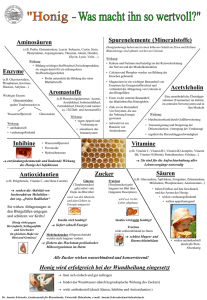
Einfluss einer Supplementierung mit beta
Werbung

Aus dem Arbeitsbereich Bestandstiermedizin der Klinik für Rinder (im Richard-Götze-Haus) Tierärztliche Hochschule Hannover Einfluss einer Supplementierung mit beta-Karotin in Form einer Injektionslösung (Carofertin) auf die Fruchtbarkeitsleistung von Milchkühen INAUGURAL-DISSERTATION Zur Erlangung des Grades einer Doktorin der Veterinärmedizin (Dr.med.vet.) durch die Tierärztliche Hochschule Hannover Vorgelegt von Natascha Gossen aus Etterbeek (Belgien) Hannover 2003 2 Wissenschaftliche Betreuung: Univ.-Prof. Dr. M. Hoedemaker, Ph.D. 1. Gutachter: Univ.-Prof. Dr. M. Hoedemaker, Ph.D. 2. Gutachter: Univ.-Prof. Dr. B. Meinecke Tag der mündlichen Prüfung: 03.06.2003 3 Meiner Familie und meinen Freunden gewidmet 4 Inhaltsverzeichnis 1 Einleitung........................................................................................................... 11 2 Literaturübersicht ............................................................................................... 12 2.1 Was ist beta-Karotin? ................................................................................. 12 2.2 Vorkommen in der Natur............................................................................. 13 2.3 Vorkommen in Futtermitteln........................................................................ 14 2.3.1 Gehalte................................................................................................ 14 2.3.2 Verwertbarkeit ..................................................................................... 15 2.4 Bedarf einer Milchkuh an beta-Karotin ....................................................... 15 2.5 Vitamin-A-Bedarf ........................................................................................ 17 2.6 Resorption von beta-Karotin nach oraler Aufnahme................................... 19 2.6.1 Tierartliche Unterschiede..................................................................... 19 2.6.2 Resorption von beta-Karotin beim Rind............................................... 19 2.7 Transport im Körper nach oraler Aufnahme................................................ 20 2.7.1 Transport im Blut ................................................................................. 20 2.7.2 Aufnahme in die Zellen........................................................................ 21 2.7.3 Versorgung von Euter und Follikeln .................................................... 21 2.8 Verteilung von beta-Karotin in den Geweben und in der Milch ................... 22 2.8.1 Blutplasma und Fettgewebe ................................................................ 22 2.8.2 Leber ................................................................................................... 22 2.8.3 Milch .................................................................................................... 23 2.9 Resorption nach parenteraler Gabe ........................................................... 24 2.10 Beta-Karotin im Blut.................................................................................... 24 2.10.1 Untersuchungsmethoden .................................................................... 24 2.10.2 Beta-Karotin-Gehalte im Blut von Milchkühen ..................................... 25 2.10.3 Individuelle Schwankungen ................................................................. 27 2.10.4 Blutspiegel im peripartalen Zeitraum ................................................... 27 2.11 Beta-Karotin als Provitamin A..................................................................... 28 2.11.1 Vitamin-A-Stoffwechsel ....................................................................... 28 2.11.2 Einige Funktionen des Vitamin A im Organismus................................ 29 5 2.12 2.12.1 Beta-Karotin und die Fruchtbarkeitsparameter.................................... 30 2.12.2 Beta-Karotin und die Gravidität ........................................................... 33 2.12.3 Beta-Karotin und Retentio secundinarum............................................ 33 2.13 Beta-Karotin und das Ovar ......................................................................... 34 2.13.1 Allgemeines......................................................................................... 34 2.13.2 Beta-Karotin-Gehalte........................................................................... 34 2.13.3 Ovarialzysten....................................................................................... 35 2.13.4 Progesteronproduktion und beta-Karotin-Gehalte im Ovar.................. 36 2.13.5 Progesteronproduktion und beta-Karotin-Gehalte im Blut ................... 36 2.13.6 Beta-Karotin in der Zelle...................................................................... 38 2.13.7 Vitamin-A-unabhängige Aufgabe des beta-Karotins im Ovar .............. 38 2.13.8 Beta-Karotin als lokale Vitamin-A-Quelle des Ovars ........................... 39 2.14 Beta-Karotin und das Immunsystem........................................................... 40 2.15 Weitere Einflussfaktoren auf die Fruchtbarkeit des Rindes ........................ 41 2.15.1 Energieversorgung .............................................................................. 41 2.15.2 Eiweißversorgung................................................................................ 42 2.15.3 Mineralstoffe........................................................................................ 43 2.15.4 Spurenelemente .................................................................................. 43 2.15.5 Vitamine .............................................................................................. 44 2.15.6 Mykotoxine .......................................................................................... 45 2.15.7 Oxidativer Stress ................................................................................. 45 2.16 3 Beta-Karotin und Fruchtbarkeit................................................................... 30 Kosten pro Trächtigkeit............................................................................... 45 Material und Methoden ...................................................................................... 48 3.1 Das Präparat: Carofertin.......................................................................... 48 3.2 Überprüfung der Vitamin-A-, Vitamin-E- und Selenversorgung .................. 49 3.2.1 Bestimmung von Vitamin A und Vitamin E im Serum.......................... 50 3.2.2 Bestimmung von Selen im Serum ....................................................... 52 3.3 Verlaufsstudie............................................................................................. 52 3.3.1 Versuchsziel ........................................................................................ 52 3.3.2 Haltung und Fütterung der Versuchstiere............................................ 53 6 3.3.3 Versuchsaufbau .................................................................................. 53 3.3.4 Untersuchung des Serums auf beta-Karotin........................................ 54 3.4 4 Langzeitstudie ............................................................................................ 55 3.4.1 Versuchsziele ...................................................................................... 55 3.4.2 Haltung der Versuchstiere ................................................................... 55 3.4.3 Fütterung der Versuchstiere ................................................................ 57 3.4.3.1 Vor dem Stallumbau..................................................................... 57 3.4.3.2 Nach dem Stallumbau .................................................................. 60 3.4.4 Versuchsaufbau: beta-Karotin-Gehalt im Serum ................................. 62 3.4.5 Versuchsaufbau: Puerperalkontrolle ................................................... 63 3.4.6 Kalbeverlauf, Krankheiten und Abgänge ............................................. 66 3.4.7 Fruchtbarkeitskennzahlen ................................................................... 66 3.5 Progesteronbestimmung............................................................................. 68 3.6 Kosten pro Trächtigkeit............................................................................... 69 3.7 Auswertung................................................................................................. 71 Ergebnisse......................................................................................................... 72 4.1 Verlauf der beta-Karotin-Konzentration (Verlaufsstudie) ............................ 72 4.2 Vitamin-A-, Vitamin-E- und Selenversorgung (Langzeitstudie)................... 75 4.3 Übersicht über den Verbleib der Versuchstiere (Langzeitstudie)................ 76 4.4 Beta-Karotin-Gehalte im Serum (Langzeitstudie) ....................................... 77 4.4.1 Alle Tiere ............................................................................................. 77 4.4.2 Kühe .................................................................................................... 78 4.4.3 Färsen ................................................................................................. 79 4.4.4 Kühe mit einem beta-Karotin-Wert unter 2000 µg/l zum Zeitpunkt I. ... 80 4.5 Puerperalkontrolle (Langzeitstudie) ............................................................ 82 4.5.1 Beginn des Zyklus ............................................................................... 82 4.5.2 Ovarialzysten....................................................................................... 82 4.5.3 Endometritis ........................................................................................ 83 4.6 Krankheiten und Abgänge (Langzeitstudie)................................................ 84 4.6.1 Peripartale Erkrankungen : Milchfieber und Nachgeburtsverhaltung... 84 4.6.2 Endometritis in der Gesamtlaktation.................................................... 84 7 4.6.3 Ovarialzysten in der Gesamtlaktation .................................................. 85 4.6.4 Einfluß des beta-Karotin-Spiegels auf die Inzidenz von Nachgeburtsverhaltung, Endometritis und Ovarialzysten .................... 86 4.6.5 4.7 Abgänge .............................................................................................. 88 Fruchtbarkeitskennzahlen (Langzeitstudie) ................................................ 89 4.7.1 Konzeptionsrate .................................................................................. 89 4.7.2 Gesamtträchtigkeitsrate....................................................................... 90 4.7.3 Erstbesamungserfolg........................................................................... 91 4.7.3.1 Auswertung nach Gruppen........................................................... 91 4.7.3.2 Auswertung nach Quartilen .......................................................... 91 4.7.4 Trächtigkeitsindex ............................................................................... 92 4.7.4.1 Auswertung nach Gruppen........................................................... 92 4.7.4.2 Auswertung nach Quartilen .......................................................... 93 4.7.5 Rast-, Güst-, Verzögerungs- und erwartete Zwischenkalbezeit........... 94 4.7.5.1 Auswertung nach Gruppen........................................................... 94 4.7.5.2 Auswertung nach Quartilen .......................................................... 96 4.7.5.2.1 Zeitpunkt I ................................................................................. 96 4.7.5.2.2 Zeitpunkt II ................................................................................ 97 4.7.5.2.3 Zeitpunkt III ............................................................................... 99 4.7.5.3 Lineare Regressionsanalyse ...................................................... 100 4.7.6 Prozentsatz der Tiere mit einer Güstzeit unter 115 Tagen ................ 100 4.7.7 200-Tage-Nichtträchtigkeitsrate ........................................................ 101 4.7.8 Prozentsatz der Tiere, die 60 Tage post partum noch nicht in Brunst gesehen wurden ................................................................................ 102 4.7.9 5 Prozentsatz abortierender Tiere ........................................................ 102 4.8 Progesteronbestimmung........................................................................... 103 4.9 Kosten pro Trächtigkeit............................................................................. 107 Diskussion ....................................................................................................... 109 5.1 Verlauf der beta-Karotin-Konzentration (Verlaufsstudie) .......................... 109 5.2 Vitamin-A, Vitamin-E- und Selenversorgung (Langzeitstudie).................. 111 5.2.1 Vitamin A ........................................................................................... 111 8 5.2.2 Vitamin E ........................................................................................... 112 5.2.3 Selen ................................................................................................. 112 5.3 Weitere Voraussetzungen für die Anwendung von Carofertin (Langzeitstudie)............................................................................................ 113 5.4 Beta-Karotin-Gehalte im Blutserum (Langzeitstudie)................................ 113 5.5 Puerperalkontrolle (4-6 Wochen p.p., Langzeitstudie).............................. 116 5.6 Krankheiten und Abgänge ........................................................................ 118 5.6.1 Peripartale Erkrankungen: Milchfieber und Nachgeburtsverhaltung.. 118 5.6.2 Endometritis in der Gesamtlaktation.................................................. 119 5.6.3 Ovarialzysten in der Gesamtlaktation ................................................ 120 5.6.4 Abgänge ............................................................................................ 121 5.7 Fruchtbarkeitskennzahlen......................................................................... 122 5.7.1 Auswertung nach Gruppen................................................................ 122 5.7.2 Einfluß des beta-Karotin-Spiegels auf die Fruchtbarkeitskennzahlen 124 5.8 Progesteronbestimmung........................................................................... 125 5.9 Kosten pro Trächtigkeit............................................................................. 126 5.10 Schlussfolgerungen .................................................................................. 127 6 Zusammenfassung .......................................................................................... 128 7 Summary ......................................................................................................... 130 8 Literatur ........................................................................................................... 132 9 Verzeichnis der verwendeten Abkürzungen a. p. ante partum Abb. Abbildung AmPreisV Arzneimittel-Preis-Verordnung ATP Adenosintriphosphat bzw. beziehungsweise d Tag d.h. das heißt DNS Desoxyribonukleinsäure etc. et cetera evtl. eventuell FM Frischmasse GnRH Gonadotropin Releasing Hormon GOT Gebührenordnung für Tierärzte h Stunde HCG Humanes Chorion Gonadotropin HDL High Density Lipoprotein HPLC High Pressure Liquid Chromatography I.E. Internationale Einheiten i.m. intramuskulär incl. inklusive KB künstliche Besamung kg Kilogramm KOH Kaliumhydroxid KGW Körpergewicht LDL Low Density Lipoprotein LH Luteinisierungshormon mg Milligramm MLF Milchleistungsfutter PGF2α Prostaglandin F2α 10 p.i. post inseminationem p.p. post partum RIA Radioimmunoassay Tab. Tabelle TM Trockenmasse TMR Total Mixed Ration u.a. unter anderem v.a. vor allem VLDL Very Low Density Lipoprotein Wo. Wochen z.B. zum Beispiel z.T. zum Teil 11 1 Einleitung In den letzten Jahrzehnten wurde der Einfluß von beta-Karotin auf die Fruchtbarkeit des Rindes schon mehrfach untersucht. Man stellte neben der Wirkung als Provitamin A eine von Vitamin A unabhängige Wirkung auf die Fruchtbarkeit fest (Lotthammer et al. 1976). Ein Mangel an beta-Karotin führt zu Stillbrünstigkeit, Ovarialzysten und schlechten Besamungsergebnissen. Diese negativen Auswirkungen konnten durch die Gabe ausreichender beta-Karotin-Mengen behoben werden. Es wurde aber bereits damals darauf hingewiesen, dass beta-Karotin in der Praxis nur unter extremen Bedingungen als alleinige Ursache für ein Fruchtbarkeitsproblem in Frage kommt. So ist heutzutage in hochleistenden Herden, die entsprechend gefüttert werden, ein beta-Karotin-Mangel nahezu ausgeschlossen. In dieser Untersuchung wurde die Wirkung von Carofertin in einer hochleistenden Milchviehherde getestet. Dieses Präparat wurde zur Prophylaxe und Unterstützung der Therapie von Fruchtbarkeitsstörungen bei Rind und Schwein entwickelt. Carofertin ist eine 1 %-ige beta-Karotin-Lösung zur intramuskulären Injektion, die laut der Produktinformation des Herstellers Alvetra GmbH zur Verbesserung der Fruchtbarkeit einer Herde beitragen soll. Durch die gezielte Gabe des beta-Karotins über eine Injektion wurde in unserer Untersuchung sichergestellt, dass jedes Versuchstier die gleiche Menge an beta-Karotin erhielt. In früheren Untersuchungen wurde beta-Karotin dagegen meist über das Futter supplementiert. Es handelte sich dabei um Kraft- bzw. Mineralmineralfuttermittel, denen beta-Karotin zugesetzt worden war, oder um spezielle Diäten mit Futtermitteln, die bekanntermaßen viel beta-Karotin enthalten, wie zum Beispiel Möhren oder Luzerne (Flachowsky 1999). Die Aufnahme einer definierten Menge an beta-Karotin war auf diese Weise schwer zu verwirklichen. 12 2 Literaturübersicht 2.1 Was ist beta-Karotin? Chemisch gesehen gehört beta-Karotin in die Gruppe der Kohlenwasserstoffe. Das sind Verbindungen, die nur aus Kohlenstoff- und Wasserstoffatomen bestehen. Von ihnen leiten sich alle anderen organischen Verbindungen ab, indem einzelne HAtome durch andere funktionelle Gruppen ersetzt werden. Eine Untergruppe der Kohlenwasserstoffe sind die Alkene. Sie zeichnen sich durch eine oder mehrere C=C-Doppelbindungen aus und werden deshalb auch als ungesättigte Kohlenwasserstoffe bezeichnet. Sie entstehen unter anderem durch Dehydrierung von Alkanen und durch Dehydratisierung von Alkoholen (Zeeck et al. 1992). Verbindungen, die mehrere C=C-Doppelbindungen enthalten, werden als Polyene bezeichnet. Zu dieser Gruppe gehören die Karotine. Von ihnen leiten sich wieder zahlreiche hydroxylierte Verbindungen, die Karotinoide, ab. Beta-Karoten gehört zu dieser Gruppe. Es ist ein Polyen mit elf C=C-Doppelbindungen. Die Strukturformeln von beta-Karoten und Vitamin A sind Abbildung 1 zu entnehmen. Abbildung 1: Strukturformel von beta-Karotin und Vitamin A. Quelle: Friesecke (1978) 13 Dieses Molekül ist sehr lipophil. Die Endung „–en“ zeigt, dass es zu den Alkenen gehört (Zeeck et al. 1992). Von den meisten Autoren wird für diese Verbindung der Begriff beta-Karotin verwendet. Um keine Verwirrung zu stiften, wird im Folgenden ebenfalls die Bezeichnung beta-Karotin verwendet. 2.2 Vorkommen in der Natur Die Karotinoide sind fettlösliche, gelbe bis rotviolette Polyenfarbstoffe, die in der Natur weit verbreitet sind. Zu den Karotinoiden gehört zum Beispiel der rote Farbstoff der Möhre (beta-Karotin) und der gelbe Farbstoff des Maises (Kryptoxanthin). Von den über 400 natürlich vorkommenden Karotinoiden gehören über 40 zu den AProvitaminen, d.h. der Organismus kann daraus Vitamin A bilden. In der Nahrung findet man in der Regel nur vier bis sechs A-Provitamine. Praktische Bedeutung haben alpha-, beta- und gamma-Karotin. Beta-Karotin ist die bedeutsamste und mengenmäßig am häufigsten vorkommende Vitamin-A-Vorstufe und wird deshalb oft selbst zu den fettlöslichen Vitaminen gezählt. Vertreter der Karotinoide, die nicht zur Bildung von Vitamin A eingesetzt werden können, sind z. B. der rote Farbstoff der Tomate (Lycopin) sowie das Lutein (Eigelb, Laubblätter) und das Canthaxanthin (Pfifferlinge). Karotin und Karotinoide sind in grünen Pflanzenzellen in größerer Konzentration in den Chloroplasten und Chromoplasten vorhanden. Sie sind für die Photosynthese und für den Schutz vor Sauerstoff-Radikalen von Bedeutung (Kolb 1998). 14 2.3 Vorkommen in Futtermitteln 2.3.1 Gehalte Blattreiche Grünfuttermittel wie Weidegras, Grassilage, Rübenblattsilage und Heu sind relativ reich an beta-Karotin. Gesamtkarotingehalte von über 300 mg/kg TM (=Trockenmasse) sind keine Seltenheit. Der beta-Karotin-Anteil betrug bei Analysen bis zu 80 %. Die Gehalte schwanken in der Regel stark, weil sie vom Erntezeitpunkt, d.h. dem Pflanzenalter, der Konservierungsart und der Dauer der Lagerung, abhängig sind. In Tabelle 1 sind die beta-Karotin-Gehalte einiger Futtermittel aufgeführt. Tabelle 1: Beta-Karotin-Gehalt verschiedener Futtermittel in mg/kg Frischmasse (FM). Quelle: Kolb (1998) Futtermittel Beta-Karotin mg/kg FM Futtermittel Beta-Karotin mg/kg FM Grünfutter, frisch Grassilage, frisch Kleesilage Maissilage Anwelksilage, länger gelagert 180-250 10-40 10-70 4-20 7-15 Rübenblattsilage Luzernemehl, frisch Heu, frisch Heu, gelagert Möhren 5-10 200-300 10-30 1-5 60-80 Bei der Heuwerbung betrugen die Karotinverluste 70-90 %. Getreide enthielt betaKarotin nur in geringen Mengen und im Getreidestroh war der Karotingehalt fast Null (Flachowsky 1999). Die beta-Karotin-Gehalte von Silagen waren in der Regel höher als die von Heu, aber auch hier kam es je nach Konservierungsmethode und Lagerungszeit zu Verlusten bis zu 90 % (Kirchgessner 1987). Wenn frisches Gras siliert wurde, waren die beta-Karotin-Verluste gering. War das Gras angewelkt oder 15 unter ungünstigen Witterungsbedingungen (zu feucht) eingebracht worden, stiegen die Verluste (Grummer u. Clark 1980). 2.3.2 Verwertbarkeit Die Verwertbarkeit, das heißt in welchem Umfang die Tiere das beta-Karotin nutzen konnten, war ebenfalls von Futtermittel zu Futtermittel unterschiedlich. Die Verwertung von beta-Karotin aus der Maissilage war geringer als die aus der Grassilage. Mit steigender Verholzung der Pflanzen nahm die Verwertbarkeit ab. Aus Heu wurde beta-Karotin zu 25 % und aus Grassilage zu 32 % verwertet (Nehring u. Hoffmann 1967). Fernandez et al. (1976) beschrieben, dass beim Wiederkäuer nur circa 5-10 % des beta-Karotins des Futters durch die Mikroorganismen im Pansen abgebaut werden. 2.4 Bedarf einer Milchkuh an beta-Karotin Die Ableitung des Bedarfs an Vitaminen wird im Allgemeinen durch die unterschiedlichen Umwandlungsraten von Vitamin-Vorstufen in die entsprechenden Vitamine erschwert. Der Abbau im Pansen ist bei den meisten Vitaminen nicht genau bekannt. Bei beta-Karotin beträgt er 5-10 % (Fernandez et al. 1976). Oftmals bleibt deshalb der Vitamingehalt der Grundration bei der Bedarfsableitung unberücksichtigt. Eine weitere Schwierigkeit bei der Ableitung von Versorgungsempfehlungen für Vitamine besteht in den wenigen oder kaum vorhandenen Dosis-Wirkungsversuchen bei Milchkühen. Man unterscheidet folgende Bedarfskategorien: Erhaltungsbedarf (Vermeidung von Mangelsymptomen), Bedarf (ausreichende Versorgung unter Berücksichtigung von Leistungshöhe, Tiergesundheit und Reproduktion), Bedarf mit Sicherheitszuschlägen (Erreichung bestimmter Gewebekonzentrationen, z. B. betaKarotin im Serum über 3000 µg/l) und bedarfsübersteigende Versorgung zur Erreichung besonderer Zusatzeffekte wie z.B. antioxidatives Potential. Ahlswede und Lotthammer (1978) nehmen an, dass der gesamte Körpervorrat einer laktierenden 16 Kuh an beta-Karotin zur Deckung des Erhaltungsbedarfs für 3 bis 4 Tage reicht. Die kontinuierliche Zufuhr über das Futter ist somit von großer Bedeutung. Insgesamt gesehen kommen typische Vitamin-Mangelerscheinungen bei Milchkühen in Deutschland kaum noch vor (Flachowsky 1999). Die in der Praxis gegenwärtig üblichen Dosierungen gehen meist über das Niveau herkömmlicher Empfehlungen (Bedarf plus Sicherheitszuschlag) hinaus. So werden heute in Deutschland > 15 mg beta-Karotin/kg TM in Rationen von Aufzuchtrindern und Milchkühen empfohlen (Flachowsky 1999). Eine laktierende Kuh bekommt auf diese Weise bei einer Trockenmasseaufnahme von 20 kg und einem beta-Karotin-Gehalt des Futters von 15 mg/kg TM 300 mg beta-Karotin pro Tag. Lotthammer (1979) nahm an, dass pro kg Milch zusätzlich 20 mg beta-Karotin benötigt werden. Somit würde der Bedarf bei einem laktierenden Tier mit einer Milchleistung von 40 kg auf 1100 mg beta-Karotin pro Tag steigen. 17 Da die Autoren sich in den Bedarfsangaben teils erheblich unterscheiden, folgt eine Auflistung der am häufigsten zitierten Angaben (Tab. 2): Tabelle 2: Beta-Karotinbedarf von Milchkühen (kg = Kilogramm, mg = Milligramm, KGW = Körpergewicht, TM = Trockenmasse) Autor (Jahr) Bedarf Ronning (1959) Seitaridis (1963) Rosenberger (1978) Lotthammer (1979) Günther (1980) 0,18 mg/kg KGW/Tag 400-500 mg/Tag 0,2 mg/kg KGW/Tag 100 mg/Tag 300-480 mg/Tag 100 mg/Tag 10 mg/kg Milch Grummer u. Clark (1980) 400-500 mg/Tag Stiewe (1984) 200-400 mg/Tag Marschang (1985) 600 mg/Tag 200 mg/Tag 20 mg/kg Milch Lotthammer (1985b) 100-200 mg/Tag 300-400 mg/Tag Herdt u. Stowe (1991) mind. 300 mg/Tag Schweigert (1995) 200-400 mg/Tag Flachowsky (1999) Besonderheiten 100 mg/Tag bei 500 kg KGW Plus 20 mg/kg Milch Bedarf einer laktierenden Kuh Erhaltungsbedarf eines Rindes Zusätzlicher Bedarf je kg Milch Bedarf einer laktierenden Kuh Bedarf einer laktierenden Kuh Bedarf einer laktierenden Kuh Erhaltungsbedarf Zusätzlicher Bedarf je kg Milch Bedarf hochtragender Kühe Bedarf einer laktierenden Kuh Bedarf einer laktierenden Kuh Empfohlene Zulage zur Verbesserung der Ovarfunktion > 15 mg/kg TM 2.5 Vitamin-A-Bedarf Da bei beta-Karotin häufig nur an seine Funktion als Vitamin-A-Vorstufe gedacht wird, geben einige Autoren lediglich den Vitamin-A-Bedarf von Milchkühen an. Wiederkäuer nehmen als reine Pflanzenfresser mit dem Grundfutter kein Vitamin A auf. In der Fütterungspraxis wird es deshalb entweder über das Kraftfutter oder ein vitaminisiertes Mineralfutter supplementiert. Es ist auch möglich, den Vitamin ABedarf über einen ausreichenden Gehalt an beta-Karotin im Futter zu decken. Bei der Rationsberechnung muss bedacht werden, dass beim ruminierenden Rind ein 18 erheblicher Teil des oral aufgenommenen Vitamin A im Pansen mikrobiell abgebaut wird. Bei Fütterungsversuchen mit Bullen kam es zu einem durchschnittliche Abbau von 50%. Der Umfang des Abbaus war vom Gehalt des Futters an Stärke und Rohprotein abhängig. Außerdem wurde umso mehr Vitamin A abgebaut je mehr Mikroorganismen im Pansen vorhanden waren (Warner et al. 1970). Im Allgemeinen wird beim Wiederkäuer mit einer Umwandlung von 1 mg beta-Karotin in 400 IE Vitamin A gerechnet (NRC 1989, NRC 2001, Ullrey 1972). Angaben verschiedener Autoren zum Vitamin-A-Bedarf von Milchkühen sind Tabelle 3 zu entnehmen. Tabelle 3: Vitamin-A-Bedarf von Milchkühen (IE = Internationale Einheiten, kg = Kilogramm, mg = Milligramm, KGW = Körpergewicht, TM = Trockenmasse) Autor/Jahr Bedarf Besonderheiten Rosenberger (1978) 20-40 IE/kg KGW/Tag 200-250 IE/kg KGW/Tag 250-300 IE/kg KGW/Tag 75000 IE/Tag 75000-100000 IE/Tag 100000-150000 IE/Tag 75000 IE/Tag Erhaltungsbedarf nicht-/frühtragende Kuh hochtragende Kuh tragende Kuh trockenstehende Kühe laktierende Kühe frisch abgekalbte Kuh (680 kg KGW) hochtragende Kuh in Transitphase hochtragende Färse in Transitphase trockenstehende Kuh NRC (1989) Kolb (1998) NRC (2001) 100000 IE/Tag 75000 IE/Tag 83000 IE/Tag 19 2.6 Resorption von beta-Karotin nach oraler Aufnahme 2.6.1 Tierartliche Unterschiede Die Resorption von beta-Karotin im Dünndarm erfolgt tierartlich sehr unterschiedlich. Schwein, Schaf, Ziege, Ratte und Fleischfresser nehmen beta-Karotin höchstens in Spuren in den Organismus auf oder wandeln es noch in der Darmwand vollständig in Vitamin A um. Diese Tiere besitzen dadurch auffallend weißes Körperfett. Bei Rind und Pferd wird das beta-Karotin aus dem Darm aufgenommen und in Blut und Geweben akkumuliert (Schweigert 1988). Dadurch erscheint das Körperfett dieser Tiere auffallend gelb. 2.6.2 Resorption von beta-Karotin beim Rind Man nimmt an, dass für die Resorption der Karotinoide im Dünndarm die Freisetzung der Karotinoide aus der Bindung an Proteine notwendig ist. Die Karotinoide müssen außerdem im Dünndarm in die Mizellen eingebaut werden. Die Mizellen bestehen aus Gallensäuren, Phospholipiden, Monoacylglyceriden und Fettsäuren. Sie ermöglichen die Resorption von Fetten und fettlöslichen Nahrungsbestandteilen. Die Mizellen werden wahrscheinlich an die Oberfläche der Epithelzellen angelagert und ihre Bestandteile durch Diffusion aufgenommen. Die Verwertung der Karotinoide ist von der Menge der gebildeten Mizellen, also auch vom Fettgehalt des Futters abhängig. In Versuchen wurde die Resorption zum Beispiel durch einen relativ hohen Lipidgehalt des Weidegrases gefördert (Kolb 1998). Durch den Zusatz von Fett zum Futter stieg die beta-Karotin-Konzentration im Blut durch die gesteigerte enterale Resorption an (Weiss et al. 1994). Eine andere Untersuchung hat ergeben, dass eine Überversorgung mit Kalium die beta-Karotin-Resorption beeinträchtigt (Lotthammer 1985a). Zusammenfassend kann gesagt werden, dass die Resorption der Karotinoide beim Rind noch nicht vollständig geklärt ist. Außerdem wurde 20 festgestellt, dass beta-Karotin die Verdauung von anderen Futterbestandteilen beeinflusst. Versuche zeigten, dass beta-Karotin seinerseits die Verwertung von langkettigen Fettsäuren und auch die Verwertung von Zellulose verbessert, in dem es das Wachstum der Pansenbakterien förderte (Hino et al. 1993). 2.7 Transport im Körper nach oraler Aufnahme 2.7.1 Transport im Blut Beta-Karotin ist eine lipophile Substanz. Sie muss an geeignete Lösungsvermittler gebunden werden, um im wässrigen Milieu der Körperflüssigkeiten transportiert werden zu können. Diese Rolle übernehmen die Lipoproteine, die in der Leber gebildet werden. Es sind Lipid-Protein-Komplexe, die aus einer Lipidkomponente und Apoproteinen bestehen. Diese Lipoproteine werden nach ihrer Dichte in vier Klassen eingeteilt. Die geringste Dichte besitzen die Chylomikronen, dann folgen die Very Low Density Lipoproteine (VLDL), die Low Density Lipoproteine (LDL) und die High Density Lipoproteine (HDL) (Eisele 1987). Die Existenz der Chylomikronen beim ruminierenden Rind ist im Gegensatz zum Milchkalb umstritten, da durch die besondere Verdauungsphysiologie der Wiederkäuer im Darm nur geringe Mengen an Lipiden zur Resorption kommen. Deshalb werden im Vergleich zu anderen Tierarten nur wenige Chylomikronen benötigt, um die Lipide abzutransportieren und diese werden rasch im Blut abgebaut (Hartmann et al. 1966). Ihr Nachweis ist somit schwierig und man ist dazu übergegangen beim Rind alle Lipoproteine mit einer Dichte unter 1,006 g/ml zur VLDL-Fraktion dazuzurechnen (Palmquist 1976). Bei Versuchen wurde beta-Karotin zu über 80 % an die HDL-Fraktion der Lipoproteine gebunden (Ashes et al. 1982, Kurz et al. 1984, Patton et al. 1980, Schweigert 1988). Die LDL-Fraktion band 11-15 % des beta-Karotins und die VLDL-Fraktion weniger als 1 % des beta-Karotins (Ashes et al. 1984, Kurz et al. 1984). Die Verteilung des beta-Karotins auf die Liporoteine war unabhängig von Rasse, Geschlecht, Ausscheidung über die Milch und von seiner Konzentration im Blut. Ashes et al. 21 (1984) stellten bei ihren Untersuchungen außerdem fest, dass das Blut von Kühen HDL von unterschiedlicher Partikelgröße enthielt. Größere HDL-Partikel enthielten mehr beta-Karotin pro Gewichtseinheit als HDL mit einem kleineren Durchmesser. Bei der Fütterung geschützter Fette entstanden vermehrt große HDL. Diese enthielten mehr beta-Karotin pro Gewichteinheit und somit war der beta-KarotinGehalt im Blut der Kühe gesteigert. Wie die Bindung des beta-Karotins an die Lipoproteine erfolgt, ist noch unklar. Ashes et al. (1984) nahmen an, dass eine unspezifische Einlagerung in den Lipidkern der Lipoproteine vorliegt. Zuvor war eine spezifische Bindungsform postuliert worden (Krinsky et al. 1958). 