Die Sehkaskade
Werbung

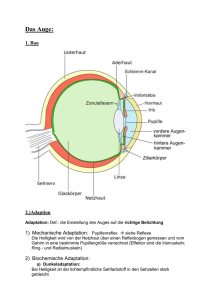
Die Sehkaskade Veranstaltung: Seminar: Visuelle Wahrnehmung Seminarleiter: Dipl. Psych. Kai Hamburger Referent: Morten Kaletsch Datum: 03.05.2005 THEMA: Photorezeptoren und Sehpigmente - Die Sehkaskade Retinale Verarbeitung Wie wird aus Licht ein elektrisches Signal, dass ans Gehirn weitergeleitet wird? Die Sehkaskade • Absorbtion schon eines einzelnen Lichtteilchens durch einen Photorezeptor im Auge löst eine Kaskade von Prozessen aus, die in einem elektrischen Signal endet • In den Photorezeptoren fängt der Prozess an Was sind Photorezeptoren und wo befinden sie sich? Die Sehkaskade • 2 Arten von Photorezeptoren in der Netzhaut: Zapfen (6 Millionen) und Stäbchen (100 – 120 Millionen) • Zapfen für Farbensehen bei hellem Licht • Stäbchen für Hell-Dunkel-, od. Dämmerungssehen • Stäbchen sind ca. 20-mal lichtempfindlicher als Zapfen Das Stäbchen Außensegment - enthält etwa 800 Membranscheiben, sog. Disks Innensegment -beherbergt Organellen, welche, die bei der Photorezeption benötigten Moleküle bereitstellen Außensegment mit Disks die Sehpigmente enthalten Die Sehkaskade • Photorezeptoren bestehen aus 2 Segmenten • Außensegmente enthalten gestapelte Disks (ca. 800 pro Stäbchen) • Sehpigmente (Rhodopsin) in den Disks absorbieren das Licht • Rhodopsin besteht aus dem Protein Opsin und dem lichtempfindlichen Molekül Retinal Wie wird das Signal entlang der Netzhautzellen und schließlich zum Gehirn weitergeleitet? Zellen der Netzhaut Ganglionzell-Schicht Übertragung der visuellen Information zum Gehirn Bipolarzell-Schicht Informationen von Fotorezeptoren zu Ganglionzellen Fotorezeptor-Schicht Zapfen und Stäbchen Außensegmente enthalten Sehpigmente In der Dunkelheit Geöffnete Natrium (Na+ u. Ca2+) Ionen-Kanäle in der Plasmamembran sorgen für eine elektrische Potentialdifferenz. Glutamat wird an den Synapsen freigesetzt In der Dunkelheit Glutamat hyperpolarisiert (hemmt) die Bipolarzelle, die somit kein Signal weiterleitet Geöffnete Natrium (Na+ u. Ca2+) Ionen-Kanäle in der Plasmamembran sorgen für eine elektrische Potentialdifferenz. Glutamat wird an den Synapsen freigesetzt In der Dunkelheit Niedrige Feuerrate der Ganglionzelle Glutamat hyperpolarisiert (hemmt) die Bipolarzelle, die somit kein Signal weiterleitet Geöffnete Natrium (Na+ u. Ca2+) Ionen-Kanäle in der Plasmamembran sorgen für eine elektrische Potentialdifferenz. Glutamat wird an den Synapsen freigesetzt Die Erregung bei Lichteinfall Licht Lichteinfall schließt Ionenkanäle; Na+ u. Ca2+ können nicht mehr eindringen Hyperpolarisation Freisetzung von Glutamat unterbunden Die Erregung bei Lichteinfall Licht Hemmung der Bipolarzelle ist aufgehoben. Neurotransmitter werden freigesetzt Lichteinfall schließt Ionenkanäle; Na+ u. Ca2+ können nicht mehr eindringen Hyperpolarisation Freisetzung von Glutamat unterbunden Die Erregung bei Lichteinfall Licht Membran der Ganglionzelle wird depolarisiert, was Aktionspotentiale der Ganglionzelle auslöst Hemmung der Bipolarzelle ist aufgehoben. Neurotransmitter werden freigesetzt Lichteinfall schließt Ionenkanäle; Na+ u. Ca2+ können nicht mehr eindringen Hyperpolarisation Freisetzung von Glutamat unterbunden Die Erregung bei Lichteinfall Registrierung von Aktionspotentialen zum Gehirn depolarisiertes Membranpotential hyperpolarisiertes Membranpotential Reiz Wie werden die Ionenkanäle im Außensegment durch Lichteinfall geschlossen? Das Stäbchen Außensegment - enthält etwa 2000 Membranscheiben, sog. Disks Innensegment -beherbergt Organellen, welche, die bei der Photorezeption benötigten Molekühle bereitstellen Außensegment mit Disks Ionenkanäle in der Segmentmembran • Im Dunkeln werden die Ionenkanäle von cyclischem Guanosin3´,5´-monophosphat (cGMP) offen gehalten • GMP gehört zur Sorte der Nucleotiden, die als Untereinheiten der Ribonucleinsäure (RNA) auftreten Cyclisches Guanosin-3´,5´-monophosphat Phosphor Wasserstoff Sauerstoff Kohlenstoff Stickstoff • Der namesgebende Ring des cyclischen GMP enthält ein Phosphoratom in einer sog. Phosphordiesterbindung • In seiner intakten Form hält es