Teil VIII - Tired Joe
Werbung

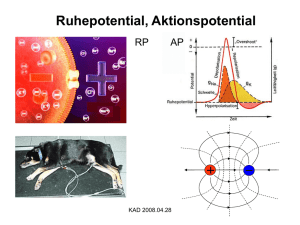
Neurophysiologie Grundstrukturen und Leistungen des Nervensystems 1. Rezeptoren (Sinneszellen) Einteilung der Rezeptoren nach Art des Reizes: Optischer Reiz Photorezeptoren (Lichtsinneszellen im Auge) Mechanischer Reiz Mechanorezeptoren (Tastkörperchen der Hand) Thermischer Reiz Termorezeptoren (Wärme-, Kälterezeptoren der Haut) Chemischer Reiz Chemorezeptoren (Geruchs- Geschmackszellen) Elektrischer Reiz Elektrorezeptoren z.B. bei Haien Die Reizart kann auch adäquat oder inadäquat sein! Beispiel Photorezeptoren: Photorezeptoren reagieren auch auf Druck „Sternchen sehen“ Farbwahrnehmung 2. Einteilung der Rezeptoren nach der Lage im Organismus: Exterorezeptoren: nach außen zur Umwelt gerichtet, Beispiel: Photorezeptoren Enterorezeptoren: im Körperinneren, Infos über den Innenzustand z.B. CO2-Rezeptor im arteriellen Blut (Halsschlagader) Neuronen und Synapsen Baustein des Nervensystems: Nervenzelle = Neuron = Ganglienzelle Einteilung nach der Zahl der Fortsätze Multi-, bi-, monopolares Neuron Einteilung nach Lage der Zellkörper bzw. Leitungsrichtung der Erregung a) Rezeptorneuron: Zellkörper in der Peripherie, Axon leitet zum ZNS = afferentes Neuron (hinführendes Neuron, z.B. Finger) b) Motoneuron: Zellkörper im Rückenmark, Axon leitet weg vom ZNS = efferentes Neuron (wegbzw. forttragendes Neuron) c) Schalt- oder Interneurone: Gesamtes Neuron innerhalb des ZNS (Hirn, Rückenmark) zeichnen können Seite 208 © Florian Zeller 08 / 09 Bau eines Motoneurons: Die Axone sind vom Gliazellen umgeben, die das Axon umhüllen, stützen und schützen, es ernähren. Bei den markhaltigen = myelinisierten Nervenfasern der Wirbeltiere heißt die Gliazelle Schwann´sche Zelle. Sie ist ganz platt und mehrfach um das Axon gewickelt (siehe Abbildung), so dass viele Membranen aufeinander zu liegen kommen = Myelinscheide (Myelin ist das Material, z.B. auch weiße Substanz im ZNS im Gegensatz zu den „kleinen grauen“ Zellkörpern). Sie isoliert das Axon elektrisch, daher hohe Geschwindigkeit der Erregungsleitung möglich!! Bei unmyelinisierten = marklosen Nervenfasern ist die Gliazelle nur eine einfache Hülle für 1 oder mehrere Axone. Leitungsgeschwindigkeit gering. Vorkommen bei Wirbellosen. Formenvielfalt von Neuronen Nur zum anschauen! Seite 209 © Florian Zeller 08 / 09 Synapsen Neuronale Versschaltung bzw. Möglichkeiten von Synapsen a) Zentrale Synapsen zwischen Nervenzellen - Axo - somatische Synapse , d.h. Synapse zwischen Axon und Soma - Axo – dentritische Synapse, d.h. Synapse zwischen Axon und Dentrit b) Synapse zwischen Axon und Muskel = neuromuskuläre Synapse = motorische Endplatte c) Synapse zwischen Axon und Drüsenzellen Neuromuskuläre Synapse = motorische Endplatte zeichnen können!! Seite 210 © Florian Zeller 08 / 09 Die unerregte Nervenfaser - Ruhepotential Versuchsanordnung zur Messung des Ruhepotentials Extracelluläre Ableitung, Potentialdifferenz = 0 intracelluläre Ableitung Potentialdifferenz - 50 mV bis - 90 mV bei Wirbeltieren = RUHEPOTENTIAL Bei Wirbeltiernervenzellen misst man eine Potentialdifferenz von ca. -70 mV, d.h. Zellinneres negativ im Vergleich zu Außen. Weitere Beobachtung: Die Potentialdifferenz tritt bereits beim Durchdringen der Membran auf, sie ist also an der Membran angelegt deswegen spricht man von Membranpotential. Die Potentialdifferenz ist an allen Punkten der Ruhenden Nervenzelle messbar! Darstellung im Diagramm: Seite 211 © Florian Zeller 08 / 09 Entstehung (Aufbau) des Ruhepotentials Untersuchungen über die Art der Ionen innen und außen an der Axonmembran ergaben folgende Ergebnisse: 1. Konzentrationsunterschiede (ungleiche Ionenverteilung) großer Buchstabe = viel vorhanden kleiner Buchstabe = wenig vorhanden sehr wichtig und zeichnen können! 2. Selektive Permeabilität Die Nervenzellmembran ist für diese Ionen unterschiedlich durchlässig. Selektive Permeabilität für K+ = 1 Cl- = 0,1 Na+ = 0,01 A- = 0 Vorstellung: in der Membran sind kleine Poren, die die leinen K+-Ionen durchtreten lassen, auf die K+-Ionen wirken zwei Kräfte: a) Bestreben nach Konzentrationsausgleich: wegen guter Permeabilität diffundieren sie nach außen b) Die nun zunehmend unkompensierte negative Ladung (Org- !!) innen hält weitere K+ durch elektrostatische Anziehung zurück, es kehren sogar manche K+ nach innen zurück, so dass es nicht zu einem Konzentrationsausgleich der Ionen kommt, sondern zu einem GGW-Zustand der treibenden Kräfte! D.h. bei einer bestimmten Spannung stellt sich ein GGW ein zwischen Konzentrationsgefälle und