2.7.2 Aufnahme in die Zellen Mit den Lipoproteinen niedriger und hoher Dichte werden die Karotine über deren Bindung an Rezeptoren der Zellmembran in die Zellen überführt und in die Membranen des Zellkerns und der Mitochondrien eingelagert. Dort sollen sie als Radikalfänger dienen (Kolb u. Seehawer 1997). Auch der intrazelluläre Transport von beta-Karotin ist ungeklärt. Versuche haben ergeben, dass dies wahrscheinlich nicht durch zytosolische Transportproteine passiert. Ob Vesikel oder andere membrangebundene Proteine dafür in Frage kommen, muss noch untersucht werden (Gugger u. Erdman 1996). 2.7.3 Versorgung von Euter und Follikeln Die Versorgung der peripheren Gewebe erfolgt nicht einheitlich, sondern je nach Gewebe wird das beta-Karotin einer bestimmten Lipoproteinfraktion verwertet. Das Euter nutzt während der Bildung des Kolostrums die LDL-Fraktion, während den Follikeln nur an HDL gebundenes beta-Karotin zur Verfügung steht (Schweigert 1986). Die wachsende Oozyte liegt im Ovar im Cumulus oophorus und dieser wiederum ist von der Follikelflüssigkeit umgeben. Die Follikelflüssigkeit besteht einerseits aus verschiedenen niedermolekularen Verbindungen aus dem Blut und 22 andererseits aus Stoffwechselprodukten der Granulosazellen (z.B. Steroide). Die Follikelflüssigkeit wird von der sehr selektiven Blut-Follikelflüssigkeitsschranke abgetrennt (Edwards 1974). Diese Schranke fungiert als Filter für Moleküle mit einem Molekulargewicht von mehr als 850000 Dalton. Dazu gehören auch die VLDL und die LDL (Shalgi et al. 1973). Das beta-Karotin im Blut gelangt über die Aufnahme der HDL in den Follikel, weil die HDL ein sehr niedriges Molekulargewicht haben und somit die Blut-Follikel-Schranke passieren können (Chew et al. 1984). 2.8 Verteilung von beta-Karotin in den Geweben und in der Milch 2.8.1 Blutplasma und Fettgewebe In Versuchen war der wichtigste Speicher von beta-Karotin im Körper des Rindes das Blutplasma. An zweiter Stelle stand die Leber und an dritter Stelle das Fettgewebe (Friesecke 1978). Die Anreicherung im Fettgewebe ist, wie bereits erwähnt, bei Rind und Pferd besonders offensichtlich, da es bei diesen Tieren durch das beta-Karotin gelb gefärbt ist. Dies führt auch zu der Bezeichnung Gelbfett-Tiere. Die größte beta-Karotin-Konzentration findet man im Gelbkörper, der dadurch seine Farbe und auch seinen Namen erhält. Durch seine geringe Masse fällt der betaKarotin-Gehalt des Gelbkörpers mengenmäßig nicht ins Gewicht. Die beta-KarotinGehalte in den Corpora lutea, in der Leber und im Fettgewebe lagen bei einer karotinarm ernährten Versuchsgruppe signifikant niedriger als diejenigen einer mit beta-Karotin ausreichend versorgten Kontrollgruppe (Ahlswede u. Lotthammer 1978). 2.8.2 Leber In Untersuchungen, die den Gehalt an Vitamin A und beta-Karotin in der Leber verglichen, wurde festgestellt, dass im Lebergewebe etwa siebenmal mehr Vitamin A als beta-Karotin enthalten war. Außerdem variierte der Gehalt in den einzelnen 23 Bioptaten derselben Leber stark (Koopman et al. 1971). Daraus ging hervor, dass beta-Karotin vom Rind wesentlich schlechter gespeichert wurde als Vitamin A. Es hat sich ebenfalls gezeigt, dass sowohl Vitamin A als auch beta-Karotin im Lebergewebe nicht gleichmäßig verteilt vorlagen. Dies ist bei der Interpretation der Ergebnisse von Leberbiopsien zu beachten. Außerdem hingen die Konzentrationen von beta-Karotin und Vitamin A im Lebergewebe von der täglichen Zufuhr beider Stoffe ab. 2.8.3 Milch In einem Versuch erhöhte eine beta-Karotin-Zulage den beta-Karotin-Gehalt in der Milch. Die beta-Karotin-Konzentration in der Milch hing eng mit der Blutkonzentration zusammen (Zucker et al. 1980). Zwei bis drei Wochen vor der Abkalbung begann die Bildung des beta-Karotin-reichen Kolostrums. Damit war ein deutlicher Abfall der beta-Karotin-Konzentration im Blut verbunden. Die Kolostralmilch enthielt 10 mal mehr beta-Karotin bzw. 5 mal mehr Vitamin A als die normale Milch. Innerhalb von 7 Tagen p.p. hatten sich diese Gehalte in der Milch wieder normalisiert. Das Kolostrum dient, neben seiner Funktion als Immunglobulinquelle, vermutlich der schnellen Versorgung der neugeborenen Kälber mit beta-Karotin und Vitamin A, da diese Substanzen die Plazentaschranke nur in sehr geringem Ausmaß durchschreiten können und der Blutspiegel bei Neugeborenen weit unter dem der Muttertiere liegt (Kolb u. Seehawer 1998). Untersuchungen zur Beziehung zwischen beta-Karotin im Blut und in der Milch bei Kühen zeigten, dass der beta-Karotin-Gehalt im Blut von der Laktationsnummer und dem Laktationsstadium abhängt. Je älter die Kuh und je fettreicher die Milch um so mehr beta-Karotin war in der Milch enthalten (Stampfer 1982). Leidl et al. (1987) kamen zu ähnlichen Ergebnissen. Zusätzlich sinkt der betaKarotin-Gehalt in der Milch bei hoher Milchleistung. Je höher der Milchfettgehalt desto höher war die beta-Karotin-Konzentration. Bei Kühen mit subklinischer Ketose stieg der Milchfettgehalt und somit auch der beta-Karotin-Gehalt an. Die betaKarotingehalte wurden ebenfalls durch die geringere Milchleistung der kranken Kühe erhöht (Larson et al. 1983). 24 2.9 Resorption nach parenteraler Gabe Eisele (1987) stellte fest, dass beta-Karotin sich nach parenteraler Gabe in gleicher Weise wie nach oraler Aufnahme auf die Lipoproteinfraktionen verteilt. Nur kurz nach der Applikation waren die lipidreichen Lipoproteinfraktionen (VLDL, LDL) vermehrt beladen. Dies beruhte wahrscheinlich auf der raschen Bindung an diese Fraktionen aufgrund der lipophilen Eigenschaften des beta-Karotins. Bei intravenöser Gabe von 100 mg beta-Karotin war es bereits nach vier Stunden vollständig an die Lipoproteine gebunden. Diese Gabe führte für zehn Tage zu einer Verdopplung des beta-KarotinSpiegels im Serum. Nach intramuskulärer Injektion von 500 mg beta-Karotin erhöhte sich der Gehalt im Blutserum im Verlauf von 26 Stunden von anfangs 910 ± 320 µg/l auf 10070 ± 1340 µg/l. Diese Verzehnfachung blieb für 10 Tage bestehen. Nach drei Wochen lagen die Serumkonzentrationen noch über 3000 µg/l. Die parenterale Gabe von beta-Karotin führte zusätzlich zu einem erhöhten beta-Karotin-Gehalt der Milch. Die vollständige Bindung des beta-Karotins an die Lipoproteine und die gesteigerte beta-Karotin-Konzentration in der Milch lassen den Schluss zu, dass parenteral verabreichtes beta-Karotin vom Organismus des Rindes genutzt werden kann (Eisele 1987). Ähnliche Versuche mit Schafen zeigten ebenfalls, dass diese parenteral (intramuskuläre Injektion) verabreichtes beta-Karotin verwerten konnten. Es kam ebenfalls zu gesteigerten beta-Karotin-Konzentrationen im Blut und in der Milch (Özpinar et al. 1995). 2.10 Beta-Karotin im Blut 2.10.1 Untersuchungsmethoden Der Blutserumgehalt an beta-Karotin lässt sich semiquantitativ mit Hilfe einer Farbtafel schätzen. Blassweißes Serum enthält unter 1500 µg/l, blassgelbes Serum enthält etwa 2500 µg/l und goldgelbes Serum enthält über 3500 µg/l beta-Karotin. 25 Dieses Verfahren eignet sich relativ gut, um einen schnellen Überblick über die Versorgung eines Tieres zu erhalten. Vergleichstudien mit einer quantitativen Methode (Photometrie) haben ergeben, dass man die Gehalte im allgemeinen etwas zu hoch einschätzt (Leidl et al. 1987). Die Spektrophotometrie ist ein quantitatives Verfahren. Dabei wird die Intensität von monochromatischem Licht beim Durchgang durch Serum-/Plasmaproben gemessen. Je gelber (karotinreicher) die Probe umso mehr Licht wird absorbiert und um so schwächer ist die Intensität des durchgegangenen Lichts. Sie liefert gute Ergebnisse unter der Voraussetzung, dass das Serum nicht hämolytisch ist. In neuerer Zeit wird der beta-Karotin-Gehalt von Serum oder Plasma über die HPLC (High Pressure Liquid Chromatography = Hochdruck-Flüssigkeits-Chromatographie) bestimmt (Cetinkaya u. Ozcan 1991, Chawla u. Kaur 2001). 2.10.2 Beta-Karotin-Gehalte im Blut von Milchkühen Im Allgemeinen wird ein beta-Karotin-Gehalt im Blutserum von Milchkühen von über 3000 µg/l angestrebt (Herdt u. Stowe 1991). In jedem Fall sollen Werte von 1000 µg/l beta-Karotin im Blutserum überschritten werden (Lotthammer 1999a). In Tabelle 4 sind einige Referenzwerte für den beta-Karotin-Gehalt im Blut von Milchkühen angegeben. 26 Tabelle 4: Referenzwerte für beta-Karotin im Blut von Milchkühen Autor Beta-Karotin µg/l Bemerkungen Friesecke (1978) 3000 Herdt u. Stowe (1991) Smith (1996) über 3000 1500-3970 unter 700 3000-5000 Mindestgehalt, bei geringeren Werten beginnt Mangel erstrebenswert normal Mangel Rind, karotinreiche Fütterung normal Fruchtbarkeit beeinträchtigt kritischer Bereich absoluter Mangel 3 Wochen ante partum bis 3 Wochen post partum übriger Zeitraum Kolb u. Seehawer (1997) Alvetra (Hersteller von Carofertin 1998) Lotthammer (1999) über 5000 unter 4000 2000 1000 über 1000 über 2000 Da der Gehalt an beta-Karotin im Blut von der Fütterung abhängt, kam es früher während der Winterfütterung zu einem erheblichen Absinken der Blutspiegel, weil Silagen selten eingesetzt wurden und Heu während der Lagerung viel an betaKarotin verliert. Man fand heraus, dass Tiere, die Grassilage bekommen, vergleichbare beta-Karotin-Serumgehalte wie Tiere auf der Weide aufweisen (Jukola et al. 1996b). Bei Untersuchungen in Kanada wurde eine positive Korrelation zwischen beta-Karotin-Gehalten im Futter, im Plasma und in der Milch nachgewiesen (Block u. Farmer 1987). Die größte beta-Karotin-Quelle war in diesem Versuch die Grassilage. Die Autoren wiesen darauf hin, dass bei Untersuchungen zur Versorgungslage der Kühe auch immer der Vitamin-A-Status der Herde erhoben werden sollte. Zusätzlich betonten sie, dass beta-Karotin bei einer nach modernen Grundregeln zusammengestellten Ration so gut wie nie der limitierende Faktor sei. Die positive Korrelation zwischen beta-Karotin-Gehalten im Futter und im Plasma bzw. Serum gilt als gesichert. 27 2.10.3 Individuelle Schwankungen Auffällig ist, dass es bei den Tieren einer Herde, die dasselbe Futter erhalten, zu großen individuellen Unterschieden in der Höhe der beta-Karotin-Werte im Blut kommt. Bei Versuchen, die den Blutserumgehalt von beta-Karotin über mehrere Wochen verfolgten, wurden neben großen individuellen Unterschieden auch periodische Schwankungen festgestellt, bei denen alle 6-8 Wochen Maximal- bzw. Minimalwerte auftraten. Aus diesen Untersuchungen schloss man, dass Kühe ein individuell bedingtes, unterschiedliches Resorptions- und Mobilisierungsvermögen besitzen (Ahlswede u. Lotthammer 1978). Ein Zusammenhang mit dem Blutcholesterinspiegel, der mit dem beta-Karotin-Spiegel positiv korrelierte, und der Schilddrüsenfunktion wurde ebenfalls vermutet (Lotthammer u. Ahlswede 1977, Lotthammer 1979). Über die Verbindung zwischen Schilddrüsenfunktion und betaKarotin beim Rind gibt es nur wenige Untersuchungen. Ein Versuch hat ergeben, dass beta-Karotin die Aktivität der 5´-Monodeiodinase in der Leber verstärkt und dadurch die Bildung von Triiodthyronin fördert. Die Autoren betonten, dass noch weitere Untersuchungen über den Schilddrüsenhormonspiegel und –stoffwechsel nötig sind, um die Rolle des beta-Karotins umfassender zu klären (Pethes et al. 1985b). Bei Versuchen mit Kühen ist zu beachten, dass die Genetik der einzelnen Tiere sehr unterschiedlich ist. Dieser Umstand begünstigt das Auftreten individueller Unterschiede. Will man eine Aussage über die beta-Karotin-Versorgung einer Herde machen, sollte immer eine genügend große Probenanzahl gewählt werden, um den Einfluss der individuellen Schwankungen zu mindern. 2.10.4 Blutspiegel im peripartalen Zeitraum Als gesichert gilt, dass die beta-Karotin- und Vitamin-A-Plasmaspiegel ante partum stark abfallen (Burgstaller et al. 1980). Dies ist auf die beginnende Kolostrumproduktion zurückzuführen. Es dauerte Wochen bis beim Muttertier die Ausgangswerte wieder erreicht wurden. Untersuchungen des Trockenstehersekrets 28 bestätigten, dass kurz vor der Geburt der Gehalt an beta-Karotin und Vitamin A stark anstieg (Krieger u. Schweigert 1993). Man fand eine positive Korrelation zwischen den beta-Karotin-Werten im Blut des Muttertieres, im Blut des Kalbes und in der Kolostralmilch (Fadle 1966). Kolb (1998) brachte dies auch mit der vieldiskutierten peripartalen Immunsuppression der Milchkühe in Verbindung. Es scheint ebenso einen Zusammenhang zwischen peripartaler Lipomobilisation und den Blutserumwerten zu geben. Bei Tieren, die peripartal gesteigert Fette mobilisierten, fand ein Abfall der beta-Karotin-Konzentration im Blut um fast 50 % statt. Bei gesunden Tieren betrug der Abfall circa 20 %. Außerdem dauerte es länger bis die Ausgangswerte wieder erreicht wurden. Die Schlussfolgerung ist, dass die gesteigerte Lipomobilisation den Transport des beta-Karotins im Blut stört, wodurch offensichtlich der stärkere Abfall der Werte und die verlängerte Phase niedriger betaKarotin-Gehalte im Blut verursacht werden. Dies könnte wiederum eine Rolle spielen bei der Verzögerung des Einsetzens eines normalen Zyklus bei energetisch schlecht versorgten Tieren (Haraszti et al. 1984). 2.11 Beta-Karotin als Provitamin A 2.11.1 Vitamin-A-Stoffwechsel Vitamin A ist der Oberbegriff für eine Reihe natürlicher und synthetischer Verbindungen, die qualitativ die biologische Aktivität des Retinols (Vitamin-A-Alkohol) aufweisen. Dazu gehören neben dem Retinol das Dehydroretinol, das Retinal (Vitamin-A-Aldehyd) und die Retinsäure (besitzt nur einen Teil der Vitamin-AWirkung). Die Ester des Retinols (Retinylacetat, -palmitat, -propionat) besitzen ebenfalls Vitamin-A-Aktivität (Flachowsky 1999). Vitamin A kommt nur in Futtermitteln tierischer Herkunft vor. Reine Pflanzenfresser wie Rinder sind also darauf angewiesen, es aus Vorstufen (vor allem dem beta-Karotin), die sie mit der Nahrung aufnehmen, selbst zu synthetisieren. Die Umwandlung des Karotins in Vitamin A erfolgt vor allem in der Dünndarmschleimhaut. Dort wird es durch ein 29 spezifisches Enzym, die beta-Karotin-15,15`-Dioxygenase, in Vitamin-A-Aldehyd gespalten (Hanck u. Kuenzle 1991). Dieses Enzym wird auch beta-Carotinase genannt. Dann wird das Retinal an Chylomikronen bzw. VLDL gebunden und gelangt über die Lymphe ins Blut. In der Milchdrüse (Brüggemann u. Niesar 1955), der Leber, der Niere (Olson u. Hayaishi 1965) und dem Ovar (Sklan 1983) ist ebenfalls beta-Carotinase-Aktivität vorhanden, so dass auch dort Vitamin A gebildet wird. Das beta-Karotin wird im Gelbkörper in Retinol umgewandelt. Es wird aber weder verestert noch gespeichert. Beta-Karotin dient somit als lokale Vitamin-A-Quelle. In vitro entstand Retinol bei der Inkubation von Gelbkörpergewebe mit beta-Karotin (Stacewicz-Sapuncakis et al. 1975). Da diese Umwandlung unter 20°C und nach Aufkochen des Materials aufhörte, nahm man an, dass diese Umwandlung enzymatisch ablief (Gawienowski et al. 1974). Wie bereits erwähnt, wird beim Rind im Mittel mit einer Umwandlung von 1 mg beta-Karotin in 400 IE Vitamin A kalkuliert (NRC 1989). Dieser Wert lag bei höherer beta-Karotin-Versorgung (z.B. 1 g/Tier/Tag) deutlich niedriger und bei reduziertem Karotinangebot höher (Cetinkaya u. Ozcan 1991). Burgstaller et al. (1980) zeigten, dass der Vitamin-A-Gehalt im Plasma ab einem Wert von 1200 I.E./l auch bei Erhöhung der beta-Karotin-Zufuhr nicht mehr steigt. Die Vitamin-A-Konzentration in den Depots (v.a. Leber), der physiologische Status (laktierend/tragend), die Leistungshöhe, verschiedene Futterinhaltsstoffe (NO3, Zellwandbestandteile) und die Rationsgestaltung bzw. die Verhältnisse im Pansen haben Einfluss auf den Umfang der beta-Karotin-Passage durch den Pansen und die Umwandlung von beta-Karotin in Vitamin A (Schlenzig et al. 1988, Weiss 1998). Diese komplexen Zusammenhänge erklären die Probleme, die bei dem Versuch auftreten, den Bedarf einer Kuh an beta-Karotin bzw. Vitamin A abzuleiten. 2.11.2 Einige Funktionen des Vitamin A im Organismus Den Namen Retinol trägt das Vitamin A wegen seiner Notwendigkeit für die Funktion der Retina (Netzhaut). So führt ein Vitamin-A-Mangel beim Menschen z.B. zu Nachtblindheit. Vitamin A wird nach Aufnahme in die Körperzellen meist in 9-cisoder all-trans-Vitamin-A-Säure (Retinsäure) überführt. Nach Bindung an Rezeptoren 30 wird es in den Zellkern eingeschleust und stimuliert dort die Transkription von zahlreichen Genen, die u.a. für Wachstum und Entwicklung notwendig sind (Kolb u. Seehawer 1997). Es wird wegen der Förderung der Sekretion von Wachstumshormon auch als Wachstumsvitamin bezeichnet. Bei Mangel an Vitamin A ist das Wachstum gehemmt, die Geschlechtsreife tritt verspätet auf und die Leistungsfähigkeit des Immunsystems ist herabgesetzt. Darauf führte man zurück, dass bei Vitamin-A-Mangel Endometritis und Mastitis gehäuft vorkamen (Kolb 1994). Vitamin-A-Mangel wird mit Fruchtbarkeitsproblemen in Verbindung gebracht. Man war lange Zeit der Meinung, dass die bei mangelhafter beta-Karotin-Versorgung auftretenden Fruchtbarkeitsprobleme auf einen sekundären Vitamin-A-Mangel zurückzuführen waren. Vitamin A soll bei der Bildung eines Enzymkomplexes zur Synthese der Steroidhormone im Ovar eine Rolle spielen. Außerdem wurde es mit der Bildung von proteolytischen Enzymen im Follikel in Verbindung gebracht. Diese Enzyme sollen für den Abbau der Follikelmembran als Voraussetzung für die Ovulation verantwortlich sein (Zerobin 1987). 2.12 Beta-Karotin und Fruchtbarkeit 2.12.1 Beta-Karotin und die Fruchtbarkeitsparameter Vor allem in der deutschsprachigen Literatur wurde Mitte der 70iger Jahre immer wieder postuliert, dass beta-Karotin eine spezifische, Vitamin-A-unabhängige Wirkung auf die Fertilität des Rindes hat. Es wurde berichtet, dass beta-Karotin die Fruchtbarkeit verbessert, auch wenn die Ration genügend Vitamin A enthält. Früher ging man nämlich davon aus, dass die Zuführung von genügend Retinylestern die Fruchtbarkeitsprobleme, die bei Vitamin-A-Mangel auftraten, beheben würde. Im Gegensatz dazu nahm man an, dass es einen Bedarf an beta-Karotin gibt, der nicht durch die alleinige Zuführung von Vitamin A gedeckt werden kann. Es gab in den letzten Jahrzehnten zahlreiche Versuche, die sich mit der Frage befassten, ob betaKarotin die Fruchtbarkeit von Milchkühen beeinflusst oder nicht. Viele Autoren haben 31 keinen Effekt auf die Fruchtbarkeit feststellen können (Akordor et al. 1986, Bindas et al. 1984, Bremel et al. 1982, Ducker et al. 1984, Folman et al. 1979, Greenberg et al. 1986, Jukola et al. 1996a, Tekpetey et al. 1987, Wang et al. 1987, Wang et al. 1988a). Andere stellten einen positiven Effekt fest (Ahlswede u. Lotthammer 1978, Ascarelli et al. 1985, Dembinski u. Bronicki 1994, Dembinski u. Bronicki 1996, Hasselmann et al. 1999, Heinz u. Herzog 1982, Inaba et al. 1986b, Jackson 1981, Lotthammer et al. 1976, Lotthammer u. Ahlswede 1977, Lotthammer 1979, Meyer et al. 1975, Rakes et al. 1985, Schams et al. 1977, Wang et al. 1982). Ein Autor berichtete, dass beta-Karotin die Fruchtbarkeit des Rindes negativ beeinflusst (Folman et al. 1987). Einige Untersucher stellten Verbesserungen einzelner Fruchtbarkeitsparameter fest, z.B. eine besser ausgeprägte Brunst oder tendenziell weniger Besamungen pro Trächtigkeit (Stolla et al. 1987). Außerdem wurden bei einer mit beta-Karotin ausreichend versorgten Versuchsgruppe höhere tägliche Zunahmen der Rinder und ein verkürzter Zeitraum zwischen LH-Peak und Ovulation festgestellt als bei einer mit beta-Karotin unterversorgten Kontrollgruppe. Die Zykluslänge, die Progesteronproduktion und die Besamungen pro Trächtigkeit blieben unbeeinflusst (Tekpetey et al. 1987). Eine Autorengruppe stellte einen Effekt im Sommer (Hitzestreß) fest. Bei einer kontinuierlichen Zufütterung von 400 mg/Tier/Tag verbesserte beta-Karotin die Fruchtbarkeitsrate bei terminierter Besamung sowie die Milchleistung. Auf die gesamte Versuchsdauer gesehen, hatte das beta-Karotin allerdings keinen Einfluss (Arechiga et al. 1998a). Im selben Jahr veröffentlichte dieser Autor eine Untersuchung, bei der beta-Karotin injiziert wurde (3 mal 800 mg: 6 Tage vor der Besamung, 3 Tage vor der Besamung + Prostaglandin F2α, am Tag der Besamung). Die Versuchstiere waren außerdem einem heißen Klima ausgesetzt. Hier hatte das beta-Karotin keinen Einfluss auf die Konzeptionsrate (Arechiga et al. 1998b). Eine Untersuchung in Deutschland ergab, dass ein Anstieg der beta-Karotin-Konzentration im Blut in den ersten 40 Tagen nach der Abkalbung zu einer signifikanten Verkürzung der Güst- und Rastzeit sowie zu einem im Durchschnitt verringerten Auftreten von Ovar- und Puerperalstörungen führte (Failing et al. 1998, Fischer 1996). Allerdings wurde diese Untersuchung an Datenmaterial aus 15 verschiedenen Betrieben durchgeführt. Insgesamt gesehen 32 liegen zu dem Thema beta-Karotin und Fruchtbarkeit sehr viele Untersuchungen mit sehr verschiedenen Versuchsanordnungen vor. Diese Unterschiede bestehen in dem Alter der Versuchstiere, der Zahl der Versuchstiere, der Haltung, dem Laktationsstadium, der Dauer der Zuführung des beta-Karotins, der Art der Applikation, der Zufütterung von Vitamin A, der Menge an verabreichtem betaKarotin und der Dauer der Fütterung der Kontrollgruppe mit beta-Karotin-armem bzw. beta-Karotin-freiem Futter (Hurley u. Doane 1989). Vor allem die Autoren aus Nordamerika konnten keinen positiven Effekt einer beta-Karotin-Supplementierung feststellen. Wenn das beta-Karotin die Fruchtbarkeit positiv beeinflusste, führte es im Vergleich zu einer karotinarm bzw. karotinfrei ernährten Kontrollgruppe zu einer gesteigerten Brunstintensität, höheren Konzeptionsrate, verringerten Anzahl an Besamungen pro Trächtigkeit, Verkürzung der Güstzeit und weniger Ovarialzysten (Ascarelli et al. 1985, Lotthammer 1979). Es ist zu beachten, dass die Autoren, die positive Effekte erzielten, diese positiven Effekte gegenüber einer Kontrollgruppe fanden, der kein beta-Karotin zugeführt wurde. Bei Untersuchern, die keine Wirkung feststellten, war das Futter der Kontrollgruppe häufig von etwas besserer Qualität als bei Untersuchern, die einen positiven Effekt durch das beta-Karotin erzielten. Lotthammer et al. (1976) betonten, dass ein beta-Karotin-Defizit in der Praxis nur unter extremen Bedingungen als alleinige Ursache für ein bestandsweise auftretendes Fruchtbarkeitsproblem in Frage kommt. Diese Auffassung wird von anderen Untersuchern bestätigt (Block u. Farmer 1987, De Kruif u. Mijten 1992). Es wird ebenfalls darauf hingewiesen, dass zuerst der Vitamin-A-Metabolismus vollständig geklärt sein muss, bevor beta-Karotin für eine optimale Fruchtbarkeit des Rindes als essenziell erklärt werden kann (Hemken u. Bremel 1982). Bei Problemen mit der Herdenfruchtbarkeit, die man im Bereich der Fütterung vermutet, sollte man zuerst an ein fehlerhaftes Energiemanagement, dann an einen Proteinüberschuss und zuletzt an einen Spurenelement- und Vitaminmangel denken (Ferguson 1996). Beta-Karotin sei keinesfalls ein Ersatz für gutes Herdenmanagement oder eine bedarfsorientierte Fütterung (Kessler et al. 1991). 33 2.12.2 Beta-Karotin und die Gravidität Ahlswede und Lotthammer (1978) fanden heraus, dass die Sekretionsphase im Uterus bei zyklischen Tieren bei beta-Karotin-Mangel verzögert eintrat und schloss daraus, dass dies die befruchtete Eizelle beeinträchtigt und zu frühembryonalem Fruchttod führen kann. Zu der Versuchsgruppe gehörten 12 Tiere und zur Kontrollgruppe 6. Lotthammer (1979) berichtet ebenfalls über die Verringerung der embryonalen Sterblichkeit und dem selteneren Auftreten von Aborten in der Frühträchtigkeit. Weigelt et al. (1988) konnten dies allerdings nicht bestätigen. Die embryonale Mortalität wurde vielmehr durch Energiemangel- und Stresssituationen beeinflusst. Durch seinen Einfluss auf die Gelbkörperdynamik (Erhöhung der Progesteronproduktion) spricht man dem beta-Karotin auch in neuerer Zeit einen positiven Einfluss auf die Aufrechterhaltung einer Gravidität zu (Aslan et al. 1998). Versuche mit Embryonen zeigten, dass beta-Karotin für die Sekretion eines Bindungsproteins von Bedeutung ist, das den Retinoltransport zum Embryo unterstützt und so indirekt das Wachstum des Embryos fördert (Ashworth 1994). 2.12.3 Über Beta-Karotin und Retentio secundinarum den Zusammenhang zwischen beta-Karotin und der Inzidenz von Nachgeburtsverhaltungen liegen keine gesicherten Erkenntnisse vor. Es wurden zwar bei Tieren mit Nachgeburtsverhaltung besonders im Winter niedrigere betaKarotin-Werte festgestellt als bei Tieren ohne Nachgeburtsverhaltung, die Unterschiede waren aber nur schwach signifikant (Inaba et al. 1986a, Spindler 1980). 34 2.13 Beta-Karotin und das Ovar 2.13.1 Allgemeines Beta-Karotin spielt wahrscheinlich eine Rolle in der Ovarfunktion, die nicht durch zugefüttertes Vitamin A ersetzt werden kann. Es hat zahlreiche Untersuchungen zu diesem Thema gegeben. Einige Autoren sind zu dem Schluss gekommen, dass beta-Karotin einen positiven Einfluss auf die Ovarfunktion besitzt (Ascarelli et al. 1985, Cooke u. Comben 1978, Graves-Hoagland et al. 1989). Diese positiven Effekte zeigten sich z.B. Gelbkörpergewebes, in im einer gesteigerten selteneren Auftreten Progesteronproduktion von Ovarialzysten und des einer Vergrößerung der Funktionskörper. Auch hier waren die Versuchsanordnungen sehr unterschiedlich. 2.13.2 Beta-Karotin-Gehalte Beim Rind waren beta-Karotin, Retinol und Retinylester in den Follikeln und im Gelbkörper in höherer Konzentration enthalten als in Schweinefollikeln bzw. Schweinegelbkörpern. Außerdem waren die Gehalte dieser Substanzen in den Rinderfollikeln positiv korreliert mit den Gehalten im Plasma. Im Gelbkörper war nur das Retinol positiv korreliert mit den Plasmagehalten (Chew et al. 1984). In einer aktuellen Untersuchung an Schlachtorganen wurde eine positive Korrelation zwischen dem beta-Karotin-Gehalt im Plasma und der Follikelflüssigkeit bzw. dem Gelbkörpergewebe gefunden. Zusätzlich war der beta-Karotin-Gehalt der Follikelflüssigkeit mit dem des Gelbkörpers positiv korreliert. Außerdem beeinflußte die beta-Karotin-Menge den Durchmesser und das Gewicht des Gelbkörpers positiv. Der beta-Karotin-Gehalt der Follikelflüssigkeit Follikeldurchmesser korreliert (Haliloglu et al. 2002). war negativ mit dem 35 2.13.3 Ovarialzysten Lotthammer et al. (1976) beobachteten bei karotinfrei ernährten Versuchstieren eine um rund einen Tag verzögerte Ovulation im Vergleich zu ausreichend versorgten Tieren. Ahlswede und Lotthammer (1978) stellten fest, dass bei Färsen mit zystösen Veränderungen die beta-Karotin-Gehalte der Ovarien am niedrigsten lagen. Allerdings wurden bei diesen Versuchen nur 20 Tiere verwendet. Die Zahl der Färsen mit Ovarialzysten lag bei drei. Die Veränderungen, die durch den betaKarotin-Mangel hervorgerufen wurden, waren alle reversibel, das heißt nach mehrwöchiger Fütterung ausreichender beta-Karotin-Mengen funktionierten die Ovarien wieder normal. In einer späteren Untersuchung mit 97 Tieren fand man keinen Zusammenhang zwischen der Inzidenz von Ovarialzysten und der betaKarotin-Zufütterung. Ebenso wurde der Behandlungserfolg von GnRH durch Zulage von beta-Karotin nicht verbessert. Da die Plasmawerte von Versuchs- und Kontrollgruppe über 3000 µg/l lagen, schloss man daraus, dass es beim Vorliegen von Ovarialzysten und ausreichender Karotinversorgung keine Vorteile bringt, betaKarotin über das Futter zu supplementieren (Marcek et al. 1985). Inaba et al. (1986b) stellten fest, dass der beta-Karotin-Gehalt im Blut bei Tieren mit Ovarialzysten niedriger lag als bei Tieren ohne Ovarialzysten. Beide Gruppen erhielten dieselbe Menge beta-Karotin über das Futter. Beim Vergleich von Schwarzbunten und Braunviehkühen zeigte sich, dass bei Braunvieh ein Zusammenhang zwischen der beta-Karotin-Konzentration im Blut und der Inzidenz von Ovarialzysten besteht. Bei Schwarzbunten gab es keinen Zusammenhang. Allerdings wurden bei den Braunviehkühen mit Ovarialzysten im Durchschnitt Werte um 2000 µg/l und bei denen ohne Ovarialzysten Werte um 3100 µg/l gemessen. Bei den Schwarzbunten hatten alle Tiere Werte über 3000 µg/l (Özpinar et al. 1988). 36 2.13.4 Progesteronproduktion und beta-Karotin-Gehalte im Ovar Ahlswede und Lotthammer (1978) fanden bei ihren Untersuchungen bei Tieren mit beta-Karotin-Mangel schwach signifikant geringere Progesteronkonzentrationen im Gelbkörpergewebe als bei ausreichend versorgten Tieren. Im Sommer zeigte sich allerdings ein steiler Anstieg der Progesteronkonzentration im Gelbkörpergewebe bis zur Zyklusmitte und danach ein steiler Abfall bis zum Zyklusende. Im Winter erfolgte ein verzögerter Anstieg, was zur Verschiebung des Maximums führte, gefolgt von einem flachen Abfall. Zwischen den Progesterongehalten der Gelbkörper aus Sommer- und Winterperiode bestand kein signifikanter Unterschied. Der durchschnittliche beta-Karotin-Gehalt der Frischsubstanz der Gelbkörper stieg zum Ende des Zyklus an, was durch einen geringeren Wassergehalt erklärt wurde. Dieser Vorgang war von der Jahreszeit unabhängig. Die beta-Karotin-Gehalte der Gelbkörper waren fütterungsbedingt im Sommer hochsignifikant höher als im Winter. Man schloss daraus, dass der langsame Anstieg der Progesteronkurven im Winter auf eine geringere Progesteronsekretion aufgrund des beta-Karotin-Mangels zurückzuführen sei (Anwandter Quentin 1974). Andere Untersuchungen bestätigten, dass die beta-Karotin-Konzentration im Gelbkörper von der beta-Karotin-Versorgung abhing. Die Masse der Gelbkörper blieb von der Karotinzulage unbeeinflusst. Es wurden in den Ovarien außerhalb der Gelbkörper orangefarbene Pigmente gefunden, die eine bis zu 70% höhere beta-Karotin-Konzentration aufwiesen als der Gelbkörper. Die Funktion dieser Pigmente konnte nicht geklärt werden (Kirsche et al. 1987). 