auf diese Weise sind eine Vielzahl von neuen Arten entstanden, die z. T. zu den Ursprungsinseln zurückkehrten und mit den Ausgangspopulationen koexistieren und sogar neue Arten gebildet haben. Eine neue Art kann Verbreitungsgebiets inmitten einer des geographischen Ausgangsart entstehen: Sympatrische Artbildung Bei der sympatrischen Artbildung entsteht eine neue Art aus der Ausgangsart inmitten des Verbreitungsgebiets. Das ist darauf zurückzuführen, dass einzelne Individuen genetisch mutieren, sodass es zur reproduktiven Isolation kommt. Das kann innerhalb einer Generation passieren. Bsp.: Viele Pflanzenarten entstehen durch Zufälle bei der Zellteilung, die zu einem zusätzlichen Chromosomensatz führen, eine Mutationsart, die Polyploidie genannt wird. Als autopolyploid 5 bezeichnet man ein Individuum mit mehr als zwei Chromosomensätzen, die alle vom selben Organismus stammen. Durch Zufall wird bei der Zellteilung beispielsweise aus einer diploiden Pflanze (2n) eine tetraploide (4n). Diese Pflanze kann sich selbst bestäuben oder sich mit anderen tetraploiden Pflanzen kreuzen. Diese Mutante können sich aber nicht mit diploiden Pflanzen kreuzen. So kann eine postzygotische Barriere innerhalb einer Generation und im Extremfall nur von einem einzigen Individuum ausgehend zur reproduktiven Isolation führen. Eine andere Form der Polyploidie, die weitaus häufiger ist als die Autopolyploidie, bezeichnet man als Allopolyploidie. Allopolyploide Pflanzen entstehen, wenn sich zwei verschiedenen Arten von Pflanzen kreuzen und Chromosomen kombinieren. Diese Hybriden sind in der Regel steril, weil bei der Meiose der haploide Chromosomensatz der einen Art sich nicht zu Tetraden paaren kann mit dem haploiden Satz der anderen Art, weil die Chromosomensätze der Eltern nicht homolog sind. Trotzdem kommt es zur Reproduktion durch ungeschlechtliche Vermehrung (s. Abb. 22.10 b). Dafür gibt es zwei Mechanismen: - Bei der mitotischen Teilung verdoppelt sich die Chromosomenzahl durch Nondisjunction (Nichttrennen der Chromosomenpaare bei der Meiose bzw. der Schwesterchromatiden bei der Mitose). Auf diese Weise können Gameten entstehen, weil es zu jedem homologen Chromosom eine Kopie gibt, mit der sich zusammenlagern kann während der Meiose. → lebensfähigere, fertilere Hybride - Die Chromosomensätze der Gameten des einen Elternteils werden nicht reduziert. Bei der Verschmelzung der Gameten beider Elternteile entsteht also eine Hybride mit diploiden und haploiden Chromosomen (tripoloide Hybride). Nun kommt es zu einer Teilmeiose, indem ein weiterer reduzierter Gamet mit der triploiden Hybride verschmilzt. → lebensfähigere, fertilere Hybride Bei beiden Mechanismen entstehen neue Arten, deren Chromosomenzahl der Summe der Chromosomen beider Elternarten entspricht. Die Beziehung allopolyploide Pflanzen und den Elternarten bezeichnet man als Netzverwandtschaft. Manche Allopolyploide sind besonders widerstandsfähig, weil diese die besten Eigenschaften der Elternarten in sich vereinigen. Das wird heutzutage von den Genforschern ausgenützt, indem sie unter Einsatz von chemischen Substanzen, die eine meiotische Nondisjunction auslösen neue polyploide Formen hervorbringen. Sympatrische Artbildung kommt auch bei Tieren vor, die Mechanismen, die Nondisjunction hervorrufen, sind jedoch anders. Es kann dazu kommen, wenn durch Mutationen auf andere Ressourcen angewiesen sind als diejenigen der Elternpopulation. Bsp.: Feigenwespen legen ihre Eier in den Blütenständen der Feigen und bestäuben so deren Blüten. Jede Feigenart wird von einer bestimmten Wespenart bestäubt. Eine Mutation könnte einen Wechsel der Feigenart und somit die Abtrennung von der Elternpopulation zur Folge haben. Damit wären die Ausgangsbedingungen für 6 eine Auseinanderentwicklung erfüllt. Sympatrische Artbildung kann auch eine Folge von Polymorphismus verbunden mit assortativer Paarung1 sein. Allopatrische Artbildung ist im Tierreich weitaus häufiger als die sympatrische, während bei den Pflanzen beide Formen gleichwertig sind. Mit Hilfe der Populationsgenetik lässt sich die Artbildung erklären Artbildung durch adaptive Divergenz Wenn sich zwei Populationen an zwei Umwelten anpasst, häufen sich Unterschiede in deren Genpools. In diesem Prozess der allmählichen adaptiven Divergenz (Auseinanderentwicklung) zweier Genpools können u. a. – muss aber nicht sein – Fortpflanzungsbarrieren entstehen (prä- und postzygotische Barrieren). Das heisst, dass reproduktive Isolation eine sekundäre Folge der adaptiven Divergenz von Populationen ist. In einigen Fällen beruht reproduktive Isolation auf sexuelle Selektion. Das