Potentialgefälle, siehe Abbildung! Das Ruhepotential (RP) ist ein „Kaliumpotential“ Seite 212 © Florian Zeller 08 / 09 Zum Thema Leckströme Die Nervenzellmembran ist etwas durchlässig für Natriumkationen und Chloridionen. Folgen: - Geringer Chloridioneneinstrom stützt das Membranpotential. Ständiges Einsickern von Natriumkationen ins Zellinnere (aufgrund des Konzentrationsgefälles und des Potentialgefälles) müsste eigentlich das Membranpotential schmälern. Kaliumionen können leichter nach außen treten, das Ruhepotential müsste irgendwann zusammenbrechen. Dies ist jedoch am intakten Axon nicht der Fall !! Ursache: gegenläufig arbeitende Natrium-Kalium-Ionenpumpe, ein aktiver Ionentransportmechanismus. Die Pumpe ist eine Membranstruktur , die unter Energieverbrauch (ATP-Verbrauch) Natriumionen aus dem zellinneren gegen Kaliumionen aus dem Außenmedium austauscht. Der Transport erfolgt gegen den jeweiligen Konzentrationsgradienten, also „bergauf“. Die erregte Nervenfaser – Auslösung und Entstehung von Aktionspotentialen Prinzip: Überschwelliger Reiz (gereizte Sinneszelle, Stromstoß) Erregung der Nervenzelle Folge: Membran wird kurzfristig umgepolt, die Umpolungswelle Pflanzt sich über das Axon fort. Diese Potentialänderung bezeichnet man als AKTIONSPOTENTIAL Seite 213 © Florian Zeller 08 / 09 Um Näheres über die Auslösebedingungen und die charakteristischen Eigenschaften eines Aktionspotentials zu erfahren, verwendet man die Versuchsanordnung zur gleichzeitigen intrazellulären Reizung und Ableitung von einem Axon. Auslösung von Aktionspotentialen 1. 2. 3. 4. 5. & 6. Bei nicht aktivem Reizgerät erfasst das Messgerät das Ruhepotential von – 70 mV. Ist die Reizelektrode mit dem negativen Pol des Reizgerätes verbunden, so wird dem Axoninneren eine gewisse Menge negativer Ladung zugeführt. Diese bewirkt eine entsprechende Verschiebung des Membranpotentials in den Bereich stärker negativer Werte. Die „Antwort“ der Membran ist passiv. Diese Verstärkung des Ruhepotentials wird als Hyperpolarisation bezeichnet. Ebenfalls passive Antwort; die Größe der Hyperpolarisation ist der Reizspannung proportional. Ist die Reizelektrode mit dem positiven Pol des Reizgerätes verbunden, so wird dem Axoninneren eine gewisse menge positiver Ladung zugeführt. Diese bewirkt eine entsprechende Verschiebung des Membranpotentials in den Bereich weniger stark negativer Werte. Die „Antwort“ der Membran ist wie bei 2. und 3. passiv. Diese Abschwächung des Ruhepotentials wird als Depolarisation bezeichnet. Ebenfalls passive Antwort; die Größe der Depolarisation ist der Reizspannung proportional. Seite 214 © Florian Zeller 08 / 09 Ab einer bestimmten Reizschwelle kommt es zur Auslösung von Aktionspotentialen. Die „Antwort“ der Membran ist aktiv. Dabei gilt die „Alles-oder-Nichts-Regel“. Sie gesagt, dass sämtliche überschwelligen Reize die Bildung eines Aktionspotentials bewirken, sämtliche Reize unterhalb der Schwelle hingegen nicht. Das Aktionspotential läuft in immer gleicher, starrer Weise ab. 10. Am Ende jedes Aktionspotentials kann es zu sog. Nachpotentialen kommen. 7. – 9. Also: Nur ein überschwelliger Reiz bewirkt eine überproportionale Depolarisation (aktive „Antwort“) der Axonmembran, und damit eine Potentialumkehr: Das Aktionspotential schnellt auf + 30 mV hoch. Die Höhe (+30 mV) des Aktionspotentials ist unabhängig von der Größe des überschwelligen Reizes Alles-oder-Nichts-Signal !!! Nur ein Aktionspotential wird über das Axon weitergeleitet !! Die Phasen eines Aktionspotentials Depolarisation: Sehr rasche positive Potentialänderung in 0,2 – 0,5 ms, negative Ruheladung geht verloren; positiver Anteil des Aktionspotential = Überschuss. Repolarisation: Normale Polarisation der Zellmembran wird wieder hergestellt, Prozess verläuft gegen Ende langsamer. Nachpotentiale: Bei der Rückkehr des Potentials von der Aktionspotentialspitze, schwingt es über den normalen Wert des Ruhepotential hinaus, erst nach hyper- & depolarisierten Nachpotential wird der Ruhewert erreicht. Ein Aktionspotential ist ein für jede Nervenzelle konstanter Ablauf von Depolarisation und Repolarisation, der immer dann selbsttätig auftritt, wenn die Membran über den Schwellenwert hinaus depolarisiert wird. Nachpotentiale: Bei der Rückkehr des Aktionspotentials von der Spitze stellen sich noch „Nachschwingungen“ ein, danach wird der Wert des Ruhepotentials wieder erreicht! Hinweis: Wenn Sie die Phasen eines Aktionspotentials skizzieren, achten Sie darauf, dass Sie die Depolarisation steiler zeichnen als die Repolarisation!! Seite 215 © Florian Zeller 08 / 09 