2.13.5 Progesteronproduktion und beta-Karotin-Gehalte im Blut Dembinski et al. (1996) bewirkten eine Erhöhung des Progesteronspiegels im Blut durch eine beta-Karotin-Zulage über das Futter. Dies konnte von anderen Autoren nicht bestätigt werden, wenn der Blutserumspiegel des beta-Karotin über 1500 µg/l lag (Bonsembiante et al. 1980, Ducker et al. 1981, Folman et al. 1983). Es lagen 37 feine Unterschiede in der Reaktion der Gelbkörper von mit beta-Karotin behandelten Tieren auf die Injektion von Humanem Chorion Gonadotropin (hCG) im Vergleich zu Kontrolltieren vor. Der Plasmaprogesterongehalt stieg nach der hCG-Injektion steiler an, glich sich aber im späteren Verlauf an die Werte der Kontrollgruppe an. Daraus schloss man, dass beta-Karotin möglicherweise Einfluß auf die endokrinen Vorgänge nimmt (Bindas et al. 1984). Diese Ergebnisse wurden von anderen Untersuchern bestätigt (Pethes et al. 1985a). Bei In-vitro-Versuchen mit Follikeln enthielten diese bei Kultivierung mit beta-Karotin weniger Östradiol als ohne beta-Karotin. Für Progesteron waren die Verhältnisse umgekehrt. Daraus wurde ebenfalls gefolgert, dass beta-Karotin die Steroidsynthese beeinflußt. Der genaue Angriffspunkt im Syntheseweg blieb ungeklärt (Bittner 1986). In-vitro-Versuche mit Gelbkörpergewebe ergaben, dass bei Gelbkörpern, die im Winter entnommen wurden (niedrigere betaKarotin-Werte) eine positive Korrelation zwischen der Progesteronproduktion in vitro und der beta-Karotin-Plasmakonzentration in vivo bestand. Bei im Sommer gewonnenen Gelbkörpern (hohe beta-Karotin-Werte) gab es diese Korrelation nicht (Graves-Hoagland et al. 1988). Haliloglu et al. (2002) zeigten, dass der Progesterongehalt im Plasma positiv mit dem beta-Karotin-Spiegel im Plasma korreliert ist. Die gleiche Korrelation liegt zwischen dem Progesterongehalt im Plasma und dem beta-Karotin-Spiegel in der Follikelflüssigkeit und dem beta-KarotinGehalt des Gelbkörpers vor. Bei den Versuchen von Schams et al. (1977) hatte eine Karotinmangelernährung keinen signifikanten Einfluss auf den Progesteronspiegel im Blut. Die Absolutwerte der präovulatorischen LH-Gipfel waren in Versuchs- und Kontrollgruppe ebenfalls nicht signifikant unterschiedlich. Es wurde lediglich eine verzögerte Ovulation festgestellt. Bei Wang et al. (1988) veränderte eine betaKarotin-Supplementierung weder die Progesteronproduktion noch die Gelbkörpergröße. Die LH-Produktion und die LH-Antwort nach GnRH-Gabe blieben ebenfalls unbeeinflusst. 38 2.13.6 Beta-Karotin in der Zelle Inaba et al. (1987) kultivierten Granulosazellen in vitro und stellten bei Zusatz von beta-Karotin eine vermehrte Progesteronproduktion fest. Bei Zusatz von Vitamin A ging die Progesteronproduktion zurück. Bei einem Versuch, der sich mit dem betaKarotin-Gehalt von Milchfettglobuli befasste, fanden Patton et al. (1980) als Nebenbefund heraus, dass intrazelluläre Membranen wie die der Mitochondrien, die der Mikrosomen und des Golgi-Apparates in einer höheren Konzentration betaKarotin enthielten als die Milchfettglobuli. Dann untersuchte man, in welchen Bestandteilen der Gelbkörperzellen das beta-Karotin vorkommt und wie fest es dort gebunden ist. An den Zellkern und die Mitochondrien war es nur lose gebunden, aber aus der Mikrosomenfraktion lies es sich nur schwer entfernen. Somit ist beta-Karotin wahrscheinlich ein Bestandteil der Mikrosomenmembran (Patton et al. 1980). Im Zytosol kam es an Proteine mit einem hohem Molekulargewicht gebunden vor (O'Fallon u. Chew 1984). Neuere Untersuchungen zeigten, dass beta-Karotin fest an die progesteronproduzierende Mikrosomenfraktion gebunden ist (Holt et al. 1995). Rapoport et al. (1998) fanden heraus, dass beta-Karotin die Bildung von Progesteron in der Mikrosomenfraktion der Gelbkörperzellen steigert. Dies wird durch neuere Studien bestätigt (Arikan u. Rodway 2000a, Arikan u. Rodway 2000b). 2.13.7 Vitamin-A-unabhängige Aufgabe des beta-Karotins im Ovar Bei Inkubation isolierter Gelbkörperzellen von Sauen mit beta-Karotin in einer Konzentration von nur 0,1 µmol/l war die Bildung von Progesteron etwa um das sechsfache größer als bei einer Inkubation mit einer äquivalenten Menge an Vitamin A (Talavera u. Chew 1988). Diese Ergebnisse zeigen, dass beta-Karotin tatsächlich eine Vitamin-A-unabhängige Wirkung am Ovar besitzt. Die genaue Wirkungsweise war damit immer noch unklar. Beta-Karotin gehört in die Gruppe der Antioxidantien, deren Aufgabe darin besteht, die Körperzellen vor freien Radikalen zu schützen. Diese entstehen bei vielen enzymatischen Reaktionen. Der größte Teil des in die 39 Zellen aufgenommenen Sauerstoffs wird zum Beispiel zur Oxidation von Wasserstoff zur Gewinnung von ATP in den Mitochondrien verbraucht. Bei diesen Reaktionen werden Sauerstoffradikale und andere reaktionsfähige Sauerstoffverbindungen gebildet. Diese Radikale werden durch Antioxidantien abgefangen, bevor sie an den Zellmembranen oder der DNS Schäden anrichten. Peripartal auftretender oxidativer Stress (vermehrtes Auftreten von freien Radikalen) kann sich auf die Entstehung von Euterödemen, Nachgeburtsverhaltung, Milchfieber, Fruchtbarkeitsproblemen und Mastitis negativ auswirken (Miller et al. 1993). Um dem vorzubeugen, wurde vorgeschlagen, Antioxidantien (Vitamin A, C, E, beta-Karotin, Gluthathion) in ausreichender Menge dem Futter zuzusetzen. Die genau benötigten Mengen dieser Substanzen sind noch unbekannt. Beta-Karotin wird über die Bindung an Lipoproteine in die Zellen der Tertiärfollikel und der Gelbkörper überführt, aber der Grund dafür war unklar, bis Biochemiker in Versuchen mit isolierten Granulosazellen herausfanden, dass bei der Progesteronproduktion ebenfalls freie Radikale entstehen, die an den beteiligten Enzymen und Proteinen Schäden verursachen. Durch Zugabe von beta-Karotin wurde dieser negative Effekt vermindert. Die Forscher schlossen aus ihren Ergebnissen, dass die hohen beta-Karotin-Gehalte im Ovar diese Schäden auch in vivo beschränken (Rodgers et al. 1995, Young et al. 1995). Das beta-Karotin wirkte hauptsächlich als Antioxidans und Radikalfänger bei der Stabilisierung der Membranen. Dies wird auch von anderen Untersuchern bestätigt (Rapoport et al. 1998). 2.13.8 Beta-Karotin als lokale Vitamin-A-Quelle des Ovars Schams et al. (1977) vermuteten, dass beta-Karotin eine lokale Vitamin-A-Quelle darstellt. Schweigert (1986) stellte fest, dass in nicht atretischen Follikeln mehr als doppelt soviel Vitamin A vorhanden war wie in atretischen Follikeln. Er nahm an, dass es durch lokale Umwandlung aus beta-Karotin entstanden war. In späteren Versuchen zeigte sich, dass die Aktiviät der Carotinase in den Tertiärfollikeln im Verlauf der Ausreifung um ein Vielfaches anstieg und auch im Gelbkörper hoch lag. 40 Das so entstandene Vitamin A förderte die Sekretion von 17-ß-Östradiol im Tertiärfollikel und die Sekretion von Progesteron im Gelbkörper. 2.14 Beta-Karotin und das Immunsystem Zahlreiche Versuche befassten sich mit der Bedeutung des beta-Karotins für das Immunsystem. In-vitro-Versuche zeigten, dass beta-Karotin die Proliferation von Blutlymphozyten in Abhängigkeit vom physiologischen Zustand des Tieres (in Laktation oder nicht in Laktation) beeinflusst (Tjoelker et al. 1988). Während des Trockenstehens hatte die beta-Karotin-Supplementierung einen stabilisierenden Effekt auf die Funktion der neutrophilen Granulozyten und auch auf die Funktion der Lymphozyten (Tjoelker et al. 1990). Zusätzlich wurde in vitro untersucht, ob betaKarotin die Funktion der Phagozyten in der peripartalen Phase beeinflusst. Es erhöhte die Fähigkeit der Blut- und Milchphagozyten zur Abtötung von Staphylococcus aureus während dieser Phase, aber es hatte keinen signifikanten Einfluss auf die Phagozytose selbst (Daniel et al. 1991a). Ebenso verbesserte betaKarotin die von Concavalin A (mitogenes Reagenz) induzierte Proliferation von Monozyten eine Woche a.p. (Daniel et al. 1991b). Dadurch soll es den Schutz des Euters vor Infektionen verbessern. Der genaue Wirkungsmechanismus ist unbekannt. Beta-Karotin verbesserte die Lymphozyten- und Phagozytenfunktion in der peripartalen Phase (Michal et al. 1994). In Feldversuchen konnte kein positiver Effekt von beta-Karotin auf die Eutergesundheit während des Trockenstehens oder der Frühlaktation festgestellt werden (Oldham et al. 1991). In der Humanmedizin ist das Interesse an den beta-Karotinen wiedererwacht, weil man Anhaltspunkte dafür gefunden hat, dass es bei der Bekämpfung von Krebs helfen kann (Ziegler 1989). An Versuchstieren wie Ratten und Kaninchen stellte man fest, dass es die Blastogenese der Lymphozyten fördert, die Entstehung von bestimmten Lymphozytengruppen verstärkt, die Zytotoxizität von Lymphozyten steigert und die Produktion von verschiedenen Zytokinen stimuliert. Zusätzlich verbessert es die Phagozytose und bakterienabtötende Fähigkeit von neutrophilen und peritonealen Makrophagen. Dies 41 gilt als gesicherte Funktion des beta-Karotins, die unabhängig von seiner Eigenschaft als Provitamin A ist. Beim Rind sieht man Möglichkeiten durch diese immunstimulierenden Fähigkeiten des beta-Karotins, die Gesundheit von Milchkühen zu verbessern (Chew 1993). 2.15 Weitere Einflussfaktoren auf die Fruchtbarkeit des Rindes Die Fruchtbarkeit von Rindern wird von vielen Faktoren beeinflusst, die zum Teil auch noch interagieren. Will man ein Fruchtbarkeitsproblem mit einem Mangel an beta-Karotin in Verbindung bringen bzw. es durch die Gabe von beta-Karotin beheben, sind zuerst alle anderen Ursachen ausschließen. 2.15.1 Energieversorgung Bei Milchkühen wird zwischen den Folgen einer Energieüberversorgung und eines Energiemangels unterschieden. Eine Energieüberversorgung kommt bei einer hochleistenden Milchkuh nur am Ende der Laktation bzw. in der Trockenstehphase vor. Die Energiebilanz sollte in dieser Zeit leicht positiv sein. Ein Energiedefizit ante partum führt bereits in diesem Stadium zu subklinischen Ketosen und Leberschäden. Eine verzögerte Uterusinvolution und Ovarfunktion post partum sind die Folge. Diese Fütterungsfehler in der Transitphase können nicht mehr durch eine bedarfsgerechte Energieversorgung unmittelbar post partum ausgeglichen werden (Markusfeld et al. 1997). Eine Verfettung der Tiere ist allerdings auch zu vermeiden, da sie sonst post partum häufiger an Milchfieber, Labmagenverlagerung, Stoffwechsel- und Fortpflanzungsstörungen leiden (Morrow et al. 1979, Swanson 1989). Größere Probleme bereitet die Deckung des Energiebedarfs zu Beginn der Laktation. Wegen der zum Teil erheblichen Milchleistung direkt nach der Abkalbung (> 40 l Milch/Tag) ist der Energiebedarf mit einer wiederkäuergerechten Ration nicht zu decken. Die Tiere haben eine negative Energiebilanz und mobilisieren Fett. Ob diese negative Energiebilanz die Fruchtbarkeit von Rindern negativ beeinflusst, ist umstritten. Den 42 zu diesem Thema Versuchsanordnungen erfolgten Untersuchungen zugrunde. Hinzu lagen kommen die sehr unterschiedliche teils unzureichenden Tierzahlen. Deshalb sind die Ergebnisse nur schwer zu vergleichen. Es spricht aber einiges dafür, dass die negative Energiebilanz Einfluss auf die Fruchtbarkeit nimmt. Die Trächtigkeitsraten lagen in hochleistenden Herden bei der zweiten Besamung zum Beispiel höher als bei der ersten. Zum Zeitpunkt der Zweitbesamung hatten die Kühe schon wieder eine positive Energiebilanz (Butler u. Smith 1989). Außerdem neigen Kühe mit starkem Gewichtverlust vermehrt zu Anöstrie (Perkins 1985). Man nimmt an, dass die aus dem energetischen Defizit resultierende Hypoglykämie die Synthese von gonadotropen Hormonen beeinträchtigt (Kolb u. Elze 1995). Die Folge sind Azyklie, stille Brunst, verzögerte Ovulation bis hin zur Zystenbildung (Staples et al. 1990). 2.15.2 Eiweißversorgung Eine mangelhafte Eiweißversorgung kommt in den Rationen von hochleistenden Milchviehherden praktisch nicht vor. Das Futter enthält meist große Mengen an Rohprotein, um hohe Milchleistungen zu erreichen. Außerdem sind eiweißreiche Rationen schmackhafter und die Trockenmasseaufnahme der Tiere steigt. Solche Rationen bereiten keine Probleme, solange sie genügend Energie enthalten. Bei ausreichender Energieversorgung wird der Ammoniak, der im Pansen aus dem Protein entsteht, zur Synthese mikrobieller Proteine verwendet (hepatoruminaler Kreislauf) (Martens et al. 2000). Liegt ein Energiemangel vor, wird der Ammoniak im Pansen resorbiert und in der Leber zu Harnstoff verstoffwechselt. Der Harnstoff wird dann über die Milch und die Nieren ausgeschieden. Dieser Vorgang führt auf die Dauer zu einer erheblichen Leberbelastung. Als Folge wurden stille Brunst, unregelmäßige Brunstintervalle und eitrige Genitalkatarrhe beobachtet (Lotthammer 1985b). Hohe Ammoniak- und Harnstoffkonzentrationen veränderten die uterinen Sekretionsvorgänge und den pH-Wert der Uterusflüssigkeit (Reksen u. Ropstad 2002). Die Produktion von Prostaglandin F2α steigerte sich ebenfalls. 43 Zusammenfassend kann gesagt werden, dass der Eiweißgehalt einer Ration die Fruchtbarkeit nicht beeinflusst, so lange der Rohproteinanteil niedriger als 18 % pro kg TM liegt. Bei höheren Eiweißgehalten können Fruchtbarkeitsprobleme entstehen, wenn die Ration nicht genügend Energie enthält und unzureichend strukturiert ist (Ferguson u. Chalupa 1989). 2.15.3 Mineralstoffe Früher wurde eine mangelhafte Phosphorversorgung mit Fruchtbarkeitsstörungen in Verbindung gebracht. Versuche unter definierten Bedingungen konnten aber keinen signifikanten Einfluß auf die Fertilität feststellen (Noller et al. 1977). Ein Phosphormangel führt demnach nur zu einer geringeren Futteraufnahme und kann durch den daraus resultierenden Energiemangel Fruchtbarkeitsstörungen hervorrufen. Bei der Verwendung von qualitativ hochwertigem Kraftfutter ist eine mangelhafte Phosphorversorgung sehr unwahrscheinlich. Ob das Kalzium- /Phosphor-Verhältnis eine Rolle in der Herdenfruchtbarkeit spielt ist noch nicht geklärt (Hurley u. Doane 1989). Im Gegensatz dazu gilt der negative Einfluss eines Kaliumüberschusses in Verbindung mit einem Natriummangel in der Ration als gesichert (Lotthammer 1985a). Als Folge treten Genitalkatarrhe I. und II. Grades, Azyklie und schlecht ausgebildete Gelbkörper auf. Zusätzlich wird angenommen, dass Kalium die Resorption von beta-Karotin stört, da bei einem Kaliumüberschuss erniedrigte beta-Karotin-Werte im Serum gefunden wurden. In neuerer Zeit wird ein Überschuss an Kalium auch mit dem vermehrten Auftreten von peripartalen Erkrankungen wie Milchfieber, Nachgeburtsverhaltung und Endometritis in Verbindung gebracht (Chandler 1997, Horst et al. 1997). 2.15.4 Spurenelemente In Gegenden mit Moorböden ist ein Selenmangel als Ursache von vermehrt auftretenden Nachgeburtsverhaltungen von praktischer Relevanz. Die daraus häufig 44 resultierenden Endometritiden verschlechtern die Fruchtbarkeit der Herden erheblich (Kappel et al. 1984). Man fand deutlich schlechtere Besamungsergebnisse bei Tieren, die Selengehalte unter 80 µg/l Serum während des Trockenstehens und bis 60 Tage post partum aufwiesen, als bei Tieren, die zu diesen Zeitpunkten Werte über 80 µg/l hatten (Boehnke et al. 1993). Ebenso wurde nachgewiesen, dass die Supplementierung von Selen bei Selenmangel die Fruchtbarkeit signifikant verbessert (Jukola et al. 1996a). Lotthammer (1999) geht dagegen bei Selenwerten von über 40 µg/l Serum in allen Stadien von einer ausreichenden Selenversorgung aus. Es liegen Hinweise vor, das eine mangelhafte Versorgung mit Mangan zu Anaphrodisie, mangelhafter Kontraktionsbereitschaft des Uterus und Genitalkatarrhen führt (Anke 1971). Kupfermangel soll gehäuft auftretenden embryonalen Frühtod mit daraus resultierender Subfertilität verursachen. Zuvor sind oft schon andere Symptome des Kupfermangels deutlich wie zum Beispiel die „Kupferbrille“ (grauweißer Ring um die Augen) (Seekles u. Claessens 1967). Die Rolle des Kupfers ist in der Literatur umstritten, zumal die Nutzung des Kupfers durch ein Überangebot von anderen Mineralien wie z. B. Molybdän, Eisen, Zink oder Kalzium stark negativ beeinflusst wird. Besonders ein hoher Molybdängehalt kombiniert mit einem niedrigen Kupfergehalt beeinflusst die Fertilität ungünstig (Whitaker 1999). Zusammenfassend ist zu sagen, dass ein Spurenelementmangel mit Ausnahme des Selens im Zusammenhang mit Fruchtbarkeitsstörungen keine praktische Bedeutung besitzt. Außerdem ist bei hochleistenden Herden davon auszugehen, dass die Tiere über Kraft- und Mineralfutter ausreichend mit Spurenelementen versorgt werden. 2.15.5 Vitamine Mit Ausnahme von beta-Karotin und Vitamin A (siehe oben) spielen Vitamine im Fruchtbarkeitsgeschehen des Rindes keine Rolle. Früher wurde dem Vitamin E eine Bedeutung im Fruchtbarkeitsgeschehen zugesprochen. In neueren Untersuchungen konnte aber immer nur ein positiver Einfluß auf die Inzidenz von Mastitiden 45 nachgewiesen werden. Positive Auswirkungen auf die Fruchtbarkeit waren nicht festzustellen (Allison u. Laven 2000, LeBlanc et al. 2002). 2.15.6 Werden Mykotoxine beim Rind verschimmelte Futtermittel eingesetzt, kann es zu Fruchtbarkeitsstörungen kommen. Bei den Toxinen handelte es sich meist um Ochratoxin, Zearalenon und Trichotecene (Höltershinken et al. 1996). Besonders Zearalenon führte zu Nymphomanie, Anöstrie, Ovarialzysten und Aborten (Weaver et al. 1986). 2.15.7 Oxidativer Stress Peripartal auftretender oxidativer Stress führte zu einer Häufung von Euterödemen, Nachgeburtsverhaltung und Mastitiden (Miller et al. 1993). Außerdem gab es negative Auswirkungen auf die Fruchtbarkeit (s.o.). Die Autoren betonten, dass die Kühe ausreichend mit Antioxidantien versorgt werden müssen. Genaue Angaben über den Bedarf einer Milchkuh liegen bis heute nicht vor. Es sollte aber darauf geachtet werden, dass der Peroxidgehalt im Futter nicht zu hoch ist, da beta-Karotin sonst nur als Antioxidans verwendet wird und für seine eigenständige Wirkung am Ovar nicht mehr zur Verfügung steht. Ähnlich wirkt sich ein Mangel an Vitamin E und Selen aus. 2.16 Kosten pro Trächtigkeit Um die Wirtschaftlichkeit der Fruchtbarkeitsleistung einer Herde zu beurteilen, wurden bisher verschiedene Faktoren in die Kostenberechnung miteinbezogen. Dazu zählten die Kosten für Besamungen, die Arbeitsleistung des Tierarztes für Behandlungen bei Sterilität, die Medikamentenkosten, die Kosten für verlängerte 46 Güstzeiten und die Remontierungskosten aufgrund von Fruchtbarkeitsstörungen oder Unfruchtbarkeit. Die Tierarzt- und Medikamentenkosten stellten nur einen geringen Anteil an den Gesamtkosten dar (Jakob u. Distl 1997, Tenhagen et al. 1998). Die Kosten der nicht realisierten Gewinne durch verlängerte Güstzeiten (geringere Milchleistung und verlängerte Zwischenkalbezeit) und die Remontierungskosten wegen Unfruchtbarkeit hatten dagegen einen hohen Anteil an den Gesamtkosten. Diese indirekten Verluste waren somit ein wesentlich größerer Kostenfaktor als die Tierarztkosten (Britt 1985, Dijkhuizen et al. 1984, Lotthammer 1992, Tenhagen et al. 1998). Aus ökonomischen Gesichtpunkten wurde eine Zwischenkalbezeit von 12 Monaten empfohlen (Fetrow u. Blanchard 1987). Daraus ergab sich eine optimale Güstzeit von 85 Tagen. Die Kosten pro zusätzlichem Güsttag variierten zwischen 1 und 5 € pro Tag. Diese Schwankungen waren abhängig von den jeweils aktuellen Schlacht-, Milch- und Kälberpreisen (Britt 1975, Dijkhuizen et al. 1984, Dohoo et al. 1984, Esselmont u. Peeler 1993, Fetrow u. Blanchard 1987, Stott u. DeLorenzo 1988, Tenhagen u. Heuwieser 1997). Tischer (1998) betonte, dass diese Kosten betriebsspezifisch stark variieren können. Tabelle 5 gibt einige Literaturangaben über die Kosten verlängerter Güstzeiten und die Remontierungskosten wieder. Daraus ergab sich auch der Schluß, dass die Gesamtkosten pro Trächtigkeit von vielen betriebsspezifischen Faktoren abhängen und so von Betrieb zu Betrieb stark schwanken. Tischer (1998) gab pro Trächtigkeit Kosten von 321,60 – 380,91 € an. Drillich (1999) kam auf 152,21 – 162,65 €. Bei Klindworth (2000) kostete eine Trächtigkeit zwischen 196,71 und 278,10 € und bei Surholt (2001) ergaben sich Beträge von 295,52 – 308,74 €. 47 Tabelle 5: Kosten für verlängerte Güstzeiten und remontierte Tiere Autor Kosten pro Güsttag Währung Britt (1975) Dohoo et. al. (1984) Dijkhuizen et al. (1985) Fetrow u. Blanchard (1987) Stott u. DeLorenzo (1988) Esselmont u. Peeler (1993) Tenhagen u. Heuwieser (1997) Tischer (1998) 0,44-2,48 1,90 0,62-1,7 2 2,14 3 0-6,50 8,32/10,47 US Dollar US Dollar Holl. Gulden US Dollar US Dollar Brit. Pfund DM DM Remontierungskosten 500 590 755 48 3 Material und Methoden Ziel dieser Arbeit war es, den Einfluß einer Supplementierung mit beta-Karotin in Form der Injektionslösung Carofertin auf die Fruchtbarkeitsleistung von Milchkühen zu untersuchen. Die Versuche setzten sich aus einer Verlaufs- und einer Langzeitstudie zusammen und fanden in der Zeit von Januar 2001 bis September 2002 statt. 3.1 Das Präparat: Carofertin Der Hersteller von Carofertin ist die Firma Alvetra GmbH in Neumünster. Bei Carofertin handelt es sich um eine 1%ige Beta-Karotin-Injektionslösung. 1 ml der Injektionslösung enthält 10 mg natürliches beta-Karotin als arzneilich wirksamen Bestandteil. Sonstige Konservierungsmittel, Bestandteile sind Ascorbylpalmitat und 10 mg Benzylalkohol als all-rac-alpha-Tocopherol als Antioxidantien, Poly-8-oxyethylen-x-12-hydroxystearat (Kosolvent), Isopropylmyristat und Wasser für Injektionszwecke als Lösungsmittel. Das Präparat ist für Rinder und Schweine zugelassen. Die Einhaltung einer Wartezeit ist nicht erforderlich. Die Anwendung während der Laktation und Trächtigkeit ist möglich. Der Nutzen des Präparats ergibt sich laut Hersteller aus der eigenständigen Wirkung des betaKarotins auf die Fruchtbarkeit, seiner Wirkung als Antioxidans (Membranschutz), seiner Wirkung als Provitamin-A und der Immunitätssteigerung. Anwendungsgebiete beim Rind sind laut Hersteller Prophylaxe und Unterstützung der Therapie von Fruchtbarkeitsstörungen, Verstärkung der Brunstsymptome durch Erhöhung des Östrogenspiegels, Beschleunigung der Uterusinvolution, Senkung des Risikos von Nachgeburtsverhaltung und Endometritis, Stabilisierung der Corpora lutea, Anhebung des Progesteronspiegels, Verbesserung der Nidation durch Stimulierung der Endometrium-Drüsen, Stabilisierung der Gravidität, Immunsteigerung der Neugeborenen sowie Verbesserung der Aufzucht, Unterstützung der Therapie und 49 Prophylaxe von Endometritiden und Substitution der Beta-Karotin-Versorgung während der Mangelperiode von November bis März oder ganzjährig bei Heu oder Silagefütterung. Vor der Anwendung von Carofertin soll allerdings ein Vitamin-AMangel ausgeschlossen werden, da das beta-Karotin sonst komplett in Vitamin A umgewandelt wird und seine eigenständige Wirkung nicht entfalten kann. Ebenso sind ein hoher Peroxidgehalt (oxidativer Stress) im Futter und ein Mangel an Vitamin E und Selen auszuschließen, da beta-Karotin sonst als Antioxidant verwendet wird. Andere negative Effekte auf die Fruchtbarkeit z. B. durch Mykotoxine im Futter oder einen Mangel an Spurenelementen (P, Mn, Zn, Mo, J) sollte man vor der Anwendung von Carofertin ebenfalls beseitigen. Laut Herstellerangabe liegt die Einzeldosis für eine Kuh oder Färse bei 20-25ml. Das Präparat ist zur tiefen intramuskulären oder zur subkutanen Injektion geeignet. Bei der Sterilitätstherapie werden 1-3 Injektionen im Abstand von 2 Wochen empfohlen. Im Puerperium soll die erste Injektion 1-2 Wochen und die zweite Injektion 6-8 Wochen post partum erfolgen. Zur Immunitätssteigerung bei Neugeborenen wird eine Injektion 1-2 Wochen ante partum empfohlen. Die Injektionsmenge wird nach Möglichkeit auf maximal 10 ml pro Injektionsstelle aufgeteilt. Der Hersteller weist darauf hin, dass es vereinzelt zu Schwellungen an der Injektionsstelle kommen kann, die gewöhnlich ohne Behandlung wieder abklingen. Solche Schwellungen wurden aber während des ganzen Versuches nur zweimal beobachtet. Das Präparat ist unangebrochen bei einer Temperatur unter 25 °C und mit Lichtschutz 2 Jahre haltbar. Nach Anbruch einer Flasche sollte das Medikament laut Hersteller innerhalb von 3 Tagen aufgebraucht werden und bei 2-8 °C gelagert werden. 3.2 Überprüfung der Vitamin-A-, Vitamin-E- und Selenversorgung Vor der Anwendung von Carofertin soll die Versorgung der zu behandelnden Kühe mit Vitamin A, Vitamin E und Selen überprüft werden, da ein Mangel die Fruchtbarkeit ebenfalls negativ beeinflusst. Die Vitamin-A-Konzentration im Serum sollte bei ausreichend versorgten Kühen über 0,3 mg/l liegen (Herdt u. Stowe 1991). 50 Bei Vitamin E sollte die Serumkonzentration über 3,0 mg/l sein. Für Selen gelten Werte über 40,0 µg/l im Serum als physiologisch (Lotthammer 1999, Scholz 1989). Zur Überprüfung der Versorgung wurden von 8 Färsen und 12 Kühen jeweils aus beiden Gruppen, verteilt auf alle vier Jahreszeiten, Serumproben ausgewählt. Das Serum stammte zu gleichen Teilen von den Entnahmezeitpunkten I (3-4 Wochen a.p.) und Entnahmezeitpunkt II (1-2 Wochen p.p.). Insgesamt wurden somit 80 Serumproben untersucht. 3.2.1 Bestimmung von Vitamin A und Vitamin E im Serum Die Bestimmung von Vitamin A und Vitamin E erfolgte im klinischen Labor der Klinik für Rinderkrankheiten der Tierärztlichen Hochschule Hannover. Beide Vitamine wurden in Anlehnung an die Methode von Rammel et al. (1983) untersucht. Nach der Verseifung der Probe und anschließender Extraktion der unverseifbaren Substanzen mit organischen Lösungsmitteln erfolgte die Separation und Bestimmung der Vitamine A und E durch HPLC und fluoreszenzphotometrische Detektion. Zur Durchführung dieser Analyse wurden jeweils 1 ml Serum + 1 ml Ascorbinsäure 10 %1 + 1 ml Methanol2 + 2 ml Ethanol3 + 1 ml KOH4 in mit Plastikstopfen verschlossenen 10-ml-Reagenzglasröhrchen gut durchgemischt und danach bei 60 °C verseift. Nach 10-minütiger Abkühlung im Eisbad wurden zu den Proben 5 ml Hexan5 hinzugefügt. Die Hexanphase wurde nach Mischen und Zentrifugieren (1000 g, 5 min) in kleine Kolben abpipettiert. Diese Hexanextraktion wurde zweimal wiederholt. Danach erfolgte die Eindampfung der vereinigten Hexanphasen im Rotationsverdampfer1 bis nur noch ein trockener Rückstand zurückblieb (Druck 150 mbar, Temperatur 35°C). Daraufhin wurde der Extrakt in 1 ml Methanol gelöst. Danach folgte die 1 Merck, Art.Nr. 500074, Darmstadt 2 Fisons, Art.Nr. XM 4056, Loughorough, Großbritannien 3 Chromatographie Handel Müller GmbH, Art.Nr. E/0665/17, Fridolfingen 4 Merck, Art.Nr. 5033, Darmstadt 5 Fisons, Art. Nr. XM/0406/17, Loughorough, Großbritannien 51 Probenanalyse über Autoinjektor mittels HPLC-Apperatur. Die genauen Daten der HPLC-Apperatur sind Tabelle 6 zu entnehmen. Tabelle 6: Daten der HPLC-Apperatur Messtechnik HPLC-Anlage Säule Vorsäule Autoinjektor HPLC-Pumpe 2150 250 x 4,6 Nucleosil C18 10 x 4,6 Nucleosil C18 SIL 9 A Fa. Pharmacia LKB, Freiburg Fa. Grom, Herrenburg Fa. Grom, Herrenburg Fa. Shimadzu Europa GmbH, Duisburg Fisons, Art.Nr. XM 4056, Loughorough, Großbritannien Fließmittel Aqua bidest. Methanol p.a. (v/v) Fließgeschwindigkeit 2 ml/min Detektion Spectrofluorometer RF-551 Exzitation Integrator 296 nm C-R5A Fa. Shimadzu Europa GmbH, Duisburg Fa. Shimadzu Europa GmbH, Duisburg Vor jeder Meßreihe wurde der Vitamin-A-/Vitamin-E-Gehalt einer Standardlösung dreimal bestimmt. Anhand der gemittelten Ergebnisse wurden die Chromatogramme der Proben ausgewertet. Durch sechsmaligen Ansatz einer Serumprobe wurde ein Intraassay-Variationskoeffizient von 4,22 % ermittelt. Der Interassay- Variationskoeffizient lag bei Vitamin E bei 6,37 % und bei Vitamin A bei 8,40 %. 1 Fa. Heidolph-Elektro GmbH & CoKG, Kelheim 52 3.2.2 Bestimmung von Selen im Serum Die Selenkonzentration im Serum wurde mit Hilfe der Graphitrohr-Atom-AbsorptionsSpektrometrie (Unicam solaar 6961) bestimmt. Diese Methode wurde im Institut für Tierernährung der Tierärztlichen Hochschule Hannover entwickelt und ist als die Hausmethode dieses Instituts anzusehen. Es erfolgte zuerst eine Verdünnung des Serums mit destilliertem Wasser zu gleichen Teilen. Anschließend wurde der zehnte Teil des Tensids Triton-X2 zugegeben, um die Viskosität des Gemisches zu senken. Die Messung fand in einer elektrisch beheizten Graphitrohr-Einheit statt. Um ab 600 °C flüchtiges Selen zu stabilisieren, kam Nickelnitratlösung3 (2g/l Aqua dest.) als Modifier hinzu. Die Atomisierung des Selens in der Messphase erfolgt bei 2600 °C. Die Messung in Form einer Standardaddition mit zwei Selenstandards von 25 µg und 50 µg wurde mit einer Deuterium-Untergrundkorrektur durchgeführt, da Selen sonst nicht nachweisbar ist. Der Intraassay-Variationskoeffizient betrug 5,10 % und der Interassay-Variationskoeffizient lag bei 4,80 %. 3.3 Verlaufsstudie 3.3.1 Versuchsziel Es sollte ermittelt werden, ob die Injektion von 20 ml Carofertin den Serumspiegel von beta-Karotin bei trockenstehenden Milchkühen bzw. Färsen erhöht und wie lange diese Erhöhung anhält. 1 Fa. Unicam, Offenbach 2 Fa. Fluka, Deisenhofen 3 Fa. Fluka, Deisenhofen 53 3.3.2 Haltung und Fütterung der Versuchstiere Zu dieser Studie gehörten 6 Färsen und 14 Kühe der Rasse Deutsche Schwarzbunte mit Einkreuzung von Holstein Friesian (DSBxHF). Die Tiere wurden auf dem Lehrund Forschungsgut Ruthe der Tierärztlichen Hochschule Hannover gehalten. Die Tiere waren hochtragend (3 bis 4 Wochen vor dem errechneten Kalbetermin) und die Kühe standen trocken. Sie wurden in einem Laufstall mit Spaltenboden gehalten. Den Tieren standen Hochboxen zur Verfügung, die zum größten Teil mit Gummimatten ausgelegt waren. Der Rest der Hochboxen war mit Wasserbetten ausstattet. Das Futter bestand aus einer TMR (Total Mixed Ration) aus 40 % Maissilage, 10 % Gerstenstroh und 50 % Grassilage und wurde zweimal am Tag vorgelegt. 3.3.3 Versuchsaufbau Die 20 Versuchstiere wurden nach Kühen und Färsen getrennt durch ein Losverfahren paarweise in Kontroll- und Versuchstiere eingeteilt. Allen Versuchstieren wurde am ersten Versuchstag 20 ml Blut aus der gestauten V. jugularis externa entnommen und auf zwei 10 ml-Serumröhrchen mit Gerinnungshilfe verteilt1. Die 3 Färsen und 7 Kühe der Gruppe der Versuchstiere bekamen am ersten Versuchstag je 20 ml Carofertin, Alvetra GmbH, in die Ankonäenmuskulatur injiziert2. Das Injektionsvolumen wurde auf zwei Stellen verteilt. Die 3 Färsen und 7 Kühe der Kontrollgruppe bekamen stattdessen 20 ml 0,9%ige Kochsalzlösung3 in gleicher Weise injiziert. Allen Tieren des Versuchs wurde dann 14 Tage lang einmal täglich Blut in gleicher Weise wie oben beschrieben abgenommen. Danach erfolgten weitere vier Blutentnahmen in zweitägigen Abständen, so dass die letzte Blutentnahme am 22. Versuchstag erfolgte. Wenn die Tiere vorzeitig abkalbten, 1 Sarstedt, Nümbrecht 2 Kanüle: TSK Sterijekt 1,40x40, Ehrhardt-Söhne, Geislingen; sterile Einmalspritze: Injekt Luer Solo 20 ml, B. Braun Melsungen AG, Melsungen 3 Isotonische Natrium-Chlorid-Lösung, Delta-Pharma, Pfullingen 54 wurde auf weitere Blutentnahmen verzichtet. Das entnommene Blut wurde bei 4°C ein bis sechs Stunden aufbewahrt und nach dem vollständigen Gerinnen in einer Megafuge 1.0 R1 bei 3000 Umdrehungen/Minute (entspricht einer Beschleunigung von 1900 g) und 4°C für 20 Minuten zentrifugiert. Hämolytisches Serum wurde verworfen. Einwandfreies Serum wurde durch Dekantieren gewonnen, in 8 ml Röhrchen2 abgefüllt und bei –20 °C eingefroren. Alle zwei Monate wurden die eingefrorenen Proben in mit Trockeneis gekühlten Behältern per Kurier zur Untersuchung auf beta-Karotin zum VetMedLab Ludwigsburg geschickt. 