bedeutet, dass bei der Partnerwahl auf morphologische und / oder Verhaltenseigenschaften geachtet wird. Diese Art der Selektion steigert den Fortpflanzungserfolg innerhalb einer Population und ist keineswegs als Schutz gegen Kreuzungen mit anderen Populationen zu verstehen. In diesem Fall geht man von einem auf Erkennung beruhenden Artbegriff aus, was den biologischen Artbegriff präzisiert. Da ist also die reproduktive Isolation auch ein Nebenprodukt. Artbildung durch Verschiebungen in Anpassungsgipfeln Ein Anpassungsgebirge – auch adaptive Landschaft genannt – ist eine symbolische Landschaft mit verstreuten Gipfeln. Diese Gipfel werden Anpassungsgipfel genannt und stellen Genpools dar, in denen besonders viele Allelfrequenzen vorkommen, die eine optimierte Anpassung an die Umwelt ermöglichen. Bewegt sich der Genpool von einem Anpassungsgipfel zu einem anderen, durchläuft die Population eine Phase der geschwächten Fitness, was als Tal dargestellt wird. Die natürliche Selektion tendiert die Population bei einem bestimmten Anpassungsgipfel zu halten, obwohl rein theoretisch verschiedene Gipfel erreicht werden könnten. Im Fall einer Veränderung der Umwelt, 1 Eine Form der nichtzufälligen Partnerwahl, bei welcher die Geschlechtspartner in bestimmten phänotypischen Eigenschaften ähneln. 7 ändert sich das Anpassungsgebirge. Damit die Population überleben kann, muss diese durch Mikroevolution zu einem neuen Anpassungsgipfel gelangen. Gipfelverschiebungen sind nicht immer auf Umweltveränderungen zurückzuführen (adaptive Divergenz), sondern auch auf nichtadaptive Veränderungen in der genetischen Struktur einer Population. Bsp. für eine Gipfelverschiebung: Gründer-, Flaschenhalseffekt. Hybridzonen und der auf Kohäsion (Zusammenhalt) beruhende Artbegriff Wenn zwischen zwei verwandten allopatrischen Populationen nach einer bestimmten Zeitspanne, in der es zur Artbildung kam, die geographischen Barrieren wieder verschwinden, kann es sein, dass sich die zwei benachbarten Arten durch Introgression wieder fortpflanzen können. Wenn das nur dort geschieht, wo sich die Verbreitungsgebiete angrenzen bzw. überlappen, spricht man von einer Hybridzone. Die Hybridzone bleibt also stabil. In diesem Fall ist der biologische Artbegriff nicht ausreichend, um dieses Phänomen zu erklären, denn trotz Hybridisierung bleiben die Arten erhalten. Somit ist der auf Kohäsion beruhende Artbegriff eine weitere Alternative zum biologischen Artbegriff. Kohäsion könnte eine gemeinsame, verbesserte Anpassung an die Umwelt beinhalten, die für eine Art Charakteristisch ist. Durch Selektion wird also eine phänotypische Abwechslung verhindert bzw. auf einen Bereich beschränkt, der ein Übergang zwischen zwei Umwelten darstellt. Das Konzept des Punktualismus regte Forschungen über die Geschwindigkeit der Artbildung an Evolutionsstammbaum: bildliche Darstellung der Abstammung von Arten von altertümlichen Spezies, wobei jede neue Art ständig über lange Zeit evolviert und sich zu neuen Arten auseinanderentwickelt durch Anhäufung von Veränderungen der Allelfrequenzen im Genpool. Diese Theorie wird als Gradualismus bezeichnet. Fossile Befunde belegen aber, dass eine graduelle Evolution nicht so häufig vorkommt. Vielmehr ist ein plötzliches Auftauchen einer neuen Art durch eine relativ schnelle Auseinanderentwicklung von der Ausgangsart. Danach verändert sich jedoch diese geringfügig. Diese Theorie wird als Punktualismus bezeichnet. Obwohl die Veränderung einer Art einige Tausenden Generationen dauern kann, ist diese Zeitspanne verglichen mit der Überlebenszeit der Art sehr klein. Aus fossilen Befunden weiss man, dass eine Spezies wenige Millionen Jahre existiert, bevor sie ausstirbt, was aufzeigt, dass die Phase, 8 in der die Art die Umwandlung durchmacht nur ein Bruchteil der gesamten Existenz ausmacht. Das erklärt auch, wieso Paläontologen selten graduell evolvierte Fossilen entdecken. Diese Tendenz der morphologischen Stasis ist wahrscheinlich auf eine stabile bzw. sich nur geringfügig ändernde Umwelt während der Existenz einer bestimmten Art, was zu einer stabilisierenden Selektion führt. D. h., dass Umwandlungen des Genpools unterdrückt werden. Allerdings muss man sagen, dass Fossile kein guter Nachweis sind für den Punktualismus, da ja viele Veränderungen im Verhalten, in der inneren Morphologie, in der Physiologie oder auf molekularer Ebene auftreten. Zur Zeit sind sich die Forscher aber nicht einig über die relative Bedeutung von gradueller und punktueller Evolution. Für genaue Aussagen weitere eingehende morphologische Studien von gut erhaltenen Fossilen notwendig. 9