Refraktärzeiten Kommen zwei Reize zeitlich knapp hintereinander, so beobachtet man Refraktärzeiten. Absolute Refraktärzeit: Bei sehr enger zeitlicher Aufeinanderfolge (2 ms und weniger) löst der eigentlich überschwellige zweite Reiz kein Aktionspotential aus. Relative Refraktärzeit: In der Zeitspanne zwischen 5 und 2 ms nach der Auslösung eines Aktionspotentials Kann nur ein abgeschwächtes Aktionspotential ausgelöst werden. Die ursprüngliche Ionenverteilung wird durch die Na-K-Pumpe wiederhergestellt. Hinweis: Die Zahl der Ionen, die während eines Aktionspotentials die Membran passieren ist im Vergleich zur Gesamtzahl relativ gering. Deshalb können auch bei blockierter Na-K-Pumpe noch ca. 1000 Aktionspotentiale ausgelöst werden. Die Na-K-Pumpe hat also keinen Einfluss auf das Aktionspotential. Alles-oder-Nichts-Regel: Alle überschwelligen Reize lösen ein Aktionspotential aus, das für eine bestimmte Nervenzelle immer die gleiche Größe besitzt. Verschlüsselung der Reizstärke: Starke Reize lösen pro Zeiteinheit eine größere Zahl von Aktionspotentialen aus als schwache Reize. „Die Reizstärke ist durch die Frequenz der Aktionspotentiale codiert“. (max. Frequenz ca. 500 Aktionspotentiale pro Sekunde, begrenzt durch Refraktärzeit) Seite 216 © Florian Zeller 08 / 09 Die Rolle der Ionen beim Aktionspotential Erinnerung: Das Ruhepotential ist ein „Kaliumpotential“ Das Aktionspotential ist ein „Natriumpotential“! Sie müssen jetzt wissen, dass die Nervenzellmembran neben den stets offenen Kaliumionenporen auch spannungsgesteuerte Ionenporen (Ionenkanäle) für Natriumkationen enthält !!! Und auch spannungsgesteuerte K+-Poren ! Was passiert bei der Depolarisationsphase? Ein überschwelliger Reiz depolarisiert die Membran über den Schwellenwert hinaus, Spannungsgesteuerte Natriumionenporen öffnen sich schlagartig, Natriumkationen strömen schlagartig in das Axon ein (wegen Konzentrationsgefälle und Potentialgefälle (innen - )), führt zur Potentialumkehr an der Membran, die Membraninnenseite wird positiv geladen. Bei +30 mV schließen sich die Natriumionenkanäle. (Ein Ausgleich der Natriumionenkonzentration auf beiden Seiten der Membran wird dabei nicht erreicht.) Jetzt setzt die Repolarisation ein: Spannungsgesteuerte K+-Poren Kaliumionen diffundieren verstärkt aus der Zelle durch die Membran, (wegen Konzentrationsgefälle und negativer Ladung außen), bis das Ruhepotential (nach den Schwankungen der Nachpotentiale) wieder hergestellt ist. Tatsächlich ist experimentell eine unterschiedliche Membranleitfähigkeit für die Ionen während des Aktionspotentials messbar. Die Dauer eines Aktionspotentials (Depolarisation, Umpolung, Repolarisation, Nachpotential) beträgt 4 – 5 ms. Während der Depolarisation und der Repolarisation (ca. 2 ms) kann auch durch einen überschwelligen Reiz kein weiteres Aktionspotential ausgelöst werden (absolute Refraktärphase). Der Natriumionenporen-Öffnungsmechanismus ist ganz einfach nicht funktionsfähig. Während der relativen Refraktärzeit (von ca. 2 – 5 ms) ist sowohl die Natriumionen- als auch die Kaliumionenpermeabilität gebremst. Seite 217 © Florian Zeller 08 / 09 Zum Anschauen ! Ionenverteilung nach einem Aktionspotential mehr Na+ innen mehr K+ außen als ursprünglich Doppelte Aufgabe der K+/Na+ - Pumpe Sie transportiert die während eines Aktionspotentials eingeströmten Na+ wieder nach außen und die K+ nach innen. Mit dem PORENMODELL lassen sich weitere Sachverhalte für die Auslösung von Aktionspotentialen erklärt. Alles-oder-Nichts-Regel unterschwelliger Reiz: Zahl der geöffneten Na+ - Poren reicht nicht aus, um Entladung der Membran zu bewirken. überschwelliger Reiz: schlagartige Öffnung aller Na+ - Poren konstante Amplitudenhöhe von Aktionspotential. Eine noch höhere Reizstärke kann keine Amplitudenerhöhung bewirken, da die Zahl der Na+ - Poren begrenzt ist. Refraktärzeiten absolute Refraktärzeit: Nach einem ausgelösten Aktionspotential ist der Na+ - Porenöffnungsmechanismus 1 – 2 ms nicht funktionsfähig. relative Refraktärzeit: Weitere 3 ms ist die Erregbarkeit der Nervenfaser noch herabgesetzt. Nur bei erhöhter Reizschwelle öffnen aller Poren. Seite 218 © Florian Zeller 08 / 09 Latenzzeit ≠ Reflektärzeit Weiterleitung von Aktionspotentialen Erregungsleitung in einer marklosen Nervenfaser skizzieren können (1) An der Stelle, an der gerade ein Aktionspotential auftritt, wird das Membranpotential durch einen starken Einstrom von Na+ - Ionen umgepolt. (2) An beiden Seiten der Membran stoßen damit positive und negative Ladungen direkt aufeinander. (3) Da sich entgegengesetzte Ladungen