3.3.4 Untersuchung des Serums auf beta-Karotin Beta-Karotin wurde nach einer Proteinfällung mit 2-Propanol direkt aus dem Serum mittels High Pressure Liquid Chromatography (HPLC) basierend auf der Methode nach Miller et al. (1984) bestimmt. Stark hämolytisches Serum war zur Untersuchung nicht geeignet (Miller et al. 1984). Zur Beta-Karotin-Bestimmung wurden je 200 µl 2Propanolol in einem Eppendorf-Gefäß vorgelegt und jeweils 100 µl Serum dazupipettiert. Das Gefäß wurde kurz geschüttelt und dann zentrifugiert. Der Überstand wurde in ein Probengefäß überführt und dann zur HPLC-Analyse verwendet. Es wurde ein HPLC System LC 1 mit UV-Vis-Detektor3 verwendet. Als HPLC-Säule wurde eine Purospher Star RP18-encapped Säule (3 µm (30x4mm))4 benutzt. Das Laufmittel war Methanol. Das Injektionsvolumen betrug 20 µl mit einer Flussrate von 1 ml/min bei einer Detektionswellenlänge von 452 nm. Die Substanzidentifizierung erfolgte über den Vergleich der Retentionszeiten des Peaks auf dem Chromatogramm der Serumprobe mit denjenigen des Chromatogramms der Standardlösungen (500, 1000, 2500 µg/l beta-Karotin). Die Quantifizierung erfolgte über die erstellte Eichkurve mit Hilfe der Externalstandard-Methode. Der Intraassay- 1 Heraeus Instruments GmbH, Hanau 2 8 ml, 84x14,5 mm, Sarstedt, Nümbrecht 3 SPD-6AV, Shimadzu, Duisburg 4 Fa. Varian 2010, Darmstadt 55 Variationskoeffizient betrug 5,00 %. Der beta-Karotin-Gehalt des Serums wurde vom Labor in der Einheit µg/l angegeben. Der vom Labor angegebene Normalbereich liegt bei über 700 µg/l. 3.4 Langzeitstudie 3.4.1 Versuchsziele Es sollte ermittelt werden, welchen Einfluß die Gabe von Carofertin auf verschiedene Fruchtbarkeitskennzahlen (Rastzeit, Güstzeit, Erstbesamungserfolg, Trächtigkeitsindex, Gesamtträchtigkeitsrate, erwartete Zwischenkalbezeit, Konzeptionsrate) hat. Außerdem wurde untersucht, ob die Behandlung einen Einfluß auf das Auftreten von häufig vorkommenden Erkrankungen wie Milchfieber, Nachgeburtsverhaltung, Endometritis, Stillbrünstigkeit und Ovarialzysten hatte. Zusätzlich wurden die Abgangsgründe und die Abgangsraten ausgewertet. Zum Schluß sollten die Kosten pro Trächtigkeit errechnet werden. 3.4.2 Haltung der Versuchstiere Die 201 Versuchtiere (hauptsächlich Tiere der Rasse Deutsche Schwarzbunte mit Einkreuzung von Holstein Friesian (DSBxHF), einige Rotbunte und eine Jersey) gehörten zu einem Betrieb in Wasserleben (Sachsen-Anhalt). Die Herde umfasste ca. 220 laktierende Tiere plus Trockensteher und Jungvieh. Die durchschnittliche Herdenleistung lag bei 8500-9000 kg Milch mit 4,2% Fett und 3,41 % Eiweiß. Die laktierenden Tiere wurden zu Versuchsbeginn in einem Freßliegeboxenstall mit Stroheinstreu gehalten. Die laktierenden Tiere wurden ganzjährig im Stall gehalten. Die tragenden Rinder befanden sich am anderen Ende des Dorfes in einem Laufstall mit Tiefeinstreu. Von Mai bis Oktober hatten sie freien Zugang zur Weide. Ungefähr 8 Wochen vor dem errechneten Kalbetermin wurden die Tiere in ein separates 56 Stallabteil des Hauptgebäudes umgestallt. In diesem Stall wurden ebenso die trockenstehenden Kühe gehalten. Es handelte sich dabei um einen ehemaligen Anbindestall mit Fressgitter (Mittellangstand mit Kotstufe und Stoheinstreu), in dem die Tiere aber frei liefen. Zur Abkalbung standen 2 Abkalbeboxen zur Verfügung. Frischabgekalbte Tiere, kranke und schwermelkende Tiere wurden in einem Teil dieses Stalles in Anbindung auf Stoh gehalten. Festliegende Tiere oder Tiere mit Gliedmaßenerkrankungen wurden zum Teil auch in den Abkalbeboxen untergebracht. Ungefähr ein halbes Jahr nach Versuchsbeginn wurde der Stall mit den Fressliegeboxen wegen Mastitisproblemen aus Hygienegründen umgebaut. Die altmelkenden Tiere kamen für 3 Monate auf die Weide. Die frischlaktierenden Tiere wurden auf dem Nachbarbetrieb in einem Fressliegeboxenstall mit Stroheinstreu gehalten. Aus Platzgründen wurden die trockenstehenden Kühe zu den Rindern gebracht und erst 4 Wochen vor der Abkalbung in den ehemaligen Anbindestall umgestallt. Der alte Fressliegeboxenstall wurde entkernt und an der Längsseite über die ganze Länge des Stalles Außenfressplätze angebaut. Der Innenraum wurde in 3 Abteile aufgeteilt. Es wurden nun Hochboxen mit Gummimatten eingebaut. Der Stall wurde mit planbefestigten Laufflächen, einer Faltschieberentmistung und Trogtränken ausgestattet. Nach der Fertigstellung befanden sich die altmelkenden Kühe und Färsen im ersten Abteil, im zweiten Abteil die frischabgekalbten Färsen und niedrigleistenden Kühe und im dritten Abteil die frischabgekalbten Tiere mit hoher Leistung. Jedes Abteil besaß zwei Tore mit Zugang nach draußen. Die tragenden Rinder und die trockenstehenden Kühe wurden wie vor dem Umbau gehalten. 57 3.4.3 Fütterung der Versuchstiere 3.4.3.1 Vor dem Stallumbau Vom Versuchsbeginn bis zum Stallumbau bekamen die laktierenden Kühe eine Mischration in zwei Leistungsgruppen, die durch Kraftfutter über Transponder in Abhängigkeit von der Milchleistung aufgewertet wurde. Die Ration für die „Frischmelker“ war für eine Milchleistung von 30 kg ausgelegt; die Tiere bekamen allerdings schon ab einer Milchleistung von 27 kg 1 kg Milchleistungsfutter (Frischmelker: Rohprotein 20 %, Energiestufe 4; Altmelker: Rohprotein 18 %, Energiestufe 3) pro 2 kg Milch zugefüttert. Die Grundfutterration der altmelkenden Kühe war für eine Leistung von 22,5 kg Milch berechnet und wurde ebenfalls durch eine Kraftfuttergabe nach Leistung ergänzt. Die Tiere wurden ungefähr 8 Wochen vor der Abkalbung trockengestellt und erhielten dann eine Mischration, die für 8 kg Milch ausgelegt war. Zwei Wochen vor der Abkalbung begann die Vorbereitungphase, in der die Tiere die Ration der frischmelkenden Kühe ohne Mineralfutterzusatz erhielten. Nach der Abkalbung bekamen die Tiere weiterhin die Ration der Frischmelker und 2 kg Milchleistungsfutter. Nach ein paar Tagen wurde die Kraftfuttergabe, wie oben beschrieben, der Milchleistung angepasst. Etwa 3-4 Monate nach der Abkalbung wurden die Tiere in die Gruppe der Altmelker umgestallt und erhielten die Altmelkerration plus Milchleistungsfutter nach Leistung. Die genaue Zusammensetzung der Ration ist Tabelle 7 zu entnehmen. 58 Tabelle 7 Futterrationen vor dem Stallumbau Futtermittel (FM in kg) Maissilage Grassilage Gerstenstroh Heu Biertreber, frisch Pressschnitzel, siliert Kartoffeln Gerste/Weizen Trockenschnitzel 10-15 % Propylenglykol Sojaextraktionsschrot 44 Rapsschrot 35% Rohprotein Viehsalz Kohlens. Futterkalk Vilomin 11462 Viloprem Frischmelker Altmelker Trockensteher 20,0 9,0 0,3 15,0 13,0 11,6 4,4 1,1 2,2 5,5 4,0 4,5 1,0 0,5 0,2 1,8 1,5 0,08 0,07 0,1 5,5 5,0 5,0 0,5 0,9 0,6 0,06 0,02 0,15 0,08 Die Zusammensetzung der Milchleistungsfuttermittel ist in Tabelle 8 und 9 aufgeführt: Tabelle 8: Zusammensetzung der Milchleistungsfutter (MLF) Inhaltsstoffe (%) Rohprotein Rohfett Rohfaser Rohasche Calcium Phosphor Natrium MJ NEL/kg MLF Frischmelker MLF Altmelker 20,0 3,5 7,0 7,5 0,8 0,5 0,25 7,0 18,0 4,0 11,0 8,0 0,8 0,5 0,25 6,7 59 Tabelle 9: Zusatzstoffe im Milchleistungsfutter (MLF) Zusatzstoffe je kg Vitamin A (I.E.) Vitamin D3 (I.E.) Kupfer (mg) Selen (mg) MLF Frischmelker MLF Altmelker 10000 1000 10 0,5 10000 1000 10 0,5 Die Zusammensetzung der Mineralfuttermittel ist Tabelle 10 zu entnehmen: Tabelle 10: Zusammensetzung der Mineralfuttermittel Inhaltsstoffe Calcium (%) Phosphor (%) Natrium (%) Magnesium (%) Vitamin A (I.E.) Vitamin D3 (I.E.) Vitamin E (mg) Eisen (mg) Kupfer (mg) Mangan (mg) Zink (mg) Jod (mg) Selen (mg) Kobalt (mg) Vilomin 11462 Viloprem 11430 18,0 3,0 10,0 3,0 1000000 100000 2000 1000 800 4000 6000 50 50 30 2,0 7,0 10,0 38,0 600000 120000 1000 900 60 Eine Analyse der Mischrationen durch die LUFA Hameln ergab folgendes Ergebnis (Tab. 11). Tabelle 11: Ergebnis der Futtermittelanalyse durch die LUFA Hameln (FM=Frischmasse, TM=Trockenmasse) Parameter Frischmelker FM TM Altmelker FM TM Trockensteher FM TM pH-Wert Konservierungsqualität Trockenmasse (%) Rohasche (%) Rohprotein (%) Rohfaser Stärke (%) Gasbildung ml/200 mg OS Karotin mg/kg NEL (MJ/kg) ME (MJ/kg) Nutzbares Rohprotein (g/kg) Rum. N-Bilanz (g/kg) 4,4 sehr gut-gut 33,0 2,3 7,1 4,8 14,5 6,2 18,9 6,5 19,7 18,0 54,7 16,8 50,9 2,2 6,7 3,6 11,0 48,9 148,9 4,2 sehr gut-gut 30,5 2,0 6,4 2,8 9,2 5,3 17,3 7,1 23,2 17,3 56,7 16,3 56,6 2,1 6,7 3,4 11,0 42,1 137,9 4,7 sehr gut-gut 41,0 3,3 8,1 4,4 10,8 11,1 27,1 4,4 10,7 16,6 40,5 8,6 20,9 2,2 5,3 3,7 8,9 47,2 115,2 -0,2 -2,2 -0,5 -0,6 -7,3 -1,1 3.4.3.2 Nach dem Stallumbau Nach dem Stallumbau erhielten die Tiere eine TMR (Total Mixed Ration) unterteilt in drei Leistungsgruppen. Die Ration der hochleistenden Frischmelker (Abteil III) reichte aus für eine Milchleistung von vorgegebenen 37,8 kg bei 4,11 % Fett und 3,26 % Eiweiß. Die Ration der Altmelker (Abteil I) wurde berechnet für eine Leistung von 24,3 kg Milch bei 4,49 % Fett und 3,56 % Eiweiß. Die Ration der frischabgekalbten Färsen und der nicht so hochleistenden Kühe (Abteil II) bestand zu 50 % aus der Ration der Frischmelker und zu 50 % aus der Ration der Altmelker und reichte für eine Milchleistung von ca. 30 kg. Die Ration der Vorbereitungsgruppe bestand pro Kuh aus 13,8 kg der Frischmelker-TMR, 15,0 kg der Altmelker-TMR, 0,1 kg Propylenglykol und 20 g Dicalciumphosphat. Bei laktierenden Tieren würde sie für 13 61 kg Milch ausreichen. Die Trockensteher erhielten 9,1 kg Gras- und Maissilage, 2,9 kg Gerstenstroh und 0,1 kg Viloprem 11430. In Tabelle 12 ist die Zusammensetzung der Futterrationen getrennt nach Frischmelkern, Altmelkern und Trockenstehern aufgelistet. Tabelle 12: Zusammensetzung der Futterrationen nach dem Umbau, Angabe der Futtermittel in kg Frischmasse (FM) Futtermittel (FM kg) Frisch- Färsen/nicht Altmelker Trocken- Vorbereitungsmelker hochleistende gruppe steher Kühe Maissilage Grassilage Gerstenstroh Gemisch aus Kartoffeln, Biertreber, Presschnitzeln (1:1:1) Körnermais Gerste/Weizen MLF 18/3 Sojaextraktionsschrot 44 Rapsschrot 35% Rohprotein Trockenschnitzel 1015% Viehsalz Vilomin 11462 Viloprem 11430 Kohlensaurer Futterkalk Dicalciumphosphat 40 Propylenglykol 21,0 7,5 0,3 15,0 19,5 8,0 0,6 13,5 18,0 8,5 0,8 12,0 9,1 9,1 2,9 12,0 5,0 0,4 8,2 1,0 1,0 0,5 2,5 0,5 0,75 0,75 1,5 0,5 1,0 0,5 0,3 0,5 0,5 1,1 1,5 1,2 0,9 0,7 0,5 0,7 0,35 0,25 0,14 0,03 0,1 0,05 0,1 0,02 0,07 0,1 0,08 0,25 0,1 0,05 0,04 0,02 0,03 20,0 0,3 0,2 62 Unter Berücksichtigung der Grundfuttermittelanalysen für Gras- und Maissilage wurden die Kennzahlen der TMR, die nach dem Umbau an die verschiedenen Gruppen (Frischmelker, abgekalbte Färsen bzw. nicht so hochleistende Kühe, Altmelker, Trockensteher und Vorbereitungsgruppe) gefüttert wurde, errechnet (Tab. 13). Tabelle 13: Gehalte der TMR nach dem Umbau (TM=Trockenmasse) Parameter (pro kg TM) Trockenmasse (%) Frischmasse (kg) Rohprotein (g) Rohfaser (%) Stärke (g) Karotin (mg) NEL (MJ) Nutzbares Rohprotein (g) Ruminale Stickstoffbilanz (g) Frisch- Färsen/nicht Altmelker Trocken- Vorbereitungsmelker hochleistende steher gruppe Kühe 43,3 51,2 178,0 15,8 49,7 40,8 7,1 42,8 47,0 165,0 17,3 46,1 41,0 6,9 42,2 42,8 152,0 18,8 42,4 41,2 6,6 54,7 21,2 123,2 26,8 79,0 66,7 5,7 43,1 29,0 161,5 17,0 45,9 41,0 7,0 163,9 156,6 149,3 122,0 156,4 2,3 1,2 0 -1,5 1 3.4.4 Versuchsaufbau: beta-Karotin-Gehalt im Serum Es wurden 201 Milchkühe für den Zeitraum von 4 Wochen ante partum bis zum Ende der folgenden Laktation beobachtet. Die Einteilung in Versuchs- und Kontrollgruppe erfolgte ebenfalls paarweise und nach dem Losverfahren. Die 100 Tiere der Versuchsgruppe (71 Kühe und 29 Färsen) erhielten je drei Injektionen mit 20 ml Carofertin1 intramuskulär und die 101 Tiere der Kontrollgruppe (72 Kühe und 29 Färsen) erhielten je drei Injektionen von 20 ml 0,9 % NaCl-Lösung2 intramuskulär in 1 Alvetra GmbH, Neumünster 2 Delta-Pharma, Pfullingen 63 die Ankonäenmuskulatur auf zwei Stellen verteilt. Die erste Injektion erfolgte 3-4 Wochen vor dem errechneten Kalbetermin. Die zweite Injektion wurde 1-2 Wochen nach der Abkalbung und die dritte Injektion 6-8 Wochen nach der Abkalbung vorgenommen. Um den Gehalt des Serums an beta-Karotin zu bestimmen, wurde den Tieren unmittelbar vor der ersten und zweiten Injektion Blut aus der V. jugularis externa entnommen und wie bei der Verlaufsstudie zum VetMedLab Ludwigsburg zur beta-Karotin-Bestimmung geschickt. Eine dritte Blutentnahme fand kurz nach der Besamung statt. Bei bewusst güsten Tieren, die z.B. wegen Euterproblemen oder geringer Leistung nicht mehr besamt wurden, fand die dritte Blutentnahme ca. 90 Tage post partum statt. 3.4.5 Versuchsaufbau: Puerperalkontrolle Bei allen Tieren wurde eine Puerperalkontrolle 4-6 Wochen post partum durchgeführt. Die Puerperalkontrolle bestand aus einer vaginalen und rektalen Untersuchung, bei der die Befunde über Scheidenbild, Ovarien (Zyklus ja/nein, Ovarialzysten ja/nein) und Uterus (Endometritis ja/nein) dokumentiert wurden. 64 Bei der Dokumentation kam der Schlüssel nach Grunert (1990) zur Anwendung (Tab. 14). Tabelle 14: Befunde der rektalen Untersuchung (Grunert 1990) Gebärmuttergröße GI G II G III G IV GV G VI Uterus unter der Hand zu versammeln, Hörner etwa fingerstark Uterus unter der Hand zu versammeln, Hörner etwa zweifingerstark Uterus unter der Hand zu versammeln, Hörner etwa drei- bis vierfingerstark Uterus mit der Hand abzugrenzen, d.h. die große Kurvatur des etwa brotlaibgroßen Organs lässt sich abtasten Uterus fast mit der Hand abzugrenzen, d.h. die große Kurvatur lässt sich nicht mehr vollständig abgrenzen Uterus nicht mit der Hand abzugrenzen, d.h. die große Kurvatur befindet sich außerhalb der Reichweite der untersuchenden Hand Symmetrie S As Beide Hörner gleich groß Hörner unterschiedlich groß (z.B. As+ = rechtes Horn größer) Konsistenz und Kontraktilität KI K II K III Uterus schlaff, wenig kontraktil Uterus mäßig kontraktil Starke Kontraktionsbereitschaft 65 Bei der vaginalen Untersuchung kam folgender Schlüssel zur Anwendung (Tab. 15): Tabelle 15: Befunde der vaginalen Untersuchung (Grunert 1990) Form der Portio vaginalis cervicis Z R V S Zapfenförmig Rosettenförmig Breit verlaufen Schlaff-lappig überhängend Öffnungsgrad des Zevikalkanals 0 1 2 3 4 5 Vollständig geschlossen Strohhalmstark geöffnet Bleistiftstark geöffnet Fingerstark geöffnet Zweifingerstark geöffnet Dreifingerstark geöffnet Farbe der Schleimhaut A B C D E Blass Blaßrosa Hyperämisch Deutlich krankhafte Rötung Sehr starke, schmutzig verwaschene Rötung Feuchtigkeitsgrad der Schleimhaut I II III IV V Trocken, klebrig Wenig feucht Mäßig feucht Sehr feucht Flüssigkeitsansammlung in der Scheide (Schl:Schleim, Bl:Blut, Ei:Eiter) 66 3.4.6 Kalbeverlauf, Krankheiten und Abgänge Der Kalbeverlauf (ohne Zughilfe, leichte bzw. starke Zughilfe), die Anzahl der Kälber, Totgeburten und Geburtsverletzungen dokumentierten die Betreiber des Hofes ebenso wie auftretende Erkrankungen und die dazugehörigen Behandlungen. Die Behandlungen wurden vom Haustierarzt durchgeführt. Zu den erfassten Erkrankungen zählten Milchfieber, Nachgeburtsverhaltung, Klauenerkrankungen, Endometritis und Ovarialzysten. Bei Abgängen notierten die Betreiber das Abgangsdatum und die Abgangsgründe. Als Abgang wegen mangelnder Fruchtbarkeit wurden Tiere gewertet, die besamt wurden und nach 200 Tagen post partum noch nicht tragend waren oder krankhafte Veränderungen am Genitale hatten wie z.B. Uterusverwachsungen oder therapieresistente Ovarialzysten. Tiere, die nach 3 Besamungen nicht tragend waren und vom Betriebsleiter aus diesem Grund von der Zucht ausgeschlossen wurden, gehörten ebenfalls zu dieser Gruppe. Weitere Abgangsursachen waren geringe Leistung, Euterprobleme, Klauen- und Gliedmaßenerkrankungen, Verenden nach Krankheit bzw. Euthanasie wegen Krankheit und sonstige Abgangsgründe (Paratuberkulose-Sanierung, Aggressivität, Spaltenlieger). 3.4.7 Fruchtbarkeitskennzahlen Zur Berechnung der Fruchtbarkeitskennzahlen wurden die Daten von Brunsten, Besamungen und Trächtigkeitsuntersuchungen dokumentiert. Die Brunstbeobachtung, die Besamungen und die Trächtigkeitsuntersuchungen (im Allgemeinen 5 Wochen nach der Besamung) wurden von einem der Betreiber durchgeführt. Die Brunstbeobachtung erfolgte zweimal am Tag und die freiwillige Wartezeit betrug ca. 60 Tage. Wenn eine Kuh bewusst güst blieb, das heißt, wenn sie aus betrieblichen Gründen oder wegen einer Krankheit nicht mehr besamt wurde, dokumentierte man dies mit Angabe des Grundes. Der Versuchszeitraum endete mit 67 der Trockenstellung der Kühe. Zur Beurteilung der Fertilität wurden folgende Kennzahlen herangezogen (Tab. 16): Tabelle 16: Fruchtbarkeitskennzahlen Parameter Formel Zielwert Gesamtträchtigkeitsrate (Berchtold 1982) Anz. tragende Tiere x 100 / Anzahl bes. Tiere Erstbesamungserfolg (De Kruif et al. 1998) Anz. trag. Tiere nach 1. KB x 100 / Anzahl Erstbesamungen Konzeptionsrate (Folman et al. 1990) Anz. tragende Tiere x 100 / Anzahl Besamungen insgesamt Trächtigkeitsindex (De Kruif et al. 1998) Anz. Besamungen bei tragenden Tieren < 1,6 200-Tage-Nichtträchtigkeitsrate (Morton u. McGowan 2002) Abgekalbte, nicht trächtige Tiere minus (bewusst güste Tiere + Tiere, die vor 200 Tagen p.p. abgegangen sind) ≤7% % der Tiere mit einer Güstzeit unter 115 Tagen (De Kruif et al. 1998) Anteil tragender Tiere mit einer Güstzeit unter 115 Tagen > 75 % Rastzeit (De Kruif et al. 1998) Tage von der Abkalbung bis zur ersten KB ∅ 80 - 85 Tage Güstzeit (De Kruif et al. 1998) Tage von der Abkalbung bis zum ersten Trächtigkeitstag ∅ 105 Tage Verzögerungszeit (De Kruif et al. 1998) Erwartete Zwischenkalbezeit Güstzeit minus Rastzeit ∅ 18 Tage Mittlere Güstzeit plus Tragzeit ∅ 385 Tage Abgänge wegen mangelnder Fruchtbarkeit % der Kühe mit Nachgeburtsverhaltung % der Kühe mit eitrigem Ausfluß post partum % der Kühe mit Ovarialzysten % der Kühe, die 60 Tage nach dem Abkalben noch nicht in Brunst gesehen wurden % abortierender Kühe % <7% % < 15 % % < 15 % % < 15 % % < 15 % Verwerfen zwischen 45.265. Tag <8% > 55 % 68 3.5 Progesteronbestimmung Von jeweils 10 Tieren pro Gruppe (jeweils 5 Kühe und 5 Färsen) wurden ab dem 40. Tag p.p. zweimal wöchentlich Blutproben aus der V. coccygea media entnommen. Es wurden dabei etwa 10 ml Blut in einem mit Lithium-Heparin beschichteten Zentrifugen-Röhrchen1 aufgefangen. Die Proben wurden ebenfalls bei 3000 U/min für 20 Minuten bei 4 °C zentrifugiert. Das Plasma wurde gewonnen und bis zur Untersuchung auf den Progesteronbestimmung Progesterongehalt bei erfolgte Radioimmunoassay mittels –20 °C eingefroren. (RIA) Die im endokrinologischen Labor der Klinik für Geburtshilfe und Gynäkologie des Rindes der Tierärztlichen Hochschule Hannover an frisch aufgetauten Proben mit geringen Modifikationen nach Hamburger (1974). Bei dieser Methode findet eine modifizierte Antigen-Antikörper-Reaktion nach dem Prinzip der Verdrängungsreaktion statt. Das in der Probe enthaltene Progesteron bindet sich an die spezifischen Antikörper und verdrängt so ein markiertes Substrat, dessen Menge dann quantitativ bestimmt werden kann. Die Ergebnisse werden in nmol/l Plasma angegeben. In der Literatur wird häufig noch die Einheit ng/ml verwendet. 1 ng/ml entspricht 3,18 nmol/l. Die Nachweisgrenze lag bei 0,318 nmol/ Blutplasma. Als Intra- und InterassayVariationskoeffizient wurden Werte von 6,6 % (n=10) und 8,0 % (n=16) errechnet. Ab einem Wert von 3-4 nmol Progesteron/l Plasma kann man davon ausgehen, dass ein funktionstüchtiger Gelbkörper vorliegt. Die Beprobung wurde so lange fortgeführt, bis von jedem Tier eine komplette Gelbkörperphase aufgezeichnet war. Entwickelte ein Tier eine Ovarialzyste oder kam es zu einer Azyklie schied es aus dem Versuch aus und wurde durch ein neues Tier derselben Gruppe ersetzt. Nachdem 20 Gelbköperphasen (10 Karotingruppe, 10 Kontrollgruppe) komplett waren, wurde jeweils der Entnahmetag, an dem die Progesteronkonzentration im Serum 1 nmol/l überschritt mit Entnahmezeitpunkte Entnahmezeitpunkt wurden 1 fortlaufend benannt. Die durchnummeriert. darauffolgenden Als letzter Entnahmezeitpunkt galt der Tag, an dem die Progesteronkonzentration im Serum 1 Sarstedt, Nümbrecht 69 wieder 1 nmol/l unterschritt. Da die Probennahme zweimal wöchentlich erfolgte, lag im Schnitt ein Abstand von 3,5 Tagen zwischen jedem Entnahmezeitpunkt. Die Progesteronkonzentrationen pro Entnahmezeitpunkt wurden getrennt nach Karotinund Kontrollgruppe gemittelt und in Abbildung 3 graphisch dargestellt. 3.6 Kosten pro Trächtigkeit Um die Wirtschaftlichkeit der Carofertin-Behandlung zu ermitteln, ist es notwendig, die Gesamtkosten pro erzielter Trächtigkeit zu errechnen und die beta-KarotinGruppe mit der NaCl-Gruppe zu vergleichen. Dafür wurden die Gesamtkosten pro Gruppe durch die Anzahl der tragenden Tiere (=erzielte Trächtigkeiten plus wegen mangelnder Fruchtbarkeit remontierte Tiere) geteilt (Surholt 2001). Damit erhält man die durchschnittlichen Gesamtkosten pro Trächtigkeit für beide Gruppen. Die Kosten pro Trächtigkeit setzten sich aus folgenden Teilbeträgen zusammen: • Tierarzt und Medikamentenkosten (laut Gebührenordnung für Tierärzte (GOT) vom 28.07.1999 einfacher Satz bzw. Arzneimittelpreisverordnung (AmPreisV) vom 14.11.1980 zuletzt geändert durch das 9. Euro-Einführungsgesetz vom 10.11.2001) bei Nachgeburtsverhaltung, Stillbrünstigkeit, Ovarialzysten, Endometritis) • Durchschnittliche Spermakosten (durchschnittlicher Preis einer Spermaportion auf diesem Betrieb 2001) = 7,50 € • Kosten für Trächtigkeitsuntersuchung (laut GOT 1999 einfacher Satz) = 7,67 € • 2,50 € pro Güsttag >85 Tage p.p. (Durchschnittswert laut Literatur) • Kosten für wegen mangelnder Fruchtbarkeit remontierte Kühe = 425,20 €/Tier 70 Die Kosten durch remontierte Kühe berechnen sich wie folgt: Verkaufspreis je Färse (∅ 2001 VIT Verden) 1100,00 € - Schlachterlös je remontierte Kuh 350,00 € = Differenz 750,00 € 750 € / 2,2 Laktationen1 341,00 € + Kosten durch geringere Milchleistung der Färsen 1052 kg x 0,08 Grenzkosten2/kg = Remontierungskosten pro Kuh 84,20 € 425,20 € 1 Durchschnittl. Nutzungsdauer: Durchschnittsalter (4,4 Jahre)-Erstkalbealter (2,2 Jahre) 2 Grenzkosten: Auszahlungspreis Milch: 0,37 €/kg -Variable Kosten (TA, Futter, Tiereinsatz) 0,22 €/kg -Lebendige Arbeit 0,07 €/kg = 0,08 €/kg => betriebsspezifische Werte laut Betriebskostenanalyse des Versuchbetriebes für das Jahr 2001 71 3.7 Auswertung Mit Hilfe des Tabellenkalkulationsprogramms EXCEL 20001 wurden die Daten erfasst und dann mit dem Statistikprogramm SAS2 basierend auf Standardmethoden (Steel u. Torrie 1980) ausgewertet. Alle Daten wurden auf Normalverteilung getestet. Für die Auswertung der beta-Karotin-Gehalte und die Fruchtbarkeitsparameter kamen nichtparametrische Tests zur Anwendung. Die Ergebnisse wurden jeweils als Mittelwert mit Standardabweichung (±SD) angegeben. Zum Vergleich der Mittelwerte wurde der Wilcoxon`s Rangtest herangezogen. Häufigkeiten wurden mit dem Χ2-Test verglichen. Beziehungen zwischen zwei Parametern wurden mit Hilfe einer Regressionsanalyse ermittelt. Zur Berechnung der Progesteronmenge während einer Lutealphase (Progesteronversuch) wurden die Progesteronkonzentrationen der einzelnen Entnahmezeitpunkte als Kurve aufgezeichnet und die Fläche unter der Kurve mittels Integralrechnung bestimmt (Fischer 1999). Bei allen Tests wurde eine Irrtumswahrscheinlichkeit von P < 0,05 vorgegeben. 1 Microsoft Corp., Redmond WA, USA 2 Version 8.02, Statistical Analysis Institute, Cary, NC, USA 72 4 Ergebnisse 4.1 Verlauf der beta-Karotin-Konzentration (Verlaufsstudie) Die Verlaufsstudie wurde mit 20 Tieren durchgeführt. Es handelte sich um 14 Kühe und 6 Färsen, die zu gleichen Teilen auf die Karotin- bzw. Kontrollgruppe verteilt wurden. Zunächst war auffällig, dass die beta-Karotin-Serumkonzentration von Tier zu Tier und von Tag zu Tag stark schwankten. Die Einzelwerte aller Tiere sind dem Anhang zu entnehmen (Tab. 64). Vergleicht man die Mittelwerte der beiden Gruppen zu den verschiedenen Zeitpunkten miteinander, so lagen die Werte bei der Karotingruppe meist höher als bei der Kontrollgruppe. Die Unterschiede waren aber nicht signifikant (P>0,05). In Abbildung 2 wurden die Mittelwerte der beta-KarotinKonzentrationen zu den einzelnen Entnahmezeitpunkten dargestellt. Bei einem nach Karotin- und Kontrollgruppe getrennten Vergleich der Ausgangskonzentrationen (Entnahmezeitpunkt I) mit allen weiteren Entnahmezeitpunkten, ergab sich in der Karotingruppe zum Entnahmezeitpunkt IV eine signifikant höhere beta-KarotinKonzentration im Serum als zum Zeitpunkt I (P<0,05). Zu allen anderen Zeitpunkten und innerhalb der Kontrollgruppe traten keine signifikanten Unterschiede (P>0,05) auf. Die Mittelwerte mit Standardabweichung der beta-Karotin-Konzentrationen zu den einzelnen Entnahmezeitpunkten sind Tabelle 17 zu entnehmen. 73 Tabelle 17: Mittlere beta-Karotinkonzentrationen (µg/l) zu den verschiedenen Entnahmezeitpunkten (1-14 = alle 24 h; 14-18 = alle 2061 a 10 1 10 1234,27 2549 10 2 822,95 684,84 786,42 964,02 529,07 2431 10 2670 10 3 2257 10 2668 b 10 4 2121 10 2442 10 5 2482 10 2562 10 6 1818 10 2385 10 7 1552,32 2304 10 1653,38 2969 10 8 887,63 1991 10 409,06 2146 10 9 631,61 2058 10 1159,64 2489 10 10 647,56 438,93 381,00 337,55 326,63 379,66 351,19 397,83 1968 10 2514 10 11 1744 10 1927 10 12 1639 10 1827 10 13 1663 10 1930 10 14 1619 10 1919 9 15 1600 10 1846 8 16 1543 10 1632 7 17 1460 9 1485 5 18 48 h) (MW = Mittelwert, SD = Standardabweichung, n = Anzahl der Versuchtiere, C = Karotingruppe, NaCl = Kontrollgruppe) 557,90 2138 Zeitpunkt MW 10 502,78 n SD 2033 937,26 499,43 536,89 463,97 412,72 603,67 528,06 432,57 n 732,21 931,05 602,96 745,95 880,20 775,14 NaCl MW C STD a vs b signifikant höherer Mittelwert der beta-Karotinkonzentration zum Zeitpunkt IV verglichen mit Zeitpunkt I (P<0,05) 73 74 Beta-Karotin-Serumkonzentration (µg/l) 5000 4500 4000 3500 b 3000 a 2500 2000 1500 1000 500 0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 Entnahm ezeitpunkte Karotin NaCl Abbildung 2: Verlauf der beta-Karotin-Serumkonzentrationen nach einmaliger Injektion von 20 ml Carofertin; Entnahmezeitpunkte siehe Legende Tabelle 17; für die Karotingruppe: a,b = Mittelwerte unterscheiden sich signifikant (P<0,05) 75 4.2 Vitamin-A-, Vitamin-E- und Selenversorgung (Langzeitstudie) Zur Überprüfung der Vitamin- und Spurenelementversorgung der Versuchstiere der Langzeitstudie wurden von 8 Färsen und 12 Kühen verteilt auf alle vier Jahreszeiten Serumproben ausgewählt. Die Ergebnisse sind der Tabelle 18 zu entnehmen. Tabelle 18: Vitamin-A-, Vitamin-E- und Selenkonzentrationen im Blutserum (t = Entnahmezeitpunkt, t1 = 3-4 Wochen a.p., t2 = 1-2 Wochen p.p., n = Anzahl der Versuchstiere, SD = Standardabweichung) Parameter Vitamin A (mg/l) Vitamin A (mg/l) Vitamin E (mg/l) Vitamin E (mg/l) Selen (µg/l) Selen (µg/l) t n Mittelwert ±SD Anteil der ausreichend versorgten Tiere (%) 1 40 0,54 0,20 100,00 > 0,3 2 40 0,52 0,25 95,00 > 0,3 1 40 5,18 2,42 82,50 > 3,0 2 40 3,11 1,09 55,00 > 3,0 1 2 40 40 49,40 60,60 13,00 16,70 70,00 90,00 > 40,0 > 40,0 Normalwert Die Mittelwerte der Vitamin-A-, Vitamin-E- und Selenkonzentrationen liegen für alle Parameter über den allgemein anerkannten Normalwerten. Betrachtet man die Werte der Einzeltiere, so sind zum Zeitpunkt 1 (3-4 Wochen a.p.) 82,5 % und zum Zeitpunkt 2 (1-2 Wochen p.p.) 55 % der Tiere ausreichend mit Vitamin E versorgt. Die Selenversorgung ist zum Zeitpunkt 1 bei 70 % der Tiere und zum Zeitpunkt 2 für 90 % der Tiere ausreichend. Die Vitamin A-Versorgung ist zum Zeitpunkt 1 bei allen Tieren gut. Zum Zeitpunkt 2 weisen 95 % der Tiere Blutserumkonzentrationen über 0,3 mg/l auf. 76 4.3 Übersicht über den Verbleib der Versuchstiere (Langzeitstudie) Es folgt eine Übersicht über den Verbleib der Versuchstiere während des Versuchszeitraums. Die Aufstellung erfolgt nach Karotin- und Kontrollgruppe getrennt. Karotingruppe: 100 Tiere → 5 Tiere vor BP II abgegangen (davon 1 Tier bei der Geburt verendet) ↓ 95 Tiere → 2 Tiere vor Puerperalkontrolle abgegangen ↓ 93 Tiere → 4 Tiere vor BP III abgegangen ↓ 89 Tiere → 85 Tiere besamt (4 bewußt güst) → 60 tragend geworden ↓ 33 Tiere während der Laktation abgegangen (5 davon tragend) ↓ 56 Tiere waren zum Versuchende noch im Bestand Kontrollgruppe: 101 Tiere → 7 Tiere vor BP II abgegangen ↓ 94 Tiere → 5 Tiere vor Puerperalkontrolle abgegangen ↓ 89 Tiere → 4 Tiere vor BP III abgegangen ↓ 85 Tiere → 85 Tiere besamt → 67 tragend geworden ↓ 23 Tiere während der Laktation abgegangen (5 davon tragend) ↓ 62 Tiere waren zum Versuchende noch im Bestand 77 4.4 Beta-Karotin-Gehalte im Serum (Langzeitstudie) Der beta-Karotingehalt im Serum wurde bei 201 Milchkühen an 3 Zeitpunkten bestimmt. Die Karotingruppe bestand aus 100 Tieren (71 Kühe und 29 Färsen) und die Kontrollgruppe (NaCl) bestand aus 101 Tieren (72 Kühe und 29 Färsen). Im Folgenden erfolgt die Auswertung der beta-Karotin-Serumkonzentration zuerst für alle Versuchstiere zusammen und dann für Kühe und Färsen getrennt. 4.4.1 Alle Tiere Es ergaben sich bei der Auswertung der gesamten Karotingruppe gegen die gesamte Kontrollgruppe zu den drei Entnahmezeitpunkten keine signifikanten Unterschiede (P>0,05) hinsichtlich der beta-Karotin-Konzentration im Blutserum. Vergleicht man die beta-Karotin-Serumkonzentrationen in der gesamten Karotingruppe zwischen den Zeitpunkten, d.h. Zeitpunkt 1 gegen Zeitpunkt 2, Zeitpunkt 1 gegen Zeitpunkt 3 und Zeitpunkt 2 gegen Zeitpunkt 3, so ergeben sich zwischen allen Zeitpunkten signifikante Unterschiede (P<0,05). Die beta-KarotinKonzentration ist zum Zeitpunkt 1 höher als zum Zeitpunkt 2 bzw. 3. Ebenso ist die beta-Karotin-Konzentration zum Zeitpunkt 2 höher als zum Zeitpunkt 3. Bei der Kontrollgruppe sind die beta-Karotin-Konzentrationen des Zeitpunkts 1 signifikant höher (P<0,05) als die Werte zum Zeitpunkt 2 bzw.3. Die Zeitpunkte 2 und 3 unterscheiden sich nicht signifikant (P>0,05). Die Daten zu dieser Auswertung sind in Tabelle 19 dargestellt. 78 Tabelle 19: Mittlere beta-Karotin-Konzentration aller Tiere (n = Anzahl der Versuchstiere, Zeitpunkt 1 = 3-4 Wochen a.p., Zeitpunkt 2 = 1-2 Wochen p.p., Zeitpunkt 3 = zur Zeit der Besamung, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe C Zeitpunkt n Mittelwert ± SD 1 2 3 100 95 89 1948,51 a 1123,47 b 1013,90 c 864,87 386,44 385,80 1 101 1943,40 a 696,58 NaCl 2 94 1063,30 b 392,53 3 85 1024,15 b 417,63 Innerhalb jeder Gruppe: a,b,c = Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant (P<0,05) 4.4.2 Kühe Es ergaben sich bei der Auswertung der Kühe der Karotingruppe gegen die Kühe der Kontrollgruppe zu den drei Entnahmezeitpunkten keine signifikanten Unterschiede (P>0,05) hinsichtlich der beta-Karotin-Konzentration im Blutserum. Vergleicht man die beta-Karotin-Serumkonzentrationen in der Karotingruppe zwischen den Zeitpunkten, d.h. Zeitpunkt 1 gegen Zeitpunkt 2, Zeitpunkt 1 gegen Zeitpunkt 3 und Zeitpunkt 2 gegen Zeitpunkt 3, so war die beta-Karotin-Konzentration zum Zeitpunkt 1 höher als zum Zeitpunkt 2 und 3. Zwischen Zeitpunkt 2 und Zeitpunkt 3 war die Differenz nicht signifikant (P<0,05). Bei der Kontrollgruppe waren die beta-KarotinKonzentrationen des Zeitpunkts 1 signifikant höher (P<0,05) als die Werte zum Zeitpunkt 2 bzw.3. Die Zeitpunkte 2 und 3 unterschieden sich ebenfalls nicht signifikant (P>0,05). Die Daten zu dieser Auswertung sind in Tabelle 20 dargestellt. 