die Ionen-Kanäle offen Aktivierung der Phosphodiesterase • Lichteinfall setzt Prozesse in Gang die den cyclischen GMP Ring sprengen • Die Ionenkanäle schließen sich • Welche Prozesse gehen voraus? Fangen wir bei der Absorbtion eines einzelnen Lichtquants an! Stäbchen-Rhodopsin - Sehfarbstoff Opsin Lipid-Doppelschicht 11-cis-Retinal Retinal • 11-cis-Retinal ändert mit Absorbtion eines Photons seine Gestalt • Absorbtion bewirkt Drehung der Kette zwischen Kohlenstoffatomen • Kette richtet sich gerade aus, sie streckt sich alltrans-Retinal • Durch die Streckung ist nicht mehr genug Platz innerhalb des Opsins und es muss seine räumliche Struktur verändern • Aus inaktivem Rhodopsin (R) wird aktives (R*) • Das aktive Rhodopsin aktiviert nun ein Protein namens Transducin • Transducin besteht aus drei Proteinuntereinheiten, Alpha, Beta und Gamma • Transducin bindet Guanosindiphosphat (GDP) und Guanosintriphosphat (GTP) • Durch R* wird GDP an der αUntereinheit gegen GTP ausgetauscht • Transducin zerfällt in die TαGTP- und Tβγ-Untereinheit • Ein einziges Rhodopsin Molekül produziert Hunderte aktive Tα-GTP-Komplexe (T*) • Aktiviertes Transducin wandert zum nächsten Enzym der Kaskade, zur PDE (cGMP-Phosphodiesterase) • PDE besteht aus drei – wieder mit Alpha, Beta und Gamma bezeichneten – Einheiten • Die Gamma-Untereinheit fungiert als Inhibitor der PDE, im Ruhezustand hemmt sie die enzymatische Aktivität der PDE α γ β • Aktiviertes Transducin bindet die Gamma-Untereinheit der Phosphodiesterase und hebt somit diese Blockade auf • Ein aktiviertes Transducin aktiviert eine Phosphodiesterase (1:1) α γ β • Aktiviertes Transducin bindet die Gamma-Untereinheit der Phosphodiesterase und hebt somit diese Blockade auf • Ein aktiviertes Transducin aktiviert eine Phosphodiesterase (1:1) • Aktivierte PDE spaltet bis zu 2000 cGMP-Moleküle pro Sekunde α γ β Aktivierung der Phosphodiesterase • Phosphodiesterase sprengt den Ring des cyclischen GMP (es baut ein Wassermolekühl in ihn ein, hydrolysiert ihn also). Aus cyclischem Guanosin-3´,5´-monophosphat wird Guanosin-5´monophosphat • Die Ionenkanäle schließen sich. Die elektrischen Eigenschaften der Membran ändern sich, so dass ein Nervenimpuls entsteht. Aber wir wollen ja nicht nur einmal sehen! Abschalten der Enzymkaskade und Regeneration • Chemische Schaltuhr in der α-Einheit inaktiviert Transducin durch Umwandlung des GTP in GDP • Transducin gibt die hemmende GammaEinheit der Phosphodiesterase frei • Die Gamma-Einheit heftet sich an die Phosphodiesterase und stellt den Ruhezustand wieder her • Die Alpha-Untereinheit des Transducins vereinigt sich ihrerseits mit dem BetaGamma-Komplex • Somit liegt Transducin wieder in seiner GDPForm vor • Rhodopsin wird durch ein Enzym, eine spezifische Kinase inaktiviert. • Die Bindung von Transducin wird blockiert und das System in den Dunkelzustand versetzt Zusammenfassung • Stäbchen haben ein Höchstmaß an Empfindlichkeit erreicht, es kann von einem einzigen Photon – der kleinstmöglichen Lichtmenge – erregt werden • 1 Rhodopsin-Molekül aktiviert bis zu 3000 Transducin-Moleküle, jedes davon eine Phosphodiesterase • Jede PDE kann bis zu 2000 cGMP-Moleküle pro Sekunde sprengen Zusammenfassung • Theoretisch können durch ein Photon 6 000 000 cGMP-Moleküle inaktiviert werden. • Na+ und Ca2+-Ionen-Einstrom in die Zelle wird verhindert. • Die Zelle hyperpolarisiert • Dies führt zum Verschluss der spannungsgesteuerten Ca2+-Ionenkanäle in der Synapse • Es wird kein Transmitter (Glutamat) freigesetzt Zusammenfassung • Die nachfolgende Zelle Bipolarzelle ist nicht mehr gehemmt und setzt ihrerseits Transmitter frei • Ganglionzelle wird depolarisiert • Ein Aktionspotential entsteht • Es erfolgt ein Signal ans Gehirn • Das war‘s! Fragen über Fragen 1. Wo werden die Aktionspotentiale ausgelöst? • Ganglienzelle 2. In welchem Teil der Photorezeptoren wird das Licht absorbiert? • Außensegment Rhodopsin 11-cis-Retinal 3. Durch welche zwei Prozesse wird die Wirkung des Lichteinfalls verstärkt? • • Rhodopsin aktiviert bis zu 3000 Transducin-Moleküle Phosphodiesterase sprengt bis zu 2000 cGMP-Moleküle