anziehen, kommt es zu einer Verschiebung der Ionen in Längsrichtung des Axons. Diese Ionenströme („Kreisströmchen“) bewirken eine Veränderung des Membranpotentials in der unmittelbaren Nachbarschaft der gerade umgepolten Stelle. (4) Die Nachbarstelle wird auf diese Weise überschwellig depolarisiert, und es bildet sich an ihr ein Aktionspotential aus. (5) An der ursprünglich umgepolten Stelle stellt sich wieder das Ruhepotential ein. Seite 219 © Florian Zeller 08 / 09 (6 & 7) Ionenströme bewirken wiederum die überschwellige Depolarisation der Membran in der unmittelbaren Nachbarschaft der aktuell umgepolten Stelle. (8) An der Nachbarstelle entsteht erneut ein Aktionspotential, das in der beschriebenen Form auf die neue Nachbarstelle einwirkt. Das Aktionspotential breitet sich unter Beibehaltung seiner vollen Stärke entlang der gesamten Nervenfaser aus (kontinuierliche Erregungsleitung). Die Leitungsgeschwindigkeit (ca. 10 𝑚 𝑠 ) ist abhängig von der Dicke des Axons. Aufgabe: In welcher Richtung kann die Erregung a) bei künstlicher Reizung eines Axons laufen?(vom Soma zum Endknöpfchen) b) im Körper nehmen? (in beide) Erregungsleitung in einer markhaltigen Nervenfaser Bei der markhaltigen Nervenfaser sind weite Bereiche des Axons durch eine Markscheide isoliert. Nur an einem RANVIER-Schnürring können sich Ionen durch die Membran bewegen. (1) An der Einschnürung, an der gerade ein Aktionspotential auftritt, wird das Membranpotential durch den Einstrom von Na+ - Ionen umgepolt. (2) Die Ionenströme (Kreisströmchen) können wegen der isolierend wirkenden Markscheide erst am nächsten Schnürring eine Depolarisation bewirken. (3) Dort wird ein Aktionspotential gebildet, während sich an der ursprünglich umgepolten Stelle wieder das Ruhepotential einstellt. (4) Die Ionenströme führen dann zur Weitergabe des Aktionspotentials an den nächstfolgenden Schnürring. Die Erregung springt gleichermaßen von Schnürring zu Schnürring. Man spricht deshalb auch von saltatorischer Erregungsleitung. Die Leitungsgeschwindigkeit beträgt ca. 100 𝑚 𝑠. Aufgabe: Geben Sie drei Gründe an, weshalb sich markhaltige Nervenfasern gegenüber marklosen Nervenfasern im Verlauf der Evolution durchgesetzt haben? - Materialersparnis bei gleicher Leistung (Platzersparnis) - ATP-Verbrauch geringer weniger Na+ / K+ - Pumpen werden benötigt (Energieersparnis) - Schneller (Zeitersparnis) Seite 220 © Florian Zeller 08 / 09 Vorgänge an der neuromuskulären Synapse, chemische Erregungsübertragung durch Transmitter 1. Ein Aktionspotential kommt an der präsynaptischen Membran an, löst über Calciumioneneinstrom die Ausschüttung des Transmitterstoffs Acetylcholin, das in den synaptischen Bläschen gespeichert ist, aus. 2. Die Acetylcholinmoleküle diffundieren über den synaptischen Spalt (Dauer: max. 1 ms). 3. Die Acetylcholinmoleküle setzen sich an der subsynaptischen Membran weist wie die Membran der Nervenzelle ein Ruhepotential auf. 4. Durch eine Konformationsänderung der Rezeptormoleküle kommt es zu einer Permeabilitätsänderung der Membran für Natrium- und Kaliumkationen. 5. Als Folge wird die subsynaptische Membran depolarisiert, es entsteht ein sog. „Endplattenpotential“, das sich in den postsynaptischen Membranbereich der Muskelfasermembran hinein ausbreitet und letzendlich die Kontraktion einer Muskelfaser auslöst. 6. Ein an der subsynaptischen Membran befindliches Enzym – die Cholinesterase – löst die Acetylcholinmoleküle augenblicklich wieder ab, indem es sie in Cholin und Essigsäure spaltet. 7. Diese Moleküle diffundieren zur präsynaptischen Membran zurück, werden ins Endknöpfchen aufgenommen, wieder zu Acetylcholin verbunden und in die synaptischen Bläschen verpackt! Seite 221 © Florian Zeller 08 / 09 Störung der Synapsenfunktion durch Gifte Seite 222 © Florian Zeller 08 / 09 Beeinflussung der synaptischen Erregungsübertragung 1. 2. Hemmung der Erregungsübertragung durch Giftstoffe Hemmstoff Angriffsort Wirkungsart Botulinus-Toxin präsynaptische Membran Hemmt die Acetylcholinfreisetzung Blockade der Erregung Lähmung Curare und Derivate subsynaptische Membran Besetzt Ach-Rezeptoren, ohne eine Erregung auszulösen, kann nicht von Enzymen gespalten werden Lähmung E 606 Sarin Acetylcholinesterase Hemmung der Acetylcholinesterase, erst Erregung Krämpfe, dann Lähmung (Kompetitiver Hemmstoff) Wirkung von Psychopharmaka und Drogen Substanzen, die die Aktivität des Nervensystems, insbesondere des Gehirns beeinflussen. Sie wirken dadurch auf psychische Prozesse ein. (nur zum anschauen) Seite 223 © Florian Zeller 08 / 09 Das Gehirn (Wirbeltiere) Bildung der Neurula mit Verschluss des Neuralrohres. V = VorderM = MittelH = Hinterhirnbläschen Die Entwicklung des menschlichen Gehrins Seite 224 © Florian Zeller 08 / 09 Seite 225 © Florian Zeller 08 / 09 Längsschnitt durch das Gehirn des Menschen 1 = Großhirn = Vorderhirn 2 = Balken 3 = Zwischenhirn 4 = Mittelhirn 5 = Kleinhirn = Hinterhirn 6 = Hypophyse (Hirnanhangsdrüse) 7 = Nachhirn = verlängertes Mark Zwischen-, Mittel- & Nachhirn wird als Stammhirn bezeichnet. Das Gehirn des Menschen „Die Größe des Gehirns sagt nichts über die Intelligenz aus!