79 Tabelle 20: Mittlere beta-Karotinkonzentration der Kühe (n = Anzahl der Versuchstiere, Zeitpunkt 1 = 3-4 Wochen a.p., Zeitpunkt 2 = 1-2 Wochen p.p., Zeitpunkt 3 = zur Zeit der Besamung, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe Zeitpunkt n Mittelwert ± SD C 1 2 3 71 67 61 2143,62 a 1111,37 b 1073,30 b 901,74 314,77 411,51 NaCl 1 2 3 72 68 60 1974,21 a 1033,44 b 986,93 b 720,53 353,49 392,61 a, b = Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 4.4.3 Färsen Es ergab sich bei der Auswertung der Färsen der Karotingruppe gegen die Färsen der Kontrollgruppe zum Zeitpunkt 1 ein signifikanter Unterschied hinsichtlich der beta-Karotin-Konzentration im Blutserum (P<0,05). Die Färsen der Karotingruppe wiesen im Mittel deutlich niedrigere beta-Karotin-Serumkonzentrationen als die Färsen der Kontrollgruppe auf. Vergleicht man die beta-Karotin- Serumkonzentrationen in der Karotingruppe zwischen den Zeitpunkten, d.h. Zeitpunkt 1 gegen Zeitpunkt 2, Zeitpunkt 1 gegen Zeitpunkt 3 und Zeitpunkt 2 gegen Zeitpunkt 3, so ergeben sich zwischen allen Zeitpunkten signifikante Unterschiede (P<0,05). Die beta-Karotin-Konzentration ist zum Zeitpunkt 1 höher als zum Zeitpunkt 2 bzw. 3. Ebenso ist die beta-Karotin-Konzentration zum Zeitpunkt 2 höher als zum Zeitpunkt 3. Bei der Kontrollgruppe sind die beta-Karotin-Konzentrationen des Zeitpunkts 1 signifikant höher (P<0,05) als die Werte zum Zeitpunkt 2 bzw.3. Die Zeitpunkte 2 und 3 unterscheiden sich nicht signifikant (P>0,05). Die Daten zu dieser Auswertung sind in Tabelle 21 dargestellt. 80 Tabelle 21: Mittlere beta-Karotinkonzentration der Färsen (n = Anzahl der Versuchstiere, Zeitpunkt 1 = 3-4 Wochen a.p., Zeitpunkt 2 = 1-2 Wochen p.p., Zeitpunkt 3 = zur Zeit der Besamung, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe C Zeitpunkt n Mittelwert 1 2 3 29 28 28 1470,83 a, 1 1152,43 b 884,50 c ± SD 528,24 525,82 288,92 1 29 1866,90 a, 2 638,74 NaCl 2 26 1141,38 b 479,09 3 25 1113,48 b 468,78 Innerhalb jeder Gruppe: a,b,c = Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant (P<0,05). Zwischen den Gruppen: 1,2 = Mittelwerte mit unterschiedlichen Zahlenindices unterscheiden sich signifikant (P<0,05) 4.4.4 Kühe mit einem beta-Karotin-Wert unter 2000 µg/l zum Zeitpunkt I. Um einen eventuellen Effekt einer beta-Karotin-Injektion auf die beta- Karotinkonzentration im Serum bei Vorliegen einer beta-Karotin-Unterversorung bzw. einem durch das individuelle Resorptionsvermögen niedrigen beta-Karotin-Wert zu untersuchen, wurden die Versuchstiere in Abhängigkeit vom beta-Karotinwert in Gruppen eingeteilt. Bei der Gruppe der Kühe, die zum Zeitpunkt 1 einen betaKarotin-Wert unter 2000 µg/l beta-Karotin im Serum hatte, lag ein signifikanter Gruppenunterschied zur Kontrollgruppe zum Zeitpunkt 3 vor (P<0,05). Die betaKarotin-Konzentration im Blutserum der beta-Karotin-Tiere war, wie in Tabelle 22 zu sehen, zum Zeitpunkt der Besamung höher als bei der Kontrollgruppe. Vergleicht man die beta-Karotin-Serumkonzentrationen in der gesamten Karotingruppe zwischen den Zeitpunkten, so war die beta-Karotin-Konzentration zum Zeitpunkt 1 höher als zum Zeitpunkt 2 (P<0,05). Zwischen Zeitpunkt 2 und Zeitpunkt 3 war die Differenz nicht signifikant (P>0,05). Bei der Kontrollgruppe waren die beta-KarotinKonzentrationen des Zeitpunkts 1 signifikant höher (P<0,05) als die Werte zum Zeitpunkt 2 bzw. 3. Die Zeitpunkte 2 und 3 unterscheiden sich ebenfalls nicht 81 signifikant (P>0,05). Beim Vergleich der Färsen, die zum Zeitpunkt I eine betaKarotin-Konzentration unter 2000 µg/l hatten, traten keine Gruppenunterschiede auf. Ebenso zeigte die Auswertung der Tiere unter 1500 bzw. 1000 µg/l beta-KarotinSerumkonzentration (alle Tiere, Kühe bzw. Färsen) keine signifikanten Gruppenunterschiede zu den drei Zeitpunkten. Die Differenzen zwischen den Zeitpunkten waren jeweils so wie bei den gesamten Untergruppen (alle Tiere siehe Tab. 19, Kühe siehe Tab. 20, Färsen siehe Tab. 21). Tabelle 22: Mittlere beta-Karotinkonzentration der Kühe unter 2000 µg/l beta-Karotin zum Zeitpunkt 1 (n = Anzahl der Versuchstiere, Zeitpunkt 1 = 3-4 Wochen a.p., Zeitpunkt 2 = 1-2 Wochen p.p., Zeitpunkt 3 = zur Zeit der Besamung, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe C Zeitpunkt n Mittelwert 1 2 3 37 37 37 1513,49 a 994,86 b 1128,23 b, 1 ± SD 359,69 241,40 427,90 1 40 1470,98 a 333,55 NaCl 2 40 988,43 b 374,66 3 40 853,66 b, 2 340,24 Zwischen den Gruppen: 1,2 = Mittelwerte mit unterschiedlichen Zahlenindices unterscheiden sich signifikant (P<0,05). Innerhalb einer Gruppe: a,b = Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 82 4.5 Puerperalkontrolle (Langzeitstudie) 4.5.1 Beginn des Zyklus Der Anteil der Tiere, die bei der Puerperalkontrolle einen Gelbkörper hatten, wurde zwischen den Gruppen verglichen. Der Gelbkörper war dabei Beweis für einen laufenden Ovarialzyklus. Die Auswertung zeigte, dass in der Kontrollgruppe signifikant mehr Tiere einen Gelbkörper bzw. Zyklus aufwiesen als in der Karotingruppe (P<0,05). Die Ergebnisse sind Tabelle 23 zu entnehmen. Tabelle 23: Tiere, die 4-6 Wochen p.p. einen Zyklus hatten (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe Tiere n Tiere mit Zyklus in % (n) C NaCl alle 93 89 49,46 (46) a 64,04 (57) b C NaCl Kühe 65 64 55,38 (36) 65,63 (42) C 28 35,71 (10) Färsen NaCl 25 60,00 (15) a,b = Prozentzahlen mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 4.5.2 Ovarialzysten Die Inzidenz von Ovarialzysten wurde zwischen den Gruppen verglichen (alle Tiere, nur Kühe, nur Färsen). 12,90 % der Tiere der gesamten Karotingruppe und 14,61 % der Kontrolltiere hatten zum Zeitpunkt der Puerperalkontrolle Ovarialzysten. Bei der Feststellung der Ovarialzysten fand keine keine Unterteilung in Follikel-Theka- bzw. -Luteinzysten statt. Beim Vergleich der Gruppen traten in keinem Fall signifikante Unterschiede auf (P>0,05). Die Ergebnisse sind Tabelle 24 zu entnehmen. 83 Tabelle 24: Tiere mit Ovarialzysten 4-6 Wochen p.p. (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe Tiere n Tiere mit Ovarialzysten in % (n) C NaCl alle 93 89 12,90 (12) 14,61 (13) C NaCl Kühe 65 64 18,46 (12) 17,19 (11) C NaCl Färsen 28 25 0,00 (0) 8,00 (2) 4.5.3 Endometritis Carofertin soll laut Hersteller über eine Verbesserung der Uterusinvolution und eine Stimulation der Endometrium-Drüsen das Endometritisrisiko senken bzw. die Endometritistherapie unterstützen. Daraufhin wurde der Anteil der Tiere, die 4-6 Wochen p.p. eine Endometritis hatten, zwischen den Gruppen verglichen (alle Tiere, nur Kühe, nur Färsen). Dabei traten in keinem Fall signifikante Unterschiede zwischen den Gruppen auf (P>0,05). Die Ergebnisse sind Tabelle 25 zu entnehmen. Tabelle 25: Tiere mit Endometritis 4-6 Wochen p.p. (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe Tiere n Tiere mit Endometritis in % (n) C NaCl alle 93 89 30,11 (28) 22,47 (20) C NaCl Kühe 65 64 26,15 (17) 21,88 (14) C NaCl Färsen 28 25 39,29 (11) 24,00 (6) 84 4.6 Krankheiten und Abgänge (Langzeitstudie) 4.6.1 Peripartale Erkrankungen : Milchfieber und Nachgeburtsverhaltung Bei der Analyse wurde vor und nach der Abkalbung auftretendes Milchfieber gleich bewertet. Die Inzidenz von Milchfieber bzw. Nachgeburtsverhaltung wurde verglichen (alle Tiere, nur Kühe, nur Färsen). Es kam in keinem Fall zu signifikanten Unterschieden zwischen den beiden Gruppen (P>0,05). Die Ergebnisse sind Tabelle 26 zu entnehmen. Tabelle 26: Tiere mit Milchfieber bzw. Nachgeburtsverhaltung (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe Tiere n Tiere mit Milchfieber in % (n) Tiere mit Nachgeburtsverhaltung in % (n) C NaCl alle 100 101 17,00 (17) 20,79 (21) 11,00 (11) 15,84 (16) C NaCl Kühe 71 72 23,94 (17) 27,78 (20) 12,68 (9) 19,44 (14) C NaCl Färsen 29 29 0,00 (0) 3,45 (1) 6,90 (2) 6,90 (2) 4.6.2 Endometritis in der Gesamtlaktation Das Auftreten von Endometritis in der Gesamtlaktation wurde ebenfalls erfaßt (alle Tiere, nur Kühe, nur Färsen). Diese Auswertung enthält auch alle Fälle von Endometritis, die bei der Puerperalkontrolle diagnostiziert wurden. Beim Vergleich der Gruppen sind keine signifikante Unterschiede (P>0,05) aufgetreten (Tab. 27). Um die zum Teil sehr unterschiedliche Laktationsdauer und damit das Risiko zu erkranken miteinzubeziehen, wurden die Krankheitsfälle pro 100 Kuhtage „at risk“ 85 ermittelt ((Anzahl der Krankheitsfälle : Gesamtzahl der Laktationstage) × 100 Tage). Beim Vergleich der Endometritisinzidenz in Abhängigkeit von der Höhe der betaKarotinkonzentration zu den drei Zeitpunkten (Einteilung der Tiere nach Quartilen siehe Tabelle 35) traten in keinem Fall signifikante Gruppenunterschiede auf (P>0,05). Ebenso gab es keinen signifikanten Unterschied zwischen den Quartilen (P>0,05). Tabelle 27: Tiere mit Endometritis in der Gesamtlaktation (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) n Tiere mit Endometritis in % (n) Fälle pro 100 Kuhtage „at-risk“ alle 100 101 28,00 (28) 26,73 (27) 0,098 0,098 C NaCl Kühe 71 72 23,94 (17) 27,78 (20) 0,086 0,104 C NaCl Färsen 29 29 37,93 (11) 24,14 (7) 0,126 0,086 Gruppe Tiere C NaCl 4.6.3 Ovarialzysten in der Gesamtlaktation Das Auftreten von Ovarialzysten wurde zwischen den Gruppen verglichen (alle Tiere, nur Kühe, nur Färsen). Diese Auswertung enthält auch alle Fälle von Ovarialzysten, die bei der Puerperalkontrolle diagnostiziert wurden. Erkrankte ein Tier mehrmals wurde dies trotzdem als ein Fall gezählt. Beim Vergleich der Gruppen sind in keinem Fall signifikante Unterschiede aufgetreten. Die Ergebnisse sind Tabelle 28 zu entnehmen. Die Fälle pro 100 Kuhtage „at-risk“ wurden ebenfalls berechnet. Beim Vergleich der Ovarialzysteninzidenz in Abhängigkeit von der beta- Karotinkonzentration zu den drei Zeitpunkten trat nur in einen Fall ein signifikanter Gruppenunterschied auf. Die Karotintiere, die zum Zeitpunkt III zum Quartil III gehörten (beta-Karotinkonzentration zwischen 947 und 1176 µg/l) erkrankten signifikant weniger an Ovarialzysten (0,00 %) als die Kontrolltiere des Quartils III 86 (19,05 %, P<0,05). Ein signifikanter Unterschied zwischen den Quartilen war nicht vorhanden (P>0,05). Tabelle 28: Tiere mit Ovarialzysten in der Gesamtlaktation (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) n Tiere mit Ovarialzysten in % (n) Fälle pro 100 Kuhtage „at-risk“ alle 100 101 14,00 (14) 11,88 (12) 0,049 0,044 C NaCl Kühe 71 72 19,72 (14) 13,89 (10) 0,071 0,052 C NaCl Färsen 29 29 0,00 (0) 6,90 (2) 0,000 0,025 Gruppe Tiere C NaCl 4.6.4 Einfluß des beta-Karotin-Spiegels auf die Inzidenz von Nachgeburtsverhaltung, Endometritis und Ovarialzysten Beim Vergleich der Höhe der mittleren beta-Karotin-Serumkonzentration zu den drei Zeitpunkten zwischen erkrankten (Nachgeburtsverhaltung, Endometritis oder Ovarialzysten) und nicht-erkrankten Tieren lagen keine signifikanten Unterschiede vor (Tab. 29, P>0,05). 87 Tabelle 29: Mittlere beta-Karotin-Serumkonzentration bei an Nachgeburtsverhaltung, Endometritis oder Ovarialzysten erkrankten und nicht-erkrankten Tieren (n = Anzahl der Tiere) Erkrankung NachgeburtsVerhaltung keine Nachgeburtsverhaltung Endometritis keine Endometritis Ovarialzysten keine Ovarialzysten Zeitpunkt n MW ± SD I 27 1910,04 886,31 II 25 1061,40 404,86 III 25 1052,00 322,71 I 174 1951,51 768,28 II 164 1098,45 388,29 III 149 1013,36 412,82 55 I II III 55 55 55 1899,35 1138,64 939,72 835,23 464,12 350,60 146 I II III 146 134 119 1963,49 1075,04 1054,54 764,48 354,87 417,56 26 I II III 26 26 26 1978,00 1046,73 1081,81 780,07 315,42 485,42 175 I II III 175 163 148 1941,18 1101,01 1007,86 785,40 400,50 384,59 n 27 174 88 4.6.5 Abgänge Die Tabelle 30 enthält eine Aufstellung der Tiere, die im Versuchszeitraum aus verschiedenen Gründen abgegangen sind. Insgesamt sind von den 100 Tieren der Versuchsgruppe 33 Kühe und 11 Färsen abgegangen. Von den 101 Kontrolltieren sind 30 Kühe und 10 Färsen abgegangen Tabelle 30: Aufstellung der Abgangsgründe mit den dazugehörigen Tierzahlen (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe Tiere n geringe UnfruchtEuter Verendet Gliedmaßen Sonstiges Leistung barkeit C NaCl alle 100 101 5 1 19 12 10 6 4 4 4 6 2 11 C NaCl Kühe 71 72 5 1 12 8 9 4 3 4 2 6 2 7 C NaCl Färsen 29 29 0 0 7 4 1 2 1 0 2 0 0 4 Daraus ergibt sich für den Versuchszeitraum bei der Versuchsgruppe eine Remontierungsrate von 44,00 % und bei der Kontrollgruppe eine Remontierungsrate von 39,60 %. Vergleicht man die Remonierungsraten der Gruppen miteinander, so ergeben sich keine signifikanten Unterschiede. Tabelle 31 zeigt eine Auswertung der Abgänge im Bezug auf die Gruppen. Abgänge aufgrund von Verenden und Abgänge aus sonstigen Gründen wurden nicht einzeln ausgewertet, da diese Abgänge zum Teil aus sehr verschiedenen Gründen erfolgten. Diese Abgänge gingen nur in die Rubrik Abgänge insgesamt ein. In keinem Fall gab es signifikante Unterschiede zwischen den Gruppen (P>0,05). 89 Tabelle 31: Verteilung der verschiedenen Abgangsgründe im Bezug auf die einzelnen Gruppen, Angaben in % (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) n Abgänge insgesamt (%) alle 100 101 44,00 39,60 5,00 0,99 19,00 11,88 10,00 5,94 4,00 5,94 C NaCl Kühe 71 72 46,48 41,67 7,04 1,39 16,90 11,11 12,67 5,56 2,82 8,33 C NaCl Färsen 29 29 37,93 34,48 0,00 0,00 24,14 13,79 3,44 6,90 6,90 0,00 Gruppe Tiere C NaCl geringe Unfrucht- Euter Leistung barkeit (%) (%) (%) Gliedmaßen (%) 4.7 Fruchtbarkeitskennzahlen (Langzeitstudie) Es sollte untersucht werden, ob Carofertin zu einer Verbesserung der Fruchtbarkeit der Versuchstiere führte. In der Karotingruppe wurden 85 Tiere besamt. Davon wurden 60 Tiere tragend. In der Kontrollgruppe wurden ebenfalls 85 Tiere besamt, von denen 67 Tiere tragend wurden. Um die Fruchtbarkeitsleistung der Versuchstiere zu objektivieren, wurden folgende Fruchtbarkeitskennzahlen beurteilt: 4.7.1 Konzeptionsrate Die Konzeptionsrate der Karotingruppe wich nicht (alle Tiere, nur Kühe, nur Färsen) signifikant von der Konzeptionsrate der Kontrollgruppe ab (P>0,05). Die genauen Werte sind Tabelle 32 zu entnehmen. 90 Tabelle 32: Konzeptionsrate (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe Tiere n = tragende Tiere n = Anzahl aller Besamungen Konzeptionsrate (%) C NaCl alle 60 67 188 173 31,91 38,73 C NaCl Kühe 42 47 131 128 32,06 36,72 C NaCl Färsen 18 20 57 45 31,58 44,44 4.7.2 Gesamtträchtigkeitsrate Die Gesamtträchtigkeitsrate der Karotingruppe unterschied sich in keinem Fall (alle Tiere, nur Kühe, nur Färsen) signifikant von der Gesamtträchtigkeitsrate der Kontrollgruppe (P>0,05). Die genauen Werte sind Tabelle 33 zu entnehmen: Tabelle 33: Gesamtträchtigkeitsrate (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe Tiere n Gesamtträchtigkeitsrate in % (n) C NaCl alle 85 85 70,59 (60) 79,76 (67) C NaCl nur Kühe 58 60 72,41 (42) 78,33 (47) C NaCl nur Färsen 27 25 66,67 (18) 80,00 (20) 91 4.7.3 Erstbesamungserfolg 4.7.3.1 Auswertung nach Gruppen Der Erstbesamungserfolg der Karotingruppe unterschied sich in keinem Fall (alle Tiere, nur Kühe, nur Färsen) signifikant vom Erstbesamungserfolg der Kontrollgruppe (P>0,05, Tab. 34). Tabelle 34: Erstbesamungserfolg (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe Tiere n Erstbesamungserfolg in % (n) C NaCl alle 85 85 37,65 (32) 42,36 (36) C NaCl Kühe 58 60 37,93 (22) 41,67 (25) C NaCl Färsen 27 25 37,04 (10) 44,00 (11) 4.7.3.2 Auswertung nach Quartilen Um herauszufinden, ob die Höhe des beta-Karotin-Spiegels einen Einfluß auf den Erstbesamungserfolg hatte, wurden die gesamten Versuchstiere nach den Quartilen der beta-Karotin-Werte zu den drei Entnahmezeitpunkten unterteilt. Die Grenzen der Wertebereiche sind Tabelle 35 zu entnehmen. Beim Vergleich des Erstbesamungserfolges zwischen der Karotingruppe und der Kontrollgruppe innerhalb der Quartile lagen zu den drei Zeitpunkten keine signifikanten Unterschiede vor (Anhang: Tab. 57-59, P>0,05). Um festzustellen, ob die Höhe des beta-Spiegels allein eine Wirkung auf den Erstbesamungserfolg hat, wurden die Werte der Quartile (Karotin- und Kontrollgruppe) zusammengefasst und die Quartile miteinander verglichen. Ein signifikanter Unterschied lag nur zum Zeitpunkt II vor. Die Tiere der 92 Quartile I, III und IV hatten einen signifikant höheren Erstbesamungserfolg als die Tiere des Quartils II (Tab. 36, P<0,05). Tabelle 35: Übersicht über die Quartile der beta-Karotinkonzentrationen zu den drei Entnahmezeitpunkten Quartile Zeitpunkt I (µg/l) Zeitpunkt II (µg/l) Zeitpunkt III (µg/l) I ≤ 1414 ≤ 838 ≤ 751 II > 1414 ≤ 1850 > 838 ≤ 1044 > 751 ≤ 947 III > 1850 ≤ 2320 > 1044 ≤ 1290 > 947 ≤ 1176 IV > 2320 ≤ 5196 > 1290 ≤ 2870 > 1176 ≤ 2626 Tabelle 36: Erstbesamungserfolg zum Zeitpunkt III ausgewertet nach Quartilen (n = Anzahl der Versuchstiere) Quartil n Erstbesamungserfolg in % (n) I 44 45,45 (20) a II 42 23,81 (10) b III 42 42,86 (18) a IV 42 47,62 (20) a a,b = Prozentzahlen mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 4.7.4 Trächtigkeitsindex 4.7.4.1 Auswertung nach Gruppen Der Trächtigkeitsindex der Karotingruppe unterschied sich in keinem Fall (alle Tiere, nur Kühe, nur Färsen) signifikant von dem Trächtigkeitsindex der Kontrollgruppe (P>0,05). Die genauen Werte sind Tabelle 37 zu entnehmen. 93 Tabelle 37: Mittlerer Trächtigkeitsindex (n = Anzahl der Versuchstiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Gruppe Tiere n Trächtigkeitsindex ± SD C NaCl alle 60 67 1,72 1,76 0,92 0,99 C NaCl Kühe 42 47 1,71 1,83 0,92 1,07 C NaCl Färsen 18 20 1,72 1,60 0,95 0,75 4.7.4.2 Auswertung nach Quartilen Beim Vergleich der Trächtigkeitsindices der Karotinguppe mit denen der Kontrollgruppe innerhalb der Quartile der drei Zeitpunkte traten in keinem Fall signifikante Gruppenunterschiede auf (Anhang: Tab. 60-62, P>0,05). Zwischen den Quartilen gab es nur zum Zeitpunkt III eine signifikante Differenz. Der Trächtigkeitsindex der Tiere des Quartils III lag signifikant niedriger als der Trächtigkeitsindex der Tiere des Quartils II (Tab. 38, P<0,05). Tabelle 38: Mittlerer Trächtigkeitsindex zum Zeitpunkt III ausgewertet nach Quartilen (n = Anzahl der Versuchstiere, SD = Standardabweichung) Quartil n I 34 1,71 a, b 1,00 II 29 2,03 a 0,94 III 29 1,55 b 0,87 Trächtigkeitsindex ± SD IV 35 1,69 a, b 0,96 a,b = Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 94 4.7.5 Rast-, Güst-, Verzögerungs- und erwartete Zwischenkalbezeit 4.7.5.1 Auswertung nach Gruppen Der Versuch sollte zeigen, ob eine Gabe von Carofertin einen Einfluß auf die Rastzeit (RZ), die Güstzeit (GZ), die Verzögerungszeit (VZ) und daraus resultierend auf die erwartete Zwischenkalbezeit (ZKZ) hat. Die freiwillige Wartezeit betrug 60 Tage. Es kam zu keinen signifikanten Gruppenunterschieden (P>0,05). In den folgenden Tabellen sind die erreichten Werte zunächst für alle Tiere gemeinsam (Tab. 39) und dann nach Kühen (Tab. 40) und Färsen (Tab. 41) getrennt aufgeführt. Tabelle 39: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) aller Tiere (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Gruppe Variable n Mittelwert ± SD C RZ GZ VZ ZKZ 85 60 60 60 88,47 107,02 24,10 385,00 29,42 39,47 30,93 36,45 NaCl RZ GZ VZ ZKZ 85 67 67 67 84,46 107,96 25,43 385,95 25,58 37,24 32,57 37,24 95 Tabelle 40: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) der Kühe (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Gruppe Variable n Mittelwert ± SD C RZ GZ VZ ZKZ 58 42 42 42 85,91 107,19 23,60 385,17 26,66 34,98 30,17 34,95 NaCl RZ GZ VZ ZKZ 60 47 47 47 82,23 106,11 26,34 384,11 25,82 37,21 34,85 37,21 Tabelle 41: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) der Färsen (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Gruppe Variable n Mittelwert ± SD C RZ GZ VZ ZKZ 27 18 18 18 93,96 106,61 25,11 384,61 34,52 40,80 33,52 40,80 NaCl RZ GZ VZ ZKZ 25 20 20 20 90,04 112,30 23,30 390,30 24,61 37,90 27,19 37,90 Da die Kühe der Karotingruppe, die zum Zeitpunkt I einen beta-Karotin-Wert unter 2000 µg/l hatten, einen signifikant höheren beta-Karotin-Wert zum Zeitpunkt III aufwiesen als die Kühe der Kontrollgruppe, wurden diese Kühe mit denen der Kontrollgruppe bezüglich ihrer Rast-, Güst-, Verzögerungs- und Zwischenkalbezeit verglichen. Es traten keine signifikanten Gruppenunterschiede auf (P>0,05, Tab. 42). 96 Tabelle 42: Auswertung der Kühe (Tiere zum Zeitpunkt I unter 2000 µg/l betaKarotin): Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Gruppe Variable n Mittelwert ± SD C RZ GZ VZ ZKZ 32 22 22 22 86,56 105,14 22,55 383,14 24,22 32,91 33,74 32,91 NaCl RZ GZ VZ ZKZ 32 25 25 25 82,78 110,28 33,70 388,28 29,68 38,25 35,89 38,25 4.7.5.2 Auswertung nach Quartilen Um herauszufinden, ob die Höhe des beta-Karotin-Spiegels einen Einfluß auf die Rastzeit, Güstzeit, Zwischenkalbezeit oder die Verzögerungszeit hatte, wurden die gesamten Versuchstiere nach den Quartilen der beta-Karotin-Werte zu den drei Entnahmezeitpunkten unterteilt (Tab. 35). 4.7.5.2.1 Zeitpunkt I Es erfolgte eine Auswertung der Rastzeit, der Güstzeit, der Verzögerungszeit und der Zwischenkalbezeit nach Gruppen (Karotingruppe gegen Kontrollgruppe) innerhalb der vier Quartile. Dabei traten keine signifikanten Unterschiede auf. Danach folgte ein Vergleich der Quartile gegeneinander innerhalb der Karotingruppe. Dabei war die Verzögerungszeit der Tiere des Quartils IV signifikant niedriger als die Verzögerungszeit der Tiere der Quartile II und III (Anhang: Tab. 63, P<0,05). Beim Vergleich der Quartile der Kontrollgruppe traten keine signifikanten Unterschiede auf (Anhang: Tab. 64, P>0,05). Beim Vergleich der Quartile gegeneinander (Karotin- und Kontrollgruppe der Quartile zusammengenommen) gab es ebenfalls keine 97 signifikanten Unterschiede (Tab. 43, P>0,05). Auffällig war allerdings, dass die mittlere Rastzeit und die mittlere Güstzeit der Tiere des Quartils I deutlich höher waren als die Rastzeit bzw. Güstzeit der Tiere des Quartils IV. Tabelle 43: Auswertung der Quartile gegeneinander zum Zeitpunkt I: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Variable n Mittelwert ± SD I RZ GZ VZ ZKZ 47 36 36 36 90,40 110,59 24,86 388,65 28,61 39,96 35,99 40,13 II RZ GZ VZ ZKZ 41 32 32 32 85,32 109,59 28,72 387,59 28,40 39,36 32,50 39,36 III RZ GZ VZ ZKZ 40 29 29 29 87,15 113,21 26,69 391,21 32,59 36,03 30,77 36,03 IV RZ GZ VZ ZKZ 42 30 30 30 81,23 100,27 20,83 382,65 19,42 37,91 31,78 44,60 Quartil 4.7.5.2.2 Zeitpunkt II Es wurde wie beim Zeitpunkt I verfahren. Beim Vergleich der Gruppen nach Quartilen traten keine signifikanten Unterschiede auf (P>0,05). In der Karotingruppe war die Rastzeit der Tiere im Quartil III signifikant niedriger als im Quartil II (Anhang: Tab. 65, P<0,05). In der Kontrollgruppe lag die Verzögerungszeit der Tiere im Quartil 98 IV signifikant niedriger als in den Quartilen I und III. Dasselbe gilt für die Verzögerungszeit der Tiere im Quartil II zum Quartil I (Anhang: Tab. 66, P<0,05). Beim Vergleich der Quartile gegeneinander war die Verzögerungszeit der Tiere im Quartil IV signifikant niedriger als im Quartil I (P<0,05, Tab. 50). In den übrigen Fällen lagen keine signifikanten Unterschiede vor. Tabelle 44: Auswertung der Quartile gegeneinander zum Zeitpunkt II: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Quartil Variable n Mittelwert ± SD I RZ GZ VZ ZKZ 46 38 38 38 83,72 116,56 34,67 a 394,54 26,82 41,56 37,77 41,54 II RZ GZ VZ ZKZ 40 26 26 26 92,70 110,38 20,85 a,b 393,07 30,51 37,95 30,13 44,48 III RZ GZ VZ ZKZ 42 32 32 32 79,63 107,16 28,75 a,b 385,16 26,19 37,65 31,93 37,65 RZ 42 88,02 28,98 GZ 31 98,23 34,09 IV VZ 31 16,29 b 26,81 ZKZ 31 376,29 34,36 a,b Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 99 4.7.5.2.3 Zeitpunkt III Beim Vergleich der Gruppen nach Quartilen traten keine signifikanten Gruppenunterschiede auf (P>0,05). In der Karotingruppe (Anhang: Tab. 67) war die Zwischenkalbezeit der Tiere im Quartil III signifikant niedriger als im Quartil II (P<0,05). Die Rastzeit der Tiere war im Quartil II, III und IV signifikant niedriger als im Quartil I (P<0,05). In der Kontrollgruppe gab es keine signifikanten Unterschiede (Anhang: Tab. 68, P>0,05). Wertet man die Quartile beider Gruppen gegeneinander aus, lag die mittlere Rastzeit der Tiere der Quartile III und IV signifikant niedriger als die mittlere Rastzeit der Tiere des Quartils I (P<0,05, Tab. 45). Tabelle 45: Auswertung der Quartile gegeneinander zum Zeitpunkt III: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD Standardabweichung) Quartil Variable n Mittelwert ± SD I RZ GZ VZ ZKZ 43 34 34 34 93,16 a 111,88 24,03 389,88 27,94 35,11 33,03 35,11 II RZ GZ VZ ZKZ 41 28 28 28 88,85 a,b 117,14 31,00 399,13 28,57 34,02 30,01 39,95 III RZ GZ VZ ZKZ 43 29 29 29 81,31 b 100,76 21,57 378,76 31,97 40,08 32,27 40,08 IV RZ GZ VZ ZKZ 43 36 36 36 80,21 b 104,64 26,97 382,67 22,81 42,59 35,73 42,74 a,b Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant = 100 4.7.5.3 Lineare Regressionsanalyse Die Höhe des beta-Karotin-Spiegels der Versuchstiere zum Zeitpunkt II hatte einen signifikanten Einfluß auf die Güstzeit, die Verzögerungszeit und die Zwischenkalbezeit der Versuchstiere (alle Tiere, keine Gruppenunterteilung, P<0,05). Dasselbe galt für den beta-Karotin-Spiegel zum Zeitpunkt III und die Rastzeit der Versuchstiere. Mit zunehmendem beta-Karotinspiegel nahmen die Rastzeit, die Güstzeit, die Verzögerungszeit und die Zwischenkalbezeit ab (Tab. 46). Weitere Korrelationen lagen nicht vor. Tabelle 46: Einfluß der Höhe des beta-Karotin-Spiegels auf Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und Zwischenkalbezeit (ZKZ) RZ GZ VZ ZKZ Korrelationskoeffizient Zeitpunkt II Korrelationskoeffizient Zeitpunkt III kein signifikanter Einfluß - 0,19 - 0,19 - 0,19 - 0,16 kein signifikanter Einfluß kein signifikanter Einfluß kein signifikanter Einfluß 4.7.6 Prozentsatz der Tiere mit einer Güstzeit unter 115 Tagen Der Prozentsatz der Tiere, die eine Güstzeit unter 115 Tagen hatten, war für beide Gruppen in keinem Fall (alle Tiere, nur Kühe, nur Färsen) signifikant unterschiedlich (P>0,05, Tab. 47). 101 Tabelle 47: Anteil der Tiere mit einer Güstzeit unter 115 Tagen (n = Anzahl der Tiere) Gruppe Tiere n Güstzeit unter 115 Tagen in % (n) C NaCl alle 60 67 60,00 (36) 59,70 (40) C NaCl Kühe 42 47 61,90 (26) 61,70 (29) C NaCl Färsen 18 20 55,56 (10) 55,00 (11) 4.7.7 200-Tage-Nichtträchtigkeitsrate Die 200-Tage-Nichtträchtigkeitsrate beinhaltet alle abgekalbten Tiere, die 200 Tage p.p. noch nicht tragend waren. Tiere, die bewusst güst gesetzt worden sind und Tiere, die schon vor 200 Tagen p.p. abgegangen waren, gehörten nicht dazu. Es lagen in beiden Gruppen in keinem Fall (alle Tiere, nur Kühe, nur Färsen) signifikante Unterschiede vor (P>0,05, Tab. 48). Tabelle 48: Anteil der Tiere, die innerhalb von 200 Tagen post partum nicht tragend waren (n = Anzahl der Tiere, p.p. = post partum) Gruppe Tiere n Anteil nicht tragender Tiere 200 Tage p.p. in % (n) C NaCl alle 100 101 17,00 (17) 14,85 (15) C NaCl Kühe 71 72 15,49 (11) 13,89 (10) C NaCl Färsen 29 29 20,69 (6) 17,24 (5) 102 4.7.8 Prozentsatz der Tiere, die 60 Tage post partum noch nicht in Brunst gesehen wurden In diese Analyse wurden alle Tiere miteinbezogen abzüglich der Tiere, die bewusst güst waren, und abzüglich der Tiere, die unter 60 Tagen p.p. abgegangen sind. Es ergab sich nur bei den Färsen ein signifikanter Unterschied. In der Gruppe der Färsen, die Karotin erhalten hatten, waren signifikant mehr Tiere erst über 60 Tage post partum in Brunst gesehen worden als in der Kontrollgruppe (Tab. 49). Tabelle 49: Anteil der Tiere, die 60 Tage p.p. noch nicht in Brunst gesehen wurden (n = Anzahl der Tiere, p.p. = post partum Gruppe Tiere n 1. Brunst gesehen über 60 Tage p.p. in % (n) C NaCl alle 85 85 58,82 (50) 47,06 (40) C NaCl Kühe 58 60 58,62 (34) 55,00 (33) C 27 59,26 (16) a Färsen NaCl 25 28,00 (7) b a,b Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 4.7.9 Prozentsatz abortierender Tiere Diese Auswertung bezieht sich auf alle besamten Versuchstiere, die im Versuchszeitraum abortiert haben. Unter Abort wurde dabei ein Verwerfen zwischen dem 45. und 265. Tag der Trächtigkeit verstanden. Diese Tiere waren also schon als tragend befunden worden (rektale Palpation ca. 5 Wochen post inseminationem) und fielen dann durch Auftreten einer Brunst bzw. Beobachten des Aborts auf. Daraufhin erfolgte eine Nachuntersuchung dieser Tiere mit Feststellung des Aborts. Zwischen den Gruppen (alle Tiere, nur Kühe, nur Färsen) gab es keine signifikanten Unterschiede (P>0,05, Tab. 50). 103 Tabelle 50: Anteil der Tiere, die zwischen dem 45. und 265. Tag der Trächtigkeit abortiert haben (n = Anzahl der Tiere) Gruppe Tiere n Abortierende Tiere in % (n) C NaCl alle 60 67 8,33 (5) 10,45 (7) C NaCl Kühe 42 47 11,90 (5) 8,51 (4) C NaCl Färsen 18 20 0,00 (0) 15,00 (3) 4.8 Progesteronbestimmung Carofertin soll durch eine Stabilisierung der Corpora lutea zu einer Anhebung des Progesteronspiegels führen. Zur Überprüfung dieses Sachverhaltes wurden die Lutealphasen von 10 Tieren aus der Karotingruppe und 10 Tieren aus der Kontrollgruppe ausgewertet. Bei den aufgezeichneten Gelbkörperphasen wurden die mittleren Progesteronserumkonzentrationen zu den einzelnen Entnahmezeitpunkten berechnet. Der Beginn und das Ende der Lutealphase wurde bei 1 nmol/l Progesteron festgelegt. Die mittlere Progesteronkonzentration Lutealphase ist der Abbildung 3 zu entnehmen. während der Progesteronkonzentration im Plasma (nmol/l) 104 25,00 20,00 15,00 10,00 5,00 0,00 1 2 3 4 5 6 7 8 9 Entnahmezeitpunkte Karotin Abbildung 3: Verlauf der mittleren NaCl Progesteronkonzentration während der Lutealphase (Definition Entnahmezeitpunkte siehe Seite 68) Um die produzierte Gesamtprogesteronmenge (Progges) zu ermitteln, wurde bei jedem Tier die Fläche unter der Kurve mittels Intergralrechnung ermittelt. Zusätzlich wurde für jedes Tier die Progesteronmenge pro Tag (Progtag) berechnet, indem die in der Lutealphase produzierte Progesteronmenge durch die Anzahl der Tage, die die Lutealphase gedauert hat, geteilt wurde. Zusätzlich wurde der höchste Progesterontageswert (Progmax), der in dieser Lutealphase vom jeweiligen Tier an einem Tag erreicht wurde, aufgezeichnet (Tab. 51). Bei der Auswertung der Mittelwerte dieser drei Parameter ergaben sich keine signifikanten Unterschiede zwischen den Gruppen (P>0,05). Die Mittelwerte mit Standardabweichung sind der Tabelle 52 zu entnehmen. 105 Tabelle 51: Progesteronparameter aller Tiere (C = Karotingruppe, NaCl = Kontrollgruppe) Gruppe Tiere Gesamtprogesteron- Progesteron Maximale Progesterontagesmenge (nmol/l) pro Tag (nmol/l) konzentration (nmol/l) Kühe C Färsen Kühe NaCl Färsen 133,05 7,39 17,10 247,50 11,79 22,30 129,00 7,59 12,20 153,10 6,96 11,80 212,40 9,23 25,30 353,20 16,05 35,30 251,65 11,44 19,30 233,20 11,10 20,20 214,90 10,75 15,90 223,20 8,59 12,60 247,95 13,78 28,20 189,35 9,02 18,90 250,87 9,29 17,40 331,20 17,43 29,00 186,35 9,32 17,30 307,30 15,37 28,40 228,70 12,70 23,30 236,40 13,13 29,90 127,95 7,53 11,90 162,55 9,03 18,30 106 Tabelle 52: Mittlere Gesamtprogesteronmenge (Progges), mittlere Progesteronkonzentration pro Tag (Progtag) und mittlere maximale Progesteronkonzentration (Progmax) während der Lutealphase (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Gruppe C NaCl C NaCl C NaCl Tiere alle Tiere alle Tiere Kühe Kühe Färsen Färsen n Variable Mittelwert ± SD 10 Progges Progtag Progmax 215,12 10,09 19,20 66,54 2,75 7,22 10 Progges Progtag Progmax 226,86 11,66 22,26 62,75 3,28 6,33 5 Progges Progtag Progmax 175,01 8,59 17,74 52,47 1,98 6,01 5 Progges Progtag Progmax 241,14 11,77 22,16 59,03 3,73 5,92 5 Progges Progtag Progmax 255,23 11,59 20,66 56,46 2,73 8,72 5 Progges Progtag Progmax 212,58 11,55 22,36 69,76 3,20 7,42 107 4.9 Kosten pro Trächtigkeit Die Kosten-Nutzen-Analyse erfolgte in Anlehnung an die Dissertation von Tischer (1998). Die Kosten für tierärztliche Tätigkeiten und Medikamente sind in der Tabelle 53 aufgelistet und richteten sich nach der GOT (einfacher Satz) bzw. nach der Arzneimittelpreisverordnung. Anfahrtskosten wurden nicht berechnet. Tabelle 53: Kosten der Fruchtbarkeitsbehandlungen und Trächtigkeits- untersuchungen (incl. 16 % Mwst.) Verrichtung Injektion von 20 ml Carofertin Trächtigkeitsuntersuchung Versuchte Ablösung der Nachgeburt und Einführung von 2 Tetra-SleecolUterusstäben Nachgeburt abgelöst und Einführung von 2 Tetra-Sleecol-Uterusstäben Einführung von 2 Tetra-SleecolUterusstäben zur Nachbehandlung Rektale Untersuchung und intramuskuläre Injektion von 2 ml PGF Veyx forte Rektale Untersuchung und intramuskuläre Injektion von 5 ml Receptal Uterusinstillation mit 1 Injektor Metricure Preis (incl. 16 % Mwst.) 8,71 € 7,67 € 18,90 € 26,56 € 6,11 € 17,14 € 18,41 € 21,70 € Die Spermakosten pro Portion lagen bei 7,50 € und ein Güsttag über 85 Tage p.p. wurde mit 2,50 € veranschlagt. Die Remontierungskosten pro Kuh beliefen sich auf 425,20 €. Die Kosten pro Trächtigkeit der Karotin- und der Kontrollgruppe sind den Tabellen 54 und 55 zu entnehmen. 108 Tabelle 54: Kosten pro Trächtigkeit in der Karotingruppe Kostenfaktor Kosten für das Carofertin Kosten für Fruchtbarkeitsbehandlungen Kosten durch Güsttage über 85 Tage p.p. Spermakosten Kosten durch Trächtigkeituntersuchungen Abgänge wegen Unfruchtbarkeit Gesamtkosten : 79 Tiere (60 tragende + 19 remontierte Tiere) Kosten pro Trächtigkeit pro Tier Preis 2613,00 € 2070,00 € 4075,00 € 1410,00 € 853,76 € 8078,80 € 19100,56 € : 79 241,78 € Tabelle 55: Kosten pro Trächtigkeit in der Kontrollgruppe Kostenfaktor Fruchtbarkeitsbehandlungen Kosten durch Güsttage über 85 Tage p.p. Spermakosten Kosten durch Trächtigkeitsuntersuchungen Abgänge wegen Unfruchtbarkeit Gesamtkosten : 79 Tiere (67 tragende + 12 remontierte Tiere) Kosten pro Trächtigkeit pro Tier Preis 1683,98 € 4682,50 € 1305,00 € 767,05 € 5102,40 € 13540,93 € : 79 171,40 € 109 5 Diskussion 5.1 Verlauf der beta-Karotin-Konzentration (Verlaufsstudie) Als Ausgangspunkt der vorliegenden Untersuchungen sollte zunächst abgeklärt werden, ob sich der Blutserumspiegel der trockenstehenden Versuchstiere durch eine einmalige intramuskuläre Injektion von 20 ml Carofertin erhöht. Dies entsprach einer Menge von 200 mg natürlichem beta-Karotin. Desweiteren wurde durch tägliche Überprüfung der Verlauf des beta-Karotin-Spiegels im Blutserum dokumentiert (Tab. 17). Es zeigte sich, dass die Ausgangskonzentrationen des betaKarotins von Tier zu Tier stark schwankten. Der Mittelwert lag in der Karotingruppe bei 2061 ± 557,90 µg/l und in der Kontrollgruppe bei 2033 ± 732,21 µg/l. Nach der Injektion des Carofertin kam es nur beim Vergleich des Ausgangswertes der Karotingruppe mit dem Entnahmezeitpunkt IV der Karotingruppe zu einer signifikant höheren beta-Karotin-Konzentration im Serum. Am Entnahmezeitpunkt IV lag die mittlere beta-Karotin-Konzentration in der Karotingruppe bei 2668 ± 602,96 µg/l. Beim Vergleich der mittleren beta-Karotin-Konzentration der einzelnen Zeitpunkte zwischen den Gruppen, gab es zu keinem Zeitpunkt einen signifikanten Unterschied. Dasselbe galt für den Vergleich der Ausgangskonzentrationen mit den anderen Zeitpunkten (außer Zeitpunkt IV in der Karotingruppe) sowohl in der Karotingruppe als auch in der Kontrollgruppe. Auffällig war, dass die mittleren beta-KarotinSerumkonzentrationen in der Karotingruppe trotzdem zu allen Zeitpunkten höher waren als die der Kontrollgruppe (Abb. 2). 12 Tage nach der Injektion war die Ausgangskonzentration in der Karotingruppe wieder erreicht (1927 ± 499,43 µg/l). Danach fielen die beta-Karotin-Spiegel aller Versuchtiere langsam ab, was mit dem näherrückenden Kalbetermin in Zusammenhang zu sehen war. Friesecke (1978) zeigte, dass der wichtigste Speicher für beta-Karotin im Körper des Rindes das Blutplasma ist. In den Versuchen von Lothammer (1978) wurde das betaKarotin oral verabreicht und führte im Vergleich zu einer beta-Karotin-arm ernährten 110 Tiergruppe zu höheren Blutspiegeln. Eisele (1987) bewies, dass Rinder auch parenteral verabreichtes beta-Karotin verwerten können. Nach einer intramuskulären Injektion von 500 mg beta-Karotin erhöhte sich der Blutserumspiegel bei Ausgangswerten von ca. 900 µg/l um das Zehnfache. Diese Verzehnfachung blieb für 10 Tage bestehen und nach 3 Wochen lagen die Blutserumkonzentrationen noch über 3000 µg/l. Burgstaller et al. (1980) machte deutlich, dass der Blutserumspiegel des beta-Karotins durch die einsetzenden Kolostrumproduktion ante partum stark abfällt. Lotthammer et al. (1977) berichteten von großen individuelle Schwankungen der beta-Karotin-Serumkonzentration ihrer Versuchstiere. Man erklärte diese mit einem unterschiedlichem Resorptions- und Mobilisierungsvermögen der Tiere. Ein Zusammenhang mit der Schilddrüsenfunktion wurde vermutet. Dies ist aber bislang nicht näher untersucht worden. Lotthammer (1999) hielt 1000 µg/l beta-Karotin im Blutserum für den Zeitraum von 3 Wochen ante bzw. post partum für ausreichend. Das VetMedLab Ludwigsburg ging basierend auf amerikanischer Literatur ab einem Blutserumspiegel von 700 mg/l beta-Karotin unabhängig vom Laktationsstadium von einer ausreichenden beta-Karotin-Versorgung aus (Smith 1996). Bei Friesecke (1978) gelten Werte unter 3000 µg/l generell als beta-Karotin-Mangel. In dieser Untersuchung wurde keine starke Erhöhung der beta-Karotin-Spiegel über einen längeren Zeitraum erreicht. Die mittleren beta-Karotin-Serumkonzentrationen lagen zu den Entnahmezeitpunkten allerdings für 12 Tage immer höher als die der Kontrollgruppe. Die Unterschiede waren aber nicht signifikant (P>0,05). Dies kann zum einen an der um mehr als die Hälfte geringeren Dosis an beta-Karotin als in dem Versuch von Eisele (1987) liegen und zum anderen an der Tatsache, dass in unserer Untersuchung trockenstehende Tiere verwendet wurden. Wahrscheinlich lies sich der Mehrbedarf an beta-Karotin durch die einsetzende Kolostrumbildung nicht durch die Injektion von 200 mg beta-Karotin decken, so dass die Reserven im Blutplasma angegriffen wurden und der beta-Karotin-Spiegel zwangsläufig sank. Die stark unterschiedlichen Werte innerhalb der Versuchsgruppen, aber auch bei denselben Tieren von Tag zu Tag bestätigten die Ergebnisse von Lotthammer et al. (1977). Eine Erklärung dafür konnte aus den vorliegenden Untersuchungen nicht abgeleitet werden, so dass weiterhin von einem individuell unterschiedlichen 111 Resorptions- und Mobilisierungsvermögen der Kühe ausgegangen werden muß. Bezüglich der Einschätzung der Versorgungslage unserer Versuchstiere kamen wir unter zu Hilfenahme der Referenzwerte nach Lotthammer (1999) und Smith und Bradford (1996) zu dem Schluß, dass unsere Versuchtiere der Verlaufsstudie zufriedenstellend über das Futter mit beta-Karotin versorgt waren. 5.2 Vitamin-A, Vitamin-E- und Selenversorgung (Langzeitstudie) Der Hersteller des Carofertins wies in seiner Produktinformation daraufhin, dass vor der Anwendung dieses Präparats andere Negativfaktoren auf die Fruchtbarkeit ausgeschlossen werden müssen. An erster Stelle wurde dabei ein Vitamin-A-Mangel erwähnt, dicht gefolgt von einer schlechten Versorgung mit Vitamin E und Selen. 5.2.1 Vitamin A Bei einer Stichprobe unserer Versuchstiere der Langzeitstudie (jeweils 20 Tiere aus jeder Gruppe und an Zeitpunkt I und Zeitpunkt II, Tab. 18) waren bei mittleren Vitamin-A-Spiegeln von 0,54 ± 0,20 mg/l in der Karotingruppe und 0,52 ± 0,25 mg/l in der Kontrollgruppe über 95 % der Tiere ausreichend mit Vitamin A versorgt (Normalwert > 0,3 mg/l). Nach Kolb (1994) führte ein Vitamin-A-Mangel zu einer Häufung von Endometritisfällen im Bestand. Außerdem soll Vitamin A für die Bildung von proteolytischen Enzymen, die durch Auflösung der Follikelmembran Voraussetzung für die Ovulation sind, wichtig sein (Zerobin 1987). Da unsere Versuchstiere zu über 95 % ausreichend mit Vitamin A versorgt waren, konnte ein Vitamin-A-Mangel ausgeschlossen werden. 112 5.2.2 Vitamin E Die Vitamin-E-Konzentration im Serum lag bei den Tieren der Stichprobe mit 5,18 ± 2,42 mg/l bei 82,5 % der Tiere zum Zeitpunkt I (3-4 Wochen a.p.) über dem Normalwert von >3,0 mg/l. Zum Zeitpunkt II erreichten dies nur 55 % der Versuchstiere (MW 3,11 ± 1,09 mg/l). Da in neueren Untersuchungen kein positiver Effekt von Vitamin E auf die Herdenfruchtbarkeit mehr festgestellt wurde (Allison u. Laven 2000, LeBlanc et al. 2002), wurde diese etwas kritische Versorgungslage der Versuchstiere mit Vitamin E bei der Interpretation der anderen Versuchsergebnisse vernachlässigt. 5.2.3 Selen Bei den Tieren der Stichprobe war die Selenversorgung mit 90 % ausreichend versorgten Tieren zum Zeitpunkt II (1-2 Wochen p.p.) zufriedenstellend (MW 60,60 ± 16,70 µg/l, Normalwert > 40,0 µg/l). Bei den trockenstehenden Tieren waren bei einer mittleren Serumkonzentration von 49,40 ± 13,00 µg/l 70 % der Tiere ausreichend mit Selen versorgt. Dieser geringere Prozentsatz war durch die Ration der Trockensteher bedingt. Die Trockensteher erhielten 3-4 Wochen kein Milchleistungsfutter und auch kein selenhaltiges Mineralfutter, so dass sie ihren Selenbedarf allein aus dem Grundfutter decken mussten. Ein Selenmangel hatte in vielen Untersuchungen negative Folgen auf die Fruchtbarkeit. Es kam zu einer Häufung von Nachgeburtsverhaltung und dadurch zum vermehrten Auftreten von Endometritis (Kappel et al. 1984). Eine Supplementierung von Selen verbesserte die Fruchtbarkeit bei Selenmangel deutlich (Jukola et. al. 1996a). Bei der Abkalbung war der Selenbedarf bei 90 % unserer Versuchstiere gedeckt. Somit war von keiner direkten negativen Beeinflussung in den eigenen Untersuchungen über die Fruchtbarkeit der Herde durch einen Selenmangel auszugehen. Da der Prozentsatz an Tieren mit Nachgeburtsverhaltung in der Karotingruppe mit 11,11 % und in der 113 Kontrollgruppe mit 15,84 % im Normalbereich lag (in der Gesamtherde < 15 %), kann die mangelhafte Selenversorgung der Trockensteher in diesem Zusammenhang auch keine gravierende Rolle gespielt haben. 5.3 Weitere Voraussetzungen für die Anwendung von Carofertin (Langzeitstudie) Es sollte laut der Firma Alvetra ebenso ein zu hoher Peroxidgehalt und ein zu hoher Mykotoxingehalt im Futter ausgeschlossen werden. Da bei der Analyse der Mischration durch die Lufa Hameln eine sehr gute bis gute Qualität des Futters bestätigt wurde und sensorisch zu keinem Zeitpunkt der Untersuchung Hinweise auf einen Verderb des Futters vorlagen, wurde auf eine Abklärung dieser Parameter im Labor verzichtet. Ein Mangel an Phosphor, Mangan, Zink, Molybdän und Jod wurde ebenfalls nicht überprüft, da einerseits ein negativer Einfluß eines Mangels dieser Spurenelemente auf die Fruchtbarkeit umstritten ist (Whitaker 1999) und andererseits bei den an unsere Versuchstiere verfütterten Rationen von einer ausreichenden Supplementierung dieser Stoffe (außer Molybdän) über das Milchleistungsfutter (Phosphor) bzw. das Mineralfutter (Phosphor, Zink, Mangan, Jod) ausgegangen werden konnte. Zusammenfassend ist zu sagen, dass bei den Versuchstieren die laut Hersteller geforderten Rahmenbedingungen für eine Anwendung des Carofertin erfüllt waren. 5.4 Beta-Karotin-Gehalte im Blutserum (Langzeitstudie) Zunächst wurde der beta-Karotin-Spiegel vor der ersten Injektion bei allen Versuchstieren bestimmt. 3-4 Wochen ante partum lag die mittlere Blutserumkonzentration des beta-Karotins bei der Versuchsgruppe bei 1948,51 ± 864,87 µg/l und bei der Kontrollgruppe bei 1943,40 ± 696,58 µg/l (Tab. 19). Bei den 114 Kühen der Versuchsgruppe war die mittlere beta-Karotin-Serumkonzentration 2143,62 ± 901,74 µg/l und bei den Färsen der Versuchsgruppe 1470,83 ± 528,24 µg/l. In der Kontrollgruppe lagen die Werte bei 1974,21 ± 720,53 µg/l bei den Kühen und 1866,90 ± 638,74 µg/l bei den Färsen. Zu allen drei Entnahmezeitpunkten kam es bei allen Versuchstiere zu großen individuellen Schwankungen. Diese Tatsache wird bei der Betrachtung der jeweiligen Standardabweichungen deutlich. Zum Zeitpunkt II lag die mittlere beta-Karotin-Konzentration bei der Versuchsgruppe bei 1123,47 ± 386,44 µg/l und bei der Kontrollgruppe bei 1063,30 ± 392,53 µg/l. Zum Zeitpunkt III war die mittlere beta-Karotin-Serumkonzentration der Karotingruppe 1013,90 ± 385,80 µg/l und die der Kontrollgruppe 1024,15 ± 15 µg/l. Bis auf zwei Ausnahmen gab es keine signifikanten Unterschiede zwischen der Karotin- und der Kontrollgruppe. Zum Zeitpunkt I hatte die Kontrollgruppe der Färsen eine signifikant höhere mittlere beta-Karotin-Serumkonzentration als die Karotingruppe. Die zweite Ausnahme stellte die Untergruppe der Kühe, die zum Zeitpunkt I einen beta-KarotinWert unter 2000 µg/l hatten dar. In diesem Fall lag die beta-KarotinSerumkonzentration zum Zeitpunkt III in der Karotingruppe signifikant höher als in der Kontrollgruppe. Innerhalb der Gruppen lagen die Werte in der Karotingruppe zum Zeitpunkt I stets signifikant höher als zum Zeitpunkt II und III. Dasselbe gilt für die Kontrollgruppe. Die Werte des Zeitpunkts II waren in der Karotingruppe bei der Auswertung aller Tiere und bei der Auswertung der Färsen signifikant höher als die Werte des Zeitpunkts III. Bei der getrennten Betrachtung der Kühe gab es keinen signifikanten Unterschied zwischen den Werten des Zeitpunkts II und III. In der Kontrollgruppe trat in keinem Fall ein signifikanten Unterschied zwischen Zeitpunkt II und III auf. Es gilt als gesichert, dass eine positive Korrelation zwischen den beta-KarotinGehalten im Futter und den beta-Karotin-Gehalten im Blutplasma besteht (Block u. Farmer 1987). Über die Referenzwerte für beta-Karotin im Blut von Milchkühen herrscht allerdings keine Einigkeit. Kolb (1997) ging beim Rind bei einer karotinreichen Fütterung von Gehalten von 3000-5000 µg/l aus. Smith und Bradford (1996) halten Werte über 3000 µg/l für erstrebenswert, Werte von 1500-3970 µg/l für normal und Werte unter 700 µg/l für mangelhaft. Lotthammer (1999) stellte einen 115 Zusammenhang zwischen dem Laktationsstadium bzw. der Trächtigkeit her. Er hielt Werte über 1000 µg/l in einen Zeitraum von 3 Wochen ante partum bis 3 Wochen post partum für ausreichend. In der übrigen Zeit sollten die Blutserumgehalte über 2000 µg/l liegen. Burgstaller et al. (1980) zeigten, dass die Blutserumspiegel ante partum stark abfielen und es Wochen dauerte, bis die Ausgangskonzentrationen wieder erreicht wurden. Hinzukommen noch die bei der Verlaufsstudie bereits erwähnten starken individuellen Schwankungen (Lotthammer u. Ahlswede 1977). In den vorliegenden Untersuchungen wurde sich an den Referenzwerten von Lothammer (1999) orientiert. Zum Zeitpunkt I wurde von einer ausreichenden betaKarotin-Versorgung der Versuchtiere ausgegangen. Die Kühe der Karotingruppe waren dabei im Mittel etwas besser versorgt (2143,62 ± 901,74 µg/l) als die Färsen der Karotingruppe (1470,83 ± 528,24 µg/l). In der Kontrollgruppe war der Unterschied zwischen Kühen (1974,21 ± 720,53) und Färsen (1866,90 ± 638,74) wesentlich geringer. Da die Tiere zufällig in die Gruppen eingeteilt wurden, waren diese unterschiedlichen Ausgangswerte sicherlich durch die zum Teil erheblichen individuellen Unterschiede bedingt, die schon bei der Verlaufsstudie auffällig waren. Die Standardabweichungen sämtlicher Mittelwerte machten dies zu jedem Entnahmezeitpunkt und bei jeder Gruppe (Karotingruppe, Kontrollgruppe, alle Tiere, nur Kühe, nur Färsen) deutlich. Der signifikant höhere mittlere beta-Karotin-Gehalt der Färsen der Kontrollgruppe im Vergleich mit der Karotingruppe war deshalb auch auf diese Schwankungen zurückzuführen. Somit wurden die Ergebnisse von Lotthammer und Ahlswede (1977) im Bezug auf die starken individuellen Unterschiede in der beta-Karotin-Resorption und –Mobilisation auch in der Langzeitstudie bestätigt. Der Vergleich der drei Entnahmezeitpunkte zeigte wie von Burgstaller et al. (1980) beschrieben einen Abfall der beta-Karotin-Konzentration zwischen Zeitpunkt I und Zeitpunkt II um fast Hälfte der Ausgangskonzentration. Außerdem wurden die Ausgangswerte bis zur Besamung nicht wieder erreicht. Zum Zeitpunkt II lagen die mittleren beta-Karotin-Serumkonzentrationen durchweg über den geforderten 1000 µg/l. Es gab keinen signifikanten Unterschied zwischen den Gruppen. Das Carofertin hatte also keinen Einfluß auf den beta-Karotin-Spiegel kurz nach der Abkalbung. Zum Zeitpunkt der ersten Besamung (meist ungefähr 3 116 Monate p.p.) waren die mittleren beta-Karotin-Serumkonzentrationen bei den meisten Tieren weit unter den von Lotthammer (1999) geforderten 2000 µg/l, aber über den von Smith und Bradford (1996) als Mangel deklarierten 700 µg/l. In der Karotingruppe war die mittlere beta-Karotin-Serumkonzentration zum Zeitpunkt III signifikant niedriger als zum Zeitpunkt II. In der Kontrollgruppe gab es keinen weiteren Abfall der Werte. Dass Carofertin führte also im Allgemeinen zu keiner Erhöhung des beta-Karotin-Spiegels um den Besamungszeitpunkt herum. Bei der Auswertung der Kühe, die zum Zeitpunkt I einen beta-Karotin-Gehalt von unter 2000 µg/l aufwiesen (also jenen Tieren, die beta-Karotin aus individuellen Gründen schlechter verwerten konnten), lag die mittlere beta-Karotin-Konzentration der Karotingruppe mit 1128,23 ± 427,90 µg/l zum Zeitpunkt III (Besamung) signifikant höher als bei der Kontrollgruppe (853,66 ± 340,24 µg/l, s. Abb.6). Bei diesen Tieren hatte das Carofertin also offensichtlich einen positiven Effekt auf den beta-KarotinSerumspiegel. 5.5 Puerperalkontrolle (4-6 Wochen p.p., Langzeitstudie) Der Beginn des Zyklus wurde zwischen der Karotingruppe und der Kontrollgruppe verglichen. Als Beweis für einen stattgefundenen Zyklus konnte nur das Vorhanden sein eines Gelbkörpers auf den Ovarien gewertet werden und so wurde der Anteil der Tiere, die zum Zeitpunkt der Puerperalkontrolle einen Gelbkörper hatten, zwischen den beiden Versuchsgruppen verglichen. Bei der Auswertung aller Tiere hatten die Kontrolltiere signifikant häufiger einen Gelbkörper als die Karotintiere (Karotingruppe 49,46 %, Kontrollgruppe 64,04 %,Tab. 23). Außerdem wurden in unserer Untersuchung zum einen die Ovarialzysten ausgewertet, die in der Puerperalkontrolle festgestellt wurden; zum anderen erfolgte eine Analyse der Ovarialzysten in der Gesamtlaktation (s. Kap. 5.5.3). 12,90 % der Tiere der Karotingruppe hatten zum Zeitpunkt der Puerperalkontrolle Ovarialzysten (Tab. 24). In der Kontrollgruppe lag der Anteil bei 14,61 %. Die Gruppenunterschiede waren nicht signifikant. Während der Puerperalkontrolle wurden die Tiere ebenfalls auf das 117 Vorliegen einer Endometritis untersucht, da Carofertin laut Hersteller über eine Verbesserung der Uterusinvolution und eine Stimulation der Endometrium-Drüsen das Endometritisrisiko senken soll. In der Karotingruppe erkrankten 30,11 % der Tiere an einer Endometritis und in der Kontrollgruppe 22,47 % (Tab. 25). Der Unterschied in der Endometritisinzidenz war nicht signifikant. Beta-Karotin war sowohl durch seine Wirkung als Provitamin A (Zerobin 1987) als auch durch seine Vitamin-A-unabhängige Funktion als Radikalfänger (Rapoport et al. 1998) für das reibungslose Ablaufen des Zyklus am Ovar wichtig. Failing et al. (1998) und Fischer (1996) bewirkten über den Zusatz von beta-Karotin zum Futter eine Verkürzung der Rast- und Güstzeiten bei ihren Versuchstieren. Ein weiterer Maßstab für eine zufriedenstellende Ovarfunktion ist die Inzidenz von Ovarialzysten. Inaba et al. (1986b) hatten bei Tieren mit Ovarialzysten geringere beta-Karotin-Blutspiegel festgestellt und Lotthammer et al. (1976) fanden bei ihren karotinfrei ernährten Versuchstieren einen um einen Tag verzögerte Ovulation. Marcek et al. (1985) stellten dagegen keine positive Beziehung zwischen der Gabe von beta-Karotin bei einer ausreichend versorgten Herde und dem Auftreten von Ovarialzysten fest. Über den Zusammenhang zwischen Endometritisinzidenz und beta-Karotin war in der mir zugänglichen Literatur nichts Konkretes zu finden. Allerdings stellten Markusfeld et al. (1997) fest, dass eine mangelhafte, peripartale Energieversorgung zu einer verzögerten Uterusinvolution post partum führt. Ebenso hatte ein Energiemangel in Verbindung mit hoher Eiweißversorgung (hohe Ammoniak- und Harnstoffkonzentrationen im Blut) eine Häufung von stiller Brunst und eitrigen Genitalkatarrhen (Lotthammer 1985b) zur Folge. Ein Kaliumüberschuß in der Ration bewirkte ebenfalls ein vermehrtes Auftreten von peripartalen Erkrankungen (Chandler 1997, Horst et al. 1997). In der vorliegenden Untersuchung hatten die Kontrolltiere zur Zeit der Puerperalkontrolle signifikant häufiger einen laufenden Zyklus als die Karotingruppe (Karotingruppe 49,46 %, Kontrollgruppe 64,04 %,Tab. 23). Dieser Unterschied war allerdings nicht durch die Gabe von Carofertin bedingt, sondern zeigt deutlich, dass in das Fruchtbarkeitsgeschehen viele Faktoren eingreifen und auch viele Faktoren zusammenspielen (De Kruif u. Mijten 1992). Ein Einfluß der Carofertin-Gabe auf 118 die Inzidenz von Ovarialzysten war ebenfalls nicht festzustellen. Da der Anteil an Tieren mit Ovarialzysten zum Zeitpunkt der Puerperalkontrolle bei den Versuchstieren bei unter 15 % lag, ist aber von keinem gehäuften Auftreten von Ovarialzysten in unserer Versuchsherde auszugehen. Dieses Ergebnis stützt also die Untersuchung von Marcek et al. (1985), da die Versuchstiere im allgemeinen auch ausreichend mit beta-Karotin versorgt waren. Der Unterschied in der Endometritisinzidenz der Gruppen war nicht signifikant. Wünschenswert ist eine Endometritisinzidenz von unter 15 % im Bezug auf die Gesamtherde (De Kruif et al. 1998). Mit 30,11 % in der Karotingruppe bzw. 22,47 % in der Kontrollgruppe war der Anteil von Tieren mit Endometritis in der Versuchsherde deutlich zu hoch. Auf diesen Aspekt wird bei der Besprechung der Endometritisinzidenz in der Gesamtlaktation noch eingegangen. An dieser Stelle wird aber deutlich, dass die Kalium-, Energieund Eiweißversorgung der Versuchstiere wahrscheinlich einen größeren Einfluß auf das Auftreten von Endometritis hat als die beta-Karotin-Versorgung. 5.6 Krankheiten und Abgänge 5.6.1 Peripartale Erkrankungen: Milchfieber und Nachgeburtsverhaltung In dieser Untersuchung waren weder bei Milchfieber (17,17 % der Karotingruppe, 20,79 % Kontrollgruppe) noch bei der Nachgeburtsverhaltung (11,11 % der Karotingruppe, 15,84 % der Kontrollgruppe) signifikante Gruppenunterschiede in der Häufigkeit dieser Krankheiten vorhanden (Tab.26). Bei 2 Kühen der Karotingruppe und 7 Kühen der Kontrollgruppe traten dabei beide Erkrankungen zusammen auf. Sowohl Inaba et al. (1986a) als auch Spindler (1980) stellten bei Tieren mit Nachgeburtsverhaltung niedrigere beta-Karotin-Serumkonzentrationen fest. Die Unterschiede waren aber nur schwach signifikant. In der vorliegenden Untersuchung lagen im Bezug auf den Blutserumspiegel des beta-Karotins keine signifikanten Unterschiede zwischen den Tieren, die eine Nachgeburtsverhaltung hatten, und den 119 Tieren, die nicht an Nachgeburtsverhaltung erkrankten, vor. Deshalb können die Ergebnisse von Inaba et al. (1986a) und Spindler (1980) nicht bestätigt werden. In einer gut geführten Herde sollte der Prozentsatz der Tiere mit Nachgeburtsverhaltung bei unter 15 % im Bezug auf die Gesamtherde liegen (De Kruif et al. 1998). Mit 11,11 % bei der Karotingruppe und 15,84 % bei der Kontrollgruppe war der Anteil an Nachgeburtverhaltung bei den Versuchstieren insgesamt gesehen noch im Normalbereich. Carofertin soll laut Hersteller die Uterusinvolution beschleunigen und so das Risiko einer Nachgeburtsverhaltung senken. Ob die Uterusinvolution durch das Carofertin beschleunigt wurde, konnte anhand der vorliegenden Ergebnisse nicht geklärt werden. Ein positiver Einfluß auf die Häufigkeit von Nachgeburtsverhaltung zeigte sich jedenfalls nicht statistisch absicherbar, obwohl die absolute Zahl in der Karotingruppe etwas niedriger lag als in der Kontrollgruppe. 5.6.2 Endometritis in der Gesamtlaktation Das Auftreten von Endometritiden bzw. pathologischem Scheidenausfluß wurde über die gesamte Laktation verfolgt und dokumentiert. In dieser Auswertung sind also auch alle Endometritisfälle enthalten, die während der Puerperalkontrolle diagnostiziert wurden. Rezidive wurden als ein Fall gewertet. In der Karotingruppe hatten 28,00 % der Tiere während der Gesamtlaktation eine Endometritis. In der Kontrollgruppe lag der Prozentsatz bei 26,73 % (Tab. 27). Die Differenz war auch in diesem Fall nicht signifikant. Pro 100 Kuhtage traten dabei in beiden Gruppen jeweils 10 Endometritisfälle auf. Der Normalwert von < 15 % Endometritiserkrankungen in der Gesamtherde wurde bei weitem überschritten und diese Tatsache hat die Fruchtbarkeitsleistung der Versuchstiere sicherlich erheblich negativ beeinflusst. Über einen Zusammenhang zwischen Endometritis und beta-Karotin war in der mir zugänglichen Literatur, wie bereits erwähnt, nichts zu finden. Die Firma Alvetra führt in ihren Produktinformationen an, dass die antioxidative Wirkung des beta-Karotins für die beschleunigte Uterusinvolution und somit für eine Verringerung des 120 Endometritisrisikos verantwortlich sei. In dieser Untersuchung führte die Injektion des Carofertins jedenfalls nicht zu einem reduzierten Auftreten der Endometritiden bei den Versuchstieren. Es fand keine weitere Abklärung der gehäuft auftretenden Endometritiden statt. Im Allgemeinen werden Infektionen während der Abkalbung (mangelhafte Hygiene bei der Geburtshilfe), Nachgeburtsverhaltung, mangelhafte Hygiene bei der Besamung und Besamungen während des Diöstrus für bestandsweise gehäuftes Auftreten von Endometritiden verantwortlich gemacht (De Kruif et al. 1998). Zusätzlich wurde der Einfluß des beta-Karotin-Spiegels auf die Endometritisinzidenz untersucht. Zu diesem Zweck wurden die beta-Karotin Spiegel der erkrankten Tiere mit denen nicht-erkrankter Tiere verglichen. Bei der Betrachtung der beta-KarotinSpiegel getrennt nach den drei Entnahmezeitpunkten lagen keine signifikanten Unterschiede vor. Zusätzlich wurde die Endometritisinzidenz zwischen den Quartilen der beta-Karotin-Serumkonzentration verglichen. Es traten in keinem Fall signifikante Unterschiede auf, so dass in der vorliegenden Untersuchung kein Zusammenhang zwischen dem beta-Karotin-Spiegel und der Endometritisinzidenz festgestellt werden konnte. 5.6.3 Ovarialzysten in der Gesamtlaktation Diese Auswertung enthält alle Fälle von Ovarialzysten in der Laktation der Versuchstiere. Rezidive wurden als ein Fall gewertet. In der Karotingruppe traten bei 14,00 % der Tiere Ovarialzysten auf und in der Kontrollgruppe bei 11,88 % (Tab. 28). Der Gruppenunterschied war nicht signifikant. Der Normalwert von unter 15 % wurde somit nicht überschritten. Das Risiko pro 100 Kuhtage an einer Ovarialzyste zu erkranken lag in beiden Gruppen bei 4 %. Grundsätzlich kann gesagt werden, dass Ovarialzysten bei Fütterungsmängeln (Energiemangel, Kaliumüberversorgung, beta-Karotinmangel), einer hohen Milchleistung und auch bei genetisch prädisponierten Tieren gehäuft auftreten (Aurich et al. 1996). Untersuchungen von Ascarelli et al. (1985) und Lotthammer 121 (1979) hatten bei beta-Karotin-arm ernährten Versuchstieren vermehrt Ovarialzysten gefunden. In dieser Untersuchung waren aber alle Versuchtiere weitgehend ausreichend mit beta-Karotin über das Futter versorgt und die Ovarialzysteninzidenz lag im Normalbereich. Das Carofertin führte zu keiner weiteren Verringerung der Ovarialzystenfälle. Insofern können wir auch in der Gesamtlaktation die Ergebnisse von Marcek et al. (1985) bestätigen. Wie schon bei der Endometritis wurde auch der Einfluß des beta-Karotin-Spiegels auf die Inzidenz von Ovarialzysten untersucht. Inaba et al. (1986b) hatten festgestellt, dass der beta-Karotin-Gehalt im Blut bei Tieren mit Ovarialzysten niedriger lag als bei Tieren ohne Ovarialzysten. Beim Vergleich der beta-Karotin Spiegel der erkrankten Tiere mit denen nicht-erkrankter Tiere lagen in dieser Untersuchung keine signifikanten Unterschiede vor. Bei der Gegenüberstellung der Quartile gab es zum Zeitpunkt III im Quartil III (947-1176 µg/l beta-Karotin) signifikant weniger Karotintiere (0 %) mit Ovarialzysten als Kontrolltiere (19,05 %, P<0,05). Falls dies auf die Gabe von Carofertin zurückzuführen ist, so gibt es jedenfalls keinen Zusammenhang mit der Höhe des beta-Karotin-Spiegels, da es zu keinem signifikanten Unterschied beim Vergleich der Quartile gegeneinander kam. 5.6.4 Abgänge In der Karotingruppe lag die Remontierungsrate bei 38,61 % und in der Kontrollgruppe bei 44,00 %. Die Abgänge wegen Unfruchtbarkeit betrugen in der Karotingruppe 19,00 % und 10,89 % in der Kontrollgruppe (Tab. 31). Es war kein signifikanter Unterschied vorhanden, so dass das Carofertin auch im Bezug auf die Abgänge wegen Unfruchtbarkeit keinen positiven Einfluss hatte. Aus ökonomischer Sicht macht es Sinn, eine unrentabel gewordene Kuh schnell durch eine leistungsfähigere Färse zu ersetzen (Fetrow 1994) und so sind Remontierungsraten in dieser Größenordnung heute an der Tagesordnung. Im allgemeinen sollen die Abänge wegen Unfruchtbarkeit bezogen auf die mittlere 122 Zuchtherde bei unter 7 % liegen (De Kruif et al. 1998). Es muss allerdings berücksichtigt werden, dass in der vorliegenden Untersuchung alle Tiere, die 200 Tage post partum noch nicht tragend waren, als Abgang wegen Unfruchtbarkeit in diese Statistik miteingingen und die tatsächliche Anzahl an Abgängen etwas geringer war, da einige Tiere später doch noch tragend geworden sind. Dies geschah allerdings auf Wunsch der Betreiber und war rein wirtschaftlich betrachtet nicht sinnvoll. Bei den Abgängen wegen geringer Leistung, Euterkrankheit und Gliedmaßenerkrankungen gab es ebenfalls keine signifikanten Gruppenunterschiede. 