“ Das Gehirn ist Teil des Zentralnervensystems, der bei Wirbeltieren im Schädel liegt. Beim Menschen ist es etwa eine 1,3 kg schwere Masse aus rosa-grauem Gewebe, mit ca. 10 Milliarden Nervenzellen, die untereinander verknüpft sind. 1. Großhirn (= Vorderhirn) Es besteht aus 2 Hälften, der linken und der rechten Großhirnhemisphäre, und macht etwa 85 % der gesamten Hirnmasse aus. Hier liegt der Sitz der Gedächtnisleistung (Motivation, Lernen/Speicher) und Intelligenzleistung, sowie des Bewusstseins und des Willens. 2. Balken (Corpus Callosum) Ein Faserbündel, das die beiden Großhirnhemisphären miteinander verbindet und Informationsaustausch untereinander ermöglicht. 3. Zwischenhirn Es umfasst den Thalamus, Hypothalamus und die Hypophyse. Thalamus: spielt bei allen Sinnesreizen außer dem Geruchssinn eine Rolle. Vielerlei für die Großhirnrinde bestimmte Signale durchlaufen diese Struktur. Verbindung zwischen Sinnesorganen und Großhirn (Filter!) Hypothalamus: steuert die Anpassung von Funktionen wie Herzfrequenz und Körpertemperatur. Darüber hinaus bestimmt er Schlafrhythmus und Hormonkonzentration. Er ist das Steuerzentrum für das vegetative Nervensystem. 4. Mittelhirn Leitet sensorische Impulse aus dem Rückenmark in andere Hirnabschnitte und steuert Reflexantworten auf bestimmte Reize. Steuerung von Aktivität und Ruhe. Seite 226 © Florian Zeller 08 / 09 5. Kleinhirn (= Hinterhirn) Ist für Gleichgewicht und Haltung verantwortlich, es verarbeitet Informationen über das Raumgefühl und koordiniert alle Bewegungen des Menschen. 6. Hypophyse (Hirnanhangsdrüse) Setzt unter der Kontrolle des Hypothalamus Hormone frei, die im Körper zirkulieren und wiederum die Tätigkeit anderer Drüsen steuern. 7. Nachhirn (verlängertes Mark) Unterstützt die Regulation von Atmung, Schlucken, Blutdruck und Reaktionen wie Niesen ebenso wie den Schlaf. Sitz vieler Reflexzellen. 8. Rückenmark Übermittelt Impulse aus dem Gehirn an andere Körperabschnitte und leitet von dort aus Nachrichten zurück an verschiedene Hirnstrukturen. „ Das menschliche Gehirn ist eine großartige Sache. Es funktioniert vom Moment der Geburt an – bis zu dem Zeitpunkt, wo du aufstehst, um eine Rede zu halten.“ (Mark Twain) Seite 227 © Florian Zeller 08 / 09 Rückenmark: Querschnitt (im Bereich zwischen zwei Wirbeln) Rückenmarkwurzeln: Hinterwurzel: Vorderwurzel: Spinalnerv: alle afferenten Nervenfasern treten über sie ein; Zellkörper der afferenten Nervenfasern befinden sich im Hinterwurzel- oder Spinalganglion. alle efferenten Nervenfasern treten über sie aus! Zellkörper der efferenten Nervenfasern liegen im vorderem Teil der grauen Substanz des Rückenmarks. beide Wurzeln vereinigen sich zum „gemischten“ Nerv, teilt sich aber bald wieder auf und versorgt somatisches und vegetatives Nervensystem. Das Rückenmark Seite 228 © Florian Zeller 08 / 09 Das Nervensystem der Wirbeltiere und des Menschen Zentrales Nervensystem Nervensystem Gehirn Rückenmark Motorische Nervenbahnen Afferenzen somatisches Nervensystem Peripheres Nervensystem animales Nervensystem Sensorische Nervenbahnen Efferenzen vegetatives Nervensystem Versorgt die inneren Organe vegetatives Nervensystem Autonomes = vegetatives Nervensystem Das vegetative Nervensystem innerviert die glatte Muskulatur der inneren Organe und zwar antargonistisch. Es regelt unter anderem die Funktion von: - Atmung - Kreislauf - Verdauung - Stoffwechsel - Drüsen Die zentralen Bereiche liegen im Gehirn, Rückenmark und dem sog. Grenzstrang. Sie sind über den peripheren Bereich mit den inneren Organen verbunden. 