5.7 Fruchtbarkeitskennzahlen 5.7.1 Auswertung nach Gruppen Einer der Hauptaspekte dieser Untersuchung war die Analyse der Fruchtbarkeitskennzahlen, da man anhand dieser Daten einen guten Überblick über die Herdenfruchtbarkeit bekommt. Zusammenfassend kann gesagt werden, dass die Fruchtbarkeitskennzahlen bis auf eine Ausnahme keinen signifikanten Gruppenunterschied aufwiesen. Diese Ausnahme war, dass die Färsen der Karotingruppe signifikant später (über 60 Tage p.p.) in Brunst gesehen wurden als die Färsen der Kontrollgruppe. In der Auswertung der Gesamtgruppe war der Anteil an Tieren, bei denen die erste Brunst über 60 Tagen p.p. gesehen wurde, mit 58,82 % in der Karotingruppe deutlich höher als in der Kontrollgruppe (46,51 %). Der Unterschied war allerdings nicht signifikant. Akodor et al. (1986), Bindas et al. (1984), Bremel et al. (1982), Ducker et al. (1984), Wang et al (1987) und Jukola et al. (1996a) konnten keine positiven Effekte einer beta-Karotin-Supplementierung auf die Herdenfruchtbarkeit feststellen. Positive Effekte wurden bisher nur bei Tieren mit einem nachgewiesenen beta-KarotinMangel aufgrund von karotinarmer Nahrung erzielt (Lothammer 1979, Ascarelli et al. 1985). Sowohl Block und Farmer (1987) als auch De Kruif und Mijten (1992) 123 betonten, dass ein beta-Karotin-Mangel nur unter extremen Bedingungen als alleinige Ursache von Fruchtbarkeitsstörungen in Frage kommt. Ferguson (1996) und Kessler et al. (1991) machten bereits deutlich, dass ein gutes Herdenmanagement und eine bedarfsorientierte Fütterung einen viel größeren Effekt auf die Fruchtbarkeit hat als ein Vitamin- oder Spurenelementmangel. Stolla et al. (1987) und Ascarelli et al. (1985) berichteten dagegen über eine gesteigerte Brunstintensität durch betaKarotin. Es konnten keine positiven Effekte der Carofertin-Gabe auf die Fruchtbarkeitskennzahlen festgestellt werden. Im Bezug auf die erst nach 60 Tagen p.p. in Brunst gesehenen Färsen schnitt die Karotingruppe sogar schlechter ab als die Kontrollgruppe, so dass die vorliegenden Ergebnisse auch gegen eine durch das Carofertin gesteigerte Brunstintensität sprechen. Man darf allerdings nicht vergessen, dass die Prozentsätze der Versuchstiere mit Endometritis (Karotingruppe 28,00 %, Kontrollgruppe 26,73 %) unabhängig davon, ob die Tiere Carofertin bekommen hatten oder nicht, deutlich zu hoch lagen. Dieser Sachverhalt dürfte eine Erklärung für die hohe Abgangsrate wegen Unfruchtbarkeit, die schlechte Konzeptionsrate (Karotingruppe 31,91 %, Kontrollgruppe 38,73 %) bzw. den schlechten Erstbesamungserfolg (Karotingruppe 37,65 %, Kontrollgruppe 41,86 %) sein. Damit lagen die Werte der Versuchstiere deutlich unter den geforderten 55 % erfolgreichen Erstbesamungen. Der Trächtigkeitsindex war mit ca. 1,7 ebenfalls etwas zu hoch. Wünschenswert ist ein Trächtigkeitsindex von < 1,6. Das Carofertin führte in keinem Fall zu einer signifikanten Verbesserung der Fruchtbarkeitskennzahlen (P>0,05). Insgesamt gesehen liegen die Rastzeit, Güstzeit, Verzögerungszeit und die erwartete Zwischenkalbezeit in den erwünschten Bereichen (Tab. 45), obwohl die Tiere erst spät in Brunst gesehen wurden und schlecht aufnahmen. Dies machte deutlich, dass man die Fruchtbarkeit einer Herde nie an einzelnen Kennzahlen beurteilen sollte, sondern immer alle zusammen auswerten sollte. Diese Untersuchung bestätigte somit die Vermutung, dass mit einer Supplementierung von beta-Karotin in einer Herde, die über das Futter ausreichend mit beta-Karotin versorgt wurde, keine Verbesserung der Fruchtbarkeit zu erreichen ist. 124 5.7.2 Einfluß des beta-Karotin-Spiegels auf die Fruchtbarkeitskennzahlen Wie bereits erwähnt, waren die beta-Karotin-Serumkonzentrationen der Versuchstiere unabhängig davon, zu welcher Gruppe sie gehörten, zu den drei Zeitpunkten sehr unterschiedlich. Um herauszufinden, ob die Tiere mit niedrigeren Blutserumkonzentrationen eine schlechtere Fruchtbarkeit aufwiesen, wurden die Versuchstiere nach den Quartilen ihrer beta-Karotin-Serumkonzentrationen unterteilt (Tab. 35). Die Höhe des beta-Karotin-Spiegels hatte keinen Einfluß auf die Inzidenz von Nachgeburtsverhaltung, Endometritis und Ovarialzysten. Der Erstbesamungserfolg war bis auf einen Fall nicht signifikant von der Höhe der betaKarotinwerte beeinflusst. Die Tiere, die zum Zeitpunkt III Werte zwischen 751 und 947 µg/l aufwiesen (Quartil II), hatten einen signifikant niedrigeren Erstbesamungserfolg als die Tiere der drei übrigen Quartile. Ebenso lag der Trächtigkeitsindex des Quartils II (Zeitpunkt III) signifikant niedriger als im Quartil III (P<0,05). Bei der Auswertung der Rastzeit, Güstzeit, Verzögerungszeit und Zwischenkalbezeit gab es zum Zeitunkt I keinen Quartilunterschied. Zum Zeitpunkt II lag die Verzögerungszeit im Quartil IV (16,29 Tage) signifikant niedriger als im Quartil I (34,54 Tage). Zum Zeitpunkt III war die Rastzeit in den Quartilen II und IV signifikant niedriger als im Quartil I. Außerdem war die Zwischenkalbezeit in Karotinund Kontrollgruppe im Quartil III signifikant niedriger als im Quartil II. Die lineare Regressionsanalyse ergab, dass die Höhe des beta-Karotin-Spiegels zum Zeitpunkt II negativ korreliert war mit der Güstzeit, der Verzögerungszeit und der Zwischenkalbezeit. Die Rastzeit war zum Zeitpunkt III negativ mit der beta-KarotinSerumkonzentration korreliert (Tab. 54). Allerdings waren diese Korrelationen nur sehr gering und außerdem lag eine breite Streuung der Werte vor. Zusätzlich ist zu bedenken, dass die Tiere mit niedrigen beta-Karotin-Spiegeln zum Teil bessere Fruchtbarkeitsleistungen aufwiesen als die Tiere mit hohen beta-Karotin-Spiegeln. Signifikante Unterschiede waren allerdings nicht vorhanden. Zusammenfassend ist zu sagen, dass die Tiere mit niedrigeren beta-Karotin-Serumkonzentrationen in 125 Teilbereichen eine schlechtere Fruchtbarkeit aufwiesen als die Tiere mit höheren beta-Karotin-Spiegeln. Die vorliegenden Ergebnisse lassen aber durch die breite Streuung der Werte unabhängig vom beta-Karotin-Spiegel darauf schließen, dass beta-Karotin nur ein Faktor von vielen ist, die die Fruchtbarkeit einer Herde beeinflussen. Warum die beta-Karotin-Werte bei einigen Tieren viel niedriger lagen als bei anderen Versuchstieren und wie diese individuellen Schwankungen auf Herdenbasis vermieden werden können, konnte nicht geklärt werden. 5.8 Progesteronbestimmung In dieser Untersuchung gab es keine signifikanten Gruppenunterschiede in der während der Lutealphase produzierten Progesteronmenge. Die Progesteronmenge pro Tag und der höchste Progesteronwert der untersuchten Lutealphasen waren ebenfalls nicht signifikant unterschiedlich zwischen den Gruppen (Tab. 60). Inaba et al. (1987) hatten durch den Zusatz von beta-Karotin zu in vitro kultivierten Granulosazellen eine vermehrte Progesteronproduktion ausgelöst. Auch Rapoport et al. (1998) und Arikan et al. (2000a, b) stellten fest, dass beta-Karotin in vitro die Progesteronproduktion der Mikrosomenfraktion der Gelbkörperzellen steigert. In vivo allerdings konnten nur Dembinski et al. (1996) eine Erhöhung der Progesteronkonzentration im Blutserum bewirken. Bei Bonsembiante et al. (1980), Ducker et al. (1981) und Folman et al. (1983) fand keine Steigerung der Progesteronkonzentration im Blutserum statt, wenn die Tiere beta-KarotinSerumkonzentrationen über 1500 µg/l hatten. Carofertin soll durch Stabilisierung der Corpora lutea zu einer Anhebung des Progesteronspiegels führen. Das war in dieser Untersuchung nicht der Fall. Die vorliegende Untersuchung bestätigt hingegen die Ergebnisse von Bonsembiante et al. (1980), Ducker et al. (1981) und Folman et al. (1983). Das beta-Karotin in Form von Carofertin beeinflusste die Menge des produzierten Progesterons nicht. 126 5.9 Kosten pro Trächtigkeit Bei der Gegenüberstellung der Kosten pro Trächtigkeit (Tab. 62 und 63) schneiden die Karotintiere (241,78 €) deutlich schlechter ab als die Kontrolltiere (171,40 €). Insgesamt gesehen lagen die Kosten pro Trächtigkeit bei den Versuchstieren etwas höher als bei Drillich (1999). Klindworth (2000) kam zu ähnlichen Ergebnissen wie in dieser Untersuchung. Tischer (1998) und Surholt (2001) errechneten etwas höhere Beträge. Dies zeigt deutlich, dass die Kosten pro Trächtigkeit stark von betriebsspezifischen Faktoren abhängen (Tischer 1998) und somit keine generellen Empfehlungen über die Höhe der Kosten pro Trächtigkeit gegeben werden können. Die Kosten pro Trächtigkeit lagen in dieser Untersuchung bei den Karotintieren höher als in der Kontrollgruppe, weil die Carofertin-Behandlung erhebliche Kosten verursacht hat (Kosten pro Trächtigkeit ohne Carofertin in der Karotingruppe: 208,87 €). Hinzu kommt, dass in der Karotingruppe 7 Tiere mehr wegen Unfruchtbarkeit abgegangen sind als in der Kontrollgruppe. Die Kosten für Fruchtbarkeitsbehandlungen, Güsttage über 85 Tage p.p., Spermakosten und die Kosten durch Trächtigkeitsuntersuchungen sind in beiden Gruppen annähernd gleich. In der Karotingruppe waren die größten Kostenfaktoren die Abgänge bzw. die Kosten für remontierte Tiere (8078,80 €) gefolgt von den Kosten für Güsttage über 85 Tage p.p. (4075,00 €). In der Kontrollgruppe waren die Remontierungskosten (5102,40 €) ebenfalls an erster Stelle und die Güsttage über 85 Tage p.p. (4682,50 €) an zweiter. Dies bestätigt die Ergebnisse von Jakob und Distel (1997) und Tenhagen et al. (1998), wonach die Tierarztkosten und Medikamentenkosten nur einen geringen Anteil an den Gesamtkosten haben und die Kosten für Güsttage über 85 Tage p.p. und Remontierung den Hauptteil ausmachen. In der vorliegenden Untersuchung wurde somit keine Verringerung der Kosten pro Trächtigkeit durch die Carofertin-Gabe erreicht. 127 5.10 Schlussfolgerungen Die Gabe von Carofertin in der vom Hersteller vorgeschlagenen Dosierung beeinflusste weder die beta-Karotin-Serumkonzentration noch die Fruchtbarkeit der Versuchstiere positiv. Dies kann zum einen an der relativ geringen beta-KarotinMenge pro Injektion im Vergleich zu den über das Futter aufgenommenen Mengen gelegen haben und zum anderen daran, dass die Versuchstiere im allgemeinen ausreichend über das Futter mit beta-Karotin versorgt waren. Es besteht allerdings in der Literatur große Uneinigkeit im Bezug auf die Referenzwerte für die beta-KarotinBlutserumkonzentrationen, so dass es schwierig ist, den Versorgungsstatus einer Herde anhand der beta-Karotin-Serumkonzentrationen zu beurteilen. Auffällig war die große Streuung der beta-Karotin-Serumkonzentrationen zu allen drei Zeitpunkten. Weitere Untersuchungen sollten klären, ob es wirklich ein individuelles Resorptions- und Mobilisationsvermögen für beta-Karotin bei Kühen gibt und ob es sich beeinflussen lässt, da die Tiere mit niedrigen beta-Karotin- Serumkonzentrationen tatsächlich zum Teil eine schlechtere Fruchtbarkeit aufwiesen als die Tiere mit höheren Blutspiegeln. Eine Anhebung des Progesteronspiegels durch das Carofertin wurde nicht erreicht. Die Kosten pro Trächtigkeit lagen in der Karotingruppe deutlich über denen der Kontrollgruppe. Die größten Kostenfaktoren waren aber gruppenunabhängig die Remontierungskosten und die Kosten durch Güsttage über 85 Tagen p.p. Aufgrund der Ergebnisse dieser Untersuchung kann der Einsatz von Carofertin zur Verbesserung der Fruchtbarkeit weder zu prophylaktischen noch zu therapeutischen Zwecken empfohlen werden. 128 6 Zusammenfassung Natascha Gossen (2003): Einfluss einer Supplementierung mit beta-Karotin in Form einer Injektionslösung (Carofertin) auf die Fruchtbarkeitsleistung von Milchkühen Ziel dieser Arbeit war, den Einfluß einer Supplementierung von beta-Karotin in Form einer Injektionslösung (Carofertin) auf die Fruchtbarkeitsleistung von Milchkühen zu untersuchen. Dazu wurde zunächst eine Verlaufsstudie an 20 Tieren, die am ersten Versuchstag 20 ml Carofertin (n=10) bzw. 0,9 % NaCl-Lösung (n=10) i.m. injiziert bekamen, durchgeführt. Alle Tiere wurden unmittelbar vor der Injektion und dann einmal täglich 14 Tage lang beprobt. Die Verlaufstudie sollte zeigen, ob Carofertin in der vom Hersteller empfohlenen Dosierung die beta-Karotin-Serumkonzentration erhöht. An der Langzeitstudie nahmen von Januar 2001 bis September 2002 201 Tiere teil, die drei i.m.-Injektionen mit je 20 ml Carofertin (n=100) bzw. 0,9 % NaClLösung (n=101) (3-4 Wo. a.p., 1-2 Wo. p.p., 6-8 Wo. p.p.) erhielten. Um die betaKarotin-Serumkonzentration zu bestimmen, wurde den Versuchstieren unmittelbar vor der ersten und zweiten Injektion und kurz nach der ersten Besamung Blut entnommen. Bei allen Versuchstieren wurden die auftretenden Krankheiten, die Behandlungen, die Besamungen, die Abgänge und die Abgangsursachen dokumentiert und eine Puerperalkontrolle durchgeführt. Weiterhin wurden die Fruchtbarkeitskennzahlen aus den erhobenen Daten ermittelt. Von einer Stichprobe von Kühen wurde ab dem 40. Tag p.p. die Progesteronserumkonzentration zweimal wöchentlich bestimmt, bis die Tiere eine Lutealphase durchlaufen hatten. Der Versuch endete mit dem Abgang oder der Trockenstellung der Tiere. 1. In der Verlaufsstudie kam es nur beim Vergleich des Ausgangswertes der Karotingruppe mit dem Entnahmezeitpunkt IV zu einer signifikant höheren betaKarotin-Serumkonzentration. Insgesamt waren die Mittelwerte der beta-KarotinSerumkonzentration der Karotingruppe für 12 Tage höher als die der Kontrollgruppe. Die Unterschiede waren aber nicht signifikant. Innerhalb und zwischen den Tieren bestanden große Schwankungen der beta-Karotin-Konzentration. 2. Es lagen keine signifikanten Gruppenunterschiede in der Höhe der mittleren betaKarotin-Serumkonzentration zu den drei Entnahmezeitpunkten vor. 129 3. Innerhalb der Gruppen lagen die beta-Karotin-Werte in beiden Gruppen zum Zeitpunkt I signifikant höher als zu den Zeitpunkten II und III. Die Werte des Zeitpunkts II waren in der Karotingruppe bei der Auswertung aller Tiere signifikant höher als zum Zeitpunkt III. In der Kontrollgruppe gab es keine signifikanten Unterschiede. Somit konnte der starke Abfall der beta-Karotin-Werte zur Zeit der Abkalbung bestätigt werden. Die Carofertin-Gabe hatte auf dieses Verhalten keinen Einfluß. 4. Bei der Puerperalkontrolle hatten die Kontrolltiere signifikant häufiger einen intakten Zyklus als die Karotintiere. Ein positiver Einfluß des Carofertins auf das Einsetzen der Ovartätigkeit kann folglich nicht bestätigt werden. 5. Weder bei der Inzidenz von Krankheiten noch bei den Abgangsursachen traten signifikante Gruppenunterschiede auf. 6. Bei der Auswertung der Fruchtbarkeitskennzahlen wurden keine signifikanten Gruppenunterschiede festgestellt. Es war somit kein positiver Einfluß des Carofertins auf die Herdenfruchtbarkeit vorhanden. 7. Die Höhe des beta-Karotin-Spiegels hatte keinen Einfluß auf die Krankheitsinzidenz. Die Auswertung der Fruchtbarkeitskennzahlen führte zu widersprüchlichen Ergebnissen. Somit besteht sicherlich ein gewisser Zusammenhang zwischen beta-Karotin und der Fruchtbarkeit von Milchkühen. Die Herdenfruchtbarkeit wird aber durch viele Faktoren beeinflusst und beta-Karotin spielt dabei wahrscheinlich nur eine untergeordnete Rolle. 8. In dieser Untersuchung hatte Carofertin auf die Progesteronkonzentration im Blut bzw. auf die produzierte Progesteronmenge keinen Einfluß. 9. Bei der Gegenüberstellung der Kosten pro Trächtigkeit schnitten die Karotintiere (241,78 €) deutlich schlechter ab als die Kontrolltiere (171,40 €). Hauptursache waren die Mehrkosten durch das Carofertin (8,71 € pro Injektion). Die größten Kostenfaktoren waren in beiden Gruppen die Remontierung aufgrund von Unfruchtbarkeit und die Kosten durch Güsttage über 85 Tagen p.p.. 10. Aufgrund der Ergebnisse dieser Untersuchung kann der Einsatz von Carofertin bei Milchkühen weder zu prophylaktischen noch zu therapeutischen Zwecken empfohlen werden. 130 7 Summary Natascha Gossen (2003): Effect of parenteral supplementation with ß-carotin (Carofertin) on the reproductive performance in dairy cows It was the aim of this study to investigate the effect of a supplementation with ßcarotin administered systemically (Carofertin) on the reproductive performance in dairy cows. In a short-term study, cows were injected with 20 ml Carofertin (n=10) or 0.9 % NaCl-solution (n=10) intramuscularly on the first day of the trial. Serum concentration of ß-carotin was determined before the treatment and then daily for 14 days in order to show a positive effect of Carofertin in a dosage recommended by the manufacturer on serum concentrations of ß-carotin. In a long-term study which lasted from January 2001 until September 2002, 100 cows received three injections of 20 ml Carofertin 3-4 weeks ante partum, 1-2 weeks post partum and 6-8 weeks post partum. Control cows (n=101) were treated with 20 ml 0.9 % NaCl-solution. Serum concentrations of ß-carotin were determined before the first and the second ß-carotin injection and at the time of first insemination. For all animals, diseases, treatments, inseminations, cullings and culling reasons were recorded, and a clinical examination was performed during the puerperal period (4-6 weeks post partum). Furthermore, fertility parameters were calculated. From a subgroup of cows, blood samples were taken twice weekly starting around Day 40 post partum, and serum concentrations of progesterone were measured throughout the luteal phase. The trial period lasted until the day of drying off or until a cow was culled. 1. In the short-term study, mean serum concentration of ß-carotin in the ß-carotin group was only higher on Day 4 following injection compared with the preinjection value (P<0.05). Overall, mean concentrations of ß-carotin were higher in the ßcarotin group than in the controll group for 12 days. However, the differences between groups were not statistically significantly different (P>0.05). Large variations of ß-carotin concentrations were observed within animals and between animals. 131 2. In the long-term study, mean serum concentrations of ß-carotin at the three sampling points did not differ between groups (P>0.05). 3. In both groups, ß-carotin concentrations were higher at the first sampling than at the second and third sampling (P<0.05). In the ß-carotin group, concentration of ßcarotin was also higher at the second sampling compared with the third sampling (P<0.05). This difference was not observed in the controll group. Hence, the marked decrease in the ß-carotin concentration at the time of calving observed by other authors could be confirmed. Supplementation with Carofertin did not have an influence on this decrease. 4. The clinical examination during the puerperal phase revealed that cyclicity had started statistically significantly more often in control cows than in the cows receiving Carofertin, i.e. a positive effect of Carofertin could not be verified. 5. There were no group differences with regard to the incidence of diseases, culling rate and culling reasons (P>0.05). 6. Fertility parameters did not differ between groups, i.e. a positive effect of Carofertin on herd fertility could not be detected. 7. Whereas the ß-carotin concentration in serum did not affect incidence of diseases, the analysis of the fertility parameters revealed contradictory results indicating that there might be a slight interrelationship between ß-carotin and reproductive performance in dairy cows. However, as the herd fertility is influenced by many factors, ß-carotin apparently seems to be of minor importance. 8. Progesterone concentration in serum and amount of progesterone produced during the luteal phase were not influenced by Carofertin. 9. Cost per pregnancy were higher in the Carofertin group (241.78 €) than in the control group (171.40 €). The main reason for the higher cost were the cost of Carofertin (8.71 € per injection). In both groups, replacement cost for cows culled due to infertility and cost of days open beyond 85 days post partum had the highest economical impact on cost per pregnancy. 10. These results suggest that the use of Carofertin in dairy cows for therapy or prophylaxis might not be recommendable. 132 8 Literatur Ahlswede, L. u. K. H. Lotthammer (1978): Untersuchungen über eine spezifische, Vitamin-A-unabhängige Wirkung des ßCarotins auf die Fertilität des Rindes, 5. Mitteilung: Organuntersuchungen (Ovarien, Corpora lutea, Leber, Fettgewebe, Uterussekret, Nebennieren) - Gewichts- und Gehaltsbestimmungen. Dtsch. Tierärztl. Wschr. 85, 7-12 Akordor, F. Y., J. B. Stone, J. S. Walton, K. E. Leslie u. J. G. Buchanan Smith (1986): Reproductive performance of lactating Holstein cows fed supplemental betacarotene. J. Dairy Sci. 69, 2173-2178 Allison, R. D. u. R. A. Laven (2000): Effect of vitamin E supplementation on the health and fertility of dairy cows. Vet. Rec. 147, 703-708 Anke, M. (1971): Der Einfluß von Mangan auf die Fruchtbarkeit des Wiederkäuers. Wiss. Z. Karl-Marx-Univ. Leipzig, Math.-naturwiss. R. 20, 420-478 Anwandter Quentin, C. G. (1974): Progesteron- und ß-Carotingehalt des bovinen Corpus luteum während der verschiedenen Phasen des Sexualzyklus in Abhängigkeit von der Jahreszeit. Hannover, Tierärztl. Hochsch., Diss. Arechiga, C. F., C. R. Staples, L. R. McDowell u. P. J. Hansen (1998a): Effects of timed insemination and supplemental beta-carotene on reproduction and milk yield of dairy cows under heat stress. J. Dairy Sci. 81, 390-402 Arechiga, C. F., S. Vazquez-Flores, O. Ortiz, J. Hernandez-Ceron, A. Porras, L. R. McDowell u. P. J. Hansen (1998b): Effect of injection of beta-carotene or vitamin E and selenium on fertility of lactating dairy cows. Theriogenology 50, 65-76 Arikan, S. u. R. G. Rodway (2000a): Effects of high density lipoprotein containing high or low beta-carotene concentrations on progesterone production and beta-carotene uptake and depletion by bovine luteal cells. Anim. Reprod. Sci. 62, 253-263 133 Arikan, S. u. R. G. Rodway (2000b): Effect of cyclodextrin-encapsulated beta-carotene on progesterone production by bovine luteal cells. Anim. Reprod. Sci. 64, 149-160 Ascarelli, I., Z. Edelman, M. Rosenberg u. Y. Folman (1985): Effect of dietary carotene on fertility of high-yielding dairy cows. Anim. Prod. 40, 195-207 Ashes, J. R., R. W. Burley, J. B. Davenport u. G. S. Sidhu (1982): Effects of dietary supplements of protected lipids on the concentration and transport of beta-carotene and cholesterol in bovine blood and milk: unusual chromatographic behaviour of the high-density lipoprotein with high levels of beta-carotene. J. Dairy Res. 49, 39-49 Ashes, J. R., R. W. Burley, G. S. Sidhu u. R. W. Sleigh (1984): Effect of particle size and lipid composition of bovine blood high density lipoprotein on its function as a carrier of beta-carotene. Biochim. Biophys. Acta 797, 171-177 Ashworth, C. J. (1994): Nutritional factors related to embryonic mortality in the domestic species. In: M. T. Zavy u. R. D. Geisert (Hrsg.): Embryonic mortality in domestic species. CRS Press, Boca Raton, 179-210 Aslan, S., J. Handler u. K. Arbeiter (1998): Frühgravidität und embryonale bzw. frühfetale Mortalität bei der Kuh Gelbkörperdynamik, Progesteron-, Vitamin E-, Vitamin-B12-, ß-Carotin- und Folsäurekonzentrationen im peripheren Blut. Wien. Tierärztl. Mschr. 85, 141- 147 Aurich, J. E., D. Ahlers u. P. Andresen (1996): Ovarialzysten. In: E. Grunert (Hrsg.): Buiatrik. Verlag M. & H. Schaper Alfeld, Band I, 235-238 Berchtold, M. (1982): Fruchtbarkeitsüberwachung auf Herdenbasis. In: E. Grunert u. M. H. Berchtold (Hrsg.): Fertilitätsstörungen beim weiblichen Rind. 1. Aufl., Verlag Paul Parey, Berlin, 441-449 Bindas, E. M., F. C. Gwazdauskas, R. J. Aiello, J. H. Herbein, M. L. McGilliard u. C. E. Polan (1984): Reproductive and metabolic characteristics of dairy cattle supplemented with betacarotene. J. Dairy Sci. 67, 1249-1255 134 Bindas, E. M., F. C. Gwazdauskas, M. L. McGilliard u. C. E. Polan (1984): Progesterone responses to human chorionic gonadotropin in dairy cattle supplemented with beta-carotene. J. Dairy Sci. 67, 2978-2985 Bittner, W. (1986): In vitro-und in vivo-Untersuchungen über den Einfluß des ß-Carotins auf die ovarielle Steroidhormonsynthese. Gießen, Univ., Vet. med. Fak., Diss. Block, E. u. B. Farmer (1987): The status of beta-carotene and vitamin A in Quebec dairy herds: factors affecting their status in cows and their effects on reproductive performance. Can. J. Anim. Sci. 67, 775-788 Boehnke, H.-J., J. Ehlers u. K. H. Lotthammer (1993): Blutuntersuchungen zur Spurenelementversorgung beim Rind. Milchpraxis 31, 16-19 Bonsembiante, M., G. Bittante u. I. Andrighetto (1980): Effect of beta-carotene on the fertility of cows given diets supplemented with vitamin A. Zootecnica e Nutrizione Animale 6, 47-58 Bremel, D. H., R. W. Hemken, G. Heersche, L. A. Edgerton u. D. Olds (1982): Effects of ß-carotene on metabolic and reproductive parameters in lactating dairy cows. J. Dairy Sci. 65, Suppl.1, 178 (Abst.) Britt, J. H. (1975): Early postpartum breeding in dairy cows. A review. J. Dairy Sci. 58, 266-271 Britt, J. H. (1985): Enhanced reproduction and its economic implications. J. Dairy Sci. 68, 1585-1592 Brüggemann, J. u. K.-H. Niesar (1955): Der Nachweis eines den Vitamin A-Gehalt der Milch beeinflussenden Faktors im Eutergewebe des Rindes. Milchwiss. 10, 223-225 Burgstaller, G., P. Matzke, W. Hollwich, J. Kogel, O. Kreutzberg u. H. Zucker (1980): Untersuchungen zur Versorgung von Milchkühen mit ß-Carotin und Vitamin A, 1. Mitteilung: Einfluß auf den Blutspiegel. Zbl. Vet. Med. A 27, 485-494 135 Butler, W. R. u. R. D. Smith (1989): Interrelationships between energy balance and post partum reproductive function in dairy cattle. J. Dairy Sci. 72, 767-783 Cetinkaya, N. u. H. Ozcan (1991): Investigation of seasonal variations in cow serum retinol and beta-carotene by high performance liquid chromatographic method. Comp. Biochem. Physiol. A 100, 1003-1008 Chandler, P. (1997): Milk fever may be caused by potassium and sodium, not calcium. Feedstuffs 69, 15-23 Chawla, R. u. H. Kaur (2001): Isocratic HPLC method for simultaneus determination of ß-carotene, retinol and alpha-tocopherol in feeds and blood plasma. Ind. J. Dairy Sci. 54, 84-90 Chew, B. P., D. M. Holpuch u. J. V. O'Fallon (1984): Vitamin A and beta-carotene in bovine and porcine plasma, liver, corpora lutea, and follicular fluid. J. Dairy Sci. 67, 1316-1322 Chew, B. P. (1993): Role of carotenoids in the immune response. J. Dairy Sci. 76, 2804-2811 Cooke, B. C. u. N. Comben (1978): A study of the relationships between beta-carotene and fertility problems in dairy cows. British Society of Animal Production. Paper summaries. Winter Meeting 4, 45 (Abst.) Daniel, L. R., B. P. Chew, T. S. Tanaka u. L. W. Tjoelker (1991a): ß-Carotene and vitamin A effects on bovine phagocyte function in vitro during the peripartum period. J. Dairy Sci. 74, 124-131 Daniel, L. R., B. P. Chew, T. S. Tanaka u. L. W. Tjoelker (1991b): In vitro effects of beta-carotene and vitamin A on peripartum bovine peripheral blood mononuclear cell proliferation. J. Dairy Sci. 74, 911-915 De Kruif, A. u. P. Mijten (1992): Das Verhältnis zwischen Fütterung und Fruchtbarkeit beim Rind. Berl. Münch. Tierärztl. Wschr. 105, 271-279 136 De Kruif, A., R. Mansfeld u. M. Hoedemaker (1998): Tierärztliche Bestandsbetreuung beim Milchrind. Ferdinand Enke Verlag, Stuttgart Dembinski, Z. u. M. Bronicki (1994): Progesterone P-4 level in blood and the values of selected fertility indexes in cows fed various doses of carotenes. Bull. Vet. Inst. Pulawy 38, 115-118 Dembinski, Z. u. M. Bronicki (1996): Influence of different amounts of beta-carotene in food on progesterone level in blood of dairy cows. Zeszyty Naukowe Akademii Rolniczej w Szczecinie 33, 15-18 Dijkhuizen, A. A., J. Stelwagen u. J. A. Renkema (1984): Economic aspects of reproductive failure in dairy cattle, 1. Financial loss at farm level. Prev. Vet. Med. 3, 251-263 Dohoo, J. R., S. W. Martin u. A. H. Meek (1984): Disease, production and culling in Ontario dairy cattle. Can. Vet. J. 25, 106-110 Drillich, M. (1999): Vergleich des strategischen Einsatzes von Prostaglandin F2alpha mit konventionellen Methoden des Fruchtbarkeitsmanagements in zwei Milchviehherden. Berlin, Univ., Tierärztl. Fak., Diss. Ducker, M. J., R. A. Haggett, W. J. Fisher, S. V. Morant, G. A. Bloomfield, M. I. Gurr u. J. D. Edwards Webb (1981): An investigation of the effect of level of beta -carotene on the fertility of dairy cows and heifers. National Institute for Research in Dairying, UK Report 1981, 42 (Abst.) Ducker, M. J., N. H. Yarrow, G. A. Bloomfield u. J. D. Edwards Webb (1984): The effect of beta-carotene on the fertility of dairy heifers receiving maize silage. Animal Prod. 39, 9-16 Edwards, R. G. (1974): Follicular fluid. Reprod. Fert. 37, 189-219 Eisele, W. (1987): ß-Carotin und Vitamin A in Blut und Milch des Rindes nach parenteraler und oraler Applikation von ß-Carotin. München, Univ., Tierärztl. Fak., Diss. 137 Esselmont, R. J. u. E. J. Peeler (1993): The scope for raising margins in dairy herds by improving fertility and health. Br. Vet. J. 149, 537-547 Fadle, W. S. A. (1966): Vergleichende Untersuchungen über den Vitamin A- und ß-Carotin-Gehalt des Blutplasmas von Muttertieren und Kälbern sowie des Kolostrums. Hannover, Tierärztl. Hochsch., Diss. Failing, K., K. Fischer u. B. Hoffmann (1998): Erhebungen zum Fruchtbarkeitsstatus von Milchkühen, Teil 2: Multifaktorielle Beziehungen zu Blutparametern sowie dem allgemeinen und gynäkologischen Status. Tierärztl. Umschau 53, 467-472 Ferguson, J. D. u. W. Chalupa (1989): Impact of protein nutrition on reproduction in dairy cows. J. Dairy Sci. 72, 746-766 Ferguson, J. D. (1996): Diet, production and reproduction in dairy cows. Anim. Feed Sci. Techn. 59, 172-184 Fernandez, S. C., P. Budowski, I. Ascarelli, H. Neumark u. A. Bondi (1976): Pre-intestinal stability of beta-carotene in ruminants. Int. J. Vitam. Nutr. Res. 46, 439-445 Fetrow, J. u. T. Blanchard (1987): Economic impact of use of prostaglandin in induce estrus in dairy cows. J. Am. Vet. Assoc. 190, 163-168 Fetrow, J. (1994): Culling and genetic improvement programs for dairy herds. In: O. M. Radostits, K. E. Leslie u. J. Fetrow (Hrsg.): Herd Health - Food animal production medicine, 2. Aufl. W.B. Saunders Company, Philadelphia, 159-182 Fischer, K. (1996): Erhebungen zum Fruchtbarkeitsstatus von Milchkühen, Beziehungen zu Blutparametern sowie dem allgemeinen und gynäkologischen Status-Eine explorative Datenanalyse. Gießen, Univ., Vet. med. Fak., Diss. Fischer, K.-F. (1999): Taschenbuch der Technischen Formeln. Fachbuchverlag, Leibzig 138 Flachowsky, G. (1999): Zur Vitaminversorgung von Milchkühen. Übers. Tierernährung 27, 29-64 Folman, Y., I. Ascarelli, Z. Herz, M. Rosenberg, M. Davidson u. A. Halevi (1979): Fertility of dairy heifers given a commercial diet free of beta-carotene. Br. J. Nutr. 41, 353-359 Folman, Y., M. Rosenberg, I. Ascarelli, M. Kaim u. Z. Herz (1983): The effect of dietary and climatic factors on fertility, and on plasma progesterone and oestradiol-17 beta levels in dairy cows. J. Steroid Biochem. Mol. Biol. 19, 863-868 Folman, Y., I. Ascarelli, D. Kraus u. H. Barash (1987): Adverse effect of beta-carotene in diet on fertility of dairy cows. J. Dairy Sci. 70, 357-366 Folman, Y., M. Kaim, Z. Herz u. M. Rosenberg (1990): Comparison of methods for the synchronisation of estrous cycles in dairy cows. 2. Effects of progesterone and parity on conception. J. Dairy Sci. 73, 2817-2825 Friesecke, H. (1978): Beta-Carotin und die Fruchtbarkeit des Rindes. Hoffman-LaRoche AG, Grenzach-Wyhlen Gawienowski, A. M., M. Stacewicz-Sapuncakis u. R. Longley (1974): Biosynthesis of retinal in bovine corpus luteum. J. Lipid Res. 15, 375-379 Graves-Hoagland, R. L., T. A. Hoagland u. C. O. Woody (1988): Effect of beta-carotene and vitamin A on progesterone production by bovine luteal cells. J. Dairy Sci. 71, 1058-1062 Graves-Hoagland, R. L., T. A. Hoagland u. C. O. Woody (1989): Relationship of plasma beta-carotene and vitamin A to luteal function in postpartum cattle. J. Dairy Sci. 72, 1854-1858 Greenberg, L. G., F. Bristol, B. D. Murphy u. B. Laarveld (1986): Beta-carotene does not influence fertility in beef heifers. Theriogenology 26, 491-508 139 Grummer, R. R. u. J. H. Clark (1980): Effekt of nitrogen solubility on milk yield, milk composition and ruminal degradation of protein. J. Dairy Sci. 63, Suppl., 139 (Abst.) Grunert, E. (1990): Weiblicher Geschlechtsapparat. In: Rosenberger, G. (Hrsg.): Die klinische Untersuchung des Rindes. 3. Aufl., Verlag Paul Parey, Berlin, 472-549 Gugger, E. T. u. J. W. Erdman, Jr. (1996): Intracellular beta-carotene transport in bovine liver and intestine is not mediated by cytosolic proteins. J. Nutr. 126, 1470-1474 Günther (1980): Vitamin-A- und ß-Carotinversorgung in der Rinderfütterung. Bayr. Landwirtsch. Jahrb. 