2 Systeme Sympathikus - Mobilisiert den Körper (Angriff, Kampf, Flucht) tagaktiv „aggressiv“ Transmitter: - Adrenalin - Noradrenalin Parasympathikus - fördert Erholung + Verdauung Ausgleich des inneren Gleichgewichts nachtaktiv „friedlich“ Acetylcholin Seite 229 © Florian Zeller 08 / 09 Wenn wir schnell viele Treppen hochsteigen, kommen wir leicht „außer Atem“. Unser Herz klopft und wir atmen sehr hastig. Nach wenigen Minuten haben sich Herz- und Lungentätigkeit wieder normalisiert. Unser Wille war an diesen Vorgängen nicht beteiligt. Die Steuerung erfolgte über selbstständig arbeitende Nervenzentren und Nervenbahnen, die man unter der Bezeichnung „vegetatives Nervensystem“ zusammenfasst. Oberstes Steuerungszentrum des vegetativen Nervensystems ist ein Teil des Zwischenhirns, der Hypothalamus. Von hier aus ziehen zwei Nervenstränge links und rechts der Wirbelsäule entlang, die man als Grenzstränge bezeichnet. Neben jedem Wirbelkörper verdicken sie sich zu Nervenknoten, von denen Nervenfasern zu den inneren Organen ziehen. Im Bauchraum und im Unterbauch werden ganze Geflechte von Nervenknoten gebildet. Bekannt ist der „Solarplexus“ („Sonnengeflecht“) im Bauchraum, bei Boxern ein Ziel für einen möglichen K.O.Schlag. Man nennt den bisher beschriebenen Teil des vegetativen Nervensystems den Sympathikus. Ein zweiter Teil des vegetativen Nervensystems besteht aus mehreren Einzelnerven, die im Hypothalamus und anderen Teilen des Stammhirns entspringen und zu den inneren Organen führen. Der bekannteste Nerv in diesem System ist der Vagus. Man nennt das System den Parasympathikus. Wieso besteht das vegetative Nervensystem aus zwei Teilen? Jedes innere Organ ist sowohl mit dem Sympathikus als auch mit dem Parasympathikus verbunden. Der Sympathikus treibt z. B. den Herzschlag an, der Parasympathikus verlangsamt ihn. Andererseits regt der Vagus (Teil des Parasympathikus) die Magendrüsen zur Sekretion an, und der Sympathikus bremst diesen Prozess. Die beiden Systeme sind also Gegenspieler und regeln so die Tätigkeit der inneren Organe unabhängig vom Großhirn. Der Sympathikus fördert die Aktivierung des Körpers; er bringt den Körper „in Schwung“. Der Parasympathikus bremst dagegen viele Organe, er fördert die Erholung und damit diejenigen Organe, die in der Erholungsphase aktiv sind, wie z. B. der Verdauungstrakt. Wieso läuft es uns bei Angst „kalt den Rücken hinunter“? Jeder kennt diese Redensart, doch kaum einer weiß, dass sie ihre Ursache in der Sympathikus-Tätigkeit hat. Das vegetative Nervensystem ist nicht völlig unabhängig vom Großhirn. Wenn wir plötzlich erschrecken, wird über das Großhirn der Sympathikus aktiviert. Die Impulse breiten sich relativ langsam (1 bis 2 𝑚 𝑠) über das Rückenmark nach unten aus und bewirken von oben nach unten absteigend eine Kontraktion der peripheren Blutgefäße. Dies wird von Nervenendigungen in der Haut registriert. Seite 230 © Florian Zeller 08 / 09 + = aktiv - = nicht aktiv Beispiele: Alle Emotionen z.B. Angst, Aggression, Freude usw. finden immer mit Beteiligung des vegetativen Nervensystems statt. Jäger in der Steinzeit liegt am Lagerfeuer, als er plötzlich ein Knacken hört: Knacken löst Erbrecken und Stress aus; es kommt zur Denkblockade; die Wahrnehmung „Angst“ wird an das Zwischen (Hypothalamus) gesendet; der Sympathikus wird aktiviert – Nebenniere schüttet Adrenalin und Noradrenalin aus; dies bewirkt eine Beschleunigung des Herzschlags, der Blutdruck steigt, der Kreislauf verändert sich; Jäger spring auf! Zucker und Fettreserven werden angezapft, um die Muskelversorgung zu decken; Jäger rennt davon oder ist nun auch bereit zu kämpfen; (= akute Phase). Danach folgt Ruhezustand = sog. Erholungsphase. Ein Mensch aus der heutigen Zeit befindet sich in einem Dauerstresszustand; Der permanente Stress ist nicht abbaubar, dies bedeutet einen nicht lösbaren Widerstand; auf die akute Phase kann keine Erholungsphase folgen, daher kommt es zur Erschöpfung (Magengeschwüre, Herzinfarkt) Lösungsmöglichkeit: z.B. durch Entspannungstraining = eine willentliche Einflussnahme auf das vegetative Nervensystem. Besonders Atemübungen haben eine gute Wirkung auf den Parasympathikus („Abschalten“ üben, Verspannungen lösen) Jedoch erfolgt dadurch keine Ursachenänderung, sondern nur eine Anpassung an den Stress. Ebenso wichtig eine Veränderung der Ursachen, soweit als möglich. Seite 231 © Florian Zeller 08 / 09 Reflexbogen = Bezeichnung für die Nervenschaltung eines Reflexes Eine über Nervenzellen laufende Verbindung zwischen Sinnesorganen und Muskeln bezeichnet man als Reflexbogen, die durch den Reiz ausgelöste Reaktion als Reflex. Reflexe Reflexe sind einfache Reaktionen, die dann in immer gleicher, stereotyper und rascher Weise auf spezifische Reize hin eintreten, wenn der Reiz eine bestimmte Stärke aufweist. Reflexe sind stets funktionsbereit. Es scheint für Reflexe ein genetisch fixiertes Programm zu geben, daher bezeichnet man sie als angeboren. Zu den Schutzreflexen zählt man z.B. den Kratz-, Rückzieh-, Liedschluss-, Nieß-, Husten-, Brechreflex