57, 238-251 Haliloglu, S., N. Baspinar, B. Serpek, H. Erdem u. Z. Bulut (2002): Vitamin A and ß-carotene levels in plasma, corpus luteum and follikular fluid of cyclic and pregnant cattle. Reprod. Dom. Anim. 37, 96-99 Hamburger, R. (1973): Progesteronbestimmung in der Milch. München, Univ., Tierärztl. Fak., Diss. Hanck, A. B. u. C. C. Kuenzle (1991): Vitamin A. Verlag Paul Parey, Berlin Haraszti, J., G. Huszenicza, L. Molnar u. A. Blaskovits (1984): Effect of periparturient lipid mobilization on serum total carotene and vitamin A concentrations in cattle. Acta Vet. Hung. 32, 193-203 Hartmann, P. E., J. G. Harris u. A. K. Lascelles (1966): The effect of oil-feeding and starvation on the composition and output of lipid in the thoracic duct lymph in the lactating cow. Aust. J. Biol. Sci. 19, 635-644 Hasselmann, L., H. Munchow, V. Manzke, W. Schneeweiss, F. Ahrens, R. Schubert, G. Flachowsky, R. Bitsch u. G. Jahreis (1999): Zum Einfluß von ß-Carotinzulagen auf den Zellgehalt der Rohmilch sowie auf die Fruchtbarkeitsleistung bei Milchkühen unter Praxisbedingungen. In: Symp. Vit. Zusatzst. Ernähr., 22.-23.9.1999, Jena (Thüringen), 134-139 140 Heinz, E. u. A. Herzog (1982): Erste Praxiserfahrungen mit dem Einsatz eines synthetischen ß-Carotins im Rahmen der zuchthygienisch-prophylaktischen Betreuung von Milchkuhherden. Berl. Münch. Tierärztl. Wschr. 95, 214-217 Hemken, R. W. u. D. H. Bremel (1982): Possible role of beta-carotene in improving fertility in daily cattle. J. Dairy Sci. 65, 1069-1073 Herdt, T. H. u. H. D. Stowe (1991): Fat-soluble vitamin nutrition for dairy cattle. Veterinary Clinics of North America, Food Animal Practice 7, 391-415 Hino, T., N. Andoh u. H. Ohgi (1993): Effects of beta-carotene and alpha-tocopherol on rumen bacteria in the utilization of long-chain fatty acids and cellulose. J. Dairy Sci. 76, 600-605 Holt, A. J., R. G. Rodway u. J. B. Findley (1995): Studies on ß-carotene in bovine corpus luteum. J. Reprod. Fertil. 15, 46 (Abst.) Höltershinken, M., K. Maiworm u. H. Scholz (1996): Mykotoxikose beim Rind - Probleme auch in Norddeutschland. Prakt. Tierarzt 77, 9-14 Horst, R. L., J. P. Goff, T. A. Reinhardt u. D. R. Buxton (1997): Strategies for preventing milk fever in dairy cattle. J. Dairy Sci. 80, 1269-1280 Hurley, W. L. u. R. M. Doane (1989): Recent developments in the roles of vitamins and minerals in reproduction. J. Dairy Sci. 72, 784-804 Inaba, T., A. Inoue, R. Shimizu, Y. Nakano u. J. Mori (1986a): Plasma concentrations of progesterone, estrogens, vitamin A and beta-carotene in cows retaining fetal membranes. Jap. J. Vet. Sci. 48, 505-508 Inaba, T., M. Mezan, R. Shimizu, Y. Nakano u. J. Mori (1986b): Plasma concentrations of beta-carotene and vitamin A in cows with ovarian cyst. Jap. J. Vet. Sci. 48, 1275-1278 Inaba, T., M. Mezan, R. Shimizu, T. Ono u. J. Mori (1987): Effects of beta-carotene and vitamin A on progesterone production by cultured bovine granulosa cells. Jap. J. Vet. Sci. 49, 1021-1025 141 Jackson, P. S. (1981): A note on a possible association between plasma beta-carotene levels and conception rate in a group of winter-housed dairy cattle. Anim. Prod. 32, 109-111 Jakob, H. u. O. Distl (1997): Tierarztkosten beim Milchvieh, 1. Mitteilung: Analyse von systematischen Variationsursachen. Züchtungskunde 69, 334-348 Jukola, E., J. Hakkarainen, H. Saloniemi u. S. Sankari (1996a): Blood selenium, vitamin E, vitamin A, and beta-carotene concentrations and udder health, fertility treatments, and fertility. J. Dairy Sci. 79, 838-845 Jukola, E., J. Hakkarainen, H. Saloniemi u. S. Sankari (1996b): Effect of selenium fertilization on selenium in feedstuffs and selenium, vitamin E, and beta-carotene concentrations in blood of cattle. J. Dairy Sci. 79, 831-837 Kappel, L. C., E. B. Ingraham, J. M. Morgan, J. M. Dixon, L. Zeringue, D. Wilson u. D. K. Babcock (1984): Selenium concentrations in feeds and effects of treating pregnant Holstein cows with selenium and vitamin E on blood selenium values and reproductive performance. Am. J. Vet. Res. 45, 691-694 Kessler, J., H. Friesecke u. P. Kunz (1991): Beta-Carotin: Versorgung der Milchkuh während der Winterfütterung. Landwirtschaft Schweiz 4, 203-206 Kirchgessner, M. (1987): Tierernährung: Leitfaden für Studium, Beratung und Praxis. 7. Aufl., DLG-Verlag, Frankfurt Kirsche, B., M. Schlenzig, W. I. Ochrimenko u. G. Flachowsky (1987): Influence of beta-carotene supplementation on carotene content of ovaries in heifers. Arch. Anim. Nutr. 37, 995-999 Klindworth, H.-P. (2000): Ovulationssynchronisation (OVSYNCH) in hochleistenden Milchvieherden. Hannover, Tierärztl. Hochsch., Diss. Kolb, E. (1994): The problem of a high content of vitamin A in the liver of calves, cattle, sheep and swine for the consumer. Amount of accumulation and mechanism of teratogenic effect (review article). Berl. Münch. Tierärztl. Wschr. 107, 342-347 142 Kolb, E. u. K. Elze (1995): Durch Energiemangel beim Rind ausgelöste Fortpflanzungsstörungen. Prakt. Tierarzt 76, 623-626 Kolb, E. u. J. Seehawer (1997): Die Bedeutung der Carotine und des Vitamin A für die Fortpflanzung bei Rindern, Pferden und Schweinen - eine Übersicht. Prakt. Tierarzt 78, 783-789 Kolb, E. (1998): Verwertung und Anwendung von Vitaminen bei Haustieren. Hoffmann-LaRoche AG, Grenzach-Wyhlen, 25-34 Kolb, E. u. J. Seehawer (1998): Die Entwicklung des Immunsystems und des Vitamingehalts beim Rinderfetus und beim Kalb sowie die Förderung der Aktivität durch Verabreichung von Vitaminen eine Übersicht. Tierärztl. Umschau 53, 723-730 Koopman, J. J., A. Wijbenga u. A. C. M. Bloem (1971): Enkele waarnemingen over het caroteen- en vitamine A-gehalte van levers bij rundvee. Tijdschr. Diergeneesk. 96, 1019-1027 Krieger, K. u. F. J. Schweigert (1993): Verhalten der fettlöslichen Vitamine A, E und des ß-Carotins während des Trockenstehens beim Rind. In: Symp. Vit. Zusatzst. Ernähr., 30.09.-01.10.1993, Jena (Thüringen), 235-238 Krinsky, N. I., D. G. Cornwell u. J. L. Oncley (1958): The transport of vitamin a and carotinoids in human plasma. Arch. Biochem. Biophys. 73, 233-246 Kurz, E., W. A. Rambeck u. H. Zucker (1984): Verteilung des ß-Carotins auf die Serumlipoproteine bei Milchkühen. Zbl. Vet. Med. A 31, 174-181 Larson, L. L., S. E. Wallen, F. G. Owen u. S. R. Lowry (1983): Relation of age, season, production, and health indices to iodine and beta-carotene concentrations in cow's milk. J. Dairy Sci. 66, 2557-2562 LeBlanc, S. J., T. F. Duffield, K. E. Leslie, K. G. Bateman, J. TenHag, J. S. Walton u. W. H. Johnson (2002): The effect of prepartum injection of vitamin E on health in transition dairy cows. J. Dairy Sci. 85, 1416-1426 143 Leidl, W., R. Porzig u. R. Stolla (1987): Untersuchungen über die Wirkungen einer ß-Carotinzufütterung auf die Fruchtbarkeit von Rindern, 1. Mitteilung: Blutkonzentrationen und diagnostische Möglichkeiten. Berl. Münch. Tierärztl. Wschr. 100, 1-6 Lotthammer, K. H., L. Ahlswede u. H. Meyer (1976): Untersuchungen über eine spezifische Vitamin-A-unabhängige Wirkung des ßCarotins auf die Fertilität des Rindes. 2. Mitteilung: Weitere klinische Befunde und Befruchtungsergebnisse. Dtsch. Tierärztl. Wschr. 83, 353-358 Lotthammer, K. H. u. L. Ahlswede (1977): Untersuchungen über eine spezifische, Vitamin-A-unabhängige Wirkung des ßCarotins auf die Fertilität des Rindes, 3. Mitteilung: Blutserumuntersuchungen (ßCarotin, Vitamin A, SGOT, Gesamtcholesterin, Glukose, anorganischer Phosphor). Dtsch. Tierärztl. Wschr. 84, 220-226 Lotthammer, K. H. (1978): Importance of beta -carotene for fertility of dairy cattle. Tierzüchter 30, 520-522 Lotthammer, K. H. (1979): Importance of beta-carotene for the fertility of dairy cattle. Feedstuffs 51, 37-38 Lotthammer, K. H. (1985a): Auswirkungen einer erhöhten Aufnahme einiger Mengen- und Spurenelemente sowie von Nitrat über wirtschaftseigene Futtermittel auf die Gesundheit und Fruchtbarkeit von Milchkühen. Dtsch. Tierärztl. Wschr. 92, 205-244 Lotthammer, K. H. (1985b): Ursachen und Maßnahmen beim primär nicht infektiösen Genitalkatarrh des Rindes. Prakt. Tierarzt 66, 79-84 Lotthammer, K. H. (1992): Anforderungen an den Tierarzt in der Rinderpraxis der Zukunft. Prakt. Tierarzt 62, 305-307 Lotthammer, K. H. (1999a): Klinisch-chemische Untersuchungen bei bestandsweise auftretenden Fruchtbarkeitsstörungen. In: E. Grunert u. A. De Kruif (Hrsg.): Fruchtbarkeitsstörungen beim weiblichen Rind. Parey Buchverlag, Berlin, 62-66 144 Lotthammer, K. H. (1999b): Umweltbedingte Fruchtbarkeitsstörungen. In: E. Grunert u. A. De Kruif (Hrsg.): Fertilitätsstörungen beim weiblichen Rind. Parey Buchverlag, Berlin, 320-321 Marcek, J. M., L. H. Appell, C. C. Hoffman, P. T. Moredick u. L. V. Swanson (1985): Effect of supplemental beta-carotene on incidence and responsiveness of ovarian cysts to hormone treatment. J. Dairy Sci. 68, 71-77 Markusfeld, O., N. Galon u. E. Ezra (1997): Body condition score, health, yield and fertility in dairy cows. Vet. Rec. 141, 67-72 Marschang (1985): Carotinversorgung beim Rind - Einfluß auf das Karankheitsgeschehen. Tierzüchter 37, 267-269 Martens, H., G. Breves u. S. Leonhard-Marek (2000): Funktionen der Vormägen. In: W. v. Engelhardt u. G. H. Breves (Hrsg.): Physiologie der Haustiere. Enke Verlag, Stuttgart, 359-360 Meyer, H., L. Ahlswede u. K. H. Lotthammer (1975): Untersuchungen über eine spezifische Vitamin-A-unabhängige Wirkung des ßCarotins auf die Fertilität des Rindes. 1. Mitteilung: Versuchsanstellung, Körperentwicklung und Eierstocksfunktion. Dtsch. Tierärztl. Wschr. 82, 444-449 Michal, J. J., L. R. Heirman, T. S. Wong, B. P. Chew, M. Frigg u. L. Volker (1994): Modulatory effects of dietary beta-carotene on blood and mammary leukocyte function in periparturient dairy cows. J. Dairy Sci. 77, 1408-1421 Miller, J. K., E. Brzezinska-Slebodzinska u. F. C. Madsen (1993): Oxidative stress, antioxidants, and animal function. J. Dairy Sci. 76, 2812-2823 Miller, K. W., N. A. Lorr u. C. S. Yang (1984): Simultaneus determination of plasma retinol, alpha-tocopherol, lyopene, alphacarotene and beta-carotene by high-performance liquid chromatography. Anal. Biochem. 138, 340-345 Morrow, D. A., D. Hillman, A. W. Dade u. H. Kitchen (1979): Clinical investigation of dairy herd with fat cow syndrome. J. Am. Vet. Med. Assoc. 174, 161-165 145 Morton, J. M. u. M. R. McGowan (2002): Herd-, cow-,lactation- and insemination-level factors affecting reproductive performance in dairy herds. In: M. Kaske, H. Scholz u. M. Höltershinken (Hrsg.): Recent Developments and Perspectives in Bovine Medicine - Keynote Lectures, XXII World Buiatrics Congress, 18.-23. August 2002, Hannover, Germany. Hildesheimer Druck- und Verlags-GmbH, Hildesheim, 324-333 Nehring, K. u. M. Hoffmann (1967): Die Carotinverdaulichkeit beim Wiederkäuer. Arch. Tierernähr. 17, 263-279 Noller, C. H., W. E. Castro, W. E. Wheeler, D. H. Hill u. N. J. Moeller (1977): Effect of phosphorus supplementation on growth rate, blood minerals and conception rate of dairy heifers. J. Dairy Sci. 60, 1932-1940 NRC (1989): Nutrient Requirements of Dairy Cattle. National Academy of Science, Washington, DC NRC (2001): Nutrient Requirements of Dairy Cattle. National Academy of Science, Washington, DC O'Fallon, J. V. u. B. P. Chew (1984): The subcellular distribution of beta-carotene in bovine corpus luteum. Proc. Soc. Exp. Biol. Med. 177, 406-411 Oldham, E. R., R. J. Eberhart u. L. D. Muller (1991): Effects of supplemental vitamin A or beta-carotene during the dry period and early lactation on udder health. J. Dairy Sci. 74, 3775-3781 Olson, J. A. u. O. Hayaishi (1965): The enzymatic cleavage of ß-carotene into vitamin A by soluble enzymes of rat liver and intestine. Proc. Nat. Acad. Sci. 54, 1364-1370 Özpinar, H., E. Cekgul, E. Eggenberger u. H. S. Senel (1988): Zusammenhang zwischen Ovarialzysten und den Konzentrationen von ß-Carotin, Vitamin E und Vitamin A im Serum von Milchkühen. Schweiz. Arch. Tierheilk. 130, 263-273 Özpinar, H., A. Özpinar, T. Bilal, I. Türkalp u. A. Firat (1995): Pharmacokinetics of intramuscular administered beta-carotene and its effects on reproduction in sheep. Wien. Tierärztl. Mschr. 82, 229-231 146 Palmquist, D. L. (1976): A kinetic concept of lipid transport in ruminants - a review. J. Dairy Sci. 59, 355-363 Patton, S., J. J. Kelly u. T. W. Keenan (1980): Carotene in bovine milk fat globules: observations on origin and high content in tissue mitochondria. Lipids 15, 33-38 Perkins, B. L. (1985): Production, reproduction, health and liver function following overconditioning in dairy cattle. Diss. Abstr. Int. B 46, 10 Pethes, G., E. Horvath, M. Kulcsar, G. Huszenicza, G. Somorjai, B. Varga u. J. Haraszti (1985a): In vitro progesterone production of corpus luteum cells of cows fed low and high levels of beta-carotene. Zbl. Vet. Med. A 32, 289-296 Pethes, G., P. Rudas, G. Huszenicza u. G. Somorjai (1985b): Conversion of thyroxine to triiodothyronine in liver biopsy samples of beta-carotene and vitamin-A supplemented dairy cows. Zbl. Vet. Med. A 32, 512-517 Rakes, A. H., M. P. Owens, J. H. Britt u. L. W. Whitlow (1985): Effects of adding beta-carotene to rations of lactating cows consuming different forages. J. Dairy Sci. 68, 1732-1737 Rammel, C. G., B. Cunliffe u. A. J. Kienoom (1983): Determination of alpha-tocopherol in biological specimens by HPLC. J. Liquid Chromatogr. 6, 1123-1130 Rapoport, R., D. Sklan, D. Wolfenson, A. Shaham-Albalancy u. I. Hanukoglu (1998): Antioxidant capacity is correlated with steroidogenic status of the corpus luteum during the bovine estrous cycle. Biochim. Biophys. Acta 1380, 133-140 Reksen, O. u. E. Ropstad (2002): Influence of dietary energy and protein on reproductive performance in dairy cattle. Norsk-Veterinaertidsskrift 114, 21-25 Rodgers, R. J., T. C. Lavranos, H. F. Rodgers, F. M. Young u. C. A. Vella (1995): The physiology of the ovary: maturation of ovarian granulosa cells and a novel role for antioxidants in the corpus luteum. J. Steroid Biochem. Mol. Biol. 53, 241-246 147 Ronning, M., E. R. Berousek, J. R. Griffiths u. W. D. Gallop (1959): Carotene requirement of dairy cattle. Oklahoma Agric. Exp. Tech. Bull. Oklahoma St. Univ., Stillwater 76, 30 Rosenberger, G. (1978): Krankheiten des Rindes. Verlag Paul Parey, Berlin Schams, D., B. Hoffmann, K. H. Lotthammer u. L. Ahlswede (1977): Untersuchungen über eine spezifische, Vitamin-A-unabhängige Wirkung des ßCarotins auf die Fertilität des Rindes. 4. Mitteilung: Auswirkungen auf hormonale Parameter während des Zyklus. Dtsch. Tierärztl. Wschr. 84, 307-310 Schlenzig, M., G. Flachowsky, W. I. Ochrimenko, G. H. Richter u. D. Geinitz (1988): Einfluß unterschiedlicher ß-Karotin und Vitamin-A-Versorgung auf Wachstum und Fruchtbarkeit weiblicher Jungrinder. Tierzucht 42, 178-181 Scholz, H. (1989): Selen-/Vitamin-E-Mangel; Realität auch in unserer Rinderpraxis. Prakt. Tierarzt 70, Coll. Vet. XIX, 22-27 Schweigert, F. J. (1986): ß-Carotin-Stoffwechsel des Rindes: Verteilung auf die Serumlipoproteine, Transfer in die Milch und in die Folliklflüssigkeit sowie Funktion im Follikel. München, Univ., Tierärztl. Fak., Diss. Schweigert, F. J. (1988): ß-Carotinstoffwechsel des Rindes und seine Bedeutung für die Fruchtbarkeit. Tierernährung 16, 223-246 Schweigert, F. J. (1995): Recent findings on the metabolism and function of beta-carotene and vitamin A. Kraftfutter 6, 271-272 Seekles, L. u. J. Claessens (1967): Kupfer und Fertilität beim Rind. Schweiz. Arch. Tierheilk. 109, 76-84 Seitaridis, K. (1963): Untersuchungen über den Vitamin-A- und ß-Carotingehalt des Blutplasmas in Rinderherden mit Störungen der Fruchtbarkeit unter besonderer Berücksichtigung der Carotinversorgung. Hannover, Tierärztl. Hochsch., Diss. 148 Shalgi, R., P. Kraicer, A. Rimon u. M. Pinto (1973): Proteins of human follicular fluid: The blood-follicle barrier. Fertil. Steril. 24, 429-434 Sklan, D. (1983): Carotene Cleavage Activity in the corpus luteum of cattle. Int. J. Vitam. Nutr. Res. 53, 23-26 Smith, B. P. (1996): Vitamin A Deficiency. In: B. P. Smith (Hrsg.): Large animal internal medicine: diseases of horses, cattle, sheep and goats. 2. Aufl., Mosby, St. Louis, 1064-1067 Spindler, H.-G. (1980): Untersuchungen über Ursachen der Nachgeburtsverhaltung beim Rind unter besonderer Berücksichtigung des ß-Carotin- und Vitamin-A-Gehaltes im Blutserum sowie der Zellstruktur an den Plazentomen. Hannover, Tierärztl. Hochschule, Diss. Stacewicz-Sapuncakis, M., H. H. Chang Wang u. A. M. Gawienowski (1975): Biosynthesis of retinol in bovine corpus luteum tissue. Biochim. Biophys. Acta 380, 264-269 Stampfer, R. (1982): Untersuchungen zur Beziehung zwischen ß-Carotin im Blut und in der Milch bei Kühen. München, Univ., Tierärztl. Fak., Diss. Staples, C. R., W. W. Thatcher u. J. H. Clark (1990): Relationship between ovarian activity and energy status during the early post partum period of high producing dairy cows. J. Dairy Sci. 73, 938-947 Steel, L. u. S. Torrie (1980): Principles and procedures of statistics: A biometrical approach. 2. Auflage. McGraw-Hill Book Company, New York Stiewe, H. (1984): Fruchtbarkeit kann gefüttert werden. Tierzüchter 36, 17-19 Stolla, R., R. Porzig u. W. Leidl (1987): Effects of feeding beta-carotene on the fertility of cattle. 2. Effects on fertility. Berl. Münch. Tierärztl. Wschr. 100, 90-95 149 Stott, A. W. u. M. A. DeLorenzo (1988): Factors affecting profitability of Jersey and Holstein lactations. J. Dairy Sci. 71, 275-283 Surholt, R. (2001): Vergleich dreier Fruchtbarkeitsprogramme zur Verbesserung der Herdenfruchtbarkeit in Milchviehbetrieben am Beispiel einer Milchviehanlage mit Fruchtbarkeitsstörungen. Berlin, Univ., Tierärztl. Fak., Diss. Swanson, L. V. (1989): Discussion: Interactions of nutrition and reproduction. J. Dairy Sci. 72, 805-814 Talavera, F. u. B. P. Chew (1988): Comparative role of retinol, retinoic acid and beta-carotene on progesterone secretion by pig corpus luteum in vitro. J. Reprod. Fertil. 82, 611-615 Tekpetey, F. R., W. M. Palmer u. J. R. Ingalls (1987): Reproductive performance of prepubertal dairy heifers on low or high ß-carotene diets. Can. J. Anim. Sci. 67, 477-489 Tekpetey, F. R., W. M. Palmer u. J. R. Ingalls (1987): Seasonal variation in serum ß-carotene and vitamin A and their association with postpartum reproductive performance of holstein cows. Can. J. Anim. Sci. 67, 491-500 Tenhagen, B.-A. u. W. Heuwieser (1997): Wirtschaftliche Einbußen durch verlängerte Güstzeiten - Einfluß der Laktationspersistenz. In: DVG-Tagung "Fortpflanzung und ihre Störungen", 27.06.1997, Gießen, 14-22 Deutsche Veterinärmedizinische Gesellschaft, Gießen Tenhagen, B.-A., M. Tischer, W. Heuwieser u. H. Blum (1998): Influence of puerperal endometritis on the economy of reproduction in dairy herds. In: Proc. 10th Middle-European Buiatrics Congress, 21.-23.05.1998, Siofok, Hungary, 136-139 Tischer, M. (1998): Vergleich von intrauterinen Arzneimittelapplikationen mit einem strategischen Prostaglandinprogramm zur Behandlung von chronischen Endometritiden in einer Milchviehherde. Berlin, Univ., Tierärztl. Fak., Diss. 150 Tjoelker, L. W., B. P. Chew, T. S. Tanaka u. L. R. Daniel (1988): Bovine vitamin A and beta-carotene intake and lactational status. 2. Responsiveness of mitogen-stimulated peripheral blood lymphocytes to vitamin A and beta-carotene challenge in vitro. J. Dairy Sci. 71, 3120-3127 Tjoelker, L. W., B. P. Chew, T. S. Tanaka u. L. R. Daniel (1990): Effect of dietary vitamin A and beta-carotene on polymorphonuclear leukocyte and lymphocyte function in dairy cows during the early dry period. J. Dairy Sci. 73, 1017-1022 Ullrey, D. E. (1972): Biological availability of fatsoluble vitamins: vitamin A and carotene. J. Anim. Sci. 35, 648 Wang, J. Y., L. L. Larson u. F. G. Owen (1982): Effect of beta-carotene supplementation on reproductive performance of dairy heifers. Theriogenology 18, 461-473 Wang, J. Y., C. B. Hafi, F. G. Owen u. L. L. Larson (1987): Effect of beta-carotene supplementation on periparturient health and reproduction of Holstein cows. Anim. Reprod. Sci. 15, 1-2 Wang, J. Y., C. B. Hafi u. L. L. Larson (1988a): Effect of supplemental beta-carotene on luteinizing hormone released in response to gonadotropin-releasing hormone challenge in ovariectomized Holstein cows. J. Dairy Sci. 71, 498-504 Warner, R. L., G. E. Mitchell, C. O. Little u. N. E. Alderson (1970): Preintestinal disappearance of vitamin A in steers fed different levels of corn. Int. J. Vitam. Nutr. Res. 40, 585-588 Weaver, G. A., H. J. Kurtz, J. C. Behrens, T. S. Robinson, B. E. Seguin, F. Y. Bates u. C. J. Mirocha (1986): Effect of zearalenone on the fertility of virgin dairy heifers. Am. J. Vet. Res. 47, 1395-1397 Weigelt, B., R. Weigelt, T. Barth, S. Bach, K. Eulenberger u. J. Schulz (1988): Studies into embryonic mortality in a dairy herd. Mh. Vet. Med. 43, 157-160 Weiss, W. P., J. S. Hogan, K. L. Smith u. S. N. Williams (1994): Effect of dietary fat and vitamin E on alpha-tocopherol and beta-carotene in blood of peripartum cows. J. Dairy Sci. 77, 1422-1429 151 Weiss, W. P. (1998): Requirements of fat-soluble vitamins for dairy cows: a review. J. Dairy Sci. 81, 2493-2501 Whitaker, D. A. (1999): Trace elements - the real role in dairy cow fertility? Cattle Practice 7, 239-241 Young, F. M., W. B. Luderer u. R. J. Rodgers (1995): The antioxidant beta-carotene prevents covalent cross-linking between cholesterol side-chain cleavage cytochrome P450 and its electron donor, adrenodoxin, in bovine luteal cells. Mol. Cell. Endocrinol. 109, 113-118 Zeeck, A., S. Eick, B. Krone u. K. Schröder (1992): Chemie für Mediziner. Urban & Schwarzenberg, München Zerobin, K. (1987): Physiologie der Fortpflanzung. In: A. Scheunert u. A. Trautmann (Hrsg.): Lehrbuch der Veterinärphysiologie. Verlag Paul Parey, Berlin, 215-221 Ziegler, R. G. (1989): A review of epidemiologic evidence that carotinoids reduce the risk of cancer. J. Nutr. 119, 116-122 Zucker, H., O. Kreutzberg, W. Hollwich, D. Matzke, J. Kogel u. G. Burgstaller (1980): Supply of dairy cows with beta-carotene and vitamin A. II. Effect on concentration in milk and clinico-gynaecological and hormone analytical findings. Zbl. Vet. Med. A 7, 525-533 152 Anhang 153 Tabelle 56: Beta-Karotin-Konzentrationen aller Tiere der Verlaufsstudie (Einheit µg/l, Entnahmezeitpunkte I-XVIII, Nr. = Nummer, 17 16 15 14 13 12 11 10 9 8 7 6 5 4 3 1 2 1 2 2 2 1 1 2 1 2 1 1 2 1 2 1 2 1 2 1 1 1 2 1 1 2 1 1 1 2 1 1 1 1 1 2 2 2 1189 2082 1903 1560 2243 1541 1421 1727 2409 2688 2276 1137 2170 1263 1963 1522 2357 3189 3161 3134 I 1781 2973 1520 1398 2505 1858 1233 1912 2957 2763 2262 2268 1739 2233 1833 2390 3642 1456 5610 2542 II 1772 2295 1898 1574 2106 1176 1923 2048 2592 3479 2841 2249 3573 2202 1284 2936 4038 3234 3914 3880 III 1710 2267 2154 1429 2295 1107 1790 2407 2604 3011 2557 2732 2665 1943 2284 3013 3564 3118 3808 2796 IV 1327 1514 2080 1221 1943 1160 1574 2305 2430 2757 1786 2003 2367 2133 2406 2840 3315 3490 4097 2877 V 1971 2235 1377 1369 1860 1295 1757 2248 2732 2396 2442 2445 3010 2307 1930 2785 3959 4238 4264 3823 VI 2081 2301 1788 1316 2084 1016 1965 1235 1590 2101 2458 2609 3052 1840 2302 2107 2226 2529 4266 1160 VII 2544 2199 2088 1314 1992 365 1464 1310 1623 2701 2586 2248 3190 2632 2429 6271 6831 1842 5228 1867 VIII 1710 1964 2240 1142 1920 1296 2827 1355 1436 2694 4045 1813 2141 1895 2408 3034 2123 1839 1898 1596 IX 2159 1746 1937 1074 1970 1285 1785 1723 1590 2797 2542 2093 2363 2214 5598 3086 1538 2733 3308 1929 X 2208 2360 1284 1195 1872 1337 2304 1610 1731 1569 1424 2309 3673 2160 2328 2002 3361 2300 4346 3455 XI 2127 1862 1302 1187 1855 1174 1562 1396 2012 2045 1410 1892 2217 1363 2802 1727 1478 2353 2713 2234 XII 1729 1663 1827 1058 1668 1206 1474 1050 1447 1663 1515 1891 1909 1724 2986 2451 2510 1846 1298 1743 XIII 1937 2004 2289 1300 1479 1234 1242 1101 1510 1775 1577 2040 2252 1623 1749 2395 2732 1748 2120 1822 XIV 1694 1788 1218 2076 1171 1659 999 1305 1944 1861 2542 1937 1922 2157 1927 2314 1599 1932 1419 XV 1975 1142 957 1574 1077 1241 1246 1378 1643 2112 2088 1939 2316 1802 2989 1625 2104 1565 XVI 1253 1617 1711 1514 1251 1908 1206 1166 1351 1419 813 1236 2247 2531 1903 2001 1729 XVII 1611 1373 943 1711 1225 1114 783 1610 1433 1070 1949 2258 1759 1725 XVIII Gruppe: 1 = Karotin, 2 = NaCl, Tiere: 1 = Kühe, 2 = Färsen) (I-XIV = alle 24 h; XIV-XVIII = alle 48 h) 18 1 1 Gruppe Tiere 19 2 Nr. 20 153 154 Tabelle 57: Erstbesamungserfolg zum Zeitpunkt I (n = Anzahl der Versuchstiere, C = Karotingruppe, NaCl = Kontrollgruppe) Quartil Gruppe n Erstbesamungserfolg in % (n) I C NaCl 27 20 40,74 (11) 50,00 (10) II C NaCl 22 21 36,36 (8) 28,57 (6) III C NaCl 18 21 27,78 (5) 42,86 (9) IV C NaCl 18 23 44,44 (8) 47,83 (11) Tabelle 58: Erstbesamungserfolg zum Zeitpunkt II (n = Anzahl der Versuchstiere, C = Karotingruppe, NaCl = Kontrollgruppe) Quartil Gruppe n Erstbesamungserfolg in % (n) I C NaCl 22 24 45,45 (10) 29,17 (7) II C NaCl 17 24 23,54 (4) 45,83 (11) III C NaCl 23 19 39,13 (9) 42,11 (8) IV C NaCl 23 18 39,13 (9) 55,56 (10) 155 Tabelle 59: Erstbesamungserfolg zum Zeitpunkt III (n = Anzahl der Versuchstiere, C = Karotingruppe, NaCl = Kontrollgruppe) Quartil Gruppe n Erstbesamungserfolg in % (n) I C NaCl 23 21 52,17 (12) 38,10 (8) II C NaCl 21 21 19,05 (4) 28,67 (6) III C NaCl 21 21 38,10 (8) 47,62 (10) IV C NaCl 20 22 40,00 (8) 54,55 (12) Tabelle 60: Mittlerer Trächtigkeitsindex zum Zeitpunkt I (n = Anzahl der Versuchstiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Gruppe n Trächtigkeitsindex ± SD I C NaCl 18 18 1,44 1,78 0,062 0,06 II C NaCl 17 15 2,00 1,80 1,17 0,86 III C NaCl 12 17 1,92 1,82 0,99 1,01 IV C NaCl 13 17 1,54 1,65 0,78 1,06 Quartil 156 Tabelle 61: Mittlerer Trächtigkeitsindex zum Zeitpunkt II (n = Anzahl der Versuchstiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Gruppe n Trächtigkeitsindex ± SD I C NaCl 18 21 1,67 2,14 0,91 1,15 II C NaCl 8 18 1,88 1,56 1,13 0,78 III C NaCl 17 15 1,76 1,80 0,97 1,01 IV C NaCl 17 13 1,65 1,38 0,86 0,77 Quartil Tabelle 62: Mittlerer Trächtigkeitsindex zum Zeitpunkt III (n = Anzahl der Versuchstiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Gruppe n Trächtigkeitsindex ± SD I C NaCl 17 17 1,53 1,88 0,94 1,05 II C NaCl 14 15 2,07 2,00 0,92 1,00 III C NaCl 15 14 1,67 1,43 0,90 0,85 IV C NaCl 14 21 1,64 1,71 0,93 1,01 Quartil 157 Tabelle 63: Auswertung der Karotingruppe nach Quartilen für Zeitpunkt I: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Quartil I II III IV Variable n Mittelwert ± SD RZ 27 95,15 29,14 GZ 18 111,26 39,20 VZ ZKZ 18 18 23,36 a,b 389,37 34,02 39,56 RZ 22 83,05 26,72 GZ 17 108,76 40,71 VZ 17 ZKZ 17 386,76 40,71 RZ 18 91,11 40,90 GZ 12 120,25 44,61 VZ 12 ZKZ 12 398,25 44,61 RZ 18 82,44 16,39 GZ 13 96,23 35,41 VZ 13 17,69 a 26,20 ZKZ 13 28,56 b 31,33 b 384,21 35,79 35,49 50,66 Zwischen den Quartilen: a,b Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 158 Tabelle 64: Auswertung der Kontrollgruppe nach Quartilen für Zeitpunkt I: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Quartil I II III IV Variable n Mittelwert ± SD RZ 20 84,00 27,29 GZ 18 109,89 41,87 VZ ZKZ 18 18 26,50 387,89 38,89 41,87 RZ 19 87,79 30,38 GZ 15 110,53 39,18 VZ 15 28,93 29,31 ZKZ 15 388,53 29,31 RZ 22 83,76 23,86 GZ 17 108,24 28,99 VZ 17 23,41 27,63 ZKZ 17 386,24 28,99 RZ 24 81,57 22,36 GZ 17 103,35 40,51 VZ 17 23,24 35,11 ZKZ 17 381,35 40,51 159 Tabelle 65: Auswertung der Karotingruppe nach Quartilen für Zeitpunkt II: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Quartil I II III IV Mittelwert ± SD Variable n RZ 22 GZ 17 112,50 36,80 VZ ZKZ 17 17 28,56 390,44 33,44 36,74 RZ 17 101,94 a 30,71 GZ 8 125,25 39,68 VZ 8 30,88 41,05 ZKZ 8 415,67 52,58 RZ 23 GZ 17 103,76 41,97 VZ 17 22,65 32,14 ZKZ 17 381,76 41,97 RZ 23 GZ 18 103,67 41,52 VZ 18 21,44 30,95 ZKZ 18 381,78 41,89 85,36 a,b 79,91 b 90,04 a,b 29,55 25,87 29,79 Zwischen den Quartilen: a,b Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 160 Tabelle 66: Auswertung der Kontrollgruppe nach Quartilen für Zeitpunkt II: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Quartil I II III IV Variable n Mittelwert ± SD RZ 24 82,21 24,60 GZ 21 120,05 45,86 VZ ZKZ 21 21 39,90 b,c 398,05 41,19 45,86 RZ 23 85,74 28,80 GZ 18 103,78 36,32 VZ 18 ZKZ 18 381,78 36,32 RZ 19 83,33 20,79 GZ 15 111,00 33,09 VZ 15 ZKZ 15 389,00 33,09 RZ 19 85,44 28,55 GZ 13 90,69 18,91 VZ 13 ZKZ 13 15,83 c,d 30,80 b,c 9,15 a,d 368,69 22,70 30,77 18,59 18,91 Zwischen den Quartilen: a,b,c,d Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 161 Tabelle 67: Auswertung der Karotingruppe nach Quartilen für Zeitpunkt III: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Quartil I II III IV Variable n Mittelwert ± SD RZ 23 102,04 a 25,90 GZ 17 112,53 30,26 VZ ZKZ 17 17 15,41 390,53 a,b 25,26 30,26 RZ 21 88,19 b 33,19 GZ 13 120,86 33,73 VZ 13 34,00 32,49 ZKZ 13 406,60 b 44,22 RZ 21 77,86 b 27,99 GZ 15 92,53 40,40 VZ 15 24,07 32,28 ZKZ 15 370,53 a 40,40 RZ 20 84,30 b 26,45 GZ 15 110,93 50,88 VZ 15 29,00 40,33 ZKZ 15 389,00 a,b 51,18 Zwischen den Quartilen: a,b Mittelwerte mit unterschiedlichen Buchstabenindices unterscheiden sich signifikant 162 Tabelle 68: Auswertung der Kontrollgruppe nach Quartilen für Zeitpunkt III: Rastzeit (RZ), Güstzeit (GZ), Verzögerungszeit (VZ) und erwartete Zwischenkalbezeit (ZKZ) (n = Anzahl der Tiere, C = Karotingruppe, NaCl = Kontrollgruppe, SD = Standardabweichung) Quartil I II III IV Variable n Mittelwert ± SD RZ 20 82,80 26,83 GZ 17 111,24 40,32 VZ ZKZ 17 17 32,65 389,24 37,90 40,32 RZ 20 89,55 23,63 GZ 15 113,67 35,10 VZ 15 28,20 28,35 ZKZ 15 391,67 35,10 RZ 22 88,24 31,31 GZ 14 109,57 39,26 VZ 14 13,71 29,65 ZKZ 14 387,57 39,26 RZ 23 76,50 18,78 GZ 21 100,14 36,22 VZ 21 25,43 32,75 ZKZ 21 378,14 36,22 163 Danksagung Frau Univ.-Prof. Dr. M. Hoedemaker, Ph.D. danke ich herzlichst für die Überlassung des Themas und die hervorragende Betreuung. Ihre konstruktive Kritik und ihre geduldige Beantwortung aller meiner Fragen hat mir sehr geholfen. Meiner Kollegin Alexandra Conrads danke ich aufrichtig für die freundschaftliche Zusammenarbeit und die selbstlose Unterstützung in einer schweren Zeit. Besonderer Dank gilt den Betreibern und allen Mitarbeitern „unseres Hofes“ in Wasserleben. Obwohl wir zum Teil erhebliche Mehrarbeit verursacht haben, wurde uns unermüdlich und immer gut gelaunt Hilfestellung geleistet. Herrn Marx und Herrn Becker danke ich vor allem für das mühselige und oft stundenlange Einfangen der Kühe. Frau Gallun möchte ich für die Bereitstellung der Daten und die leckere Verpflegung danken. Meiner Familie danke ich dafür, dass sie mir ermöglicht hat, Tierärztin zu werden und dafür, dass ich mich immer auf sie verlassen kann. Lutz danke ich für das geduldige Ertragen meiner Verzweiflungsanfälle und dafür, dass er mir immer Halt und Selbstvertrauen gegeben hat. Meinen Kollegen in der Klinik für Geburtshilfe und Gynäkologie des Rindes danke ich für den Beistand in Motivationstiefs und für das ausgezeichnete Arbeitsklima. Frau Erika Berger gebührt besonderer Dank für die Untersuchung unzähliger Progesteronproben. Die Firma Alvetra GmbH, Neumünster stellte freundlicherweise kostenlos das benötigte Präparat Carofertin zur Verfügung.