und Kniesehnenreflex. Im Zusammenhang mit der Ernährung wirkt der Schluck- und Speichelflussreflex. Lebenserhaltender Reflex ist z.B. der Atemreflex. Reflexe kann man unterteilen in Fremdreflexe und Eigenreflexe Fremdreflex bedeutet, dass der Weg von einem Rezeptororgan über normalerweise mehrere afferente und efferente Nervenbahnen zum Effektororgan führt. Rezeptor und Effektor liegen in verschiedenen Organen, meist polysynaptisch, z.B. Rückziehreflex bei heißer Herdplatte Eigenreflex bedeutet, dass von der Muskelspindel als Rezeptor der Weg zum gleichen Muskel als Effektor zurükführt. (Rezeptor und Effektor im selben Organ, meist monosynaptisch!) Ein beliebtes Beispiel ist der Kniesehnenreflex Dieser monosynaptische Reflex wird an einem „Dehnungsfühler“, den Muskelspindel der Wade wahrgenommen. Er wird über afferente Nervenbahnen durch elektrische Impulse (Rezeptorneuron) zum zentralen Nervensystem übertragen und dort verarbeitet. Auf efferenten Nervenbahnen gelangt die Antwort auf diesen Reiz zu einem Effektor (Muskel, Drüse), der dadurch aktiviert wird und eine Reaktion bedingt. Den gesamten Weg bezeichnet man als Reflexbogen Reflexe von Neugeborenen sind z.B. Lippen-, Saug-, Hand- bzw. Fußgreifreflex; Augenschluss-, Stütz- und Gehreflex. Einige dieser Neugeborenen Reflexe verlieren sich in den ersten Wochen bis Monaten. Seite 232 © Florian Zeller 08 / 09 = Interneuron Verschaltungen in Reflexbögen Seite 233 © Florian Zeller 08 / 09 Sarko = Fleisch (Sarkorak = Fleischfresser) Seite 234 © Florian Zeller 08 / 09 Seite 235 © Florian Zeller 08 / 09 Bau der Quergestreiften Muskulatur Eine Muskelfaser ist aus vielen Myofibrillen zusammengesetzt. Bau einer Myofibrille: Die Myofibrille wird durch sog. Z-Scheiben in ca. 2,5 𝜇m lange Abschnitte gegliedert, die Sarkomere. Die Sarkomere sind die kontraktilen Einheiten des Muskels. Sie bestehen aus Eiweißfilamenten: dicke Eiweißfilamente = Myosin dünne Eiweißfilamente = Actin Die besondere Anordnung der Myosin- und Actinfilamente ergibt eine Streifung quergestreifte Muskulatur. Aktinfilamente Myoisnfilamente (zeichnen können!) Mechanismus der Muskelkontraktion Eine Muskelzelle besitzt wie jede Zelle Organellen, besonders auffällig sind jedoch - viele Mitochondrien (hoher Stoffwechselumsatz) - zwei Hohlraumsysteme, die nicht ineinander übergehen und verschiedene Aufgaben erfüllen: a) Endoplasmatisches Reticulum, hier Sarkoplasmatisches Reticulum genannt, mit Ca 2+-Speicher b) T-System = Transversales Tubuli-System, die tubuli verlaufen quer auf Höhe der Z-Scheiben, leiten Depolarisation ins Innere Feinstruktur des Sarkomers: Aktinfilamente: Aktin: zwei Keten von Nonomeren Tropomyosin: stäbchenförmiges Molekül Troponin: am Ende des Tropomyosins Myosinfilamente: langgestreckte Moleküle mit Köpfchen, die seitlich herausragen Molekularer Mechanismus der Kontraktion Die Länge der Actin- und Myosinfilamente ändert sich dabei nicht!!! Seite 236 © Florian Zeller 08 / 09 Ruhender Muskel: Die Interaktion zwischen Myosinköpfchen und Aktin ist durch das Troponin behinder. Aktiver Muskel: Über die Muskelfasermembran trifft eine Erregung ein, das T-System leitet die Depolarisation ins Innere Ca2+-Permeabilität der Sarkomer-Membran wird erhöht, Ca2+ diffundiert aus dem Sarkomer zu den Filamenten und startet den Bewegungsmechanismus = Gleitfilamentmechanismus: Die hundertfach erhöhte Ca2+-Konzentration bewirkt eine Konformationsänderung des Troponins, das eine Strukturänderung des Tropomyosins, nun können Myosin und Aktin Kontakt aufnehmen. Die Myosinköpfchen heften am Aktin an, winkeln sich um 45° ab (ATP notwendig !!) und ziehen dadurch das Aktin über sich hinweg. Unter ATP-Verbrauch und mit Hilfe von Mg2+ lösen sich die Köpfchen und schwingen zurück 1. „Ruderschlag“ ist beendet. Solange die Ca2+-Konzentration erhöht ist, erfolgten viele „Ruderschläge“ das Sarkomer verkürzt sich der Muskel verkürzt sich. Nach Reizende werden die Ca2+ wieder ins Sarkomer zurückgepumpt, das Troponin-Hemmsystem wird wieder wirksam. Weichmacherwirkung von ATP / Totenstarre: Die Bindung der Myorinköpfchen ans Aktin ist starr! ATP löst diese starre Bindung! Fehlt ATP oder sinkt der ATP-Spiegel bleiben die Muskeln hart. Das ist auch nach dem Tod der Fall. Seite 237 © Florian Zeller 08 / 09 Biologische Regelung Begriffe zum Regelkreisschema (am Beispiel des Kühlschranks) Wer kennt das nicht? Sie reißen die Kühlschranktüre auf, danach brummt der Motor!!! Führungsgröße: bestimmt den Sollwert Sollwert: (das sind Sie, sie stellen den Thermostat ein) ist die Information über den Endzustand der Regelgröße (also wie es sein soll, z.B. wenn Sie unentschlossen die Kühlschranktüre aufreißen und während Ihrer kulinarischen Überlegungen die Temperatur im Kühlschrank auf 15 °C ansteigt, dann soll nachher die Temperatur im Kühlschrank Doch wieder ca. 8°C sein, oder?) Seite 238 © Florian Zeller 08 / 09 Regelgröße: Regelglied: ist ein konstant zu haltender Vorgang oder Zustand (also was soll geregelt werden, hier die 8°C) vergleicht Sollwert und Istwert und löst eine Steuerung aus (der Thermostat im Kühlschrank vergleicht 8°C mit 15°C und löst letztendlich den Kühlschrankmechanismus aus) Istwert: augenblicklicher Zustand der Regelgröße (im Moment Temperatur von 15°C) Fühler: Messeinrichtung für den Istwert, leitet den Istwert an das Regelglied weiter (Temperaturfühler im Kühlschrank) Störgröße: störender Außeneinfluss auf die Regelgröße (Erwärmung der normalen Kühlschranktemperatur) Stellwert: Information, was das Stellglied zu tun hat, oder welche Tätigkeit das Stellglied auszulösen hat (Info, um welchen Betrag die Temperatur verstellt werden muss) Stellglied: ist der Korrekturmechanismus Stellgröße: Tätigkeit des Korrekturmechanismus Regelstrecke: best. Bereich, innerhalb dessen die Regelgröße verändert werden kann (der Motor des Kühlschranks) (Laufzeit des Motors des Kühlschranks) (Der Kühlschrank kann innerhalb eines bestimmten Temperaturbereiches kühlen, aber Sie können ihn nicht als Backofen verwenden) Seite 239 © Florian Zeller 08 / 09 Regelung der Pupillenöffnung Durch die Beleuchtungsstärke + Pfeildiagramm: Lichteinfall (Ursache) Verengung der Pupille (Wirkung) Reitz und Reaktion bzw. Ursache und Wirkung sind zu einem Kausalkreis mit negativer Rückkoppelung geschlossen Regelung über das Nervensystem 1. Regelung der Atemgaskonzentration im Blut Pfeildiagramm: + CO2-Gehalt im Blut Atemtätigkeit Kausalkreis mit negativer Rückkoppelung Regelstrecke Blut Seite 240 © Florian Zeller 08 / 09 2. Regelung der Körpertemperatur Pfeildiagramm: zweiseitige Regelung (Körpertemp. Erhöhen & senken) Außentemperatur Muskelzittern zur Wärmeproduktion Verengung der Blutgefäße (Grundumsatz erhöht sich) + + Körpertemperatur + - Schweißabsonderung Erweiterung der BlutKapillaren (Senkung des Grundumsatzes (GU) ) Seite 241 © Florian Zeller 08 / 09 Hormone Hormone sind Botenstoffe, die der Körper selbst erzeugt. Sie sind bereits in geringsten Mengen wirksam. Sie stellen neben dem Nervensystem ein zweites Informationssystem dar. Chemischer Bau 1. Steroide 2. Aminosäurederivate 3. Peptide / Proteine (Sexualhormone) (Thyroxin, Adrenalin) (Insulin, Glucagon) uneinheitlich Orte der Produktion: 1. Endokrine (innersekretorische) Drüsen (Hypophyse, Schilddrüse, Thymus, Pankreas, Nebennieren, Keimdrüsen) 2. Gewebshormone Hypothalamus bildet Releasing-Hormone Haut bildet Histamine Drüsen Hypophyse Schilddrüse Hormon Tyhreotropin (TSH) Thyroxin Nebennierenrinde Nebennierenmark Pankreas Glucocorticoide Adrenalin Insulin Glucagon Funktion Stimuliert Schilddrüse Stimuliert Stoffwechselprozesse Wirkt auf Grundumsatz heben glucosespiegel im Blut Stresshormon, erhöht Stoffwechsel senkt Glucosespiegel im Blut hebt Glucosespielgel im Blut Zusammenhang zwischen Hormonsystem und Nervensystem Hormonsystem und Nervensystem sind bei der Aufrechterhaltung der Homöostase (inneres Gleichgewicht), der Entwicklung und der Fortpflanzung oft untrennbar verwoben. 1. 2. 3. Strukturelle Beziehungen: Der Hypothalamus und die Hypophyse bestehen aus Nervengewebe, das Nebennierenmark hat sich aus Nervengewebe entwickelt. Chemische Beziehung: Mehrere Hormone werden auch als Neurotransmitter verwendet (z.B. Adrenalin). Funktionelle Beziehung: Viele Körperfunktion werden von beiden Systemen reguliert (z.B. Stressreaktionen, Säugen der Jungen). Seite 242 © Florian Zeller 08 / 09 Regelung über das Hormonsystem Regelung des Grundumsatzes bzw. des Schilddrüsen-Hormons Thyroxin Pfeildiagramm: Hypothalamus Neurohormon TRH - + Hypophyse - Grundumsatz, Höhe des Energiestoffwechsels und der Wärmeproduktion Thyroxinkonzentration im Blut Steuerhormon TSH + + + SCHILDDRÜSE THYROXIN Regelung der Zuckerkonzentration im Blut durch Adrenalin Glucagon Glycogenabbau zu Glucose Insulin: Glycogenaufbau (holt Zucker aus dem Blut) Pfeildiagramm: Muskeltätigkeit Glucagonfreisetzung Kohlenhydratverdauung (Zufuhr von Glucose) + + Insulinfreisetzung Glucosespiegel im Blut + Glycogenzerlegung + + Glycogenaufbau 2 Regler beteiligt: 1. Hypothalamus + Hypophyse 2. Bauchspeicheldrüse Seite 243 © Florian Zeller 08 / 09 Seite 244 © Florian Zeller 08 / 09